 Ni Luh Suriani1*
Ni Luh Suriani1* Dewa Ngurah Suprapta2
Dewa Ngurah Suprapta2 I. Nyoman Suarsana3
I. Nyoman Suarsana3 Ni Made Delly Resiani4
Ni Made Delly Resiani4 Kahkashan Perveen5
Kahkashan Perveen5 Najat A. Bukhari5
Najat A. Bukhari5 Hesham Ali Ei Enshasy6Ting Seng Ho7
Hesham Ali Ei Enshasy6Ting Seng Ho7 R. Z. Sayyed8
R. Z. Sayyed8- 1Biology Study Program, Mathematics, and Natural Sciences, Udayana University, Denpasar, Bali, Indonesia
- 2Biopesticide Laboratory, Agriculture Faculty, Udayana University, Denpasar, Bali, Indonesia
- 3Faculty of Veterinary Medicine, Udayana University, Denpasar, Bali, Indonesia
- 4National Research and Innovation Agency, Central Jakarta, Indonesia
- 5Department of Botany and Microbiology, College of Science, King Saud University, Riyadh, Saudi Arabia
- 6Innovation Center in Agritechnology for Advanced Bioprocessing (ICA), Universiti Teknologi Malaysia (UTM), Pagoh, Johor, Malaysia
- 7Back2Nature Organic Farm, Kuala Pilah, Malaysia
- 8Department of Microbiology, PSGVP Mandal's S. I. Patil Arts, G. B. Patel Science and STKV Sangh Commerce College, Shahada, India
Introduction: Developing organic herbal crops provides chemical-free herbs to support environmentally friendly and sustainable farming. One way in agricultural biotechnology to grow herbal organic crops is to use rhizobacteria. The herbal plant used in this study is the Javanese ginseng plant (Talinum paniculatum), which contains phytochemicals that increase stamina.
Methods: The study used four rhizobacteria to enhance the growth and phytochemistry of T. paniculatum leaves; the target phytochemical compounds analyzed in Javanese ginseng leaves were total flavonoids, total phenols, and antioxidants (IC50%). The four selected rhizobacteria can produce indole-3-acetic acid (IAA), fix nitrogen, and dissolve phosphate. Thus, high-quality T. paniculatum leaves were obtained as herbal tea ingredients. The pattern used is a random group pattern in the greenhouse.
Results: The results of the study showed that the use of rhizobacteria at 2% concentrations had a real effect on plant growth parameters such as plant height, leaf area, root length, wet weight and dry weight of the plant, and phytochemical content like total flavonoids, total phenols, IC50%, total chlorophyll, content of T. paniculatum leave plant when compared to the control. The four rhizobacteria used can produce the IAA, fix nitrogen, and dissolve phosphorus.
Discussion: The four rhizobacteria can also colonize the root of the T. paniculatum plant. The four Rhizobzcteria used significantly affected the growth and phytochemical content of T. paniculatum leaves as an ingredient in herbal tea at a concentration of 2% compared to controls. The four rhizobacteria can produce IAA hormones, fix nitrogen, and dissolve phosphorus. All four rhizobacteria can colonize the roots of T. paniculatum plants. These four rhizobacteria can be used as alternative methods in developing organic farming systems and can also be used practically in the field by farmers. It is necessary to research the application of rhizobacteria to other crops to support sustainable agriculture.
Introduction
Ginseng plants are trendy for herbal plants (Ahmad et al., 2023), especially Korean or Chinese ginseng plants contain ginsenosides as the major bioactive components known to have complex and multiple pharmacological effects (Wang et al., 2009; Xiang et al., 2023), like as an aphrodisiac because it contains saponins, triterpenes or steroids, polyphenols, and essential oils, Javanese ginseng shows its efficacy as a stimulant. It increases the threshold of fatigue and is safe based on acute toxicity tests. Javanese ginseng extract at doses of 5 and 10 mg/40 g BW can improve fitness and extend the sleep time of mice (Ekawati, 2015). However, Korean or Chinese ginseng is expensive and sometimes not affordable for the community. In addition to Korean ginseng, Javanese ginseng (Talinum paniculatum) leaves and roots contain phytochemicals. They can function as an aphrodisiac as well. Hence, the Javanese ginseng (T. paniculatum) plant is very suitable for development as an herbal plant, where this plant is cheaper and easier to grow (Rizki et al., 2023). Many herbal plants developed so far are still non-organic, so their function is less than optimal. Developing herbal plants, such as ginseng plants, is essential as a health drink, considering their function as an aphrodisiac. Rizki et al. (2023) found that Javanese ginseng leaf extract can improve sperm quality and quantity in mice. Javanese ginseng leaf extract also contains 43.78% antioxidants, which can capture free radicals in the body (Lestario et al., 2009). Javanese ginseng may also reduce the effects of contractions (Sukwan et al., 2014). Developing organic herbal plants is desirable because they produce chemical-free herbal plants that are safer and healthier for consumption (Marcelino et al., 2023). Consuming organic herbs is very beneficial for health when compared to inorganic herbs, such as functioning as anti-imflamacy; the main health benefits mentioned included anti-inflammatory properties (45.0%), prevention of cardiovascular diseases (41.6%), and prevention of high cholesterol (39.9%), safe for the body because it does not contain chemical substances that are chemical pesticide residues (Mendes et al., 2023).
One of the biotechnologies that can be developed to support organic herbal plants is developing rhizobacteria methods, which use root bacteria that can function as biostimulants, biopesticides, and biopesticides (de Andrade et al., 2023). Rhizobacteria can also increase growth, phytochemicals, and antioxidants in Piper caninum plants because rhizobacteria can produce IAA hormones, fix nitrogen, and dissolve phosphate, which can increase growth and production in plants (Suriani et al., 2021). Rhizobacteria can also increase growth and biological activity in Lessertia frutescens plants (Hlongwane et al., 2023); likewise, plant growth Plant Growth – Promoting Rhizobacteria (PGPR) can increase lycopene in tomato plants (de la Osa et al., 2021). (PGPR) can also increase biomass and secondary metabolites in Cannabis sativa plants (Lyu et al., 2023). Inoculation of rhizobacteria in Mentha x piperita plants can increase phenolics (Del Rosario Cappellari et al., 2020). The interaction of PGPR with the rhizosphere of soil is a positive interaction with plant roots, where microorganisms in the roots can fix nitrogen, dissolve phosphates, produce siderophores, produce phytohormones, produce enzymes antagonistic to plant pathogens and plant stress (Chandran et al., 2021). The auxin hormone produced by Synecchoccus sp. bacteria can influence plant growth, so nutrient uptake in the soil (especially N) will also increase. More N uptake will encourage enzymes that form flavon compounds to run more optimally, which can increase the phenolic and flavonoid content in soybean leaves (Kurniawan et al., 2014); the addition of organic fertilizer can increase the phenolic, flavonoid content and antioxidant activity in basil plants (Taie et al., 2010). Mahdavikia et al. (2019) stated that organic fertilizer and rhizobacteria can increase the catalase enzyme so that the antioxidant content in basil plants increases, and the use of Bacillus lentus rhizobacteria can increase minerals and proline in basil plants under stress conditions. Organic fertilizer and rhizobacteria can increase antioxidant activity in wheat plants subjected to water stress (Khalilzadeh et al., 2016). PGPR produce growth hormones, fix nitrogen, dissolve phosphate, and produce valuable enzymes. All these features help achieve a sustainable agriculture system. PGPR inoculation is also one of the methods in agricultural development to overcome crop stress and sustainably increase crop productivity (Ojuederie et al., 2019).
The treatment of 4 rhizobacteria in this study (Brevibacillus agri, Bacillus velezensis, Paenibacillus polymyxa, and Pseudomonas monteilii), where rhizobacteria taken from plant roots have been selected from 40 isolates, shows that these 4 rhizobacteria can produce IAA hormones, can fix nitrogen and can also dissolve phosphate. The use of rhizobacteria in this study is thought to increase growth and the content of phytochemicals and antioxidants in Javanese ginseng (T. paniculatum) plants, which will later be used as organic herbal tea ingredients.
Materials and methods
Time and location of research
Between January 2023 and October 2024, the current study was conducted at the greenhouse at Munduk Paku Village in Senganan Penebel, Tabanan, Bali, Indonesia (88°22′49.3′ 1,115°09′43.2′) at an altitude of 600 meters from the sea, sandy loam type (Figure 1) and Udayana University in Bali, Indonesia, Biopesticide laboratory, This area has a Type A climate, according to Schmidt and Ferguson, with an average of 155.6 rainy days and 2,000 to 2,800 mm of annual rainfall. The region has four to ten wet and five dry months yearly. Furthermore, according to Suriani et al. (2021), the average air temperature is between 25°C and 28°C.
Figure 1. Map of research location (red color).
Research design
A randomized group design was used in the greenhouse, consisting of five treatments and five duplicates. This resulted in a total of 25 experimental units, each containing three clumps, amounting to a total of 75 clusters. The control, or untreated soil, is represented by F0. The treatments are represented by F1, F2, F3, and F4, which correspond to 2% Brevibacillus agri, 2% Bacillus velezensis, 2% Paenibacillus polymyxa, and 2% Pseudomonas monteilii, respectively. Each polybag contains a single ginseng jawa plant ready to be planted (Suriani et al., 2021). These four bacteria were chosen as they produce IAA hormones, fix nitrogen, and dissolve phosphate.
Test for indole acetic acid-producing ability
The specimens were cultured for 48 h at a temperature of 28°C, in the absence of light, within a 5 mL test tube containing tryptic soy broth. The color of the isolates changed after adding one cc of Salkowski’s solution to a 5 mL test tube containing one cc of tryptic soy broth. The pink liquid indicates that the rhizobacteria isolate can synthesize indole-3-acetic acid (IAA). The data was quantified using spectrophotometry at a wavelength of 520 nm or more (Delgado-Ramírez et al., 2021).
Test nitrogen fixation
The bacterial isolates were cultivated in bromothymol blue malate medium at 28°C for 48 h. Once the colonies were yellow, the rhizobacteria demonstrated active nitrogen fixation (Tang et al., 2018).
Test for phosphate solubilization
On Pikovoskaya media, rhizobacteria are produced; a clear zone suggests that the bacteria can dissolve phosphate. 31.3 g of Pikovoskaya’s medium was combined with 1,000 mL of aquadest, and the mixture was then put into test tubes holding 10 mL each. The test tubes were autoclaved for 15 min at 1210°C, cooled, and inoculated with rhizobacteria before being incubated for 48 h at 280°C (Kuan et al., 2016).
Mass production of rhizobacteria
The following bacteria isolates are grown in Nutrient Agar (NA) media: Rhizobacteria Brevibacillus agri F1 (Accession No. OM510267), Bacillus velezensis F2 (Accession No. OR244032), Paenibacillus polymyxa F3 (Accession No. OR244033), and Pseudomonas monteilii F4 (Accession No. OR225822). To produce one liter of bacteria, one liter of potato dextrose broth (PDB) medium is made, and 500 mg of Nystatin is used per liter of media. After that, five B. agri Ose culture needles are incubated for 3 days at 28°C (Ezrari et al., 2021).
N, P, K soil and leave analysis
A 0.5 g of specimens were measured after being flattened and inserted into a Kjeldahl flask. Subsequently, 25 mL of sulfuric-salicylic acid solution was introduced, agitated, and undisturbed for the night. Later, the concoction was subjected to gentle heating until the effervescence subsided by adding 4 g of Na2S2O2.5H2O. The temperature was incrementally raised until it reached a peak of 300°C (about 2 h) and then allowed to cool. The solution was put into a 500 mL measuring flask, diluted with distilled water, agitated, and adjusted to the appropriate level. The distillation process was halted after the distillation output reached a volume of 100 mL. Afterward, 25 mL of liquid was transferred using a pipette and combined with 150 mL of distilled water in a distillation flask.
Additionally, 10 mL of a 40% sodium hydroxide solution and 20 mL of a 1% boric acid solution were added. Finally, three drops were included. The solution underwent titration using a 0.05 N H2SO4 solution until the titration achieved its endpoint, indicated by a color change from green to pink. Simultaneously, efforts were made to address the unresolved issue in the solution. In addition, nitrogen levels were quantified using a UV–Vis spectrophotometer set at a wavelength of 400 nm (Liu et al., 2022).
P analysis
0.5 g of soil undergoes the ashing process by adding concentrated H2SO4 and concentrated HNO3, followed by heating it on a hot plate. Subsequently, 2.5 mL of concentrated H2SO4 was introduced, causing the substance to change color like ash. Following this, concentrated HNO3 was added until the emission of smoke from the sample ceased and the color turned black. The HNO3 was added incrementally until the sample ended to produce black smoke upon the addition of HNO3. Once the ashing process is finished, a sample is combined with 50 cc of distilled water and vigorously mixed. Subsequently, the mixture was filtered, and 54 units were transferred into the Erlenmeyer flask, followed by another transfer into the same flask. Dispense 2.5 mL of vanadate molybdate, resulting in the formation of a yellow hue. In addition, phosphorus levels were measured using a UV–Vis spectrophotometer set at a wavelength of 400 nm (Javaid et al., 2023).
A total of 2.5 g of test-ready samples were measured using a 250 mL flask. To perform the K analysis, 50 mL of a 4% solution of (NH4)2C2O4 and 125 mL of distilled water were utilized. The concoction was heated until it reached its boiling point, maintained at 30°C for 30 min, and subsequently allowed to cool. After the combination reached the desired level on the flask, it was moved to a 250 mL measuring flask and thinned out with distilled water. The 15 mL solution was filtered or allowed to settle until it became clear. It was then put into a 100 mL measuring flask to make the analytical solution. In addition, 2 mL of sodium hydroxide solution (20%), 5 mL of formaldehyde, and 1 mL of sodium tetraphenylborate were added for every 1% potassium oxide. Following the addition of distilled water to the designated level in the flask and stirring for a duration of 5 to 10 min, the solution underwent filtration using Whatman filter paper No. 12. Approximately 50 mL of the resulting filtrate was extracted for subsequent analysis (Liang et al., 2022).
Scanning electron microscopy test of rhizobacteria on the roots of Javanese ginseng (Talinum paniculatum)
The study employed scanning electron microscopy (SEM) to examine the impact of rhizobacteria treatment on bacterial colonization of plant roots. Control samples were generated by utilizing the roots of ginseng jawa, while treatment samples were produced by submerging the roots in a 2% solution of rhizobacteria for 3 days. The samples were subjected to an 8 h dehydration process, followed by a 1 week ventilation period at 50°C until a stable weight was achieved. The root samples were analyzed using a ZEISS Merlin field-emission scanning electron microscope (FE-SEM) with an energy-dispersive X-ray spectrometer (EDS) (He et al., 2023). The microscope operated at a beam current ranging from 0.2 to 30 kilovolts (kV) with a current intensity as low as a few picoamperes (pA) to 300 nanoamperes (nA) under a vacuum condition. The study employed a 3 kilovolt (kV) acceleration for imaging purposes. In contrast, a 15 kV acceleration was utilized for EDX investigations, except for the skin, where a 10 kiloelectronvolt (keV) acceleration proved adequate. The analysis was performed at the Laboratory of Universitas Gadjah Mada (UGM) (Maulina et al., 2022).
Greenhouse trials
Preparation of planting medium
The media used for planting ginseng plants is a mixture of soil and compost with a ratio of 3:1. This media was filled in a polybag (20 cm × 20 cm) at 3/4th of the capacity of the polybag.
Planting
Rhizobacteria are employed to prepare and treat the seedlings before they are used. The pest- and disease-free seedlings are uniformly 20 cm tall and healthy. According to Suriani (2019), planting is done perpendicularly at a depth of ±5 cm.
Application
Following the previously planned schedule, rhizobacteria were applied at 1, 2, 3, and 4 weeks following planting. The control was only watered, and each treatment was watered with rhizobacteria set at a concentration of 2%, as much as 200 mL for each Javanese ginseng plant.
Maintenance
Numerous crucial duties, including watering, weeding, fertilizing, and trimming, are involved in plant management. Creating plant designs with embroidery is usually done on plants that develop consistently and without deviations. These are pre-prepared plants to guarantee consistent development (Vafa et al., 2021).
Measured parameters
The field measurements encompass plant height, root length, leaf area, laboratory examination of chlorophyll content, and determining N, P, K, Cu, Cd, and Pb levels in leaves. Additionally, the phenolic content, flavonoids, and antioxidant activity were evaluated (Du et al., 2024).
Harvest
Within 2 months of the ginseng Jawa plant, harvesting takes place. Following harvesting, the leaves were cleaned, dried indoors using a clean wind, and left for 8 hours before baking.
Extract manufacturing
Before chemical analysis, the ginseng Jawa leaves are cut into 2 mm thick segments and subjected to an 8 h drying process in a sanitary and arid environment. After the leaves have dried, they are exposed to a temperature of 50°C for 10 h until they reach a consistent weight and moisture content of 4%. Subsequently, the leaves are crushed and soaked in ethanol, and the resulting mixture is then subjected to evaporation using a rotary evaporator (Lee et al., 2021). The leaves were examined to determine their phenolic, flavonoid, and antioxidant levels.
Polyphenols
Fifty milliliters of distilled water dissolved 3.5 grams of Na2CO3 to make a 7% Na2CO3 solution. Using gallic acid (GAE) as the reference standard, the colorimetric method was utilized to quantify the total phenolic compounds. After that, 10 mg of gallic acid was dissolved in 10 mL of ethanol to create a standard gallic acid solution with a 1,000 ppm concentration. 2.5 mL of the original solution and 25 mL of ethanol were combined to reach a concentration of 100 ppm. After that, 10 mL of ethanol was combined with 1, 2, 3, 4, and 5 mL of the solution to create concentrations of 10, 20, 30, and 50 parts per million (ppm). 0.4 mL of the Folin-Ciocalteau reagent was added to each concentration of 10, 20, 30, 40, and 50 ppm to test the gallic acid standard solution. After stirring the mixture for 4 to 8 min, 4.0 milliliters of a 7% Na2CO3 solution was added and stirred until a smooth consistency was reached. After that, up to 10 milliliters of distilled water were added, and the mixture remained at room temperature for 2 hours. A calibration curve was created by measuring the absorbance at 744.8 nm and comparing it to the gallic acid concentration (g/mL). The Javanese ginseng (T. paniculatum) extract solution was made by measuring and dissolving 10 milligrams of the extract in 10 mL of ethanol. A pipette with up to 1 mL of the solution was added to the mixture to measure the total phenol levels.
Furthermore, 4 to 8 min were spent stirring the solution after adding 0.4 mL of the Folin-Ciocalteau reagent and then combining it with 4.0 mL of the 7% Na2CO3 solution. After adding 10 milliliters of distilled water, the mixture was stirred and allowed to stand at room temperature for 2 hours. The wavelength at which the maximal absorption was measured was 744.8 nm. The process was repeated three times, and the amount of phenol in each gram of extract was measured in milligrams of gallic acid equivalent (Costea et al., 2022).
Flavonoids
Quercetin (QE) and steps were used in a colorimetric method to assess the total flavonoid levels. Every hour, 10 mg of standard quercetin were dissolved in 10 mL of ethanol to create quercetin solutions with a 1,000 ppm concentration. To make a solution with a concentration of 100 ppm, 10 milliliters of high-purity (p.a.) ethanol were combined with a normal quartzine solution with a concentration of 1,000 ppm. Subsequently, a series of solutions with concentrations of 10 ppm, 20 ppm, 30 ppm, 40 ppm, and 50 ppm were prepared using this solution. Each standard was supplemented with 3 mL of quercetin, 0.2 mL of 10% AlCl3, 0.2 mL of potassium acetate, and up to 10 mL of distilled water. The sample was subjected to an additional 30 min incubation period at room temperature. Subsequently, its absorbance was measured using UV–Vis spectrophotometry at a precise wavelength of 431 nm. A 100 mg cadaver extract solution was diluted in 10 mL of ethanol, and 0.2 mL of 10% AlCl3, 0.2 mL of potassium acetate, and 10 mL of distilled water were added to the solution to determine its total flavonoid concentration. After the combination was incubated for 30 min at room temperature in a light-restricted environment, the absorbance was measured using UV–Vis spectrophotometry at a wavelength of 431 nm. Concurrently, three duplicate samples of the solution were generated to determine the flavonoid contents regarding quercetin equivalents (Perisoara et al., 2022).
Antioxidant
Between 0 and 2 mg/L of gallic acid were used to create a range of concentrations. The sample was measured to be 0.05 grams, then diluted with 99.9% ethanol to fill a measuring flask to a capacity of 5 mL. After that, it was centrifuged at 3000 revolutions per minute for 15 min. Pipetting was used to add 0.5 mL of DPPH 0.1 mm (dissolved in 99.9% ethanol solvent) into the test tube after the standard and supernatants had been added. The sample was incubated for 30 minutes at a temperature of 25°C, which provided enough time for the DPPH to react with the hydrogen atoms in the antioxidants in the sample. Furthermore, an assessment of its absorbance at 517 nm was carried out. The linear regression equation, y = ax + b, was used to calculate the value of y, which indicated the antioxidant capacity (Kreatsouli et al., 2019).
Heavy metal analysis Pb, Cd, Cu
Both treated and controlled Javanese ginseng (T. paniculatum) leaves were analyzed using 0.5 g leaf samples put in a Kjeldahl flask with 5 mL of concentrated HNO3 and H2SO4. After that, the material was wet-digested until a dark, slightly yellow powdered solution was produced. Using a 100 mL measuring flask, the resultant solution was diluted with ion-free water and filtered until a filtrate was obtained. Additionally, AAS was used to examine the filtrates for mineral standards and metal grades (Aslanidis and Golia, 2022).
Data analysis
A study of variance (ANOVA) was used to study the collected data quantitatively. To determine if the therapy causes statistically significant changes in the observed variables, the analysis is expanded using the Duncans Multiple Range Test (DMRT) at a significance threshold of 5% (Hosseini et al., 2022).
Results
IAA hormone analysis, nitrogen test, and phosphate test
The analysis results (Table 1) indicated that all four rhizobacteria exhibited plant growth-promoting features, such as IAA production, nitrogen fixation, and phosphate solubilization. Moreover, F3 (P. polymyxa) produced the highest level of IAA hormone, measuring 689 ppm. B. agri is a bacterium that can synthesize the hormone IAA, perform nitrogen fixation, and enhance the development and concentration of phytochemicals and antioxidants in Piper caninum herbal plants (Suriani et al., 2021). B. velezensis, a type of rhizobacteria, acts as a biostimulant by synthesizing indole-3-acetic acid (IAA) hormones; this bacterium promotes plant growth by enhancing root development, expanding plant height, and augmenting leaf count in tomato plants (Chen et al., 2022). P. polymyxa is a plant growth-promoting rhizobacterium (PGPR) that enhances plant growth by effectively hydrolyzing phosphorus, including those found in watermelons. Additionally, it acts as a biocontrol agent against fusarium disease, as Yaoyao et al. (2017) demonstrated. The use of rhizobacteria Bacillus subtilis, Bacillus amyloliquefaciens, and Pseudomonas monteilii can enhance the ripening process in strawberries by promoting nitrogen fixation from the atmosphere, hence increasing the availability of nitrogen for strawberry plants (Nam et al., 2023).
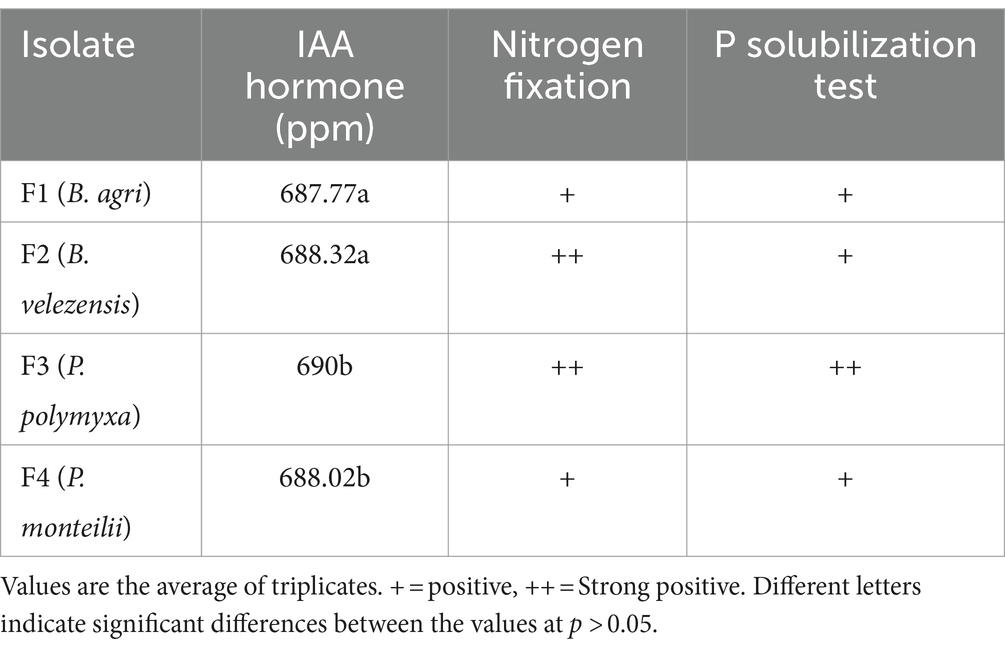
Table 1. IAA hormone content, nitrogen fixation, phosphorus solubility in rhizobacteria isolate.
The analysis results (Table 1) indicated that all four rhizobacteria exhibited positive results in the nitrogen fixation test. Additionally, they showed positive results in the phosphating test, with the positive test observed in F2, F2, F3, and F4 (Table 1). Furthermore, F3 (P. polymyxa) produced the highest level of IAA hormone, measuring 689 ppm. B. agri is a bacterium that can synthesize the hormone IAA, perform nitrogen fixation, and enhance the development and concentration of phytochemicals and antioxidants in Piper caninum herbal plants (Suriani et al., 2021). Rhizobacteria B. velezensis is a bacterium that acts as a biostimulant by producing IAA hormones, which promote growth by enhancing root development, raising plant height, and augmenting leaf count in tomato plants (Chen et al., 2022). Paenibacillus polymyxa is a PGPR that enhances plant growth by effectively solubilizing phosphates, including those found in watermelon. Additionally, it acts as a biocontrol agent against fusarium disease, as Yaoyao et al. (2017) demonstrated. The application of rhizobacteria such as Bacillus subtilis, Bacillus amyloliquefaciens, and Pseudomonas monteilii can enhance the ripening process in strawberries by promoting nitrogen fixation from the atmosphere, hence increasing the availability of nitrogen for strawberry plants (Nam et al., 2023).
Effect of treatment growth of Talinum paniculatum
The findings from Table 2 and Figure 2 indicate a notable disparity between the treatment and control groups. The treatment involving P. monteilii, specifically F4, exhibited the tallest plant height at 64.34 cm. The widest leave area was observed in the F3 treatment, measuring 14.43 cm. F2 treatment also had the longest root length and the highest wet and dry weight. The growth in the treatment group was superior to that in the control group due to the ability of F1, F2, F3, and F4 rhizobacteria to synthesize hormones, solubilize phosphorus, and perform nitrogen fixation, as indicated in Table 1. IAA hormones stimulate growth, whereas rhizobacteria’s phosphorus and nitrogen are plant nutrients, promoting growth (de Andrade et al., 2023). PGPR can enhance soil nutrition, facilitate the cycling of soil nutrients, and augment nutrient availability for plants. The growth hormone synthesized by PGPR can enhance the growth and productivity of vegetables, playing a crucial role in ensuring the accessibility of nutritious food (Kumar et al., 2021). Root exudates consist of intricate combinations of chemo effectors comprising several diverse chemicals (Xiang et al., 2023). Adding PGPR to Macadamia integrifolia plants enhances the soil’s nitrogen availability and minimizes nitrogen loss (Gallart et al., 2021). Rhizobacteria, such as Bacillus subtilis, Bacillus amyloliquefaciens, and cyanobacteria, can increase antioxidant potential, biochemical content, growth, and productivity (Plants et al., 2022). PGPR treatment in potatoes can improve, and treatment with NPK can increase by 111% (Ekin, 2019).
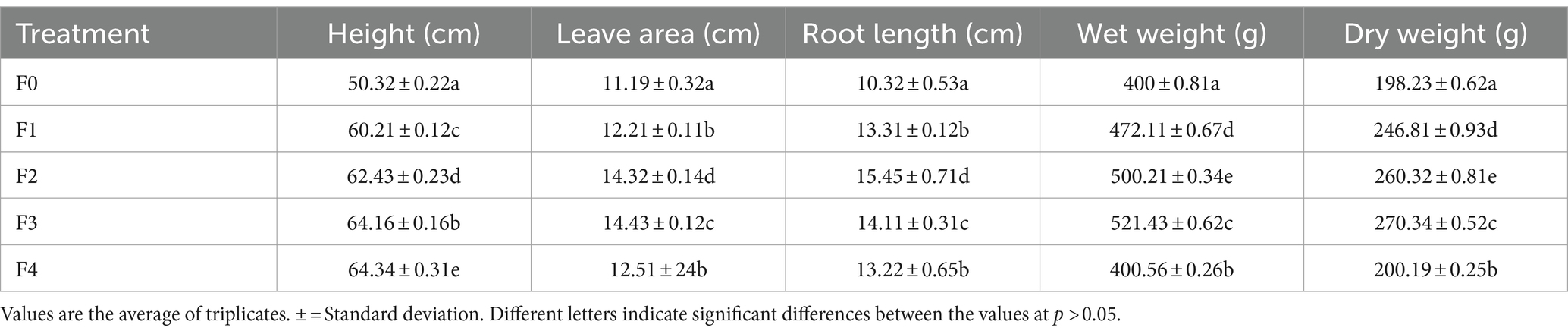
Table 2. Growth of ginseng plants after 1.5 months of planting.

Figure 2. Growth of ginseng plants from right to left: control, F1, F2, F3, and F4.
Analysis of phytochemicals, chlorophyll, antioxidants, and heavy metals of ginseng leave
Between the treatment groups and the control group, the findings of phytochemical, chlorophyll, and antioxidant tests showed significant differences (Table 3). The F2 treatment exhibited the highest concentration of total flavonoids and total phenols. Additionally, it demonstrated the smallest 50% IC value, indicating the strongest antioxidant action. The F4 treatment had the highest chlorophyll content, with the most chlorophyll a and b. According to Suriani et al. (2021), using B. agri at 1–2% can enhance the phenolic content, flavonoids, antioxidant activity, and chlorophyll content in Piper caninum herbal plants. Applying rhizobacteria derived from rice roots on toman cantos plants has enhanced the bioactive lycopene content and antioxidant activity (de la Osa et al., 2021). The combination of PGPR and salicylic acid can improve the levels of secondary metabolites, precisely phenolic compounds and monoterpenes, in Mentha x piperita plants (Del Rosario Cappellari et al., 2020; de la Osa et al., 2021). Inoculating Mucilaginibacter sp. and Pseudomonas sp. at the flowering stage resulted in a 23 and 18% augmentation in overall terpene accumulation, respectively. In general, introducing beneficial bacteria into the plants’ vegetative stage improved the yield and chemical characteristics of cannabis (Lyu et al., 2023). Jia et al. (2024) reported that the inoculation with biochar and arbuscular mycorrhizae influenced lead immobilization in mazie.
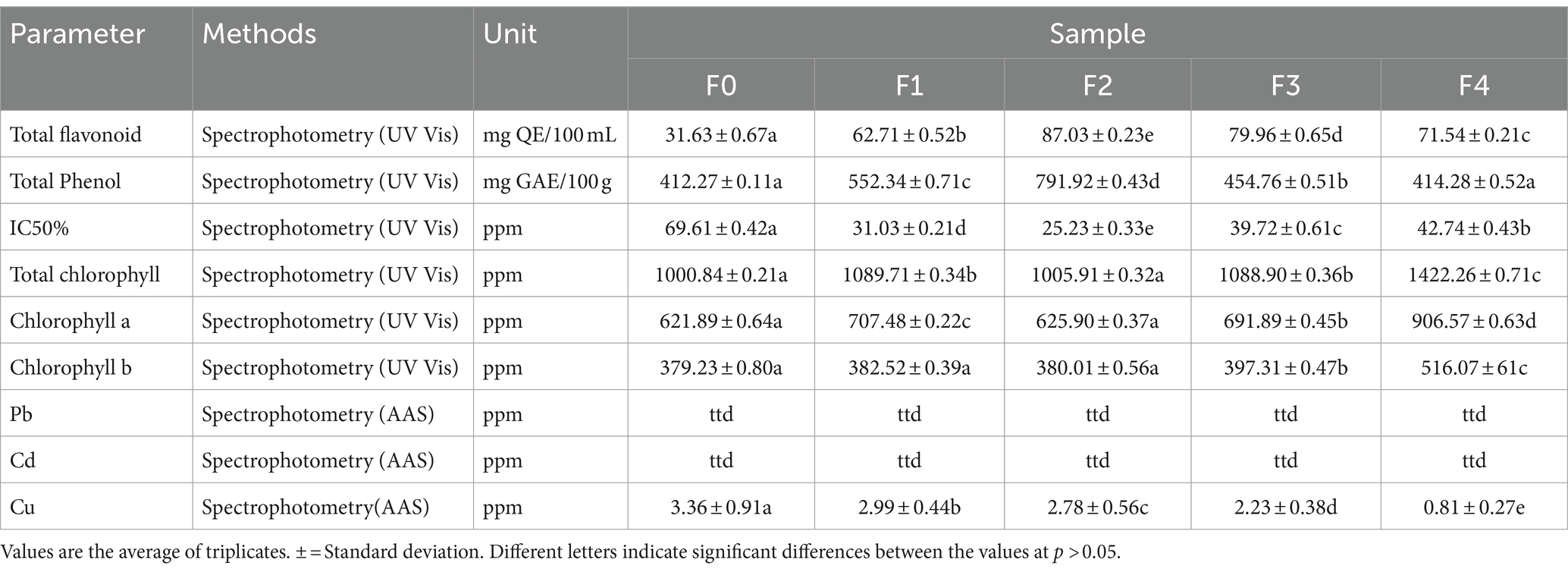
Table 3. Results of phytochemical analysis of ginseng leaves after 8 weeks of planting.
No traces of the heavy elements Pb and Cd were found. However, the controls had significant amounts of Cu, as shown in Table 3. The bacterium Pseudomonas rhizophila S211, which was found in an artichoke field contaminated with pesticides, has been shown to have the ability to promote plant development, suppress pests, and remediate contaminated soil (Lyu et al., 2023). PGPR can reduce the soil’s metal content and help the soil’s material remodeling process (Vocciante et al., 2022; Jia et al., 2024).
Analysis of N, P, K soil and leaves of Talinum paniculatum
The data in Tables 4, 5 reveal that the treatment and control groups had significantly different amounts of N, P, and K in their soil and T. paninum leaves. Rhizobacteria that promote plant growth, commonly called PGPR, are microorganisms that increase crop yield, strengthen plant immunity, and promote plant development. Based on interaction, they are divided into two groups: internal (nitrogen-fixing bacteria) and extracellular (in the soil around the cellular space, root surface, and root) (Kumar et al., 2022). Phosphate-solubilizing PGPR convert insoluble organic and inorganic phosphorus molecules into soluble forms and make them available to plants. Utilization of Plant Development It has been demonstrated that bioinoculation of Rhizobacteria (PGPR) can boost plant biomass and accelerate seed germination.
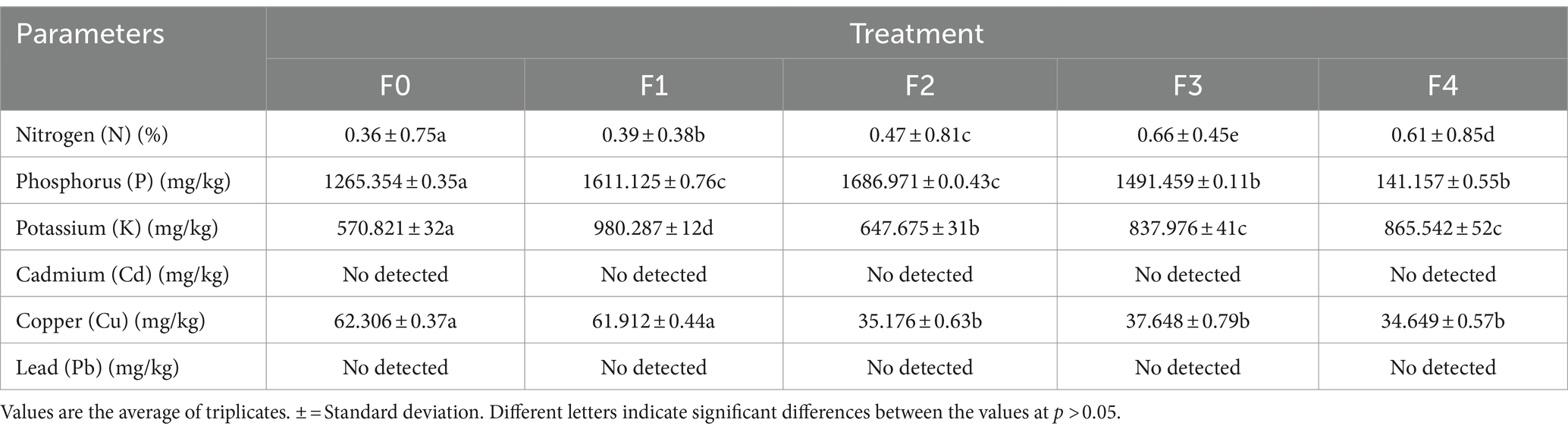
Table 4. Soil analysis.
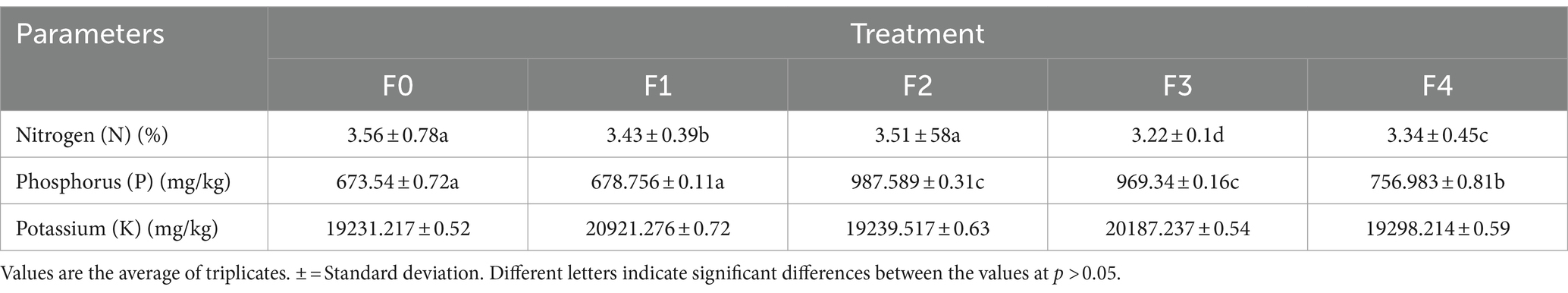
Table 5. N, P, K analysis of T. paniculatum leaves.
Furthermore, PGPR can provide essential nutrients to plant roots, including potassium (K), phosphorus (P), and nitrogen (N) (Kumar et al., 2021). The wheat plant showed an increase in its absorption of potassium and nitrogen of up to 17% when exposed to P. frederiksbergensis J158 and E. hormaechei J146. Tomatoes can have higher amounts of nitrogen and phosphorus when B. amyloliquefaciens and B. subtilis are used together (Plants et al., 2022). PGPR functions as a biofertilizer, which can improve plant development. Studies have indicated a strong correlation between the amount of chlorophyll produced by PGPR and the bacteria’s ability to enhance the host plant’s uptake of various minerals, including calcium, magnesium, phosphorus, and potassium (Khoso et al., 2024). Microbial inoculants, namely plant growth-promoting rhizobacteria (PGPR), have been shown in a recent meta-analysis to increase agricultural yield by enhancing the uptake of essential nutrients (N, P, K) and therefore elevating levels of chlorophyll, boosting agricultural output by improving the uptake of critical nutrients (nitrogen, phosphorus, and potassium) and thus raising chlorophyll levels (Du et al., 2024; Su et al., 2024). In comparison to the other rootstock, the M.9 rootstock benefited more from the bacterial treatment in terms of leaf nitrogen (N), boron (B), fruit nitrogen (N), and iron (Fe) content. Conversely, the rootstock MM.106 showed a more significant favorable impact on other nutrient content. The bacterial application benefited the cultivars in general, with the Fuji cultivar showing the most important gains in fruit manganese (Mn) content (32.1%) and leaf phosphorus (P) content (10.7%) (Yildiz et al., 2022).
Analysis scanning electron microscope colonization of rhizobacteria
In Figure 3, bacterial colonization was seen. The treatment group displayed more bacterial colonization in the ginseng plant roots than the control group, which displayed less colonization. The ginseng roots treated with B. velensensis showed the highest degree of colonization. B. agri can become established in P. caninum plant root systems (Suriani et al., 2021). The capacity of PGPR to produce biofilms shows bacterial motility, exhibit antioxidant activities, detect chemical signals and nutrients released by root exudates, and successfully evade and suppress the plant immune system, all contribute to their ability to live in the rhizosphere (Santoyo et al., 2021). Plant Growth-Promoting Rhizobacteria (PGPRs) use chemoreceptors to sense chemical attractants, which allows them to establish a mutually beneficial relationship with plants. As a result, they can form biofilms on roots and eventually colonize the rhizosphere. PGPR can colonize plant roots and can increase plant’s resistance (Wang et al., 2023).
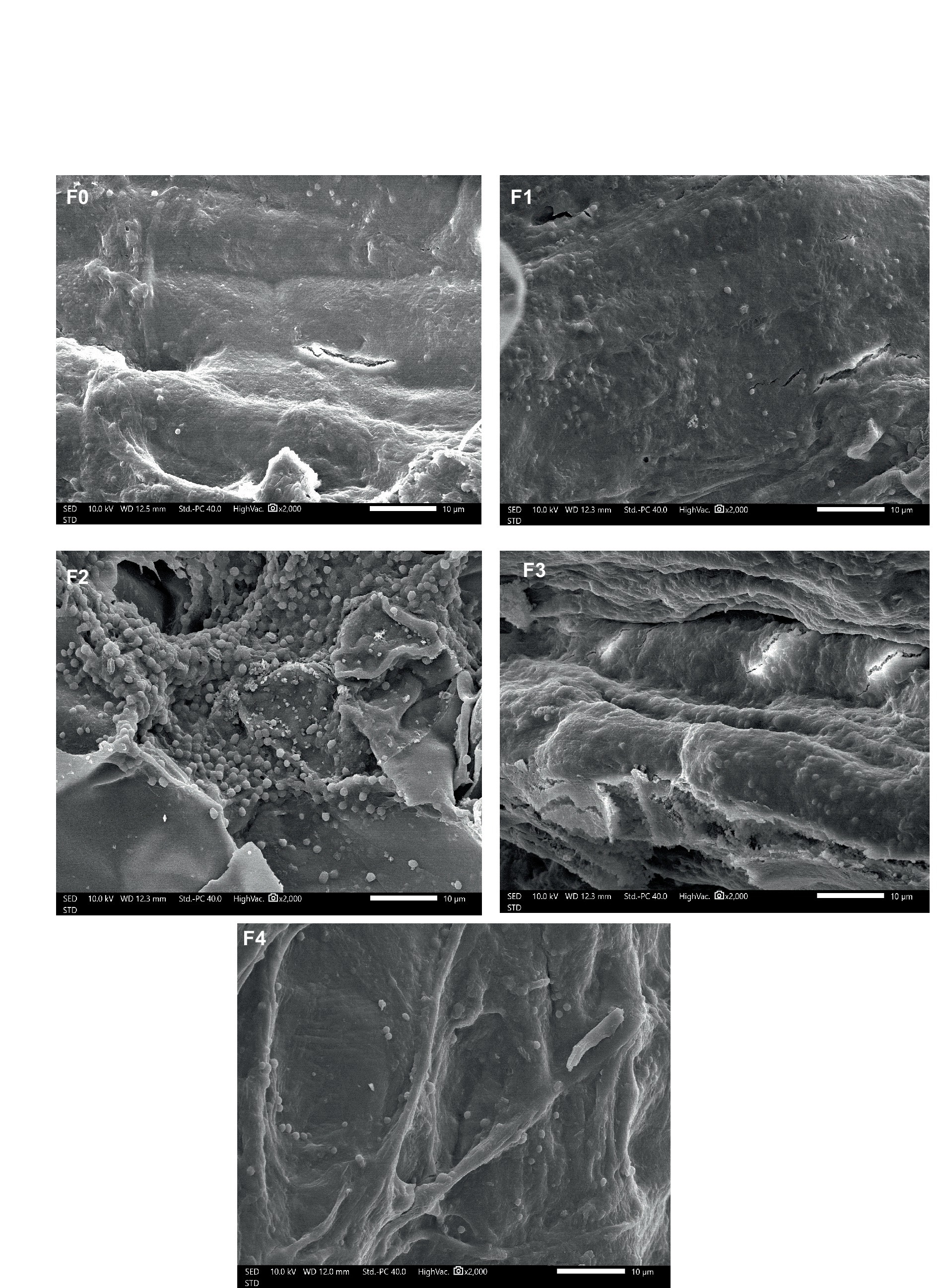
Figure 3. Colonization of rhizobacteria on the roots of T. paniculatum.
PGPR performs many functions, such as regulating hormones, nutrient balance, protecting plants from disease, and dissolving essential nutrients for plants. Furthermore, PGPR exhibits cooperative and opposing interactions with microorganisms in the rhizosphere and surrounding soil, indirectly enhancing plant growth rate (Vejan et al., 2016). PGPR enhances plant advantages via colonizing roots, synthesizing compounds, nitrogen fixation (Ahemad and Kibret, 2014), and solubilizing phosphates. The colonization of rooted rhizobacteria is intricately linked to the rhizobacteria’s capacity to synthesize hormones and enzymes (Sagar et al., 2020). The strain P. mandelii IB-Ki14, which can synthesize indole-3-acetic acid (IAA) under laboratory conditions, significantly elevated the levels of auxins in the roots by almost 100% and in the shoots by approximately 50% as compared to the control group in wheat plants (Akhtyamova et al., 2023). As per Hassan et al. (2019), certain rhizobacteria that live in plant roots can also invade the interior of the roots and proliferate. For rhizobacteria to colonize a rhizosphere, they must be able to navigate their movement in gradients of compounds that either attract or repel them. A unique environment for the interactions between plants and microbes is the rhizosphere. Root exudates are complex mixtures of chemo-effectors, which are made up of various substances. Rhizobacteria are drawn in, and connections between bacteria and roots are formed by chemotaxis towards root exudates (Feng et al., 2021).
Conclusion
The four Rhizobzcteria used significantly affected the growth and phytochemical content of T. paniculatum leaves as an ingredient in herbal tea at a concentration of 2% compared to controls. The four rhizobacteria can produce IAA hormones, fix nitrogen, and dissolve phosphorus. The four rhizobacteria can colonize the roots of T. paniculatum plants.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding authors.
Author contributions
NS: Writing – review & editing, Writing – original draft, Supervision, Project administration, Methodology, Investigation, Conceptualization. DS: Writing – original draft, Methodology, Investigation. IS: Writing – original draft, Methodology, Investigation. NR: Writing – review & editing, Investigation, Formal analysis. KP: Writing – review & editing, Validation, Formal analysis. NB: Writing – review & editing, Validation, Formal analysis. HE: Writing – review & editing, Validation, Formal analysis. TH: ___. RS: Writing – review & editing, Writing – original draft, Formal analysis, Data curation.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This research work and publication was supported by Innovation Awards, LPPM, Udayana University, No. B/1.124/UN14.4A/PT.01.03/2023 Researchers Supporting Project Number RSP2024R358, King Saud University, Riyadh, Saudi Arabia.
Acknowledgments
The authors would like to acknowledge the support provided by LPPM, Udayana University Innovation Award 2023, Udayana University, Bali Indonesia, Department of Biology Study Program at Udayana University in Bali, Indonesia, and Biopesticide Laboratory, Bali, Indonesia. The authors would like to acknowledge the support provided by Researchers Supporting Project Number RSP2024R358, King Saud University, Riyadh, Saudi Arabia.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ahemad, M., and Kibret, M. (2014). Mechanisms and applications of plant growth promoting rhizobacteria: current perspective. J. King Saud Univ. Sci. 26, 1–20. doi: 10.1016/j.jksus.2013.05.001
Ahmad, S. S., Ahmad, K., Hwang, Y. C., Lee, E. J., and Choi, I. (2023). Therapeutic applications of ginseng natural compounds for health management. Int. J. Mol. Sci. 24:17290. doi: 10.3390/ijms242417290
Akhtyamova, Z., Martynenko, E., Arkhipova, T., Seldimirova, O., Galin, I., Belimov, A., et al. (2023). Influence of plant growth-promoting Rhizobacteria on the formation of Apoplastic barriers and uptake of water and potassium by wheat Plants. Microorganisms 11:1227. doi: 10.3390/microorganisms11051227
Aslanidis, P. S. C., and Golia, E. E. (2022). Urban sustainability at risk due to soil pollution by heavy metals—case study: Volos, Greece. Land 11:1016. doi: 10.3390/land11071016
Chandran, H., Meena, M., and Swapnil, P. (2021). Plant growth-promoting rhizobacteria as a green alternative for sustainable agriculture. Sustainability (Switzerland) 13, 1–30. doi: 10.3390/su131910986
Chen, Q., Qiu, Y., Yuan, Y., Wang, K., and Wang, H. (2022). Biocontrol activity and action mechanism of Bacillus velezensis strain SDTB038 against fusarium crown and root rot of tomato. Front. Microbiol. 13, 1–18. doi: 10.3389/fmicb.2022.994716
Costea, L., Chițescu, C. L., Boscencu, R., Ghica, M., Lupuliasa, D., Mihai, D. P., et al. (2022). The polyphenolic profile and antioxidant activity of five vegetal extracts with Hepatoprotective potential. Plan. Theory 11:1680. doi: 10.3390/plants11131680
de Andrade, L. A., Santos, C. H. B., Frezarin, E. T., Sales, L. R., and Rigobelo, E. C. (2023). Plant growth-promoting Rhizobacteria for sustainable agricultural production. Microorganisms 11:1088. doi: 10.3390/microorganisms11041088
de la Osa, C., Rodríguez-Carvajal, M. Á., Gandullo, J., Aranda, C., Megías, M., Ollero, F. J., et al. (2021). Plant growth-promoting rhizobacteria modulate the concentration of bioactive compounds in tomato fruits. Separations 8, 1–16. doi: 10.3390/separations8110223
Delgado-Ramírez, C. S., Hernández-Martínez, R., and Sepúlveda, E. (2021). Rhizobacteria associated with a native solanaceae promote plant growth and decrease the effects of fusarium oxysporum in tomato. Agronomy, 11. doi: 10.3390/agronomy11030579
Del Rosario Cappellari, L., Chiappero, J., Palermo, T. B., Giordano, W., and Banchio, E. (2020). Volatile organic compounds from rhizobacteria increase the biosynthesis of secondary metabolites and improve the antioxidant status in Mentha Piperita L. grown under salt stress. Agronomy 10, 1–16. doi: 10.3390/agronomy10081094
Du, K., Huang, J., Wang, W., Zeng, Y., Li, X., and Zhao, F. (2024). Monitoring low-temperature stress in winter wheat using TROPOMI solar-induced chlorophyll fluorescence. IEEE Trans. Geosci. Remote Sens. 62, 1–11. doi: 10.1109/TGRS.2024.3351141
Ekawati, E. R. (2015). UJI EFEKTIFITAS PERASAN KUNYIT PUTIH (Curcuma Spp.) TERHADAP PERTUMBUHAN Escherechia Coli. STIGMA J. Sci. 8, 12–16.
Ekin, Z. (2019). Integrated use of humic acid and plant growth promoting rhizobacteria to ensure higher potato productivity in sustainable agriculture. Sustainability (Switzerland) 11:3417. doi: 10.3390/su10023417
Ezrari, S., Mhidra, O., Radouane, N., Tahiri, A., Polizzi, G., Lazraq, A., et al. (2021). Potential role of rhizobacteria isolated from citrus rhizosphere for biological control of citrus dry root rot. Plan. Theory 10:872. doi: 10.3390/plants10050872
Feng, H., Fu, R., Hou, X., Lv, Y., Zhang, N., Liu, Y., et al. (2021). Chemotaxis of beneficial rhizobacteria to root exudates: the first step towards root–microbe rhizosphere interactions. Int. J. Mol. Sci. 22:6655. doi: 10.3390/ijms22136655
Gallart, M., Paungfoo-Lonhienne, C., Gonzalez, A., and Trueman, S. J. (2021). Nitrogen source influences the effect of plant growth-promoting rhizobacteria (Pgpr) on macadamia integrifolia. Agronomy 11:1064. doi: 10.3390/agronomy11061064
Hassan, M. K., McInroy, J. A., and Kloepper, J. W. (2019). The interactions of rhizodeposits with plant growth-promoting rhizobacteria in the rhizosphere: a review. Agriculture (Switzerland) 9:142. doi: 10.3390/agriculture9070142
He, M., Ren, T., Jin, Z. D., Deng, L., Liu, H., Cheng, Y. Y., et al. (2023). Precise analysis of potassium isotopic composition in plant materials by multi-collector inductively coupled plasma mass spectrometry. Spectrochim. Acta B At. Spectrosc. 209:106781. doi: 10.1016/j.sab.2023.106781
Hlongwane, M. M., Mohammed, M., Mokgalaka, N. S., and Dakora, F. D. (2023). The Potential of Rhizobacteria to Mitigate Abiotic Stress in Lessertia frutescens. Plants 12:196 doi: 10.3390/plants12010196
Hosseini, A., Hosseini, M., and Schausberger, P. (2022). Plant growth-promoting Rhizobacteria enhance defense of strawberry Plants against spider mites. Front. Plant Sci. 12, 1–12. doi: 10.3389/fpls.2021.783578
Javaid, S., Mushtaq, S., Mumtaz, M. Z., Rasool, G., Naqqash, T., Afzal, M., et al. (2023). Mineral solubilizing Rhizobacterial strains mediated biostimulation of Rhodes grass seedlings. Microorganisms 11:2543. doi: 10.3390/microorganisms11102543
Jia, Q., Sun, J., Gan, Q., Shi, N., and Fu, S. (2024). Zea mays cultivation, biochar, and arbuscular mycorrhizal fungal inoculation influenced lead immobilization. Microbiol. Spectr. 12, e03427–e03423. doi: 10.1128/spectrum.03427-23
Khalilzadeh, R., Sharifi, R. S., and Jalilian, J. (2016). Antioxidant status and physiological responses of wheat (Triticum aestivum L.) to cycocel application and biofertilizers under water limitation conditions. J. Plant Interact. 11, 130–137. doi: 10.1080/17429145.2016.1221150
Khoso, M. A., Wagan, S., Alam, I., Hussain, A., Ali, Q., Saha, S., et al. (2024). Impact of plant growth-promoting rhizobacteria (PGPR) on plant nutrition and root characteristics: current perspective. Plant. Stress 11:100341. doi: 10.1016/j.stress.2023.100341
Kreatsouli, K., Fousteri, Z., Zampakas, K., Kerasioti, E., Veskoukis, A. S., Mantas, C., et al. (2019). A polyphenolic extract from olive mill wastewaters encapsulated in whey protein and maltodextrin exerts antioxidant activity in endothelial cells. Antioxidants 8:280. doi: 10.3390/antiox8080280
Kuan, K. B., Othman, R., Rahim, K. A., and Shamsuddin, Z. H. (2016). Plant growth-promoting rhizobacteria inoculation to enhance vegetative growth, nitrogen fixation, and nitrogen remobilization of maize under greenhouse conditions. PLoS One 11, 1–19. doi: 10.1371/journal.pone.0152478
Kumar, M., Giri, V. P., Pandey, S., Gupta, A., Patel, M. K., Bajpai, A. B., et al. (2021). Plant-growth-promoting rhizobacteria emerging as an effective bioinoculant to improve the growth, production and stress tolerance of vegetable crops. Int. J. Mol. Sci. 22:12245. doi: 10.3390/ijms222212245
Kumar, R., Swapnil, P., Meena, M., Selpair, S., and Yadav, B. G. (2022). Plant growth-promoting Rhizobacteria (PGPR): approaches to alleviate abiotic stresses for enhancement of growth and development of medicinal Plants. Sustainability (Switzerland) 14:15514. doi: 10.3390/su142315514
Kurniawan, I. D., Soedradjad, R., and Syamsunihar, R. (2014). Pengaruh Dosis Pupuk Organik terhadap Kandungan Fenolik dan Flavonoid Biji Tanaman Kedelai yang Berasosiasi Dengan Synechococcus sp. Berkala Ilmiah Pertanian 1, 1–3.
Lee, Y. H., Jang, H. J., Park, K. H., Kim, S. H., Kim, J. K., Kim, J. C., et al. (2021). Phytochemical analysis of the fruits of sea buckthorn (Hippophae rhamnoides): identification of organic acid derivatives. Plan. Theory 10, 1–11. doi: 10.3390/plants10050860
Liang, H., Wang, C., Lu, X., Sai, C., and Liang, Y. (2022). Dynamic changes in soil phosphorus accumulation and bioavailability in phosphorus-contaminated protected fields. Int. J. Environ. Res. Public Health 19:12262. doi: 10.3390/ijerph191912262
Liu, J., Zhang, J., Zhu, M., Wan, H., Chen, Z., Yang, N., et al. (2022). Effects of plant growth promoting Rhizobacteria (PGPR) strain Bacillus licheniformis with biochar amendment on potato growth and water use efficiency under reduced irrigation regime. Agronomy 12:1031. doi: 10.3390/agronomy12051031
Lestario, L. N., Christian, A. E., and Martono, Y. (2009). Aktivitas antioksidan daun Ginseng jawa (Talinum paniculatum Gaertn) Antioxidant Activity of Javanese Ginseng (Talinum paniculatum Gaertn) Leaves. Agritech 29, 71–78.
Lyu, D., Backer, R., Berrué, F., Martinez-Farina, C., Hui, J. P. M., and Smith, D. L. (2023). Plant growth-promoting Rhizobacteria (PGPR) with microbial growth broth improve biomass and secondary metabolite accumulation of Cannabis sativa L. J. Agric. Food Chem. 71, 7268–7277. doi: 10.1021/acs.jafc.2c06961
Mahdavikia, H., Rezaei-Chiyaneh, E., Rahimi, A., and Mohammadkhani, N. (2019). Effects of fertilizer treatments on antioxidant activities and physiological traits of basil (Ocimum basilicum L.) under water limitation conditions. J. Med. Plants Byprod. 8, 143–151.
Marcelino, S., Hamdane, S., Gaspar, P. D., and Paço, A. (2023). Sustainable agricultural practices for the production of medicinal and aromatic Plants: evidence and recommendations. Sustainability 15:14095. doi: 10.3390/su151914095
Maulina, N. M. I., Suprapta, D. N., Temaja, I. G. R. M., Adnyana, I. M., and Suriani, N. L. (2022). Rhizobacteria of Bali with obvious growth-promoting properties on corn (Zea mays L.). Front. Sustain. Food Syst. 6, 1–12. doi: 10.3389/fsufs.2022.899736
Mendes, A., Oliveira, A., Lameiras, J., Mendes-Moreira, P., and Botelho, G. (2023). Organic medicinal and aromatic Plants: consumption profile of a Portuguese consumer sample. Food Secur. 12:4145. doi: 10.3390/foods12224145
Nam, J. H., Thibodeau, A., Qian, Y. L., Qian, M. C., and Park, S. H. (2023). Multidisciplinary evaluation of plant growth promoting rhizobacteria on soil microbiome and strawberry quality. AMB Express 13:18. doi: 10.1186/s13568-023-01524-z
Ojuederie, O. B., Olanrewaju, O. S., and Babalola, O. O. (2019). Plant growth promoting rhizobacterial mitigation of drought stress in crop plants: implications for sustainable agriculture. Agronomy 9:712. doi: 10.3390/agronomy9110712
Perisoara, A., Marinas, I. C., Geana, E. I., Constantin, M., Angheloiu, M., Pirvu, L., et al. (2022). Phytostimulation and synergistic Antipathogenic effect of Tagetes erecta extract in presence of Rhizobacteria. Horticulturae 8:779. doi: 10.3390/horticulturae8090779
Plants, S. L., Alshallash, K. S., El-Taher, A. M., Azab, E. S., El-Raouf, H. S. A., and Ibrahim, M. F. M. (2022). Cyanobacteria on botanical characteristics of tomato. Plants, 11:2732. doi: 10.3390/plants11202732
Rizki, F. L., Made, N., Suarni, R., Suriani, N. L., Gusti, N., Manik, A., et al. (2023). Effect of Javanese ginseng (Talinum Paniculatum (Jacq.) Gaertn) leaf extract on spermatozoa quality of mice (Mus Musculus). East. J. Agric. Biol. Sci. 3, 1–8.
Sagar, A., Sayyed, R. Z., Ramteke, P. W., Sharma, S., Marraiki, N., Elgorban, A. M., et al. (2020). ACC deaminase and antioxidant enzymes producing halophilic Enterobacter sp. PR14 promotes the growth of rice and millets under salinity stress. Physiol. Mol. Biol. Plants 26, 1847–1854. doi: 10.1007/s12298-020-00852-9
Santoyo, G., Urtis-Flores, C. A., Loeza-Lara, P. D., Orozco-Mosqueda, M. D. C., and Glick, B. R. (2021). Rhizosphere colonization determinants by plant growth-promoting rhizobacteria (Pgpr). Biology 10, 1–18. doi: 10.3390/biology10060475
Su, F., Zhao, B., and Dhondt-Cordelier, S. (2024). Plant-growth-promoting Rhizobacteria modulate carbohydrate metabolism in connection with host plant defense mechanism. Int. J. Mol.Sci. 25:1465. doi: 10.3390/ijms25031465
Sukwan, C., Wray, S., and Kupittayanant, S. (2014). The effects of ginseng java root extract on uterine contractility in nonpregnant rats. Physiol. Rep. 2, 1–11. doi: 10.14814/phy2.12230
Suriani, N. L. (2019). Piper caninum blume leaf extract and compost to suppress blast disease and increase the production of Bali red rice (Oryza sativa) in the greenhouse. Int. Res. J. Eng. IT Sci. Res. 5, 46–54. doi: 10.21744/irjeis.v5n4.693
Suriani, N. L., Suprapta, D. N., Perveen, K., Suarsana, I. N., Made, N., Resiani, D., et al. (2021). Formulation of Brevibacillus Agri and compost to improve growth and phytochemicals compound of Piper caninum herbal plant. Food Syst. 7:1163426. doi: 10.3389/fsufs.2023.1163426.
Taie, H. A. A., Salama, Z. A. E. R., and Radwan, S. (2010). Potential activity of basil plants as a source of antioxidants and anticancer: EBSCOhost. Notulae Botanicae Horti Agrobotanici Cluj-Napoca 38, 119–127.
Tang, A., Haruna, A. O., and Majid, N. M. A. (2018). Potential PGPR properties of cellulolytic, nitrogen-fixing, and phosphate-solubilizing bacteria of a rehabilitated tropical forest soil. BioRxiv, 1–22.
Vafa, Z. N., Sohrabi, Y., Sayyed, R. Z., Suriani, N. L., and Datta, R. (2021). Effects of the combinations of rhizobacteria, mycorrhizae, seaweed, and supplementary irrigation on growth and yield in wheat cultivars. Plan. Theory 10, 1–17. doi: 10.3390/plants10040811
Vejan, P., Abdullah, R., Khadiran, T., Ismail, S., and Nasrulhaq Boyce, A. (2016). Role of plant growth promoting rhizobacteria in agricultural sustainability-a review. Molecules 21, 1–17. doi: 10.3390/molecules21050573
Vocciante, M., Grifoni, M., Fusini, D., Petruzzelli, G., and Franchi, E. (2022). The role of plant growth-promoting Rhizobacteria (PGPR) in mitigating plant's environmental stresses. Appl. Sci. (Switzerland) 12:1231. doi: 10.3390/app12031231
Wang, H., Peng, D., and Xie, J. (2009). Ginseng leaf-stem: Bioactive constituents and pharmacological functions. Chinese Medicine, 4:20. doi: 10.1186/1749-8546-4-20
Wang, D., Wang, C., Chen, Y., and Xie, Z. (2023). Developing plant-growth-promoting Rhizobacteria: a crucial approach for achieving sustainable agriculture. Agronomy 13, 1–5. doi: 10.3390/agronomy13071835
Xiang, J., Mlambo, R., Shaw, I., Seid, Y., Shah, H., He, Y., et al. (2023). Cryopreservation of bioflavonoid-rich plant sources and bioflavonoid-microcapsules: emerging technologies for preserving bioactivity and enhancing nutraceutical applications. Front. Nutr. 10:1232129. doi: 10.3389/fnut.2023.1232129
Yaoyao, E., Yuan, J., Yang, F., Wang, L., Ma, J., Li, J., et al. (2017). PGPR strain Paenibacillus polymyxa SQR-21 potentially benefits watermelon growth by re-shaping root protein expression. AMB Express 7:104. doi: 10.1186/s13568-017-0403-4
Keywords: chlorophyll, herbal crop, P solubilization, phytochemical, phytohormones, plant growth promotion, PGPR
Citation: Suriani NL, Suprapta DN, Suarsana IN, Resiani NMD, Perveen K, Bukhari NA, Ei Enshasy HA, Ho TS and Sayyed RZ (2024) Biopotential of rhizobacteria to improve growth and phytochemical content in Javanese ginseng (Talinum paniculatum) herbal plant. Front. Sustain. Food Syst. 8:1384700. doi: 10.3389/fsufs.2024.1384700
Edited by:
Muhammad Saqlain Zaheer, Khwaja Fareed University of Engineering and Information Technology (KFUEIT), PakistanReviewed by:
Hafiz Haider Ali, University of Arkansas, United StatesRihab Djebaili, Consorzio Interuniversitario Nazionale per le Scienze Ambientali, Italy
Copyright © 2024 Suriani, Suprapta, Suarsana, Resiani, Perveen, Bukhari, Ei Enshasy, Ho and Sayyed. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ni Luh Suriani, bmlsdWhzdXJpYW5pQHVudWQuYWMuaWQ=; R. Z. Sayyed, c2F5eWVkcnpAZ21haWwuY29t