 Roghayeh Rahmani1
Roghayeh Rahmani1 Shiva Khalesro
Shiva Khalesro Gholamreza Heidari
Gholamreza Heidari Ali Mokhatssi-Bidgoli
Ali Mokhatssi-Bidgoli- 1Department of Agronomy and Plant Breeding, Faculty of Agriculture, University of Kurdistan, Sanandaj, Iran
- 2Department of Agronomy, Faculty of Agriculture, Tarbiat Modares University, Tehran, Iran
The exogenous application of organic and natural inputs is a key strategy for producing healthy and high-quality crops in sustainable agricultural systems. Black cumin (Nigella sativa L.) is a highly popular plant used worldwide in the medical and food industries. According to the authors' knowledge, no research has been published to examine the effects of zeolite and vermicompost on yield components, nitrogen (N), phosphorous (P), and potassium (K) contents in seeds, essential and fixed oil contents, or the composition of black cumin in the organic agricultural system. In a semi-arid area of Iran in 2017 and 2018, an experiment with a full factorial layout was conducted using a randomized complete block design with three replications. The experimental treatments included four vermicompost rates (0, 2, 4, and 6 tons ha−1) and four zeolite rates (0, 3, 6, and 9 tons ha−1). There were linear responses between either vermicompost or zeolite application rates and the majority of the attributes studied. The integrated treatment of 6 tons of vermicompost and 9 tons of zeolite ha−1 produced the maximum seed and biological yields (466.2 and 3716.7 kg ha−1, respectively). The utilization of 6 tons of vermicompost ha−1 increased the N, P, and K contents of seed by 13.5%, 10.8%, and 14.1%, respectively, compared with the control. Seed essential oil content was enhanced by 24.1% in plots treated with 9 tons of zeolite ha−1 compared to the untreated control. The use of 6 tons of vermicompost ha−1 resulted in higher production of unsaturated fatty acids such as linoleic (53.3%), oleic (25.36%), and linolenic acid (0.6%) in oil. Overall, both the quantity and quality of black cumin improved when vermicompost and zeolite were used for 2 consecutive years. This showed the agronomic potential of both amendments in promising and environmentally friendly agricultural systems.
1. Introduction
In the 21st century, environmental degradation is one of the most critical agricultural issues. Soil organic matter decrement is a key facet of soil degradation, such as loss of soil fertility and capacity to produce crops (Karami et al., 2023). The arid and semi-arid agricultural soils do not have enough organic matter, cannot hold enough water, and are getting worse (Guo et al., 2019). The intensive use of synthetic inorganic inputs such as chemical fertilizers has resulted in soil organic matter depletion, groundwater contamination, air pollution, and production quality reductions in the regions (Hernandez et al., 2015; Zambon et al., 2017). Hence, sustainable strategies, such as organic fertilizers and natural inputs, are becoming more important to enhance soil organic matter and plant yield (Yang et al., 2013; Vafadar-Yengeje et al., 2019). Exogenous application of vermicompost and zeolite in organic systems was thought to be an ecological and environmentally friendly strategy for producing high-quality plant products and increasing yield stability (Hosseinzadeh et al., 2021).
The use of organic fertilizers such as vermicompost is a key part of organic systems. Vermicompost is a great soil amendment and a good source of nutrients for plants, especially nitrogen, phosphorus, potassium, calcium, and magnesium during the growing season (Arancon et al., 2005; Hait and Tare, 2011; Yang et al., 2013). Vermicompost not only increases soil organic matter but also decreases environmental pollution in terms of recycling waste into valuable organic fertilizer (Nagavallemma et al., 2006). Moreover, vermicompost improves soil respiration, biomass, and soil organism abundance (Xu et al., 2016). The favorable effects of vermicompost application on the yield and biochemical profile of Dracocephalum moldavica L., Moringa oleifera Lam., and nutrient uptake of Ocimum basilicum L. have been previously documented (Vafadar-Yengeje et al., 2019; Guzman-Albores et al., 2020; Celikcan et al., 2021).
Zeolites are a natural substance that can help improve soil, water storage, and the availability of soil nutrients. They do this by increasing the soil's cation exchange capacity without changing its structure much and by reducing nitrogen leaching out of the soil (Leggo, 2000; Powlson et al., 2008; Tsintskaladze et al., 2016). Nitrogen leaching losses in agroecosystems enhance the potential for human health impacts from contaminated drinking water sources as well as environmental degradation such as water body eutrophication (Diaz and Rosenberg, 2008). Hence, research on agricultural systems leading to management practices that improve nitrogen use efficiency (NUE) and reduce N losses is vital. When zeolite was incorporated into the soil, plants such as Zea mays L. (Ippolito et al., 2011), Trigonella foenum-graecum L. (Baghbani-Arani et al., 2017), and Allium cepa L. (Bybordi et al., 2017) would induce their growth and metabolism. Therefore, zeolite application, due to its properties such as water storage and slow nutrient release, could be an appropriate input in the current global water crisis, especially in Iran.
Black cumin (Nigella sativa L.) is an annual herb that is part of the Ranunculaceae family. It is currently used a lot in the food and medicine industries (Hosseini et al., 2018; Shahbazi, 2019). This crop is farmed mostly in dry and semi-arid areas of the Middle East, Western Asia, the Mediterranean, and central Europe (Randhawa and Alghamdi, 2011; Ozer et al., 2020). In Iran, black cumin is cultivated in various areas and is widely used in people's diets and traditional medicine (Rezaei-Chiyaneh et al., 2018). Essential and fixed oil, protein, saponin, and alkaloid are all found in black cumin seeds. The fixed oil has an abundance of unsaturated fatty acids, including linoleic and oleic acids. This plant is antiviral, antibacterial, antipyretic, carminative, and relaxing to the muscles (Ali and Blunden, 2003; Majdalawieh et al., 2010). The demand for Nigella sativa L. oil is growing worldwide (Asif and Ansari, 2019). Furthermore, many food and pharmaceutical firms employ components generated from plant farming methods that are organic (Fonseca-Santos et al., 2015). Hence, producing high-quality Nigella sativa L. and improving its oil in sustainable agricultural systems are of great importance.
Research studies have shown that vermicompost and zeolite can have a positive impact on plant growth in various species (Bybordi et al., 2017; Celikcan et al., 2021; HabibiSharafabad et al., 2022). However, there is limited information available regarding their effects on Nigella sativa L., including any potential interaction between the two amendments. With the negative impacts of chemical fertilizers and the increasing importance of sustainable agricultural systems, this study aimed to assess if these organic inputs can provide a natural and effective alternative for increasing Nigella sativa L. yield and improving its nutrient content and essential and fixed oils. The objective was also to confirm the potential interaction of vermicompost and zeolite on the agronomic performance and fatty acid composition of Nigella sativa L. while reducing the cost and toxicity of chemical fertilizers. According to the scientific hypothesis, exogenous application of vermicompost and zeolite at various levels can improve both the quantitative and qualitative traits of Nigella sativa L., offering a useful strategy for sustainable production in soils where soil organic matter and water availability are the main factors limiting crop growth.
2. Material and methods
2.1. Study site conditions and experimental design
A 2-year experiment was carried out from April 2017 to September 2018 at the Research Field (latitude of 35° 19′ N, longitude 47° 18′ E, and 1865 m above sea level), University of Kurdistan, Sanandaj, Iran. The long-term average temperature is 12.1°C, and the annual precipitation is 311.6 mm for the area. The monthly summary of weather data recorded each year is presented in Figure 1. Before planting, soil samples from a depth of 30 cm were taken at the site of the experiment to find out about its physical and chemical properties (Table 1). The soil was loamy in texture.
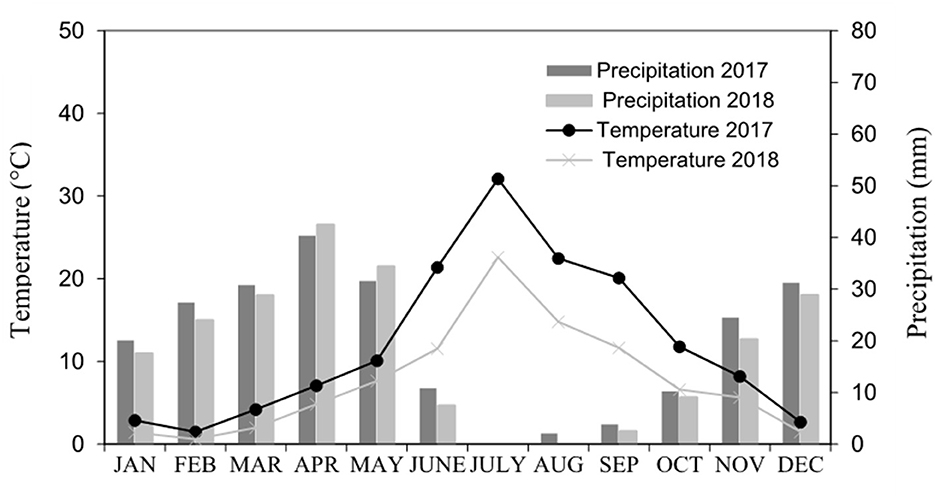
Figure 1. Monthly mean temperature (°C) and total precipitation (mm) in 2017 and 2018 growing seasons in the experimental area.
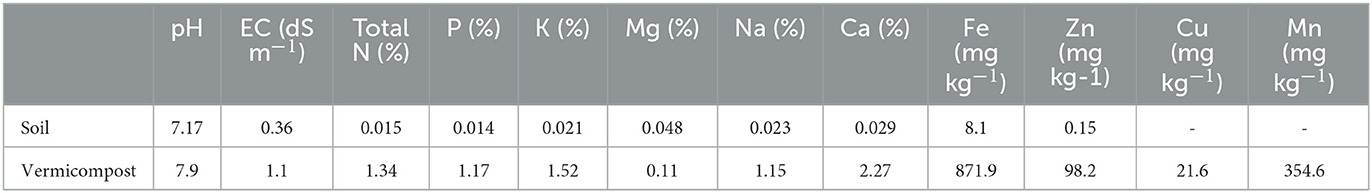
Table 1. Chemical characteristics of the soil of the experimental field and vermicompost used in the experiment.
The research was set up as a factorial experiment using a randomized complete block design with three replications. Treatment combinations included four vermicompost rates (V0 = 0, V2 = 2, V4 = 4, and V6 = 6 tons ha−1) and four zeolite rates (Z0 = 0, Z3 = 3, Z6 = 6, and Z9 = 9 tons ha−1). Each plot's vermicompost and zeolite were added into the soil before planting. Table 1 shows the chemical properties of vermicompost. The zeolite (clinoptilolite type) contained 0.01% P2O5, 0.03% TiO2, 0.04% MnO, 1.5% Fe2O3, 65% SiO2, 12.02% Al2O3, 3.0% K2O, 1.08% Na2O, 0.1% MgO, and 12.3% CaO. The black cumin seeds were sown on 26th April 2017 and 27th April 2018. Each experimental plot was comprised of six rows of 3 m each, spaced 30 cm apart, with a 2-m unseeded alley. Seeds (PakanBazr Co. Isfahan, Iran) were manually sown to a 0.5–1 cm depth at approximately 5 cm intervals. After seed sowing, irrigation was carried out immediately. The field was irrigated once a week using drip irrigation (Mirabesfahan Company, Iran). Weeds were removed by hand as required. In accordance with organic management practices, plants did not receive any chemical inputs such as fertilizers and herbicides.
2.2. Determining black cumin properties and harvesting
2.2.1. Morphological traits, yield components, and biological and seed yield
At the mature stage, five plants were chosen at random from each plot to measure the morphological variables (plant height and branch number) and yield components (1,000 seed weight, seed number per capsule, and capsule number per plant). In the center rows of each plot, 1 m2 of area was harvested at the end of each growing season on 12th September 2017 and 14th September 2018, respectively. To determine biological yield (all above-ground parts of plants), black cumin samples were dried in the shade and cool conditions for 2 weeks after harvest. Then, the seeds were separated and weighted to calculate the seed yield.
2.2.2. Nitrogen, phosphorus, and potassium seed contents
The samples were digested with H2SO4-H2O2 and then tested for total N content using the kjeldahl technique (Bremner, 1996), P content using the photometric method (yellow vanadium molybdate), and K content using a flame photometer (Jones et al., 1991).
2.2.3. Seed essential oil content and yield
The essential oil from the samples (15 g of powdered seeds from each sample) was extracted for 3 h in a Clevenger water distillation device using the specified process (Analytical Methods Committee, 1988). Essential oil production was computed using seed yield and essential oil content (Amani Machiani et al., 2018).
2.2.4. Seed fixed oil content and yield
To extract the fixed oil samples (5 g of powdered seeds from each sample), a Soxhlet extraction with n-hexane solvent was used. The extracted oil was isolated via rotary liquid solvent evaporation (Leal et al., 2009). The recovered oil was collected, weighed, and analyzed for its fatty acid profile. The fixed oil yield was calculated by multiplying the seed yield by the fixed oil percentage in the seed (Rezaei-Chiyaneh et al., 2020).
2.2.5. Fatty acid analysis
Metcalf et al. (1966) method was used to turn fatty acids into their methyl esters (FAMEs), and FAMEs were evaluated using gas chromatography (Model Agilent 7890A, GC system) with a fused silica capillary column DB WAX (60 m, 0.25 i.d.) and a flame ionization detector (Wilmington, DE, USA). The oven temperature was set to 5 min at 170°C, then 4 min at 190°C, then 15 min at 190°C. As a carrier gas, nitrogen was employed. The injector and detector had temperatures of 260°C and 220°C, respectively. By comparing their retention times to those of pure standards, the FAMEs were discovered (Sigma-Aldrich, St. Louis, MO).
2.3. Statistical analysis
SAS v. 9.4 used the MIXED procedure with the type 3 method option to analyze data (SAS Institute, Cary, NC, USA). The main impacts of vermicompost and zeolite rates, as well as their two-way interactions, were all fixed effects. Random effects were years, replicates × years, and years × vermicompost rates × zeolite rates. Using the PDIFF option for least square means (LS-means), the means were compared. When there were significant two-way interactions between experimental components, the SLICE technique in SAS was employed to examine them. A partitioned analysis of the LS-means for an interaction may be carried out using the SLICE statement, which offers a general mechanism for carrying out such an analysis. To analyze the response of black cumin to vermicompost and zeolite rates, linear and quadratic regression analyses were conducted, and significant models (P ≤ 0.05) were reported.
3. Results
3.1. Morphological traits and yield components measurement
Zeolite and vermicompost had significant impacts on plant height, number of capsules per plant, number of branches per plant, and 1,000 seed weight (Table 2). Zeolite and vermicompost had a strong two-way interaction for capsule number per plant and 1,000 seed weight (Table 2).
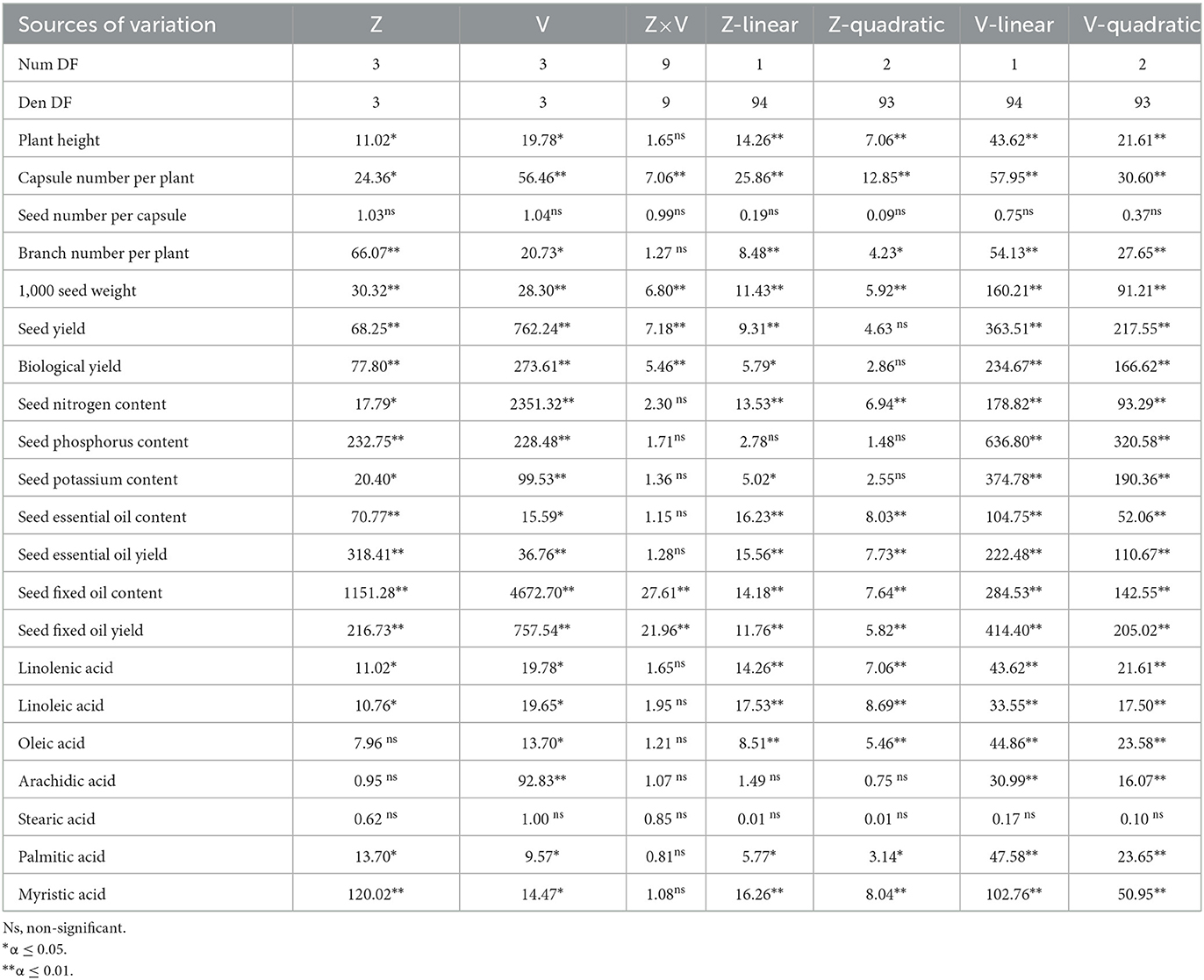
Table 2. Analysis of variance (F value) and analysis of regression (F value of linear and quadratic models) of traits of Nigella sativa L. influenced by zeolite (Z) and vermicompost rates (V).
The tallest plants (54.0 and 52.5 cm) were observed for 6 (V6) and 4 (V4) tons of vermicompost ha−1, respectively. By contrast, the shortest plants (49.6 cm) were observed in the control (V0) (Figure 2). Plant height increased significantly as vermicompost rates increased. The application of each ton of vermicompost ha−1 increased plant height by 7.5 mm (Figure 2). In addition, using the zeolite increased plant height by 3.2 mm for each ton ha−1 (Figure 2). The tallest (50.3 cm) and shortest plants (53.2 cm) belonged to the 9 (Z9) and 0 (Z0) tons of zeolite ha−1, respectively (Figure 2).
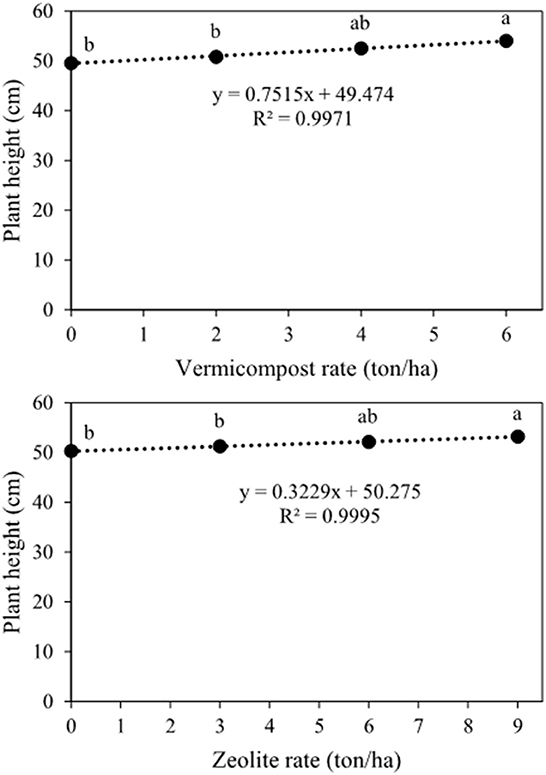
Figure 2. Main effects of vermicompost and zeolite rates on plant height of Nigella sativa L. The different letters show significantly different at the level of 0.05. Significant linear relationships are given.
The interaction of vermicompost and zeolite on the number of capsules per plant was significant (Table 2). Figure 3 shows these interactions by slicing in different rates of vermicompost and zeolite. The first chart of Figure 3A demonstrated that the lowest number of capsules per plant (4.7) belonged to the treatment without vermicompost and zeolite (V0Z0). In this figure, the number of capsules per plant for V2Z0, V4Z0, and V6Z0 was 5.9, 6.4, and 6.1, respectively. Furthermore, the maximum capsule number per plant (6.4) was predicted with the application of 4.3 tons of vermicompost ha−1. The second chart showed that the amounts of this trait for V0Z3, V2Z3, V4Z3, and V6Z3 were 5.4, 6.2, 6.8, and 7.4, respectively. Indeed, when 3 tons of zeolite ha−1 was applied, the value of this trait significantly increased by 0.3 with each ton of vermicompost ha−1 (Figure 3A). Based on the results of the third chart, the capsule numbers per plant for V0Z6, V2Z6, V4Z6, and V6Z6 were 5.4, 6.2, 6.8, and 7.4, respectively. The maximum capsule number per plant (7.6) was predicted using 5.7 tons of vermicompost ha−1 when combined with 6 tons of zeolite ha−1 (Figure 3A). The fourth chart showed that the highest capsule number per plant (8.9) was observed in the integrated treatment V6Z9 with more slope (0.49) than those of other zeolite rates (Figure 3A). Another slicing (Figure 3B) showed that the capsule numbers per plant for V0Z0, V0Z3, V0Z6, and V0Z9 were 4.7, 5.4, 5.6, and 5.9, respectively (Figure 3B). The amounts of this trait for V6Z0, V6Z3, V6Z6, and V6Z9 were 6.1, 7.4, 7.5, and 8.9, respectively. Indeed, the capsule number per plant increased by 0.1, 0.1, 0.2, and 0.3 with the application of each ton of zeolite ha−1 combined with 0, 2, 4, and 6 tons of vermicompost ha−1, respectively (Figure 3B).
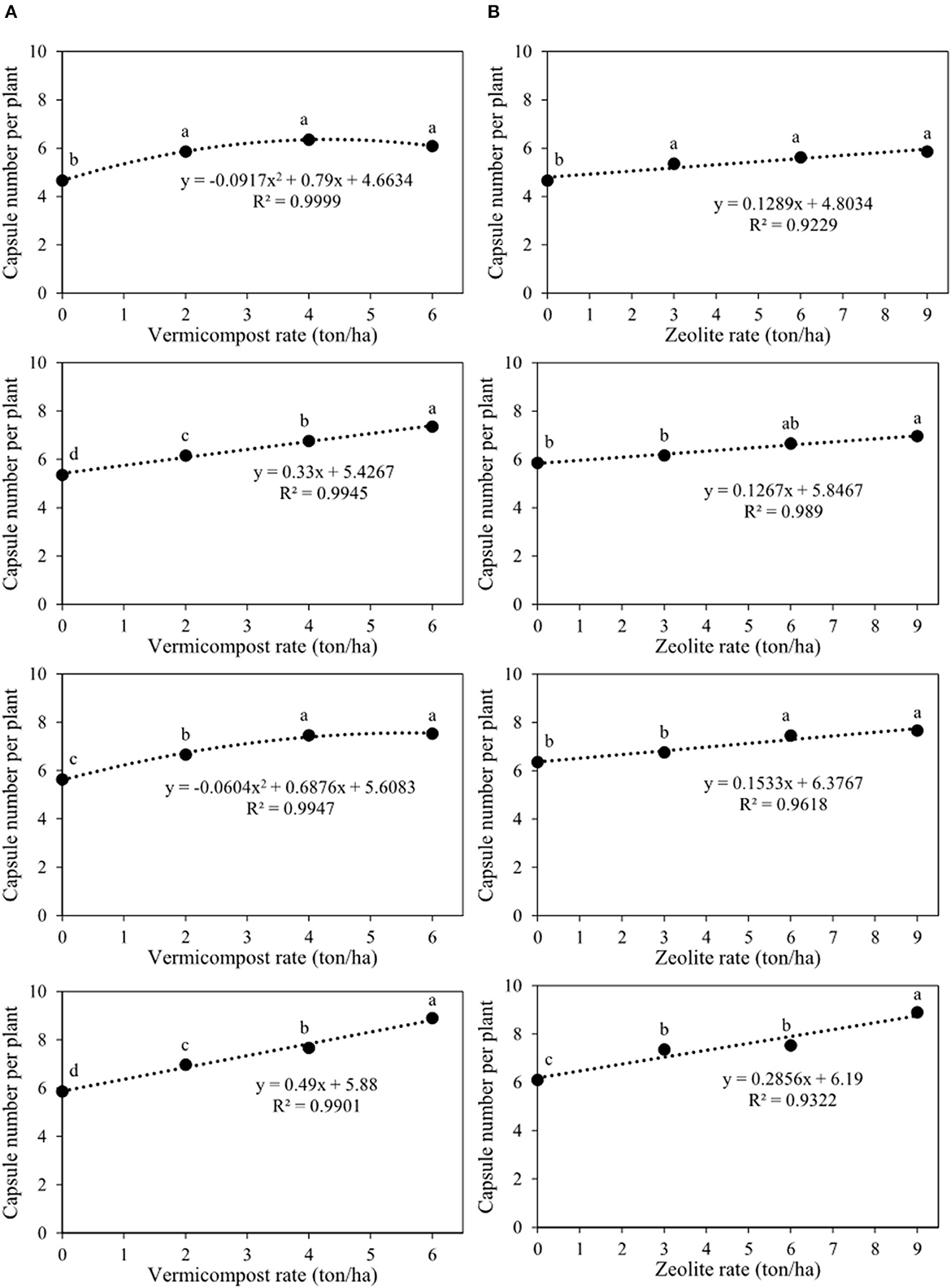
Figure 3. Interaction between vermicompost and zeolite rates on capsule number per plant of Nigella sativa L. (A) sliced by zeolite rates (0, 3, 6, 9 tons ha−1); (B) sliced by vermicompost rates (0, 2, 4, 6 tons ha−1) from top to bottom, respectively). The different letters show significantly different at the level of 0.05. Significant linear and quadratic relationships are given.
Table 2 shows that the number of seeds per capsule (66.4–82.0) did not change significantly among vermicompost and zeolite rates.
The lowest (4.0) and highest (5.8) branch numbers per plant were observed in 0 (V0) and 6 (V6) tons of vermicompost ha−1, respectively (Figure 4). There was no significant difference between V4 and V6 treatments (Figure 4). When compared to the control, applying 6, 4, and 2 tons of vermicompost ha−1 significantly increased branch number per plant by 44.2%, 34.2%, and 19.4%, respectively (Figure 4). In addition, the values for this trait increased by 17.9%, 11.2%, and 4.3%, compared with the control, when plots were treated with 3, 6, and 9 tons of zeolite ha−1 (Figure 4).

Figure 4. Main effects of vermicompost and zeolite rates on branch number per plant of Nigella sativa L. The different letters show significantly different at the level of 0.05. Significant linear relationships are given.
Figure 5A shows that the 1,000 seed weights for V0Z0, V2Z0, V4Z0, and V6Z0 were 2.05, 2.23, 2.27, and 2.29 g, respectively. Indeed, for 0 ton zeolite ha−1, the highest 1,000 seed weight was predicted with the application of 5 tons of vermicompost ha−1 (Figure 5A). The second, third, and fourth charts of Figure 5A showed that at different rates of zeolite (3, 6, and 9 tons ha−1), the highest values of this trait were observed with the application of 6 tons of vermicompost ha−1. Thus, these values were 2.31, 2.32, and 2.34 for the integrated treatments of V6Z3, V6Z6, and V6Z9, respectively (Figure 5A). In another slicing, Figure 5B, the fourth chart demonstrated that the 1,000 seed weights were 2.29, 2.31, 2.32, and 2,34 g for V6Z0, V6Z3, V6Z6, and V6Z9, respectively. There was no significant difference between V6Z3, V6Z6, and V6Z9. Indeed, for each ton added increment of zeolite in combination with 0, 2, 4, and 6 tons vermicompost ha−1, 1,000 seed weight increased by 16.9, 2.7, 4.2, and 5.5 mg, respectively.
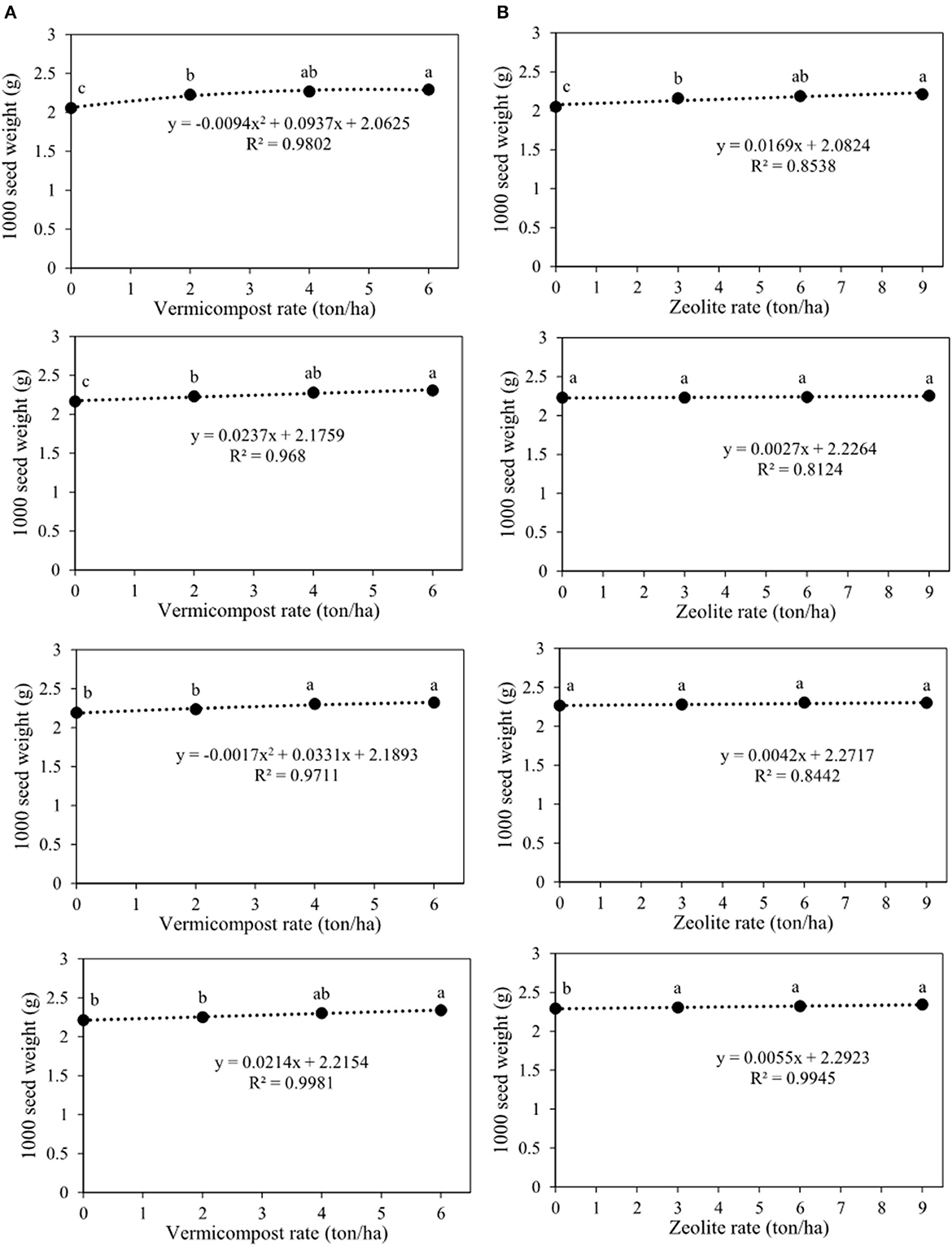
Figure 5. Interaction between vermicompost and zeolite rates on 1,000 seed weight of Nigella sativa L. (A) sliced by zeolite rates (0, 3, 6, 9 tons ha−1). (B) sliced by vermicompost rates (0, 2, 4, 6 tons ha−1) from top to bottom, respectively). The different letters show significantly different at the level of 0.05. Significant linear and quadratic relationships are given.
3.2. Seed and biological yield
Biological and seed yields were affected by all treatments and their interactions (Table 2). Seed yield significantly increased with increasing vermicompost rates at all zeolite rates (Figure 6A). Additionally, all the charts in Figure 6A showed that at different rates of zeolite (0, 3, 6, and 9 tons ha−1), the highest seed yield was observed with the application of 6 tons of vermicompost ha−1. Thus, these values were 957.4, 968.5, 1006.4, and 1112.3 kg ha−1 for the integrated treatments of V6Z0, V6Z3, V6Z6, and V6Z9, respectively (Figure 5A). Overall, the treatment without vermicompost and zeolite (V0Z0) and the integrated treatment of 6 tons of vermicompost and 9 tons of zeolite ha−1 (V6Z9) resulted in the lowest and highest seed yields (466.1 and 1112.3 kg ha−1), respectively (Figure 6A). Seed yields for each ton of zeolite ha−1 in combination with 0, 2, 4, and 6 tons of vermicompost ha−1 were increased by 25.5, 12.9, 6.8, and 16.8 kg ha−1, respectively (Figure 6B). The highest biological yields of 3424, 3417, 3536, and 3717 kg ha−1 were predicted with the application of 0, 3, 6, and 9 tons of zeolite ha−1 in combination with 6 tons of vermicompost ha−1 (Figure 7A). The lowest biological yield (1900.0 kg ha−1) belonged to the control treatment (V0Z0). The biological yield increased linearly as the zeolite rate increased in tandem with all vermicompost rates (Figure 7B).
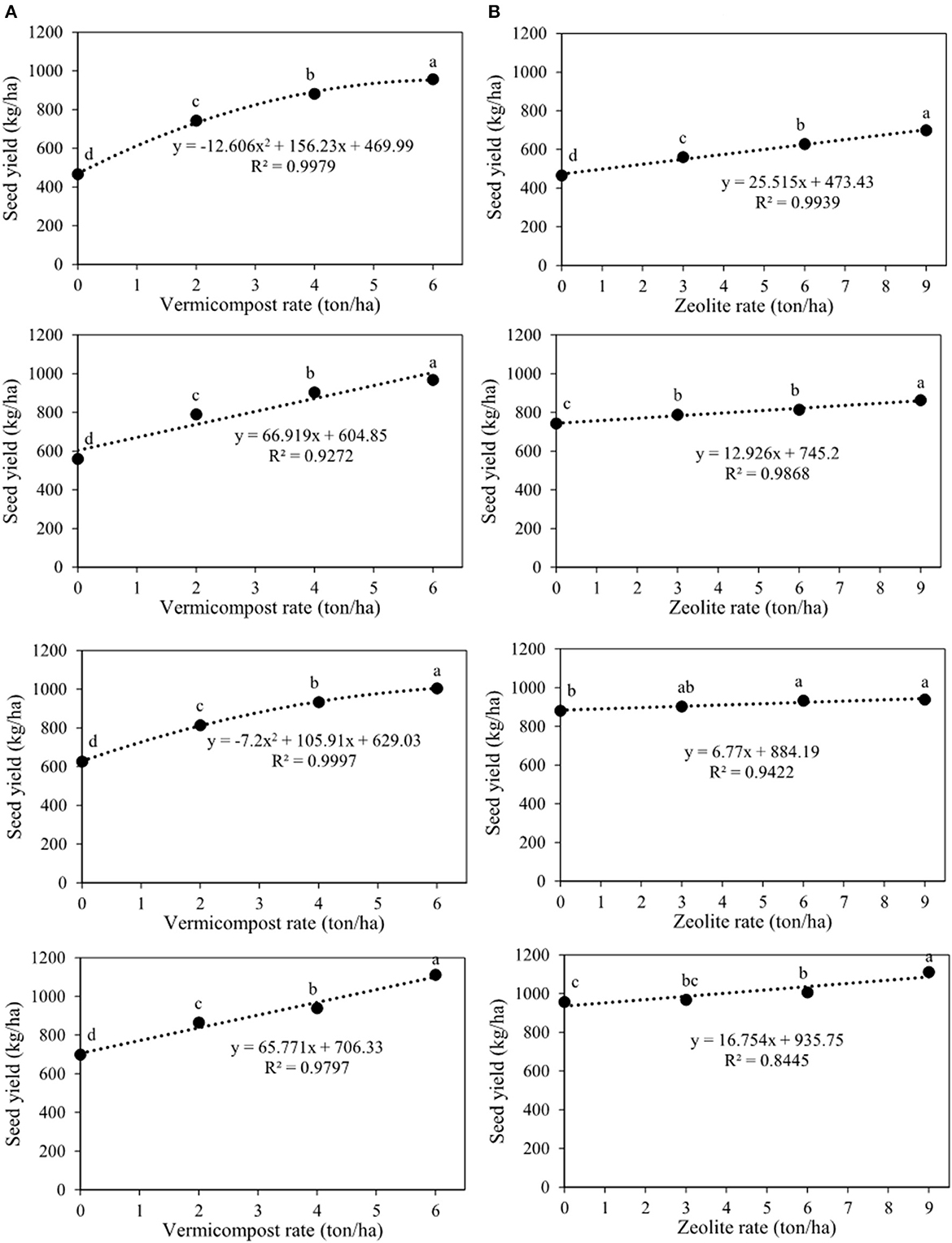
Figure 6. Interaction between vermicompost and zeolite rates on seed yield of Nigella sativa L. (A) sliced by zeolite rates (0, 3, 6, 9 tons ha−1). (B) sliced by vermicompost rates (0, 2, 4, 6 tons ha−1) from top to bottom, respectively). The different letters show significantly different at the level of 0.05. Significant linear and quadratic relationships are given.
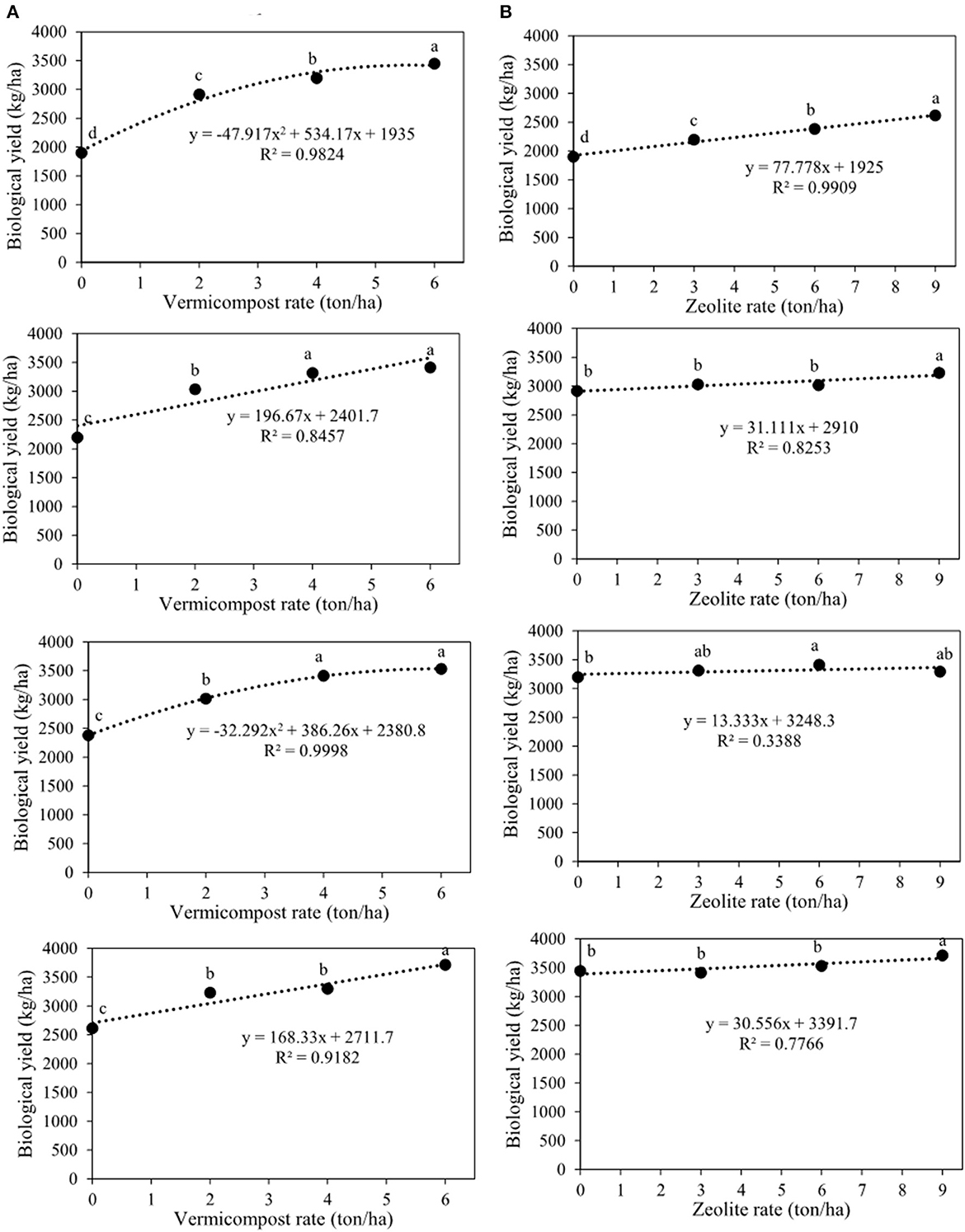
Figure 7. Interaction between vermicompost and zeolite rates on biological yield of Nigella sativa L. (A) sliced by zeolite rates (0, 3, 6, 9 tons ha−1). (B) sliced by vermicompost rates (0, 2, 4, 6 tons ha−1) from top to bottom, respectively). The different letters show significantly different at the level of 0.05. Significant linear and quadratic relationships are given.
3.3. Contents of nitrogen, phosphorus, and potassium in seed
The analysis of variance showed that the N, P, and K contents of seed were influenced by the main effects of vermicompost and zeolite (Table 2). These contents increased by increasing both zeolite and vermicompost rates (Table 3). The lowest (3.12%) and highest (3.54%) seed nitrogen content belonged to 0 and 6 tons of vermicompost ha−1, respectively. Also, the highest values of P (0.68%) and K (1.73%) seed content were observed in V6. The application of 6 tons of vermicompost ha−1 increased N, P, and K seed contents by 13.5%, 10.8%, and 14.1%, respectively, when compared with the control (Table 3). The highest seed contents of N (3.43%), P (0.65%), and K (1.65%) were reached with 9 tons of zeolite ha−1, with no significant difference from 6 tons of zeolite ha−1 (Table 3).

Table 3. Mean comparison of the main effects of vermicompost and zeolite rates on seed nitrogen, phosphorus, and potassium content and fatty acid composition of Nigella sativa L. The different letters show significantly different at the level of 0.05.
3.4. Seed essential oil content and yield
The main effect of vermicompost and zeolite on the seed essential oil content and yield was significant (Table 2). The essential oil content and yield tended to be linearly related to the application of vermicompost and zeolite (Figures 8, 9). The highest seed essential oil content (0.25%) and yield (2.52 kg ha−1) were observed in the highest rate of vermicompost (Figures 8, 9). The application of 6 tons of vermicompost ha−1 enhanced seed essential oil content by 16.0%, 28.5%, and 50.9%, respectively, when compared with 4, 2, and 0 tons of vermicompost ha−1 (Figure 8). Essential oil yield was increased by 251.5 g ha−1 for each ton of vermicompost ha−1 applied (Figure 9). The application of 9 tons of zeolite ha−1 significantly increased seed essential oil content by 8.2%, 14.4%, and 24.1%, respectively, when compared with 6, 3, and 0 tons of zeolite ha−1 (Figure 8). The highest (2.1 kg ha−1) and lowest (1.5 kg ha−1) seed essential oil yields were observed for 9 and 0 tons of zeolite ha−1, respectively (Figure 9).
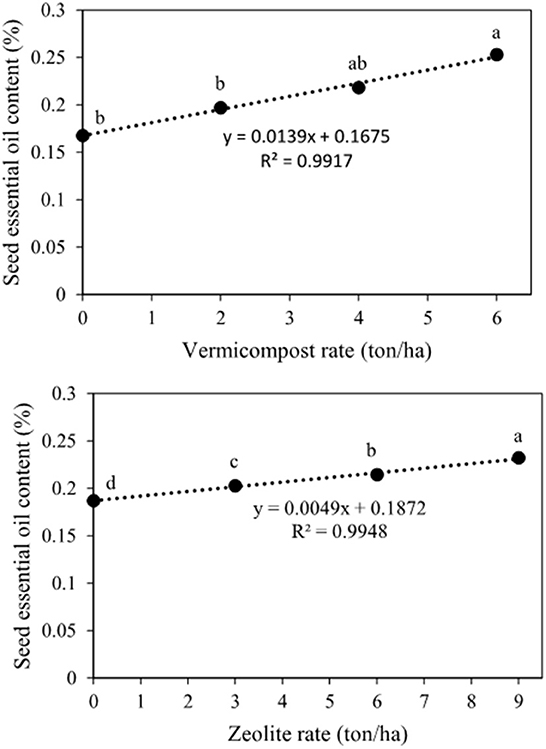
Figure 8. Main effects of vermicompost and zeolite rates on seed essential oil content of Nigella sativa L. The different letters show significantly different at the level of 0.05. Significant linear relationships are given.
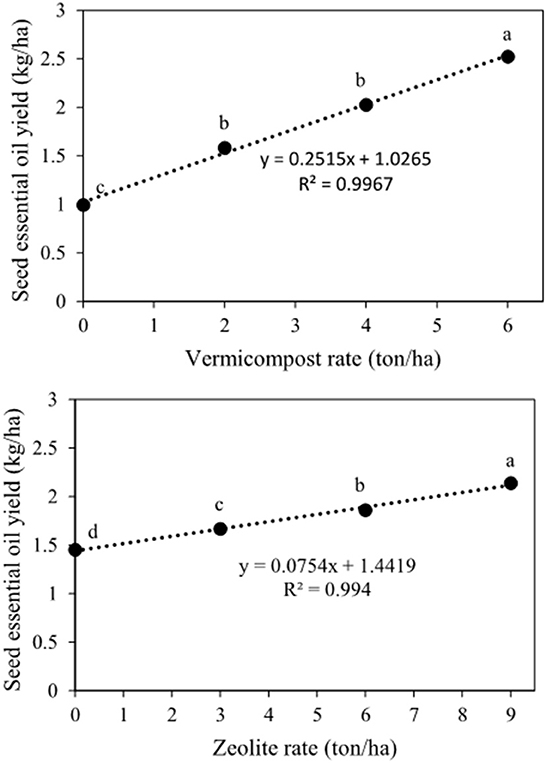
Figure 9. Main effects of vermicompost and zeolite rates on seed essential oil yield of Nigella sativa L. The different letters show significantly different at the level of 0.05. Significant linear relationships are given.
3.5. Seed fixed oil content and yield
On seed fixed oil content and yield, the primary impacts of zeolite and vermicompost and their interaction were significant (Table 2). Seed fixed oil content and yield improved with increasing vermicompost rates from 0 to 6 tons ha−1 in each of the four zeolite rates (Figures 10A, 11A). The lowest (8.9%) and highest (17.9%) seed fixed oil content belonged to V0Z0 and V6Z9, respectively (Figure 10A). The highest seed fixed oil yield using 6 tons of vermicompost ha−1 without zeolite (V6Z0) was 139.1 kg ha−1 (Figure 11A). In other zeolite rates, the highest seed fixed oil yield belonged to the V6Z3 (114.6 kg ha−1), V6Z6 (135.9 kg ha−1), and V6Z9 (200.2 kg ha−1), respectively (Figure 11A). Indeed, in these zeolite rates, response to increments of each ton of vermicompost ha−1 ranged from 14.1 to 18.7 kg of seed fixed oil yield ha−1 (Figure 11A). The other slicing showed that in all vermicompost rates except for 6 tons ha−1, there were no significant differences between the integrated treatment of vermicompost with 9 and 6 tons of zeolite ha−1 (Figure 10B). With the application of 6 tons of vermicompost ha−1, there was a significant difference among all zeolite rates (Figure 10B). Applying 9 tons of zeolite and 6 tons of vermicompost ha−1 (V6Z9) significantly increased seed fixed oil content by 15.6%, 34.0%, and 44.0% compared with control, respectively (Figure 10B). The lowest (83.0 kg ha−1) and highest (200.2 kg ha−1) seed fixed oil content belonged to V0Z0 and V6Z9, respectively (Figure 11B). Indeed, each ton of zeolite ha−1 combined with 0, 2, 4, and 6 tons of vermicompost ha−1 increased seed fixed oil yield by 4.5, 2.5, 2.0, and 6.9 kg ha−1, respectively (Figure 11B).
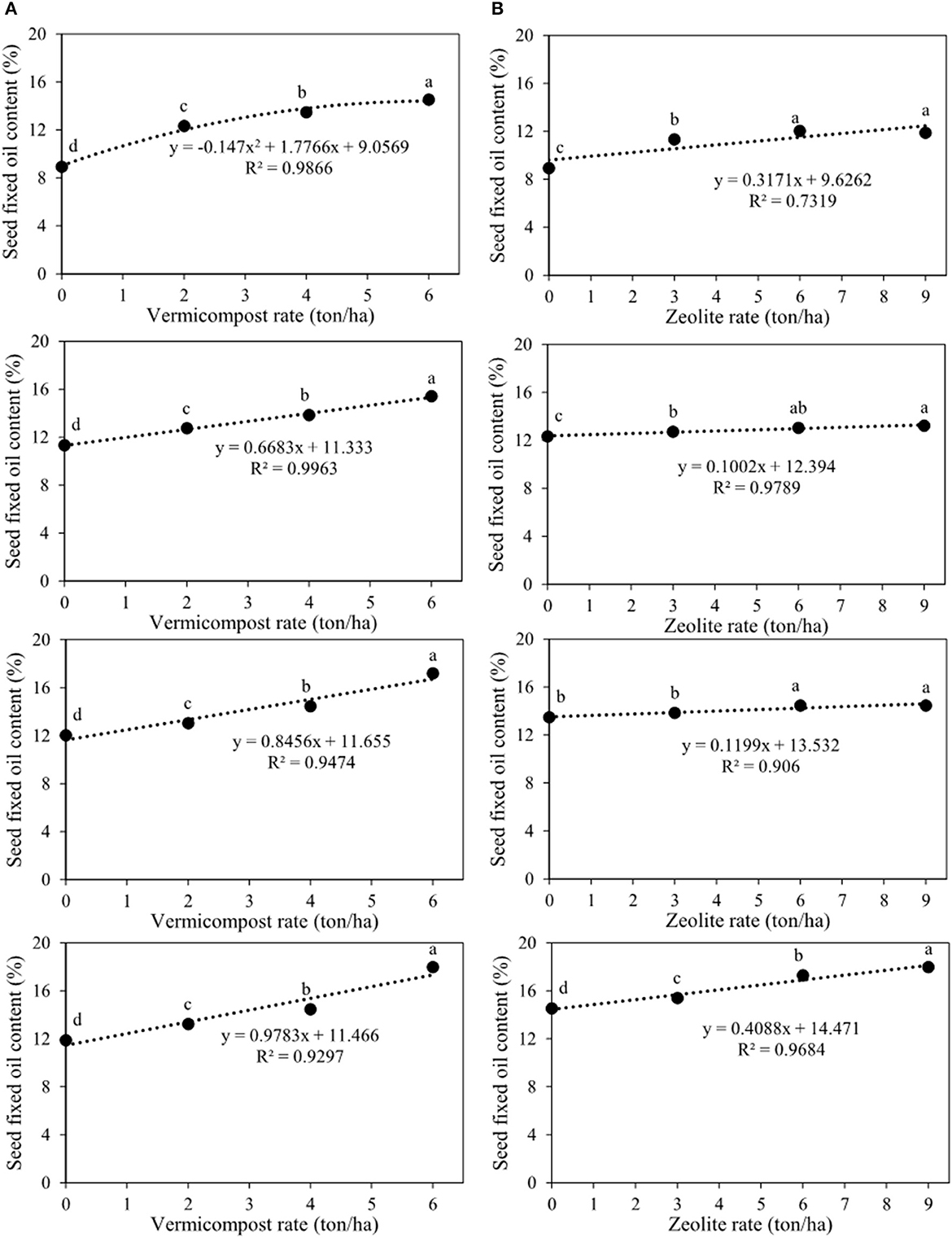
Figure 10. Interaction between vermicompost and zeolite rates on seed fixed oil content of Nigella sativa L. (A) sliced by zeolite rates (0, 3, 6, 9 tons ha−1). (B) sliced by vermicompost rates (0, 2, 4, 6 tons ha−1) from top to bottom, respectively). The different letters show significantly different at the level of 0.05. Significant linear and quadratic relationships are given.
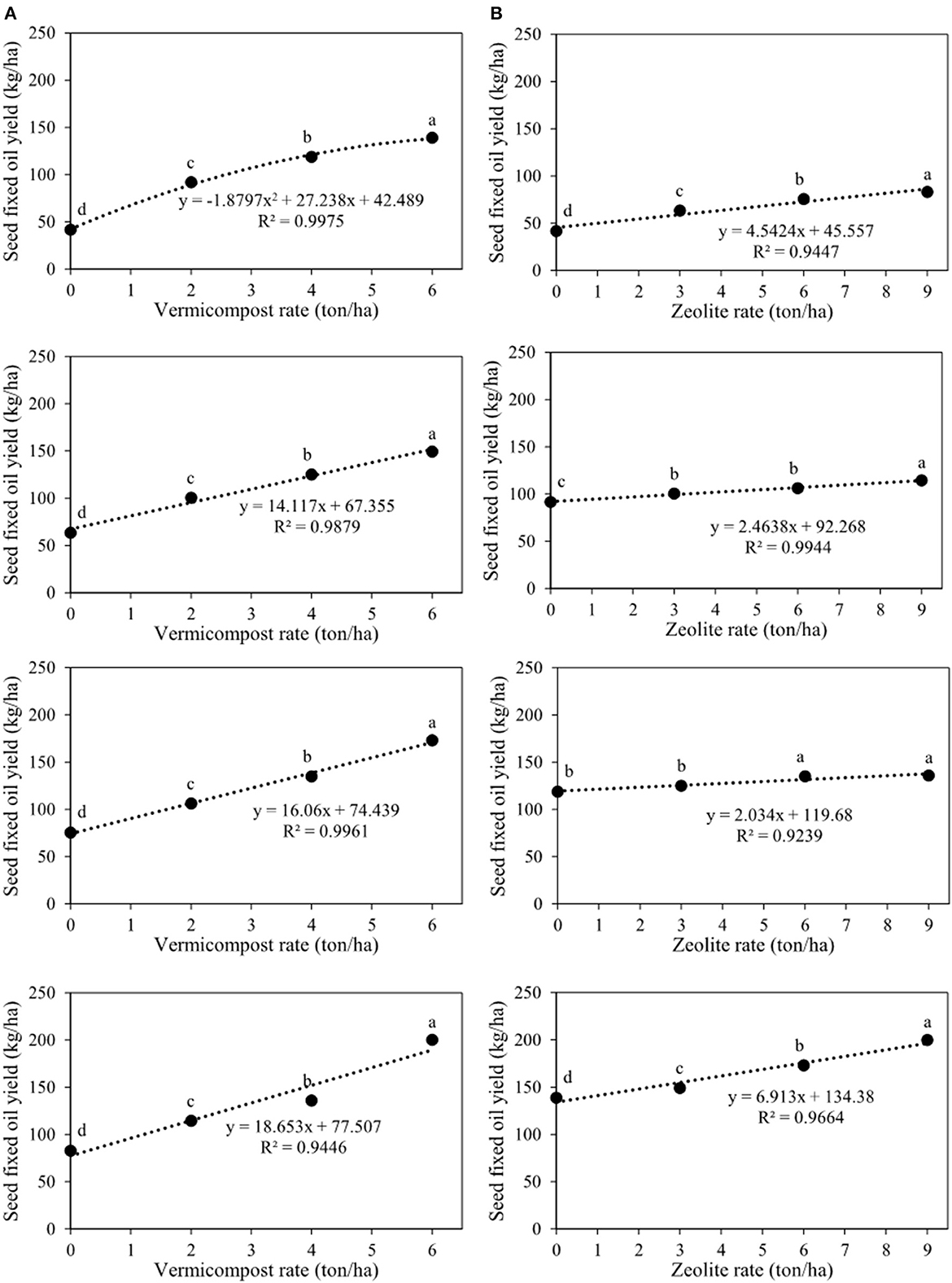
Figure 11. Interaction between vermicompost and zeolite rates on seed fixed oil yield of Nigella sativa L. (A) sliced by zeolite rates (0, 3, 6, 9 tons ha−1). (B) sliced by vermicompost rates (0, 2, 4, 6 tons ha−1) from top to bottom, respectively). The different letters show significantly different at the level of 0.05. Significant linear and quadratic relationships are given.
3.6. Fatty acid compositions
The analysis of variance showed that vermicompost significantly influenced linolenic, linoleic, oleic, arachidic, palmitic, and myristic acids (Table 2). The effect of zeolite on all fatty acids was significant except for oleic, arachidic, and stearic acids (Table 2). The highest concentrations of linolenic (0.64%), linoleic (53.33%), and oleic (25.14%) acids were found for 6 tons of vermicompost ha−1 (Table 3). There was no significant difference between 4 and 6 tons of vermicompost ha−1 for 6 tons of vermicompost ha−1. The lowest concentrations of linolenic (0.51%), linoleic (51.87%), and oleic (24.37%) acids belonged to the control treatment. Applying 9 tons of zeolite ha−1 increased linolenic and linoleic acids by 15.2% and 2.3%, respectively, when compared to the control treatment. By increasing vermicompost rates, arachidic, palmitic, and myristic acids decreased. Using 9 tons of zeolite ha−1 significantly decreased palmitic acid concentrations compared with the other rates. The most myristic acid (0.34%) was found in 0 ton of zeolite ha−1 and the least (0.23%) was found in 9 tons of zeolite ha−1 (Table 3).
4. Discussion
This study provides novel evidence on the role of vermicompost and zeolite in black cumin, for which experimental data are scarce in the literature. Although there are many studies on the effect of the mentioned treatments on different plants, generalization is restricted, and consensus exists on the need to consider specific interactions between vermicompost and zeolite on the studied plants. Based on the results, plant height and branch number of black cumin were higher in the plots treated with vermicompost. The crucial significance of organic nitrogen in boosting vegetative growth, cell divisions, and elongation may provide an explanation (Celikcan et al., 2021). On the other hand, vermicompost is an organic-rich source of macro- and micro-nutrients, especially nitrogen, which may boost plant growth and biomass by facilitating nutrient absorption over the growing season according to plant requirements. It can be concluded that vermicompost increases plant growth and biomass by absorbing and releasing nutrients. Indeed, vermicompost contains nutrients in forms that are readily taken up by the plants, such as nitrates and available phosphorus, soluble potassium, calcium, and magnesium. In addition, vermicompost regulates the growth of nitrogen-fixing microorganisms in the rhizosphere, which enhances N availability by making biologically fixed N available through the intimate mixing of ingested particles with soil (Wang et al., 2017). Furthermore, vermicompost contains biologically active substances such as plant growth regulators and a large number of functional microorganisms (Yang et al., 2013). This soil amendment could improve soil structure, optimize the nutrient cycle to improve food security, enhance moisture retention, improve soil aeration and soil-plant relations, increase photosynthesis rate, and accelerate the plant growth mechanism (Xu et al., 2016). Additionally, vermicompost may improve the environment for plant development by boosting the number and activity of soil microorganisms (Arancon et al., 2005). It can be concluded that by using vermicompost, the soil's physiochemical and biological traits and, consequently, crop growth can be improved. These results are in agreement with Roy and Singh (2006), Rekha et al. (2013), and Jami et al. (2021), who reported that vermicompost increased plant height of Hordeum vulgare L., branch number of Vigna mungo L. Hepper., and flower number of Crocus sativus L., respectively. Zeolite application increased plant height and branch number of black cumin in the present study. Zeolite is able to improve plant growth by increasing the long-term availability of water and nutrients (Ippolito et al., 2011). Due to its porous structure, zeolite can store water and develop humidity in the rhizosphere horizontally (Treacy and Higgins, 2007). Utilizing zeolite can increase vegetative growth due to selective absorption, controlled release of nutrients, decreased nitrification rate, and reduced nitrogen leaching (Leggo, 2000). The small molecular size of the open-ringed structure can physically protect NH4+ ions against microbial nitrification (Ippolito et al., 2011). So, it seems that increasing nitrogen, phosphorus, and potassium uptake helped black cumin grow taller and make more branches. In line with this result, other studies showed that zeolite increased the growth of Aleo vera L. (Hazrati et al., 2017) and Allium cepa L. (Bybordi et al., 2017), with a significant role in retaining water and soil macro- and micro-nutrients.
Based on the results, the interaction between vermicompost and zeolite had a significant positive effect on seed and biological yields. It can be said that the integrated treatments of vermicompost and zeolite increase yield components (capsule number per plant and 1000 seed weight) and morphological traits (plant height and branch number), and as a result, seed and biological yield. Because the integrated treatments had better canopy development, it is safe to say that making nitrogen more available makes plants absorb more sunlight, speed up photosynthesis, and produce more black cumin yield. In addition, yield enhancement can be attributed to an increase in soil microbial biomass after vermicompost application due to more enzymes, hormones, or humate content in the vermicompost (Wang et al., 2017; Celikcan et al., 2021). The presence of chelating siderophores in vermicompost results in these compounds chelating the minerals and consequently stabilizing mineral nutrients in the soil (Nardi et al., 2016; Jami et al., 2021). Furthermore, zeolite has high cation exchange capacity, selective absorption, and structure stability over the long term. Thus, zeolite improves vermicompost efficiency by enhancing soil cation exchange capacity (Bybordi et al., 2017). Thus, the utilization of vermicompost and zeolite improves soil physical properties in the rhizosphere by maintaining soil moisture and nutrients and indirectly enhancing crop productivity as a result. These results correspond with the findings of Baghbani-Arani et al. (2017), who reported that fenugreek yield was increased by consuming vermicompost and zeolite. Furthermore, other researchers reported the positive effect of vermicompost on the yields of Ocimum basilicum L. (Anwar et al., 2005), Sorghum bicolor L. Moench. (Kumar et al., 2005), and Zea mays L. (Liu et al., 2019).
Vermicompost and zeolite had substantial impacts on seed N, P, and K contents as well as yield and content of essential oil. Vermicompost may promote root development by retaining more water and nutrients in the soil, secreting more plant hormones, and increasing the amount of inorganic material such as N, P, and K that plants can absorb (Arancon et al., 2005). In addition, zeolite has a vital role in the repair of soil cation exchange capacity in terms of nitrogen leaching reduction and fertilizer availability increment in the rhizosphere (Ippolito et al., 2011; Hazrati et al., 2017). Hence, increasing vermicompost and zeolite rates of application led to an enhancement in seed N, P, and K contents (Table 3). The enhanced nutrients are used in the photosynthetic process of the plant. This observation is also evident from the higher transport of nitrogen to the shoots and grains and the subsequent enhancement in biological and seed yield. Other studies have found that vermicompost and zeolite increase the N content of Lavandula angustifolia L. (HabibiSharafabad et al., 2022), the P content of Vigna unguiculata L. Walp. (SailajaKumari and Ushakumari, 2002), and the K content of Achillea millefolium L. (Harb and Mahmoud, 2009). In this study, vermicompost and zeolite increased essential oil content. This may be attributed to the vital role of phosphorus in secondary metabolite synthesis (Sailo and Bagyaraj, 2005). Regarding the role of phosphorus in ATP and NADPH, it can be said that phosphorus is an essential element to supply energy for terpenoid compound synthesis. Increasing the availability of N and P stimulates the biochemical processes to produce terpene compounds of essential oil and accumulate secondary metabolites in the plants, which is in line with our findings (Table 3). Higher seed yield and seed essential oil content (Figures 6, 8) as a result of vermicompost and zeolite treatments could explain the increase in seed essential oil yield (Figure 9). Similarly, other researchers reported that zeolite and vermicompost significantly increased the essential oil content and yield of Thymus vulgaris L. (Zaghloul et al., 2016), Ocimum basilicum L. (Anwar et al., 2005), Ocimum basilicum L. (Celikcan et al., 2021), and Dracocephalum moldavica L. (Vafadar-Yengeje et al., 2019).
The combination treatments of vermicompost and zeolite in this research had the greatest seed fixed oil content and yield (Figures 10, 11). This may be explained by the fact that plants treated with vermicompost and zeolite got more water and nutrients than plants not receiving organic inputs. This enhances seed yield, growth, photosynthesis, and, consequently, fixed oil yield. Macro- and micro-nutrients, especially K, enhance the metabolism and transformation of carbohydrates and influence seed fixed oil content. In other words, proper K nutrition for the crop is a must to get superior quality, and potassium is the most important for many crop quality characteristics (Kumar et al., 2019). Thus, in the current study, vermicompost and zeolite, due to the availability of nutrients such as seed K content (Figure 10), could affect the seed fixed oil content of black cumin. These results are similar to those reported by Zahedi et al. (2009), who reported that zeolite increased the fixed oil of Brassica napus L.
Applying 6 tons of vermicompost ha−1 significantly increased unsaturated fatty acids, including linolenic, linoleic, and oleic acids, and decreased saturated fatty acids, consisting of arachidic, palmitic, and myristic acids. Vermicompost can alter the soil's biological characteristics and macro- and micro-nutrients uptake by plants. It may affect the enzymes and genes involved in the biosynthesis of fatty acids (He et al., 2020). Hence, it can be said that organic compounds can affect the biosynthesis and chain reaction of fatty acids. Furthermore, using 9 tons of zeolite ha−1 had a similar effect on these fatty acids. Regarding the higher amount of unsaturated fatty acids, it can be said that organic and natural inputs have a positive effect on the oil quality of black cumin. These results are in line with those of Hosseinzadeh et al. (2021), who reported that organic fertilizers such as vermicompost-enhanced linoleic acid in Portulaca oleracea L. This could be attributed to the good utilization of nutrients supplied for fixed oil metabolism (Schroder and Kopke, 2012). These findings were similar to those reported for the positive effects of organic fertilizer on the fatty acid composition of Nigella sativa L. (Seyyedi et al., 2015) and Foeniculum vulgare Mill. (Rezaei-Chiyaneh et al., 2020).
5. Conclusion
The plant known as black cumin (Nigella sativa L.) is a valuable plant. Current research showed that vermicompost and zeolite, as organic and natural inputs, had a favorable impact on the quantitative and qualitative features of Nigella sativa. Used together, vermicompost and zeolite treatments greatly boosted seed and biological yield, as well as their constituent parts. The application of vermicompost and zeolite enhanced the N, P, and K contents of the seed. Therefore, seed essential oil content and yield increased in plants treated with these inputs. Vermicompost and zeolite also improved seed fixed oil content and fatty acid composition. This led to more unsaturated fatty acids such as linolenic and linoleic acids.
Data availability statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding author.
Author contributions
SK conceived the idea and supervised the methodology with the assistance of GH. RR conducted the fieldwork. RR and SK collected the data. SK wrote the manuscript. AM-B analyzed the data and revised the manuscript. All authors contributed to and approved this manuscript.
Acknowledgments
The authors would like to express their gratitude to the University of Kurdistan for providing financial assistance in the form of grants.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ali, B. H., and Blunden, G. (2003). Pharmacological and toxicological properties of Nigella sativa. Phytother. Res. 17, 299–305. doi: 10.1002/ptr.1309
Amani Machiani, M., Javanmard, A., Morshedloo, M. R., and Maggi, F. (2018). Evaluation of yield, essential oil content and compositions of peppermint (Mentha piperita L.) intercropped with faba bean (Vicia faba L.). J. Clean. Prod. 171, 529–537. doi: 10.1016/j.jclepro.10062
Analytical Methods Committee. (1988). Application of gas-liquid chromatography to the analysis of essential oil. Part XIV. Monographs for five essential oils. Analyst 113, 1125–1136. doi: 10.1039/AN9881301125
Anwar, M., Patra, D. D., Chand, S., Alpesh, K., Naqvi, A. A., and Khanuja, S. P. S. (2005). Effect of organic manures and inorganic fertilizer on growth, herb and oil yield, nutrient accumulation, and oil quality of French basil. Commun. Soil Sci. Plant Anal. 36, 1737–1746. doi: 10.1081/CSS-200062434
Arancon, N. Q., Edwards, C. A., Bierman, P., Metzger, J. D., and Lucht, C. (2005). Effects of vermicompost produced from cattle manure, food waste and paper waste on the growth and yield of peppers in the field. Pedobiologia 49, 297–306. doi: 10.1016/j.pedobi.2005.02.001
Asif, A., and Ansari, M. Y. K. (2019). Generation of mutant lines of Nigella sativa L. by induced mutagenesis for improved seed yield. Ind Crop Prod. 132, 111552. doi: 10.1016/j.indcrop.2019.111552
Baghbani-Arani, A., ModarresSanavy, S. A. M., Mashhadi Akbar Boojar, M., and MokhtassiBidgoli, A. (2017). Toward improving the agronomic performance, chlorophyll fluorescence parameters and pigments in fenugreek using zeolite and vermicompost. Ind. Crops. Prod. 109, 346–357. doi: 10.1016/j.indcrop.08049
Bremner, J. M. (1996). “Nitrogen total,” in Methods of Soil Analysis. Part 3. Chemical Methods, ed S. D. L. Parks Madison, USA: SSSA and ASA, pp. 535–550.
Bybordi, A., Sadat, S., and Zargaripour, P. (2017). The effect of zeolite, selenium and silicon on qualitative and quantitative traits of onion grown under salinity conditions. Arch. Agron. Soil Sci. 64, 520–530. doi: 10.1080/03650340.2017.1373278
Celikcan, F., Kocak, M. Z., and Kulak, M. (2021). Vermicompost applications on growth, nutrition uptake and secondary metabolites of Ocimum basilicum L. under water stress: a comprehensive analysis. Ind. Crops Prod. 171, 1–12. doi: 10.1016/j.indcrop.2021.113973
Diaz, R. J., and Rosenberg, R. (2008). Spreading dead zones and consequences for marine ecosystems. Science 321, 926–929. doi: 10.1126/science.1156401
Fonseca-Santos, B., Antonio Corrêa, M., and Horill, M. (2015). Sustainability, natural and organic cosmetics: consumer, products, efficacy, toxicological and regulatory considerations. SciELO Anal. 51, 17–26. doi: 10.1590/S198482502015000100002
Guo, L., Shen, J., Li, B., and Li, Q. Q. (2019). Impacts of agricultural land use change on soil aggregate stability and physical protection of organic C. Sci. Total Environ. 707, 136049. doi: 10.1016/j.scitotenv.2019.136049
Guzman-Albores, J. M., Montes-Molina, J. A., Castan~on-Gonz'alez, J. H., Abud-Archila, M., Guti'errez-Miceli, F. A., and Ruiz-Valdiviezo, V. M. (2020). Effect of different vermicompost doses and water stress conditions on plant growth and biochemical profile in medicinal plant, Moringa oleifera Lam. J. Environ. Biol. 41, 240–246. doi: 10.22438/jeb/41/2/MRN-1164
HabibiSharafabad, Z., Abdipour, M., Hosseinifarahi, M., Kelidari, A., and Rashidi, L. (2022). Integrated humic acid and vermicompostionghanges essential oil quantity, and quality in field-grown in Lavandula angustifolia L. intercropped with Brassica nigra L. Ind. Crop. Prod. 178, 1–12. doi: 10.1016/j.indcrop.2022.114635
Hait, S., and Tare, V. (2011). Optimizing vermistabilization of waste activated sludge using vermicompost as bulking material. Waste Manage. 31, 502–511. doi: 10.1016/j.wasman.11004
Harb, E. M. Z., and Mahmoud, M. A. (2009). “Enhancing of growth, essential oil yield and components of yarrow plant (Achillea millefolium L.) growth under safe agriculture conditions using zeolite and compost,” in Proceedings of 4th Conference on Recent Technologies in Agriculture, Giza, Egypt.
Hazrati, S., Tahmasebi-Sarvestani, Z., Modarres-Sanavy, S. A. M., Mokhtassi-Bidgoli, A., Mohammadi, H., Nicola, S., et al. (2017). Effects of zeolite and water stress on growth: yield and chemical composition of Aloe vera L. Agric. Water Manage. 181, 66–72. doi: 10.1016/j.agwat.11026
He, M., Qin, C. X., Wang, X., and Ding, N. Z. (2020). Plant unsaturated fatty acids: biosynthesis and regulation. Front. Plant Sci. 11, 390. doi: 10.3389/fpls.2020.00390
Hernandez, A., Miranda, M., Arellano, E. C., Saura, S., and Ovalle, C. (2015). Landscape dynamics and their effect on the functional connectivity of a Mediterranean landscape in Chile. Ecol. Indic. 48, 198–206. doi: 10.1016/j.ecolind.08010
Hosseini, S. S., Nadjafi, F., Asareh, M. S., and Rezadoost, H. (2018). Morphological and yield related traits, essential oil and oil production of different landraces of black cumin (Nigella sativa) in Iran. Sci. Hortic. 233, 1–8. doi: 10.1016/j.scienta.01038
Hosseinzadeh, M. H., Ghalavand, A., Mashhadi-Akbar Boojar, M., Modaress Sanavy, S. A. M., and Mokhtassi-Bidgoli, A. (2021). Application of manure and biofertilizer to improve soil properties and increase grain yield, essential oil and ω3 of purslane (Portulaca oleracea L.) under drought stress. Soil Tillage Res. 205, 1–11. doi: 10.1016/j.still.2020.104633
Ippolito, A. J., Tarkalson, D. D., and Lehrsch, G. A. (2011). Zeolite soil application method affects inorganic nitrogen, moisture, and corn growth. Soil Sci. 176, 136–142. doi: 10.1097/SS.0b013e31820e4063
Jami, N., Rahimi, A., Naghizadeh, M., and Sedaghati, E. (2021). Investigating the use of different levels of mycorrhiza and vermicompost on quantitative and qualitative yield of saffron (Crocus sativus L.). Sci. Hortic. 262, 1–12. doi: 10.1016/j.scienta.2019.109027
Jones, J. A., Wolf, J. B., and Mills, H. A. (1991). Plant Analysis: A Practical Sampling, Preparation, Analysis, and Interpretation Guide. Athents, Georgia: Micro and Macro Publishing Inc., 453 p.
Karami, Z., Sharifi, Z., and Renella, G. (2023). Determining the most sensitive indicators of soil carbon transformation to the land use change from intact rangeland to cropland, western Iran. Int J Environ Res. 17:37, 1–13. doi: 10.1007/s41742-023-00527-9
Kumar, A., Pramanick, B., Mahapatra, B. S., Singh, S. P., and Shukla, D. K. (2019). Growth, yield and quality improvement of flax (Linum usitattisimum L.) grown under tarai region of Uttarakhand, India through integrated nutrient management practices. Ind. Crop. Prod. 140, 1–6. doi: 10.1016/j.indcrop.2019.111710
Kumar, S., Rawat, C. R., Dhar, S., and Rai, S. K. (2005). Dry- matter accumulation, nutrient uptake and changes in soil fertility status an influenced by different organic source of nutrients to forage sorghum (Sorghum bicolor). India J. Agric. Sci. 75, 340–342.
Leal, F., Rodrigues, A. J., Fernandes, D., Cipriano, J., Ramos, J., Teixeira, S., et al. (2009). In vitro multiplication of Calendula arvensis for secondary metabolites. Acta hortic. 812, 251–256. doi: 10.17660/ActaHortic.81233
Leggo, P. (2000). An investigation of plant growth in an organo-zeolitic substrate and its ecological significance. Plant Soil. 219, 135–46. doi: 10.1023/A:1004744612234
Liu, M., Wang, C., Wang, F., and Xie, Y. (2019). Maize (Zea mays) growth and nutrient uptake following integrated improvement of vermicompost and humic acid fertilizer on coastal saline soil. Appl. Soil Ecol. 142, 147–154. doi: 10.1016/j.apsoil.04024
Majdalawieh, A. F., Hmaidan, R., and Carr, R. I. (2010). Nigella sativa modulates splenocyte proliferation, Th1/Th2 cytokine profile, macrophage function and NK anti-tumor activity. J. Ethnopharmacol. 131, 268–275. doi: 10.1016/j.jep.06030
Metcalf, L. C., Schmitz, A. A., and Pelka, J. R. (1966). Rapid preparation of fatty acid esters from lipids for gas chromatographic analysis. Anal. Chem. 38, 514–515. doi: 10.1021/ac60235a044
Nagavallemma, K. P., Wani, S. P., Stephane, L., Padmaja, V. V., Vineela, C., Babu Rao, M., et al. (2006). Vermicomposting: Recycling wastes into valuable organic fertilizer. Global Theme on Agrecosystems Report no. 8. Patancheru 502 324, Andhra Pradesh, India. International Crops Research Institute for the Semi-Arid Tropics. 20 pp.
Nardi, S., Pizzeghello, D., Schiavon, M., and Ertani, A. (2016). Plant biostimulants: physiological responses induced by protein hydrolyzed-based products and humic substances in plant metabolism. Sci. Agric. 73, 18–23. doi: 10.1590/0103-9016-2015-0006
Ozer, H., Coban, F., Sahin, U., and Ors, S. (2020). Response of black cumin (Nigella sativa L.) to deficit irrigation in a semi-arid region: growth, yield, quality, and water productivity. Ind. Crops Prod. 144, 1–10. doi: 10.1016/j.indcrop.2019.112048
Powlson, D. S., Addiscott, T. M., Benjamin, N., Cassman, K. G., Kok, d. e., Van Grinsven, K. M. H., et al. (2008). When does nitrate become a risk for humans? J. Environ. Qual. 37, 291–295. doi: 10.2134/jeq2007.0177
Randhawa, M. A., and Alghamdi, M. S. (2011). Anticancer activity of Nigell sativa (black seed)—A review. Am. J. Chin. Med. 39, 1075–1091. doi: 10.1142/S0192415X1100941X
Rekha, G. S., Valivittan, K., and Kaleena, P. K. (2013). Studies on the influence of vermicompost and vermivash on the growth and productivity of black gram (Vigna mungo). Adv. Biol. Res. 7, 114–121. doi: 10.5829/idosi.abr.2013.7.4.73217
Rezaei-Chiyaneh, E., Amirnia, R., Amani Machiani, M., Javanmard, A., Maggi, F., Morshedloo, M. R., et al. (2020). Intercropping fennel (Foeniculum vulgare L.) with common bean (Phaseolus vulgaris L.) as affected by PGPR inoculation: a strategy for improving yield, essential oil and fatty acid composition. J. Clean Prod. 261, 1–11. doi: 10.1016/j.scienta.2019.108951
Rezaei-Chiyaneh, E., Seyyedi, S. M., Ebrahimian, E., Siavash Moghadam, S., and Damalas, C. A. (2018). Exogenous application of gamma-aminobutyric acid (GABA) alleviates the effect of water deficit stress in black cumin (Nigella sativa L.). Ind. Crop Prod. 112, 741–748. doi: 10.1016/j.indcrop.2017.12.067
Roy, D. K., and Singh, B. P. (2006). Effect of level and time of nitrogen application with and without vermicompost on yield, yield attributes and quality of malt barley (Hordeum vulgare). Indian J. Agron. 51, 40–42.
SailajaKumari, M. S., and Ushakumari, K. (2002). Effect of vermicompost enriched with rock phosphate on the yield and uptake of nutrients incowpea (Vigna unguiculata L. Walp). J. Trop. Agric. 40, 27–30.
Sailo, G. L., and Bagyaraj, D. J. (2005). Influence of different AM-fungi on the growth, nutrition and forskolin content of Coleus forskohili. Mycol. Res. 109, 795–798. doi: 10.1017/s0953756205002832
Schroder, D., and Kopke, U. (2012). Faba bean (Vicia faba L.) intercropped with oil crops—A strategy to enhance rooting density and to optimize nitrogen use and grain producution? Field. Crops Res. 135, 74–81. doi: 10.1016/j.fcr.07007
Seyyedi, S. M., RezvaniMoghaddam, P., Khajeh-Hosseini, M., and Shahandeh, H. (2015). Influence of phosphorus and soil amendments on black seed (Nigella sativa L.) oil yield and nutrient uptake. Ind.Crops Prod. 77, 167–174. doi: 10.1016/j.indcrop.08065
Shahbazi, E. (2019). Genotype selection and stability analysis for seed yield of Nigella sativa using parametric and non-parametric statistics. Sci. Hortic. 253, 172–179. doi: 10.1016/j.scienta.04047
Treacy, M. M. J., and Higgins, J. B. (2007). Collection of Simulated XRD Powder Patterns for Zeolites. Amsterdam: Elsevier.
Tsintskaladze, G., Eprikashvili, L., Urushadze, T., Kordzakhia, T., Sharashenidze, T., Zautashvili, M., et al. (2016). Nanomodified natural zeolite as a fertilizer of prolonged activity. Ann. Agrar. Sci. 14, 163–168. doi: 10.1016/j.aasci.05013
Vafadar-Yengeje, L., Amini, R., and Nasab, A. D. M. (2019). Chemical compositions and yield of essential oil of Moldavian balm (Dracocephalum moldavica L.) in intercropping with faba bean (Vicia faba L.) under different fertilizers application. J. Clean Prod. 239, 1–10. doi: 10.1016/j.jclepro.2019.118033
Wang, X., Zhao, F., Zhang, G., Zhang, Y., and Yang, L. (2017). Vermicompost improves tomato yield and quality and the biochemical properties of soils with different tomato planting history in a greenhouse study. Front. Plant. Sci. 8, 1978. doi: 10.3389/fpls.2017.01978
Xu, L., Yan, D., Ren, X., Wei, Y., Zhou, J., Zhao, H., et al. (2016). Vermicompost improves the physiological and biochemical responses of blessed thistle (Silybum marianum Gaertn.) and peppermint (Mentha haplocalyx Briq) to salinity stress. Ind. Crop Prod. 94, 574–585. doi: 10.1016/j.indcrop.09023
Yang, J., Liu, X., and Shi, Y. (2013). Effect of different mixed fertilizer on yield, quality, and economic benefits in Stevia rebaudina Bertony. Adv. J. Food. Sci. Technol. 5, 588–591. doi: 10.19026/ajfst.5.3132
Zaghloul, R. A., Mohamed, Y. F, Rasha, Y., and El-Meihy, M. (2016). Influential cooperation between zeolite and PGPR on yield and antimicrobial activity of thyme essential oil. Int. J. Plant Soil Sci. 13, 1–18. doi: 10.9734/IJPSS/2016/28688
Zahedi, H., Noor-Mohamadi, G. H., Shirani-Rad, A. H., Habibi, D., and Mashhadi-Akbar-Boojar, M. (2009). The effects of zeolite and foliar applications of selenium on growth yield and yield components of three canola cultivars under drought stress. World Appl. Sci. J. 7, 255–262.
Zambon, I., Colantoni, A., Carlucci, M., Morrow, N., Sateriano, A., Salvati, L., et al. (2017). Land quality, sustainable development and environmental degradation in agricultural districts:a computational approach based on entropy indexes. Environ. Impact. Assess. Rev. 64, 37–46. doi: 10.1016/j.eiar.01003
Keywords: black cumin, organic supplementation, fertilizer, productivity, sustainable intensification
Citation: Rahmani R, Khalesro S, Heidari G and Mokhatssi-Bidgoli A (2023) Vermicompost and zeolite improve yield, nutrient uptake, essential and fixed oil production, and composition of Nigella sativa L.. Front. Sustain. Food Syst. 7:1214691. doi: 10.3389/fsufs.2023.1214691
Received: 30 April 2023; Accepted: 03 August 2023;
Published: 24 August 2023.
Edited by:
Adnan Mustafa, Brno University of Technology, CzechiaReviewed by:
Juan Fernando Hirzel, Agricultural Research Institute, ChileMohamed Anli, Université des Comores, Comoros
Copyright © 2023 Rahmani, Khalesro, Heidari and Mokhatssi-Bidgoli. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Shiva Khalesro, c2gua2hhbGVzcm9AdW9rLmFjLmly