
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Anim. Sci. , 04 February 2025
Sec. Animal Nutrition
Volume 6 - 2025 | https://doi.org/10.3389/fanim.2025.1531761
 Farwa Mustafa1Asif Sajjad1*Muhammad Sajjad1†Mudssar Ali3Hafiza Sehrish Bashir1Muhammad Ghazanfar Abbas2Muhammad Binyameen2Raimondas Mozūratis4,5*
Farwa Mustafa1Asif Sajjad1*Muhammad Sajjad1†Mudssar Ali3Hafiza Sehrish Bashir1Muhammad Ghazanfar Abbas2Muhammad Binyameen2Raimondas Mozūratis4,5*Introduction: Insects are receiving increasing attention due to their potential to enhance farming efficiency and sustainability, mitigate vast quantities of bio-waste, and improve animal performance. It is hypothesized that replacing soybean meal with Acheta domesticus will improve broiler productive performance, health, and meat quality. Therefore, the objective of this study was to assess the effect of the gradual replacement of soybean meal (4%, 8%, and 12%) with house cricket (A. domesticus) and black soldier fly (Hermetia illucens) on productive performances, hematology, intestinal morphology, and meat quality attributes of male broiler (Ross 308).
Methods: A total of 350 1-day-old chicks (39.23 ± 0.19 g) were divided into seven groups (five pens per group and 10 chicks per pen) following a completely randomized design. The seven groups included 4%, 8%, and 12% SBM replacements with A. domesticus and H. illucens. Soybean meal was the basal diet considered the control.
Results: The broilers fed 12% A. domesticus, or 12% H. illucens had significantly higher ((p 0.05) live weight, average daily weight gain, and improved feed conversion ratio than the broilers fed basal (SBM) diet throughout starter, grower, and finisher phases. Hematology (complete blood count and serum biochemistry traits) and intestinal morphology (villus height, villus width, crypt depth, and villus height-to-crypt depth ratio) of the broilers improved when fed on 12% A. domesticus and 12% H. illucens meals as compared to the control group (p < 0.05). Feeding 12% A. domesticus and 12% H. illucens also had a significant positive impact on the meat quality traits of broilers, such as maximum redness (a*) and yellowness (b*) with minimal cooking loss and lightness (L*) (p < 0.05) than the broilers fed the control diet.
Discussion: Using insect meal as an alternative to traditional feed sources could contribute significantly to the sustainable expansion of the poultry industry.
Soybean meal (SBM) and fish meal (FM) are the predominant protein sources in poultry feed. Still, there is limited land available worldwide for soybean cultivation. Overexploitation of marine resources has led to a significant decline in small pelagic forage fish, which are critical for producing FM and fish oil (Veldkamp et al., 2022). Moreover, the increasing expenses associated with these traditional protein sources posed a threat to the sustainable future of the poultry industry (McMichael and Bambrick, 2005). Furthermore, food insecurity is a pressing issue in numerous developing countries, and there are expected challenges in supplying food for a projected population of over 9 billion people in 2050 (Makkar et al., 2014). This situation has intensified the quest for alternative protein sources to fulfill the nutritional demands of humans and animals.
Insects are emerging as a novel alternative protein source, promoting sustainable growth in the poultry sector (Sánchez-Muros et al., 2014). They have the potential as a standard protein source in animal feedings, i.e., poultry, pigs, and fish, owing to their high-quality protein content (25%–75% on a DM basis) and suitable amino acid composition and fatty acid composition (n-3 FA) (Henry et al., 2015; Kulma et al., 2020; Van Huis, 2013; Veldkamp and Bosch, 2015). Moreover, they have minimal competition with human food resources and are eco-friendly in terms of energy cost, land, and footprints (Ballitoc and Sun, 2013), which makes insects a promising option from an ecological standpoint (Makkar et al., 2014; Oonincx and De Boer, 2012; Sánchez-Muros et al., 2014).
Several types of insects, including black soldier fly (Hermetia illucens L.), yellow mealworm (Tenebrio molitor L.), house fly (Musca domestica L.), mulberry silkworm (Bombyx mori L.), and grasshopper (Caelifera spp.), have been recognized as promising sources of protein to replace conventional protein sources in animal feeds (Van Huis et al., 2013). Free-range birds voluntarily consume insects as a natural protein source (Adetunmbi, 2023; Hwangbo et al., 2009; Ijaiya and Eko, 2009; Khatun et al., 2003; Sajjad et al., 2024a). The nutritional value of insects can oscillate with the species, rearing substrate, development stages, and handling methods (Danieli et al., 2019; Gasco et al., 2020; Slimen et al., 2023). Incorporating insects into poultry feed instead of SBM or FM did not adversely affect the broilers’ growth performances (Adeniji, 2007; Oyegoke et al., 2006; Wang et al., 2005). Moreover, insect meal improves carcass traits in terms of dressing percentage, breast muscle, thigh muscle, slaughter, dressed carcass, and eviscerated weights in broilers (Ballitoc and Sun, 2013; Hwangbo et al., 2009; Khatun et al., 2003; Sajjad et al., 2024b).
Previous studies have demonstrated that the growth performance, blood hematology, gut morphology, and meat quality of broilers (Benzertiha et al., 2019), Japanese quails (Zadeh et al., 2019), and barbary partridges (Loponte et al., 2017) are improved by the inclusion of H. illucens and T. molitor in their diets (Benzertiha et al., 2020; Khan et al., 2018). The growth performance and nitrogen balance of Ross 308 are enhanced with 2.6% H. illucens meal in the starter phase (Neumann et al., 2018). Moreover, meals containing 5% to 7.5% H. illucens improved the feed efficiency and increased thigh weight while reducing meat pH, whereas 10% H. illucens boosted growth in Cobb 500 broiler chickens (Dahiru et al., 2016). Furthermore, it was noted that substituting partial or all of the soybean oil with H. illucens larvae fat had no adverse impact on the growth performance of young turkeys (Sypniewski et al., 2020). The inclusion of 20% H. illucens in the diet of Ross 308 male broilers improved meat quality by elevating levels of lauric acid, myristic acid, and eicosapentaenoic fatty acid, while also slightly decreasing the total polyunsaturated fatty acid (PUFA) content (Vilela et al., 2021). The diet with 5% H. illucens resulted in a decrease in abdominal fat; 10% H. illucens led to an increase in carcass and breast weight, while 15% inclusion of H. illucens increased the body weight, abdominal fat, meat redness, protein content in meat, and the levels of monounsaturated fatty acids (MUFAs) in breast meat while reducing PUFA in breast meat (Schiavone et al., 2019). Gut health and functioning are mainly associated with the intestinal morphology of the broilers, which is affected by the dietary protein level and digestibility of the ingested food (Kuzmuk et al., 2005; Laudadio et al., 2012; Qaisrani et al., 2014). Intestinal morphology can be assessed with the development of morphometric indices such as villus height (Vh) and crypt depth (Cd) (Franco et al., 2006; Laudadio et al., 2012).
The poultry industry depends on SBM to meet the demand for protein in animal feeds, as only a few insect species are currently utilized at an industrial scale (Sajid et al., 2023). Therefore, it is a smart choice to focus on local insect species as substantial feed ingredients in animal sectors (Sajjad et al., 2024c).
The house cricket (Acheta domesticus L.) (Orthoptera: Gryllidae) has been utilized globally as feed for insectivores (Kulma et al., 2019) because of its high protein content (60%–70% dry weight) with all essential amino acids and 10%–23% lipids (Udomsil et al., 2019). Moreover, it has high levels of omega-3 and omega-6 fatty acids and minerals such as P, Na, and Ca (Han and Heinonen, 2022; Oonincx and Finke, 2021). Furthermore, it can potentially transform organic wastes, including food scrap and agricultural by-products, into nutritious biomass that is beneficial for animal feeds, particularly for poultry and aquaculture (Van Peer et al., 2021). Cricket farming generates high-quality by-products, which can be used as organic fertilizer (Halloran et al., 2017). Furthermore, the European Union has recently authorized the inclusion of H. illucens, M. domestica, T. molitor, Alphitobius diaperinus, A. domesticus, Gryllodes sigillatus, and Gryllus assimilis as a protein source in poultry and pig feed (Schiavone and Castillo, 2024); however, there is limited information regarding the effects of partial replacement of SBM with A. domesticus on the productive performance, health, and meat quality traits of Ross 308 broiler. It is hypothesized that the replacement of SBM with A. domesticus will improve broiler productive performance, health, and meat quality. The aim of the present study was to evaluate the effects of replacing 4%, 8%, and 12% SBM with A. domesticus and H. illucens on the productive performance, hematology, intestinal morphology, and meat quality of broiler chickens.
The biological trials on broiler Ross 308 were carried out for 35 days under controlled conditions in A block, University of Animal and Veterinary Sciences (UVAS), Ravi Campus, Pattoki, Lahore, Pakistan. The Ethical Review Committee (No. DR/495) at UVAS approved all the procedures.
Initially, house crickets (A. domesticus) were collected using an aerial net from neglected kitchens of homes and hotels, and then the stock culture on chicken waste was established, while black soldier flies H. illucens were also obtained from the established culture under controlled conditions (28°C ± 2°C, relative humidity 65% ± 5%, and 16 h light:8 h dark) at the Department of Entomology, the Islamia University of Bahawalpur. Mature larvae of H. illucens (18 days old) and adult A. domesticus (70 days old) were harvested from the chicken waste and boiled at 100°C for 3 min to ensure they were killed, and feed and fecal remnants were removed and subsequently oven-dried at 60°C for 24 h (Tan et al., 2018). The dried insects were ground in a blender and stored at −20°C. The larvae of A. domesticus and H. illucens were analyzed for their nutritional compositions, energy contents, and amino acids (% on DM basis) using proximate analyses and an amino acid analyzer at the Department of Animal Nutrition at UVAS (Table 1).
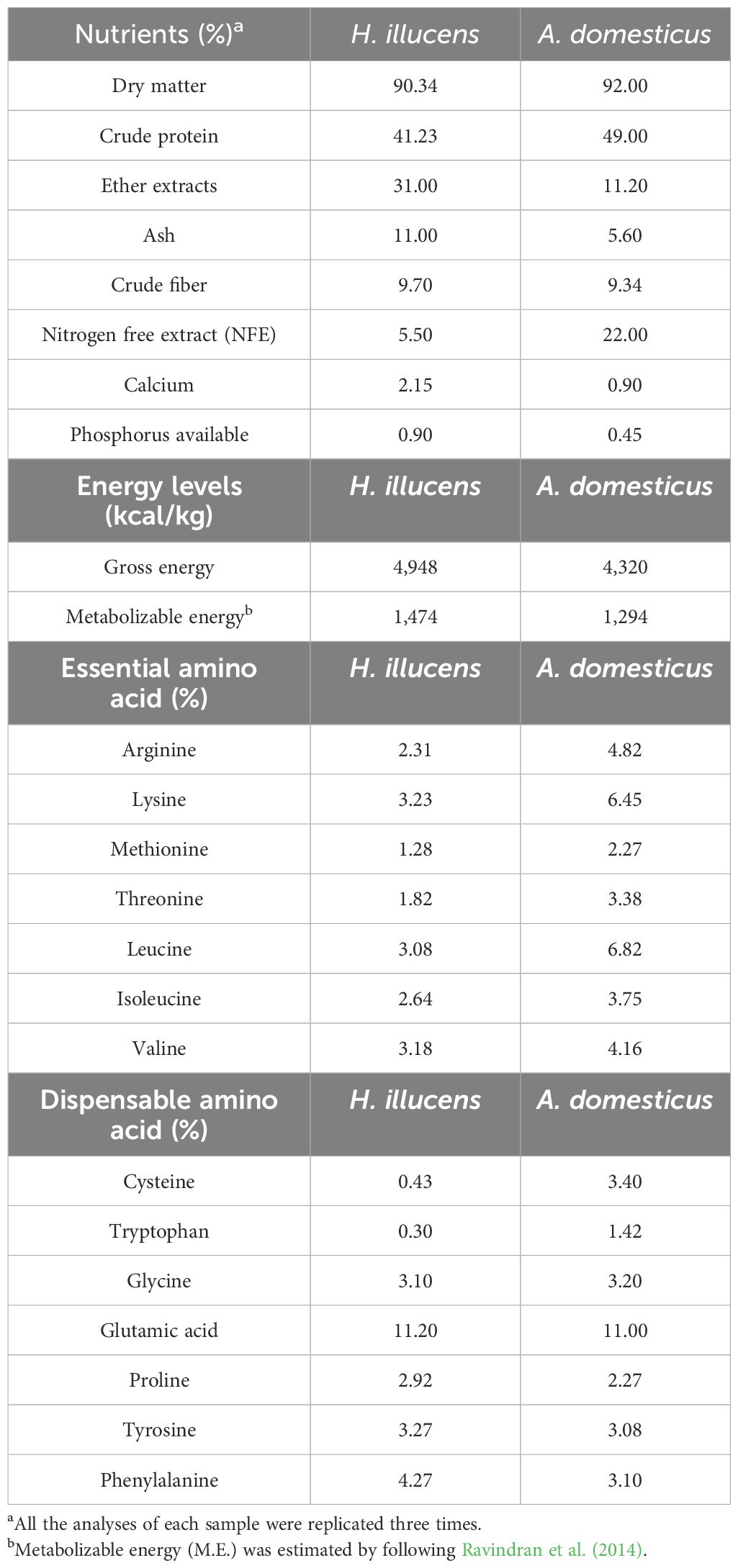
Table 1. The nutrients, energy contents, and amino acids of H. illucens and A. domesticus larvae.
Male Ross 308 was used to assess the impacts of 4%, 8%, and 12% substitution of SBM with A. domesticus and H. illucens. A total of 350 1-day-old chicks (39.23 ± 0.19 g) were allotted to seven dietary treatment groups, with five pens per treatment and 10 birds per pen. The birds were placed in spotless and disinfected pens equipped with feeders, drinkers, heat lamps, and bedding materials (3 to 4 inches of rice husk). The chicks had ad libitum feed, water, and light. Temperature was maintained at 35°C during the initial week, subsequently adjusting to 28 ± 2°C till the completion of experiments, while the relative humidity was kept at 50% ± 5%. The photoperiod was maintained at 8 h of light and 6 h of darkness using artificial lighting. The birds were vaccinated for numerous diseases as per the set protocols by the veterinarian. Enclosures were inspected daily to assess clinical signs and record any mortality among the birds.
SBM was progressively substituted at the levels of 4%, 8%, and 12% with A. domesticus and H. illucens by following the guidelines at the levels of Ross (2022). All the dietary treatments were formulated into three different phases, i.e., starter (1–10 d), grower (11–24 d), and finisher (25–35 d). Ingredients, along with the calculated and analyzed profiles of the starter, grower, and finisher meals, are presented in Tables 2–4.
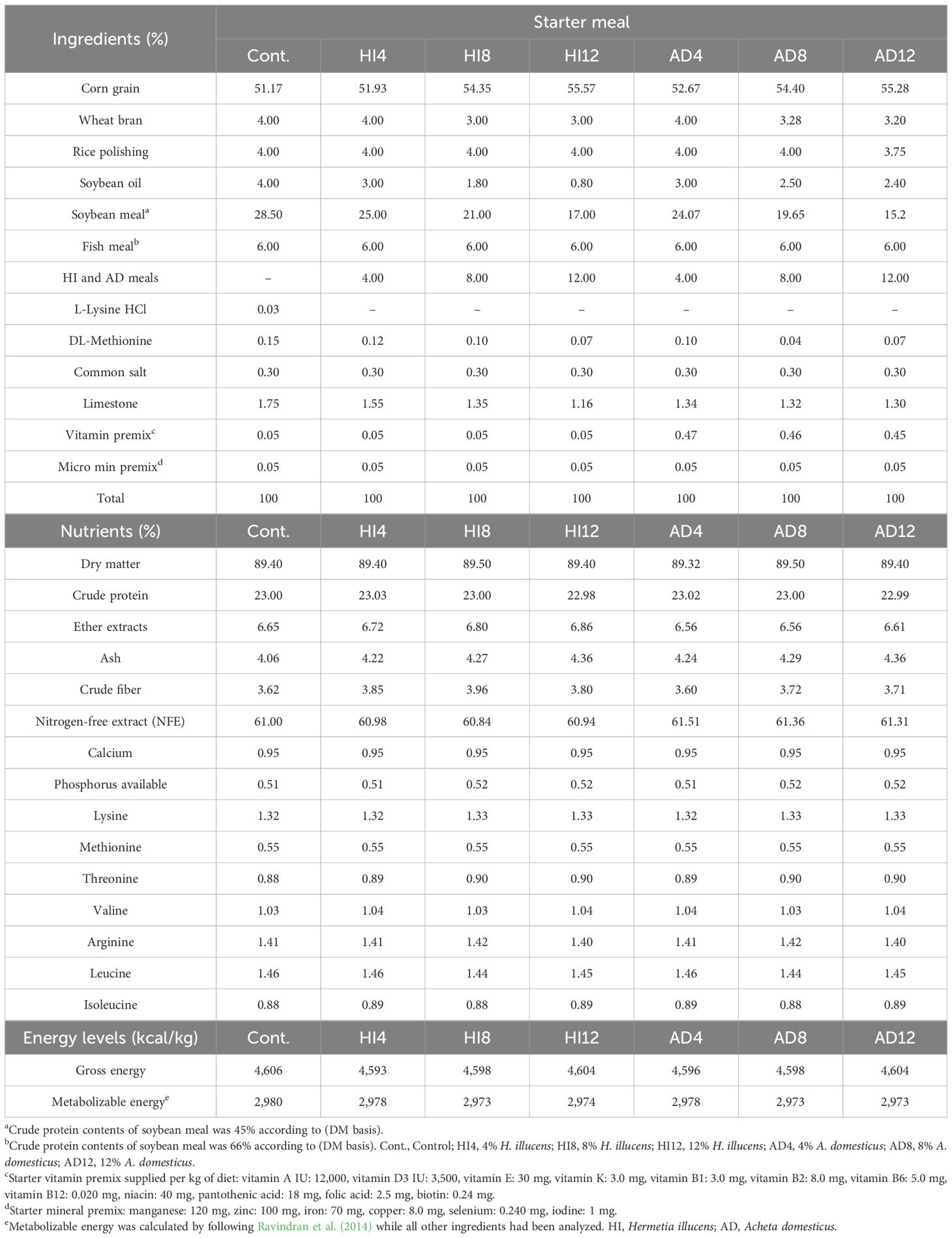
Table 2. The ingredients, chemical compositions, and energy contents of starter meals.
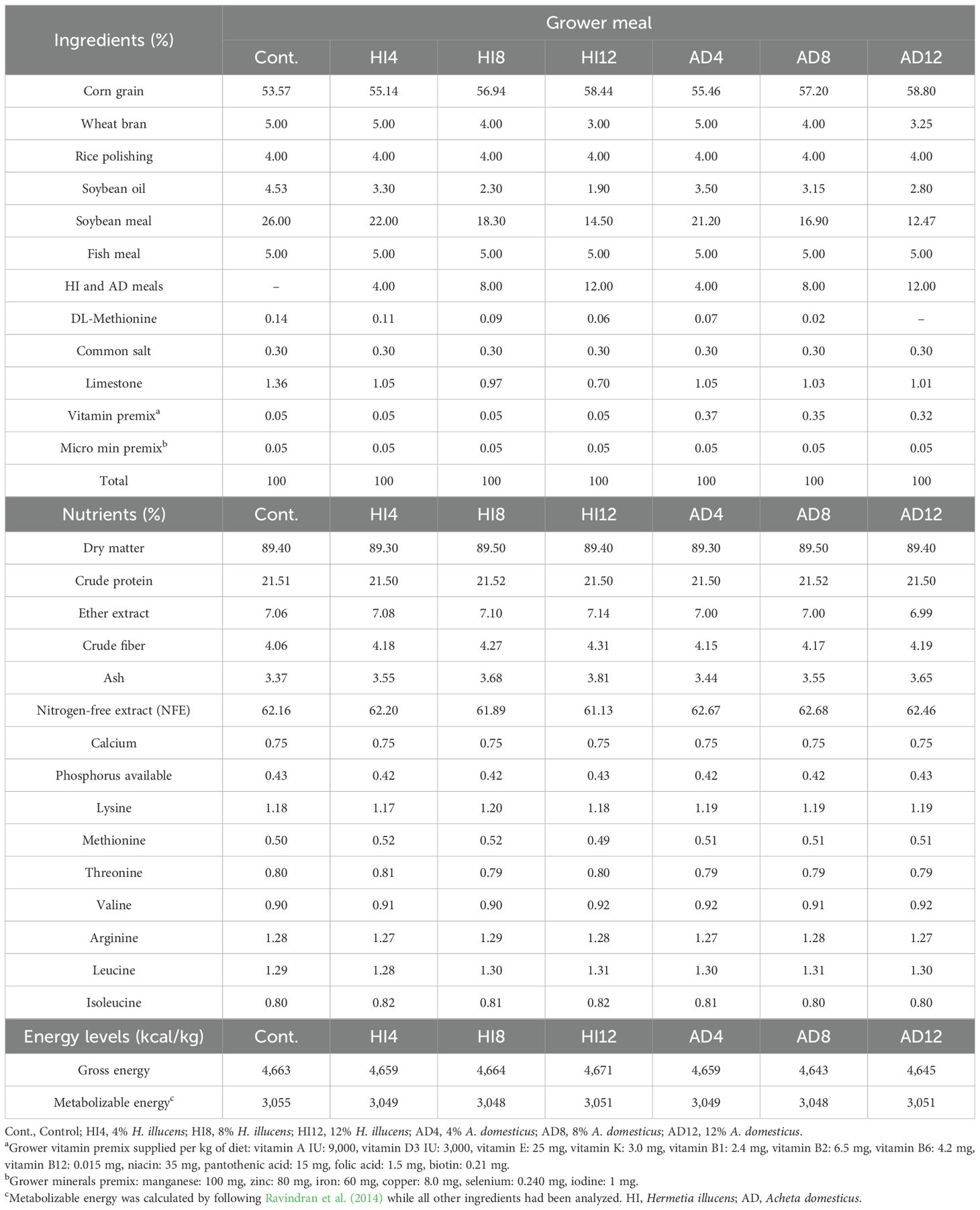
Table 3. The ingredients, chemical compositions, and energy contents of grower meals (% as fed).
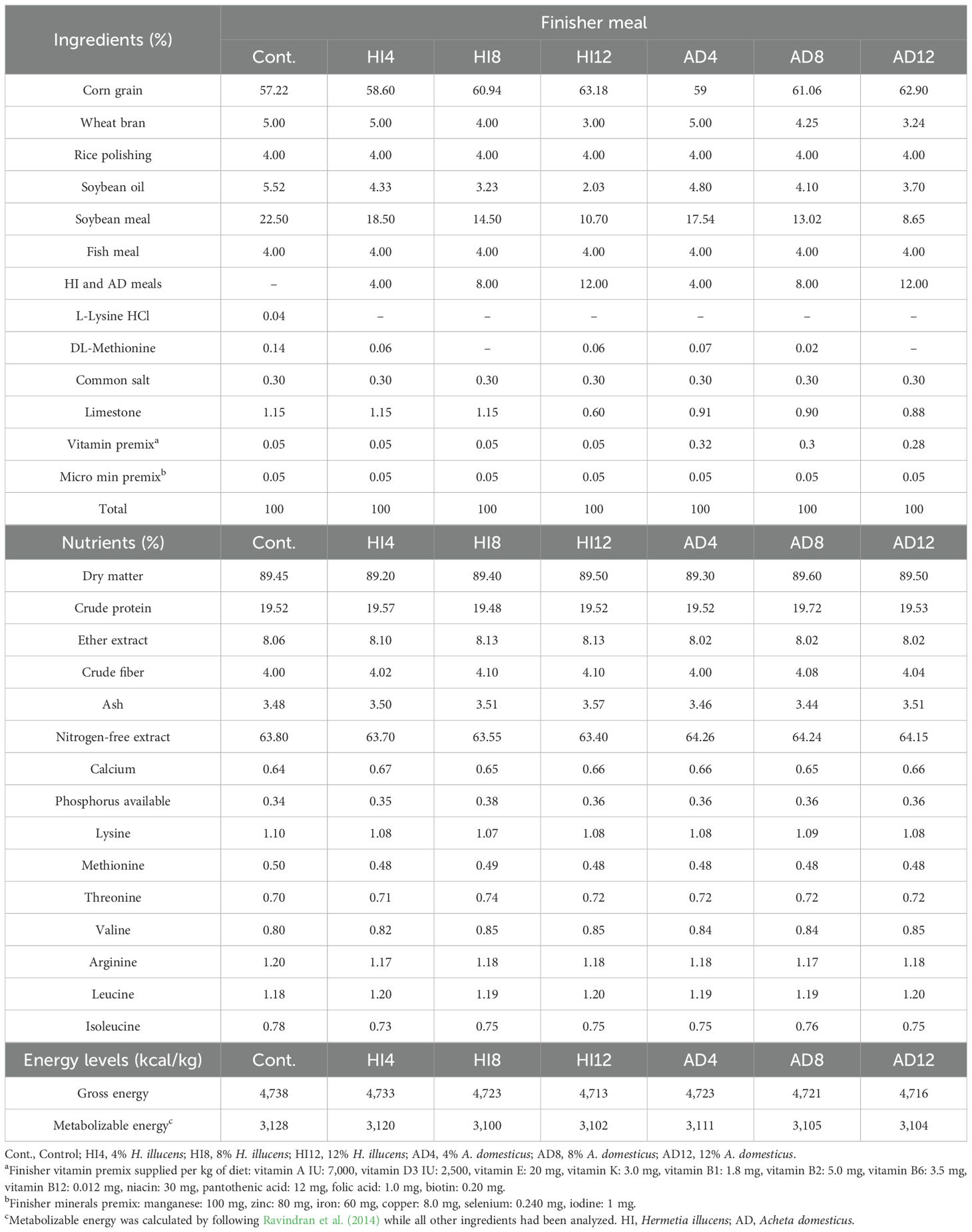
Table 4. The ingredients, chemical compositions, and energy contents of finisher meals (% as fed).
Bird’s performances, such as live weight (LW), average daily weight gain (ADG), daily feed intake (DFI), and feed conversion ratio (FCR), were calculated over a 35-day feeding period. The LW was recorded individually at the onset of the experiment and subsequently at intervals of 10 d, 24 d, and 35 d. Daily weight gain and feed intake were determined on both individual and pen bases following each growth phase, whereas the feed conversion ratio was calculated for each growth phase and for the intact intervals of the trial.
On day 35, two chicks per pen were euthanized shortly after their arrival by a skilled professional using the rapid decapitation technique. Consequently, 2.5 mL of blood was collected into EDTA and serum-separating tubes. Hematology and serum biochemistry parameters were tested by following Campbell (1995).
The small tissues (2–3 mm) from jejunum (JE) and ileum (IL) sites were excised from the slain birds, promptly washed with normal saline, and stored for 72 h in 10% formalin solution by following Laudadio et al. (2012). The transverse sections were studied by taking images under a light microscope, and villus height (Vh), villus width (Vw), crypt depth (Cd), and Vh/Cd were determined by the ImageJ software (Ferreira and Rasband, 2012).
Pectoralis major was used to evaluate meat quality parameters such as meat pH, drip loss, cooking loss, shear force, and meat color, including lightness (L*), redness (a*), yellowness (b*), chroma (C*), and hue (H*) from the broiler. The deboned meat samples were refrigerated at 4°C overnight before the analyses. Cooking and drip losses were assessed as described by Kaić et al. (2021) and Zaid et al. (2020). Meat pH was tested using pH meter 3210 SET 2 from breast samples. The CR-410 colorimeter was used to determine meat color by following Priolo et al. (2002). The breast meat was cut into a rectangular shape (1 H × 1 W × 2 L, in cm) parallel to muscle fibers using a scalpel handle blade. TAXT plus 100C texture analyzer was used to assess the Warner–Bratzler shear force (N/cm2) with the help of a V-slot blade. These analyses were carried out from UVAS, Lahore.
The pens served as the trial units for assessing productive performances, while individual birds were used to determine hematology, intestinal morphology, and meat characteristics. The significant differences among the treatment groups were analyzed using a completely randomized design (CRD) with a one-way analysis of variance followed by the Duncan Multiple Range (DMR). Significant statistical differences were accepted at p < 0.05, while values in the range of 0.05 ≤ p < 0.10 were considered to indicate a trend. Orthogonal polynomial contrasts were used to assess the linear and quadratic effects. A general linear model (GLM) was performed to assess the impacts of dietary groups, intestinal sections, and their interactions. All the analyses were performed with IBM SPSS Statistics software (version 21 for Windows, SPSS Inc., Chicago, IL, USA).
The productive performances, i.e., live weight, average daily weight gain, daily feed intake, and feed conversion ratio of the broilers fed with different meals, are summarized in Table 5. These parameters significantly differ (p < 0.05) among all the dietary treatments except for the daily feed intake (p > 0.05) across the feeding intervals. In starter and finisher phases, the LW was the highest in 12% A. domesticus and 12% H. illucens and the lowest in the control treatment. It was the maximum in 12% A. domesticus and 12% H. illucens and the minimum in 4% H. illucens in the grower phase. The LW was affected in a linear and quadratic manner among the H. illucens meals during the starter and finisher phases, while a linear response was seen during the grower phase. Similarly, LW responded linearly and quadratically to the A. domesticus meals during the feeding intervals.
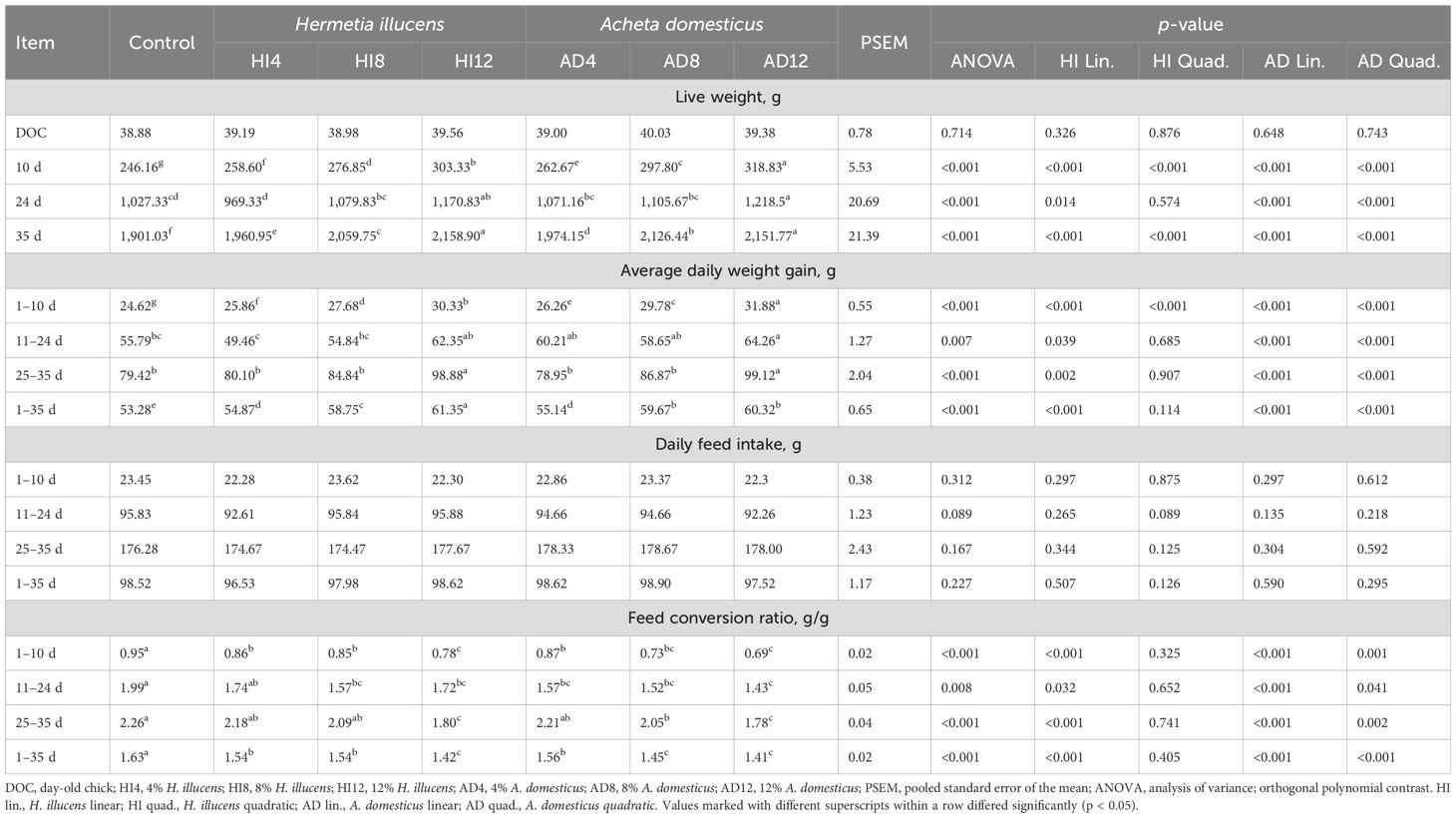
Table 5. Impacts of H. illucens and A. domesticus dietary groups on the productive performances of broilers.
During days 1–10, daily weight gain was the maximum in 12% A. domesticus and the minimum in the control group. During days 11–24, it was the highest in 12% A. domesticus and the lowest in 4% H. illucens. Similarly, maximum ADG was found in meals containing 12% A. domesticus and 12% H. illucens on days 25–35, and the minimum was found in the remaining treatments, whereas the highest daily weight gain was recorded in 12% H. illucens and the lowest in the control group. Polynomial contrast presented linear and quadratic effects in ADG during days 1–10, while a linear response was seen during the remaining feeding periods among the meals of H. illucens; likewise, there were linear and quadratic effects in ADG across the feeding periods among the A. domesticus meals.
The maximum FCR was registered in the broilers fed the control meal, while the minimum was registered when fed the 12% A. domesticus and 12% H. illucens across the feeding intervals. The H. illucens meals affected FCR in a way across the feeding periods. Likewise, there were linear and quadratic effects in FCR across the intervals among the A. domesticus meals.
Hematology and serum biochemistry traits are mentioned in Table 6. The hematological parameters showed statistical differences (p < 0.05) among all the treatments except for monocytes (p > 0.05). The maximum hemoglobin (Hb), red blood cells (RBCs), hematocrits (HCT), platelets, total leucocytes (TLC), and lymphocytes (Lyn) were registered in 12% A. domesticus and 12% H. illucens and the minimum values were registered in 4% H. illucens. Mean corpuscular volume (MCV), mean corpuscular hemoglobin (MCH), and mean corpuscular hemoglobin concentration (MCHC) were the highest in 12% A. domesticus and the lowest in the control group. HB, HCT, MCV, MCH, MCHC, TLC, heterocysts, and lymphocytes showed linear and quadratic responses, while a quadratic response to the H. illucens meals was found in RBCs and platelets. Similarly, HB, HCT, MCV, MCH, MCHC, platelets, and TLC were affected linearly and quadratically, while heterocysts and lymphocytes responded linearly to the A. domesticus meals.
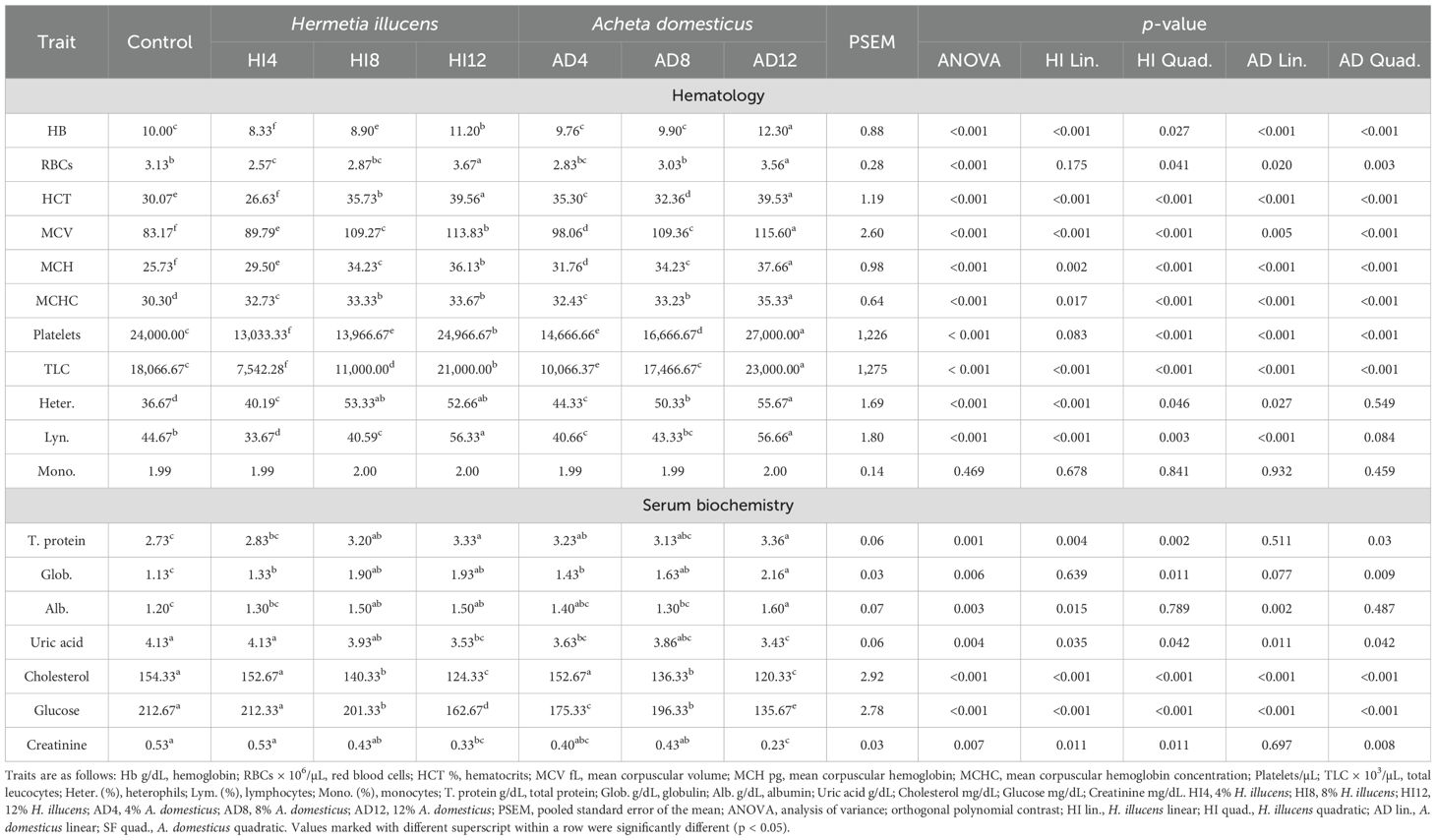
Table 6. Effects of H. illucens and A. domesticus meals on the hematology and serum biochemistry traits of broilers.
The serum biochemistry traits differed statistically (p < 0.05) among the meals. Total protein, globulin, and albumin were the highest in the broilers fed 12% A. domesticus, while they were the lowest in the control meals. The maximum concentrations of uric acid, cholesterol, glucose, and creatinine were found in the control group, and the minimum concentrations were found in 12% A. domesticus meals. The orthogonal contrast depicted linear and quadratic responses in total protein, uric acid, cholesterol, glucose, and creatinine, while a quadratic effect in globulin and a linear effect in albumin were seen among H. illucens meals. Likewise, total protein, globulin, and creatinine registered a quadratic effect; albumin showed a linear response, while uric acid, cholesterol, and glucose presented linear and quadratic effects among A. domesticus meals.
The dietary treatments, sites of the small intestine, and their interactions differed statistically (p < 0.05) (Table 7). The Vh was the longest and similar at the jejunum and ileum when broilers were fed 12% A. domesticus, and at the jejunum site fed 12% H. illucens, 8% H. illucens, and 8% AD, while it was the shortest at the ileum site in the broilers fed the control diet and 4% H. illucens (Figure 1A). The highest Cd was observed at the jejunum in those fed the control diet and the lowest at the ileum site in those fed 12% A. domesticus (Figure 1B). The Vw was the largest and comparable at both sites when fed 12% A. domesticus, and at the jejunum site when fed 12% H. illucens, 8% H. illucens, and 8% A. domesticus, while it was the smallest at the jejunum and ileum sites in the broilers fed the control diet, 4% H. illucens, and 4% A. domesticus and at the ileum site in the broilers fed 8% H. illucens and 8% A. domesticus (Figure 1C). Vh/Cd was the largest and similar at the jejunum and ileum sites in the broilers fed 12% A. domesticus, and at the jejunum site in the broilers fed 12% H. illucens, 8% H. illucens, and 8% A. domesticus. The lowest Vh/Cd ratios were recorded in the control and 4% H. illucens (Figure 1D).
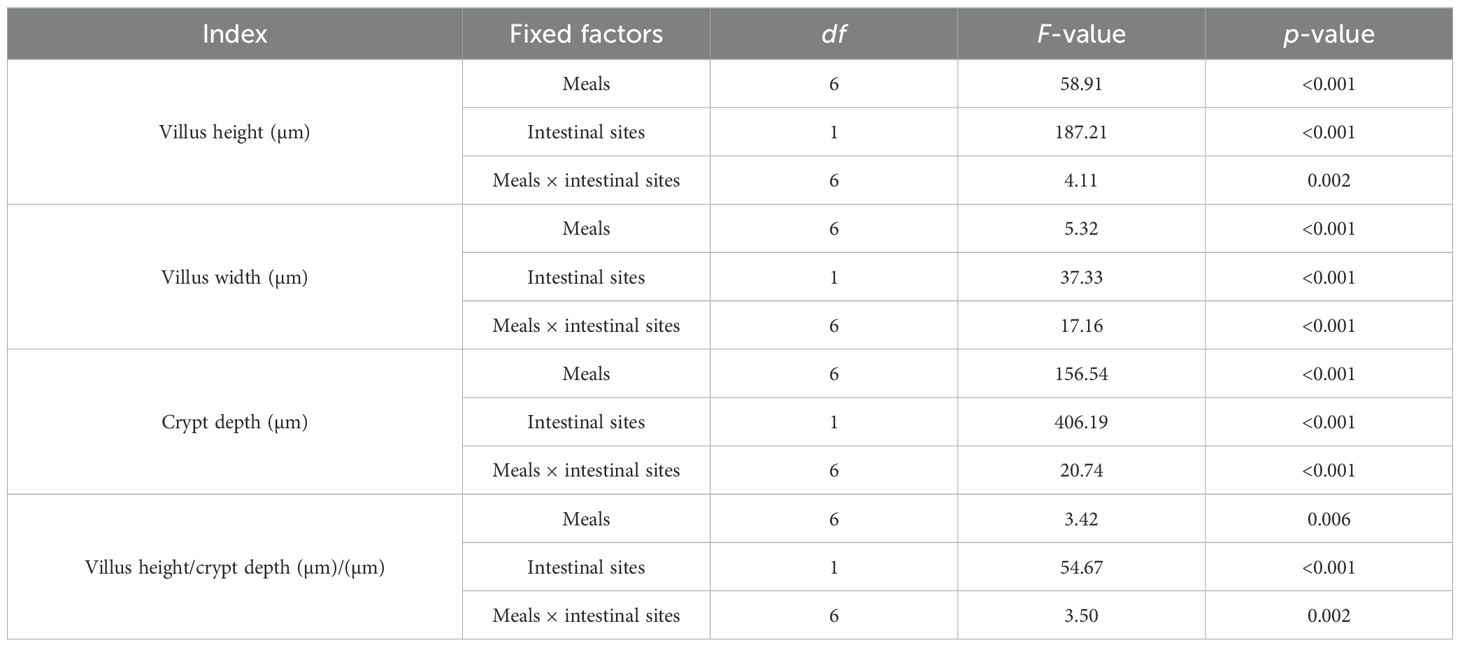
Table 7. The impact of dietary treatments, intestinal sites, and their interactions on the gut morphometric indices.
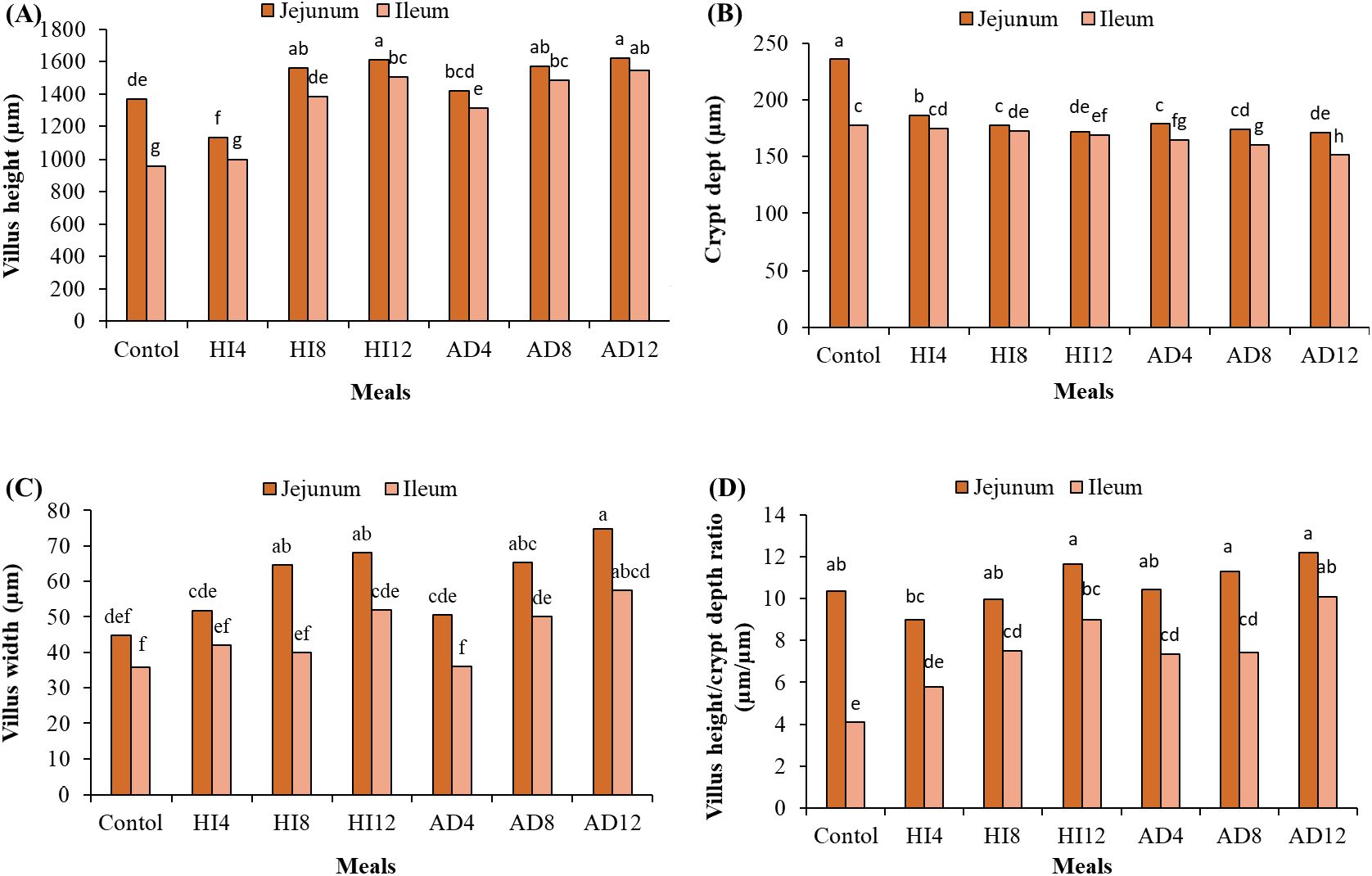
Figure 1. Interactions between intestinal sites and dietary treatments; villus height (A), crypt depth (B), villus width (C), and villus height/crypt depth ratio (D). Values marked with different letters indicate statistically significant differences (p < 0.05).
Meat quality traits are presented in Table 8. These parameters differed significantly (p < 0.05) among all the dietary treatments except for the drip loss, pH, shear force, and chroma (p > 0.05). In the control treatment, maximum cooking loss and L* were recorded, with the minimum found in the 12% A. domesticus meal. The a* was the highest in 12% A. domesticus and the lowest in the control treatment. The b* and H* were the highest in 12% A. domesticus and 12% H. illucens, and the lowest in control. Cooking loss, a*, and b* showed linear and quadratic responses, while quadratic responses were found in L* and H* in the H. illucens groups. Likewise, A. domesticus meals exhibited a quadratic effect on cooking loss, and a linear effect on L*, a*, b*, and H* of the broilers.
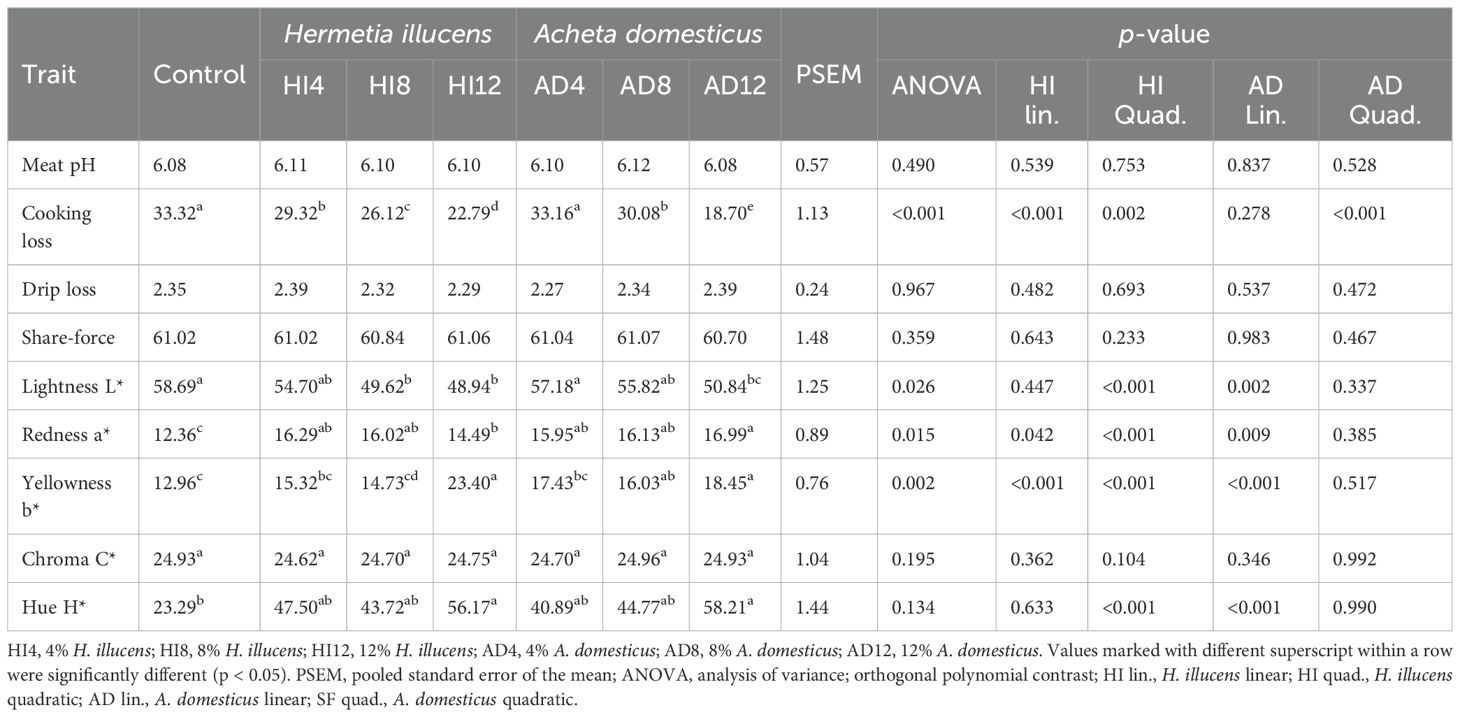
Table 8. Impact of H. illucens and A. domesticus meals on the meat quality of broilers.
The findings showed that live weight and average daily weight gain were highest in the birds fed with diets containing 12% A. domesticus and 12% H. illucens. Constituents of the diets significantly impact the efficiency of the birds in poultry farming, particularly the protein content and amino acid profiles, which are crucial for growth performances. SBM and FM have lower protein contents and lower-quality amino acids compared to insects (Fisher et al., 2020; Hasnan et al., 2023). Insects such as H. illucens have well-balanced essential amino acids, with methionine 2.2% and lysine 6.1% (García-Vaquero and García, 2024; Hobbi et al., 2022), along with vitamins, while the SBM contains methionine 0.65% and lysine 2.95% (Akhtar and Isman, 2018). Moreover, insects offer superior fatty acids than SBM (Islam et al., 2022; Lu et al., 2022). Insects possess a well-balanced nutritional profile, including essential vitamins and minerals that promote robust and fast growth (Adli, 2021; Choi et al., 2023; da-Silva et al., 2024; Rehman et al., 2019). This study is aligned with those of Loponte et al. (2017), Pieterse et al. (2019), and Dabbou et al. (2018), indicating that substituting SBM at the levels of 10%, 15%, and 50% with BSF resulted in enhanced LW and ADG in Ross 308, Cobb 500, and barbary partridges. Many other previous studies by Biasato et al. (2018) and Vasilopoulos et al. (2023) have presented a substantial increase in live weight and daily weight gain in fast-growing Ross 708 and Ross 308 as well as intermediate-growing Hubbard hybrid when fed 10% to 15% T. molitor meals. Conversely, Ramos-Elorduy et al. (2002) and Biasato et al. (2016) did not report any impact on performances on fast-growing Arbor acres and Hubbard hybrid when fed T. molitor meals.
In the present work, daily feed intake did not change significantly among diets. However, feed intake typically increases when the protein and lipid contents decrease in the feed (Van Harn et al., 2019). The feed color might also affect the feed conversion ratio (Hartinger et al., 2021). Chu et al. (2020), Dabbou et al. (2021), and Sajjad et al. (2024c) registered comparable results in broiler Ross 308.
In this study, the feed conversion ratio was improved when broilers were fed 12% A. domesticus and 12% H. illucens. The feed conversion ratio is the most imperative indicator of meal efficiency in livestock production (Davison et al., 2023; Prakash et al., 2020). It can decrease feed conversion ratio by replacing SBM with insect meal owing to the highest nutrients, balanced AAs, palatability, digestibility, and lower anti-nutritional aspects (Parrini et al., 2023; Siddiqui et al., 2024). This study’s results are analogous to the findings of Schiavone et al. (2017), who indicated that replacing soybean oil with H. illucens, either partially or completely, enhanced the feed conversion ratio in Ross 308. The study by Kim et al. (2020) and Lalev et al. (2022) revealed that the inclusion of 5% and 10% H. illucens and B. mori in broiler meals led to significant improvement in feed conversion ratio in broilers. Bovera et al. (2015) and Kierończyk et al. (2018) found an improvement in the feed conversion ratio in Ross 308 feeding 10% to 30% T. molitor. Ognik et al. (2020) reported that 15% H. illucens improved FCR in young turkeys, while Murawska et al. (2021) found no improvement in feed conversion ratio in Ross 308, even at 75% H. illucens.
In this investigation, 12% A. domesticus and 12% H. illucens resulted in the highest Hb, RBCs, HCT, MCV, MCH, MCHC, platelets, total leucocytes, and lymphocytes. Insects have a superior nutritional profile compared to traditional protein sources, which may impact the hematology of the broilers (Marono et al., 2017; Zulkifli et al., 2022). They contain higher concentrations of essential amino acids, absorbable minerals (iron and zinc), and functional lipids, which can boost blood traits by promoting RBC production, hemoglobin levels, and the overall health of the birds (Slimen et al., 2023; Zhou et al., 2022). The higher iron content in insect meal is vital for the synthesis of hemoglobin and erythropoiesis in the broiler (Elliott, 2008; Sajjad et al., 2024a). Zinc is another mineral abundant in insects that plays an essential role in enzymatic functions and the development of immune cells, potentially enhancing leukocyte production and strengthening the immune system (Vasilopoulos et al., 2024a). This study’s results are consonant with Biasato et al. (2017), who noticed that hematological traits improved in broilers Ross 708 when fed insect-based meals. Gariglio et al. (2019), Kim et al. (2020), Schiavone et al. (2017), and Ognik et al. (2020) depicted that supplanting 4% to 100% soybean oil or SBM with insects did not affect the hematological attributes. Lymphocytes were increased when the broiler was fed 20% H. illucens (De Souza Vilela et al., 2021).
In this inquiry, the diets containing 12% A. domesticus and 12% H. illucens exhibited the lowest concentrations of creatinine, uric acid, glucose, and cholesterol, while showing the highest levels of total protein, albumin, and globulin. The chitin and chitosan of insect exoskeletons could potentially have chelating effects that lower the concentrations of uric acid, glucose, and cholesterol in the blood (Hossain and Blair, 2007; Khambualai et al., 2008). This, in turn, may enhance the metabolic functioning of the bird (Alagawany et al., 2022; Sypniewski et al., 2020). The nutritional value of the feed may strengthen the immunity system of birds and increase their resistance against several infectious diseases (Schiavone et al., 2017). Total protein and globulin levels are usually influenced by isoenergetic and isoprotenic diets (Bovera et al., 2016). This study’s results are consistent with previous findings by Sedgh-Gooya et al. (2021) and Bovera et al. (2018), who revealed that 5% to 100% inclusion of T. molitor meals increased the concentrations of total protein and globulin and diminished the level of uric acid in Shaver brown broilers and Bovans white laying hens.
In the current study, the interactions between the dietary treatment and the sites of the small intestine showed significant differences. The villus height, villus width, and villus height/crypt depth ratio were the largest at the jejunum and ileum sites of the intestine using 12% A. domesticus and 12% H. illucens with the smallest crypt depth. The morphology of the intestine is affected by the consumption of insects, which have a unique composition of bio-active compounds, including chitin, antimicrobial peptides, and short-chain fatty acids (Vasilopoulos et al., 2024b). Moreover, chitin may act as a prebiotic that stimulates beneficial gut microbiota such as Lactobacillus and Bifidobacterium, enhancing the gastrointestinal tract’s health and villus growth (Mohan et al., 2023). Furthermore, antimicrobial peptides might contribute to maintaining intestinal integrity by decreasing the load of pathogenic bacteria, which decreases inflammation and promotes healthy villus development (Patyra and Kwiatek, 2023). Nutrient absorption is primarily related to histological indices of the intestine, particularly the villi and crypts (Amer et al., 2021). The longest villi and the shortest crypts play a fundamental role in the efficient digestion and absorption of nutrients (Ravindran and Abdollahi, 2021). This study’s results are aligned with Sajjad et al. (2024a) and Sajjad et al. (2024c), who found comparable effects in Ross 308 when fed 12% Spodoptera frugiperda and 12% A. diaperinus. Another study by Dabbou et al. (2018) and Dabbou et al. (2021) reported that the gut histological indices improved in broilers feeding on defatted and modified H. illucens meals.
The findings of this study presented that 12% A. domesticus and 12% H. illucens diets had the maximum a*, b*, and H* values, along with the minimum cooking loss and L* values. The physical meat and skin properties, such as color and tenderness, do not differ, as color can affect consumer acceptance (Bovera et al., 2016; Yang et al., 2011). High concentrations of lauric acid, myristic acid, and eicosapentaenoic acid, as well as the pigments of the feed ingredients, can influence the color and texture of meat (Cullere et al., 2019; Vilela et al., 2021). Replacing SBM (7.8%) with H. illucens reduced cooking loss in broiler Ross 308 (Leiber et al., 2017). Murawska et al. (2021), Kim et al. (2019), and Sajjad et al. (2024a) found that increasing the levels of H. illucens and lesser mealworm A. diaperinus in the feeds dwindled cooking loss while increasing the b* (Ross 308). However, several factors, including feed, stage, breed, and slaughtering techniques, can significantly affect the meat (Mir et al., 2017).
Incorporating local insect species as feed ingredients in the poultry sector offers several advantages. Utilizing locally sourced insects can reduce dependency on imported feed ingredients, lower feed costs, and contribute to a more sustainable and environmentally friendly poultry industry. Additionally, insect farming has a smaller carbon footprint, minimizes waste, and promotes circular economies (Madau et al., 2020).
Utilizing H. illucens and A. domesticus as a protein source significantly impacted live weight, average body weight gain, feed conversion ratio, hematological traits, gut histology, and meat quality in Ross 308. Overall, the birds fed with 12% H. illucens and 12% A. domesticus for 35 days performed better than those fed other diets. Therefore, it can be concluded that H. illucens and A. domesticus are viable alternative ingredients that can substitute SBM to develop a local-based feed ingredient for broiler chickens at the optimal inclusion rate of 12% without compromising performance and health as evidenced by blood parameters, gut histology, and meat quality of the birds. Future research should investigate how H. illucens and A. domesticus meals affect the gut microbiota of broilers when utilized as a principal protein source in poultry feed.
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding authors.
The animal study was approved by University of Animal and Veterinary Sciences (UVAS), Ravi Campus, Pattoki, Lahore, Pakistan. The Ethical Review Committee (No. DR/495) at UVAS approved all the procedures. The study was conducted in accordance with the local legislation and institutional requirements.
FM: Conceptualization, Formal analysis, Investigation, Methodology, Writing – original draft. AS: Conceptualization, Data curation, Formal analysis, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. MS: Data curation, Formal analysis, Methodology, Software, Validation, Visualization, Writing – original draft. MA: Formal analysis, Investigation, Software, Validation, Writing – original draft. HB: Conceptualization, Data curation, Validation, Visualization, Writing – original draft. MGA: Data curation, Methodology, Writing – original draft. MB: Conceptualization, Resources, Writing – review & editing. RM: Funding acquisition, Writing – review & editing.
The authors declare that financial support was received for the research, authorship, and/or publication of this article. This research was funded by the National Research Program for Universities, Higher Education Commission of Pakistan under project no. 13049 titled “Substituting traditional protein source in poultry diet with low cost and more sustainable insect protein” and by a Lithuanian state grant through the Nature Research Centre, program 2 Climate and Eco-systems, Vilnius, Lithuania, available to RM.
We thank the Department of Animal Nutrition and Poultry Production and Avian Research and Training Centre UVAS for assisting in this study.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Adeniji A. (2007). Effect of replacing groundnut cake with maggot meal in the diet of broilers. Int. J. Poult. Sci. 6, 822–825. doi: 10.3923/ijps.2007.822.825
Adetunmbi T. (2023). The potential of insects as alternative animal protein source for livestock feeding. Glob. J. Agric. Sci. 22, 47–61. doi: 10.4314/gjass.v22i1.6
Adli D. N. (2021). Use of insects in poultry feed as replacement soya bean meal and fish meal in development countries: a systematic review. Livest. Res. Rur. Dev. 33, 128–131. Available at: http://www.lrrd.org/lrrd33/10/33128danun.html (Accessed January 24, 2025).
Akhtar Y., Isman M. B. (2018). “Insects as an alternative protein source,” in Proteins in food processing (Sawston, United Kingdom: Elsevier), 263–288. doi: 10.1016/B978-0-08-100722-8.00011-5
Alagawany M., Ashour E. A., El-Kholy M. S., Abou-Kassem D. E., Roshdy T., Abd El-Hack M. E. (2022). Consequences of varying dietary crude protein and metabolizable energy levels on growth performance, carcass characteristics and biochemical parameters of growing geese. Anim. Biotechnol. 33, 638–646. doi: 10.1080/10495398.2020.1814791
Amer S. A., Beheiry R. R., Abdel Fattah D. M., Roushdy E. M., Hassan F. A., Ismail T. A., et al. (2021). Effects of different feeding regimens with protease supplementation on growth, amino acid digestibility, economic efficiency, blood biochemical parameters, and intestinal histology in broiler chickens. BMC Vet. Res. 17, 1–16. doi: 10.1186/s12917-021-02946-2
Ballitoc D. A., Sun S. (2013). Ground yellow mealworms (Tenebrio molitor L.) feed supplementation improves growth performance and carcass yield characteristics in broilers. Open Sci. Repository Agric. 18, e23050425. doi: 10.7392/OPENACCESS.23050425
Benzertiha A., Kierończyk B., Kołodziejski P., Pruszyńska–Oszmałek E., Rawski M., Józefiak D., et al. (2020). Tenebrio molitor and Zophobas morio full-fat meals as functional feed additives affect broiler chickens’ growth performance and immune system traits. Poult. Sci. 99, 196–206. doi: 10.3382/ps/pez450
Benzertiha A., Kierończyk B., Rawski M., Józefiak A., Kozłowski K., Jankowski J., et al. (2019). Tenebrio molitor and Zophobas morio full-fat meals in broiler chicken diets: Effects on nutrients digestibility, digestive enzyme activities, and cecal microbiome. Animals 9, 1128–1150. doi: 10.3390/ani9121128
Biasato I., De Marco M., Rotolo L., Renna M., Lussiana C., Dabbou S., et al. (2016). Effects of dietary Tenebrio molitor meal inclusion in free-range chickens. J. Anim. Physiol. Anim. Nutr. 100, 1104–1112. doi: 10.1111/jpn.12487
Biasato I., Gasco L., De Marco M., Renna M., Rotolo L., Dabbou S., et al. (2017). Effects of yellow mealworm larvae (Tenebrio molitor) inclusion in diets for female broiler chickens: implications for animal health and gut histology. Anim. Feed Sci. Technol. 234, 253–263. doi: 10.1016/j.anifeedsci.2017.09.014
Biasato I., Gasco L., De Marco M., Renna M., Rotolo L., Dabbou S., et al. (2018). Yellow mealworm larvae (Tenebrio molitor) inclusion in diets for male broiler chickens: effects on growth performance, gut morphology, and histological findings. Poult. Sci. 97, 540–548. doi: 10.3382/ps/pex308
Bovera F., Loponte R., Marono S., Piccolo G., Parisi G., Iaconisi V., et al. (2016). Use of Tenebrio molitor larvae meal as protein source in broiler diet: Effect on growth performance, nutrient digestibility, and carcass and meat traits. J. Anim. Sci. 94, 639–647. doi: 10.2527/jas2015-9201
Bovera F., Loponte R., Pero M. E., Cutrignelli M. I., Calabrò S., Musco N., et al. (2018). Laying performance, blood profiles, nutrient digestibility and inner organs traits of hens fed an insect meal from Hermetia illucens larvae. Res. Vet. Sci. 120, 86–93. doi: 10.1016/j.rvsc.2018.09.006
Bovera F., Piccolo G., Gasco L., Marono S., Loponte R., Vassalotti G., et al. (2015). Yellow mealworm larvae (Tenebrio molitor, L.) as a possible alternative to soybean meal in broiler diets. Br. Poult. Sci. 56, 569–575. doi: 10.1080/00071668.2015.1080815
Campbell T. W. (1995). Avian hematology and cytology (Ames, Iowa, United States: Iowa State University Press).
Choi J., Kong B., Bowker B. C., Zhuang H., Kim W. K. (2023). Nutritional strategies to improve meat quality and composition in the challenging conditions of broiler production: a review. Animals 13, 1386. doi: 10.3390/ani13081386
Chu X., Li M., Wang G., Wang K., Shang R., Wang Z., et al. (2020). Evaluation of the low inclusion of full-fatted Hermetia illucens larvae meal for layer chickens: Growth performance, nutrient digestibility, and gut health. Front. Vet. Sci. 7. doi: 10.3389/fvets.2020.585843
Cullere M., Woods M. J., Van Emmenes L., Pieterse E., Hoffman L. C., Dalle Zotte A. (2019). Hermetia illucens larvae reared on different substrates in broiler quail diets: effect on physicochemical and sensory quality of the quail meat. Animals 9, 525–543. doi: 10.3390/ani9080525
Dabbou S., Gai F., Biasato I., Capucchio M. T., Biasibetti E., Dezzutto D., et al. (2018). Black soldier fly defatted meal as a dietary protein source for broiler chickens: Effects on growth performance, blood traits, gut morphology and histological features. J. Anim. Sci. Biotechnol. 9, 1–10. doi: 10.1186/s4010401802669
Dabbou S., Lauwaerts A., Ferrocino I., Biasato I., Sirri F., Zampiga M., et al. (2021). Modified black soldier fly larva fat in broiler diet: Effects on performance, carcass traits, blood parameters, histomorphological features and gut microbiota. Animals 11, 1837. doi: 10.3390/ani11061837
Dahiru S., Azhar B., Anjas B., Asmara B. (2016). Performance of spring chicken fed different inclusion levels of black soldier fly larvae meal. Entomol. Ornithol. Herpetol. 5, 185–189. doi: 10.4172/2161-0983.1000185
Danieli P. P., Lussiana C., Gasco L., Amici A., Ronchi B. (2019). The effects of diet formulation on the yield, proximate composition, and fatty acid profile of the black soldier fly (Hermetia illucens L.) prepupae intended for animal feed. Animals 9, 178. doi: 10.3390/ani9040178
da-Silva W. C., da Silva ÉB.R., da Silva J. A. R., Martorano L. G., Belo T. S., Sousa C. E. L., et al. (2024). Nutritional Value of the Larvae of the Black Soldier Fly (Hermetia illucens) and the House Fly (Musca domestica) as a Food Alternative for Farm Animals—A Systematic Review. Insects 15, 619. doi: 10.3390/insects15080619
Davison C., Michie C., Tachtatzis C., Andonovic I., Bowen J., Duthie C.-A. (2023). Feed conversion ratio (FCR) and performance group estimation based on predicted feed intake for the optimisation of beef production. Sensors 23, 4621. doi: 10.3390/s23104621
De Souza Vilela J., Andronicos N. M., Kolakshyapati M., Hilliar M., Sibanda T. Z., Andrew N. R., et al. (2021). Black soldier fly larvae in broiler diets improve broiler performance and modulate the immune system. Anim. Nutr. 7, 695–706. doi: 10.1016/j.aninu.2020.08.014
Elliott S. (2008). Erythropoiesis-stimulating agents and other methods to enhance oxygen transport. Br. J. Pharmacol. 154, 529–541. doi: 10.1038/bjp.2008.89
Ferreira T., Rasband W. (2012). ImageJ user guide—IJ 1.46. Available online at: http://imagej.nih.gov/ij/docs/guide (Accessed 26 September 2023). imagej. nih.
Fisher H., Collins S., Hanson C., Mason B., Colombo S., Anderson D. (2020). Black soldier fly larvae meal as a protein source in low fish meal diets for Atlantic salmon (Salmo salar). Aquaculture 521, 734978. doi: 10.1016/j.aquaculture.2020.734978
Franco J. R. G., Murakami A. E., Natali M. R. M., Garcia E., Furlan A. C. (2006). Influence of delayed placement and dietary lysine levels on small intestine morphometrics and performance of broilers. Braz. J. Poult. Sci. 8, 233–241. doi: 10.1590/S1516-635X2006000400006
García-Vaquero M., García C. Á. (2024). “Nutritional value of insects and derived ingredients,” in Insects as food and food ingredients (Dublin, Ireland: Elsevier), 31–45. doi: 10.1016/B9780323955942000094
Gariglio M., Dabbou S., Crispo M., Biasato I., Gai F., Gasco L., et al. (2019). Effects of the dietary inclusion of partially defatted black soldier fly (Hermetia illucens) meal on the blood chemistry and tissue (Spleen, Liver, Thymus, and Bursa of Fabricius) histology of muscovy ducks (Cairina moschata domestica). Animals 9, 307. doi: 10.3390/ani9060307
Gasco L., Biancarosa I., Liland N. S. (2020). From waste to feed: A review of recent knowledge on insects as producers of protein and fat for animal feeds. Curr. Opin. Green Sustain. Chem. 23, 67–79. doi: 10.1016/j.cogsc.2020.03.003
Halloran A., Hanboonsong Y., Roos N., Bruun S. (2017). Life cycle assessment of cricket farming in north-eastern Thailand. J. Clean. Prod. 156, 83–94. doi: 10.1016/j.jclepro.2017.04.017
Han X., Heinonen M. (2022). Processing improves physical and oxidative stability of cricket protein emulsions. Food Chem. Adv. 1, 100125. doi: 10.1016/j.focha.2022.100125
Hartinger K., Greinix J., Thaler N., Ebbing M. A., Yacoubi N., Schedle K., et al. (2021). Effect of graded substitution of soybean meal by Hermetia illucens larvae meal on animal performance, apparent ileal digestibility, gut histology and microbial metabolites of broilers. Animals 11, 1628–1644. doi: 10.3390/ani11061628
Hasnan F. F. B., Feng Y., Sun T., Parraga K., Schwarz M., Zarei M. (2023). Insects as valuable sources of protein and peptides: production, functional properties, and challenges. Foods 12, 4243. doi: 10.3390/foods12234243
Henry M., Gasco L., Piccolo G., Fountoulaki E. (2015). Review on the use of insects in the diet of farmed fish: past and future. Anim. Feed Sci. Technol. 203, 1–22. doi: 10.1016/j.anifeedsci.2015.03.001
Hobbi P., Bekhit A. E.-D. A., Debaste F., Lei N., Shavandi A. (2022). “Insect-derived protein as food and feed,” in Alternative proteins (Boca Raton, Florida: CRC Press), 85–132.
Hossain S., Blair R. (2007). Chitin utilisation by broilers and its effect on body composition and blood metabolites. Br. Poult. Sci. 48, 33–38. doi: 10.1080/00071660601156529
Hwangbo J., Hong E., Jang A., Kang H., Oh J., Kim B., et al. (2009). Utilization of house fly-maggots, a feed supplement in the production of broiler chickens. J. Environ. Biol. 30, 609–614.
Ijaiya A., Eko E. (2009). Effect of replacing dietary fish meal with silkworm (Anaphe infracta) caterpillar meal on growth, digestibility and economics of production of starter broiler chickens. Pak. J. Nutr. 8, 845–849. doi: 10.3923/pjn.2009.845.849
Islam M., Hosain N., Islam A. T. M. F. (2022) in 22nd international biennial conference and AGM 2021, Dhaka, Bangladesh. Muhsina Yasmin, Bangladesh, 18 March 2022, Vol. 1. 23–33.
Kaić A., Janječić Z., Žanetić A., Kelava Ugarković N., Potočnik K. (2021). EZ-DripLoss assessment in chicken breast meat using different sample areas, fiber orientation, and measurement intervals. Animals 11, 1095. doi: 10.3390/ani11041095
Khambualai O., Yamauchi K.-e., Tangtaweewipat S., Cheva-Isarakul B. (2008). Effects of dietary chitosan diets on growth performance in broiler chickens. J. Poult. Sci. 45, 206–209. doi: 10.2141/jpsa.45.206
Khan S., Khan R., Alam W., Sultan A. (2018). Evaluating the nutritive profile of three insect meals and their effects to replace soya bean in broiler diet. J. Anim. Physiol. Anim. Nutr. 102, e662–e668. doi: 10.1111/jpn.12809
Khatun R., Howlider M., Rahman M., Hasanuzzaman M., Rahman M. (2003). Replacement of fish meal by silkworm pupae in broiler diets. Pak. J. Biol. Sci. 6, 955–958. Available at: https://docsdrive.com/pdfs/ansinet/pjbs/2003/955-958.pdf (Accessed January 24, 2025).
Kierończyk B., Rawski M., Józefiak A., Mazurkiewicz J., Świątkiewicz S., Siwek M., et al. (2018). Effects of replacing soybean oil with selected insect fats on broilers. Anim. Feed Sci. Technol. 240, 170–183. doi: 10.1016/j.anifeedsci.2018.04.002
Kim S., Chung T., Park H., Shin M., Park I., Choi I. (2019). Effects of diet composition on growth performance and feed conversion efficiency in Alphitobius diaperinus larvae. J. Entomol. Acarol. Res. 51, 33–37. doi: 10.4081/jear.2019.7761
Kim Y. B., Kim D.-H., Jeong S.-B., Lee J.-W., Kim T.-H., Lee H.-G., et al. (2020). Black soldier fly larvae oil as an alternative fat source in broiler nutrition. Poult. Sci. 99, 3133–3143. doi: 10.1016/j.psj.2020.01.018
Kulma M., Kouřimská L., Homolková D., Božik M., Plachý V., Vrabec V. (2020). Effect of developmental stage on the nutritional value of edible insects. A case study with Blaberus craniifer and Zophobas morio. J. Food Compos. Anal. 92, 103570–103578. doi: 10.1016/j.jfca.2020.103570
Kulma M., Kouřimská L., Plachý V., Božik M., Adámková A., Vrabec V. (2019). Effect of sex on the nutritional value of house cricket, Acheta domestica L. Food Chem. 272, 267–272. doi: 10.1016/j.foodchem.2018.08.049
Kuzmuk K. N., Swanson K. S., Tappenden K. A., Schook L. B., Fahey J. G.C. (2005). Diet and age affect intestinal morphology and large bowel fermentative end-product concentrations in senior and young adult dogs. J. Nutr. 135, 1940–1945. doi: 10.1093/jn/135.8.1940
Lalev M., Hristakieva P., Mincheva N., Oblakova M., Ivanova I. (2022). Insect meal as alternative protein ingredient in broiler feed. Bulg. J. Agric. Sci. 28, 743–751. doi: 10.5555/20220391894
Laudadio V., Passantino L., Perillo A., Lopresti G., Passantino A., Khan R., et al. (2012). Productive performance and histological features of intestinal mucosa of broiler chickens fed different dietary protein levels. Poult. Sci. 91, 265–270. doi: 10.3382/ps.2011-01675
Leiber F., Gelencsér T., Stamer A., Amsler Z., Wohlfahrt J., Früh B., et al. (2017). Insect and legume-based protein sources to replace soybean cake in an organic broiler diet: Effects on growth performance and physical meat quality. Renew. Agric. Food Syst. 32, 21–27. doi: 10.107/S1742170515000496
Loponte R., Nizza S., Bovera F., De Riu N., Fliegerova K., Lombardi P., et al. (2017). Growth performance, blood profiles and carcass traits of Barbary partridge (Alectoris barbara) fed two different insect larvae meals (Tenebrio molitor and Hermetia illucens). Res. Vet. Sci. 115, 183–188. doi: 10.1016/j.rvsc.2017.04.017
Lu S., Taethaisong N., Meethip W., Surakhunthod J., Sinpru B., Sroichak T., et al. (2022). Nutritional composition of black soldier fly larvae (Hermetia illucens L.) and its potential uses as alternative protein sources in animal diets: A review. Insects 13, 831. doi: 10.3390/insects13090831
Madau F. A., Arru B., Furesi R., Pulina P. (2020). Insect farming for feed and food production from a circular business model perspective. Sustainability 12, 5418. doi: 10.3390/su12135418
Makkar H. P., Tran G., Heuzé V., Ankers P. (2014). State-of-the-art on use of insects as animal feed. Anim. Feed Sci. Technol. 197, 1–33. doi: 10.1016/j.anifeedsci.2014.07.008
Marono S., Loponte R., Lombardi P., Vassalotti G., Pero M., Russo F., et al. (2017). Productive performance and blood profiles of laying hens fed Hermetia illucens larvae meal as total replacement of soybean meal from 24 to 45 weeks of age. Poult. Sci. 96, 1783–1790. doi: 10.3382/ps/pew461
McMichael A. J., Bambrick H. J. (2005). Meat consumption trends and health: Casting a wider risk assessment net. Public Health Nutr. 8, 341–343. doi: 10.1079/PHN2005742
Mir N. A., Rafiq A., Kumar F., Singh V., Shukla V. (2017). Determinants of broiler chicken meat quality and factors affecting them: a review. J. Food Sci. Technol. 54, 2997–3009. doi: 10.1007/s13197-017-2789-z
Mohan K., Rajan D. K., Ganesan A. R., Divya D., Johansen J., Zhang S. (2023). Chitin, chitosan and chitooligosaccharides as potential growth promoters and immunostimulants in aquaculture: A comprehensive review. Int. J. Biol. Macromol. 86, 126285. doi: 10.1016/j.ijbiomac.2023.126285
Murawska D., Daszkiewicz T., Sobotka W., Gesek M., Witkowska D., Matusevičius P., et al. (2021). Partial and total replacement of soybean meal with full-fat black soldier fly (Hermetia illucens L.) larvae meal in broiler chicken diets: impact on growth performance, carcass quality and meat quality. Animals 11, 2715. doi: 10.3390/ani11092715
Neumann C., Velten S., Liebert F. (2018). Improving the dietary protein quality by amino acid fortification with a high inclusion level of micro algae (Spirulina platensis) or insect meal (Hermetia illucens) in meat type chicken diets. Open J. Anim. Sci. 8, 12–26. doi: 10.4236/ojas.2018.81002
Ognik K., Kozłowski K., Stępniowska A., Listos P., Józefiak D., Zduńczyk Z., et al. (2020). Antioxidant status and liver function of young Turkeys receiving a diet with full-fat insect meal from Hermetia illucens. Animals 10, 1339. doi: 10.3390/ani10081339
Oonincx D. G., De Boer I. J. (2012). Environmental impact of the production of mealworms as a protein source for humans–a life cycle assessment. PloS One 7, e51145. doi: 10.1371/journal.pone.0051145
Oonincx D., Finke M. (2021). Nutritional value of insects and ways to manipulate their composition. J. Insects Food Feed 7, 639–659. doi: 10.3920/JIFF2020.0050
Oyegoke O., Akintola A., Fasoranti J. (2006). Dietary potentials of the edible larvae of Cirina forda (westwood) as a poultry feed. Afr. J. Biotechnol. 5, 1799–1802. doi: 10.5897/AJB06.189
Parrini S., Aquilani C., Pugliese C., Bozzi R., Sirtori F. (2023). Soybean replacement by alternative protein sources in pig nutrition and its effect on meat quality. Animals 13, 494. doi: 10.3390/ani13030494
Patyra E., Kwiatek K. (2023). Insect meals and insect antimicrobial peptides as an alternative for antibiotics and growth promoters in livestock production. Pathogens 12, 854. doi: 10.3390/pathogens12060854
Pieterse E., Erasmus S. W., Uushona T., Hoffman L. C. (2019). Black soldier fly (Hermetia illucens) pre-pupae meal as a dietary protein source for broiler production ensures a tasty chicken with standard meat quality for every pot. J. Sci. Food Agric. 99, 893–903. doi: 10.1002/jsfa.9261
Prakash A., Saxena V. K., Singh M. K. (2020). Genetic analysis of residual feed intake, feed conversion ratio and related growth parameters in broiler chicken: A review. World’s Poult. Sci. J. 76, 304–317. doi: 10.1080/00439339.2020.1735978
Priolo A., Micol D., Agabriel J., Prache S., Dransfield E. (2002). Effect of grass or concentrate feeding systems on lamb carcass and meat quality. Meat Sci. 62, 179–185. doi: 10.1016/S0309-1740(01)00244-3
Qaisrani S., Moquet P., Van Krimpen M., Kwakkel R., Verstegen M., Hendriks W. (2014). Protein source and dietary structure influence growth performance, gut morphology, and hindgut fermentation characteristics in broilers. Poult. Sci. 93, 3053–3064. doi: 10.3382/ps.2014-04091
Ramos-Elorduy J., González E. A., Hernández A. R., Pino J. M. (2002). Use of Tenebrio molitor (Coleoptera: Tenebrionidae) to recycle organic wastes and as feed for broiler chickens. J. Econ. Entomol. 95, 214–220. doi: 10.1603/0022-0493-95.1.214
Ravindran V., Abdollahi M. R. (2021). Nutrition and digestive physiology of the broiler chick: state of the art and outlook. Animals 11, 2795. doi: 10.3390/ani11102795
Ravindran V., Abdollahi M., Bootwalla S. (2014). Nutrient analysis, metabolizable energy, and digestible amino acids of soybean meals of different origins for broilers. Poult. Sci. 93, 2567–2577. doi: 10.3382/ps.2014-04068
Rehman A. U., Arif M., Husnain M. M., Alagawany M., Abd El-Hack M. E., Taha A. E., et al. (2019). Growth performance of broilers as influenced by different levels and sources of methionine plus cysteine. Animals 9, 1056. doi: 10.3390/ani9121056
Sajid Q. U. A., Asghar M. U., Tariq H., Wilk M., Płatek A. (2023). Insect meal as an alternative to protein concentrates in poultry nutrition with future perspectives (An updated review). Agriculture 13, 1239–1264. doi: 10.3390/agriculture13061239
Sajjad M., Binyameen M., Sajjad A., Sarmad M., Abbasi A., Khan E., et al. (2024a). Replacing soybean meal with lesser mealworm Alphitobius diaperinus improves broiler productive performances, haematology, intestinal morphology and meat quality. J. Insects Food Feed 1, 1–22. doi: 10.1163/23524588-00001190
Sajjad M., Sajjad A., Chishti G., Binyameen M., Abbasi A., Haq I., et al. (2024b). Evaluation of blow fly, Chrysomya megacephala (Calliphoridae: Diptera) as an alternate source of protein in broiler feed. J. Insects Food Feed 1, 1–19. doi: 10.1163/23524588-00001109
Sajjad M., Sajjad A., Chishti G., Khan E., Mozūraitis R., Binyameen M. (2024c). Insect larvae as an alternate protein source in poultry feed improve the performance and meat quality of broilers. Animals 14, 1–18. doi: 10.3390/ani14142053
Sánchez-Muros M.-J., Barroso F. G., Manzano-Agugliaro F. (2014). Insect meal as renewable source of food for animal feeding: a review. J. Clean Prod. 65, 16–27. doi: 10.1016/j.jclepro.2013.11.068
Schiavone A., Castillo A. (2024). Incorporating whole insect larvae into poultry diets: state of the art and future perspectives. Ital. J. Anim. Sci. 23, 1–14. doi: 10.1080/1828051X.2023.2283083
Schiavone A., Cullere M., De Marco M., Meneguz M., Biasato I., Bergagna S., et al. (2017). Partial or total replacement of soybean oil by black soldier fly larvae (Hermetia illucens L.) fat in broiler diets: Effect on growth performances, feed-choice, blood traits, carcass characteristics and meat quality. Ital. J. Anim. Sci. 16, 93–100. doi: 10.1080/1828051X.2016.1249968
Schiavone A., Dabbou S., Petracci M., Zampiga M., Sirri F., Biasato I., et al. (2019). Black soldier fly defatted meal as a dietary protein source for broiler chickens: Effects on carcass traits, breast meat quality and safety. Animal 13, 2397–2405. doi: 10.1017/S1751731119000685
Sedgh-Gooya S., Torki M., Darbemamieh M., Khamisabadi H., Abdolmohamadi A. (2021). Effect of dietary inclusion of yellow mealworm (Tenebrio molitor) larvae meal on productive performance, egg quality indices and blood parameters of laying hens. Anim. Prod. Sci. 61, 1365–1372. doi: 10.1071/AN20102
Siddiqui S. A., Elsheikh W., Ucak İ., Hasan M., Perlita Z. C., Yudhistira B. (2024). Replacement of soy by mealworms for livestock feed-A comparative review between soy and mealworms considering environmental aspects. Environ. Dev. Sustainability, 1–44. doi: 10.1007/s10668-024-04874-1
Slimen I. B., Yerou H., Larbi M. B., M’Hamdi N., Najar T. (2023). Insects as an alternative protein source for poultry nutrition: a review. Front. Vet. Sci. 10, 1–15. doi: 10.1071/AN20102
Sypniewski J., Kierończyk B., Benzertiha A., Mikołajczak Z., Pruszyńska-Oszmałek E., Kołodziejski P., et al. (2020). Replacement of soybean oil by Hermetia illucens fat in Turkey nutrition: effect on performance, digestibility, microbial community, immune and physiological status and final product quality. Br. Poult. Sci. 61, 294–302. doi: 10.1080/00071668.2020.1716302
Tan S. W., Lai K. S., Loh J. Y. (2018). Effects of food wastes on yellow mealworm Tenebrio molitor larval nutritional profiles and growth performances. Examines Mar. Biol. Oceanogr 2, 173–178. doi: 10.31031/EIMBO.2018.02.000530
Udomsil N., Imsoonthornruksa S., Gosalawit C., Ketudat-Cairns M. (2019). Nutritional values and functional properties of house cricket (Acheta domesticus) and field cricket (Gryllus bimaculatus). Food Sci. Technol. Res. 25, 597–605. doi: 10.3136/fstr.25.597
Van Harn J., Dijkslag M., Van Krimpen M. (2019). Effect of low protein diets supplemented with free amino acids on growth performance, slaughter yield, litter quality, and footpad lesions of male broilers. Poult. Sci. 98, 4868–4877. doi: 10.3382/ps/pez229
Van Huis A. (2013). Potential of insects as food and feed in assuring food security. Annu. Rev. Entomol. 58, 563–583. doi: 10.1146/annurev-ento-120811-153704
Van Huis A., Van Itterbeeck J., Klunder H., Mertens E., Halloran A., Muir G., et al. (2013). Edible insects: future prospects for food and feed security (Italy: Food and agriculture organization of the United Nations).
Van Peer M., Frooninckx L., Coudron C., Berrens S., Álvarez C., Deruytter D., et al. (2021). Valorisation potential of using organic side streams as feed for Tenebrio molitor, Acheta domesticus and Locusta migratoria. Insects 12, 796. doi: 10.3390/insects12090796
Vasilopoulos S., Giannenas I., Athanassiou G. ,. C., Rumbos C., Papadopoulos E., Fortomaris P. (2024a). Black soldier fly, mealworm and superworm: chemical composition and comparative effect on broiler growth. World’s Poult. Sci. J. 80, 681–710. doi: 10.1080/00439339.2024.2365919
Vasilopoulos S., Giannenas I., Mellidou I., Stylianaki I., Antonopoulou E., Tzora A., et al. (2024b). Diet replacement with whole insect larvae affects intestinal morphology and microbiota of broiler chickens. Sci. Rep. 14, 6836. doi: 10.1038/s41598-024-54184-9
Vasilopoulos S., Giannenas I., Savvidou S., Bonos E., Rumbos C. I., Papadopoulos E., et al. (2023). Growth performance, welfare traits and meat characteristics of broilers fed diets partly replaced with whole Tenebrio molitor larvae. Anim. Nutr. 13, 90–100. doi: 10.1016/j.aninu.2022.12.002
Veldkamp T., Bosch G. (2015). Insects: a protein-rich feed ingredient in pig and poultry diets. Anim. Front. 5, 45–50. doi: 10.2527/af.2015-0019
Veldkamp T., Dong L., Paul A., Govers C. (2022). Bioactive properties of insect products for monogastric animals–a review. J. Insects Food Feed 8, 1027–1040. doi: 10.3920/JIFF2021.0031
Vilela J. S., Alvarenga T. I., Andrew N. R., McPhee M., Kolakshyapati M., Hopkins D. L., et al. (2021). Technological quality, amino acid and fatty acid profile of broiler meat enhanced by dietary inclusion of black soldier fly larvae. Foods 10, 1–20. doi: 10.3390/foods10020297
Wang D., Zhai S. W., Zhang C. X., Bai Y. Y., An S. H., Xu Y. N. (2005). Evaluation on nutritional value of field crickets as a poultry feedstuff. Asian Australas. J. Anim. Sci. 18, 667–670. doi: 10.5713/ajas.2005.667
Yang C., Li X., Li Q., Zhang Z., Li W., Jiang X. (2011). Evaluation for meat quality performance of broiler chicken. J. Anim. Vet. Adv. 10, 949–954. doi: 10.3923/javaa.2011.949.954
Zadeh Z. S., Kheiri F., Faghani M. (2019). Use of yellow mealworm (Tenebrio molitor) as a protein source on growth performance, carcass traits, meat quality and intestinal morphology of Japanese quails (Coturnix japonica). Vet. Anim. Sci. 8, 100066. doi: 10.1016/j.vas.2019.100066
Zaid M., Hussain J., Mahmud A., Javed K., Shaheen M. S., Usman M., et al. (2020). Carcass traits, meat quality, and sensory attributes of fast-growing broilers givenoutdoor access at different ages. Turk. J. Vet. Anim. Sci. 44, 1039–1046. doi: 10.3906/vet-2003-92
Zhou Y., Wang D., Zhou S., Duan H., Guo J., Yan W. (2022). Nutritional composition, health benefits, and application value of edible insects: a review. Foods 11, 3961. doi: 10.3390/foods11243961
Keywords: Acheta domesticus, broiler, hematology, Hermetia illucens, productive performance, replacement
Citation: Mustafa F, Sajjad A, Sajjad M, Ali M, Bashir HS, Abbas MG, Binyameen M and Mozūratis R (2025) Comparative evaluation of Acheta domesticus and Hermetia illucens as alternative protein sources for the growth, health, and meat quality of the broiler. Front. Anim. Sci. 6:1531761. doi: 10.3389/fanim.2025.1531761
Received: 20 November 2024; Accepted: 09 January 2025;
Published: 04 February 2025.
Edited by:
Rayudika Aprilia Patindra Purba, Airlangga University, IndonesiaReviewed by:
Susanne Kreuzer-Redmer, University of Veterinary Medicine Vienna, AustriaCopyright © 2025 Mustafa, Sajjad, Sajjad, Ali, Bashir, Abbas, Binyameen and Mozūratis. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Asif Sajjad, asif.sajjad@iub.edu.pk; Raimondas Mozūratis, raimondas.mozuraitis@su.se
†ORCID: Muhammad Sajjad, orcid.org/0009-0008-8958-5848
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.