 Rao Muhammad Samran Gul1
Rao Muhammad Samran Gul1 Saeed Rauf
Saeed Rauf Rodomiro Ortiz
Rodomiro Ortiz
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Sustain. Food Syst. , 28 October 2024
Sec. Crop Biology and Sustainability
Volume 8 - 2024 | https://doi.org/10.3389/fsufs.2024.1477994
Introduction: Rice is a vital staple food for many countries, and its yield is known to be significantly affected by various abiotic stresses, which are expected to intensify with climate change, posing a threat to global food security. Abscisic acid (ABA), a crucial plant growth regulator, plays a crucial role in plant responses to these abiotic stresses. It influences several processes, such as seed dormancy, leaf gas exchange, reactive oxygen species (ROS) scavenging, ion toxicity reduction, and root elongation, all of which contribute to enhancing plant survival under stress.
Methods: This article reviews recent research on ABA-mediated gene responses and expressions involved in rice plant architecture and its response to abiotic stress.
Results and discussion: Abscisic acid responses were primarily driven by changes in gene expression. Expression analyses of the gene related to ABA biosynthesis or catabolism indicated several changes in plant architecture, such as changes in leaf angle, delayed flowering, and modifications in growth regulators. Additionally, tolerance-related mechanisms, such as increased ROS scavenging, reduced membrane leakage, and vacuolar compartmentation of toxic radicals, were activated under single or multiple stress conditions. While these adaptations may improve plant survival and yield sustainability under stress, they may not necessarily enhance yield potential in environments affected by drought, salinity, or heat stress. ABA expression was also associated with improved pollen viability, grain-filling potential, and seed setting under abiotic stresses such as heat, which could enhance seed yield in such challenging environments.
Crop species are often exposed to various types of abiotic stresses, such as extreme temperatures, soil salinity, UV light, hypoxia, metal toxicity, and drought. Among these, drought, salinity, and heat are the most widespread globally, affecting large areas (Figure 1). These stresses significantly reduce crop yields by causing harmful effects such as the production of reactive oxygen species (ROS), disruption of electron transport chains, and the disintegration of cellular membranes due to electrolyte leakage (Rauf et al., 2010; Hirayama and Umezawa, 2010).

Figure 1. Impact of various stresses on plants and cellular responses.
Physiological responses include the deterioration of leaf gas exchange traits, measured by stomatal conductance, photosynthesis, and transpiration rates. In response to these stresses, plants activate ROS scavengers and produce osmoprotectants such as prolines, trehalose, glycine betaine, and heat shock proteins. These osmoprotectants not only maintain osmotic potential but also help preserve the integrity of cellular structures. Plant defense responses to stress are strongly linked to stress signaling and the activation of various stress-responsive genes. These genes are responsible for transcribing antioxidant enzymes, such as superoxide dismutase (SOD), catalase (CAT), and ascorbate peroxidase (APX), via the mitogen-activated protein kinases (MAPKs) signaling pathway.
Global climate change (CC) poses a significant threat to the production potential of various crops, including rice, in various regions of the world, particularly in South Asia. The rising global temperatures have increased the risk of famine due to food shortages and surging food prices in South Asia. Heat waves and the rapid melting of glaciers, which have led to catastrophic floods, have destroyed standing rice crops. In the 2022 cropping season alone, 10 million people in Pakistan’s rice-growing areas were affected by the impacts of climate change. CC has introduced the risk of multiple simultaneous (Sinha et al., 2024), challenging rice production with issues such as UV radiations, weed pressure, supra-optimal temperatures, salinity, high transpiration rates (leading to higher water demands), reduced efficiency of fertilizers due to rapid leaching, and nitrogen loss from volatilization under extreme temperatures of 45–50°C (Hasanuzzaman et al., 2023). Additionally, high temperatures have exacerbated the infestation of insect pests, as well as bacterial and fungal pathogens.
Abscisic acid (ABA) is a common signal produced in response to various abiotic stresses such as extreme temperature, nutrient deficiency, UV radiation, drought, and salinity (Yoshida et al., 2021; Aslam et al., 2022).
Abscisic acid signals, which are generated at various growth stages of plants, modulate various functions such as seed dormancy, embryo morphogenesis, stomatal closure, and the upregulation of stress-responsive genes (Yoshida et al., 2021; Mizokami et al., 2019). ABA is known to improve stress adaptability and the survival of crop species.
Abscisic acid overexpression is associated with a reduced leaf expansion rate, the overaccumulation of osmolytes such as proline (Pro), and an increase in lateral root length in model species such as maize (Sharp et al., 2004) and Arabidopsis (Van Houtte et al., 2013).
An ABA-deficient mutant of Arabidopsis thaliana experienced more wilting than the wild-type plant (Ozfidan et al., 2013). The same study revealed that the exogenous application of ABA maintained relative water content but reduced stomatal conductance, while the exogenous application of ABA under non-stressed conditions produced stress-like symptoms (Ozfidan et al., 2013) (Figures 2, 3).
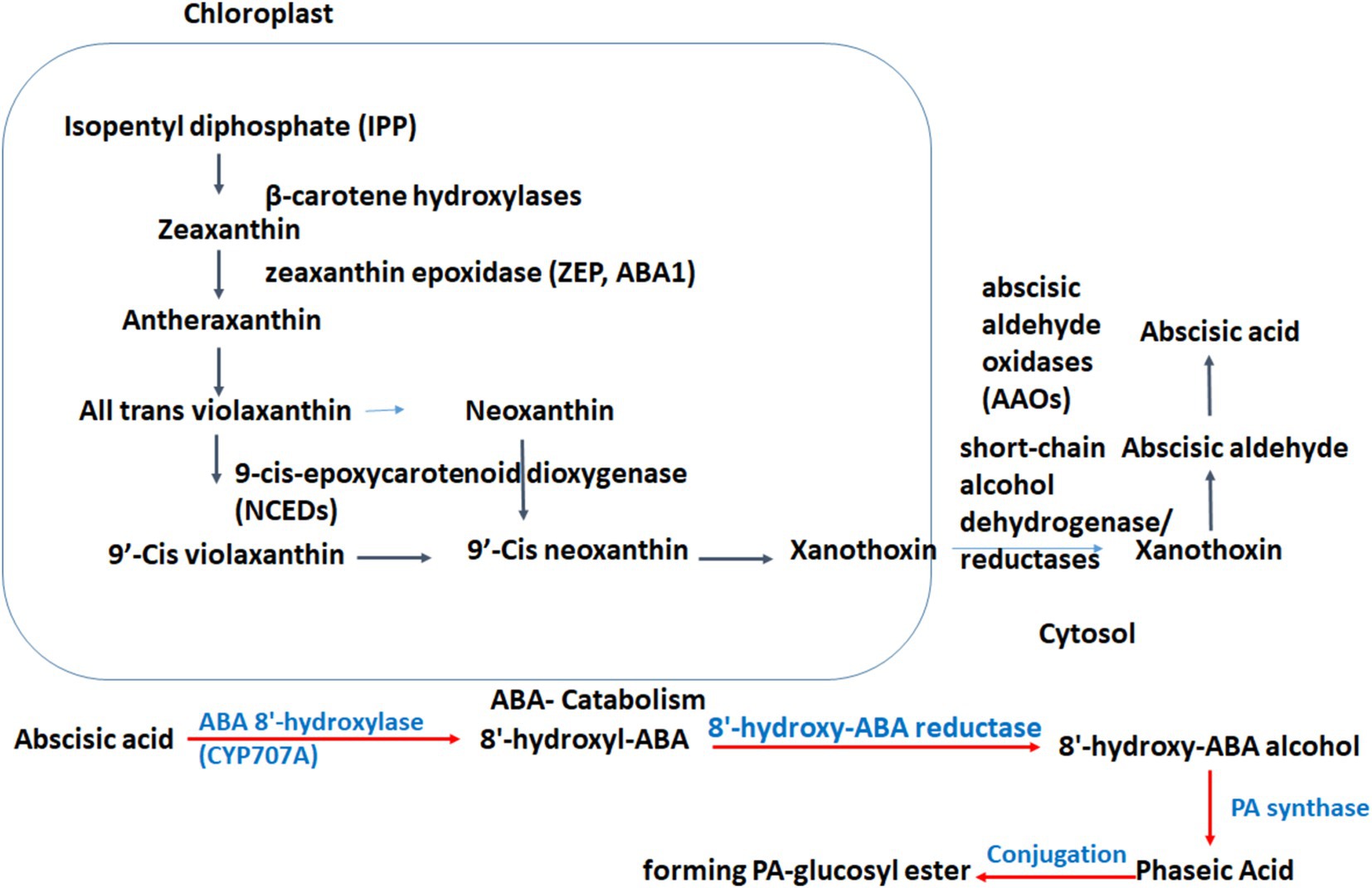
Figure 2. Abscisic acid biosynthesis pathways in crop species.
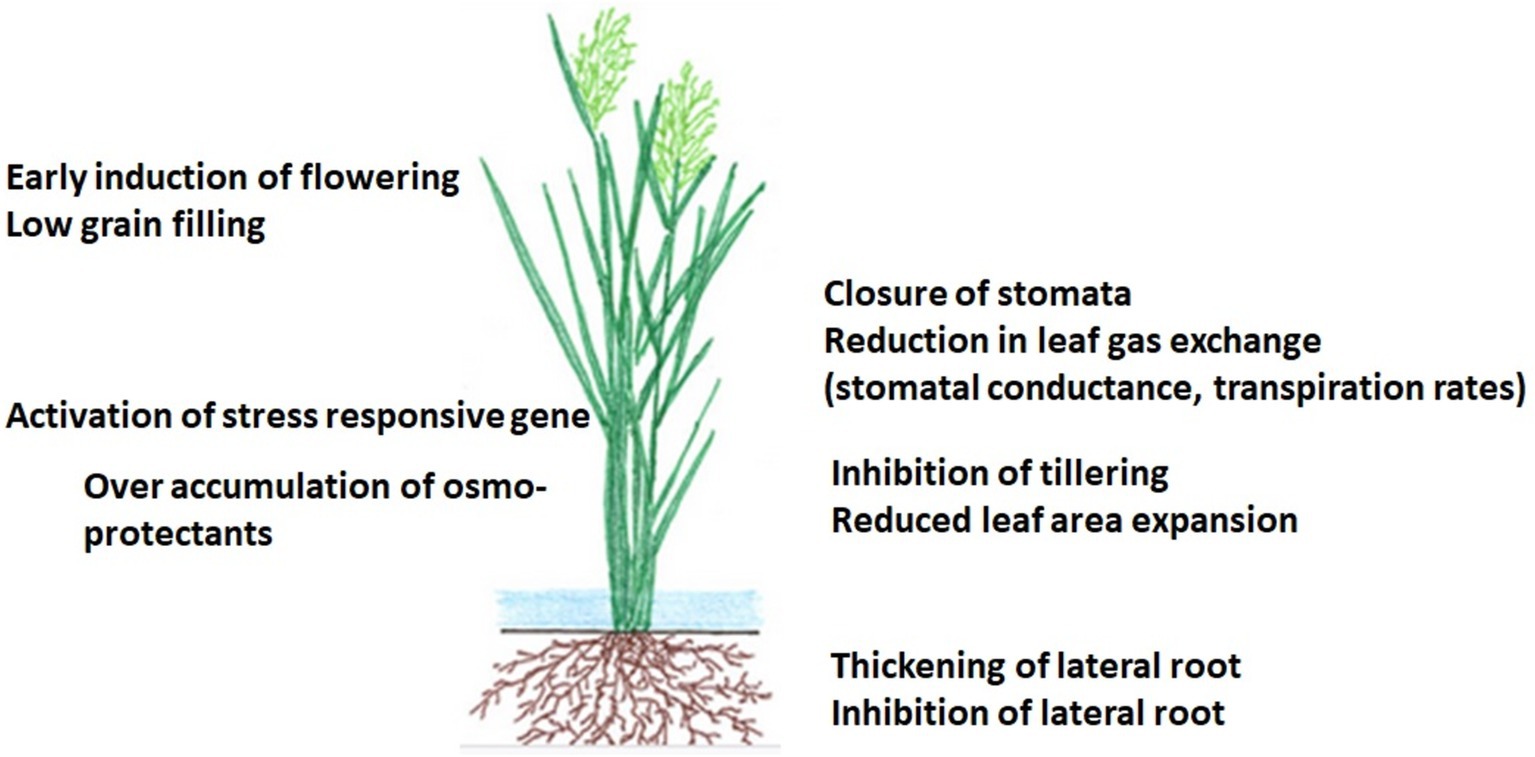
Figure 3. ABA-mediated plant signaling and responses.
Stress significantly affects plant development by modifying the expression of genes involved in normal growth and development.
Stress conditions prioritize root growth, stomata closure, activation of reactive oxygen species scavenger enzymes, overaccumulation of compatible solutes, and the production of stress-related proteins, such as chaperones or dehydrins (Figure 1). Stress responses in rice plants occur at the expense of growth, development, and production.
Genes affecting plant growth have been identified through targeted mutations induced by various molecular methods, such as RNAi or clustered regularly interspaced palindromic region (CRISPR).
The Pyrabactin resistance-like (PYL) abscisic acid receptor family has been characterized and is known to induce ABA signaling in response to abiotic stress in Arabidopsis and other plant species, including rice (Tian et al., 2015).
Ten orthologs of PYL have been isolated in rice, and overexpression of the rice PYL orthologs (Os PYL3 and Os PYL9) positively regulated ABA expression and enhanced cold and drought tolerance in rice (Tian et al., 2015).
CRISPR/Cas9-generated mutant lines demonstrated that PYL orthologs control stomatal movement, seed dormancy, and growth regulation (Miao et al., 2018). A mutant line for pyl1/4/6 exhibited increased growth and productivity in rice paddy fields while maintaining seed dormancy (Miao et al., 2018).
Abscisic acid biosynthesis antagonizes cytokinin- and auxin-related factors, which may affect normal plant architecture and development. OsNF-YA3 is a negative regulator of ABA signaling by regulating the expression of ABA catabolic genes (i.e., OsABA8ox1 and OsABA8ox3) by binding to their promoters (Jin et al., 2023). It positively upregulates gibberellic acid biosynthesis by activating the GA biosynthesis gene (OsGA20ox1). Thus, it balances the biosynthesis of GA, the promotion of growth, and the negative regulation of ABA biosynthesis under non-stress conditions (Jin et al., 2023). SAPK9m, which is a positive regulator of ABA biosynthesis, is activated under osmotic stress and phosphorylates OsNF-YA3, thereby causing its degradation under stress conditions (Jin et al., 2023) (Figure 4).
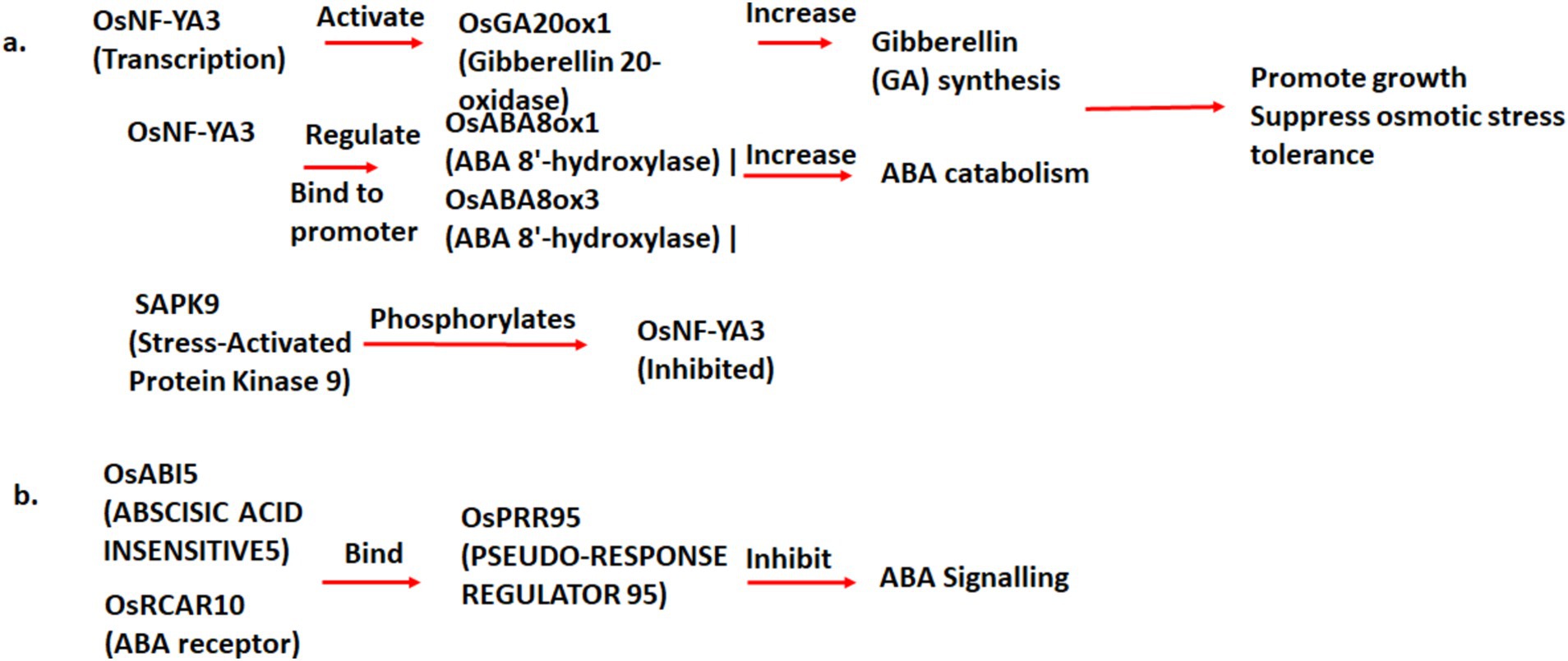
Figure 4. Conceptual illustration of inactivation of ABA signal pathways led to the increase of plant growth regulators and loss of osmotic stress tolerance.
OsPRR95 (PSEUDO-RESPONSE REGULATOR 95) is an inhibitor of ABA signaling and is associated with seed germination, seedling growth, and breaking seed dormancy. It binds to the ABA receptor gene (OsRCAR10) DNA to inhibit its expression (Wang et al., 2023). OsRCAR10 acts downstream of OsPRR95 in mediating ABA responses. OsPRR95 expression required a functional OsRCAR10 and the ABA-responsive element-binding factor ABSCISIC ACID INSENSITIVE5 (OsABI5), which bound directly to the promoter of OsPRR95 (Wang et al., 2023). OsGA20ox2 requires ABA-responsive proteins ASR5 and ABA receptor PLY5 for the upregulation, and it increases the plant height and gibberellic acid contents under non-stress conditions (Nawaz et al., 2020).
NAC054 activates OsABI5 and NYC1 by binding to the mitochondrial dysfunction motifs in their promoters. The activity of OsNAC054 is regulated by proteolytic processing of the C-terminal transmembrane domain (TMD).
The onaco54 mutant showed downregulation of ABA INSENSITIVE5 (OsABI5) and senescence-associated genes, including STAY-GREEN and NON-YELLOW COLORING1 (NYC1). Lines overexpressing OsNAC054 exhibited early leaf yellowing under dark- and ABA-induced senescence conditions (Sakuraba et al., 2020). Additionally, the OsGA20ox2 gene was mutated using a guided gRNA sequence. The CRISPR-Cas9-mediated mutagenic line showed the upregulation of abscisic stress-ripening protein 5 (ASR5) and abscisic acid receptor (PYL5). This suppression leads to an increase in gibberellins (GA1 and GA4) levels, plant height (28.72%), and flag leaf length. However, other morphological traits remained unchanged (Nawaz et al., 2020).
Several genes related to leaf senescence or staying green in rice have been induced by ABA signals and are identified through expression analyses or the development of mutants through CRISPR/Cas9 (Table 1). OsNAP is a marker of leaf senescence in rice and is induced by the ABA. It affects chlorophyll degradation and nutrient transport in rice (Liang et al., 2014). The mechanism of leaf senescence has been investigated in the psf rice mutant.
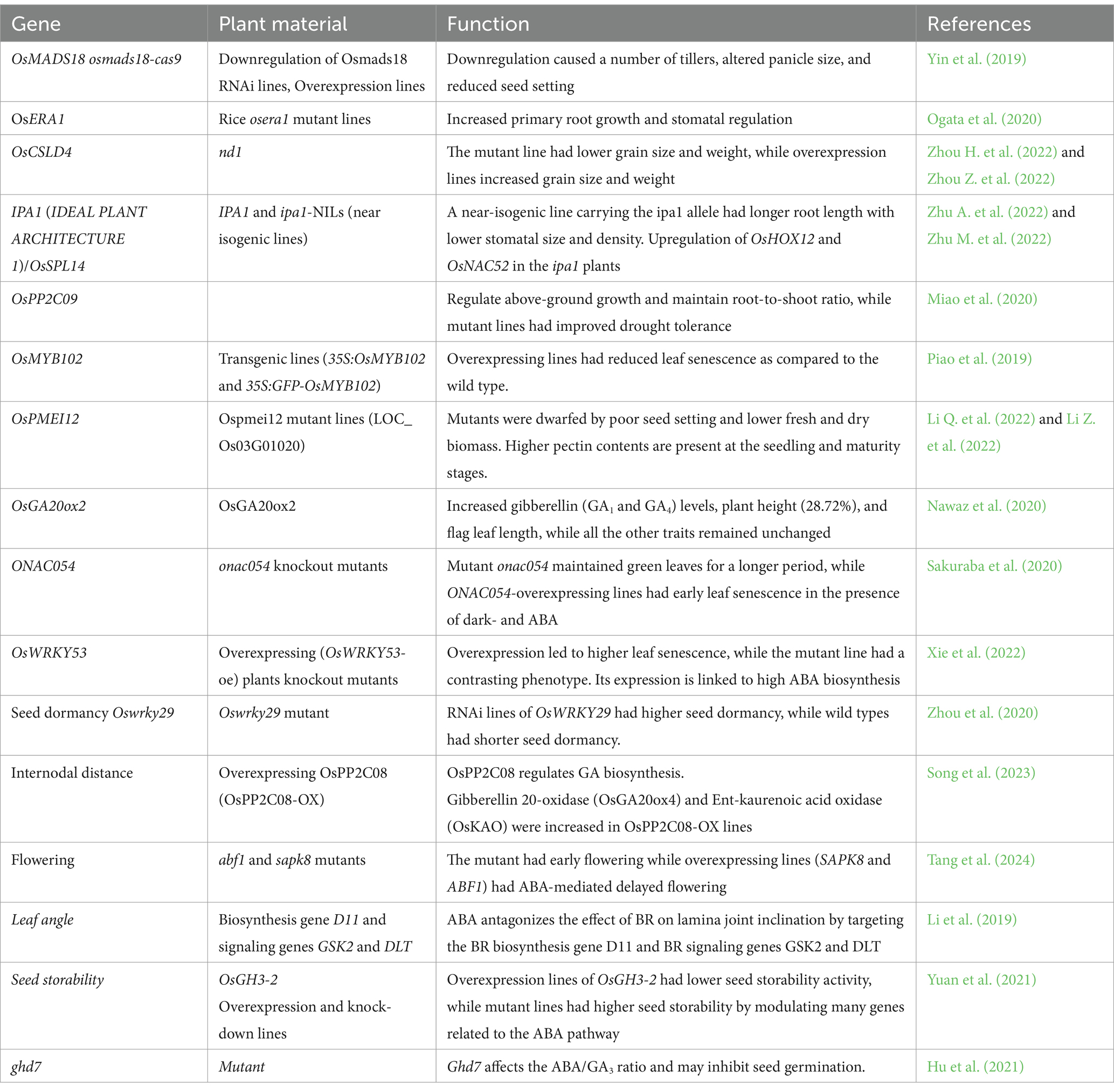
Table 1. Gene related to plant architecture affected by ABA signaling.
Sugar starvation is known to increase ABA concentration and induce premature leaf senescence in rice leaves (Asad et al., 2021).
Knocking down the OsNAP gene in the PS1-D mutant reduced leaf senescence and extended the grain-filling period, resulting in a rice yield increase of 6.3 and 10.3% in two independent RNAi-induced mutant lines (Liang et al., 2014).
The OSMADs-18 gene is known to control plant architecture, unlike its paralogs, OSMADs-14 and OSMADs-15, which induce flowering in rice. Expression of OSMAS-18 was stimulated by abscisic acid, which helps its translocation from the plasma membrane to the nucleus.
A mutant line induced by CRISPR/Cas9 in the OsMADS18 gene increased the yield component of rice, such as the number of tillers plant−1 and altered panicle size but reduced seed setting, while overexpression lines produced fewer tillers (Yin et al., 2019). Overexpression lines (nd1) for cell wall cellulose synthase-like D4 protein (OsCSLD4) had increased grain weight and width in rice (Zhou H. et al., 2022; Zhou Z. et al., 2022). The gene OsPP2C09 was rapidly induced by exogenous ABA treatments and positively affects rice growth but is a negative regulator of drought tolerance.
Exogenous application of ABA suppressed excessive ABA signaling. The OsPP2C09 gene may act as a negative regulator of ABA, helping recover from the growth arrest induced by ABA.
OsPP2C09 transcript levels were much higher in the roots than in shoots and rapidly increased with stress treatment (Tian et al., 2015). This differential expression may control the root-to-shoot ratio.
A CRISPR/Cas9-edited line carrying mutants in gn1a, dep1, and gs3 had larger grain size, erect panicles, and a higher number of grains panicle−1 (Khan et al., 2020). Another line carrying CRISPR/Cas9-mediated mutants in dep1 and gs3 produced a phenotype with long awns, large grains, and semi-dwarf plant architecture (Khan et al., 2020).
Overexpressing OsWRKY53-oe lines showed higher leaf senescence, while OsWRKY53 knockout mutants (oswrky53) remained green longer compared to the normal wild type.
OsWRKY53 is a regulatory gene that binds directly to the promoters of OsABA8ox1 and OsABA8ox2, inhibiting their expression and increasing endogenous ABA levels (Figures 5, 6). The genes OsABA8ox1 and OsABA8ox2, which are involved in ABA catabolism, had their ABA 8′-hydroxylase activity reduced by the OsWRKY53-oe (Xie et al., 2022).

Figure 5. Activation of ABA pathway signals by inactivation of ABA catabolism genes to enhance ABA contents.
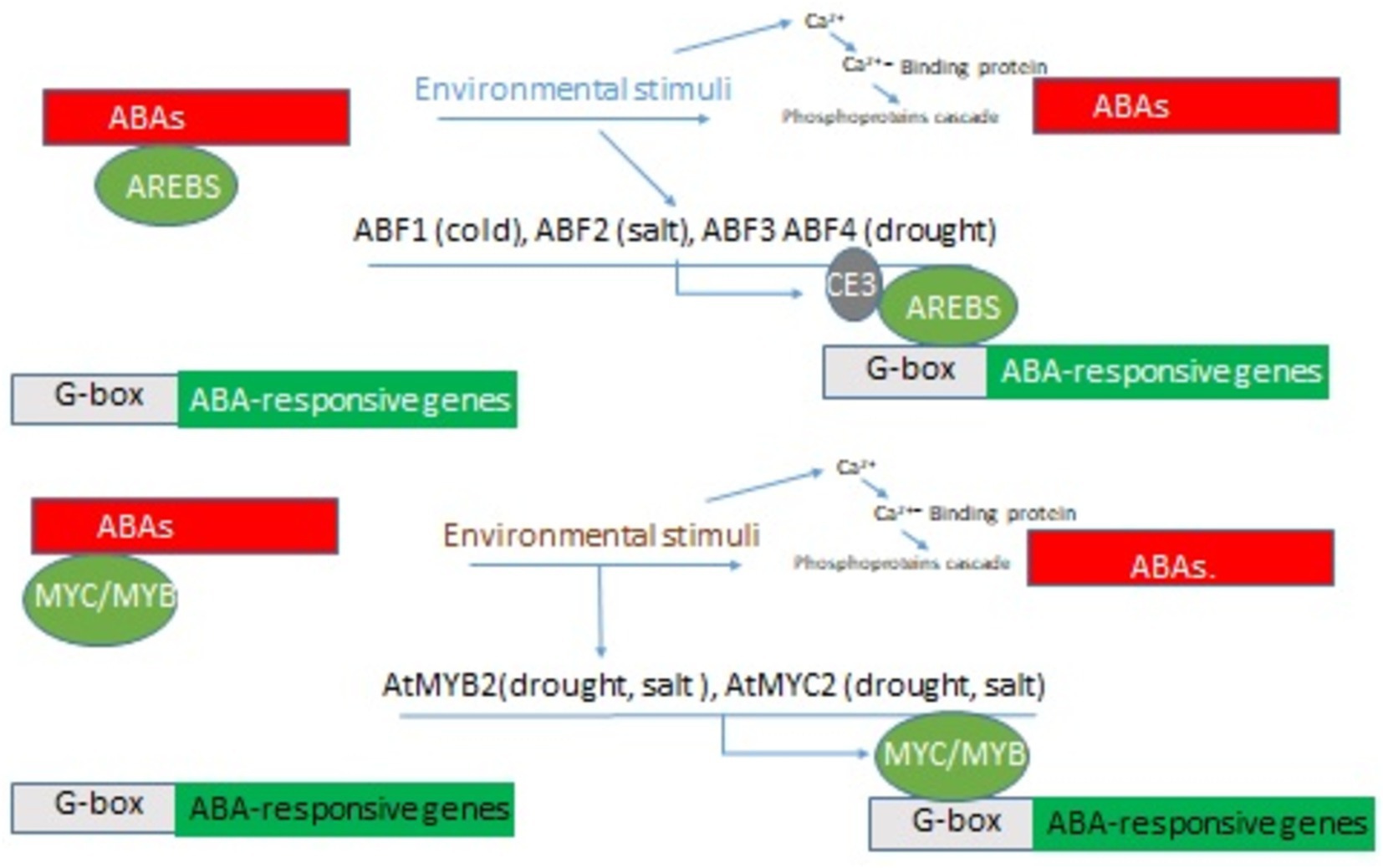
Figure 6. Activation of the ABA-responsive gene under various abiotic stress.
Moreover, Oswrky29 controls seed dormancy in rice. RNAi lines of OsWRKY29 enhanced the seed dormancy, whereas its overexpression lines showed contrasting phenotypes, i.e., lower seed dormancy. OsWRKY29 could bind to the promoters of OsABF1 and OsVP1 to inhibit their expression (Zhou et al., 2020). OsABA8ox encodes abscisic acid hydroxylase and is considered a drought stress-responsive gene. Its expression was decreased in roots due to stress treatment. CRISPR/Cas9 was used to develop a mutant for the osaba8ox. The analyses showed that the mutant had higher ABA contents and large vertical root lengths, while overexpression lines had smaller roots (Zhang et al., 2020).
Overexpression transgenic lines (35S: OsMYB102 and 35S: GFP-OsMYB102) had longer growth cycles, maintaining green leaves longer compared to the wild type. A knockout mutant of osmyb102, when treated with ABA, showed senescence at a higher rate in the dark. Near isogenic lines, i.e., IPA1 (Ideal Plant Architecture 1), controlled the tiller number, and ipa1 was phenotypically characterized. The iso line ipa1 had longer root elongation and a smaller leaf aperture, allowing them to show better survival with reduced leaf senescence under water stress conditions (Zhu A. et al., 2022). IPA1 and OsSPL14 encode a transcription factor that takes part in plant architecture and abiotic stress response by activating the ABA signaling pathway. An ipa-1 isogeneic line significantly improves the abiotic stress at the seedling stage. Ipa1 plants have better root systems and smaller leaf stomatal apertures. Under stress conditions with PEG 8000, ipa1 shows high ABA content upregulation in mutant type as compared to wild APA1, and by promoter edition and yeast two-hybrid protein interaction and dual luciferase assay find some promoter OsHOX12 and OsNAC52 and its activity support the ABA signaling pathway (Zhu A. et al., 2022). Table 1 lists some examples of various genes affected by ABA signaling.
UGT74E2 is involved in the transfer of glucose to the indole butyric acid and controls plant architecture and stress tolerance in Arabidopsis thaliana. Overexpression of UGT74E2 in rice resulted in improved seed germination under multiple treatments and stress conditions (Wang B. et al., 2020; Wang C. et al., 2020; Wang H. et al., 2020; Wang T. et al., 2020). ABA biosynthesis genes, i.e., OsABI3 and OsABI5, were, however, downregulated in overexpression lines (Wang B. et al., 2020; Wang C. et al., 2020; Wang H. et al., 2020; Wang T. et al., 2020).
The rice SnRK2 comprises 10 members from SAPK1-10 (Sn-RK2, protein kinase). These proteins are crucial for the ABA pathways inducing drought and salt tolerance (Liu et al., 2024). Simultaneous knockout of ABF1 and SAPK8 accelerated the flowering, while the overexpression line delayed the flowering. The ABA signal (SAPK8 stress-activated protein kinase) phosphorylates ABF1, which helps the binding of the promoters related to the flowering regulators (Ehd1 and Ehd2) (Tang et al., 2024). Epigenetic suppression of ABF1 by PRC2 (polycomb repressive complex 2) of flowering regulators (Ehd1 and Ehd2) by trimethylation of histone (H3K27me3 suppressive histone modification) leads to the delayed flowering in rice (Tang et al., 2024). SAPKs are an important component of the ABA signal pathways. SAPK1-3 may not have any role in the regulatory pathway of rice heading date, while SAPK4, SAPK5, and SAPK5 were positive regulators of flowering (Liu et al., 2024). The double mutant of sapk9/sapk10 had delayed flowering. Overexpression of SAPK10 resulted in the early flowering of rice. The SAPK10 phosphorylated the bZIP77. It was known that SAPK10 partially regulates the flowering through FAC-MADS15 pathways (Liu X. et al., 2019; Liu X. L. et al., 2019). SAPK2 helps to transport nitrate and assimilation by regulating the specific promoters, and mutant sapk2 had stunted growth and fewer grains (Lou et al., 2020).
When exposed to the water stress condition, several phenotypic changes were noted in the plants. These include the closure of stomata, activation of accumulation of leaf cuticular waxes and compatible solutes, and primary root elongation, among others. Most of these responses are ABA-induced to avoid water loss from the plant organs and increase the chances of plant survival under stress conditions. The general mechanism of any stress tolerance includes the following steps: (i) activation of transcriptional factors under stress conditions; (ii) binding with the ABA signal pathway to direct synthesis of ABA, and (iii) upregulation of ABA-dependent genes. CRISPR/Ca9-directed mutant lines have been developed to study the plant’s response under stress and regulation of stress-responsive genes. Gene expression of SAPK2 was upregulated by stress factors such as drought, salinity, or osmotic (polyethylene glycol). However, a mutant line for sapk2 showed ABA insensitivity during seed germination and ABA-mediated seed dormancy (Lou et al., 2017).
Moreover, mutant sapk2 was more sensitive to drought tolerance and ROS, thus showing the role of SAPK2 in the enhancement of drought tolerance (Lou et al., 2017).
The gene has been shown to assist plant survival by reducing water loss through stomatal closure, activating ROS scavengers, and upregulating the expression of genes such as OsRab16b, OsRab21, OsbZIP23, OsLEA3, and OsOREB1 (Lou et al., 2017). The gene SAPK9 has a role in osmolyte (sugar and proline) accumulation, which helps to maintain osmotic potential under water stress conditions and retain water content by stomatal closure and reduction in the electrolyte leakage and ion detoxification (Dey et al., 2016a,b).
The gene SAPK9 was originally cloned from Oryza rufipogon, and overexpression lines were developed in Oryza sativa (rice crop), which showed higher drought tolerance than wild-type and RNAi lines (Dey et al., 2016a). SAPK9 physically interacted with the OsMADS23 for transcriptional activation that upregulated the ABA and proline biosynthesis genes such as OsNCED2, OsNCED3, OsNCED4, and OsP5CR (Li et al., 2021). The expression of gene OsbZIP23 was induced during the reproductive stage and improved the yield component by overaccumulating the osmotica such as sugar proline and decreasing the lipid oxidation when the gene was cloned in the drought-susceptible lines IR20 (Dey et al., 2016b). Overexpression transgenic lines in rice have also been developed to confirm stress tolerance in the rice. Overexpression of the OsPYL6 gene reduced plant transpiration by 25% compared to the control plant (Santosh Kumar et al., 2021). To cope with water stress and to avoid stress, plants produce several metabolites, i.e., sugars, free amino, heat shock proteins, and acids, to cope with the changing osmotic potential of the cell. These metabolites are collectively called osmolytes, which protect the cellular membrane and organelles at the cost of energy. Metabolites may, however, be utilized during stress recovery of the plant.
BA1 (ERA1) plays a role in plants during osmotic stress. It encodes a β-subunit of farnesyltransferase, which regulates the ABA signaling and dehydration response; therefore, ABA1 (ERA1) is an important candidate gene to enhance drought tolerance in numerous crops. Frameshift mutant cas9 line shows a similar phenotype of leaf shape while positively increasing the root growth, which helps the plant to uptake water from deep soil profile and enhance the drought tolerance capacity (Ogata et al., 2020). It also enhances both the ABA sensitivity and drought response through stomatal regulation. As a result, during no-stress conditions, the role is opposite for primary root growth. However, the study of the ABA1 gene benchmark assists in understanding the role of ABA in osmotic stress, and such knowledge may improve rice drought tolerance for caryopsis yield.
OsPYL/RCAR7 was a novel ABA receptor linked with drought tolerance. It has weak ABA signal activity but a strong dependency on ABA. This receptor-induced drought tolerance with a minimum penalty for yield (Bhatnagar et al., 2020). Overexpression of the ABA receptor (i.e., OsPYL/RCAR3) induced better growth under osmotic stress. Clade A Type 2C protein phosphatases are a negative regulator of ABA signals but activate the ABA-independent pathway via cis-acting drought-responsive elements. Overexpression of the OsPYL/RCAR3 interacting protein, i.e., OsPP2C09, resulted in plant sensitivity to the stress (Min et al., 2021).
SiMYB56 was related to ABA biosynthesis and upregulates drought tolerance genes. Overexpression lines SiMYB56 had lower monoaldehyde contents and high lignin contents, which may be related to the transpiration reduction without affecting the plant’s normal morphology (Xu et al., 2020).
Leaf morphology (such as leaf cuticular waxes and leaf rolling) is also important to avoid dehydration.
Homozygous mutants were developed using CRISPR-mediated targeted gene knockdown of SRL1 and SRL2 in rice (Liao et al., 2019).
The mutants exhibited lower chlorophyll content, impaired leaf gas exchange characteristics, and a reduced number of panicles (Liao et al., 2019). However, under drought conditions, the mutants showed lower monoaldehyde contents and a higher panicle-filling percentage compared to wild-type plants. The mutants also demonstrated greater defense against ROS by activating ROS scavenging enzymes, such as superoxide dismutase (SOD), and catalase (CAT), and showed higher ABA content (Liao et al., 2019). Proteomic analysis of the semi-rolled lead mutant showed that LBD proteins were downregulated, while stress-responsive proteins were upregulated under drought conditions (Liao et al., 2019).
Drought drastically changes the expression of genes. It activates genes related to adaptability under water stress (Table 2). Functional genomics and the development of mutant or overexpression lines resulted in understanding the genes related to water stress tolerance in rice. Genes related to osmotic adjustment and reactive oxygen species (ROS), e.g., OsHSFA3, are known to be activated by ABA signals (Zhu et al., 2020). OsPYL6 upregulated the epoxy carotenoid (NCED) gene to direct ABA biosynthesis. Disruption of the Os3BGlu6 gene caused dwarfism and decreased the ABA contents of the cell, lowered the photosynthesis rate, and increased the overall sensitivity to drought (Wang B. et al., 2020; Wang C. et al., 2020; Wang H. et al., 2020; Wang T. et al., 2020). Contrastingly, overexpression lines had higher drought tolerance and maintained higher ABA contents. Overexpression lines also had higher activity of the β-glucosidase enzyme (Wang B. et al., 2020; Wang C. et al., 2020; Wang H. et al., 2020; Wang T. et al., 2020). Similarly, disruption of the OsASLRK gene resulted in drought sensitivity and lower ABA contents in the cell (Du et al., 2022). Mutants for gene Osao3- had lower drought tolerance with high yield potential due to increased number of panicles, spikelet number, and spikelet fertility.
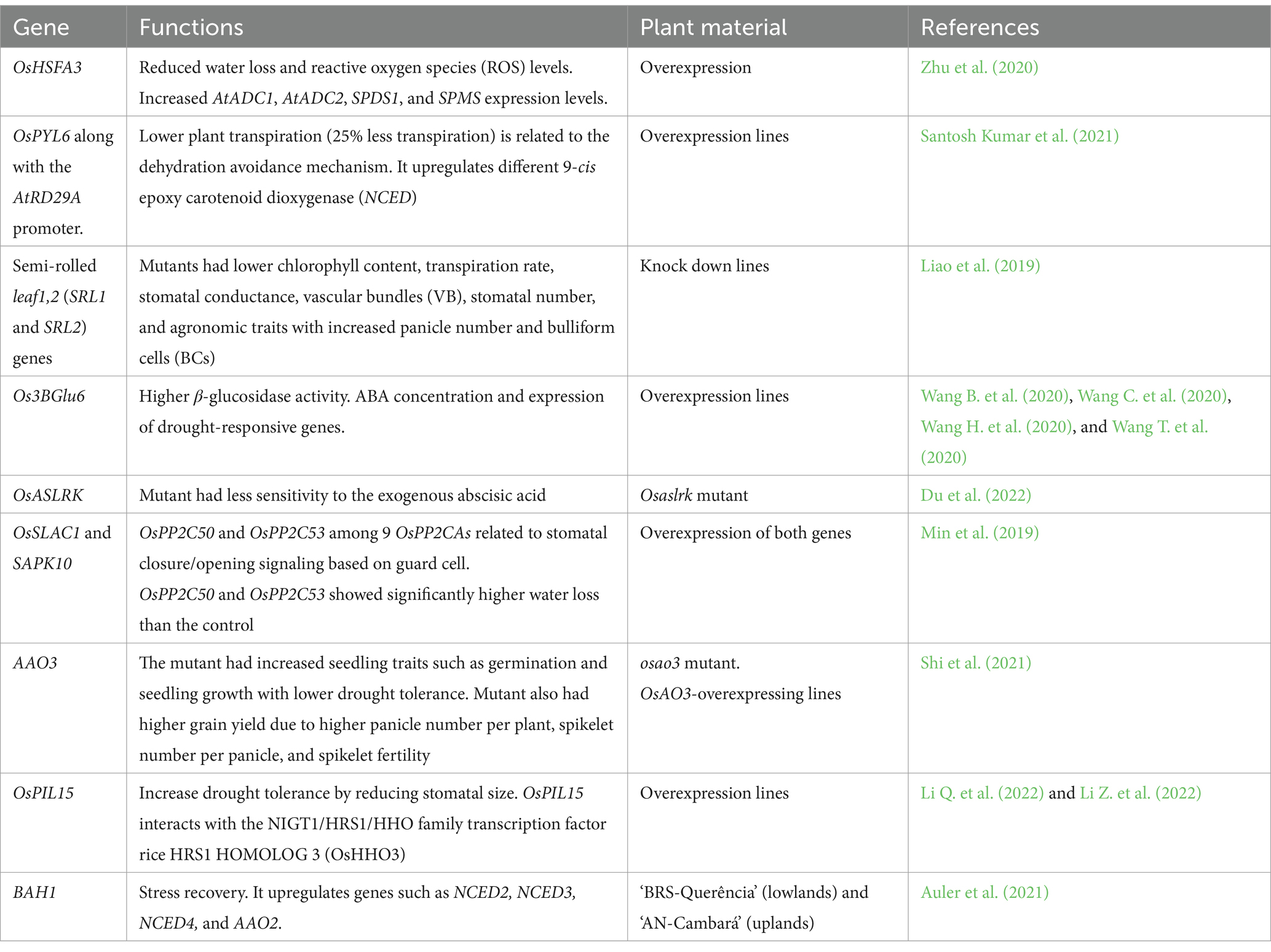
Table 2. ABA signaling pathways for genes related to drought tolerance in rice identified in overexpression six or knockdown lines.
OsFBX257 is co-expressed in a network of the protein kinases and phosphatases. Its knockdown oxfbx257 has reduced root growth and depth, panicle size, and survivability under stress (Sharma et al., 2023). Overexpression lines for OsFBX257, on the other hand, had deeper root, grain yield, and panicle size (Sharma et al., 2023).
Dehydration or osmotic stress and mineral toxicity due to the accumulation of salts in response to brackish water irrigation induce several plant responses, such as vacuolar compartmentation, activation of reactive oxygen species, and osmotic adjustment. Many of the plant defense responses against salt stress are induced by the ABA signaling. The gene OsMFT1 was involved in the ABA and GA metabolism in rice under salinity stress and also modulates seed germination (Lu et al., 2023). Suppression of catabolic ABA gene OsABA8ox1-kd maintained a higher level of ABA, and plants exhibited reduced plasma lemma injury, ROS accumulation, NA+/K+ ratio, and less mortality under mineral stress induced by the various levels of Na2CO3 (Liu et al., 2022a,Liu et al., 2022b).
The structural and functional diversity of OsNHX1-OSNHX5 was investigated in rice, as these genes are involved in Na+/K+ antiporters for the vacuolar compartmentation of ions (Fukuda et al., 2011). The activity of the genes OSNHX1 was localized in the specialized tissues such as guard cells and trichomes of rice leaf, whereas OSNHX1 activity was observed in the pollen grains and root tip (Fukuda et al., 2011).
Downregulation of the miR528 (1-ascorbate oxidase) resulted in the increase of ascorbic acid and ABA contents of the cell, which caused a significant decrease in reactive oxygen species (ROS), inducing salt tolerance in rice (Wang G. et al., 2021; Wang M. et al., 2021; Wang T. et al., 2021). The regulatory gene OsJRL40 was known to have higher activity in roots and internodes under salt stress (Gao et al., 2023). It controlled the expression of genes related to the Na+/K+ transporter and antioxidant activity of the cell (Gao et al., 2023). Overexpression of OsMADS25 activated the genes OsGST4 (glutathione S–transferase) and OsP5CR involved in the biosynthesis of proline by directly binding to the promoter of the respective gene (Xu et al., 2011). It increased the plant sensitivity for the ABA application and promoted the expression of the ABA-dependent gene. The expression of OsMADS25 also resulted in the scavenging of ROS (Xu et al., 2011). Many other genes (e.g., SalT, OsNAC9, OsAKT1, and OsHKT) were involved in the transcription of reactive oxygen scavengers, i.e., superoxide dismutase, catalase, and peroxidase that reduced the damage due to alkalinity (Liu X. et al., 2019; Liu X. L. et al., 2019).
OsNAC genes play an important role in rice development under salt stress, with OsNAC3 specifically associated with ABA stress signaling and salt tolerance. Knocking out of OSNAC3 decreased the sensitivity for ABA and increased sensitivity for the salt stress. Knock outline for osnac3 showed a higher accumulation of Na+ in the shoot (Zhang A. et al., 2021; Zhang X. et al., 2021). Knock outline also showed downregulation of the OsHKT1;4, OsHKT1;5, OsLEA3–1, OsPM-1, OsPP2C68, and OsRAB-21, which were related to the ABA pathway and salt tolerance (Zhang A. et al., 2021; Zhang X. et al., 2021). Overexpression lines of OsERF19 induce salt tolerance and hypersensitivity for the ABA. It was found that the transcriptional factor of OsERF19 binds with the promoters of OsOTS1 and OsNCED5 to induce the synthesis of ABA. OsERF19 also upregulates the genes OsLEA3, OsNHX1, OsHKT6, and OsOTS1 under salinity stress, while genes such as OsRAB21, OsNCED5, and OsP5CS1 were upregulated under ABA treatment (Huang et al., 2021). OsSAPK7 modulates ion homeostasis, redox reaction, and photosynthesis. It increased seedling survival, chlorophyll and proline contents, catalase, and superoxide dismutase activity. The knockout line of ossapk7 had a lower survival rate, higher Na+/K+ uptake, and ion toxicity under salt stress, which showed their role in salinity tolerance (Dan et al., 2021).
In conclusion, it was noticed that suppression of the ABA catabolic gene (i.e., OsABA8ox1-kd) increased the ABA contents, which also improved salt tolerance. Gene OsNHX1-5 may be considered for further research and use in breeding due to their role in the vacuolar compartmentation. The gene OSNAC3 was related to ABA contents and stress signaling under salinity. Gene OsSAPK7 and other series kinase proteins within the same group are also of particular interest due to their positive role in stress signaling and other responses under stress (Table 3).
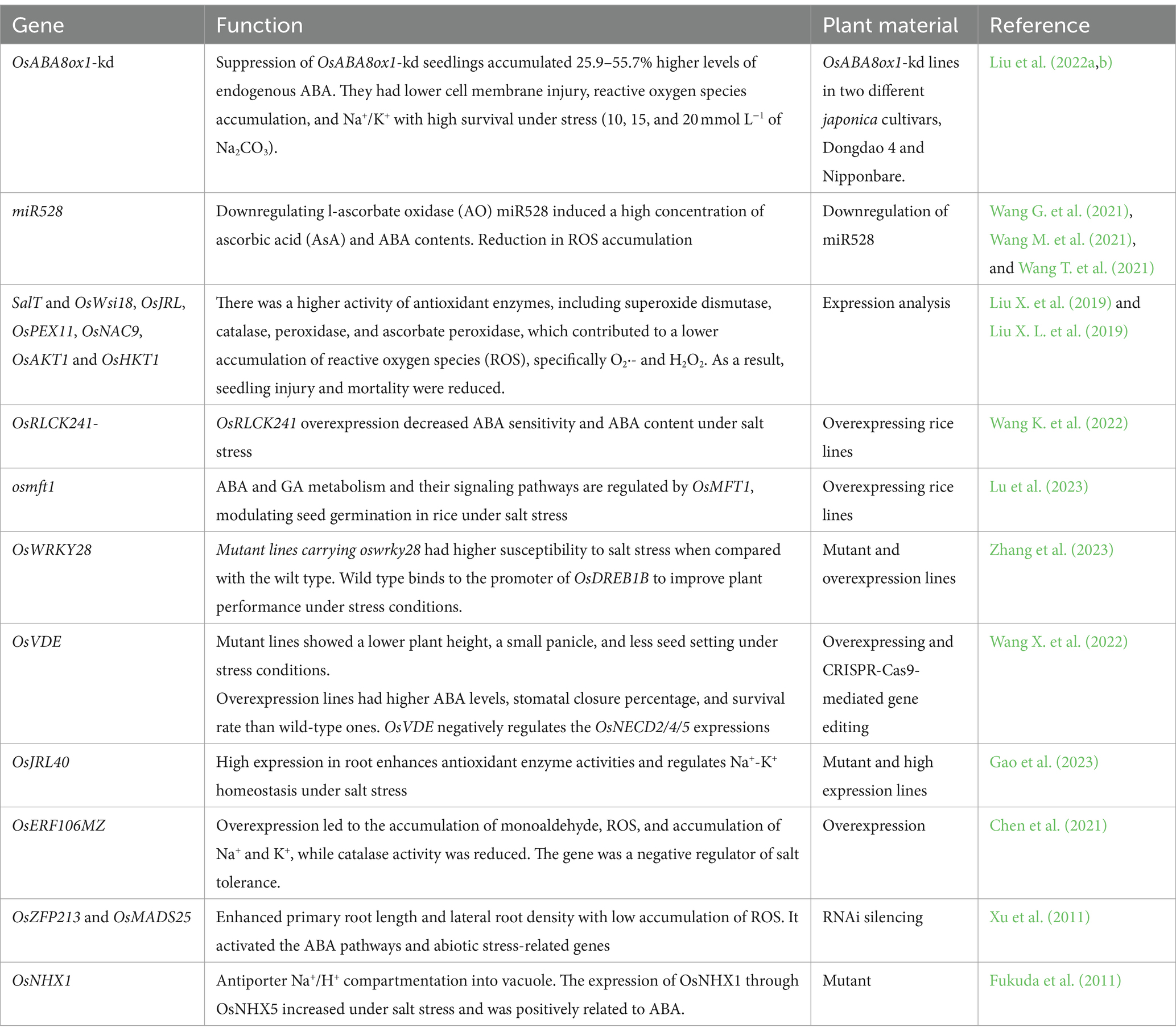
Table 3. ABA stress signaling in rice for plant responses and tolerance mechanism under mineral stress.
The ABA is a common stress signal under various abiotic stress conditions. The common stress-responsive genes were identified when cells were exposed to multiple stress factors. Transcriptional factor OsbZIP72 bonds to the promoter of the OsHKT1 for the activation of ABA biosynthesis. Overexpression of OsbZIP72 induced tolerance against water and salinity stress (Baoxiang et al., 2021). OsbZIP72 binds to the promoters of genes like OsSWEET13 and OsSWEET15 to activate sugar transport under multiple stressors (Mathan et al., 2021). OsbZIP86 encodes a transcriptional factor that binds to the ABA biosynthesis gene (e.g., OsNCED3 to mitigate drought tolerance). OsbZIP86 itself was regulated through miR2105, which directs its cleavage (Gao et al., 2023).
Moreover, OsSAPK10 activates the expression of OsbZIP86 to increase the expression of OsNCED3. Overexpression lines of OsbZIP86 caused the enhancement of drought tolerance by reducing the water losses from stomata and controlling the transpiration loss (Gao et al., 2023). Similarly, OsNAC006 induces regulatory transcriptional factors for the activation of abiotic-resistant genes under various treatments, i.e., H2O2, cold, heat, abscisic acid (ABA), indole-3-acetic acid (IAA), gibberellin (GA), NaCl, and polyethylene glycol (PEG) 6,000 (Wang B. et al., 2020; Wang C. et al., 2020; Wang H. et al., 2020; Wang T. et al., 2020). OsMSR2 induced drought and salinity tolerance by modulating the ABA acid pathway (Xu et al., 2011). Overexpression of the OsSMP1 (stress membrane protein) induced tolerance against drought, cold, and salinity heavy metals and was dependent on the ABA pathways (Zheng et al., 2021). Overexpression of OsMLP423 increased the sensitivity to the ABA and water use efficiency and reduced the membrane damage and reactive oxygen species under salinity and drought stress (Zhou H. et al., 2022; Zhou Z. et al., 2022). Auxin/IAA genes were induced in response to the drought and other abiotic stresses. RNAi lines for OsIAA20 showed reduced tolerance to drought and salt tolerance (Zhang A. et al., 2021; Zhang X. et al., 2021). This stress-sensitive line had lower proline content and gradually lost chlorophyll content.
Moreover, the susceptible mutant line for osiaa20 had a higher monoaldehyde value and Na+/K+ ratio. The overexpression line had increased stomatal closure and decreased transpiration loss, helping rice plants to survive under salt and drought stress (Zhang A. et al., 2021; Zhang X. et al., 2021). RNAi line also had downregulated OsRab21, which was an ABA-responsive gene. The overall result showed that OsIAA20 induced salt and drought tolerance via ABA stress signals (Zhang A. et al., 2021; Zhang X. et al., 2021). The overexpression line for the glycosyltransferase gene UGT3 enhanced the salt and drought tolerance in rice. There was high expression of antioxidant enzymes and very low production of reactive oxygen species within the overexpression line of UGT3 (Wang G. et al., 2021; Wang M. et al., 2021; Wang T. et al., 2021).
HDA710 expression levels accumulate in response to multiple stressors and phytohormones, i.e., the jasmonic and abscisic acids. Knock-down line oshda710 showed enhanced salt tolerance and reduced ABA sensitivity (Sharma et al., 2023). Mutant hda710 also upregulated the ABA and salt-tolerant genes such as OsLEA3, OsABI5, OsbZIP72, and OsNHX1 (Sharma et al., 2023). Expression of these genes was epigenetically controlled by acetylation of H4 histones.
OsABAR1 confers drought and salt tolerance in ABA-dependent pathways and is a novel GRAM-encoding gene. Gene expression was highest in roots under drought and salt stress and upregulated by ABA (Zheng et al., 2020). Overexpression lines of OsABAR1 had a high tolerance to salinity and drought, while knock-out mutant lines had the opposite phenotype (Zheng et al., 2020).
OsRF1-OE is a small ring-type E3 ligase that is involved in the increased endogenous level of the ABA and confers tolerance to salt and drought stress. Overexpression lines of OsRF1-OE had enhanced ABA level, salt, and drought tolerance. Overexpression lines showed faster transcriptional activation of genes related to ABA biosynthesis, e.g., ZEP, NCED3, and ABA4. OsRF1 exhibited E3 ligase activity and was involved in the ubiquitination and degradation of the OsPP2C09 protein. It was identified that the OsRF1 facilitates ABA biosynthesis by modifying and degrading the clade A PP2C proteins (Kim et al., 2022). OsDIP1 and ZFP36 activated the ABA-induced antioxidative defense system in rice. Overexpression lines had a high activity of superoxide dismutase and catalase enzyme, i.e., there was a decrease in monoaldehyde contents. In contrast, mutant lines OsDIP1 had decreased expression of genes such as SODCc2 and CatB and increased levels of monoaldehyde contents (Min et al., 2021) (Table 4).
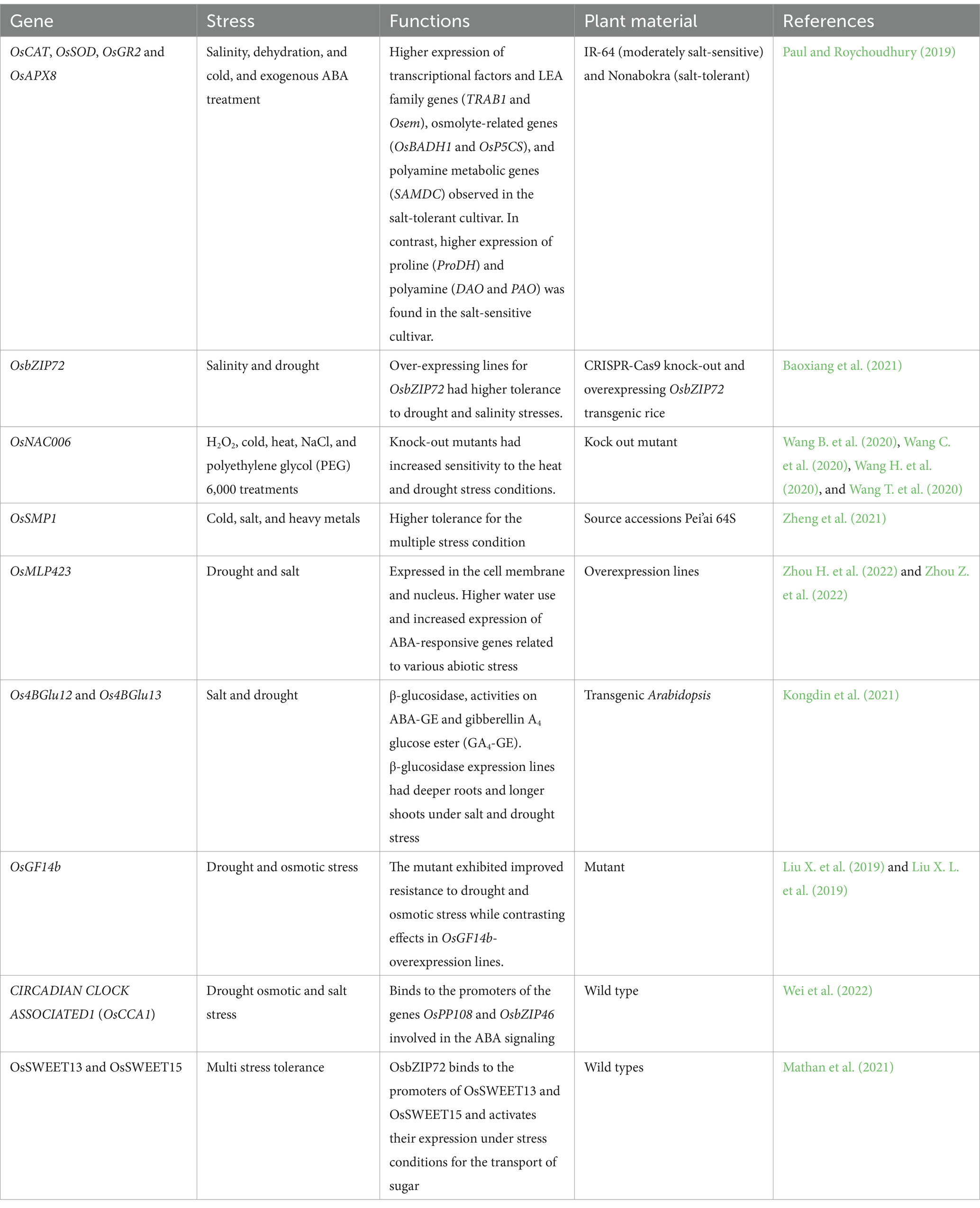
Table 4. ABA-modulated genes under a multi-stress environment in rice.
Heat stress causes significant damage to rice plant development by increasing respiration, accelerating senescence, causing membrane leakage, and generating reactive oxygen species (ROS). High pollen sterility has been associated with elevated ROS levels (Zhao et al., 2023). ABA-induced gene responses under heat stress have been shown to improve plant tolerance (Wang B. et al., 2020; Wang C. et al., 2020; Wang H. et al., 2020; Wang T. et al., 2020). Direct foliar application of ABA at 40°C also increases heat tolerance and boosts yield due to increased pollen viability (Rezaul et al., 2019).
Increases of 14 and 22% in seed setting were observed in heat-resistant and heat-susceptible rice lines, respectively, following exogenous application under heat stress (Zhou et al., 2019). ABA application under stress conditions induced heat shock proteins (HSP24.1 and HSP71.1) and triggered the expression of sucrose transporter (SUT) genes, sucrose synthase (SUS) genes, and invertase (INV) genes (Rezaul et al., 2019). Several ABA-associated genes, such as OsIAA13, OsIAA20, ILL8, OsbZIP12, OsPP2C51, OsDi19-1, and OsHOX24, were validated under high temperatures of 37 and 42°C during the seedling stage (Sharma et al., 2021).
The ABA pretreatment reduced cell injury, monoaldehyde, ROS, and the expression of programmed cell death genes such as OsKOD1, OsCP1, and OsNAC4 while upregulating OsBI1, a suppressor of programmed cell death (Liu et al., 2020). Additionally, ABA pretreatment increased the antioxidant capacity of cells by reducing ROS and monoaldehyde levels (Liu et al., 2020). Overexpression of OsNCED1, a key factor in ABA biosynthesis, increased pollen viability, grain filling, and ROS scavenger activity (catalase, superoxide dismutase, and peroxidase) compared to controls while reducing relative electrolyte leakage and monoaldehyde content in transgenic plants carrying the OsNCED1 gene (Zhou et al., 2019) (Table 5).
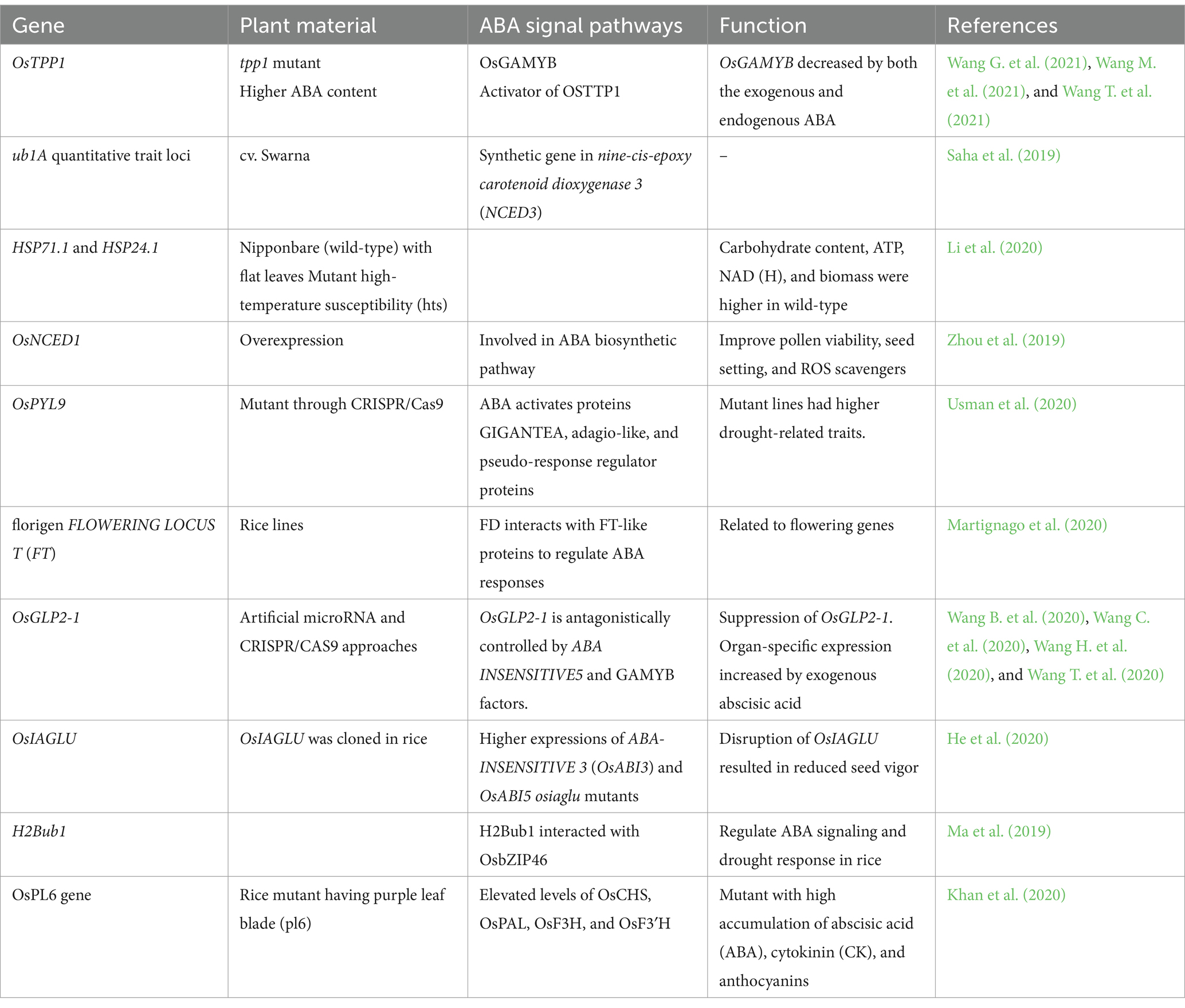
Table 5. ABA-mediated genes under heat stress.
RG: Writing – review & editing, Writing – original draft, Conceptualization. SR: Writing – review & editing, Writing – original draft, Supervision, Project administration, Conceptualization. RO: Writing – review & editing, Resources, Project administration. MK: Writing – review & editing. YK: Writing – review & editing.
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Asad, M. A. U., Wang, F., Ye, Y., Guan, X., Zhou, L., Han, Z., et al. (2021). Contribution of ABA metabolism and ROS generation to sugar starvation-induced senescence of rice leaves. Plant Growth Regul. 95, 241–257. doi: 10.1007/s10725-021-00718-4
Aslam, M., Waseem, M., Jakada, B. H., Okal, E. J., Lei, Z., Saqib, H. S. A., et al. (2022). Mechanisms of abscisic acid-mediated drought stress responses in plants. Int. J. Mol. Sci. 23:1084. doi: 10.3390/ijms23031084
Auler, P. A., Nogueira do Amaral, M., Rossatto, T., Lopez Crizel, R., Milech, C., Clasen Chaves, F., et al. (2021). Metabolism of abscisic acid in two contrasting rice genotypes submitted to recurrent water deficit. Physiol. Plant. 172, 304–316. doi: 10.1111/ppl.13126
Baoxiang, W., Yan, L., Yifeng, W., Jingfang, L., Zhiguang, S., Ming, C., et al. (2021). OsbZIP72 is involved in the transcriptional gene-regulation pathway of abscisic acid signal transduction by activating rice high-affinity potassium transporter OsHKT1; 1. Rice Sci. 28, 257–267. doi: 10.1016/j.rsci.2021.04.005
Bhatnagar, N., Kim, R., Han, S., Song, J., Lee, G. S., Lee, S., et al. (2020). Ectopic expression of OsPYL/RCAR7, an ABA receptor having low signaling activity, improves drought tolerance without growth defects in rice. Int. J. Mol. Sci. 21:4163. doi: 10.3390/ijms21114163
Chen, K., Du, K., Shi, Y., Yin, L., Shen, W. H., Yu, Y., et al. (2021). H3K36 methyltransferase SDG708 enhances drought tolerance by promoting abscisic acid biosynthesis in rice. New Phytol. 230, 1967–1984. doi: 10.1111/nph.17290
Dan, Z., Chunchao, W., Junpin, X., Fan, Z., Jialing, L., Xiaorong, S., et al. (2021). Stress-activated protein kinase OsSAPK7 regulates salt-stress tolerance by modulating diverse stress-defensive responses in rice. Rice Sci. 28, 547–556. doi: 10.1016/j.rsci.2021.09.003
Dey, A., Samanta, M. K., Gayen, S., and Maiti, M. K. (2016a). The sucrose non-fermenting 1-related kinase two gene SAPK9 improves drought tolerance and grain yield in rice by modulating cellular osmotic potential, stomatal closure, and stress-responsive gene expression. BMC Plant Biol. 16:158. doi: 10.1186/s12870-016-0845-x
Dey, A., Samanta, M. K., Gayen, S., Sen, S. K., and Maiti, M. K. (2016b). Enhanced gene expression rather than natural polymorphism in the coding sequence of the OsbZIP23 determine drought tolerance and yield improvement in rice genotypes. PLoS One 11:e0150763. doi: 10.1371/journal.pone.0150763
Du, C., Cai, W., Lin, F., Wang, K., Li, S., Chen, C., et al. (2022). Leucine-rich repeat receptor-like kinase OsASLRK regulates abscisic acid and drought responses via cooperation with S-like RNase OsRNS4 in rice. Environ. Exp. Bot. 201:104949. doi: 10.1016/j.envexpbot.2022.104949
Fukuda, A., Nakamura, A., Hara, N., Toki, S., and Tanaka, Y. (2011). Molecular and functional analyses of rice NHX-type Na+/H+ antiporter genes. Planta 233, 175–188. doi: 10.1007/s00425-010-1289-4
Gao, Q., Yin, X., Wang, F., Hu, S., Liu, W., Chen, L., et al. (2023). OsJRL40, a jacalin-related lectin gene, promotes salt stress tolerance in rice. Int. J. Mol. Sci. 24:7441. doi: 10.3390/ijms24087441
Hasanuzzaman, M., Zhou, M., and Shabala, S. (2023). How does stomatal density and residual transpiration contribute to osmotic stress tolerance? Plan. Theory 12:494. doi: 10.3390/plants12030494
He, Y., Zhao, J., Yang, B., Sun, S., Peng, L., and Wang, Z. (2020). Indole-3-acetate beta-glucosyltransferase OsIAGLU regulates seed vigour through mediating crosstalk between auxin and abscisic acid in rice. Plant Biotech. J. 18, 1933–1945. doi: 10.1111/pbi.13353
Hirayama, T., and Umezawa, T. (2010). The PP2C–SnRK2 complex: the central regulator of an abscisic acid signaling pathway. Plant Signal. Behav. 5, 160–163. doi: 10.4161/psb.5.2.10460
Hu, Y., Song, S., Weng, X., You, A., and Xing, Y. (2021). The heading-date gene Ghd7 inhibits seed germination by modulating the balance between abscisic acid and gibberellins. Crop J. 9, 297–304. doi: 10.1016/j.cj.2020.09.004
Huang, S., Ma, Z., Hu, L., Huang, K., Zhang, M., Zhang, S., et al. (2021). Involvement of rice transcription factor OsERF19 in response to ABA and salt stress responses. Plant Physiol. Biochem. 167, 22–30. doi: 10.1016/j.plaphy.2021.07.027
Jin, X., Zhang, Y., Li, X., and Huang, J. (2023). OsNF-YA3 regulates plant growth and osmotic stress tolerance by interacting with SLR1 and SAPK9 in rice. Plant J. 114, 914–933. doi: 10.1111/tpj.16183
Khan, A., Jalil, S., Cao, H., Tsago, Y., Sunusi, M., Chen, Z., et al. (2020). The purple leaf (pl6) mutation regulates leaf color by altering the anthocyanin and chlorophyll contents in rice. Plant 9:1477. doi: 10.3390/plants9111477
Kim, S., Park, S. I., Kwon, H., Cho, M. H., Kim, B. G., Chung, J. H., et al. (2022). The rice abscisic acid-responsive RING finger E3 ligase OsRF1 targets OsPP2C09 for degradation and confers drought and salinity tolerance in rice. Front. Plant Sci. 12:797940. doi: 10.3389/fpls.2021.797940
Kongdin, M., Mahong, B., Lee, S. K., Shim, S. H., Jeon, J. S., and Ketudat Cairns, J. R. (2021). Action of multiple rice β-glucosidases on abscisic acid glucose ester. Int. J. Mol. Sci. 22:7593. doi: 10.3390/ijms22147593
Li, Q. F., Lu, J., Zhou, Y., Wu, F., Tong, H. N., Wang, J. D., et al. (2019). Abscisic acid represses rice lamina joint inclination by antagonizing brassinosteroid biosynthesis and signaling. Int. J. Mol. Sci. 20:4908. doi: 10.3390/ijms20194908
Li, Z., Rao, M. J., Li, J., Wang, Y., Chen, P., Yu, H., et al. (2022). CRISPR/Cas9 mutant rice Ospmei12 involved in growth, cell wall development, and response to phytohormone and heavy metal stress. Int. J. Mol. Sci. 23:16082. doi: 10.3390/ijms232416082
Li, X., Yu, B., Wu, Q., Min, Q., Zeng, R., Xie, Z., et al. (2021). OsMADS23 phosphorylated by SAPK9 confers drought and salt tolerance by regulating ABA biosynthesis in rice. PLoS Genet. 17:e1009699. doi: 10.1371/journal.pgen.1009699
Li, G., Zhang, C., Zhang, G., Fu, W., Feng, B., Chen, T., et al. (2020). Abscisic acid negatively modulates heat tolerance in rolled leaf rice by increasing leaf temperature and regulating energy homeostasis. Rice 13:18. doi: 10.1186/s12284-020-00379-3
Li, Q., Zhou, L., Chen, Y., Xiao, N., Zhang, D., Zhang, M., et al. (2022). Phytochrome interacting factor regulates stomatal aperture by coordinating red light and abscisic acid. Plant Cell 34, 4293–4312. doi: 10.1093/plcell/koac244
Liang, C., Wang, Y., Zhu, Y., Tang, J., Hu, B., Liu, L., et al. (2014). OsNAP connects abscisic acid and leaf senescence by fine-tuning abscisic acid biosynthesis and directly targeting senescence-associated genes in rice. Proc. Natl. Acad. Sci. 111, 10013–10018. doi: 10.1073/pnas.1321568111
Liao, S., Qin, X., Luo, L., Han, Y., Wang, X., Usman, B., et al. (2019). CRISPR/Cas9-induced mutagenesis of semi-rolled leaf1, 2 confers curled leaf phenotype and drought tolerance by influencing protein expression patterns and ROS scavenging in rice (Oryza sativa L.). Agronomy 9:728. doi: 10.3390/agronomy9110728
Liu, X., Hu, Q., Yan, J., Sun, K., Liang, Y., Jia, M., et al. (2020). ζ-Carotene isomerase suppresses tillering in rice through the coordinated biosynthesis of strigolactone and abscisic acid. Mol. Plant 13, 1784–1801. doi: 10.1016/j.molp.2020.10.001
Liu, X., Ji, P., Yang, H., Jiang, C., Liang, Z., Chen, Q., et al. (2022a). Priming effect of exogenous ABA on heat stress tolerance in rice seedlings is associated with the upregulation of antioxidative defense capability and heat shock-related genes. Plant Growth Regul. 98, 23–38. doi: 10.1007/s10725-022-00828-7
Liu, X., Li, Z., Hou, Y., Wang, Y., Wang, H., Tong, X., et al. (2019). Protein interactomic analysis of SAPKs and ABA-inducible bZIPs revealed key roles of SAPK10 in rice flowering. Int. J. Mol. Sci. 20:1427. doi: 10.3390/ijms20061427
Liu, X., Xie, X., Zheng, C., Wei, L., Li, X., Jin, Y., et al. (2022b). RNAi-mediated suppression of the abscisic acid catabolism gene OsABA8ox1 increases abscisic acid content and tolerance to saline–alkaline stress in rice (Oryza sativa L.). Crop J. 10, 354–367. doi: 10.1016/j.cj.2021.06.011
Liu, X. L., Zhang, H., Jin, Y. Y., Wang, M. M., Yang, H. Y., Ma, H. Y., et al. (2019). Abscisic acid primes rice seedlings for enhanced tolerance to alkaline stress by upregulating antioxidant defense and stress tolerance-related genes. Plant Soil 438, 39–55. doi: 10.1007/s11104-019-03992-4
Liu, Y., Zhou, W., He, M., Sui, J., Tian, X., Guan, Q., et al. (2024). Comprehensive analysis of stress-activated protein kinase genes (OsSAPKs) in rice flowering time. Planta 259:149. doi: 10.1007/s00425-024-04431-0
Lou, D., Chen, Z., Yu, D., and Yang, X. (2020). SAPK2 contributes to rice yield by modulating nitrogen metabolic processes under reproductive stage drought stress. Rice 13:35. doi: 10.1186/s12284-020-00395-3
Lou, D., Wang, H., Liang, G., and Yu, D. (2017). OsSAPK2 confers abscisic acid sensitivity and tolerance to drought stress in rice. Front. Plant Sci. 8:993. doi: 10.3389/fpls.2017.00993
Lu, K., Guo, Z., Di, S., Lu, Y., Muhammad, I. A. R., Rong, C., et al. (2023). OsMFT1 inhibits seed germination by modulating abscisic acid signaling and gibberellin biosynthesis under salt stress in rice. Plant Cell Physiol. 64, 674–685. doi: 10.1093/pcp/pcad029
Ma, S., Tang, N., Li, X., Xie, Y., Xiang, D., Fu, J., et al. (2019). Reversible histone H2B monoubiquitination fine-tunes abscisic acid signaling and drought response in rice. Mol. Plant 12, 263–277. doi: 10.1016/j.molp.2018.12.005
Martignago, D., Siemiatkowska, B., Lombardi, A., and Conti, L. (2020). Abscisic acid and flowering regulation: many targets, different places. Int. J. Mol. Sci. 21:9700. doi: 10.3390/ijms21249700
Mathan, J., Singh, A., and Ranjan, A. (2021). Sucrose transport in response to drought and salt stress involves ABA-mediated induction of OsSWEET13 and OsSWEET15 in rice. Physiol. Plant. 171, 620–637. doi: 10.1111/ppl.13210
Miao, J., Li, X., Li, X., Tan, W., You, A., Wu, S., et al. (2020). OsPP2C09, a negative regulatory factor in abscisic acid signalling, plays an essential role in balancing plant growth and drought tolerance in rice. New Phytol. 227, 1417–1433. doi: 10.1111/nph.16670
Miao, C., Xiao, L., Hua, K., Zou, C., Zhao, Y., Bressan, R. A., et al. (2018). Mutations in a subfamily of abscisic acid receptor genes promote rice growth and productivity. Proc. Natl. Acad. Sci. 115, 6058–6063. doi: 10.1073/pnas.1804774115
Min, M. K., Choi, E. H., Kim, J. A., Yoon, I. S., Han, S., Lee, Y., et al. (2019). Two clade a phosphatase 2Cs expressed in guard cells physically interact with abscisic acid signaling components to induce stomatal closure in rice. Rice 12:37. doi: 10.1186/s12284-019-0297-7
Min, M. K., Kim, R., Hong, W. J., Jung, K. H., Lee, J. Y., and Kim, B. G. (2021). OsPP2C09 is a bifunctional regulator in both ABA-dependent and independent abiotic stress signaling pathways. Int. J. Mol. Sci. 22:393. doi: 10.3390/ijms22010393
Mizokami, Y., Noguchi, K., Kojima, M., Sakakibara, H., and Terashima, I. (2019). Effects of instantaneous and growth CO2 levels and abscisic acid on stomatal and mesophyll conductances. Plant Cell Environ. 42, 1257–1269. doi: 10.1111/pce.13484
Nawaz, G., Usman, B., Zhao, N., Han, Y., Li, Z., Wang, X., et al. (2020). CRISPR/Cas9 directed mutagenesis of OsGA20ox2 in high yielding basmati rice (Oryza sativa L.) line and comparative proteome profiling of unveiled changes triggered by mutations. Int. J. Mol. Sci. 21:6170. doi: 10.3390/ijms21176170
Ogata, T., Ishizaki, T., Fujita, M., and Fujita, Y. (2020). CRISPR/Cas9-targeted mutagenesis of OsERA1 confers enhanced responses to abscisic acid and drought stress and increased primary root growth under non-stressed conditions in rice. PLoS One 15:e0243376. doi: 10.1371/journal.pone.0243376
Ozfidan, C., Turkan, I., Sekmen, A. H., and Seckin, B. (2013). Time course analysis of ABA and non-ionic osmotic stress-induced changes in water status, chlorophyll fluorescence and osmotic adjustment in Arabidopsis thaliana wild-type (Columbia) and ABA-deficient mutant (aba2). Environ. Exp. Bot. 86, 44–51. doi: 10.1016/j.envexpbot.2010.09.008
Paul, S., and Roychoudhury, A. (2019). Transcript analysis of abscisic acid-inducible genes in response to different abiotic disturbances in two indica rice varieties. Theor. Exp. Plant Physiol. 31, 249–272. doi: 10.1007/s40626-018-0131-4
Piao, W., Kim, S. H., Lee, B. D., An, G., Sakuraba, Y., and Paek, N. C. (2019). Rice transcription factor OsMYB102 delays leaf senescence by down-regulating abscisic acid accumulation and signaling. J. Exp. Bot. 70, 2699–2715. doi: 10.1093/jxb/erz095
Rauf, S., da Silva, J. T., Khan, A. A., and Naveed, A. (2010). Consequences of plant breeding on genetic diversity. Int. J. Plant Breed 4, 1–21.
Rezaul, I. M., Baohua, F., Tingting, C., Weimeng, F., Caixia, Z., Longxing, T., et al. (2019). Abscisic acid prevents pollen abortion under high-temperature stress by mediating sugar metabolism in rice spikelets. Physiol. Plant. 165, 644–663. doi: 10.1111/ppl.12759
Saha, I., Sarkar, B., Ghosh, A., De, A. K., and Adak, M. K. (2019). Abscisic acid induced cellular responses of sub1A QTL to aluminium toxicity in rice (Oryza sativa L.). Eco. Environ. Safet. 183:109600. doi: 10.1016/j.ecoenv.2019.109600
Sakuraba, Y., Kim, D., Han, S. H., Kim, S. H., Piao, W., Yanagisawa, S., et al. (2020). Multilayered regulation of membrane-bound ONAC054 is essential for abscisic acid-induced leaf senescence in rice. Plant Cell 32, 630–649. doi: 10.1105/tpc.19.00569
Santosh Kumar, V. V., Yadav, S. K., Verma, R. K., Shrivastava, S., Ghimire, O., Pushkar, S., et al. (2021). The abscisic acid receptor OsPYL6 confers drought tolerance to indica rice through dehydration avoidance and tolerance mechanisms. J. Exp. Bot. 72, 1411–1431. doi: 10.1093/jxb/eraa509
Sharma, E., Bhatnagar, A., Bhaskar, A., Majee, S. M., Kieffer, M., Kepinski, S., et al. (2023). Stress-induced F-box protein-coding gene OsFBX257 modulates drought stress adaptations and ABA responses in rice. Plant Cell Environ. 46, 1207–1231. doi: 10.1111/pce.14496
Sharma, E., Borah, P., Kaur, A., Bhatnagar, A., Mohapatra, T., Kapoor, S., et al. (2021). A comprehensive transcriptome analysis of contrasting rice cultivars highlights the role of auxin and ABA responsive genes in heat stress response. Genomics 113, 1247–1261. doi: 10.1016/j.ygeno.2021.03.007
Sharp, R. E., Poroyko, V., Hejlek, L. G., Spollen, W. G., Springer, G. K., Bohnert, H. J., et al. (2004). Root growth maintenance during water deficits: physiology to functional genomics. J. Exp. Bot. 55, 2343–2351. doi: 10.1093/jxb/erh276
Shi, X., Tian, Q., Deng, P., Zhang, W., and Jing, W. (2021). The rice aldehyde oxidase OsAO3 gene regulates plant growth, grain yield, and drought tolerance by participating in ABA biosynthesis. Biochem. Biophys. Res. Commun. 548, 189–195. doi: 10.1016/j.bbrc.2021.02.047
Sinha, R., Peláez-Vico, M. Á., Shostak, B., Nguyen, T. T., Pascual, L. S., Ogden, A. M., et al. (2024). The effects of multifactorial stress combination on rice and maize. Plant Physiol. 194, 1358–1369. doi: 10.1093/plphys/kiad557
Song, J., Ga, E., Park, S., Lee, H., Yoon, I. S., Lee, S. B., et al. (2023). PROTEIN PHOSPHATASE 2C08, a negative regulator of abscisic acid signaling, promotes internode elongation in rice. Int. J. Mol. Sci. 24:10821. doi: 10.3390/ijms241310821
Tang, L., Li, G., Wang, H., Zhao, J., Li, Z., Liu, X., et al. (2024). Exogenous abscisic acid represses rice flowering via SAPK8-ABF1-Ehd1/Ehd2 pathway. J. Adv. Res. 59, 35–47. doi: 10.1016/j.jare.2023.06.012
Tian, C., Zuo, Z., and Qiu, J. L. (2015). Identification and characterization of ABA-responsive microRNAs in rice. J. Genet. Genom. 42, 393–402. doi: 10.1016/j.jgg.2015.04.008
Usman, B., Nawaz, G., Zhao, N., Liao, S., Liu, Y., and Li, R. (2020). Precise editing of the OsPYL9 gene by RNA-guided Cas9 nuclease confers enhanced drought tolerance and grain yield in rice (Oryza sativa L.) by regulating circadian rhythm and abiotic stress responsive proteins. Int. J. Mol. Sci. 21:7854. doi: 10.3390/ijms21217854
Van Houtte, H., Vandesteene, L., López-Galvis, L., Lemmens, L., Kissel, E., Carpentier, S., et al. (2013). Overexpression of the trehalase gene AtTRE1 leads to increased drought stress tolerance in Arabidopsis and is involved in abscisic acid-induced stomatal closure. Plant Physiol. 161, 1158–1171. doi: 10.1104/pp.112.211391
Wang, C., Chen, S., Dong, Y., Ren, R., Chen, D., and Chen, X. (2020). Chloroplastic Os3BGlu6 contributes significantly to cellular ABA pools and impacts drought tolerance and photosynthesis in rice. New Phytol. 226, 1042–1054. doi: 10.1111/nph.16416
Wang, M., Guo, W., Li, J., Pan, X., Pan, L., Zhao, J., et al. (2021). The miR528-AO module confers enhanced salt tolerance in rice by modulating the ascorbic acid and abscisic acid metabolism and ROS scavenging. J. Agric. Food Chemist. 69, 8634–8648. doi: 10.1021/acs.jafc.1c01096
Wang, T., Li, P., Mu, T., Dong, G., Zheng, C., Jin, S., et al. (2020). Overexpression of UGT74E2, an Arabidopsis IBA glycosyltransferase, enhances seed germination and modulates stress tolerance via ABA signaling in rice. Int. J. Mol. Sci. 21:7239. doi: 10.3390/ijms21197239
Wang, K., Li, S., Tian, H., Chen, C., Hu, Z., Zhao, Q., et al. (2022). Receptor-like cytoplasmic kinase OsRLCK241 functions as an important regulator of abscisic acid synthesis and response in rice. Environ. Exp. Bot. 194:104744. doi: 10.1016/j.envexpbot.2021.104744
Wang, G., Li, X., Ye, N., Huang, M., Feng, L., Li, H., et al. (2021). OsTPP1 regulates seed germination through the crosstalk with abscisic acid in rice. New Phytol. 230, 1925–1939. doi: 10.1111/nph.17300
Wang, T., Ma, Y. Q., Huang, X. X., Mu, T. J., Li, Y. J., Li, X. K., et al. (2021). Overexpression of OsUGT3 enhances drought and salt tolerance through modulating ABA synthesis and scavenging ROS in rice. Environ. Exp. Bot. 192:104653. doi: 10.1016/j.envexpbot.2021.104653
Wang, X., Ren, P., Ji, L., Zhu, B., and Xie, G. (2022). OsVDE, a xanthophyll cycle key enzyme, mediates abscisic acid biosynthesis and negatively regulates salinity tolerance in rice. Planta 255:6. doi: 10.1007/s00425-021-03802-1
Wang, Y., Wu, F., Lin, Q., Sheng, P., Wu, Z., Jin, X., et al. (2023). A regulatory loop establishes the link between the circadian clock and abscisic acid signaling in rice. Plant Physiol. 191, 1857–1870. doi: 10.1093/plphys/kiac548
Wang, H., Zhang, Y., Xiao, N., Zhang, G., Wang, F., Chen, X., et al. (2020). Rice GERMIN-LIKE PROTEIN 2-1 functions in seed dormancy under the control of abscisic acid and gibberellic acid signaling pathways. Plant Physiol. 183, 1157–1170. doi: 10.1104/pp.20.00253
Wang, B., Zhong, Z., Wang, X., Han, X., Yu, D., Wang, C., et al. (2020). Knockout of the OsNAC006 transcription factor causes drought and heat sensitivity in rice. Int. J. Mol. Sci. 21:2288. doi: 10.3390/ijms21072288
Wei, H., Xu, H., Su, C., Wang, X., and Wang, L. (2022). Rice CIRCADIAN CLOCK ASSOCIATED 1 transcriptionally regulates ABA signaling to confer multiple abiotic stress tolerance. Plant Physiol. 190, 1057–1073. doi: 10.1093/plphys/kiac196
Xie, W., Li, X., Wang, S., and Yuan, M. (2022). OsWRKY53 promotes abscisic acid accumulation to accelerate leaf senescence and inhibit seed germination by downregulating abscisic acid catabolic genes in rice. Front. Plant Sci. 12:816156. doi: 10.3389/fpls.2021.816156
Xu, G. Y., Rocha, P. S., Wang, M. L., Xu, M. L., Cui, Y. C., Li, L. Y., et al. (2011). A novel rice calmodulin-like gene, OsMSR2, enhances drought and salt tolerance and increases ABA sensitivity in Arabidopsis. Planta 234, 47–59. doi: 10.1007/s00425-011-1386-z
Xu, W., Tang, W., Wang, C., Ge, L., Sun, J., Qi, X., et al. (2020). SiMYB56 confers drought stress tolerance in transgenic rice by regulating lignin biosynthesis and ABA signaling pathway. Front. Plant Sci. 11:785. doi: 10.3389/fpls.2020.00785
Yin, X., Liu, X., Xu, B., Lu, P., Dong, T., Yang, D., et al. (2019). OsMADS18, a membrane-bound MADS-box transcription factor, modulates plant architecture and the abscisic acid response in rice. J. Exp. Bot. 70, 3895–3909. doi: 10.1093/jxb/erz198
Yoshida, T., Fernie, A. R., Shinozaki, K., and Takahashi, F. (2021). Long-distance stress and developmental signals associated with abscisic acid signaling in environmental responses. Plant J. 105, 477–488. doi: 10.1111/tpj.15101
Yuan, Z., Fan, K., Wang, Y., Tian, L., Zhang, C., Sun, W., et al. (2021). OsGRETCHENHAGEN3-2 modulates rice seed storability via accumulation of abscisic acid and protective substances. Plant Physiol. 186, 469–482. doi: 10.1093/plphys/kiab059
Zhang, X., Long, Y., Chen, X., Zhang, B., Xin, Y., Li, L., et al. (2021). A NAC transcription factor OsNAC3 positively regulates ABA response and salt tolerance in rice. BMC Plant Biol. 21:546. doi: 10.1186/s12870-021-03333-7
Zhang, Y., Wang, X., Luo, Y., Zhang, L., Yao, Y., Han, L., et al. (2020). OsABA8ox2, an ABA catabolic gene, suppresses root elongation of rice seedlings and contributes to drought response. Crop J. 8, 480–491. doi: 10.1016/j.cj.2019.08.006
Zhang, A., Yang, X., Lu, J., Song, F., Sun, J., Wang, C., et al. (2021). OsIAA20, an aux/IAA protein, mediates abiotic stress tolerance in rice through an ABA pathway. Plant Sci. 308:110903. doi: 10.1016/j.plantsci.2021.110903
Zhang, M., Zhao, R., Wang, H., Ren, S., Shi, L., Huang, S., et al. (2023). OsWRKY28 positively regulates salinity tolerance by directly activating OsDREB1B expression in rice. Plant Cell Report. 42, 223–234. doi: 10.1007/s00299-022-02950-2
Zhao, Q., Guan, X., Zhou, L., Asad, M. A. U., Xu, Y., Pan, G., et al. (2023). ABA-triggered ROS burst in rice developing anthers is critical for tapetal programmed cell death induction and heat stress-induced pollen abortion. Plant Cell Environ. 46, 1453–1471. doi: 10.1111/pce.14551
Zheng, S., Liu, S., Feng, J., Wang, W., Wang, Y., Yu, Q., et al. (2021). Overexpression of a stress response membrane protein gene OsSMP1 enhances rice tolerance to salt, cold and heavy metal stress. Environ. Exp. Bot. 182:104327. doi: 10.1016/j.envexpbot.2020.104327
Zheng, C., Zhou, J., Zhang, F., Yin, J., Zhou, G., Li, Y., et al. (2020). OsABAR1, a novel GRAM domain-containing protein, confers drought and salt tolerance via an ABA-dependent pathway in rice. Plant Physiol. Biochem. 152, 138–146. doi: 10.1016/j.plaphy.2020.04.032
Zhou, Z., Fan, J., Zhang, J., Yang, Y., Zhang, Y., Zan, X., et al. (2022). OsMLP423 is a positive regulator of tolerance to drought and salt stresses in rice. Plant 11:1653. doi: 10.3390/plants11131653
Zhou, C., Lin, Q., Lan, J., Zhang, T., Liu, X., Miao, R., et al. (2020). WRKY transcription factor OsWRKY29 represses seed dormancy in rice by weakening abscisic acid response. Front. Plant Sci. 11:691. doi: 10.3389/fpls.2020.00691
Zhou, X., Ni, L., Liu, Y., and Jiang, M. (2019). Phosphorylation of bip130 by OsMPK1 regulates abscisic acid-induced antioxidant defense in rice. Biochem. Biophys. Res. Commun. 514, 750–755. doi: 10.1016/j.bbrc.2019.04.183
Zhou, H., Wang, Y., Zhang, Y., Xiao, Y., Liu, X., Deng, H., et al. (2022). Comparative analysis of heat-tolerant and heat-susceptible rice highlights the role of OsNCED1 gene in heat stress tolerance. Plant 11:1062. doi: 10.3390/plants11081062
Zhu, M., He, Y., Zhu, M., Ahmad, A., Xu, S., He, Z., et al. (2022). ipa1 improves rice drought tolerance at seedling stage mainly through activating abscisic acid pathway. Plant Cell Rep. 41, 221–232. doi: 10.1007/s00299-021-02804-3
Zhu, A., Li, J., Fu, W., Wang, W., Tao, L., Fu, G., et al. (2022). Abscisic acid improves rice thermo-tolerance by affecting trehalose metabolism. Int. J. Mol. Sci. 23:10615. doi: 10.3390/ijms231810615
Keywords: altered gene expression, dormancy, electrolyte leakage, genome editing, ion toxicity, omics, plant architecture, reactive oxygen species
Citation: Gul RMS, Rauf S, Ortiz R, Waqas Khalid M and Kaya Y (2024) Understanding abscisic acid-mediated stress signaling to affect rice development under stress. Front. Sustain. Food Syst. 8:1477994. doi: 10.3389/fsufs.2024.1477994
Edited by:
Pushp Sheel Shukla, Sea6 Energy Private Limited, IndiaReviewed by:
Avishek Dey, KRYSP-R LLC, IndiaCopyright © 2024 Gul, Rauf, Ortiz, Waqas Khalid and Kaya. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rodomiro Ortiz, cm9kb21pcm8ub3J0aXpAc2x1LnNl
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.