 Sejin Cheong1
Sejin Cheong1 Carolyn Chandler-Khayd1
Carolyn Chandler-Khayd1 Sequoia R. Williams2
Sequoia R. Williams2 Amélie C. M. Gaudin2
Amélie C. M. Gaudin2 Peiman Aminabadi3
Peiman Aminabadi3 Michele T. Jay-Russell3
Michele T. Jay-Russell3 Emily Evans4Lee Klossner4
Emily Evans4Lee Klossner4 Paulo Pagliari4
Paulo Pagliari4 Patricia Millner5
Patricia Millner5 Annette Kenney6
Annette Kenney6 Fawzy Hashem6
Fawzy Hashem6 Amber R. Sciligo7
Amber R. Sciligo7 Alda F. A. Pires1*
Alda F. A. Pires1*- 1Department of Population Health and Reproduction, University of California-Davis, Davis, CA, United States
- 2Department of Plant Sciences, University of California-Davis, Davis, CA, United States
- 3Western Center for Food Safety, University of California-Davis, Davis, CA, United States
- 4Department of Soil, Water, and Climate, College of Food, Agriculture and Natural Resources Sciences, University of Minnesota, Saint Paul, MN, United States
- 5ARS-Environmental Microbial and Food Safety Laboratory, U.S. Department of Agriculture, Beltsville, MD, United States
- 6Department of Agriculture, Food, and Resource Sciences, University of Maryland Eastern Shore, Princess Anne, MD, United States
- 7The Organic Center, Washington, DC, United States
Introduction: Integrated crop-livestock systems (ICLS) improve soil health and productivity but may lead to the transfer of foodborne pathogens to fresh produce from soil contaminated with the feces of grazing animals.
Methods and results: Over 2 years (2021–2022), organic ICLS field trials were conducted in California (CA) and Minnesota (MN) to examine the presence of foodborne pathogens (Escherichia coli O157, non-O157 Shiga toxin producing E. coli (STEC), and Listeria monocytogenes) across three treatments (fallow, cover crop without grazing, and cover crop with grazing by small ruminants) in soils and produce. A Random Forest (RF) analytical approach was used to determine potential associations between meteorological and soil chemical factors, and the generic E. coli (gEc) presence in soil. The persistence of gEc in soil, as an indicator of fecal contamination, was evaluated using mixed effect zero-inflated negative binomial (ZINB) models, considering top-ranked meteorological factors identified from RF analyses. One produce sample (cucumber) tested positive for non-O157 STEC (0.6%, 1/157) from grazed soil in MN, with no other foodborne pathogens detected in produce. Soil contamination by non-O157 STEC increased in 2022 (2.7%, 15/552) compared to 2021 (0%, 0/504) from both states, aligning with the increased prevalence of non-O157 STEC in post-grazed fecal samples from sheep in CA (41.7%, 5/12) and goats (42.5%, 17/40) in MN in 2022. Concentrations (Most Probable Number/100 g) of gEc in grazed soil returned to levels comparable to non-grazed or fallow treatments within 87–147 days post-graze (DPG) in both years, with a significant decrease predicted after 32 DPG. Interestingly, non-O157 STEC was detected in soil even after concentrations of gEc declined 115–147 DPG.
Discussion: Although the effects of meteorological factors and soil chemical characteristics were not as influential as treatment or sampling day effects, ZINB analyses with identified meteorological factors in grazed soil suggested that regional differences in gEc counts were likely influenced by maximum air/soil temperatures on the sampling day. Further investigation is needed to evaluate the adoption of the NOP 90-120-day interval rules between manure application by grazing and harvest, considering a wider range of environmental regions and the potential cumulative effect of continuous ICLS in the same field on pathogen loads.
1 Introduction
Agroecological practices focus on building the resilience of ecosystems and supporting more sustainable food production systems (Schipanski et al., 2016; Newton et al., 2020). These practices include minimizing soil disturbance and enhancing biodiversity to build fertility, soil health, and pest control mechanisms (Rhodes, 2017; LaCanne and Lundgren, 2018). Organic and regenerative farms have increasingly coupled the use of cover crops and livestock grazing known as integrated crop-livestock systems (ICLS), which aim to enhance soil fertility and improve carbon storage and water retention (Franzluebbers et al., 2011; Hilimire, 2011; Lemaire et al., 2014; Schipanski et al., 2014). This study focused on a rotational form of ICLS in organic fresh produce production, which involves animal grazing of winter cover crops seeded between growing produce within a single field. In this system, manure deposited by the animals and remaining cover crop residue are incorporated into the soil as organic sources of fertilizer before planting produce crops.
Despite various ecological benefits, animal grazing in ICLS poses a risk of transferring foodborne pathogens to produce crops via direct contact with soil (Kudva et al., 1998; Park et al., 2012; Nazareth et al., 2021). Raw manure from livestock naturally carries enteric pathogenic bacteria including Shiga toxin-producing Escherichia coli (STEC, such as E. coli O157:H7), Salmonella spp., and Listeria spp., which can cause severe illness and even death in humans (Hutchison et al., 2005; Franz et al., 2008; Alegbeleye et al., 2018). Application of livestock manure, including biological soil amendments of animal origin (BSAAO), is recognized as one of the major sources of preharvest bacterial contamination in fresh produce production (Park et al., 2015; Sharma and Reynnells, 2016; Pires et al., 2018). These types of soil amendments are also important inputs for the United States Department of Agriculture (USDA) certified organic producers who are not allowed to use synthetic fertilizers (USDA, 2024). Certified organic farms operate under the standards established by the USDA National Organic Program (NOP) when applying BSAAO onto their fields. The NOP stipulates a time-interval of 90–120 days between the application of raw manure and the harvest of fresh produce to minimize the risk of bacterial contamination (USDA-AMS NOP, 2011). The US Food and Drug Administration (FDA) also addresses concern regarding the use of BSAAOs applied to fresh produce fields through the Produce Safety Rule of the Food Safety Modernization Act (FSMA; FDA, 2015). However, the FDA has not explicitly established the differences and rules between raw manure applications and grazing due to lack of scientific evidence. Recent studies have begun to assess the risk and persistence of bacterial contamination in raw manure-amended fields (Sharma et al., 2019; Tran et al., 2020; Ramos et al., 2021; Pires et al., 2023).
Previous studies have shown that meteorological factors (i.e., precipitation, air temperature, and wind speed) and soil characteristics (i.e., soil type, soil moisture content, and soil chemical properties) by geographical regions are risk factors for contamination of foodborne pathogens in fresh produce fields (Ivanek et al., 2009; Ongeng et al., 2011; Strawn et al., 2013; Sharma et al., 2019; Weller et al., 2020; Litt et al., 2021; Cook et al., 2023). In particular, multiple studies reported that average precipitation (i.e., rainfall and snowfall) before soil or produce sample collection showed a significant association with the contamination and survival of STEC, Listeria spp., and generic E. coli (gEc) (Park et al., 2015; Pang et al., 2017; Litt et al., 2021; Pires et al., 2023). According to a meta-analysis compiling 42 studies investigating environmental factors regulating persistence of E. coli in manure-amended soils, temperature was the most influential factor affecting decline rates of E. coli under field conditions (Tran et al., 2020). Among soil characteristics, soil moisture content and temperature were the most determinant factors on the duration of manure-borne pathogen survival in soil (Park et al., 2016). Soil pH and temperature were also identified as critical factors influencing survival of both commensal and pathogenic E. coli in soil (Franz et al., 2014).
Although studies related to the application of raw manure or BSAAO in fields have been conducted in various regions, little research is present regarding the potential risk associated with directly implementing grazing in ICLS fields (Ingham et al., 2004; Nazareth et al., 2021; Goodwyn et al., 2023). Our previous observational and experimental ICLS studies conducted in California revealed that foodborne pathogen contamination of produce by grazing events was minimal, and the concentration of gEc in soil, serving as an indicator of fecal contamination, returned to the baseline levels within 120 days post-grazing (Patterson et al., 2018; Cheong et al., 2024). However, these grazing studies did not consider various agricultural and environmental factors such as meteorological elements and soil characteristics, which may influence the persistence and survival of bacterial pathogens in soil.
To provide detailed guidelines for minimizing contamination of foodborne pathogens in ICLS, more research is needed across diverse regions to clarify various environmental risk factors. In this study, we examined the risk of grazing on the contamination of soil and fresh produce by foodborne pathogens under two different environmental conditions in California (CA) and Minnesota (MN). Two-year ICLS experimental studies were conducted to evaluate the presence of foodborne pathogens (E. coli O157, non-O157 STEC, and L. monocytogenes) in soil and produce, as well as in fecal samples from grazing animals, across three different treatments (fallow, cover crop without grazing, and cover crop with grazing by small ruminants). Persistence of gEc was further assessed through monthly quantification of gEc in grazed soil, while potential risk factors related to meteorological conditions and soil health influencing the presence of gEc in soil were examined using a machine learning approach.
2 Materials and methods
2.1 Study design
Replicated field trials were established as randomized complete block designs with four replicates in organically certified fields in two states (CA and MN) over two growing seasons (2020–2022). Fields were located at the Russell Ranch Sustainable Agriculture Facility, University of California, Davis (UC Davis; 38° 32′ 36.87″, −121° 52′ 11.89″) and at the University of Minnesota Southwest Research and Outreach Center (44° 14′ 29.4858″, −95° 19′ 1.5312″). Three treatments—fallow as a control, cover crop tilled without grazing (non-graze CC), and cover crop grazed by sheep or goats (graze CC) were randomly allocated within each block. Each block corresponded to 9 beds (13.5 m wide), and 3 beds in a row were assigned to each treatment. A middle bed (1.5 m wide) of each treatment was used for sampling, as the two side beds functioned as a buffer between treatments.
The timing of cover crop mix seeding, sheep/goats grazing, and planting of the main produce crops (spinach in 2021, and cucumber in 2022) depended on each state (CA and MN) climate and growing seasons. Both states followed the cropping sequence that is generally adopted by regional organic vegetable growers. The cover crop mix was seeded in November for the CA trial and in late August or early September for the MN trial. The cover crop mix in CA consisted of cereal rye (60 lbs./ac), crimson clover (10–12 lb./ac), and daikon radish (10 lbs/ac), while in MN the mix comprised winter rye (1 lbs/ac), berseem clover (10 lbs/ac), and daikon radish (10 lbs/ac). Grazing was initiated when the cover crops height was 25 cm. After grazing events, beds of graze CC and non-graze CC treatments were terminated, and the main crops of spinach/cucumber were transplanted/seeded in March–April in CA, and in May–June in MN.
For grazing, CA and MN trials used sheep and goats, respectively. The sheep originated from the sheep facility located at UC Davis, and goats were from a goat rental company in MN, which was located 2 h from the field. Electrical fences were installed around the areas of the graze CC treatment before sheep or goats were transported to the field. The number of grazing animal units and period of grazing were decided based on the amount of cover crop biomass available for grazing each time and location (USDA-Natural Resources Conservation Service, 2013). As a result of growing conditions, grazing was conducted once in CA but twice in MN per year. While grazing at CA, a flock of 49 sheep grazed once during March 2–4, 2021, and 25 sheep grazed once on February 17, 2022. The flock was composed of ewes (e.g., Suffolk, Hampshire, Dorset, and crossbreed) with ages from 1 to 5 years old. For MN, in the first year of the study, a herd of 40 goats grazed on November 4, 2020, and 115 goats grazed on April 28–30, 2021. In the second year, grazing took place twice and 160–170 goats grazed during October 6–8, 2021, and again on May 20, 2022. The herd comprised Spanish goat breed in age from yearlings to up to 10 years old. Sheep or goats grazed rotationally from one paddock (13.5 × 22.9 m) to another to achieve equalized levels of biomass (approximately 10 cm). Grazing animals were provided ad libitum water, and lights (CA) or a guard dog (MN) to protect them from wildlife when they stayed overnight at the field. The research protocol was approved by the Institutional Animal Care and Use Committees (IACUC) of the University of California, Davis (IACUC protocol #22700) and the University of Minnesota (IACUC protocol #2008-38348A).
2.2 Sample collection
A total of 1,056 soil samples were collected during the seven or eight field visits each year, including the baseline after seeding winter cover crops, just before grazing [0 Day Post-Graze (DPG)], 7 DPG, and monthly after the grazing event until approximately 120 or 150 DPG. Figure 1 describes the specific soil sampling and grazing dates of the field trials in CA and MN, with the number of samples collected on each date. When weather conditions did not allow (i.e., heavy rain or snow), sampling was performed within 1–2 weeks from the planned DPG dates. Grazing events were conducted twice in the MN trials each year; thus, DPG dates were calculated based on the second grazing event. On each sampling day, 36 soil samples were collected as three composite soil samples per bed (12 beds × 3 composites), except for the baseline (i.e., 24 samples as two composite soil samples per bed) and 7 DPG in 2021 in MN (i.e., 60 samples as five composite soil samples per bed). Soil samples were collected from the middle bed of each treatment using a stainless-steel soil core sampling probe (15 cm deep × 2.54 cm diameter) and placed in sterile Whirl-Pak bags (Nasco, Modesto, CA). Gloves were changed and the soil core was cleaned then sanitized with 70% ethanol between each bed.
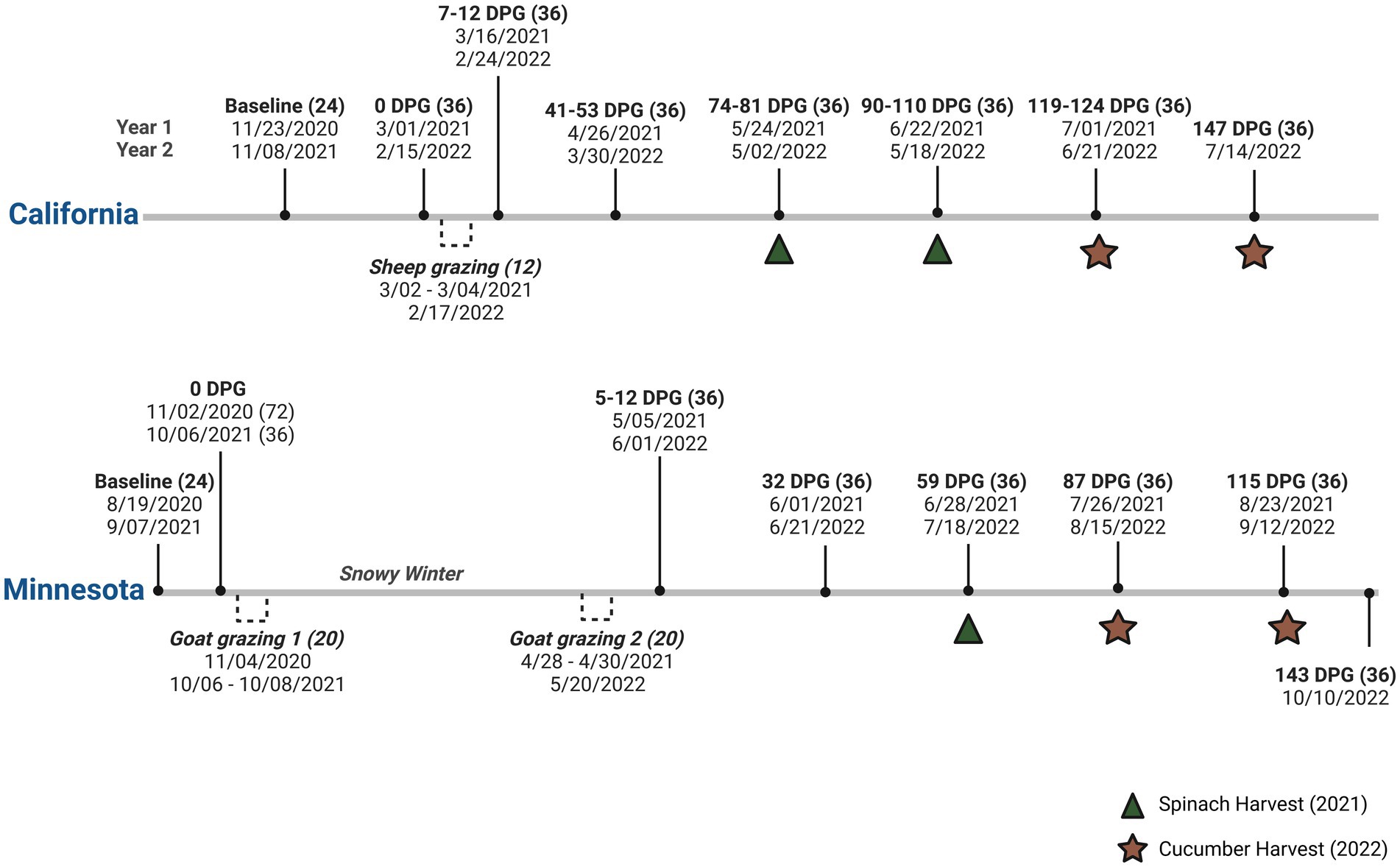
Figure 1. Soil sampling dates and grazing dates in organic integrated crop-livestock systems field trials in California and Minnesota (2021–2022). Days post-graze (DPG) was calculated based on the last day of the grazing event in each year, and the number of collected soil samples was specified next to each date (Biorender).
Pre-and post-grazing fecal samples were collected to assess the presence of foodborne pathogens (i.e., Non-O157 STEC, L. monocytogenes, and E. coli O157). Pre-graze fecal samples were collected from the barn floor (CA) or trailer floor (MN) before sheep or goats were transported to the field. Post-graze fecal samples were collected immediately after sheep or goats were removed from the graze CC treatment plots on the same day. A total of 36 sheep fecal samples from CA (i.e., 12 samples per each sampling) and 160 goat fecal samples from MN (i.e., 20 samples per each sampling) were collected over the two growing seasons. Pre-graze samples from the second growing season of the CA trial were not collected because the grazing event lasted less than a day. Fresh fecal samples (approximately 30 g of composite fecal material) were aseptically scooped from the barn/transporter or field using gloves and placed into a sterile Whirl-pack bag.
Fresh produce samples (approximately 100 g for spinach and 400–500 g for cucumber per sample) were collected once or twice each year depending on produce maturity. A total of 48 spinach and 47 cucumbers samples were harvested twice each year from CA (Figure 1). In Minnesota, severe heat and drought conditions limited spinach harvest to a total of 14 samples in the first year. In the second year, conditions were normal and a total of 48 cucumber samples were collected from the MN site. On each sampling day, up to two samples per bed (12 beds × 2 samples) of produce were collected from the middle bed of each treatment, as had been done for soil sampling.
Irrigation water (2 L) was sampled one time during each growing season directly into two sterile bottles from tap water linked to irrigation lines.
All samples were transported in a cooler or shipped in insulated containers with ice to the Pires laboratory at UC Davis within 24 h of collection, and processed within 48 h.
2.3 Sample preparation, and isolations of foodborne pathogens and generic E. coli (gEc)
Sample preparation, microbial analyses for the isolations of non-O157 STEC, E. coli O157:H7, L. monocytogenes, and quantification of gEc were described in our prior manuscript (Cheong et al., 2024).
In brief, each soil (30 g per sample) and fecal sample (10 g per sample) was enriched in a 24 oz. Whirl-Pak bag filled with 270 ml Tryptic Soy Broth (TSB; BD BactoTM, Heidelberg, Germany) to detect non-O157 STEC, E. coli O157:H7, and gEc, and Listeria Enrichment Broth (LEB; Neogen Culture Media, Lansing, MI, United States) to detect L. monocytogenes. To quantify the concentration of gEc (Most Probable Number, MPN/g) in the soil and produce samples, a 48-well reservoir (E&K Scientific, Santa Clara, CA, United States) was filled with 5 ml of the samples in TSB medium, with serial dilution up to 10−6 with 4 replications per sample. For the TSB enrichment, samples were incubated at 25 °C for 2 h followed by 42 °C for 8 h with 50 rpm shaking, then held at 6 °C with no shaking in a Multitron programmable shaking incubator (Eppendorf, Hauppauge, NY, United States). For the LEB enrichment, samples were incubated for 18 h at 30 °C with 100 rpm shaking, then held at 6 °C with no shaking.
To isolate non-O157 STEC, 1 ml of enriched TSB sample was put into a tube with 9 ml of modified enterohemorrhagic E. coli (mEHEC) selective media (Biocontrol, Bellevue, WA, United States), and incubated for 12 h at 42 °C. Then, mEHEC solution was streaked onto ChromSTEC agar (CHROMagarTM, Paris, France) using a 10 μl inoculation loop, and incubated for 24 h at 37 °C. Presumptive positive isolates (i.e., purple colonies that fluoresced under ultraviolet light) on ChromSTEC agar were re-streaked onto secondary and tertiary ChromSTEC agar. A final pure presumptive positive colony was streaked onto Tryptic Soy Agar (TSA) to confirm as non-O157 STEC by conducting a standard polymerase chain reaction (PCR) assay targeting the stx1 and stx2 genes.
To detect E. coli O157:H7 and L. monocytogenes, Immunomagnetic separation (IMS) was performed with enriched TSB and LEB samples using Dyna anti-E. coli O157 or Listeria beads on the automated Dyna Bead Retriever (Invitrogen, Carlsbad, CA), respectively. For E. coli O157:H7, an IMS product of 50 μl washed beads streaked onto both Rainbow agar (Biolog, Hayward, CA, United States) with novobiocin (20 mg/L) and tellurite (0.8 mg/L; MP Biomedicals, Solon, OH, United States), and MacConkey II Agar with sorbitol supplemented with potassium tellurite (2.5 mg/L) and cefixime (0.05 mg/L); both plates were incubated for 24 h at 37 °C. For L. monocytogenes, an IMS product of 30 μl washed beads streaked onto Brillance Listeria Agar (BLA) with its selective and differential supplements (Oxoid, Hants, United Kingdom), and that of 100 μl was added into 5 ml of Fraser broth (BD, Sparks, MD); both BLA plates and Fraser broths were incubated for 48 h at 37 °C. With sequential insolation process, a final pure presumptive positive colony was streaked onto TSA, and PCR targeting the eaeA and hylA gene was performed to confirm the presence of E. coli O157:H7 and L. monocytogenes, respectively.
To determine the presence and enumeration of gEc, 10 μl from the enriched TSB bag and 4 μl from each diluted well of the reservoir were streaked onto CHROMagar E. coli (ECC; CHROMagar Microbiology, Paris, France), followed by incubation for 24 h at 37 °C. After re-streaking presumptive positive colonies (i.e., blue colonies) onto secondary and tertiary ECC plates, the pure isolates on TSA were confirmed as gEc by PCR targeting the uspA gene.
2.4 Collection and modification of meteorological and soil health data
Meteorological data were obtained from the weather stations located approximately 1 km from the experimental sites in CA and MN. Additionally, solar radiation data for CA were retrieved from the Davis station of California Irrigation Management Information Systems (not recorded at the field weather station). The 80 meteorological variables consisted of 16 original weather variables on each soil sampling day (Table 1) and 64 calculated mean or sum variables depending on different time intervals. Different time intervals including 0–3 days, 0-7 days, 0–20 days, and 0–30 days before the soil sampling days were chosen to evaluate whether meteorological variables had immediate or long-lasting effects on the presence of gEc in soil (Ivanek et al., 2009; Park et al., 2015; Pang et al., 2017).
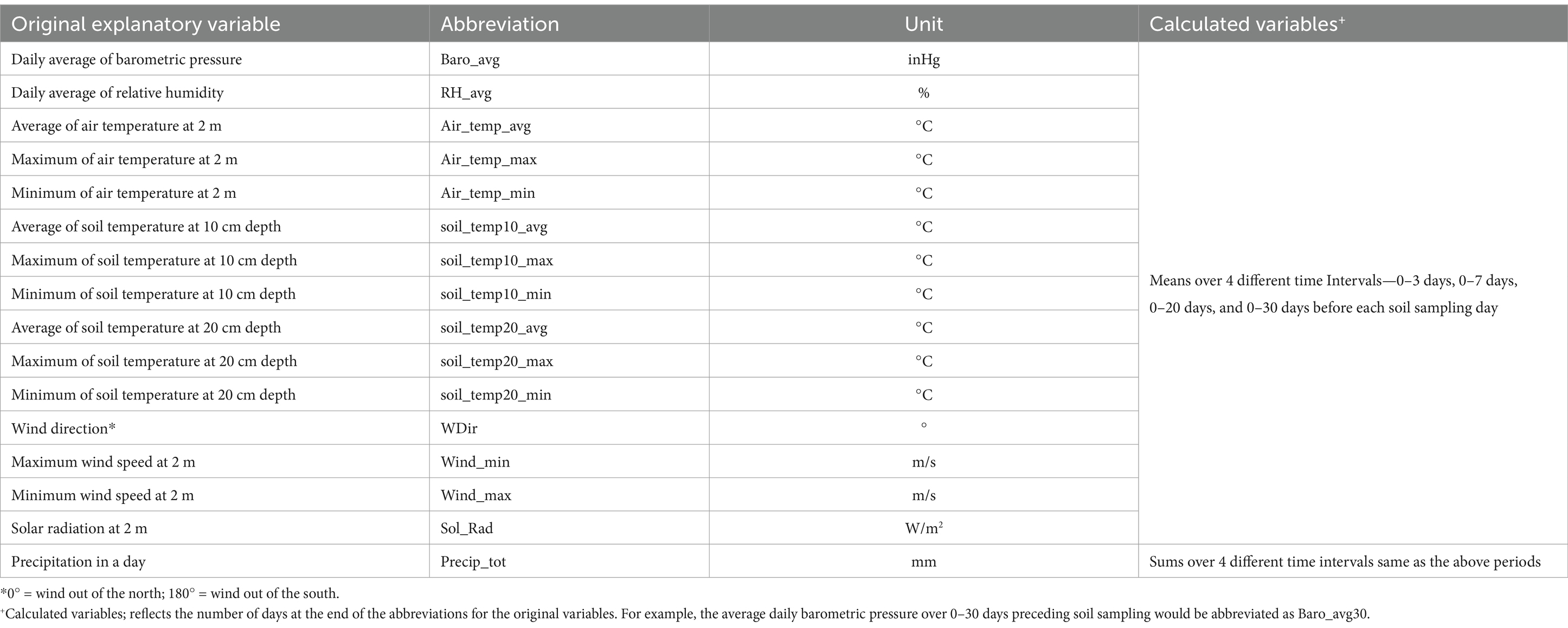
Table 1. Explanatory meteorological variables considered as potential risk factors for generic E. coli presence in soil from organic integrated crop-livestock system field trials in California and Minnesota (2021–2022).
To investigate soil characteristics, soil sampling was conducted from each of 12 beds (i.e., 3 treatments × 4 blocks) five times in CA including the baseline (December 1, 2020) and twice during each growing season (March 29 and May 3, 2021, and April 8 and June 6, 2022), and twice (June 28, 2021, and August 9, 2022) in MN over 2 years. Soil samples (~ 700 g per sample) were submitted to the University of Maine Analytical Laboratory and Maine Soil Testing Service to obtain routine soil bio-physical and chemical properties. Soil chemical properties used for further analyses included 20 variables—soil pH, percentages (%) of organic matter (OM), potassium (K), magnesium (Mg), and calcium (Ca), and concentrations (ppm) of nitrate (NO3), ammonium (NH4), aluminum (Al), boron (B), copper (Cu), iron (Fe), Manganese (Mn), sodium (Na), sulfur (S), zinc (Zn), cation exchange capacity (CEC), and the concentrations (lb) of phosphorus (P), K, Mg, and Ca.
The moisture content of each soil sample was calculated based on the weights of wet and dry soil (g) with the Equation 1. The dry weight of soils reflects oven (105 °C) grams dry weight (gdw) of 10 g fresh weight of soil sample.
2.5 Statistical analysis
Proportions of foodborne pathogens (E. coli O157:H7, non-O157 STEC, and L. monocytogenes) positive in soil, produce, and fecal samples were summarized using descriptive statistics. For soil and produce samples, concentrations of gEc (log10 MPN/100gdw soil) were reported as median and range (min–max) after transforming the values under the limit of quantification (LOQ = 0.089 MPN/gdw) as 0.0445 MPN/gdw (LOQ/2). To compare the median concentrations of gEc in soils across the three treatment groups on each sampling day, Kruskal-Wallis tests were used as non-parametric tests, due to non-normal distributions observed in gEc concentrations.
Two random forest (RF) model analyses (1) based on 80 meteorological factors (Table 1) and (2) 20 soil chemical factors were conducted to identify and rank risk factors associated with the presence of gEc in soils. As additional explanatory variables, state, year, treatment and sampling days as categorical variables and soil moisture content as a continuous variable were added into each model. The dataset for the first model incorporating meteorological factors included 29 timepoints with 1,032 soil sample profiles; the dataset for the second model integrating soil characteristics factors included seven timepoints with 257 soil sample profiles. The presence of gEc data for the second model were extracted from sample profiles of the first model corresponding to the nearest dates of soil characteristics testing, because microbial analyses to investigate the soil pathogens were not conducted simultaneously with soil samplings for characteristics analyses. Each dataset was randomly split into training (80%) and testing sets (20%), and stratified sampling was performed based on the presence of gEc to make the sets balanced. The RF models were run using the randomForest package in R software (v.4.2.0), and the importance of the explanatory variables in the models was evaluated by permutation feature importance scores (Liaw and Wiener, 2002). To decrease the number of variables in the model, feature elimination was performed based on permutation feature importance scores until the out-of-bag error rate ceased to decrease, and none of the variables were highly correlated (i.e., less than 0.85) by dropping the use of variables from the same explanatory categories but within different time intervals. To run each model, ntree (i.e., number of trees) and mtry (i.e., number of randomly drawn candidate variables) were set to 500 and the square root of the number of explanatory variables, respectively. The predictive performance of each model was assessed by the remaining 20% testing sets and subsequently evaluated with a receiver operating characteristic (ROC) curve.
To evaluate the effect of sheep/goats grazing on the soil concentrations of gEc over time, particularly within the graze CC treatment (i.e., 319 samples), mixed effect zero-inflated negative binomial (ZINB) models were used after excluding one extreme outlier (i.e., 94,923 MPN/gdw). The outcome variable of gEc concentration (MPN/100gdw) was log10–transformed and rounded off to the nearest integer if the data were non-zero. Explanatory variables considered in the model were state, year, and DPG. State and year were categorical variables, and the DPG variable was a spline linear continuous variable with the knot at 32 DPG because the highest concentrations of gEc was mostly observed at 32 DPG, followed by a decrease thereafter. Beds of the fields (i.e., 144 total beds) in each year of each state were added as random effects in the models to explain repeated sampling of soil over time. Final multivariable modeling was conducted with variables showing p-value <0.2 in univariable analyses by forward selection and included a two-way interaction between state and DPG. All the modeling was run using the glmmTMB (generalized linear mixed models Template Model Builder) package with nbinom1 negative binomial distribution, in which the variance increases linearly with the mean (Brooks et al., 2017). The data collected before each growing season (i.e., November in CA and August/September in MN) as the baselines were reported separately and were not included in the modeling. Furthermore, the seven top-ranked meteorological variables from the RF analysis were individually added into the final ZINB model to evaluate their associations. All the statistical analyses were performed using R software (v.4.3.0) and the significance level was p-value <0.05.
3 Results
3.1 Presence of foodborne pathogens
None of the collected soil nor fecal samples from either state tested positive for E. coli O157:H7 during the 2 years of the study period. Table 2 summarizes the presence of non-O157 STEC and L. monocytogenes in the soil and fecal samples for each treatment group. Of the collected post-graze fecal samples in CA and MN, 20.8% (5/24) and 22.5% (18/80) were non-O157 STEC positive, respectively. Whereas none of the sheep fecal samples collected from CA tested positive for L. monocytogenes, 15.0% (12/80) of the post-graze fecal samples from goats in MN trials tested positive for L. monocytogenes.
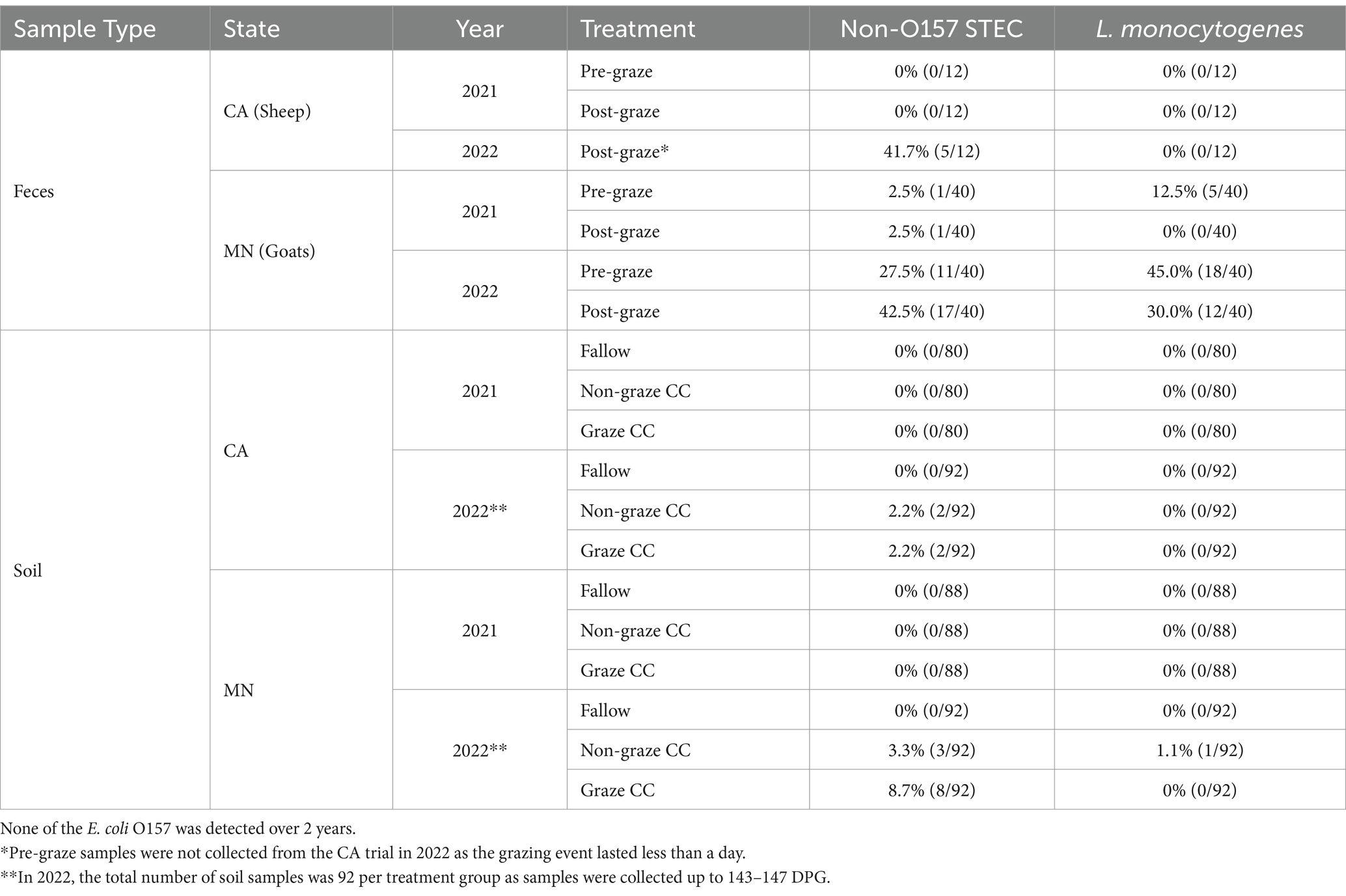
Table 2. Presence of foodborne pathogens (non-O157 STEC and L. monocytogenes) in fecal and soil samples (%) collected from the organic integrated crop-livestock system field trials (2021–2022) in California and Minnesota.
In the first year (2021), none of the soil samples tested positive for foodborne pathogens. However, in 2022 trials, 1.4% (4/276) and 4.0% (11/276) of the soil samples tested positive for non-O157 STEC from CA and MN, respectively. Of four non-O157 STEC positive samples from the CA field, two from the graze CC treatment plots were collected at 0 and 147 DPG, and the other two from the non-graze CC treatment plots were also collected at 0 and 147 DPG. Of 11 non-O157 STEC positive samples from MN field, eight samples from the graze CC treatment plot were collected at 32 DPG, and the three samples from the non-graze CC treatment plots were collected at 32 and 115 DPG. In addition, one sample (0.4%, 1/276) from MN tested positive for L. monocytogenes, which was from the non-graze CC treatment plot at 12 DPG.
One cucumber sample (0.6%, 1/157) tested positive for non-O157 STEC collected from one graze CC treatment plot at 115 DPG in MN trials. None of the other produce samples (i.e., spinach and cucumber) from either state tested positive for foodborne pathogens (i.e., non-O157 STEC, E. coli O157, L. monocytogenes).
3.2 Presence and quantification of generic E. coli (gEc) in soil and produce
3.2.1 Soil
Over the 2 years of the field trials, 39.8% (86/216) and 35.7% (90/252) of soil samples from CA, and 25.4% (61/240) and 44.8% (113/252) of soil samples from MN tested positive for gEc. The concentrations of gEc (log10 MPN/100gdw) in graze CC soil were similar between CA (mean = 1.35, sd = 1.07) and MN (mean = 1.49, sd = 1.16). Sampling days that showed the highest soil concentration of gEc among three treatment groups (i.e., fallow, non-graze CC, and graze CC) were between 12 and 53 DPG in the graze CC treatment in both states (Figure 2). No statistically significant differences were observed in the median concentrations of gEc (log10 MPN/100gdw) among three treatment groups after 53–59 DPG in both states in the 2021 trials. In the 2022 trials, CA trial showed significant differences in the median concentrations of gEc among treatment groups until 124 DPG, whereas MN trial showed the difference at 12, 32, and 87 DPG.
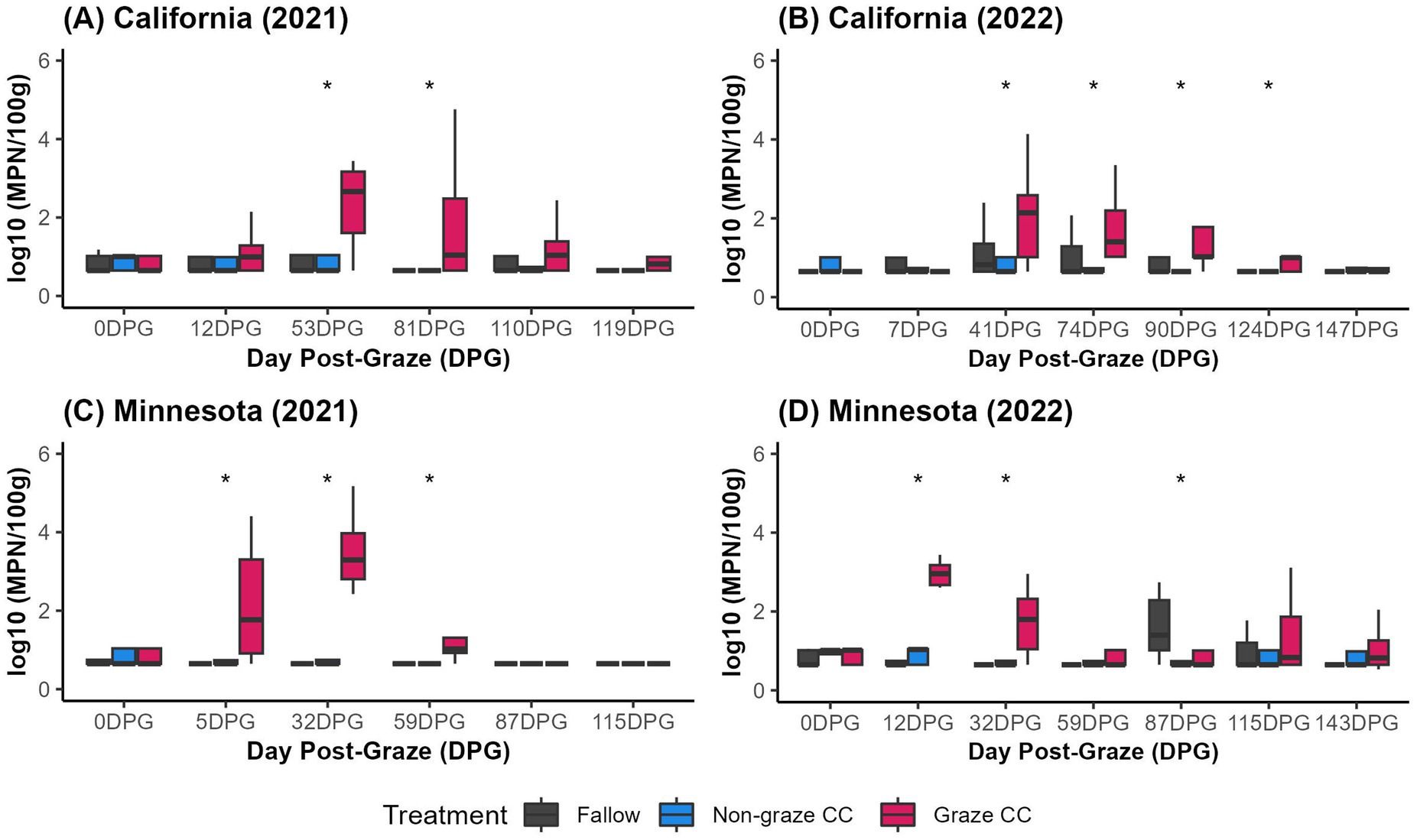
Figure 2. Soil concentrations of generic E. coli (log10 MPN/100gdw) by Day Post-Graze (DPG) from the organic crop-livestock integration systems field trials in (A) California in 2021, (B) California in 2022, (C) Minnesota in 2021, and (D) Minnesota in 2022 (*p-value <0.05).
Interestingly, the baseline soil samples, collected after seeding winter cover crops in the fields 1–4 months ahead of conducting grazing, showed higher proportions of gEc than soil samples collected at 0 DPG right before grazing events. In CA, proportions of gEc positive at baseline were 75.0% (36/48) although those at 0 DPG were 29.2% (21/72) for 2 years. Similarly, proportions of gEc positive in MN were 70.8% (34/48) at baseline, and 39.6% (38/96) at 0 DPG.
3.2.2 Produce
A higher proportion (79.0%, 49/62) of gEc positive was observed in produce samples (i.e., spinach and cucumber) from MN compared to those from CA (5.3%, 5/95; Table 3). The median concentration of gEc (log10 MPN/100grams fresh weight, gfw) in gEc-positive samples from MN was 1.78 (0.51–4.66), higher than that from CA which was 0.51 (0.51–1.78). However, the median concentrations of gEc between the types of produce were similar [i.e., spinach: 1.56 (0.51–2.38), cucumber: 1.36 (0.51–4.66)]. Among the five gEc positive samples from CA trials, two samples were from the graze CC treatment, and the three were from the non-graze CC and fallow treatments. In MN trials, out of 49 gEc positive samples, 17 were from the graze CC treatment, 17 from the non-graze CC treatment, and 15 from the fallow treatment.
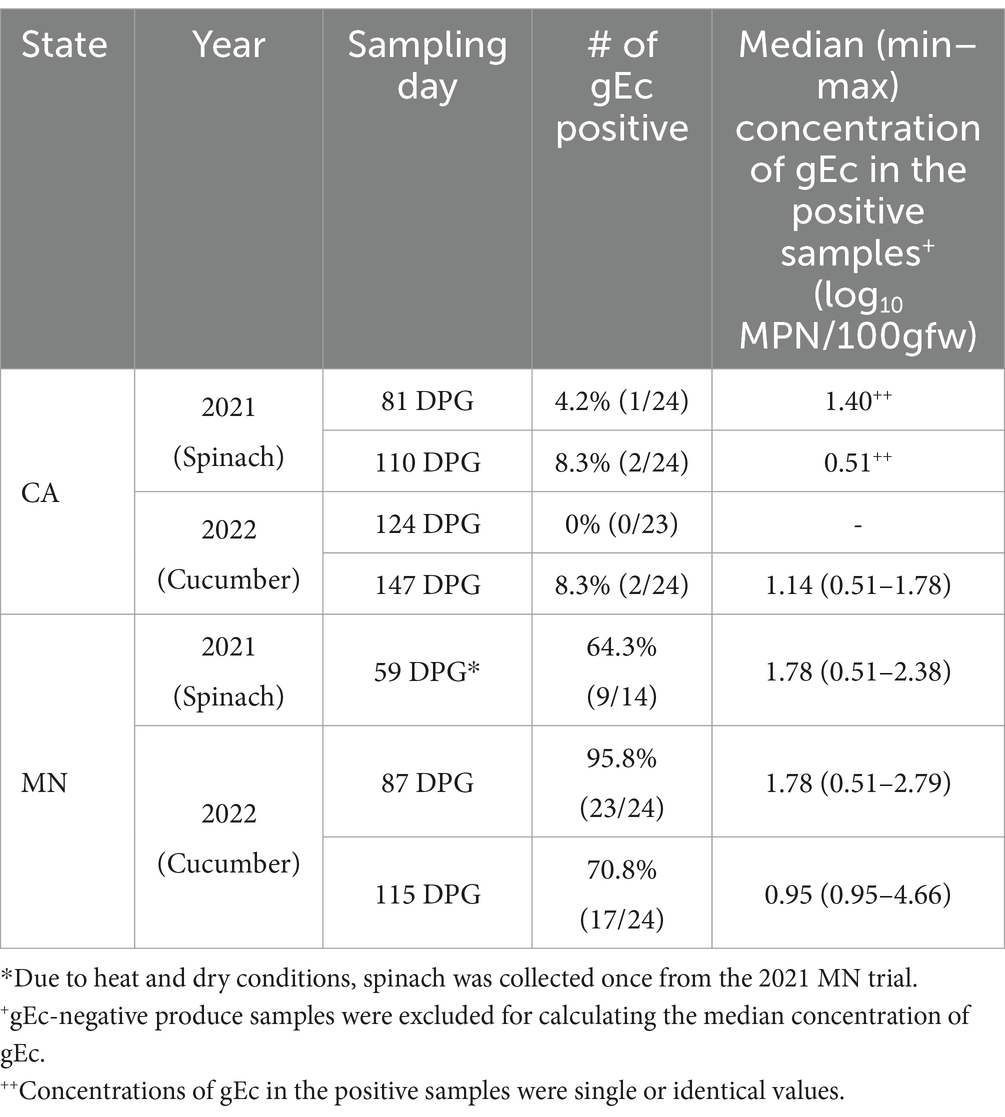
Table 3. Proportion and median (min-max) concentrations (log10 MPN/100gfw) of generic E. coli positive (gEc) in produce samples by sampling day [Day Post-Graze (DPG)] from the organic crop-livestock integration system field trials in California and Minnesota (2021–2022).
3.3 Associations between presence of generic E. coli (gEc) in soil and meteorological/soil chemical factors
In the RF analyses, the influential explanatory variables in the final models were ranked based on ‘MeanDecreaseAccuracy’, representing the percentage of gEc presence incorrectly classified by removing each variable from the models (Figure 3). In the soil sample profile used for investigating meteorological factors, 39.6% (409/1,032) of the profile were gEc positive. The most influential meteorological factors associated with the presence of gEc in the soil, according to the final RF classifier, included maximum air temperature (Air_temp_max), maximum soil temperature at a 10 cm depth (soil_temp10_max), maximum wind speed (Wind_max) on the sampling day, average wind direction for the 3 days before the sampling day (WDir3), average minimum air temperature for 30 days before sampling day (Air_temp_min30), and total precipitation for 7 days before the sampling day (Precip_tot7; Figure 3A). In the soil sample profile for soil chemical factors, 45.9% (118/257) of the profile were gEc positive. The top-ranked factors associated with the presence of gEc in the soil determined by the RF classifier were CEC, percentage of Ca, organic matter, and concentration of Na, Mn, NO3, and NH4, along with soil moisture content (Figure 3B). In both meteorological and soil chemical models, the treatment effect was identified as the most detrimental factor affecting the presence of gEc in the soil. Although the sampling day effect was not evaluated in the soil chemical RF models due to insufficient data, it was ranked second among meteorological factors (Figure 3A). Notably, in the RF models, the rank of treatment and sampling day consistently appeared as top predictors. The order of other meteorological and soil chemical factors varied yet exhibiting similar predictive performances across the training sets. When assessing the two model performances with ROC curves, the values of area under the curve (AUC) were 0.76 for the model with meteorological factors, and 0.78 for the model with soil chemical factors.
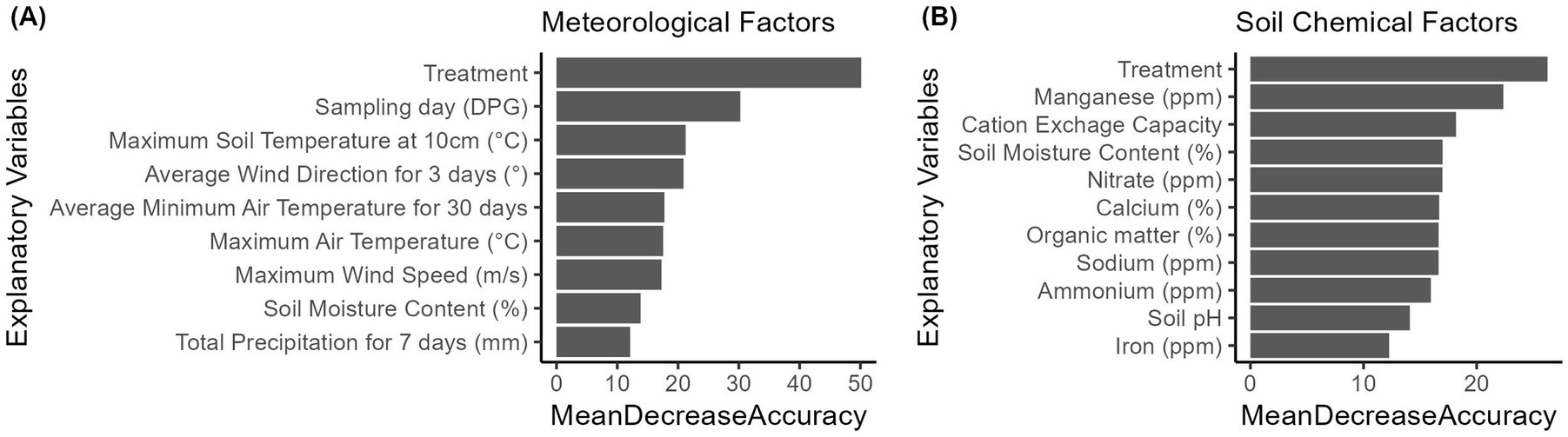
Figure 3. Results of random forest analyses to identify (A) meteorological and (B) soil chemical factors associated with the presence of generic E. coli (gEc) in the soil of the organic crop-livestock integration system field trials in California and Minnesota (2021–2022). State, year, and treatment as categorical variables and soil moisture content were added into each model, and sampling day (DPG) was additionally added into the model of (A) meteorological risk factors.
3.4 Effects of state, sampling day (DPG), and meteorological factors on soil concentrations of generic E. coli (gEc) from graze CC treatment
The effect of grazing on the concentrations of gEc over time was examined within the graze CC treatment because median concentrations (log10 MPN/gdw) of gEc in the fallow and non-graze CC treatments were consistently zero on each sample day across two growing seasons. Of the outcome variable, 43.5% were zero (i.e., gEc negative) with the high variance as 1.3 (mean = 1.1) in gEc count. State and year were not associated with the probability of zero count (i.e., zero-inflated component) and concentration (i.e., conditional component) of gEc in the univariable mixed-effect zero inflated negative binomial models. The DPG as a spline linear variable showed significant association with the concentration of gEc (i.e., log10 MPN/100gdw rounded off to the nearest integer; p < 0.001). Although state alone did not exhibit the significant association, the interaction between sampling day and state showed a significant association with the concentration of gEc (p < 0.001).
The final multivariable model included state and sampling day variables with its interaction term (Table 4). None of the variables showed significant associations with the probability of zero in gEc, but all the variables were significantly related to the concentrations (i.e., counts as integer) of gEc. Soil samples collected from MN had higher expected counts of gEc (RR = 3.43, 95% CI = 1.77–6.62) compared to CA. The application of a piecewise linear spline variable for the DPG variable showed a significant increase (RR = 1.07, 95% CI = 1.05–1.09) in mean gEc counts per day before 32 DPG and a significant decrease (RR = 0.98, 95% CI = 0.97–0.99) per day after 32 DPG in CA. Similarly, MN exhibited a significant increase (RR = 1.02, 95% CI = 1.01–1.03) and decrease (RR = 0.98, 95% CI = 0.97–0.99) per day in mean counts of gEc before and after 32 DPG.
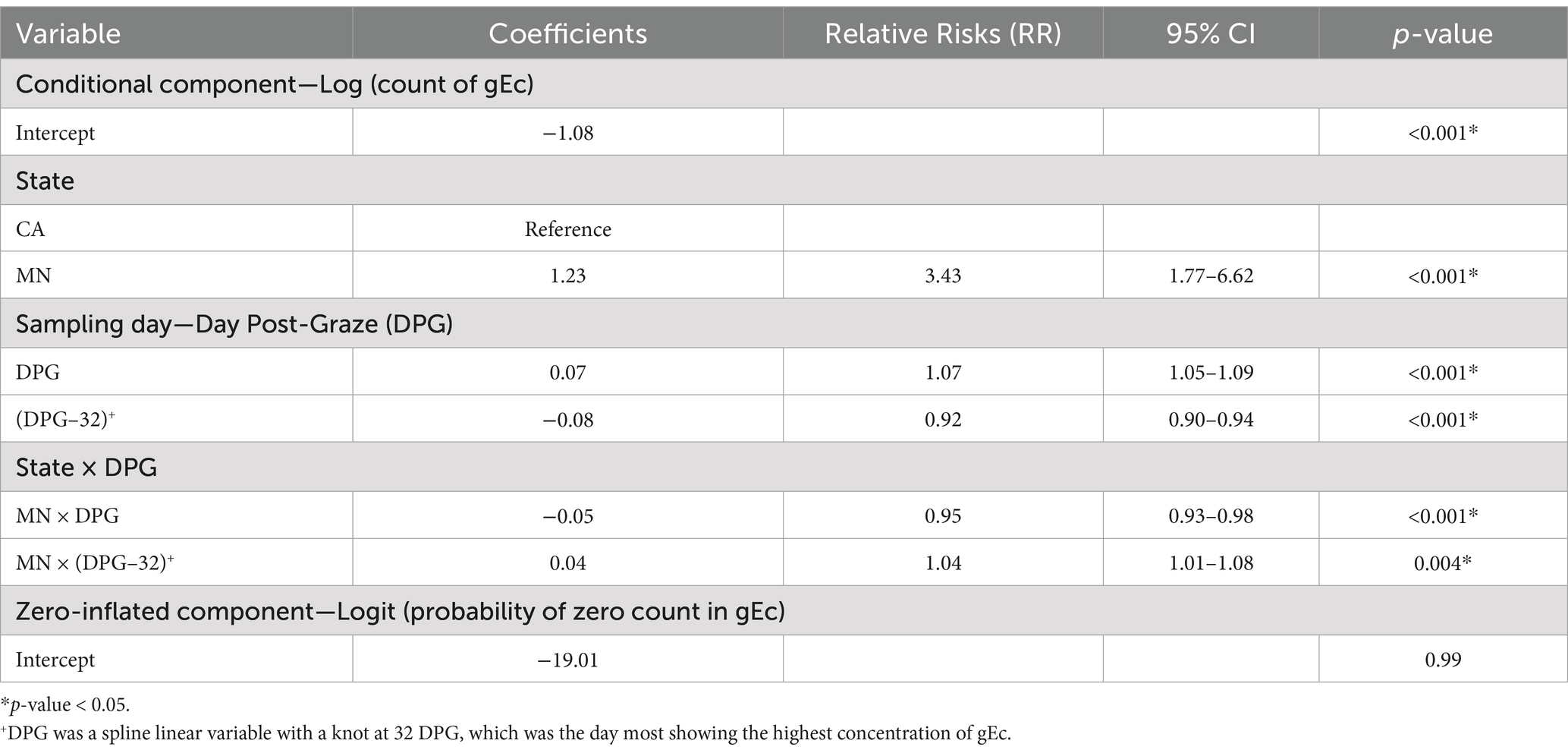
Table 4. Multivariable analyses using a mixed-effect zero-inflated negative binomial model with row as a random effect for the concentrations (i.e., log10 MPN/100 gdw rounded off to the nearest integer) of generic E. coli (gEc) in soil samples collected from graze CC treatment of the organic crop-livestock integration system field trials in California and Minnesota (2021–2022).
When the seven top-ranked meteorological explanatory variables from the RF analyses (Figure 3A) were individually added into the final model, Air_temp_max, soil_temp10_max, Wind_max, and Air_temp_min30 were all significantly negatively associated with the concentrations (i.e., counts as integer) of gEc (p < 0.001). Among those four variables, two variables—Air_temp_max and soil_temp10_max—also showed significant associations with the probability of having a zero count in gEc (i.e., absence of gEc; Supplementary Table 1). The absence of gEc in the soil was significantly associated with lower Air_temp_max (OR = 0.75, 95% CI = 0.59–0.96) and soil_temp10_max (OR = 0.64, 95% CI = 0.48–0.85), which was the opposite direction of the associations when considering the concentrations of gEc in the conditional components of the models. Interestingly, when Air_temp_max and soil_temp10_max was separately added to the final model, the interaction terms between MN and DPG or (DPG—32) became non-significant. This suggests that the regional differences in the changes in the concentrations of gEc over time may be attributed to air and soil temperatures on the sampling days. After adding the meteorological variables in the final model, the changes in the coefficient of MN were larger for Air_temp_max (31%) than for soil_temp10_max (16%), which implies that air temperature may have more influence on the concentration of gEc than soil temperature (Supplementary Table 1).
4 Discussions
In this study, ICLS field trials were conducted simultaneously under two different environmental conditions in CA and MN over 2 years (2021–2022). Among the investigated foodborne pathogens in produce and soils, a cucumber collected from MN trial and 1.4% of soil samples from both states tested positive for non-O157 STEC. The concentration of gEc in the graze CC soil returned to levels comparable to the non-graze CC or fallow treatments within 87–147 DPG over 2 years, showing a decrease after 32–53 DPG. Maximum air and soil temperatures on the sampling day were related to presence of gEc in the soil.
4.1 Presence of foodborne pathogens in the fresh produce, soil, and fecal samples
Only one non-O157 STEC (0.6%, 1/157) positive cucumber sample originated from the graze CC treatment at 115 DPG in MN, but none of the soil samples taken at the same time from these treatment plots were non-O157 STEC positive. In contrast, one soil sample from the non-graze CC treatment tested positive for non-O157 STEC at 115 DPG. Based on this, the positive cucumber sample found in the graze CC treatment may have been transferred from adjacent treatment plots by wind or originated from wildlife intrusions. Other studies noted that wind-driven dust or observed wildlife pose important foodborne pathogen transmission routes in fresh produce farms (Strawn et al., 2013; Pang et al., 2017). In soil, the three positive non-O157 STEC isolates were still detected at 115–147 DPG, despite none being identified between 32 and 115 DPG. The 115–147 DPG falls outside the NOP stipulated the waiting period (i.e., 90–120 days) between manure application and harvest. However, two out of three non-O157 STEC positive isolates were even collected from the non-graze CC treatment plots, making it difficult to conclude that positive isolates found in soil were solely from the feces deposited by the grazed animals. Similarly, the unexpected increase of non-O157 STEC prevalence in the soil after 120-days post-manure application has also been observed in a previous study (Ramos et al., 2021). Therefore, to distinguish the origin of the pathogen between the isolates from soil and cucumber, further genetic investigation (e.g., serotype, virulence factors) is needed. On the other hand, an increased presence of non-O157 STEC in soil (2.7%) was observed in the second year compared to that in year one (0%). This finding aligns with the increased prevalence of non-O157 STEC in feces deposited by grazing animals during the second year (i.e., 41.7% in CA and 45% in MN), which implies that shedding status of grazing animals may affect the presence of foodborne pathogens in the soil. These were also higher prevalences compared to a previous study; the prevalence of non-O157 STEC in sheep fecal samples in the three-year ICLS study conducted in CA was 26%, which had no detection of non-O157 STEC in grazed soil (Cheong et al., 2024). In addition, fecal pathogen shedding may have been influenced by the variable number of grazing animals in the field, which was not consistent across years and was primarily determined by the density of the cover crop biomass.
The notable difference between the two states was the higher prevalence of L. monocytogenes (37.5%, 30/80) in goat fecal samples from MN, whereas no L. monocytogenes was detected in sheep fecal samples from CA over 2 years. This disparity in fecal shedding of pathogens could be due to different species of animals, stress induced by long transportation and diets of grazing animals prior to the trials, along with different environments in the two states. Studies have shown that decreasing dietary fiber or using silage, as well as exposure to stress of long transport or cold temperature, are associated with increased fecal shedding of pathogens such as E. coli O157 and L. monocytogenes in sheep, goats and cattle (Lema et al., 2002; Bach et al., 2004; Nightingale et al., 2005; Ivanek et al., 2007; Schoder et al., 2011). In this study, sheep were transported from an adjacent sheep barn located 5.5 km away from the field in CA, while goats were transported from a goat rental company located in Faribault, MN, approximately 2 h away from the field in MN. Grazing events mostly occurred in spring in CA, while in MN, they were conducted twice per growing season, once in winter (before cover crop was covered in snow) and once in spring. Diet differences could have contributed to the shedding pattern, either because of species-specific habits or feed available to the flocks in the different states. The diet of sheep in the barn was mainly alfalfa hay with a grain rotation, although the goat diet was variable with a base of hay and other biomass that were available at the moment.
4.2 Persistence of generic E. coli and associations with meteorological and soil chemical factors
In previous studies conducted in CA, the presence of non-O157 STEC in soil samples collected under ICLS trials was rarely reported. Patterson et al. (2018) and Cheong et al. (2024) observed that none of the soil samples tested positive for non-O157 STEC. Accordingly, the persistence of gEc has been evaluated as a surrogate of pathogenic strains of E. coli in soil as other studies supported a similar survival die-off pattern and influencing factors between them (Naganandhini et al., 2015; Pang et al., 2020). However, based on the observation from the present study, 10 positive non-O157 STEC isolates from soil samples were detected during the earliest and latest days of the growing seasons, including 0, 32, and 147 DPG, from the graze CC treatment, when gEc concentrations in the soil were low. Similarly, a study measuring the prevalence of E. coli O157 and gEc in water sources used in leafy greens farms located in CA suggested that gEc concentration was not associated with the presence of E. coli O157 (Benjamin et al., 2013). A meta-regression analysis noted that the dynamics of pathogenic E. coli in the farm environment cannot be inferred from that of gEc when soil characteristics and temperatures are taken into account (Franz et al., 2014). On the other hand, a risk factors analysis study investigating the association of non-O157 STEC presence in manured soil with weather factors reported significant positive associations with gEc concentrations in the soil (Pires et al., 2023). Thus, further studies are needed on survival of gEc and pathogenic E. coli under diverse environments considering intricate interactions in nature.
When modeling the gEc concentration in soil over time in this study, the piecewise linear spline variable of DPG in the ZINB model demonstrated that a significant decrease in the mean gEc counts in soil occurred after 32 DPG within the graze CC treatment for both states, although the sampling day with the highest soil concentrations of gEc varied each year in each state (i.e., 53 DPG and 41 DPG in CA each year, and 32 DPG in MN for both years). Similarly, a two-stage survival kinetic approach was used in a meta-analysis aggregating 70 studies to evaluate the survival of manure-borne pathogen after introduction into soil (Park et al., 2016); the study supported the presence of distinct patterns of pathogen decline charactered by two different slopes. In contrast to MN, in the second year of CA trial, the graze CC treatment had higher gEc concentration than the other treatments until 124 DPG. The period of elevated gEc concentration in the grazed soil lasted longer than in our previous study in CA as well, where the graze CC treatment showed higher concentrations of gEc than the other treatments only until 61–82 DPG (Cheong et al., 2024). Given that both trials in CA were conducted in the same field, and that higher gEc concentrations in the graze CC was shorter in MN, where ICLS had not been conducted in the field before, a potential cumulative effect of repeating ICLS might be present. Therefore, the effect of continuing the ICLS over multiple years on gEc concentration in the soil and produce should be considered for future modeling.
Interestingly, despite the persistently higher concentration of gEc in the soil in CA, the presence and concentration of gEc on produce was higher in MN. Splash events after rainfall or transfer of soil pathogens to produce via irrigation (i.e., spray irrigation was used in MN fields, as opposed to drip lines in CA fields) may depend on different agricultural practices and environments (Franz et al., 2008; Litt et al., 2021). Indeed, CA fields had lower average precipitation during harvest (i.e., 0–0.01 inches) in a semi-arid climate compared to MN fields (i.e., 0.64–5 inches) in a humid continental climate. According to a quantitative microbial contamination model of E. coli in spinach at the field level, solar radiation and rain events were identified as important factors in contamination at harvest (Allende et al., 2017). Solar radiation is linked to bacterial inactivation, while rain events are associated with the splashing of soil pathogens. Therefore, the transfer of pathogens from soil to produce and their persistence in produce may be more influenced by environmental factors than by pathogen concentrations in the soil itself.
The RF model in the present study identified treatment as an agricultural factor and sampling day (i.e., DPG) as a temporal factor, both highly ranked for the soil presence of gEc. A study evaluating factors affecting survival durations of gEc in manure-amended soils in the Mid-Atlantic United States found that combined agricultural and spatiotemporal factors were better predictors than weather variables (Sharma et al., 2019). However, the state, considered as a spatial factor in this study, was not listed among the influential factors in our final RF model. This observation corresponds to findings from a repeated cross-sectional study; the state factor was also excluded from their final multivariable model with the concentration of gEc, suggesting it may serve as a proxy for weather effects (Park et al., 2015). Similarly, in the present study, the inclusion of weather-related factors (i.e., maximum air and soil temperature) in the final ZINB model led to the interaction terms between the state and sampling day becoming non-significant.
Among meteorological factors considered for the RF analyses, maximum air and soil temperatures on the sampling day were influential factors on the soil presence of gEc as well as on the counts of gEc in the ZINB model. Similarly, a meta-analysis of 42 studies on the decline of E. coli in manure-amended soil revealed that temperature, including soil and air temperatures, was the most significant factor affecting persistence of gEc under field conditions among all investigated environmental factors (Tran et al., 2020), as well as the detrimental factor on longer persistence of gEc before showing decline in soil (Park et al., 2016). This aligns with the study identifying environmental factors associated with the count of gEc on spinach at preharvest in Colorado and Texas; maximum air temperature over 9 days preceding the sampling was also the factor that determined both contamination and counts of gEc in their final model (Park et al., 2015). Interestingly, our final ZINB model in the graze CC treatment by adding individual factors of maximum air and soil temperature indicated the opposite effects of maximum air and soil temperature on the probability of gEc absence and on the concentration (i.e., counts) of gEc. In short, an excess of zero counts in gEc concentrations was observed in lower temperatures, while lower concentrations of gEc-positive were observed in higher temperatures. This implies that a specific temperature threshold may exist that determines the presence of the gEc in soil, with an optimal temperature for bacterial proliferation when present.
In the present study, the linear model analysis was not performed with soil chemical characteristic factors due to the lack of sample profile dates and non-correspondence between the dates of pathogen testing and soil chemical testing. Soil moisture content, soil pH, and soil temperature have been frequently highlighted as detrimental factors to soil survival of pathogens (Lang et al., 2007; Franz et al., 2014; Xing et al., 2019; Litt et al., 2021). Specifically, higher soil moisture and pH have been linked to longer pathogen survival in soil (Lang et al., 2007; Cook et al., 2023). Although the association between the presence of gEc and soil pH or water content was not further evaluated in this study, they consistently remained as influential in either the RF models with meteorological factors or soil chemical factors. Regarding chemical nutrient factors, eight elements listed in our final RF model (e.g., Mn, CEC, NH4, Na, OM, NO3, Fe, and Ca) appeared as important factors influencing the presence of gEc in the soil. However, intricate interactions of soil micronutrients made it difficult to interpret the results with limited research on the topic (Chandler-Khayd et al., 2023; Cook et al., 2023). Additionally, soil type is a known important factor on soil survival of bacteria, as fine soils (e.g., clay) demonstrated prolonged survival times compared to coarse soils (e.g., sandy; Franz et al., 2008, 2014; Brennan et al., 2014), as different classification of soil types between CA and MN may have effect on the presence of gEc.
5 Conclusion
The foodborne pathogens (E. coli O157, non-O157 STEC, and L. monocytogenes) were rarely or completely not detected in soil and produce from organically certified production fields where small ruminant grazing was directly implemented in the current study. Our findings suggest that screening for the presence of foodborne pathogens in animals prior to grazing and exclusion of individuals determined positive offer a strategy to mitigate potential contamination risk in ICLS, as the presence of foodborne pathogens in soil was higher when high levels were detected in grazing animal feces. The application of the NOP 90-and 120-day interval rules between manure application by grazing and harvest should consider other management and environmental factors such as the cumulative effect of continuous ICLS in the same field, the foodborne pathogen shedding status of grazing animals, and meteorological characteristics during the growing season. Our analysis of meteorological factors in the grazed soil showed that regional differences in gEc concentrations were most likely affected by maximum air and soil temperatures on the sampling day, although the effects of meteorological factors and soil chemical characteristics were not as influential as the effects of treatment or sampling days. As regional differences on pathogen survival may be attributed to those factors, further studies will be required to consider a wider range of environmental factors across multiple regions in the future.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Ethics statement
The research protocol was approved by the Institutional Animal Care and Use Committees (IACUC) of the University of California, Davis (IACUC protocol #22700) and the University of Minnesota (IACUC protocol 157 #2008-38348A).
Author contributions
SC: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Validation, Visualization, Writing – original draft, Writing – review & editing. CC-K: Data curation, Methodology, Writing – review & editing. SW: Data curation, Methodology, Writing – review & editing. AG: Conceptualization, Funding acquisition, Methodology, Project administration, Resources, Writing – review & editing. PA: Data curation, Methodology, Writing – review & editing. MJ-R: Conceptualization, Funding acquisition, Methodology, Resources, Writing – review & editing. EE: Data curation, Writing – review & editing. LK: Data curation, Methodology, Writing – review & editing. PP: Conceptualization, Funding acquisition, Resources, Writing – review & editing. PM: Conceptualization, Funding acquisition, Resources, Writing – review & editing. AK: Writing – review & editing. FH: Conceptualization, Funding acquisition, Resources, Writing – review & editing. AS: Conceptualization, Funding acquisition, Methodology, Writing – review & editing. AP: Conceptualization, Funding acquisition, Methodology, Project administration, Resources, Supervision, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. Project and publication was made possible by the U.S. Department of Agriculture’s (USDA) Agricultural Marketing Service through grant AM200100XXXXG032 and the USDA-NIFA hatch project# CA-D-PLS-2332-H. Its contents are solely the responsibility of the authors and do not necessarily represent the official views of the USDA.
Acknowledgments
We acknowledge Thomas Luder, Kimberly Aguirre, Jannie Xu, Everardo Sanchez, and Elizabeth Isenhower for assistance in the laboratory and field work, and Jose Pablo Gomez-Vazquez and Shuyi Feng for advice on statistical analysis. We would like to thank Gaudin’s lab personnel for their assistance with field work. Figure 1 was created with Biorender.com.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fsufs.2024.1464018/full#supplementary-material
References
Alegbeleye, O. O., Singleton, I., and Sant’Ana, A. S. (2018). Sources and contamination routes of microbial pathogens to fresh produce during field cultivation: A review. Food Microbiol. 73, 177–208. doi: 10.1016/j.fm.2018.01.003
Allende, A., Castro-Ibáñez, I., Lindqvist, R., Gil, M. I., Uyttendaele, M., and Jacxsens, L. (2017). Quantitative contamination assessment of Escherichia coli in baby spinach primary production in Spain: effects of weather conditions and agricultural practices. Int. J. Food Microbiol. 257, 238–246. doi: 10.1016/j.ijfoodmicro.2017.06.027
Bach, S. J., McALLISTER, T. A., Mears, G. J., and Schwartzkopf-Genswein, K. S. (2004). Long-haul transport and lack of preconditioning increases fecal shedding of Escherichia coli and Escherichia coli O157:H7 by calves†. J. Food Prot. 67, 672–678. doi: 10.4315/0362-028X-67.4.672
Benjamin, L., Atwill, E. R., Jay-Russell, M., Cooley, M., Carychao, D., Gorski, L., et al. (2013). Occurrence of generic Escherichia coli, E. coli O157 and Salmonella spp. in water and sediment from leafy green produce farms and streams on the Central California coast. Int. J. Food Microbiol. 165, 65–76. doi: 10.1016/j.ijfoodmicro.2013.04.003
Brennan, F. P., Moynihan, E., Griffiths, B. S., Hillier, S., Owen, J., Pendlowski, H., et al. (2014). Clay mineral type effect on bacterial enteropathogen survival in soil. Sci. Total Environ. 468-469, 302–305. doi: 10.1016/j.scitotenv.2013.08.037
Brooks, M., Kristensen, K., Benthem, K. J., Magnusson, A., Berg, C. W., Nielsen, A., et al. (2017). glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 9:378. doi: 10.32614/RJ-2017-066
Chandler-Khayd, C., Di Francesco, J., Baron, J. N., Ramos, T. D. M., Aminabadi, P., Jay-Russell, M. T., et al. (2023). Risk factors associated with the prevalence of Listeria monocytogenes in manured soils on certified organic farms in four regions of the United States. Front. Sustain. Food Syst. 7:1222192. doi: 10.3389/fsufs.2023.1222192
Cheong, S., Jay-Russell, M. T., Chandler-Khayd, C., Di Francesco, J., Haghani, V., Aminanadi, P., et al. (2024). Presence of foodborne pathogens and survival of generic Escherichia coli in an organic integrated crop-livestock system. Front. Sustain. Food Syst. 8:1343101. doi: 10.3389/fsufs.2024.1343101
Cook, C., Diekman, C. M., Weller, D. L., Murphy, C. M., Hamilton, A. M., Ponder, M., et al. (2023). Factors associated with foodborne pathogens and indicator organisms in agricultural soils. Front. Sustain. Food Syst. 7:1269117. doi: 10.3389/fsufs.2023.1269117
FDA (2015). FSMA final rule on produce safety - standards for the growing, harvesting, packing, and holding of produce for human consumption. Available at: https://www.federalregister.gov/documents/2015/11/27/2015-28159/standards-for-the-growing-harvesting-packing-and-holding-of-produce-for-human-consumption (Accessed, August 18, 2023).
Franz, E., Schijven, J., de Roda Husman, A. M., and Blaak, H. (2014). Meta-regression analysis of commensal and pathogenic Escherichia coli survival in soil and water. Environ. Sci. Technol. 48, 6763–6771. doi: 10.1021/es501677c
Franz, E., Semenov, A. V., and van Bruggen, A. H. C. (2008). Modelling the contamination of lettuce with Escherichia coli O157:H7 from manure-amended soil and the effect of intervention strategies. J. Appl. Microbiol. 105, 1569–1584. doi: 10.1111/j.1365-2672.2008.03915.x
Franzluebbers, A. J., Sulc, R. M., and Russelle, M. P. (2011). “Opportunities and challenges for integrating north-American crop and livestock systems” in Grassland productivity and ecosystem services. eds. G. Lemaire, J. Hodgson, and A. Chabbi (UK: CABI), 208–218.
Goodwyn, B., Millner, P., Punchihewage-Don, A. J., Schwarz, M., Hashem, F., Bowers, J., et al. (2023). Survival and persistence of foodborne pathogenic and indicator bacteria in spatially separated integrated crop-livestock farms. Front. Sustain. Food Syst. 7:1287371. doi: 10.3389/fsufs.2023.1287371
Hilimire, K. (2011). Integrated crop/livestock agriculture in the United States: A review. J. Sustain. Agric. 35, 376–393. doi: 10.1080/10440046.2011.562042
Hutchison, M. L., Walters, L. D., Avery, S. M., Munro, F., and Moore, A. (2005). Analyses of livestock production, waste storage, and pathogen levels and Prevalences in farm manures. Appl. Environ. Microbiol. 71, 1231–1236. doi: 10.1128/AEM.71.3.1231-1236.2005
Ingham, S. C., Losinski, J. A., Andrews, M. P., Breuer, J. E., Breuer, J. R., Wood, T. M., et al. (2004). Escherichia coli contamination of vegetables grown in soils fertilized with noncomposted bovine manure: garden-scale studies. Appl. Environ. Microbiol. 70, 6420–6427. doi: 10.1128/AEM.70.11.6420-6427.2004
Ivanek, R., Gröhn, Y. T., Jui-Jung Ho, A., and Wiedmann, M. (2007). Markov chain approach to analyze the dynamics of pathogen fecal shedding—example of Listeria monocytogenes shedding in a herd of dairy cattle. J. Theor. Biol. 245, 44–58. doi: 10.1016/j.jtbi.2006.09.031
Ivanek, R., Gröhn, Y. T., Wells, M. T., Lembo, A. J., Sauders, B. D., and Wiedmann, M. (2009). Modeling of spatially referenced environmental and meteorological factors influencing the probability of Listeria species isolation from natural environments. Appl. Environ. Microbiol. 75, 5893–5909. doi: 10.1128/AEM.02757-08
Kudva, I. T., Blanch, K., and Hovde, C. J. (1998). Analysis of Escherichia coli O157:H7 survival in ovine or bovine manure and manure slurry. Appl. Environ. Microbiol. 64, 3166–3174. doi: 10.1128/AEM.64.9.3166-3174.1998
LaCanne, C. E., and Lundgren, J. G. (2018). Regenerative agriculture: merging farming and natural resource conservation profitably. PeerJ 6:e4428. doi: 10.7717/peerj.4428
Lang, N. L., Bellett-Travers, M. D., and Smith, S. R. (2007). Field investigations on the survival of Escherichia coli and presence of other enteric micro-organisms in biosolids-amended agricultural soil: E. coli survival in sludge-amended soil. J. Appl. Microbiol. 103, 1868–1882. doi: 10.1111/j.1365-2672.2007.03489.x
Lema, M., Williams, L., Walker, L., and Rao, D. R. (2002). Effect of dietary ®ber on E. coli O157:H7 shedding in lambs. Small Rumin. Res. 43, 249–255. doi: 10.1016/S0921-4488(02)00010-X
Lemaire, G., Franzluebbers, A., de Carvalho, P. C. F., and Dedieu, B. (2014). Integrated crop–livestock systems: strategies to achieve synergy between agricultural production and environmental quality. Agric. Ecosyst. Environ. 190, 4–8. doi: 10.1016/j.agee.2013.08.009
Litt, P. K., Kelly, A., Omar, A., Johnson, G., Vinyard, B. T., Kniel, K. E., et al. (2021). Temporal and agricultural factors influence Escherichia coli survival in soil and transfer to cucumbers. Appl. Environ. Microbiol. 87, e02418–e02420. doi: 10.1128/AEM.02418-20
Naganandhini, S., Kennedy, Z. J., Uyttendaele, M., and Balachandar, D. (2015). Persistence of pathogenic and non-pathogenic Escherichia coli strains in various tropical agricultural soils of India. PLoS One 10:e0130038. doi: 10.1371/journal.pone.0130038
Nazareth, J., Shaw, A., Delate, K., and Turnbull, R. (2021). Food safety considerations in integrated organic crop–livestock systems: prevalence of Salmonella spp. and E. coli O157:H7 in organically raised cattle and organic feed. Renew. Agric. Food Syst. 36, 8–16. doi: 10.1017/S1742170519000292
Newton, P., Civita, N., Frankel-Goldwater, L., Bartel, K., and Johns, C. (2020). What is regenerative agriculture? A review of scholar and practitioner definitions based on processes and outcomes. Front. Sustain. Food Syst. 4:577723. doi: 10.3389/fsufs.2020.577723
Nightingale, K. K., Fortes, E. D., Ho, A. J., Schukken, Y. H., Grohn, Y. T., and Wiedmann, M. (2005). Evaluation of farm management practices as risk factors for clinical listeriosis and fecal shedding of Listeria monocytogenes in ruminants. Proc. Ann, Meet. Am. Vet. Med. Assoc. 227, 1808–1814. doi: 10.2460/javma.2005.227.1808
Ongeng, D., Muyanja, C., Geeraerd, A. H., Springael, D., and Ryckeboer, J. (2011). Survival of Escherichia coli O157:H7 and Salmonella enterica serovar typhimurium in manure and manure-amended soil under tropical climatic conditions in sub-Saharan Africa: fate of enteric bacteria in the tropics. J. Appl. Microbiol. 110, 1007–1022. doi: 10.1111/j.1365-2672.2011.04956.x
Pang, H., McEgan, R., Mishra, A., Micallef, S. A., and Pradhan, A. K. (2017). Identifying and modeling meteorological risk factors associated with pre-harvest contamination of Listeria species in a mixed produce and dairy farm. Food Res. Int. 102, 355–363. doi: 10.1016/j.foodres.2017.09.029
Pang, H., Mokhtari, A., Chen, Y., Oryang, D., Ingram, D. T., Sharma, M., et al. (2020). A predictive model for survival of Escherichia coli O157:H7 and Generic E. coli in soil amended with untreated animal manure. Risk Anal. 40, 1367–1382. doi: 10.1111/risa.13491
Park, S., Navratil, S., Gregory, A., Bauer, A., Srinath, I., Szonyi, B., et al. (2015). Multifactorial effects of ambient temperature, precipitation, farm management, and environmental factors determine the level of generic Escherichia coli contamination on Preharvested spinach. Appl. Environ. Microbiol. 81, 2635–2650. doi: 10.1128/AEM.03793-14
Park, Y., Pachepsky, Y., Shelton, D., Jeong, J., and Whelan, G. (2016). Survival of manure-borne Escherichia coli and fecal coliforms in soil: temperature dependence as affected by site-specific factors. J. Environ. Qual. 45, 949–957. doi: 10.2134/jeq2015.08.0427
Park, S., Szonyi, B., Gautam, R., Nightingale, K., Anciso, J., and Ivanek, R. (2012). Risk factors for microbial contamination in fruits and vegetables at the Preharvest level: A systematic review. J. Food Prot. 75, 2055–2081. doi: 10.4315/0362-028X.JFP-12-160
Patterson, L., Navarro-Gonzalez, N., Jay-Russell, M. T., Aminabadi, P., Antaki-Zukoski, E., and Pires, A. F. A. (2018). Persistence of Escherichia coli in the soil of an organic mixed crop-livestock farm that integrates sheep grazing within vegetable fields. Zoonoses Public Health 65, 887–896. doi: 10.1111/zph.12503
Pires, A. F. A., Patricia, D. M., Jerome, B., and Michele, T. J.-R. (2018). Assessment of current practices of organic farmers regarding biological soil amendments of animal origin in a multi-regional U.S. Study.
Pires, A. F. A., Ramos, T. D. M., Baron, J. N., Millner, P. D., Pagliari, P. H., Hutchinson, M., et al. (2023). Risk factors associated with the prevalence of Shiga-toxin-producing Escherichia coli in manured soils on certified organic farms in four regions of the USA. Front. Sustain. Food Syst. 7:1125996. doi: 10.3389/fsufs.2023.1125996
Ramos, T. D. M., Jay-Russell, M. T., Millner, P. D., Baron, J. N., Stover, J., Pagliari, P., et al. (2021). Survival and persistence of foodborne pathogens in manure-amended soils and prevalence on fresh produce in certified organic farms: A multi-regional baseline analysis. Front. Sustain. Food Syst. 5:674767. doi: 10.3389/fsufs.2021.674767
Rhodes, C. J. (2017). The imperative for regenerative agriculture. Sci. Prog. 100, 80–129. doi: 10.3184/003685017X14876775256165
Schipanski, M. E., Barbercheck, M., Douglas, M. R., Finney, D. M., Haider, K., Kaye, J. P., et al. (2014). A framework for evaluating ecosystem services provided by cover crops in agroecosystems. Agric. Syst. 125, 12–22. doi: 10.1016/j.agsy.2013.11.004
Schipanski, M. E., MacDonald, G. K., Rosenzweig, S., Chappell, M. J., Bennett, E. M., Kerr, R. B., et al. (2016). Realizing resilient food systems. Bioscience 66, 600–610. doi: 10.1093/biosci/biw052
Schoder, D., Melzner, D., Schmalwieser, A., Zangana, A., Winter, P., and Wagner, M. (2011). Important vectors for Listeria monocytogenes transmission at farm dairies manufacturing fresh sheep and goat cheese from raw Milk. J. Food Prot. 74, 919–924. doi: 10.4315/0362-028X.JFP-10-534
Sharma, M., Millner, P. D., Hashem, F., Vinyard, B. T., East, C. L., Handy, E. T., et al. (2019). Survival of Escherichia coli in manure-amended soils is affected by spatiotemporal, agricultural, and weather factors in the mid-Atlantic United States. Appl. Environ. Microbiol. 85, e02392–e02318. doi: 10.1128/AEM.02392-18
Sharma, M., and Reynnells, R. (2016). Importance of soil amendments: survival of bacterial pathogens in manure and compost used as organic fertilizers. Microbiol. Spectr. 4:4.4.36. doi: 10.1128/microbiolspec.PFS-0010-2015
Strawn, L. K., Gröhn, Y. T., Warchocki, S., Worobo, R. W., Bihn, E. A., and Wiedmann, M. (2013). Risk factors associated with Salmonella and Listeria monocytogenes contamination of produce fields. Appl. Environ. Microbiol. 79, 7618–7627. doi: 10.1128/AEM.02831-13
Tran, D. T. Q., Bradbury, M. I., Ogtrop, F. F. V., Bozkurt, H., Jones, B. J., and McCONCHIE, R. (2020). Environmental drivers for persistence of Escherichia coli and Salmonella in manure-amended soils: A Meta-analysis. J. Food Prot. 83, 1268–1277. doi: 10.4315/0362-028X.JFP-19-460
USDA-AMS NOP (2011). Guidance processed animal manures in organic crop production. Available at: https://www.ams.usda.gov/sites/default/files/media/5006.pdf
USDA-Natural Resources Conservation Service (2013). NRCS Pasture Stick instructions for using. Available at: https://nwdistrict.ifas.ufl.edu/phag/files/2019/11/NRCS-PASTURE-STICK-instructions.pdf
Weller, D., Belias, A., Green, H., Roof, S., and Wiedmann, M. (2020). Landscape, water quality, and weather factors associated with an increased likelihood of foodborne pathogen contamination of New York streams used to source water for produce production. Front. Sustain. Food Syst. 3:124. doi: 10.3389/fsufs.2019.00124
Keywords: grazing, food safety, sheep, goats, STEC, cover crops, fresh produce
Citation: Cheong S, Chandler-Khayd C, Williams SR, Gaudin ACM, Aminabadi P, Jay-Russell MT, Evans E, Klossner L, Pagliari P, Millner P, Kenney A, Hashem F, Sciligo AR and Pires AFA (2024) Evaluation of environmental risk factors associated with survival of generic E. coli in organic integrated crop-livestock systems in California and Minnesota. Front. Sustain. Food Syst. 8:1464018. doi: 10.3389/fsufs.2024.1464018
Edited by:
M. Leonor Faleiro, University of Algarve, PortugalReviewed by:
Eva Cunha, University of Lisbon, PortugalMassimo Pajoro, IRCAF-Catholic University, Campus Santa Monica Cremona, Italy
Copyright © 2024 Cheong, Chandler-Khayd, Williams, Gaudin, Aminabadi, Jay-Russell, Evans, Klossner, Pagliari, Millner, Kenney, Hashem, Sciligo and Pires. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alda F. A. Pires, YXBpcmVzQHVjZGF2aXMuZWR1