 Raquel J. Krach
Raquel J. Krach Cynthia A. Daley
Cynthia A. Daley Garrett C. Liles
Garrett C. Liles- Center for Regenerative Agriculture and Resilient Systems, College of Agriculture, California State University, Chico, Chico, CA, United States
This study assessed the impact of compost applications and multispecies cover crops on almond production and soil health with the working hypothesis that one or both practices would improve soil structure and fertility and therefore increase the overall productivity of the orchard. Treatments were applied to a 17-year-old organic almond orchard in a randomized complete block design for three consecutive years. Treatments included: (1) Fall-applied compost (COMP), (2) Fall-seeded multispecies cover crop (MSCC), (3) both compost and cover crop combined (COMP + MSCC), and (4) a non-treated control (CONT). Soil samples were collected in the fall of year one, before treatments were applied, and again in the fall of year three. Samples were analyzed for physical, chemical, and biological characteristics at the Center for Regenerative Agriculture & Resilient Systems’ (CRARS) Regenerative Agriculture Demonstration Lab and at a commercial soil lab. Almonds were harvested by row to determine yield within each plot and kernels were analyzed for nutrient content after the third growing season. Understory forage was collected in the spring of year three and analyzed for quality and dry matter production. After 3 years, soil in all treatment plots (COMP, MSCC, and COMP + MSCC) had higher quantities of soil organic matter (SOM), and therefore carbon, nitrogen, and other nutrients, as compared to CONT, with compost increasing SOM content more than the cover crop treatment (p < 0.05). Microbial biomass was higher in COMP + MSCC plots compared to CONT plots (p < 0.05) and microbial activity (soil respiration rate and % microbially active carbon) was higher in the COMP plots compared to CONT plots (p < 0.05). Understory forage biomass and nutritional quality were significantly higher in the MSCC and COMP + MSCC treatment plots compared with plots without cover crops (p < 0.05). This is an important benefit since orchard understory grazing with sheep is an important component of this farm’s normal operation. Finally, almond yields and kernel nutrient densities were significantly higher in treatment plots compared to control plots (p < 0.05). Collectively, these results demonstrate the multiple benefits of these regenerative/climate-smart practices in an organic orchard system.
1 Introduction
Agriculture must adopt climate-smart management practices to reduce the detrimental environmental impacts of modern farming (Intergovernmental Panel On Climate Change (IPCC), 2023) mitigate climate change through soil carbon sequestration (Paustian et al., 2016) and provide resiliency for our current food production system (Altieri et al., 2015; Lal, 2010; Li et al., 2019). While modern organic farming has been shown to improve water quality (Rad et al., 2022; Sivaranjani and Rakshit, 2019) and reduce the use of pesticides and other inorganic inputs, additional soil health management practices will be needed to slow the rapid loss of topsoil, conserve water, and mitigate climate change. In fact, a recent analysis with biogeochemical modeling determined that large-scale transition to organic production with “normative” practices would result in a net loss of soil organic carbon stocks (Gaudaré et al., 2023).
Evidence is accumulating that agricultural management practices that regenerate and maintain healthy soil improve agroecosystem function, provide more farm resiliency, and mitigate environmental problems including climate change (Blanco-Canqui and Francis, 2016; Li et al., 2019; Paustian et al., 2016). The primary mechanism for this is increasing soil organic carbon, and the practices that achieve this, sometimes referred to as “Regenerative or Climate Smart,” do so by adding organic matter to the system and preventing the organic matter already in the system from being lost. This is done by minimizing chemical and mechanical disturbance; growing more plants, either in time or space; importing organic matter in the form of organic amendments; increasing biological diversity; and integrating livestock.
Organic amendments such as compost and multispecies cover crops are two climate-smart practices amenable to orchard crop management. They are known to provide ecosystem services such as increased nutrient cycling, increased water use efficiency, decreased pest pressure, and increased biodiversity (Fenster et al., 2021; Khalsa et al., 2022; Lepsch et al., 2019; Wilson et al., 2022). This is critical, as almonds and other orchard crops are of high importance in California agriculture, both economically and environmentally, especially in the context of increasingly unpredictable weather and water availability.
Previous research has shown that compost may provide myriad benefits. Recent studies have found that, in general, organic matter amendments (OMA) improve soil physical, chemical and biological properties; increase crop nutrition, growth, and yield; and by increasing the soil organic matter, increase plant available nutrients and soil carbon sequestration (Diacono and Montemurro, 2011; Urra et al., 2019). While there has been less research in perennial crop systems compared to annual crops, these benefits have occurred in nectarines and citrus in Italy (Baldi et al., 2018; Canali et al., 2004), avocados in Spain (López et al., 2014), apples in Arkansas and the Pacific Northwest (Forge et al., 2013; Mays et al., 2014; Sharaf et al., 2021), vineyards in British Columbia (Hannam et al., 2016), and almonds in California and Spain (Hodson et al., 2021; Khalsa et al., 2022; Lepsch et al., 2019; Pérez-Murcia et al., 2021; Villa et al., 2021).
The use of compost in orchards and other perennial systems has been found to improve the soil physical characteristics such as compaction (Mujdeci, 2011) cation exchange capacity (Thompson and Peck, 2017; Villa et al., 2021), dissolved salts (Pérez-Murcia et al., 2021), bulk density (Mays et al., 2014), and aggregate stability (Khalsa and Brown, 2017; Peck et al., 2011). When applied in almond orchards, composted dairy manure was found to increase soil water retention which reduced tree water stress (Lepsch et al., 2019). Similarly, increased water infiltration and higher water use efficiency were achieved when compost was applied in peach production in Spain and apple orchards in New Zealand and New York (Goh et al., 2001; Lordan Sanahuja et al., 2015; Oliveira and Merwin, 2001).
Studies on compost in California almond orchards have found soil organic carbon (SOC) increased more with green waste compost compared to composted manure (Khalsa et al., 2022; Villa et al., 2021) and fall compost applications resulted in greater soil organic matter accumulation than spring compost applications (Lepsch et al., 2019). Composted food processing wastes and sheep manure applied in almond orchards in Spain led to a 25–50% increase in soil organic matter compared with plots receiving no amendments (Pérez-Murcia et al., 2021).
Compost application results in increased soil biological activity in orchard systems (Baldi et al., 2018; Forge et al., 2013; López et al., 2014; Neilsen et al., 2014; Pérez-Murcia et al., 2021; Sharaf et al., 2021). Higher microbial biomass carbon was associated with composted materials compared to non-composted materials such as raw manure, synthetic fertilizers, or no amendments (Baldi et al., 2018; Canali et al., 2004; Neilsen et al., 2014; Pérez-Murcia et al., 2021). Enzyme activity was found to be highest in the mulch treatment in avocados (López et al., 2014). When soil bacteria, fungi, and nematode abundances and diversity were evaluated, distinct communities were found to be associated with compost use compared to the use of synthetic fertilizer alone or no amendment in almond and apple orchard systems (Forge et al., 2013; Hodson et al., 2021; Neilsen et al., 2014; Peck et al., 2011; Sharaf et al., 2021; Thompson et al., 2019).
Finally, compost use has been found to have a positive or neutral effect on crop growth and production. In almonds its application led to faster young tree growth (Hodson et al., 2021), and higher soil and kernel nutrient content with no reduction in overall yield (Pérez-Murcia et al., 2021). Sharaf et al. (2021), Forge et al. (2013), and Thompson et al. (2019) found that OMA maintained yields compared to conventional production and/or led to increased above-ground apple tree growth. In other long-term studies, Nectarine yield was maintained [compared to mineral fertilizer (Baldi et al., 2018), and peach (Lordan Sanahuja et al., 2015) and avocado (López et al., 2014)] yields increased with OMA.
Cover crops are another practice shown to provide numerous benefits. They increase the health of the ecosystem and improve soil health by armoring the soil, increasing ecosystem primary productivity, and increasing biodiversity above and below ground. Planted between the tree rows, cover crops become an understory for the orchard systems like a natural system and provide vital ecosystem services for both the farmer and the planet (Blanco-Canqui and Francis, 2016; Blesh et al., 2019). The list of services provided by cover crops has been well documented: carbon sequestration; nutrient cycling; weed and pest suppression (Finney et al., 2017; Wilson et al., 2022); provision of habitat for pollinators and other beneficial animals (De Leijster et al., 2019); reduced erosion, thus reduced soil and nutrient loss (Martínez-Mena et al., 2020); and better water holding capacity as a result of improved soil structure (Blanco-Canqui and Francis, 2016; Blesh et al., 2019; Chapagain et al., 2020). There are financial benefits for the producer when the use of cover crops allows for the reduction in chemical inputs or provides additional income streams when grown for grazing, forage, seed, or straw while maintaining or increasing the primary crop’s yields (Chapagain et al., 2020; De Leijster et al., 2019; Fenster et al., 2021; McKenzie et al., 2017; Ramos et al., 2011). Because different cover crop species provide different benefits, such as biomass production, nitrogen fixation, reduction of soil compaction, weed, and pest suppression, and pollinator habitat a mixture of species from different functional groups may provide the most benefit (Blesh et al., 2019; Chapagain et al., 2020; Finney et al., 2017; Haring and Hanson, 2022; Moukanni et al., 2022).
Cover crops physically, chemically, and biologically alter soil structure. Their roots create macropores by penetrating the soil and micropores when soil particles are aggregated by the photosynthates they exude (Bacq-Labreuil et al., 2019; Haruna et al., 2018). This increased porosity results in less soil compaction and improved hydraulic conductivity and water-holding capacity (Blanco-Canqui and Francis, 2016). The soil aggregates are protected by cover crops when they intercept the direct impact of rain or irrigation water. The cover crop catches and holds on to the water, which also prevents the loss of sediment through erosion (Kaspar and Singer, 2011). The soil microbial community is fed by cover crop roots, residues, and exudates and as they decompose these substrates soil organic carbon and soil aggregation are increased (Moukanni et al., 2022).
Cover crops can facilitate carbon sequestration in the soil through increases in net primary productivity. This increased above- and below-ground biomass translates into more stored carbon in the soil organic matter (on average 12% more compared to no cover crop control) (McClelland et al., 2021; Poeplau and Don, 2015). Mixtures of cover crop species are more likely to result in greater primary productivity and biomass accumulation (Bainard et al., 2020; Isbell et al., 2017; Tilman et al., 1996). Just a handful of orchard studies have documented increases in soil organic carbon with the growth of cover crops (almond—De Leijster et al., 2019; Martínez-Mena et al., 2020; Özbolat et al., 2023; citrus—Castellano-Hinojosa and Strauss, 2020).
Cover crops enhance the soil microbiome when they create soil environments that provide more abundant and diverse resources to support soil biology (Muhammad et al., 2021). Certain types of microbes protect crops from disease and pests, facilitate the acquisition of nutrients, improve photosynthetic efficiency, and improve their tolerance for adverse environmental conditions such as drought and salinity (Fenster et al., 2021; Finney et al., 2017; Pommier, 2023). Studies of tree cropping systems with cover crops have found more nitrogen-fixing bacteria, ammonia-oxidizing bacteria, and archaea, as well as the genetic identification of enzymes needed for these processes (Castellano-Hinojosa and Strauss, 2020; De Leijster et al., 2019).
Finally, this study aims to look at the impact of stacking regenerative practices. While both organic amendments and cover crops have shown promise to benefit orchard systems, few studies have examined the effects of combining practices. The purpose of this research was to determine the impacts of applying compost and growing a multispecies cover crop in a mature organic almond orchard on productivity of the orchard and soil health as individual practices, and combined, in comparison to the farm’s standard management which included neither practice.
2 Materials and methods
2.1 Study site and system
The study system for this field experiment was the mature, 12.14 Ha, almond orchard, owned and farmed by Massa Organics, located adjacent to the Sacramento River in Glenn County, California (Figure 1). This certified organic orchard was established in 2005, planted with Nonpareil (50%), Price (25%), and Carmel (25%) varieties on Lovell peach rootstock. The trees are spaced 6.1 m apart, within and between rows and irrigated with solid set sprinklers. The orchard’s typical management practices are low disturbance, with no tillage, herbicides, pesticides or chemical fertilizers used in production. The farm’s flock of Dorper sheep are used to graze the orchard’s permanent vegetative ground cover and the almonds are harvested with above-ground equipment, i.e., nuts are shaken onto catching frames and placed in an adjacent field to dry. The soil is alluvial, type Wyo Silt Loam (Wn and Wsw), and while considered “well-drained” it belongs to the Hydrologic Group C, which is considered to have high susceptibility to erosion due to water runoff (NRCS, USDA, S.S.S., 2024). The climate at this location is characterized by hot, dry summers and cool, wet winters. The average precipitation is 22 in. (56 cm) per year (US Department of Commerce, n. d.), however, during the study (2019–2022) it averaged <9 in. (21 cm) per year (Figures 1, 2).
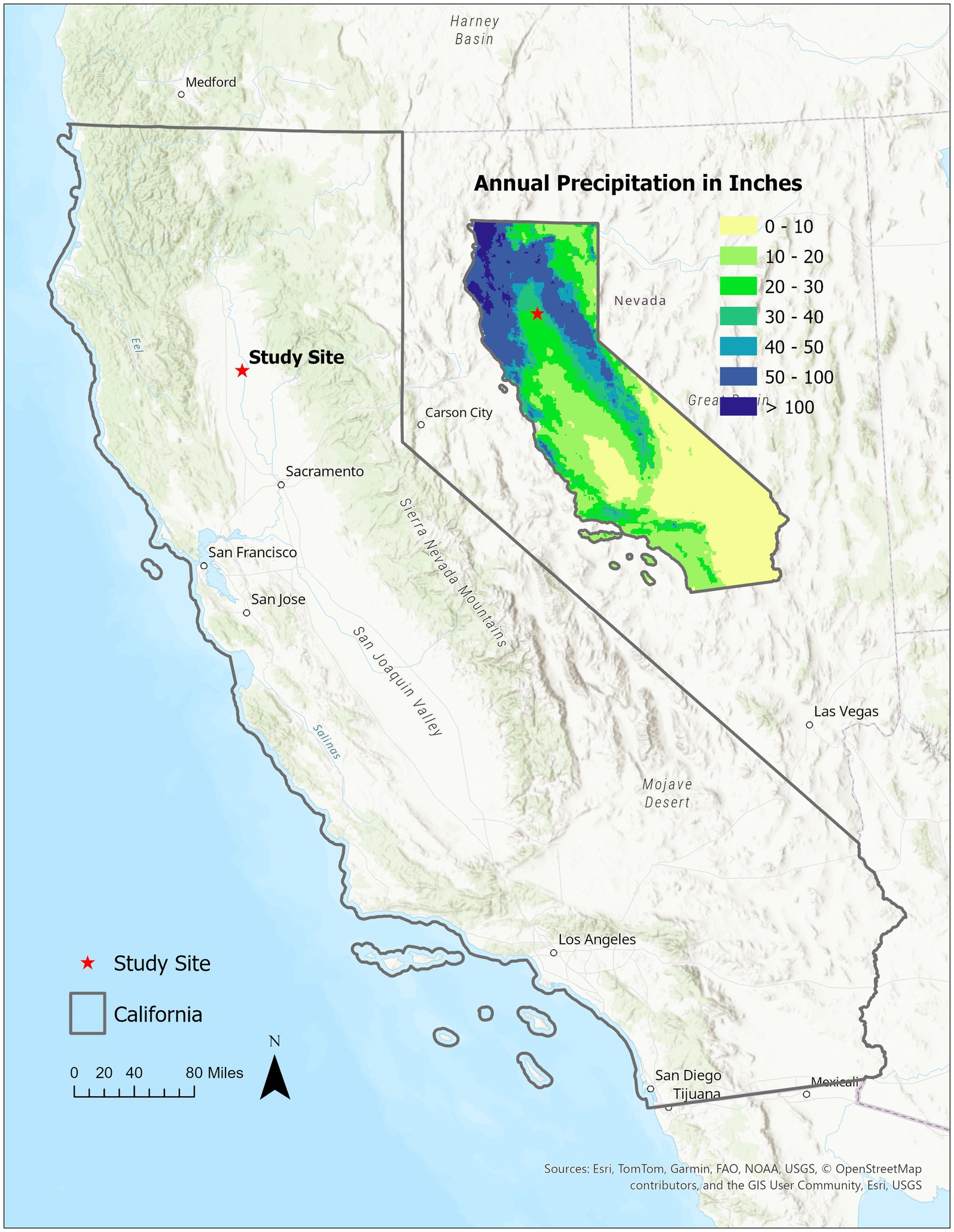
Figure 1. Map showing the location of the study site, the almond orchard of the Massa Organics Farm, adjacent to the Sacramento River in Glenn County of California; inset is map of average precipitation across the state.
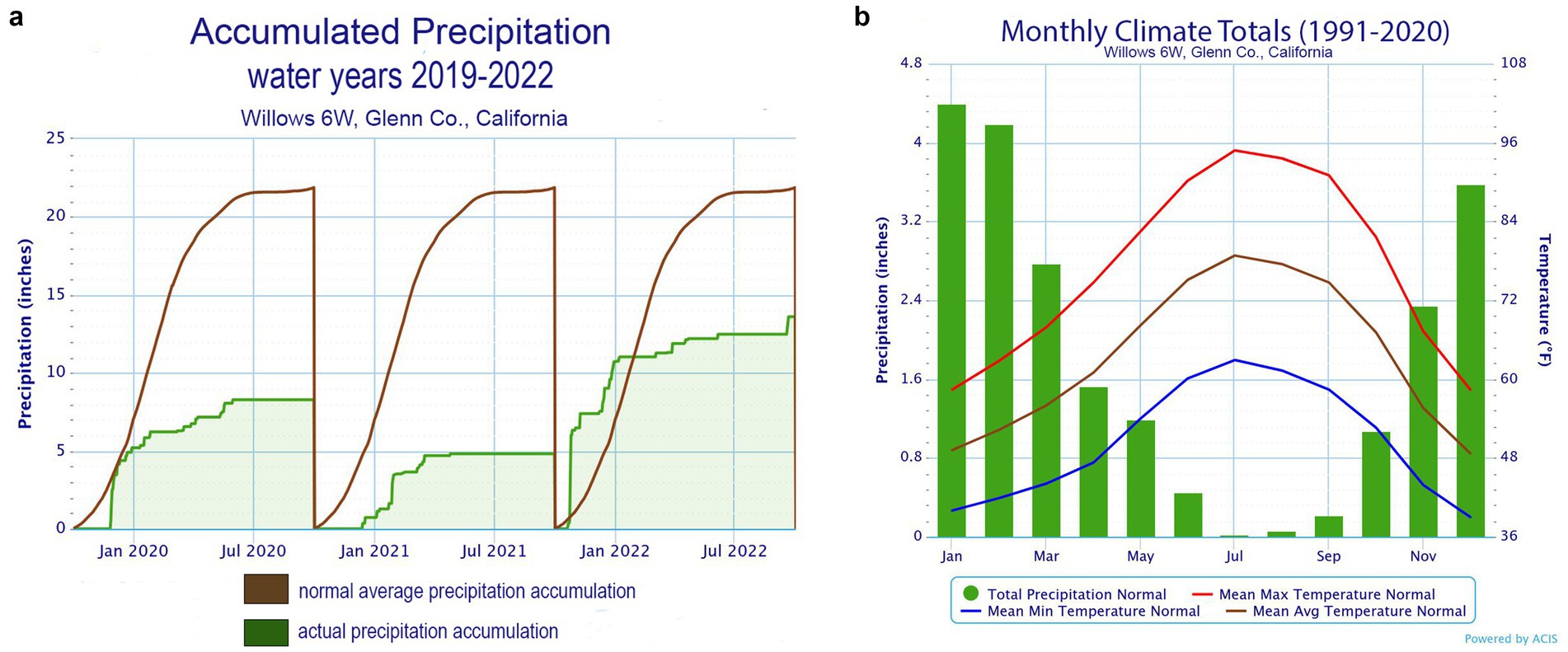
Figure 2. (A) Climate chart of accumulated precipitation, both normal and actual, over the three-year project period between initial implementation and 2022 almond harvest; (B) Normal average temperature and precipitation by month. Data is from the NOAA National Weather Service’ Willows 6 W station located approximately 10 miles from the study site (National Oceanic and Atmospheric Administration National Weather Service; https://www.weather.gov/wrh/Climate?wfo=sto accessed 2025-01-31).
2.2 Experimental design and treatments
The orchard was the focus of a California Department of Food and Agriculture Healthy Soils grant that cost-shared the climate-smart practices of growing a multispecies cover crop and applying compost from October 2019 to October 2022. To measure the impacts of these practices, individually and in combination, a 2 × 2 factorial, randomized, complete block experiment was established in a 5.7 Ha rectangular section of the orchard. The area was divided into 16 plots, each with four rows of 22–24 trees. Each plot had one of four treatments applied in October of each year: compost (COMP), multispecies cover crop (MSCC), compost plus multispecies cover crop (COMP + MSCC), and control (CONT), where neither treatment was applied (Figure 3). The compost was derived from green waste with a 16:1 C:N. It was broadcast, to include both the tree row and alleyways, at 18 metric tons per hectare. The cover crop, which included legumes, brassicas and grasses/cereals (Table 1), was planted between the tree rows using an eight-foot Great Plains No-Till drill. All plots were grazed by 75–100 sheep three times each year, February, late April/early May, and September. For each grazing event, temporary fencing was used to contain the sheep within a plot for 24 h. At the end of the 24 h, the sheep were moved to the next adjacent plot. The 24-h time frame was determined, from previous observation, to be the amount of time required for the sheep to graze approximately 50% of the forage. All plots were mechanically mowed in late July each year in preparation for August harvest.

Figure 3. Study site and experimental design. Glenn County, CA, USA. Aerial photo of almond orchard with overlay of plot map, color-coded according to treatment applied in randomized complete block design.
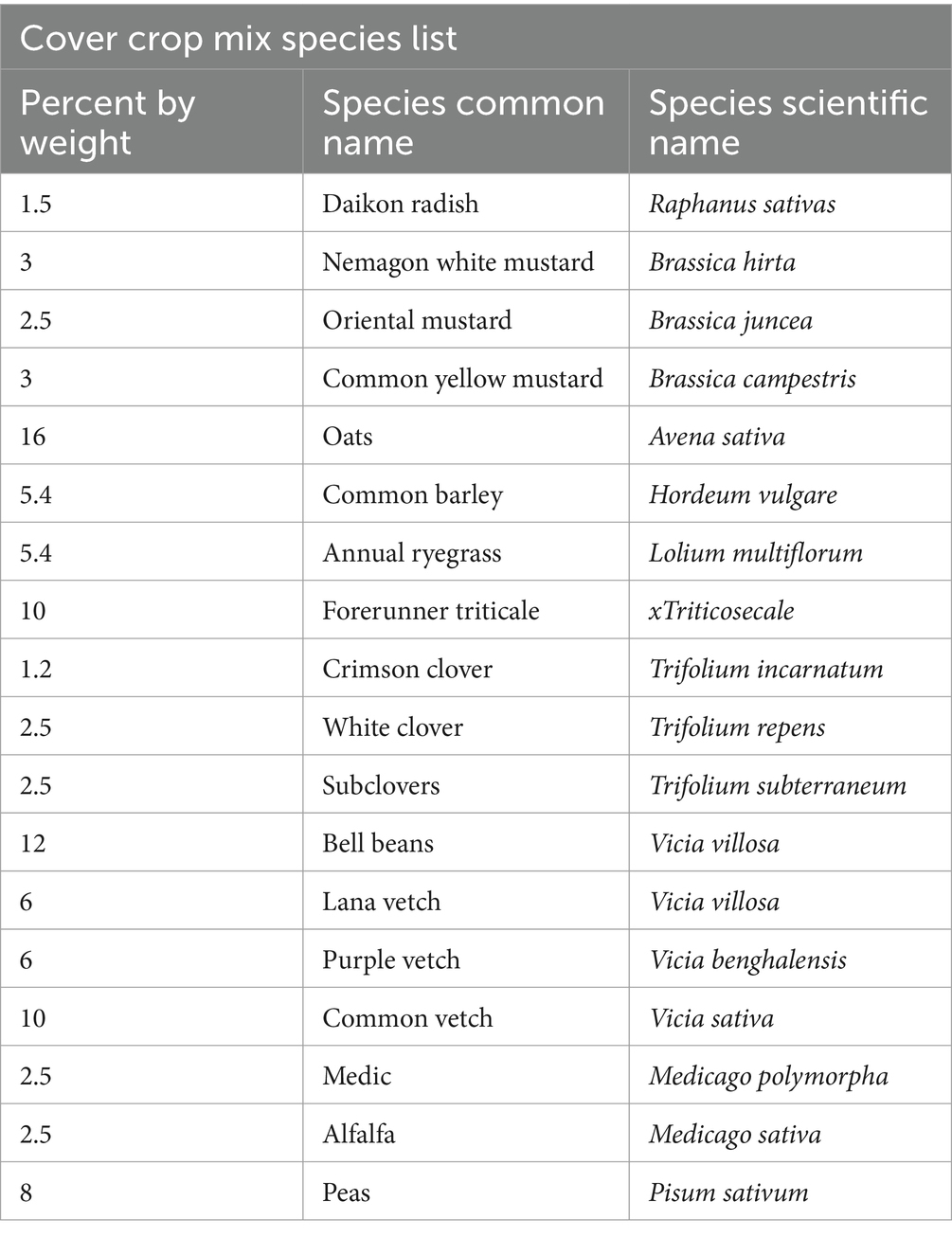
Table 1. Multispecies cover crop species list.
2.3 Data collection
2.3.1 Soil sampling
Orchard soil was collected at the end of the study in September 2022 after the treatments had been applied for 3 years. Composite samples were collected from two depths (0–15 and 15–30 cm) at three locations within each plot, between the two middle tree rows near the 6th, 12th, and 18th trees (Figure 4). A separate, 2-in soil core (8.25 cm3) was collected at each of the three sites to determine surface bulk density.
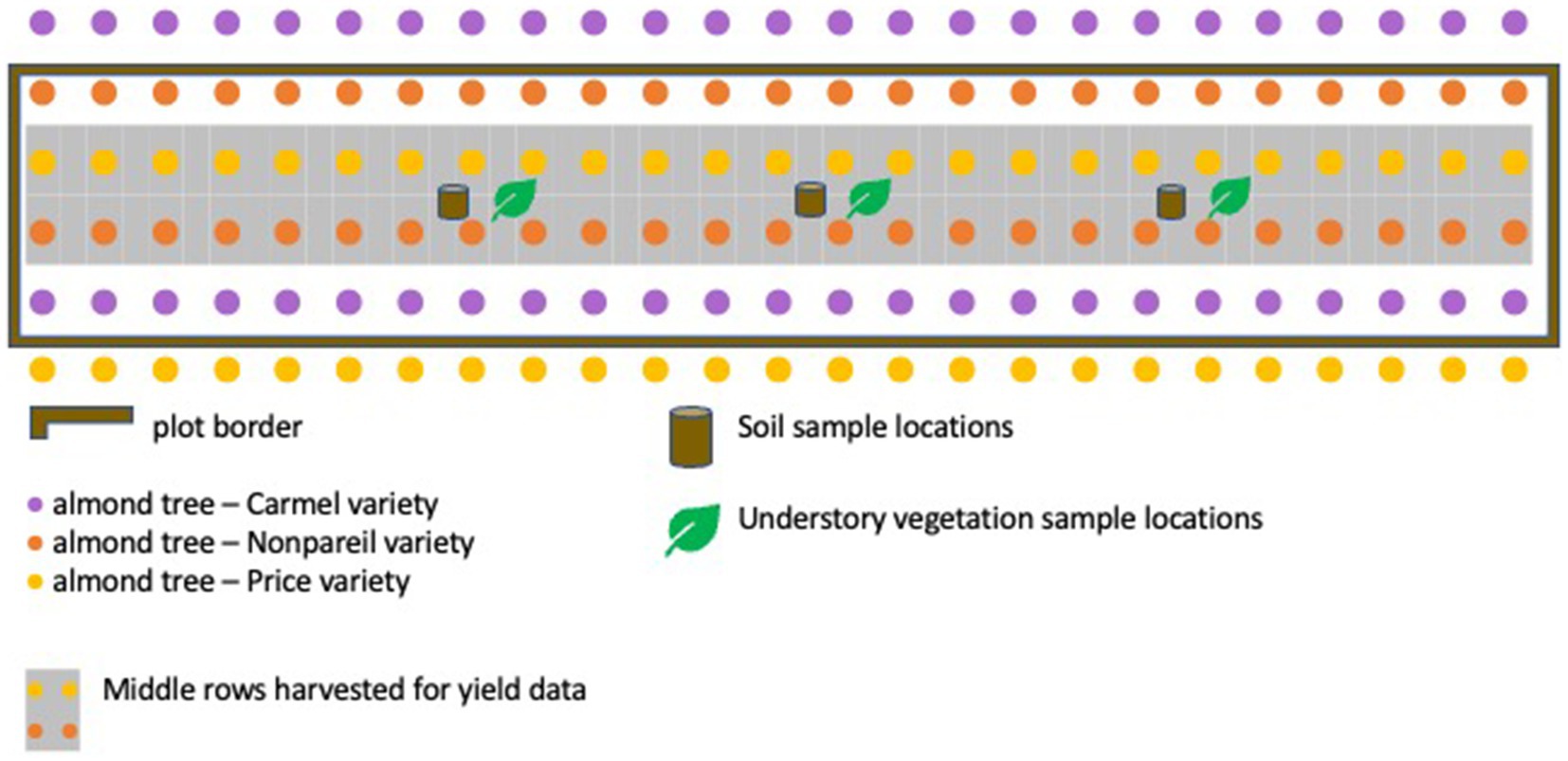
Figure 4. Schematic of a representative plot, including layout of almond trees and varieties; soil, and understory vegetation sampling.
Analyses of bulk density and wet aggregate stability were completed at the Chico State Regenerative Agriculture Demonstration Lab. To determine soil bulk density, samples were dried at 80°C and weighed to determine the mass per volume (NRCS, 2008). The percent of stable aggregates of soil from each depth was determined with a method adapted from Mikha and Rice (2004).
Further soil analyses were performed by Regen Ag Lab, LLC in Pleasanton, Nebraska, for the 0–15 cm samples, including pH, soluble salts, soil organic matter, total organic carbon, the Haney test and Phospholipid fatty acid (PLFA) analyses (Haney et al., 2006, 2008, 2018; NRCS, 2008; White, 1988; Zelles et al., 1992; Zelles and Bai, 1993). In addition to traditional chemical extraction methods to quantify soil Nitrogen and Phosphorus, the Haney soil test which uses extracts that mimic the natural soil environment: water that mimics the aqueous solution resulting from rain or irrigation and weak acid (H3A) that mimics the organic acids released from living plant roots. This test is thought to be a more accurate assessment of soil nutrients in plant-available form (Haney et al., 2018). The Haney test also measures carbon produced by respiration (CO2–C) and determines percent Microbial Active Carbon (MAC), defined as the amount of CO2–C relative to the amount of water-extractable organic carbon. Phospholipid Fatty Acid analysis determines the relative amounts of soil microbial phospholipid fatty acids as a snapshot of the quantity of microbes present in the soil (ng PLFA/g soil) (Zelles et al., 1992; Zelles and Bai, 1993).
2.3.2 Orchard understory productivity and quality analysis
To access orchard understory biomass production and forage quality, vegetation samples were collected from each plot in late April 2022. Total biomass production was determined according to the NRCS method of estimating cover crop biomass (NRCS, 2018). Vegetation within a 30 cm × 30 cm frame was cut at ground level at three sites within each plot and dried at 65°C until all moisture was removed. The combined dry weights of the three samples per plot were used to calculate dry matter per hectare for each plot.
Separate samples were collected for forage quality analysis according to the directions from the Dairy One commercial forage testing laboratory in Ithaca, New York. A composite sample of vegetation was collected from each plot by clipping vegetation at grazing height (approximately 15 cm) every 2 m along a transect from tree #6 to tree #12 between the two middle tree rows. The vegetation was chopped with clippers into small pieces, mixed thoroughly, and then a 0.45 kg sample was packaged, frozen for 12 h, and shipped for analysis at Dairy One. To assess the nutritional value of the vegetation as forage, components (Table 2) of the vegetation were analyzed using Near Infrared Reflectance Spectroscopy (NIRS) with the Foss NIRSystems Models XDS and 6,500 with ISIScan v.4.6.12 (Foss North America, 6,509 Flying Cloud Drive, Suite 130, Eden Praire, MN 55344)1 (FOSS analytical solutions for food quality improvement and control, n.d.).
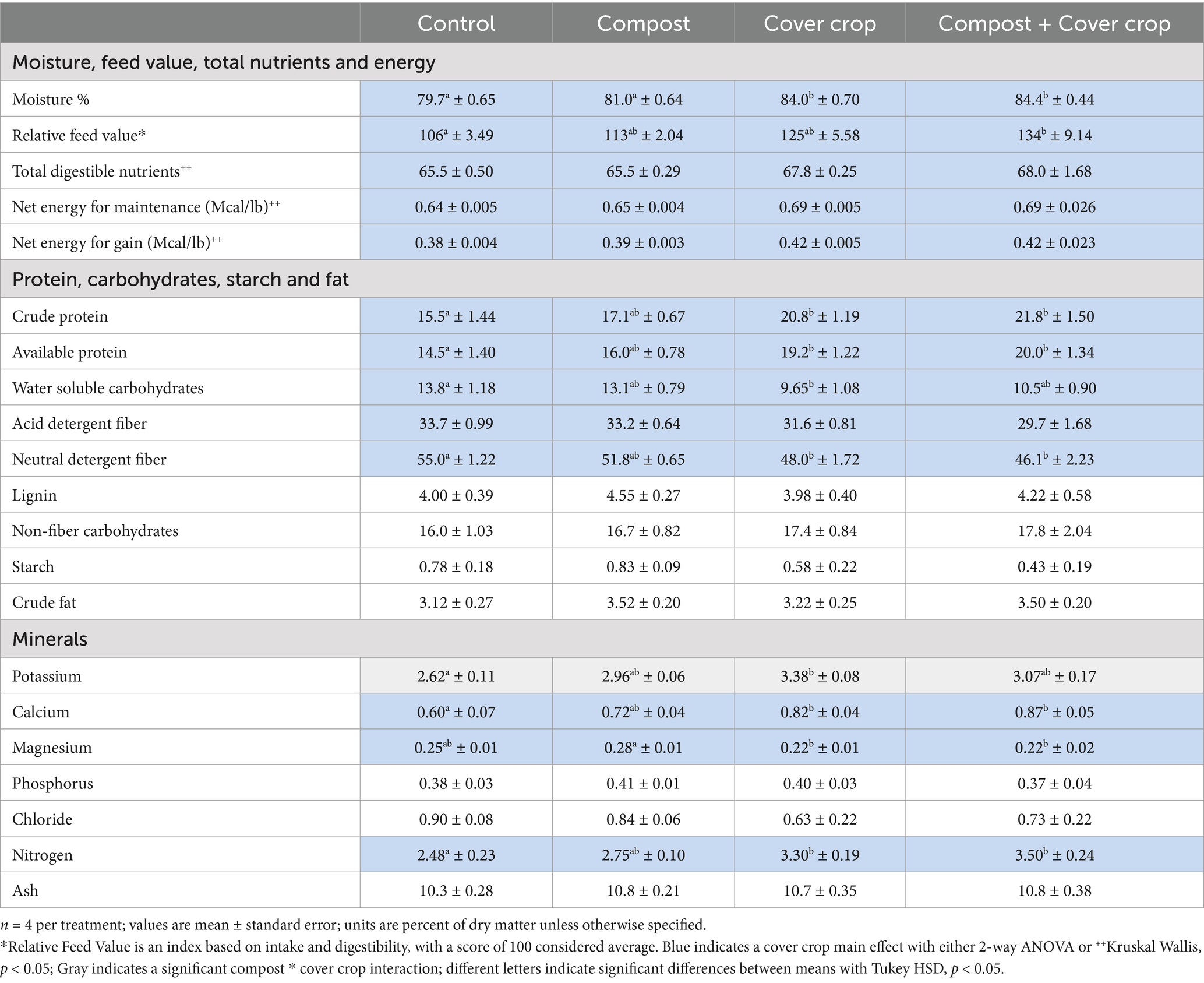
Table 2. Forage quality by treatment.
2.3.3 Almond crop yield
Almonds were harvested using catch-frame harvesting equipment and yields determined in August 2020, 2021, and 2022. The two middle rows of each plot, one row of Nonpareil and one row of Price, were harvested individually, dried, and weighed to the nearest pound (0.45 kg) using a standard weigh cart (Figure 4). The total material weighed included nut kernels, hulls and shells, and the total weight was divided by the number of trees in the row for a yield value in mass per tree.
2.3.4 Almond kernel collection and nutrient analysis
Composite samples of approximately 50 almond kernels of the Price variety were collected from each plot during harvest 2022. The kernels were analyzed for nutrient content (Ca, Mg, K, P, N, Zn, Cu, Fe, Mn, B) by Fruit Growers Laboratory in Santa Paula, California. They were treated as “whole fruit” botanical samples and analyzed primarily with a dry ash method, except for total nitrogen which was determined with an automated combustion method (Miller et al., 2013).
2.4 Statistical analysis
Statistical analyses were performed in R (Fox and Weisberg, 2018; Kassambara, 2023; R: The R Project for Statistical Computing, n.d.; Wickham et al., 2023). Comparisons of treatments were made using analysis of variance after checking for assumptions of normality of residuals (Shapiro–Wilk test) and homogeneity of variances (Levene test). Because it was a 2 × 2 factorial design, a two-way ANOVA was first performed to determine if there was an interaction effect between cover crop and compost. When no interaction was found, one-way ANOVAs were performed to determine if there were compost or cover crop main effects. Additional ANOVAs were run followed by a Tukey HSD test to determine if there were significant differences between treatment combinations. When the data did not meet the assumptions, data were transformed and retested. If the assumptions could still not be met, the Kruskal-Wallis test followed by pairwise comparisons using Wilcoxon rank sum exact test were substituted for the ANOVAs. Variables for which the data did not meet normality of residuals and homogeneity of variance assumptions are noted within the results tables.
3 Results
3.1 Soil health
3.1.1 Soil physical properties
Measures of soil structure, pH, and soluble salts are found in Table 3. While soil bulk densities did not differ significantly among treatments, there were differences in wet aggregate stability. The two-factor analysis for compost and cover crop showed a significant compost main effect for wet aggregate stability in the 0–15 cm soil samples (p < 0.05; Table 3). Plots that had compost applied (COMP & COMP + MSCC) had on average, a 9% higher percentage of stable aggregates compared to soils that did not receive compost (MSCC & CONT). There were no significant main or treatment effects for aggregate stability at the 15–30 cm soil depth, yet, the soil soluble salt concentration showed a significant interaction between compost and cover crop, with cover crop reducing the impact of compost on soluble salt concentration (p < 0.05; Table 3). The one-way analysis determined that soluble salts were significantly higher (43% higher) for COMP compared to CONT (p < 0.05; Table 3).
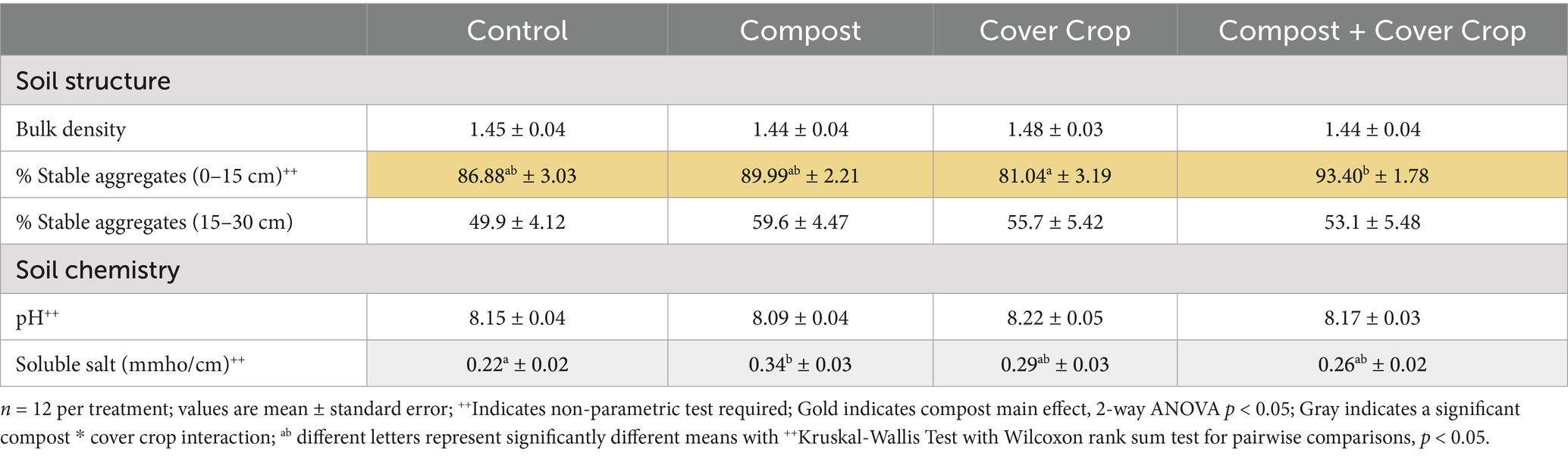
Table 3. Soil physical properties.
3.1.2 Soil organic matter and other carbon metrics
Results for soil organic matter (SOM) and other carbon (C) metrics are presented in Table 4. Compost and cover crop treatments showed (1) a significant compost main effect for SOM, percent total organic C (TOC), water extractable organic C and soil respiration (CO2–C), with up to 13, 19, 8 and 41% more, respectively, in plots with compost applied compared to those without compost; (2) a significant interaction effect between compost and cover crop for % Microbial Active Carbon (MAC); and (3) a significant cover crop main effect for C:N, where C:N was significantly lower, 4.6%, in plots with cover crops compared to plots without cover crops (p < 0.05; Table 4). The One-way analysis showed significantly higher values for COMP compared to CONT for SOM, TOC, CO2–C and MAC, and for C:N, both COMP and CONT were significantly greater than COMP + MSCC (p < 0.05; Table 4).
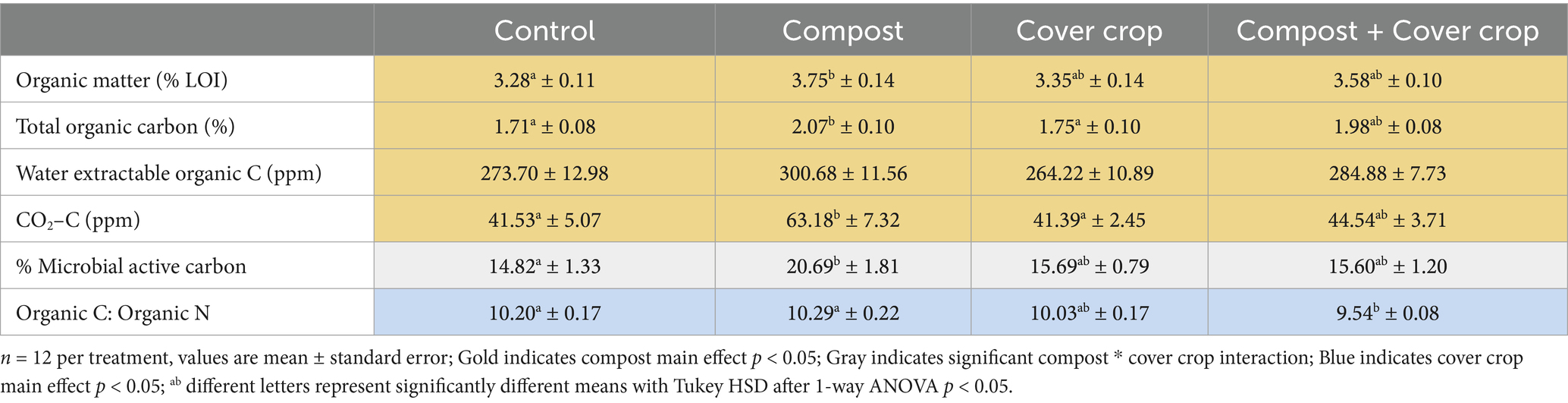
Table 4. Soil carbon metrics by treatment.
3.1.3 Soil nitrogen, phosphorus, and other macro- and micro-nutrients
Results for soil Nitrogen, Phosphorus, and other essential nutrients are presented in Tables 5, 6. For nitrogen (N), the measures included concentrations of both organic N and inorganic N, water and acid extracted N, the Haney test for available N, and the traditional test for nitrate-N−. Values for available N and traditional Nitrate-N are expressed in kg/ha to support the direct comparison of crop requirements and the quantification of the N dollar value in the soil (Figure 5). The two factor analysis for compost and cover crop showed a significant compost main effect for all measures except for the significant cover crop main effect for ammonium (p < 0.05; Tables 5, 6). There was more soil N in plots with compost applied compared to those without compost; 22, 60, 34, 60% more of Total N, Nitrate, Available N and Traditional N, respectively. There was 17% less in plots with cover crop growing compared to plots without cover crop. The one-way analysis showed: (1) water extractable total N was greater in COMP than CONT; (2) organic N Release and Available N was greater in COMP than both MSCC and CONT; (3) Nitrate, Inorganic N, and Traditional N were greater in COMP than both MSCC and CONT and COMP + MSCC was greater than both MSCC and CONT (p < 0.05; Tables 5, 6).
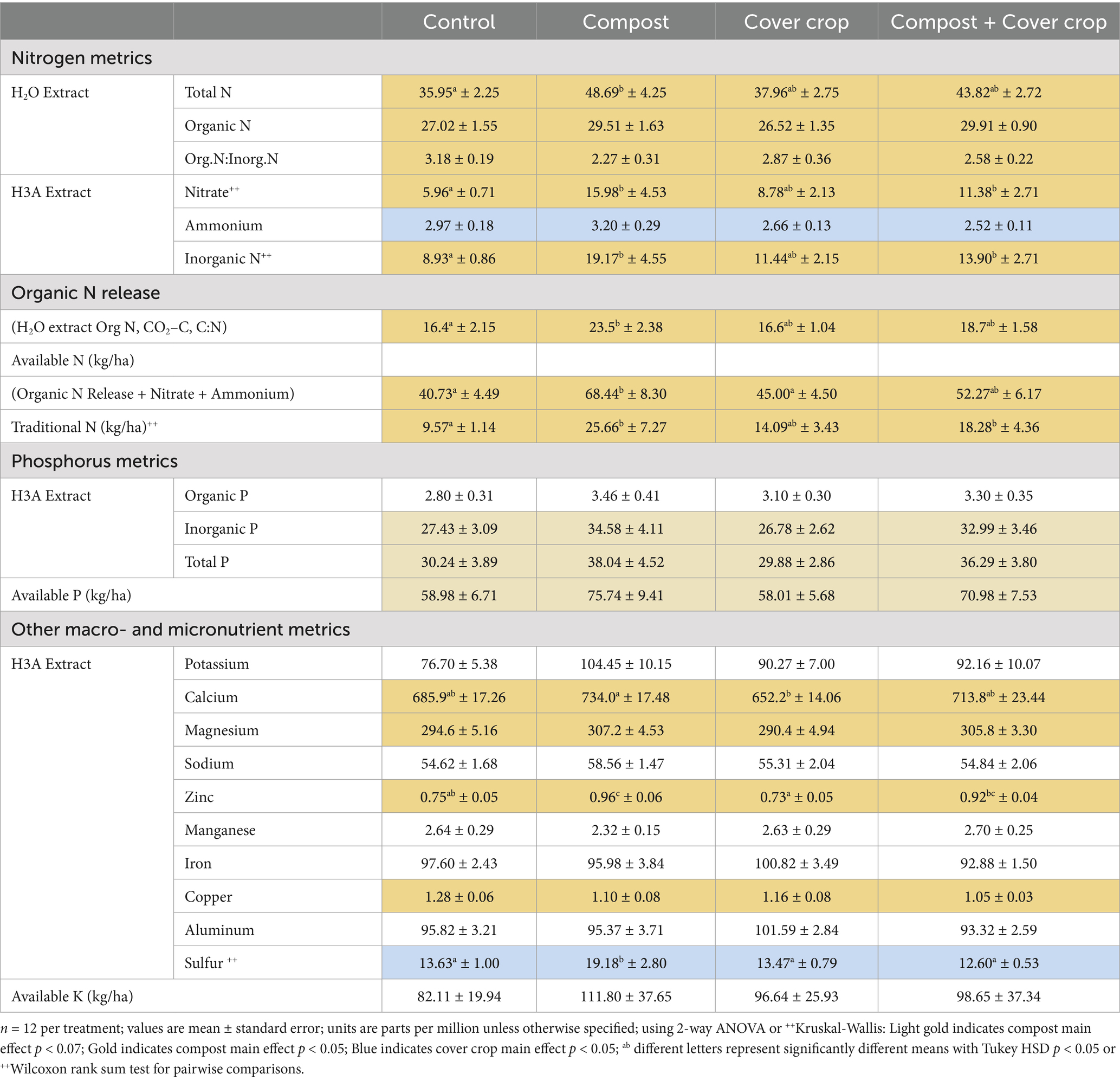
Table 5. Soil nutrient metrics by treatment.
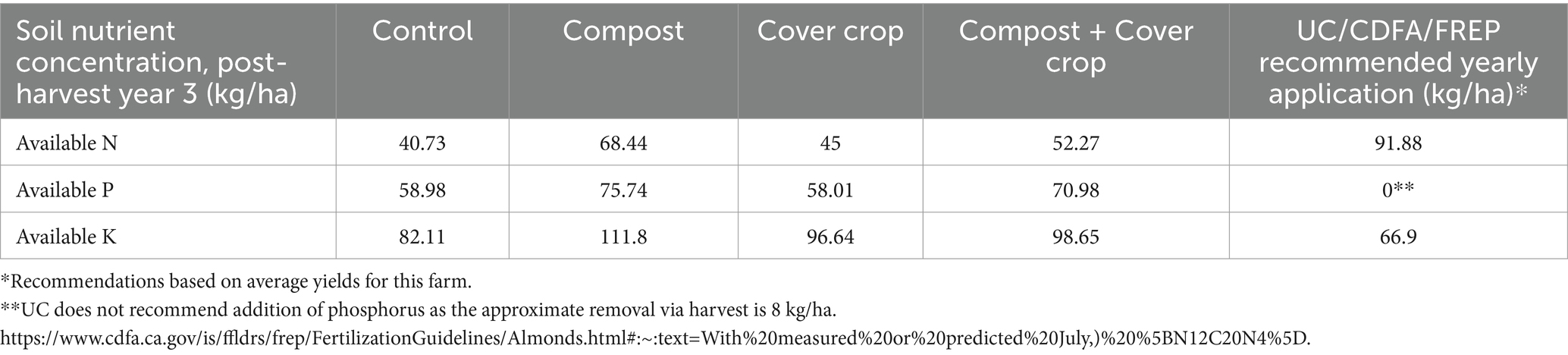
Table 6. Plant available NPK vs recommended application rates.
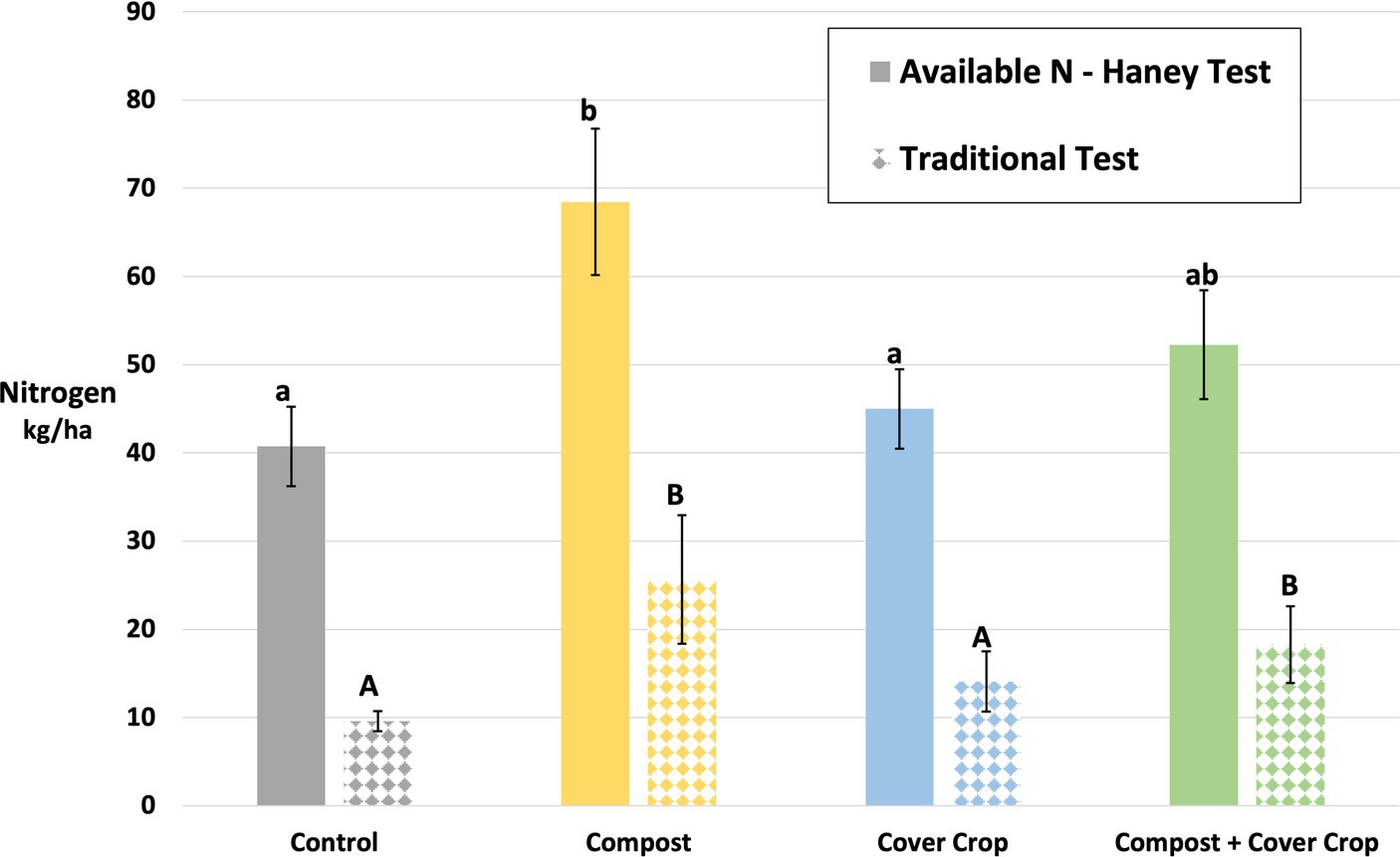
Figure 5. Nitrogen content of soils by treatment, contrasting the traditional test for soil N and the Haney Test for available N. Different letters indicate a significant difference between means (p < 0.05).
Phosphorus (P) variables (inorganic, total, and available P) trended toward a compost main effect (p < 0.07; Tables 5, 6), with 23% more soil P when compost was applied. There were no significant specific treatment differences.
Tables 5, 6 includes Potassium plus nine other nutrients. There was a significant compost main effect for Calcium, Magnesium, and Zinc, with 8, 5, 24% higher concentrations of these nutrients in composted plots compared to non-composted plots (p < 0.05) but lower for Copper in composted plots (p < 0.05) by 13%. The only significant cover crop main effect was for Sulfur, which was 23% lower with cover crops compared to without (p < 0.05). The one-way analysis showed that there were significant (p < 0.05) differences for Calcium (COMP > MSCC) Zinc (COMP > both CONT and MSCC; COMP + MSCC > MSCC) and Sulfur (COMP > CONT, MSCC, and COMP + MSCC).
3.1.4 Haney soil health score and nutrient value
The Haney Soil Health Score (SHS) (Haney et al., 2018) is a composite index incorporating the measures of water extractable organic carbon and nitrogen plus respiration. Soil Nutrient Value (NV) is a dollar per hectare value based on the nutrient content. The two factor analysis for compost and cover crop showed a significant compost main effect for both SHS and NV (p < 0.05; Table 7), where both were higher (15 and 25%, respectively) in compost-applied plots compared to non-compost plots. The One-way analysis showed that for SHS, COMP scores were significantly greater than both CONT and MSCC and the NV of COMP was greater than CONT (p < 0.05; Table 7).
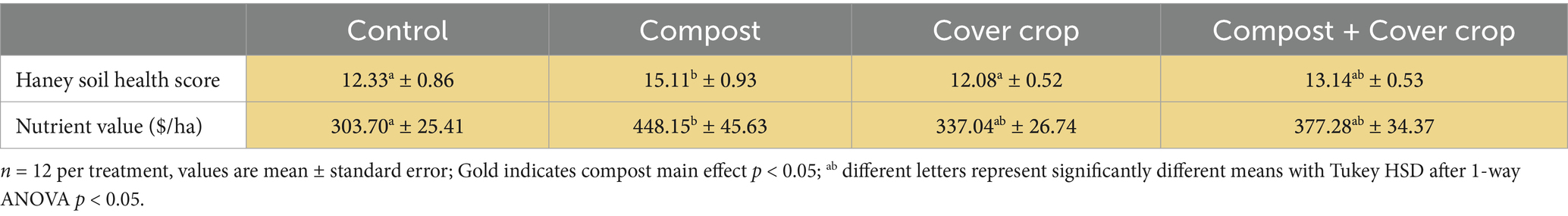
Table 7. Haney soil health score, nutrient value, and organic C:N.
3.1.5 Soil biology and the phospholipid fatty acid analysis
The soil microbiome, as characterized with the Phospholipid Fatty Acid Analysis (PFLA) is summarized in Table 8 with biomass (ng PLFA/g soil) and as the percent of the total biomass (Total Living) for each group by treatment. The two factor analysis for compost and cover crop showed a significant cover crop main effect for total biomass (Total Living); 22% greater with cover cropped versus non-cover cropped (p < 0.01; Table 8). Cover cropped plots also had significantly more gram positive, gram negative, actinomycetes, total fungi, arbuscular mycorrhizal fungi (p < 0.05). A marginally significant result was also detected for total bacteria (p = 0.06) and saprophytic fungi (p = 0.08) in cover cropped plots. There was a significant compost main effect for gram negative (p < 0.05), and a marginally significant effect for both total bacteria and saprophytic fungi (p < 0.1) was detected. There was a significant compost * cover crop interaction for the “undifferentiated” category, which includes all microorganisms that could not be identified, as there were far fewer undifferentiated microorganisms in the COMP plots than the other treatments (p < 0.05, Table 8).
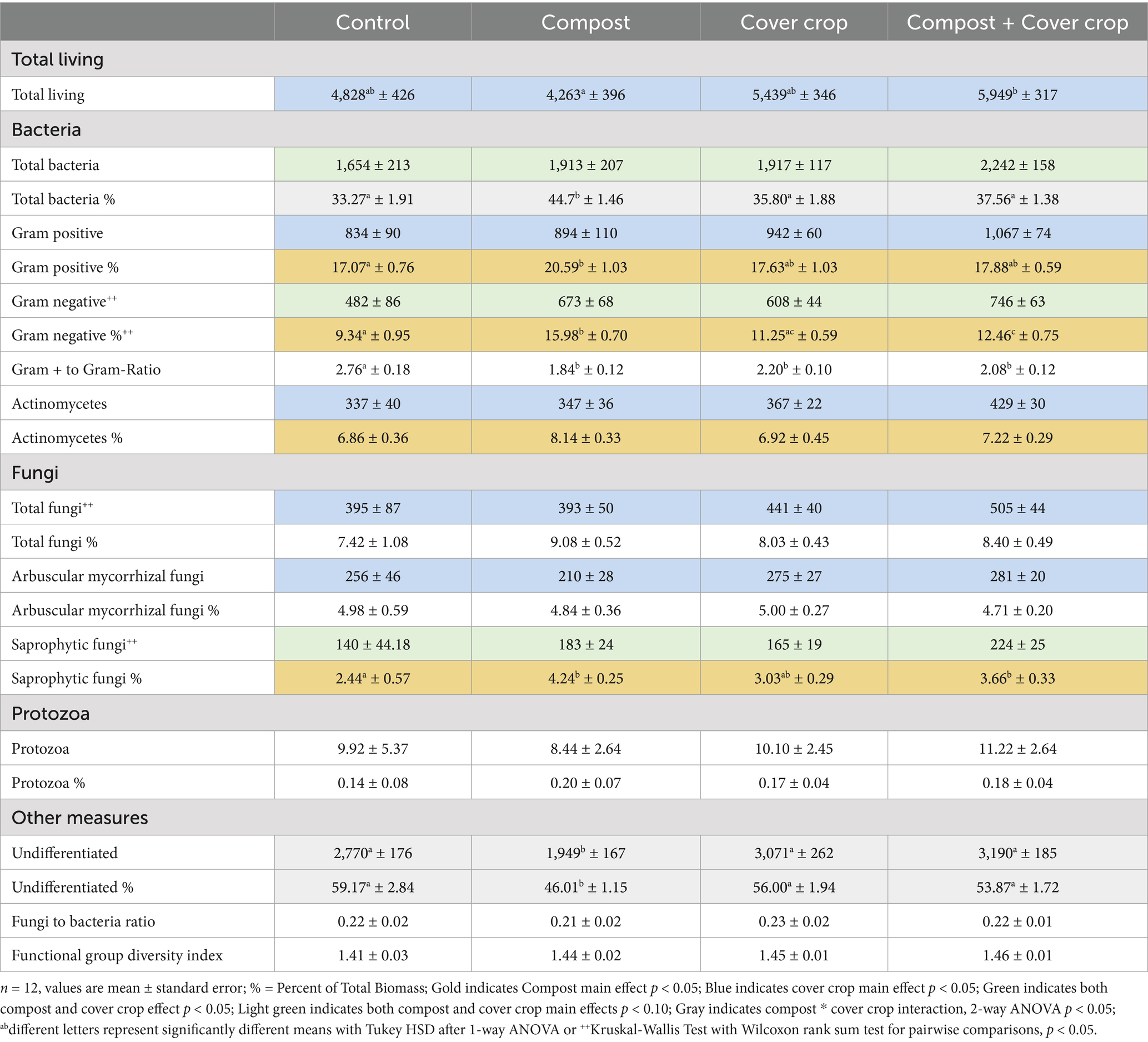
Table 8. Quantities of soil phospholipid fatty acid (ng PLFA/g soil) by treatment and group.
3.2 Orchard understory productivity and quality
3.2.1 Orchard understory primary productivity
The understory biomass of cover cropped plots was significantly greater than non-cover cropped. The two factor analysis showed a significant cover crop main effect on dry weight (p < 0.05) and the one-way analysis determined that MSCC and COMP + MSCC were both significantly higher than COMP or CONT (p < 0.05, Figure 6a). When combined, MSCC and COMP + MSCC produced 35% more understory biomass compared to CONT and COMP (8.22 Mg Ha−1 and 5.78 Mg Ha−1 respectively).
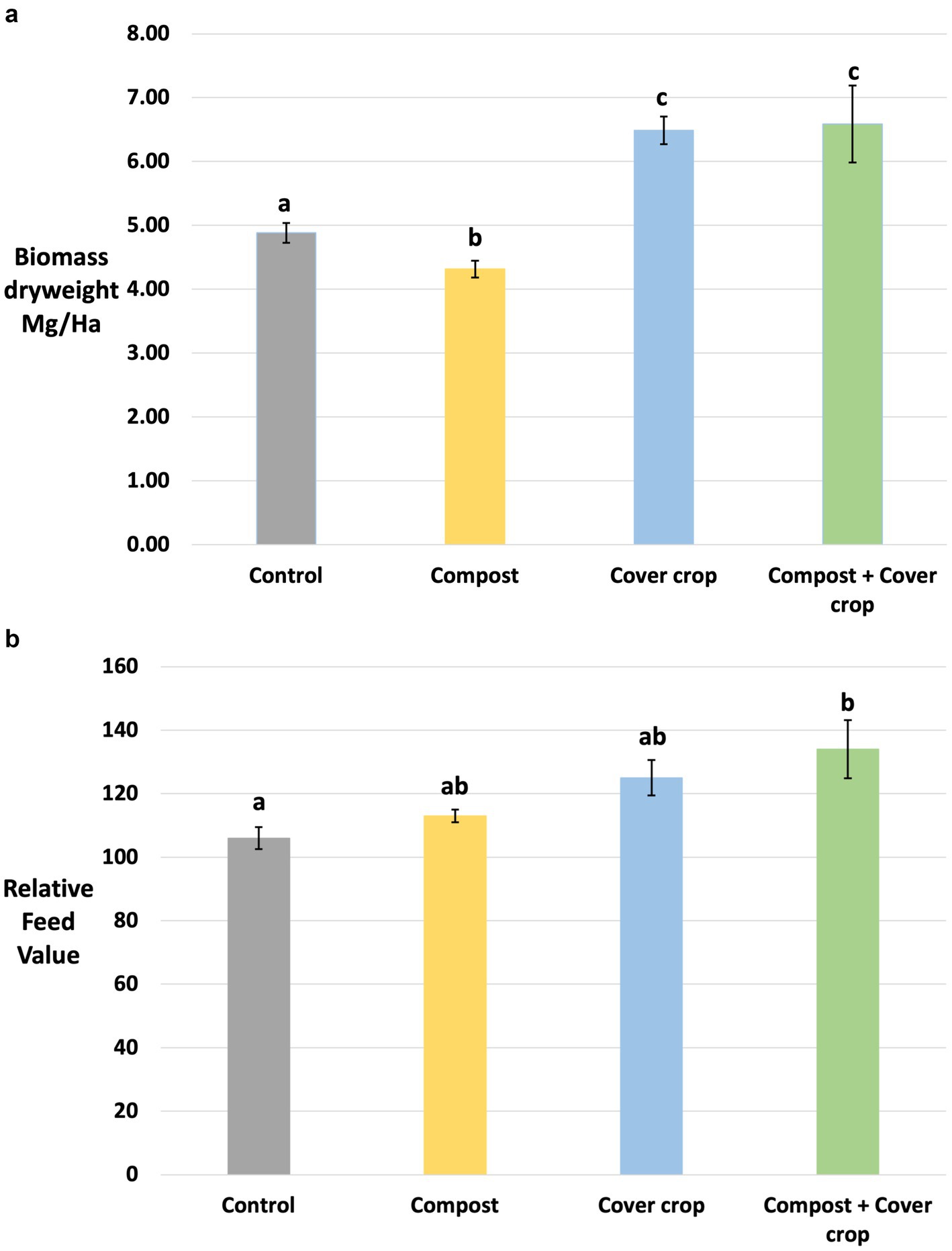
Figure 6. (a,b) Comparison of understory biomass as measured by dry weight and Relative Feed Value by treatment. Different letters indicate a significant difference between means (p < 0.05).
3.2.2 Orchard understory forage quality
Table 2 contains forage quality analysis results for quantified individual components (protein, carbohydrates, fiber, fat, and minerals), and calculated composite measures (Relative Feed Value, Total Digestible Nutrients, and Net Energy for Gain and Maintenance). In general, forage quality from cover cropped plots (MSCC and COMP + MSCC) was higher than non-cover cropped (CONT and COMP). The two factor analysis showed that cover crops had significantly higher moisture (5%), Relative Feed Value (RFV, 17%), Total Digestible Nutrients (4%), Net Energy for Gain (9%) and Maintenance (7%), crude and available protein (27 and 25%), nitrogen (26%), and calcium (25%; p < 0.05, Table 2). Water soluble carbohydrates and fiber measures (Acid Detergent Fiber and Neutral Detergent Fiber) were significantly lower with cover crop, 29, 9 and 13%, respectively, (p < 0.05, Table 2). The one-way analysis determined that MSCC had significantly higher RFV versus CONT, reflecting the greater potential intake and digestibility of the multispecies cover crop forage. The forage RFV of COMP + MSCC was 23% higher than CONT (Figure 6b). Both MSCC and COMP + MSCC had significantly more nitrogen, crude protein, and available protein compared to CONT (p < 0.05); up to 34, 34 and 32%, respectively. For minerals: Potassium was significantly higher (25%) in MSCC than CONT; Calcium was higher in both MSCC and COMP + MSCC than CONT by 34% on average; Magnesium was 27% lower in MSCC and COMP + MSCC than COMP (p < 0.05, Table 2).
3.3 Almond crop yield
The almond crop yields are presented by year and variety in Table 9 and Figure 7. While no significant differences were found in years one and two of the study, year three (2022) showed substantially higher yields from all treatment plots (COMP, MSCC and COMP + MSCC). The two-factor analysis showed a compost effect for Price (9% higher, p < 0.05) and Nonpareil (19% higher, p = 0.07) and cover crop main effect for Price (44% higher, p < 0.05). The one way analysis found that for the Price variety, yield was significantly higher in COMP + MSCC compared to CONT (49% higher, p < 0.05, Figure 7).
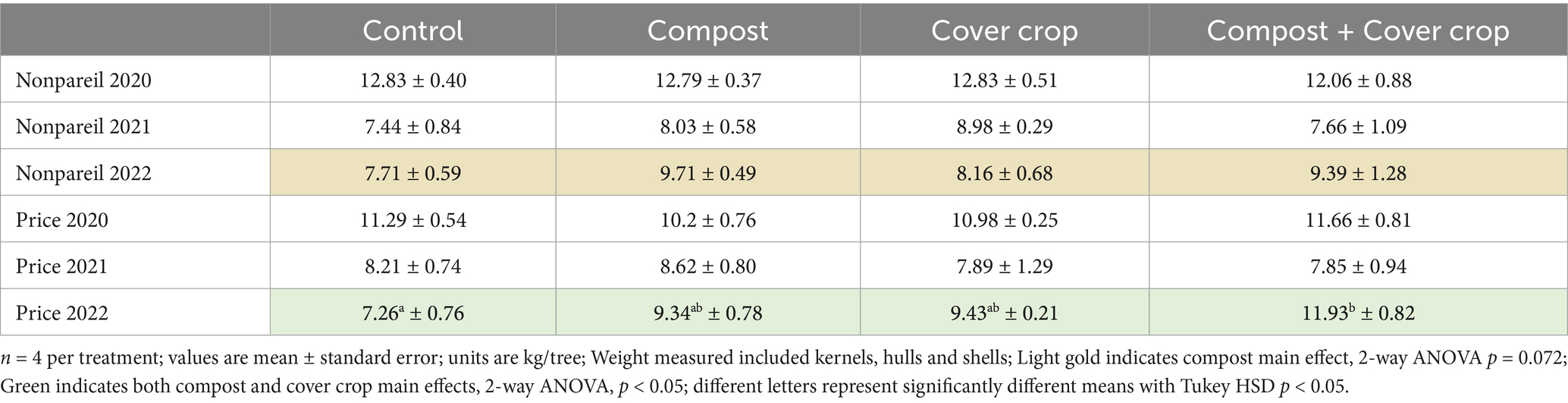
Table 9. Almond yield by variety by year by treatment.
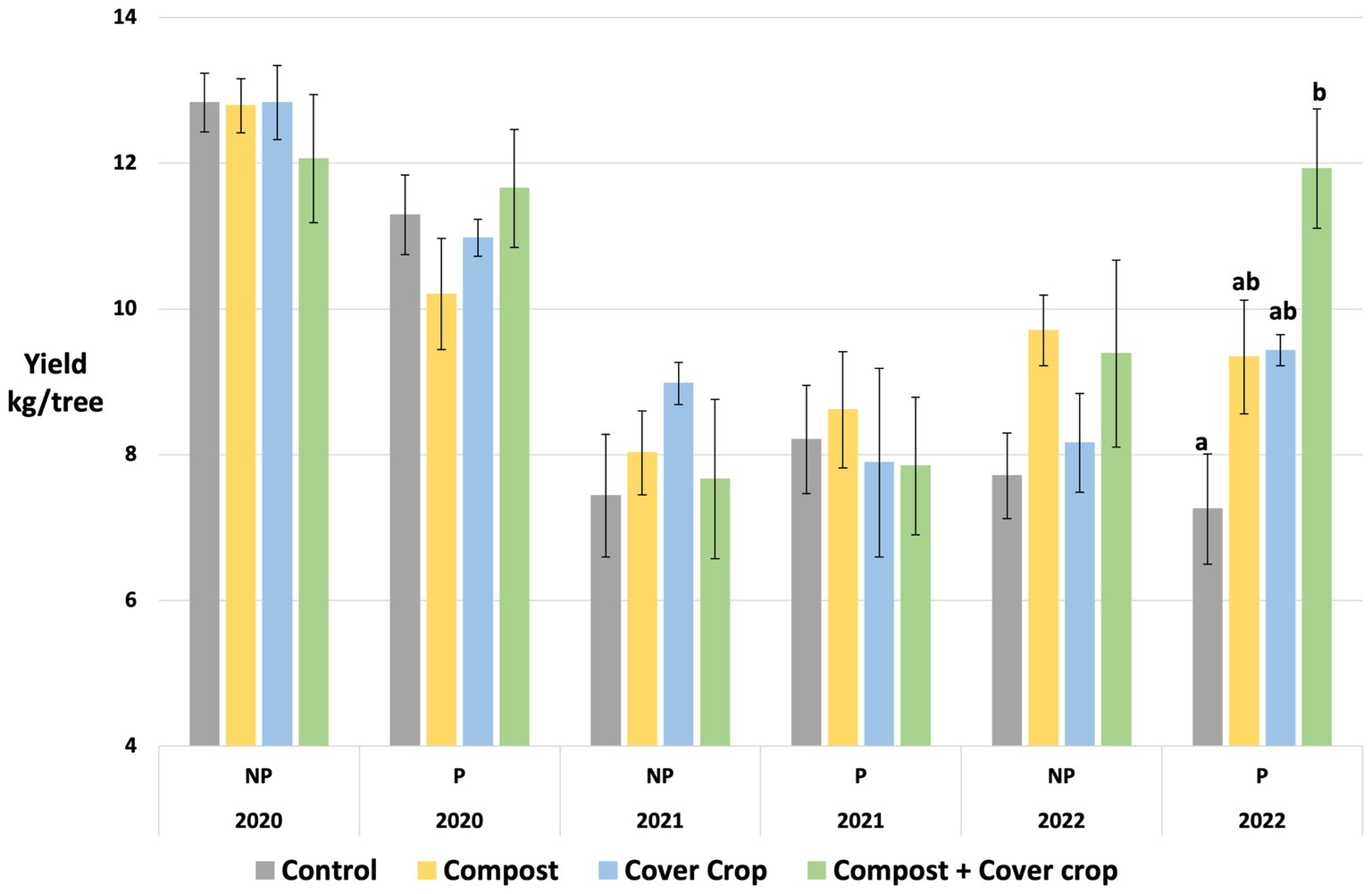
Figure 7. Almond yield by variety (NP = Nonpareil; P = Price), treatment and production year. Different letters indicate a significant difference between means (p < 0.05).
3.4 Almond kernel nutrient analysis
Almond kernel nutrient concentrations are listed in Table 10. The two-factor analysis showed compost main effects for Phosphorus, Nitrogen, and Zinc (p < 0.05) and Calcium and Magnesium (p < 0.07). Almond kernels from plots where compost was applied had 4% more Phosphorus, 8% more Nitrogen, 4% more Zinc and 14% more Ca, compared to plots without compost.
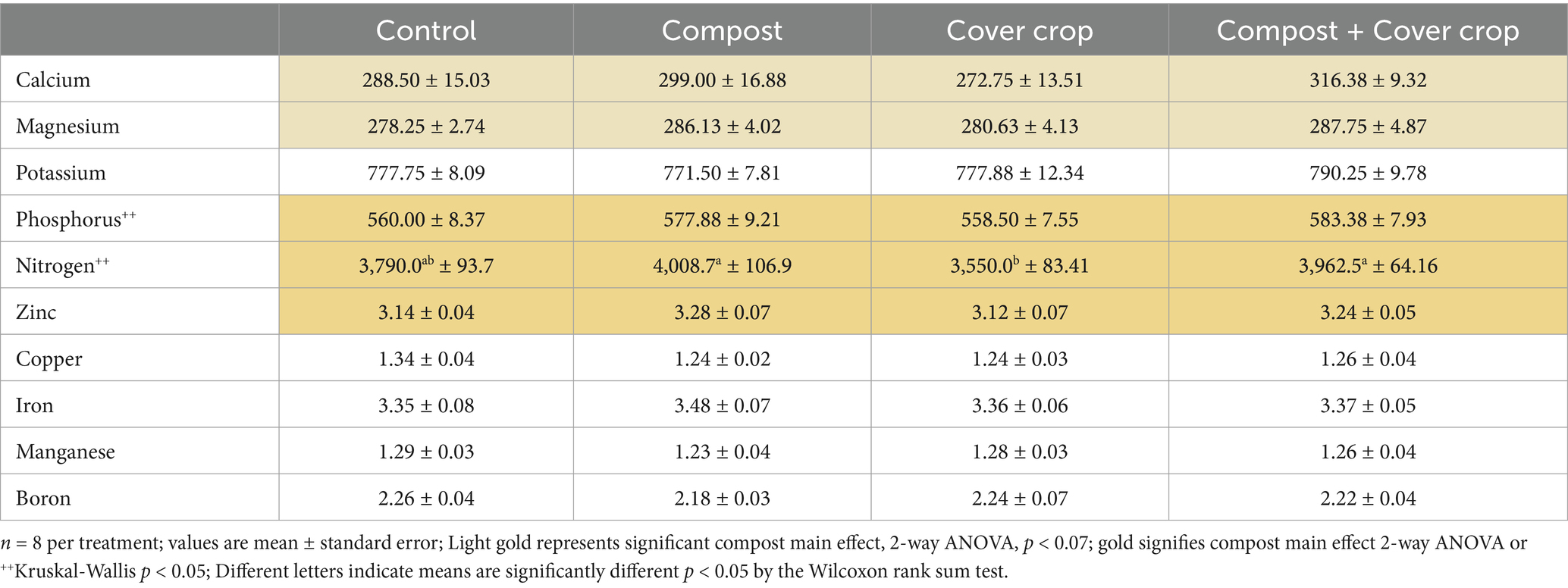
Table 10. Almond kernel nutrient density, mg/100 g, by treatment.
4 Discussion
The application of compost and/or multispecies cover crop improved soil health and overall productivity of this mature organic almond orchard. Results showed that both compost and multispecies cover crop treatments added carbon to the system, with the addition of composted organic matter and via photosynthesis-driven increases in plant and microbial biomass. The increase in carbon correlated with improved soil structure, more active soil microbiology and enhanced nutrient cycling throughout the orchard system. The multispecies cover crop produced more understory biomass with higher forage quality compared to resident vegetation, which when grazed by sheep, translates into more kg of lamb harvested per Ha. Both almond crop yield and kernel nutrient density were significantly higher after 3 years of regenerative treatment. These whole-system benefits result in a more resilient agroecosystem.
4.1 Soil health
4.1.1 Soil biology: respiration and phospholipid fatty acid analysis
In this study, soil respiration rates (CO2–C) and % microbial active carbon (%MAC) (Table 4) were substantially higher (up to 41 and 17% respectively) in the COMP plots compared to all other treatments, revealing higher microbial activity associated with compost application. In contrast, the PLFA analysis found greater quantities of microbial biomass associated with the multispecies cover crop; up to 25% more in MSCC and COMP + MSCC compared to CONT and COMP (Table 8). The different soil nutrient profiles of each treatment (Table 7) provided different resources for microbes resulting in different microbial community structures and function (Hartmann and Six, 2022; Pommier, 2023). While some research has found microbial biomass to be correlated with microbial activity (Finney et al., 2017). Pérez-Murcia et al. (2021) found that soil respiration did not correlate with soil microbial biomass quantities when different types of compost were applied in almond orchards.
Our finding that microbial biomass was greatest in the multispecies cover crop plots (MSCC and COMP + MSCC) is consistent with previous research. One meta-analysis of 81 studies comparing cover crops to no cover crop found that the quantity of PLFA was 24% higher with cover crops (Muhammad et al., 2021). Since understory vegetation in these plots was more productive and species rich, the greater soil microbial biomass found in them is consistent with two general relationships found in the agricultural and ecological literature: (1) greater plant biomass (PB) is associated with greater microbial biomass (MB) (Bainard et al., 2020; Finney et al., 2017, 2016); and (2) higher plant diversity (PD) is associated with greater MB (Lange et al., 2015; Prommer et al., 2020).
Quantities of bacteria, as well as the Saprophytic fungi, were also strongly impacted by the application of compost (Table 8). This is consistent with previous orchard-compost studies that found either higher microbial activity or microbial biomass associated with compost use (Baldi et al., 2018; Forge et al., 2013; Hodson et al., 2021; López et al., 2014; Neilsen et al., 2014; Pérez-Murcia et al., 2021; Sharaf et al., 2021; Thompson et al., 2019).
The highest PLFA in the MSCC+COMP plots might indicate that the multispecies cover crop supported an increased size of the soil MB, and that the compost provided additional resources to enhance this growth. Moukanni et al. (2022) proposed that much of the movement of carbon through the soil is directed by the microbial carbon pump. This carbon is the base of the myriad trophic interactions that facilitate the nutrient mining from the soil and generate availability to the crop (Kallenbach et al., 2016). MB was greater in COMP or MSCC versus CONT, but greatest for the combined treatment (COMP + MSCC (Table 8)). This additive, or synergistic effect of the two practices was also evident for total bacteria and total fungi. For Actinomycetes, the individual treatments led to minimal increases whereas the combined treatment resulted in 27% more biomass versus CONT (Table 8). For Arbuscular mycorrhizal fungi (AMF), COMP plots had lower biomass versus CONT, which could indicate that compost inhibited the growth of AMF. However, when combined with the multispecies cover crop this was no longer the case. Vasilikiotis et al. (2020) surveyed multiple almond orchards using different practices to determine which factors were most important for tree root AMF colonization and found the presence of cover crops or native vegetation between the tree rows to be the most important factor. They also found that colonization was not affected by compost use (Vasilikiotis et al., 2020).
Finally, the soil biology indicators highlight the added value of the multispecies cover crop over resident vegetation, which is less productive and diverse. Other studies have found similar increases in microbial biomass, activity and diversity associated with cover crops compared to resident vegetation (Castellano-Hinojosa and Strauss, 2020; Jiao et al., 2023; Moreno et al., 2009; Rodriguez-Ramos et al., 2022; Vasilikiotis et al., 2020; Wang et al., 2020).
4.1.2 Soil carbon
Compost application had the biggest impact on SOM and SOC as compared to controls over the course of this 3 years study (Table 4). The change translates to 0.64 Mg Ha−1, or 21%, more SOC due to compost application (at 18 Mt./Ha) versus control. This is consistent with previous research that has also found compost application associated with increased SOC to varying degrees depending on compost source material, soil texture, and farming practices (Khalsa et al., 2022; Villa et al., 2021). In their literature review across a range of agroecosystems, Diacono and Montemurro (2011) found that compost increased SOC by 90% when compared to non-compost treatments and up to 100% when compared to chemical fertilizer treatments. Numerous orchard system studies have documented increased SOC with compost applications (Baldi et al., 2018; De Leijster et al., 2019; Hodson et al., 2021; Khalsa et al., 2022; Lepsch et al., 2019; López et al., 2014; Mays et al., 2014; Neilsen et al., 2014; Pérez-Murcia et al., 2021; Thompson et al., 2019; Villa et al., 2021) and a 2016 meta-analysis found an average increase in SOC of 4.07 Mg Ha−1 Yr−1 with the use of organic amendments in woody crop systems (Vicente-Vicente et al., 2016).
Although the multispecies cover crop captured more atmospheric CO2 through photosynthesis, it did not yet translate into increased soil carbon stocks in the MSCC treatment plots. However, other studies have documented increases in soil carbon with the use of cover crops, the degree of which depended on cover crop species, growing period, and termination method (Moukanni et al., 2022; Poeplau and Don, 2015; Vicente-Vicente et al., 2016). Poeplau and Don (2015) found that the average increase of SOC with the use of winter cover crops was 0.32 Mg Ha−1 Yr−1 compared to fallow cropland, and Vicente-Vicente et al. (2016) found the average increase in SOC was 1.03 Mg Ha−1 Yr−1 with the use of cover crops in Mediterranean woody crop systems. The unique practices of the farm in this study make comparisons to the literature difficult. First, the orchard system used had been under no-till management with a permanent non-seeded vegetative cover, for 18 years. Most orchard studies that have shown increases in SOC have compared the impact of cover crop to a control defined by frequent tillage and bare soil (Almagro et al., 2013; Martínez-Mena et al., 2020; Özbolat et al., 2023; Ramos et al., 2011; Sastre et al., 2020). In two previous studies that observed differences between seeded cover crops and spontaneous vegetation, both documented slight increases of SOC in the top 5 cm of soil (Repullo-Ruiberriz de Torres et al., 2021; Sastre et al., 2020). Second, the farm uses sheep to terminate the cover crop. This method, where the sheep remove some of the vegetation and trample the rest, is much less common than mowing and/or incorporating the cover crop. Finally, it is possible that the three-year length of this study was not long enough for the multispecies cover crop to affect a difference in SOC. The greater production of PB of the cover crop and the associated MB represents an increase of carbon to the system that may eventually translate into more SOC.
We did not find an increase in SOC in the combination treatment compared to compost alone. Similarly, Vicente-Vicente et al. (2016) found that, on average, the combination of compost and cover crop use in woody crops produced an intermediate level of carbon sequestration of 2.62 Mg Ha−1 Yr−1, lower than compost applications, and higher than cover crop when each was an independent practice. This would indicate that cover crops can contribute to soil carbon accrual, but do so more slowly, even when combined with organic amendments.
4.1.3 Soil nitrogen and other macro- and micro-nutrients
All measures of soil nitrogen were significantly higher in COMP and COMP + MSCC compared to MSCC and CONT, except ammonium, which was only higher in COMP (Tables 5, 6). The annual application of compost resulted in the accumulation of nitrogen, both organic and inorganic, in the soil over 3 years. This is consistent with previous studies of orchards using compost or other organic amendments that have observed increases in soil nitrogen, including studies in almonds (De Leijster et al., 2019; Hodson et al., 2021; Khalsa et al., 2022; Villa et al., 2021), avocados (López et al., 2014), and apples (Thompson et al., 2019).
The use of the multispecies cover crop had mixed effects on soil N. For instance, soil nitrate levels were significantly higher while soil ammonium levels were lower in MSCC versus CONT. It is possible that the MSCC and the associated increase in total microbial biomass improved N cycling, compared to control, facilitating conversion of ammonium to nitrate. More likely, the elevated soil nitrate may be the result of nitrogen fixation by legumes within the mix; almost half of the multispecies cover crop species were nitrogen fixing legumes. Several studies have demonstrated an increase in soil nitrate levels in response to legume rich multispecies cover crops in orchard systems (Almagro et al., 2013; Castellano-Hinojosa and Strauss, 2020; Martínez-Mena et al., 2020; Özbolat et al., 2023; Repullo-Ruiberriz de Torres et al., 2021; Thompson et al., 2019).
While the treatment interaction was not statistically significant, the data suggest that the multispecies cover crop slowed the accumulation of soil nitrate (COMP > COMP + MSCC). It’s possible that excess nitrate from the compost was utilized to produce more both plant and microbial biomass, as both of these metrics were significantly higher in MSCC and COMP + MSCC plots compared to COMP and CONT. These differences also highlight enhanced nutrient cycling with multispecies cover crop compared to the control plots where only resident vegetation was present.
Soil nutrients (P, Ca, Mg, and Zn) were significantly higher in COMP and COMP + MSCC compared to MSCC and CONT (Tables 5, 6). Like this study, soil P has frequently been found to increase with compost application in perennial cropping systems (Hannam et al., 2016; López et al., 2014; Pérez-Murcia et al., 2021; Thompson et al., 2019) and Thompson et al. (2019) found that Ca, B, Mg, Mn were also higher when composts were used in apple production. Potassium (K), however, was not statistically different between treatments, but was highest in COMP. There was also a probable interaction effect (p = 0.10) where MSCC appeared to reduce the accumulation of potassium from the compost in the COMP + MSCC treatment. Potassium was still higher in MSCC versus CONT, indicating that the multispecies cover crop was facilitating more K-cycling compared to CONT’s resident vegetation. Some previous studies have found soil K to be higher with compost application in orchard systems (De Leijster et al., 2019; Pérez-Murcia et al., 2021; Thompson et al., 2019).
Finally, the multispecies cover crop did not appear to reduce soil nutrient availability for the almonds. MSCC did not have lower soil total or available N, P K, or most trace minerals (Ca, Mg, Zn, Mn, Co, Fe, S, and Al) compared to CONT (Tables 5, 6). This is not consistent with the hypothesis that cover crops compete for available nutrients with cash crops, a finding of studies of single species cover crops (Sastre et al., 2020). This could be an important advantage of using a multi-species cover crop mix (Table 1).
4.1.4 The Haney test
The Haney test provides a holistic chemical and biological assessment of soil health. Soil nutrient extractions are accomplished with water and organic acids, which more closely mimic the soil solution versus the traditional soil extractants. Values are determined by integrating one-day soil respiration values, a measure of the soil microbial activity, and water extractable C and N into the formulas for plant-available nutrients, the Soil Health Score (SHS), and the Nutrient Value (NV) (Tables 4–7; Haney et al., 2018).
Available N, P and K and Soil Health Scores were highest in COMP and COMP + MSCC plots. The soil respiration rate (CO2–C) was significantly higher for COMP compared to all other treatments. This higher level of soil respiration, the greater concentrations of water extractable organic carbon, and plant available N, P and K demonstrate that there was more biological activity, and thus, more nutrient cycling occurring in COMP and COMP + MSCC (Tables 4–7; Haney et al., 2018). Since there was also evidence of nutrient cycling in the cover crop plots (higher forage quantity and quality, higher MB, and increased almond yields), it is possible that increases in SHS for MCSS may have a 2–3-year lag compared to the COMP treatment (L. Gunderson, Regen Ag Soil Laboratory, pers. comm.). Fenster et al. (2021) found that regeneratively farmed almond orchards had higher SHS than conventionally farmed almond orchards.
The higher availability of nutrients in all treatment plots (COMP, MSCC, and COMP + MSCC) is reflected in their higher Nutrient Value (NV) dollar amounts in Table 7, compared to CONT plots. This represents 11–48% more nutrients being supplied by the compost or the multispecies cover crop. This is an example of why this measure is useful for crop nutrient management and could provide an opportunity for reductions in nutrient applications by growers. The Haney test offers an advantage over traditional chemical extraction tests that underestimate the availability of N, P and K (Figure 3). Underestimating available soil nutrient levels may lead to the overapplication of external inputs. This is not only an unnecessary financial expense, but potentially, also nutrient loss to the environment. All soils samples were collected in September, just after almond harvest, and before the annual fall compost application and winter cover crop planting. Thus, the soil nutrients detected were what remained after harvest. These concentrations represent a substantial soil nutrient reserve for the following year (Table 6), especially considering the recommended fertilizer application rates for the almond production (CDFA - FREP - CA Fertilization Guidelines - Almonds, n.d.).
4.1.5 Soil structure
Compost application and multispecies cover crop had mixed effects on soil structure. Soil bulk density (BD) was not affected by compost or cover crop, however, aggregate stability (in the top 15 cm of soil) did improve with the application of compost (Table 3). Although previous studies have found BD to be improved (reduced) with the use of compost and cover crops (Blanco-Canqui and Ruis, 2020; Mays et al., 2014), certain characteristics of this orchard may explain its observed lack of response. As a mature, organic, perennial system with clay soils (estimated at 35% clay), bulk density may not respond quickly or dramatically to soil health improvement. In addition, some compaction may result from equipment use (seeding, compost application, mowing, and harvest) and sheep traffic in the orchard. Finally, the permanent no-till status of this orchard could be a factor, as it has been found that long term no-till can lead to increases in BD (Li et al., 2020).
In contrast to BD, the aggregate stability of the top 15 cm of soil was 9% higher in plots where compost had been applied (COMP and COMP + MSCC) versus non-compost (CONT and MSCC). This result is consistent with the review by Diacono and Montemurro (2011) which showed improvement in aggregate stability due to compost application in multiple farming systems. This is also consistent with the studies by Almagro et al. (2013) and Soto et al. (2021), that found aggregate stability was improved with regenerative practices such as ground covers and organic amendments, compared to conventional practices in almond orchards in Spain. The increased microbial presence and activity in these soils likely improved soil aggregation with increased glue-like exudates and enhanced mycorrhizal and mycelial networks and that are favored by no-till management (Kabir, 2005).
4.2 Orchard productivity
4.2.1 Understory vegetation
4.2.1.1 Understory primary productivity
Growing vegetation in the alleys between tree rows is one way to maximize photosynthesis and to increase the overall orchard productivity. Research has shown that instead of competing for water and nutrients, orchard floor plants facilitate the enhanced movement and uptake of these resources through the soil via root and fungal networks (Castellano-Hinojosa and Strauss, 2020). In this study the cover crop treatments (MSCC and COMP + MSCC) produced greater plant biomass versus plots without cover crops planted (CONT and COMP). This greater PB is what we would expect with the multispecies cover crop’s greater number of plant species due to niche partitioning, complementarity, and over yielding as these effects are well documented in the literature in both natural and managed systems where increased plant species richness is correlated with higher primary productivity (Isbell et al., 2017; Tilman et al., 1996; Weisser et al., 2017).
4.2.1.2 Understory forage quality
Understory forage quality is important as a source of feed for sheep. Multispecies cover crop (MSCC and COMP + MSCC) biomass had higher protein, net energy, palatability, digestibility, K and Ca (Table 2). Others have shown the benefits of cover crops as livestock forage in both annual and perennial systems, with more diverse mixtures of species and functional groups providing the greatest benefit for cash crop production, livestock nutrition, and soil health (Bainard et al., 2020; Farney et al., 2018; McKenzie et al., 2017; Mozea, 2022; Opoku et al., 2024; Ramos et al., 2011; Wortman et al., 2012). Here, MSCC and COMP + MSCC plots had more plant species and functional groups along with larger, more diverse soil microbiomes compared to CONT or COMP which resulted in greater forage production and quality.
4.2.1.3 Influence of sheep grazing
Integrating livestock into cropping systems benefits soil health and farm resiliency through ecological intensification (Brewer and Gaudin, 2020; Cappelli et al., 2022; Franco et al., 2021; Franzluebbers and Martin, 2022; Peterson et al., 2020). The benefits that ruminant grazing provide differ from other cover crop termination strategies such as incorporation, mowing, roller-crimping, and herbicide application. Ruminants, such as sheep, reduce plant biomass while stimulating plant growth, provide excreta in the form of feces, urine, and saliva; and increase the contact between the vegetation and the soil by trampling (personal observation). With this study, every effort was made to “equalize” the sheep impact by allowing them to graze an equal amount of time in each plot. Their behavior of smashing vegetation that was not eaten made it difficult to gauge exact intake. Since sheep are known to graze preferentially, having different plants in each plot may have led to different grazing patterns. Sheep could have contributed to soil compaction despite the short durations of their grazing time. Sheep are integrated into almond orchards in Spain (Ramos et al., 2011) for grazing cover crops or “permanent natural cover,” as well as occasionally in California for the same purpose (Fenster et al., 2021). This is an example of the multispecies cover crop providing nourishment for the soil, the crop and the livestock.
4.2.2 Almond production and quality
4.2.2.1 Crop yield
While climate smart practices such as compost application and cover crops are employed because they provide multiple benefits to farms and the environment, their impact on cash crop yield is a primary consideration for their adoption (DeVincentis et al., 2020; Finney et al., 2017; Khalsa and Brown, 2017; Sastre et al., 2020; Wauters et al., 2023). There were no yield reductions due to treatments in years one and two; and by year three there were significantly higher yields in the treatment plots. For the Price variety, COMP + MSCC produced 60% more while COMP and MSCC produced 25% more almonds than CONT (Table 3; Figure 3). For the Nonpareil, COMP and COMP + MSCC yielded 20% more than CONT and MSCC.
Compost application correlated with higher almond yields, which is consistent with other orchard studies of organic amendment use where crop yields were either maintained—nectarines (Baldi et al., 2018); almonds (Pérez-Murcia et al., 2021); apples (Neilsen et al., 2014) or increased—peaches (Lordan Sanahuja et al., 2015); apples (Forge et al., 2013); avocados (López et al., 2014) compared to control treatments, including controls that included synthetic fertilizers. It took 3 years to detect, the increased nutrient availability in the compost plots’ soil likely translated into our observed higher yields (Figure 7).
Conversely, for this study, the impact of the multispecies cover crop was different for the two varieties. While there was a clear benefit to the Price variety, there was no effect on yield for Nonpareil. Previous orchard studies have observed mixed yield responses to the use of cover crops: negative—olives with grass cover (Sastre et al., 2020); neutral—olives with vetch cover (Sastre et al., 2020), almonds with oats and vetch (Özbolat et al., 2023), apples with legume and grass cover (Sharaf et al., 2021), and positive—almonds with oats and vetch (Martínez-Mena et al., 2020). No studies have addressed the impact of multispecies cover crops on orchard crop yields, but, in row crop systems. Finney et al. (2017) and Chalise et al. (2019) each saw yields of soybeans increase after 3 years in rotations with multispecies (up to six) cover crops. Additionally, a recent review concluded that increased plant diversity was associated with increases in crop and forage yield (Isbell et al., 2017), and the inclusion of the multispecies cover crop in this system increased plant diversity substantially.
The combination of compost and cover crop showed a seemingly synergistic effect on the Price yields. Few previous studies have examined this combination, but those that did found either no treatment differences [almond (De Leijster et al., 2019; Fenster et al., 2021)] or increased yields [olives (Sofo et al., 2010)]. While soil nutrient concentrations were impacted most by compost and the understory vegetation quantity and quality by multispecies cover crop seeding, soil biology, as measured by MB, was greatest for all major groups in the combined treatment plots. The synergistic effect of combined practices on soil biology may be the mechanism for the synergistic effect of the combined treatment on Price yields.
Maintaining yields while using regenerative practices will be vital to their adoption. Yield levels and consistency are important to maintain cash flow and markets for crops (personal observation) and may be more likely when using alternative practices that build soil health compared to conventional reliance on fossil fuels and chemical inputs. Li et al. (2019) found, in their review of 24 years of data, that yields of tomato were more consistent and sometimes higher with organic management that included cover crops and compost compared to conventional management. They propose that the slow, steady buildup of soil organic carbon and the associated slow, steady release of nutrients allowed tomato yields to be more resilient to environmental variation, resulting in consistent yields even when conditions would typically have reduced yields. In this study, these practices were introduced into an orchard characterized by low disturbance which may have guarded against a yield drag (due to treatment implementation) the first 2 years. The higher yields in year three may have occurred because sufficient time had passed to allow different soil physical, chemical, and biological processes to boost crop yield.
4.2.2.2 Almond kernel quality
Nuts from the compost-applied plots had higher concentrations of Ca, Mg, P, N, and Zn than nuts from non-compost-applied plots. Higher soil nutrient levels in the composted plots, suggests that more nutrient cycling is happening in the composted soil, making those nutrients available to the trees. This is consistent with data accumulating from research in Spain and California showing that regenerative practices increase the nutritional value of almonds. Cárceles Rodríguez et al. (2023) found that antioxidant activity and total polyphenol content of almonds increased with 5 years of legume cover crop; and Pérez-Murcia et al. (2021) found differences in fiber, protein, fat, sugar, organic acids, and sterols correlated with the specific composts used to produce them, and Fenster et al. (2021) found magnesium content was higher in regenerative nuts compared to conventional.
4.3 Economic assessment
Some basic financial statements can be made with respect to practice adoption: (1) The yield increase more than pays for the cost of compost application and cover crop planting. (2) The increased forage quantity and quality translates into lower feed costs for lamb production. (3) Demonstrating higher nutrient content in the nuts may translate into a price premium. (4) Soil health management practices that increase soil carbon by enhancing biological and ecological processes will promote resiliency. (5) The cost of compost and cover crop seed was subsidized by the California Department of Food and Agriculture Healthy Soils grant for three full years before the yields increased. This farm had previously applied compost and single species cover crops but stopped due to high cost and lack of immediate return on investment. With this yield increase, farmers can implement these practices at their own cost for the longer term.
5 Conclusion
This three-year trial demonstrated higher primary productivity, including almond and forage crop yields, and improved soil health metrics. Treatment plots produced more and higher quality understory forage, more and higher quality nuts, and had higher overall Soil Health Scores which reflected their soils’ higher nutrient content and biological activity. The research highlighted the systems approach undertaken by the Massa Organics farm to produce almonds and lamb while enhancing ecosystem services. Both compost and cover crop practices added carbon, through importing organic matter (compost) and capturing more sun by growing understory biomass (cover crop). Mediated by greater or more diverse microbial communities, this translated into more soil organic carbon, increased nutrient cycling, higher soil nutrient availability, and greater yields and nutrient density of both almonds and forage. Finally, combined practices generated the greatest benefit. Improved soil health can increase farm resiliency and improve environmental and human health. In the face of increasing climate uncertainty, it should be the focus of agriculture to promote the benefits to all (DeVincentis et al., 2020; Wauters et al., 2023), and encourage more farmers, organic and not, to adopt these climate smart practices.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors without undue reservation.
Author contributions
RK: Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Visualization, Writing – original draft, Writing – review & editing. CD: Conceptualization, Funding acquisition, Methodology, Resources, Supervision, Writing – review & editing. GL: Methodology, Resources, Supervision, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was funded in part by a California Department of Food and Agriculture Healthy Soils Initiative grant to Massa Organics farm. Additional support was provided by the Center for Regenerative Agriculture and Resilient Systems at California State University, Chico.
Acknowledgments
We thank the many individuals that contributed their time and labor to this project, implementing the treatments, measuring yields, and collecting and processing soil samples: Greg Massa, Aundrea Asbell, and numerous staff and student assistants.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Footnotes
1. ^www.foss.us
References
Almagro, M., De Vente, J., Boix-Fayos, C., García-Franco, N., Melgares De Aguilar, J., González, D., et al. (2013). Sustainable land management practices as providers of several ecosystem services under rainfed Mediterranean agroecosystems. Mitig. Adapt. Strateg. Glob. Change 21, 1029–1043. doi: 10.1007/s11027-013-9535-2
Altieri, M. A., Nicholls, C. I., Henao, A., and Lana, M. A. (2015). Agroecology and the design of climate change-resilient farming systems. Agron. Sustain. Dev. 35, 869–890. doi: 10.1007/s13593-015-0285-2
Bacq-Labreuil, A., Crawford, J., Mooney, S. J., Neal, A. L., and Ritz, K. (2019). Cover crop species have contrasting influence upon soil structural genesis and microbial community phenotype. Sci. Rep. 9, 7473–7479. doi: 10.1038/s41598-019-43937-6
Bainard, L. D., Evans, B., Malis, E., Yang, T., and Bainard, J. D. (2020). Influence of annual plant diversity on forage productivity and nutrition, soil chemistry, and soil microbial communities. Front. Sustain. Food Syst. 4:560479. doi: 10.3389/fsufs.2020.560479
Baldi, E., Cavani, L., Margon, A., Quartieri, M., Sorrenti, G., Marzadori, C., et al. (2018). Effect of compost application on the dynamics of carbon in a nectarine orchard ecosystem. Sci. Total Environ. 637-638, 918–925. doi: 10.1016/j.scitotenv.2018.05.093
Blanco-Canqui, H., and Francis, C. A. (2016). Building resilient soils through agroecosystem redesign under fluctuating climatic regimes. J. Soil Water Conserv. 71, 127A–133A. doi: 10.2489/jswc.71.6.127A
Blanco-Canqui, H., and Ruis, S. J. (2020). Cover crop impacts on soil physical properties: a review. Soil Sci. Soc. Am. J. 84, 1527–1576. doi: 10.1002/saj2.20129
Blesh, J., VanDusen, B. M., and Brainard, D. C. (2019). Managing ecosystem services with cover crop mixtures on organic farms. Agron. J. 111, 826–840. doi: 10.2134/agronj2018.06.0365
Brewer, K. M., and Gaudin, A. C. (2020). Potential of crop-livestock integration to enhance carbon sequestration and agroecosystem functioning in semi-arid croplands. Soil Biol. Biochem. 149:107936. doi: 10.1016/j.soilbio.2020.107936
Canali, S., Trinchera, A., Intrigliolo, F., Pompili, L., Nisini, L., Mocali, S., et al. (2004). Effect of long term addition of composts and poultry manure on soil quality of citrus orchards in southern Italy. Biol. Fertil. Soils 40, 206–210. doi: 10.1007/s00374-004-0759-x
Cappelli, S. L., Domeignoz-Horta, L. A., Loaiza, V., and Laine, A.-L. (2022). Plant biodiversity promotes sustainable agriculture directly and via belowground effects. Trends Plant Sci. 27, 674–687. doi: 10.1016/j.tplants.2022.02.003
Cárceles Rodríguez, B., Lipan, L., Durán Zuazo, V. H., Soriano Rodríguez, M., Sendra, E., Carbonell-Barrachina, Á. A., et al. (2023). Linking conventional and organic rainfed almond cultivation to nut quality in a marginal growing area (SE Spain). Agronomy 13:2834.
Castellano-Hinojosa, A., and Strauss, S. L. (2020). Impact of cover crops on the soil microbiome of tree crops. Microorganisms 8:328. doi: 10.3390/microorganisms8030328
CDFA - FREP - CA Fertilization Guidelines - Almonds. (n.d.). Available online at: https://www.cdfa.ca.gov/is/ffldrs/frep/FertilizationGuidelines/Almonds.html (Accessed October 12, 2024).
Chalise, K. S., Singh, S., Wegner, B. R., Kumar, S., Pérez-Gutiérrez, J. D., Osborne, S. L., et al. (2019). Cover crops and returning residue impact on soil organic carbon, bulk density, penetration resistance, water retention, infiltration, and soybean yield. Agron. J. 111, 99–108. doi: 10.2134/agronj2018.03.0213
Chapagain, T., Lee, E. A., and Raizada, M. N. (2020). The potential of multi-species mixtures to diversify cover crop benefits. Sustain. For. 12:2058. doi: 10.3390/su12052058
De Leijster, V., Santos, M. J., Wassen, M. J., Ramos-Font, M. E., Robles, A. B., Díaz, M., et al. (2019). Agroecological management improves ecosystem services in almond orchards within one year. Ecosyst. Serv. 38:100948. doi: 10.1016/j.ecoser.2019.100948
DeVincentis, A. J., Solis, S. S., Bruno, E. M., Leavitt, A., Gomes, A., Rice, S., et al. (2020). Using cost-benefit analysis to understand adoption of winter cover cropping in California’s specialty crop systems. J. Environ. Manag. 261:110205. doi: 10.1016/j.jenvman.2020.110205
Diacono, M., and Montemurro, F. (2011). Long-term effects of organic amendments on soil fertility. Sustain. Agric. 2, 761–786. doi: 10.1007/978-94-007-0394-0_34
Farney, J. K., Sassenrath, G. F., Davis, C. J., and Presley, D. (2018). Forage mass production, forage nutritive value, and cost comparisons of three-way cover crop mixes. Crop Forage Turfgrass Manag. 4, 1–7. doi: 10.2134/cftm2017.11.0081
Fenster, T. L., Oikawa, P. Y., and Lundgren, J. G. (2021). Regenerative almond production systems improve soil health, biodiversity, and profit. Front. Sustain. Food Syst. 5:664359. doi: 10.3389/fsufs.2021.664359
Finney, D., Buyer, J., and Kaye, J. (2017). Living cover crops have immediate impacts on soil microbial community structure and function. J. Soil Water Conserv. 72, 361–373. doi: 10.2489/jswc.72.4.361
Finney, D. M., White, C. M., and Kaye, J. P. (2016). Biomass production and carbon/nitrogen ratio influence ecosystem services from cover crop mixtures. Agron. J. 108, 39–52. doi: 10.2134/agronj15.0182
Forge, T., Neilsen, G., Neilsen, D., Hogue, E., and Faubion, D. (2013). Composted dairy manure and alfalfa hay mulch affect soil ecology and early production of ‘Braeburn’ apple on M. 9 rootstock. HortScience 48, 645–651. doi: 10.21273/HORTSCI.48.5.645
FOSS analytical solutions for food quality improvement and control. (n.d.). Available online at: https://www.fossanalytics.com/en/ (Accessed November 13, 2024)
Fox, J., and Weisberg, S. (2018). An R companion to applied regression. Thousand Oaks, CA: Sage Publications.
Franco, J. G., Berti, M. T., Grabber, J. H., Hendrickson, J. R., Nieman, C. C., Pinto, P., et al. (2021). Ecological intensification of food production by integrating forages. Agronomy 11:2580. doi: 10.3390/agronomy11122580
Franzluebbers, A. J., and Martin, G. (2022). Farming with forages can reconnect crop and livestock operations to enhance circularity and foster ecosystem services. Grass Forage Sci. 77, 270–281. doi: 10.1111/gfs.12592
Gaudaré, U., Kuhnert, M., Smith, P., Martin, M., Barbieri, P., Pellerin, S., et al. (2023). Soil organic carbon stocks potentially at risk of decline with organic farming expansion. Nat. Clim. Chang. 13, 719–725. doi: 10.1038/s41558-023-01721-5
Goh, K., Pearson, D., and Daly, M. (2001). Effects of apple orchard production systems on some important soil physical, chemical and biological quality parameters. Biol. Agric. Horticult. 18, 269–292. doi: 10.1080/01448765.2001.9754889
Haney, R. L., Brinton, W., and Evans, E. (2008). Estimating soil carbon, nitrogen, and phosphorus mineralization from short-term carbon dioxide respiration. Commun. Soil Sci. Plant Anal. 39, 2706–2720. doi: 10.1080/00103620802358862
Haney, R., Haney, E., Hossner, L., and Arnold, J. (2006). Development of a new soil extractant for simultaneous phosphorus, ammonium, and nitrate analysis. Commun. Soil Sci. Plant Anal. 37, 1511–1523. doi: 10.1080/00103620600709977
Haney, R. L., Haney, E. B., Smith, D. R., Harmel, R. D., and White, M. J. (2018). The soil health tool—theory and initial broad-scale application. Appl. Soil Ecol. 125, 162–168. doi: 10.1016/j.apsoil.2017.07.035
Hannam, K. D., Neilsen, G. H., Forge, T. A., Neilsen, D., Losso, I., Jones, M. D., et al. (2016). Irrigation practices, nutrient applications, and mulches affect soil nutrient dynamics in a young merlot (Vitis vinifera L.) vineyard. Can. J. Soil Sci. 96, 23–36. doi: 10.1139/cjss-2014-0118
Haring, S. C., and Hanson, B. D. (2022). Agronomic cover crop management supports weed suppression and competition in California orchards. Weed Sci. 70, 595–602. doi: 10.1017/wsc.2022.48
Hartmann, M., and Six, J. (2022). Soil structure and microbiome functions in agroecosystems. Nat. Rev. Earth Environ. 4, 4–18. doi: 10.1038/s43017-022-00366-w
Haruna, S. I., Anderson, S. H., Nkongolo, N. V., and Zaibon, S. (2018). Soil hydraulic properties: influence of tillage and cover crops. Pedosphere 28, 430–442. doi: 10.1016/S1002-0160(17)60387-4
Hodson, A. K., Sayre, J. M., Lyra, M. C., and Rodrigues, J. L. M. (2021). Influence of recycled waste compost on soil food webs, nutrient cycling and tree growth in a young almond orchard. Agronomy 11:1745. doi: 10.3390/agronomy11091745
IPCC (2021). “Summary for policymakers”, in Climate change 2021: The physical science basis. Contribution of working group I to the sixth assessment report of the intergovernmental panel on climate change. eds. V. Masson-Delmotte, P. Zhai, A. Pirani, S. L. Connors, C. Péan, and S. Berger, et al. Cambridge, United Kingdom and New York, NY, USA: Cambridge University Press, 3–32.
Isbell, F., Adler, P. R., Eisenhauer, N., Fornara, D., Kimmel, K., Kremen, C., et al. (2017). Benefits of increasing plant diversity in sustainable agroecosystems. J. Ecol. 105, 871–879. doi: 10.1111/1365-2745.12789
Jiao, K., Wu, Y., Guo, B., Zhang, L., Li, Y., and Song, P. (2023). Regulation of microbial community structure, function, and nutrient of apple orchard soil by interplanting red clover. Front. Sustain. Food Syst. 7:1197624. doi: 10.3389/fsufs.2023.1197624
Kabir, Z. (2005). Tillage or no-tillage: impact on mycorrhizae. Can. J. Plant Sci. 85, 23–29. doi: 10.4141/P03-160
Kallenbach, C. M., Frey, S. D., and Grandy, A. S. (2016). Direct evidence for microbial-derived soil organic matter formation and its ecophysiological controls. Nat. Commun. 7, 1–10. doi: 10.1038/ncomms13630
Kaspar, T., and Singer, J. (2011). “The use of cover crops to manage soil” in Soil management: building a stable base for agriculture. eds. J. L. Hatfield and T. J. Sauer (John Wiley & Sons, Inc.), 321–337.
Kassambara, A. (2023). ggpubr: ‘ggplot2’ Based Publication Ready Plots. R package version 0.6.0. Available at: https://rpkgs.datanovia.com/ggpubr/.
Khalsa, S. D. S., and Brown, P. H. (2017). Grower analysis of organic matter amendments in California orchards. J. Environ. Qual. 46, 649–658. doi: 10.2134/jeq2016.11.0456
Khalsa, S. D. S., Hart, S. C., and Brown, P. H. (2022). Nutrient dynamics from surface-applied organic matter amendments on no-till orchard soil. Soil Use Manag. 38, 649–662. doi: 10.1111/sum.12744
Lal, R. (2010). Beyond Copenhagen: mitigating climate change and achieving food security through soil carbon sequestration. Food Secur. 2, 169–177. doi: 10.1007/s12571-010-0060-9
Lange, M., Eisenhauer, N., Sierra, C. A., Bessler, H., Engels, C., Griffiths, R. I., et al. (2015). Plant diversity increases soil microbial activity and soil carbon storage. Nat. Commun. 6:6707. doi: 10.1038/ncomms7707
Lepsch, H. C., Brown, P. H., Peterson, C. A., Gaudin, A. C., and Khalsa, S. D. S. (2019). Impact of organic matter amendments on soil and tree water status in a California orchard. Agric. Water Manag. 222, 204–212. doi: 10.1016/j.agwat.2019.06.002
Li, Y., Li, Z., Cui, S., and Zhang, Q. (2020). Trade-off between soil pH, bulk density and other soil physical properties under global no-tillage agriculture. Geoderma 361:114099. doi: 10.1016/j.geoderma.2019.114099
Li, M., Peterson, C. A., Tautges, N. E., Scow, K. M., and Gaudin, A. C. (2019). Yields and resilience outcomes of organic, cover crop, and conventional practices in a Mediterranean climate. Sci. Rep. 9:12283. doi: 10.1038/s41598-019-48747-4
López, R., Burgos, P., Hermoso, J. M., Hormaza, J. I., and González-Fernández, J. J. (2014). Long term changes in soil properties and enzyme activities after almond shell mulching in avocado organic production. Soil Tillage Res. 143, 155–163. doi: 10.1016/j.still.2014.06.004
Lordan Sanahuja, J., Pascual Roca, M., Villar Mir, J. M., Fonseca Salcedo, F., Papió, J., Montilla, V., et al. (2015). Use of organic mulch to enhance water-use efficiency and peach production under limiting soil conditions in a three-year-old orchard. Spanish J. Agric. Res. 13:e0904. doi: 10.5424/sjar/2015134-6694
Martínez-Mena, M., Carrillo-López, E., Boix-Fayos, C., Almagro, M., Franco, N. G., Díaz-Pereira, E., et al. (2020). Long-term effectiveness of sustainable land management practices to control runoff, soil erosion, and nutrient loss and the role of rainfall intensity in Mediterranean rainfed agroecosystems. Catena 187:104352. doi: 10.1016/j.catena.2019.104352
Mays, N., Brye, K., Rom, C. R., Savin, M., and Garcia, M. (2014). Groundcover management and nutrient source effects on soil carbon and nitrogen sequestration in an organically managed apple orchard in the Ozark highlands. HortScience 49, 637–644. doi: 10.21273/HORTSCI.49.5.637
McClelland, S. C., Paustian, K., and Schipanski, M. E. (2021). Management of cover crops in temperate climates influences soil organic carbon stocks: a meta-analysis. Ecol. Appl. 31:e02278. doi: 10.1002/eap.2278
McKenzie, S. C., Goosey, H. B., O’Neill, K. M., and Menalled, F. D. (2017). Integration of sheep grazing for cover crop termination into market gardens: agronomic consequences of an ecologically based management strategy. Renew. Agric. Food Syst. 32, 389–402. doi: 10.1017/S1742170516000326
Mikha, M. M., and Rice, C. W. (2004). Tillage and manure effects on soil and aggregate-associated carbon and nitrogen. Soil Sci. Soc. Am. J. 68, 809–816. doi: 10.2136/sssaj2004.8090
Miller, R. O., Gavlak, R., and Horneck, D. (2013). Soil, plant and water reference methods for the western region. Fort Collins, CO, USA: Colorado State University, 155.
Moreno, B., Garcia-Rodriguez, S., Cañizares, R., Castro, J., and Benítez, E. (2009). Rainfed olive farming in South-Eastern Spain: long-term effect of soil management on biological indicators of soil quality. Agric. Ecosyst. Environ. 131, 333–339. doi: 10.1016/j.agee.2009.02.011
Moukanni, N., Brewer, K. M., Gaudin, A. C., and O’geen, A. T. (2022). Optimizing carbon sequestration through cover cropping in Mediterranean agroecosystems: synthesis of mechanisms and implications for management. Front. Agron. 4:844166. doi: 10.3389/fagro.2022.844166
Mozea, K. (2022). Biomass Yield and Nutritive Value of Annual Forage Mixes Compared with Monocrops (Master’s thesis, North Dakota State University).
Muhammad, I., Wang, J., Sainju, U. M., Zhang, S., Zhao, F., and Khan, A. (2021). Cover cropping enhances soil microbial biomass and affects microbial community structure: a meta-analysis. Geoderma 381:114696. doi: 10.1016/j.geoderma.2020.114696
Mujdeci, N. (2011). The effects of organic material applications on soil penetration resistance. J. Food Agric. Environ. 9, 1045–1047.
Neilsen, G., Forge, T., Angers, D., Neilsen, D., and Hogue, E. (2014). Suitable orchard floor management strategies in organic apple orchards that augment soil organic matter and maintain tree performance. Plant Soil 378, 325–335. doi: 10.1007/s11104-014-2034-8
NRCS, U. (2008). Soil quality indicators: bulk density. USA: Service UNRC. USDA Natural Resources Conservation Service.
NRCS, U. (2018). Estimating cover crop biomass. Des Moines, Iowa, USA: USDA, Natural Resources Conservation Service.
NRCS, USDA, S.S.S. (2024). Web soil survey. Available online at: https://websoilsurvey.nrcs.usda.gov/app/WebSoilSurvey.aspx (Accessed October 4, 2024).
Oliveira, M. T., and Merwin, I. A. (2001). Soil physical conditions in a New York orchard after eight years under different groundcover management systems. Plant Soil 234, 233–237. doi: 10.1023/A:1017992810626
Opoku, A., Ogunleye, A. M., Solomon, J. K., and Payne, W. A. (2024). Cover crop systems impact on biomass production, carbon-to-nitrogen ratio, forage quality, and soil health in a semi-arid environment. Heliyon 10. doi: 10.1016/j.heliyon.2024.e39600
Özbolat, O., Sánchez-Navarro, V., Zornoza, R., Egea-Cortines, M., Cuartero, J., Ros, M., et al. (2023). Long-term adoption of reduced tillage and green manure improves soil physicochemical properties and increases the abundance of beneficial bacteria in a Mediterranean rainfed almond orchard. Geoderma 429:116218. doi: 10.1016/j.geoderma.2022.116218
Paustian, K., Lehmann, J., Ogle, S., Reay, D., Robertson, G. P., and Smith, P. (2016). Climate-smart soils. Nature 532, 49–57. doi: 10.1038/nature17174
Peck, G. M., Merwin, I. A., Thies, J. E., Schindelbeck, R. R., and Brown, M. G. (2011). Soil properties change during the transition to integrated and organic apple production in a New York orchard. Appl. Soil Ecol. 48, 18–30. doi: 10.1016/j.apsoil.2011.02.008
Pérez-Murcia, M. D., Bustamante, M. Á., Orden, L., Rubio, R., Agulló, E., Carbonell-Barrachina, Á. A., et al. (2021). Use of agri-food composts in almond organic production: effects on soil and fruit quality. Agronomy 11:536. doi: 10.3390/agronomy11030536
Peterson, C. A., Deiss, L., and Gaudin, A. C. (2020). Commercial integrated crop-livestock systems achieve comparable crop yields to specialized production systems: a meta-analysis. PLoS One 15:e0231840. doi: 10.1371/journal.pone.0231840
Poeplau, C., and Don, A. (2015). Carbon sequestration in agricultural soils via cultivation of cover crops–a meta-analysis. Agric. Ecosyst. Environ. 200, 33–41. doi: 10.1016/j.agee.2014.10.024
Pommier, T. (2023). Diverse organic carbon activates soil microbiome functioning via niche modulation. Soil Ecol. Lett. 5:230180. doi: 10.1007/s42832-023-0180-8
Prommer, J., Walker, T. W., Wanek, W., Braun, J., Zezula, D., Hu, Y., et al. (2020). Increased microbial growth, biomass, and turnover drive soil organic carbon accumulation at higher plant diversity. Glob. Chang. Biol. 26, 669–681. doi: 10.1111/gcb.14777
R: The R Project for Statistical Computing. (n.d.). Available online at: https://www.r-project.org/ (Accessed November 13, 2024).
Rad, S. M., Ray, A. K., and Barghi, S. (2022). Water pollution and agriculture pesticide. Clean Technol. 4, 1088–1102. doi: 10.3390/cleantechnol4040066
Ramos, M., Altieri, M., Garcia, P., and Robles, A. (2011). Oat and oat-vetch as rainfed fodder-cover crops in semiarid environments: effects of fertilization and harvest time on forage yield and quality. J. Sustain. Agric. 35, 726–744. doi: 10.1080/10440046.2011.606490
Repullo-Ruiberriz de Torres, M. A., Moreno-García, M., Ordóñez-Fernández, R., Rodríguez-Lizana, A., Carceles Rodriguez, B., García-Tejero, I. F., et al. (2021). Cover crop contributions to improve the soil nitrogen and carbon sequestration in almond orchards (SW Spain). Agronomy 11:387. doi: 10.3390/agronomy11020387
Rodriguez-Ramos, J. C., Scott, N., Marty, J., Kaiser, D., and Hale, L. (2022). Cover crops enhance resource availability for soil microorganisms in a pecan orchard. Agric. Ecosyst. Environ. 337:108049. doi: 10.1016/j.agee.2022.108049
Sastre, B., Álvarez, B., Antón, O., Pérez, M. Á., Marques, M. J., Bienes, R., et al. (2020). Groundcovers in olive groves in semiarid climates: are they always beneficial? Water 12:2230. doi: 10.3390/w12082230
Sharaf, H., Thompson, A. A., Williams, M. A., and Peck, G. M. (2021). Compost applications increase bacterial community diversity in the apple rhizosphere. Soil Sci. Soc. Am. J. 85, 1105–1121. doi: 10.1002/saj2.20251
Sivaranjani, S., and Rakshit, A. (2019). “Organic farming in protecting water quality” in Organic farming. eds. C. Sarath Chandran, S. Thomas, and M. R. Unni (Cham: Springer International Publishing), 1–9.
Sofo, A., Palese, A. M., Casacchia, T., Celano, G., Ricciuti, P., Curci, M., et al. (2010). Genetic, functional, and metabolic responses of soil microbiota in a sustainable olive orchard. Soil Sci. 175, 81–88. doi: 10.1097/SS.0b013e3181ce8a27
Soto, R. L., Martinez-Mena, M., Padilla, M. C., and de Vente, J. (2021). Restoring soil quality of woody agroecosystems in Mediterranean drylands through regenerative agriculture. Agric. Ecosyst. Environ. 306:107191. doi: 10.1016/j.agee.2020.107191
Thompson, A. A., and Peck, G. M. (2017). Alternative fertility management for establishing new apple orchards in the mid-Atlantic. HortScience 52, 1313–1319. doi: 10.21273/HORTSCI11916-17
Thompson, A. A., Williams, M. A., and Peck, G. M. (2019). Compost and Geneva® series rootstocks increase young ‘Gala’ apple tree growth and change root-zone microbial communities. Sci. Hortic. 256:108573. doi: 10.1016/j.scienta.2019.108573
Tilman, D., Wedin, D., and Knops, J. (1996). Productivity and sustainability influenced by biodiversity in grassland ecosystems. Nature 379, 718–720. doi: 10.1038/379718a0
Urra, J., Alkorta, I., and Garbisu, C. (2019). Potential benefits and risks for soil health derived from the use of organic amendments in agriculture. Agronomy 9:542. doi: 10.3390/agronomy9090542
US Department of Commerce, (n. d.). National Oceanic and Atmospheric Administration National Weather Service. Available online at: https://www.weather.gov/wrh/Climate?wfo=sto (Accessed January 31, 2025).
Vasilikiotis, C., Li, M., Schmidt, J. E., Azimi, A., Garcia, J., Volder, A., et al. (2020). Orchard management practices affect arbuscular mycorrhizal fungal root colonisation of almond. Biol. Agric. Horticult. 36, 230–248. doi: 10.1080/01448765.2020.1802777
Vicente-Vicente, J. L., García-Ruiz, R., Francaviglia, R., Aguilera, E., and Smith, P. (2016). Soil carbon sequestration rates under Mediterranean woody crops using recommended management practices: a meta-analysis. Agric. Ecosyst. Environ. 235, 204–214. doi: 10.1016/j.agee.2016.10.024
Villa, Y. B., Khalsa, S. D. S., Ryals, R., Duncan, R. A., Brown, P. H., and Hart, S. C. (2021). Organic matter amendments improve soil fertility in almond orchards of contrasting soil texture. Nutr. Cycl. Agroecosyst. 120, 343–361. doi: 10.1007/s10705-021-10154-5
Wang, Y., Liu, L., Yang, J., Duan, Y., Luo, Y., Taherzadeh, M. J., et al. (2020). The diversity of microbial community and function varied in response to different agricultural residues composting. Sci. Total Environ. 715:136983. doi: 10.1016/j.scitotenv.2020.136983
Wauters, V. M., Jarvis-Shean, K., Williams, N., Hodson, A., Hanson, B. D., Haring, S., et al. (2023). Developing cover crop systems for California almonds: current knowledge and uncertainties. J. Soil Water Conserv. 78, 5A–11A. doi: 10.2489/jswc.2023.1109A
Weisser, W. W., Roscher, C., Meyer, S. T., Ebeling, A., Luo, G., Allan, E., et al. (2017). Biodiversity effects on ecosystem functioning in a 15-year grassland experiment: patterns, mechanisms, and open questions. Basic Appl. Ecol. 23, 1–73. doi: 10.1016/j.baae.2017.06.002
White, D. C. (1988). Validation of quantitative analysis for microbial biomass, community structure, and metabolic activity. Adv. Limnol. 31.
Wickham, H., François, R., Henry, L., Müller, K., and Vaughan, D. (2022). Dplyr: a grammar of data manipulation. 2020 [online]. Boston, MA: Posit Software, PBC, 1–18. Available at: https://cran.r-project.org/web/packages/dplyr/index.html
Wilson, H., Daane, K. M., Maccaro, J. J., Scheibner, R. S., Britt, K. E., and Gaudin, A. C. (2022). Winter cover crops reduce spring emergence and egg deposition of overwintering navel Orangeworm (Lepidoptera: Pyralidae) in almonds. Environ. Entomol. 51, 790–797. doi: 10.1093/ee/nvac051
Wortman, S. E., Francis, C., and Lindquist, J. L. (2012). Cover crop mixtures for the western Corn Belt: opportunities for increased productivity and stability. Agron. J. 104, 699–705. doi: 10.2134/agronj2011.0422
Zelles, L., and Bai, Q. (1993). Fractionation of fatty acids derived from soil lipids by solid phase extraction and their quantitative analysis by GC-MS. Soil Biol. Biochem. 25, 495–507. doi: 10.1016/0038-0717(93)90075-M
Keywords: multispecies cover crop, compost, almond, regenerative agriculture, soil health, climate-smart agriculture, organic, whole-system research
Citation: Krach RJ, Daley CA and Liles GC (2025) Climate smart management practices add value to mature organic almond production system. Front. Sustain. Food Syst. 9:1527898. doi: 10.3389/fsufs.2025.1527898
Edited by:
Gaetano Alessandro Vivaldi, University of Bari Aldo Moro, ItalyReviewed by:
Megahed Mohamed Amer, Agricultural Research Center, EgyptSubhradip Bhattacharjee, Central Coastal Agricultural Research Institute (ICAR), India
Copyright © 2025 Krach, Daley and Liles. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Raquel J. Krach, cmtyYWNoQGNzdWNoaWNvLmVkdQ==