 Eva Chatonnat
Eva Chatonnat Mariya Julien1
Mariya Julien1 Eric Jubinville
Eric Jubinville Nicole Pavio
Nicole Pavio Julie Jean
Julie Jean- 1Institute of Nutrition and Functional Food (INAF), Université Laval, Québec City, QC, Canada
- 2French Agency for Food, Environmental and Occupational Health and Safety (ANSES), Maisons-Alfort, France
The Hepatitis E virus is widespread throughout the world and causes sporadic cases of liver disease. In industrialized countries, its transmission is zoonotic, with pigs, deer, and wild boar being major reservoirs. Humans can be infected by ingesting raw or undercooked contaminated meat products. In the Canadian province of Quebec, HEV has been detected previously in pork liver and chops. In the present study, we analyzed 83 pork liver pâtés sold in local grocery stores and 79 raw pork livers obtained from Quebec hog processors. HEV RNA was extracted and detected by RT-qPCR. HEV RNA was amplified in 29% of the pâtés and 4% of the raw pork livers. Out of all amplified PCR-positive samples, only one showed a product during electrophoresis and was sent for sequencing. The resulting sequence matched a strain belonging to genotype 3 (HEV-3). This pilot study shows that HEV is present frequently in pork liver-based products in the province of Quebec.
1. Introduction
The hepatitis E virus (HEV) is a single-stranded positive RNA virus that causes hepatitis (Balayan et al., 1983). It was recognized as a new disease in the 1980s, and the genome was cloned and characterized in 1991 (Tam et al., 1991; Khuroo, 2011). Since then, HEV has become endemic in developing countries due to a fecal–oral transmission facilitating rapid spread wherever sanitation systems are deficient (Khuroo et al., 2016). HEV is now regarded as an emerging foodborne virus (Harrison and DiCaprio, 2018), and sporadic cases of hepatitis E, not related to travel in endemic countries, are increasingly reported in industrialized countries. Many of these autochthonous cases are associated with zoonotic transmission (Lewis et al., 2010) after the consumption of raw or undercooked food products from infected animals. The WHO estimates the number of HEV infections at 20 million per year worldwide and the number of associated deaths at 44,000 (World Health Organization, 2022). Although most HEV infections are asymptomatic, severe forms such as chronic hepatitis, fulminant hepatitis, and neurological illness may develop in susceptible populations (Webb and Dalton, 2019). HEV is particularly dangerous to pregnant women, with up to 25% mortality during the third trimester of pregnancy in some endemic regions (Navaneethan et al., 2008).
The hepatitis E virus (family Hepeviridae, subfamily: Orthohepevirinae, genus: Paslahepevirus) has eight known genotypes. Among these, four can infect humans (HEV-1 to HEV-4), HEV-1 and HEV-2 infect only humans, and HEV-3 and HEV-4 are transmitted back and forth between humans and animals (Smith et al., 2014). Swine are natural hosts and an important reservoir of HEV (Meng et al., 1997; Li et al., 2022), but the virus is also detected in wild animals, for example, deer, wild boar, and hare (Pavio et al., 2010; Smith et al., 2014). In zoonotic transmissions, humans become infected mainly by eating raw or undercooked meat (Raji et al., 2022).
HEV is liver-cell-tropic, and the food products most often involved in transmission are charcuteries or prepared meals containing a large proportion of pork liver. Since 2010, numerous studies on the prevalence of HEV in pork liver products have been conducted, especially in Europe. The prevalence of HEV ranges from 5 to 70% depending on the type of product and the geographical region (Pavio et al., 2014; Boxman et al., 2019; Pallerla et al., 2021). In Canada, 47% of pork liver pâtés and 10.5% of raw pork livers sampled in Ottawa, the Canadian federal capital, were found positive for HEV RNA (Mykytczuk et al., 2017). To date, no study has focused on the presence of HEV in food in the province of Quebec. Furthermore, cases of patients who developed hepatitis after liver transplantation have been reported in the province and a foodborne origin has been suspected. This hypothesis was supported by the close genomic similarity of the viral strain found in the patients with a strain isolated from swine raised in Quebec (Halac et al., 2012).
We report here the results of a pilot study on the prevalence of HEV RNA in pork liver pâtés sampled in retail stores in Quebec City (QC, Canada) and raw pork liver provided by a local hog processing operation.
2. Methods and materials
2.1. Sampling of pâtés sold commercially
Pork liver pâtés or country-style pâtés were purchased during the summer of 2022 (June 15 to August 17) at 10 grocery stores in Quebec City (QC, Canada). All 83 pâtés were from different lots and contained pork liver as one of the first three ingredients listed. Samples were aliquoted (2.0 g ± 0.1 g) and stored at −30°C until processing.
2.2. Sampling of raw pork liver
Whole raw pork livers were provided by a local business that collected them from hog processing plants throughout the province of Quebec. All 79 were from pigs aged 6 to 7 months and weighing on average 112 kg. Samples were aliquoted (3.0 g ± 0.1 g) and then stored at −30°C until processing.
2.3. Sample processing positive controls
Stocks of purified Mengo virus strain vMC0 and murine norovirus 1 (MNV-1) strain ATCC VR-1937 already in our collection were used as positive controls for sample processing. Viral titers were estimated by RT-qPCR and were 2.16 × 106 genome copies/μl for the Mengo virus and 1 × 106 genome copies/μl for MNV-1. The viral stock suspensions were stored at −80°C until processing.
2.4. Extraction of virus from pork liver pâté
A method of extracting HEV from sausages (Szabo et al., 2015) was adapted to pâté. Briefly, 2.0 g ± 0.1 g of sample was blended in a 50 ml plastic centrifugation tube with 10 μl of Mengo virus suspension and 7 ml of TRI Reagent Solution® (Thermo Fisher Scientific, Waltham, MA, USA) by vortex mixing at maximum speed for 2 min. The resulting mixture was centrifuged at 10,000 × g for 20 min (at 4°C), and the supernatant (below a thin layer of liquid fat) was diluted 1:10 with sterile water and vortexed for 15 s. Nucleic acid was extracted from 1 ml of this diluted supernatant. The negative control was obtained by repeating the protocol without pâté and with 10 μl of sterile water instead of Mengo virus suspension.
2.5. Extraction of virus from raw pork liver
Approximately 3.0 g ± 0.1 g of raw pork liver was homogenized with 10 μl of MNV-1 suspension, 7 ml of TRI Reagent Solution®, 2 μl of RNaseOUT™ (Invitrogen, Thermo Fisher Scientific, Waltham, MA, USA), and six glass beads (3 mm, VWR, Radnor, PA, USA) in a Bead Genie device (Scientific Industries, Bohemia, NY, USA) at maximum speed (4,800 strokes per minute) for 2 × 30 s. The homogenate was centrifuged at 10,000 × g for 20 min at 4°C, and the supernatant was diluted 1:10 with sterile water. The diluted supernatants were vortexed for 15 s and 1 ml was used for nucleic acid extraction. The negative control was obtained by repeating the protocol without the liver and with 10 μl of sterile water instead of MNV-1 suspension.
2.6. Nucleic acid extraction
All samples (pâté, liver, controls, 1 ml) were treated with 2 ml of NucliSENS® lysis buffer (bioMérieux, Marcy-l'Étoile, France) at room temperature for 10 min, then centrifuged at 1,800 × g for 2 min at 4°C. The semi-automated eGENE-UP® system (bioMérieux, Marcy-l'Étoile, France) was used according to the manufacturer's instructions to extract RNA. The extracts of 100 μl thus obtained were diluted 1:10 with NucliSENS® buffer 3 (bioMérieux, Marcy-l'Étoile, France) and stored at −80°C until analysis.
2.7. Detection of HEV by RT-qPCR
HEV, Mengo virus, and MNV-1 were detected using a method described previously (Trudel-Ferland et al., 2021). Genomes were amplified using an iTaq Universal Probe One-step kit (Bio-Rad, Hercules, CA, USA). The primers and probes used are listed in Table 1. The expected size of the HEV amplicon is 72 pb. To quantify HEV, a standard curve was produced in triplicate from 105 genome copies/μl to 101 genome copies/μl. The MiniGene plasmid pIDTSMART-AMP+ generated by IDT (Integrated DNA Technologies, Coralville, IA, USA) was used (GenBank number MP690161.1). RT-qPCR reactions were performed in 96-well clear PCR microplates (Applied Biosystems, Thermo Fisher Scientific, Waltham, MA, USA) sealed with an optical adhesive film (Applied Biosystems, USA). The ABI7500 real-time PCR thermal cycler (Applied Biosystems, Thermo Fisher Scientific, Waltham, MA, USA) was used as follows: 50°C for 10 min, 95°C for 3 min and 45 cycles of 95°C for 15 s, and 60°C for 30 s. ROX dye was used as the reference dye. Each sample was analyzed in duplicate. All RT-qPCR runs included a negative control (RNase-free water, VWR, Radnor, PA, USA). All qPCR standard curves had an R2 over 0.985 and efficiency between 90 and 110%. Viral RNA recovery was calculated for all samples using the following formula:
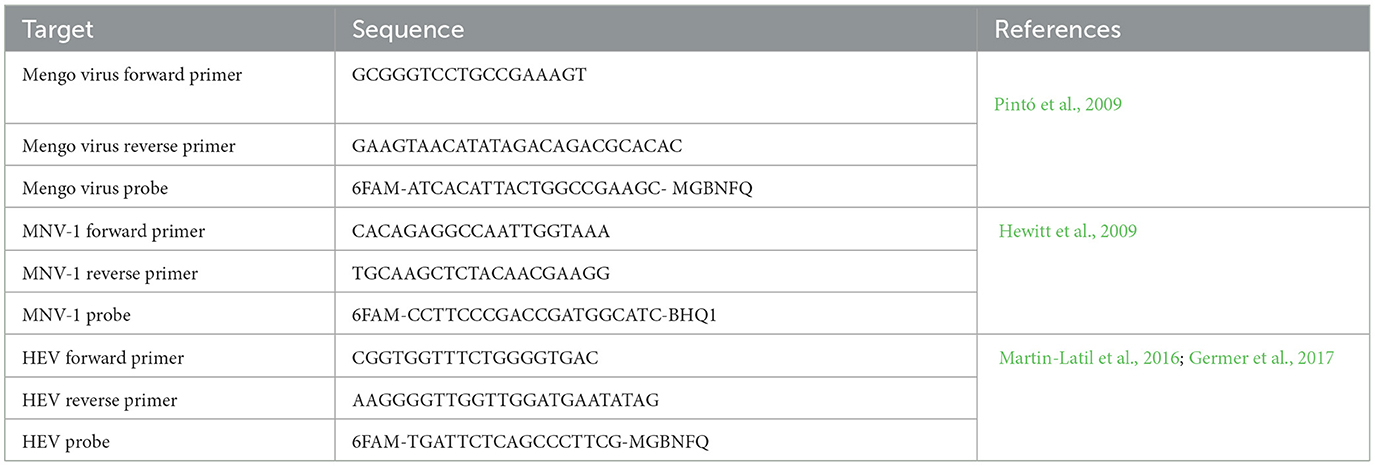
Table 1. Primers, probe sequences, quencher, and dye used for the detection of the virus by RT-qPCR.
where [RNA] is expressed in genome copies per sample.
All results were analyzed using SDS Software v1.5.1 (Applied Biosystems, Thermo Fisher Scientific, Waltham, MA, USA).
2.8. Amplification by conventional RT-PCR and sequencing of HEV
For each positive RT-qPCR sample, a conventional RT-PCR was performed using the same RNA used for RT-qPCR.
Briefly, RNA was retrotranscribed into cDNA as follows: 10 μl of RNA extract, 4 μl of 5X iScript reaction mix containing, 1 μl of reverse transcriptase, and 5 μl of PCR water (iScript cDNA synthesis kit, BioRad, Hercules, CA, USA) were mixed and thermo-cycled at 25°C for 5 min, 46°C for 20 min, and 95°C for 1 min in an Eppendorf® Mastercycler gradient device (Millipore Sigma, Darmstadt, Germany).
Conventional PCR was performed under the recommended conditions as follows: 5 μl of cDNA, 10 μl of iQ Supermix, 1 μl of forward primer (10 μM), 1 μl of reverse primer (10 μM), and 3 μl of PCR Water (iQ Supermix, Bio-Rad, Hercules, CA, USA) were mixed and amplified under the following conditions: 95°C for 5 min, 45 cycles of 95°C for 30 s, 55°C for 30 s, and 72°C for 1 min, and 72°C for 10 min. Specific primer sequences targeting the HEV ORF2 region were used for the amplification: forward primer GTYATGYTYTGCATACATGGCT and reverse primer AGCCGACGAAATYAATTCTGTC (Mykytczuk et al., 2017). The amplicons with an expected size of 348 bp were stored at −80 °C until processing.
An agarose gel (2%) in tris acetate EDTA buffer 1 × with 0.01% SYBR® Safe (Invitrogen, Thermo Fisher Scientific, Waltham, MA, USA) was used. Each well was loaded with 3 μl of sample and 2 μl of TriTrack DNA loading dye (Thermo Scientific, Thermo Fisher Scientific, Waltham, MA, USA). The amplicon size was compared to a 100 bp DNA ladder (Thermo Scientific, Thermo Fisher Scientific, Waltham, MA, USA) after 45 min of migration at 120 V. A UV-light photo of the gel was taken with the ChemiDocTM MP imaging system (Bio-Rad, Hercules, CA, USA) and processed using Quantity One® software (Bio-Rad, Hercules, CA, USA).
PCR products were Sanger-sequenced after the gel purification at the IBIS Genomic Analysis Platform, Université Laval (Quebec City, QC, Canada) following their recommendations. The Chromas software v2.6.6 was used, and the obtained sequences were aligned using BioEdit v7.0.5.3. The consensus sequence was analyzed with BLAST.
3. Results
3.1. Prevalence of HEV RNA
Of the 83 samples of pork liver pâté, 24 tested positive for HEV RNA (Table 2), indicating a prevalence of 29% with genome copy quantities per gram ranging from 3.34 × 102 to 1.84 × 107 (Table 3). A total of three samples of country-style pâté (P21, P22, and P23) contained conspicuously larger amounts of HEV RNA, and eight samples were withdrawn from the study because the control virus could not be detected. Most of these were of the same brand and one of two products. Of the 79 raw pork livers tested, three were positive for HEV (Tables 2, 3). The estimated prevalence of HEV in raw pork liver produced in the province of Quebec is therefore 4%.
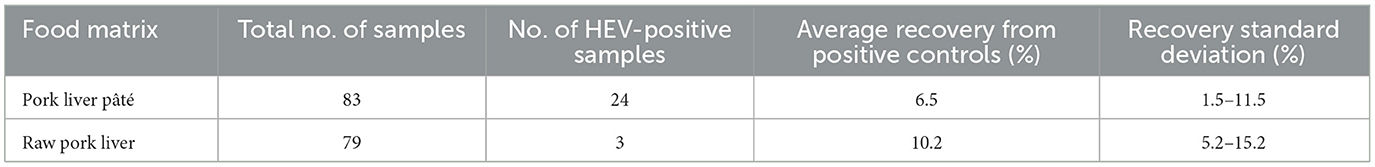
Table 2. Detection of HEV in pork liver pâtés collected from grocery stores and in raw livers produced in the province of Quebec.
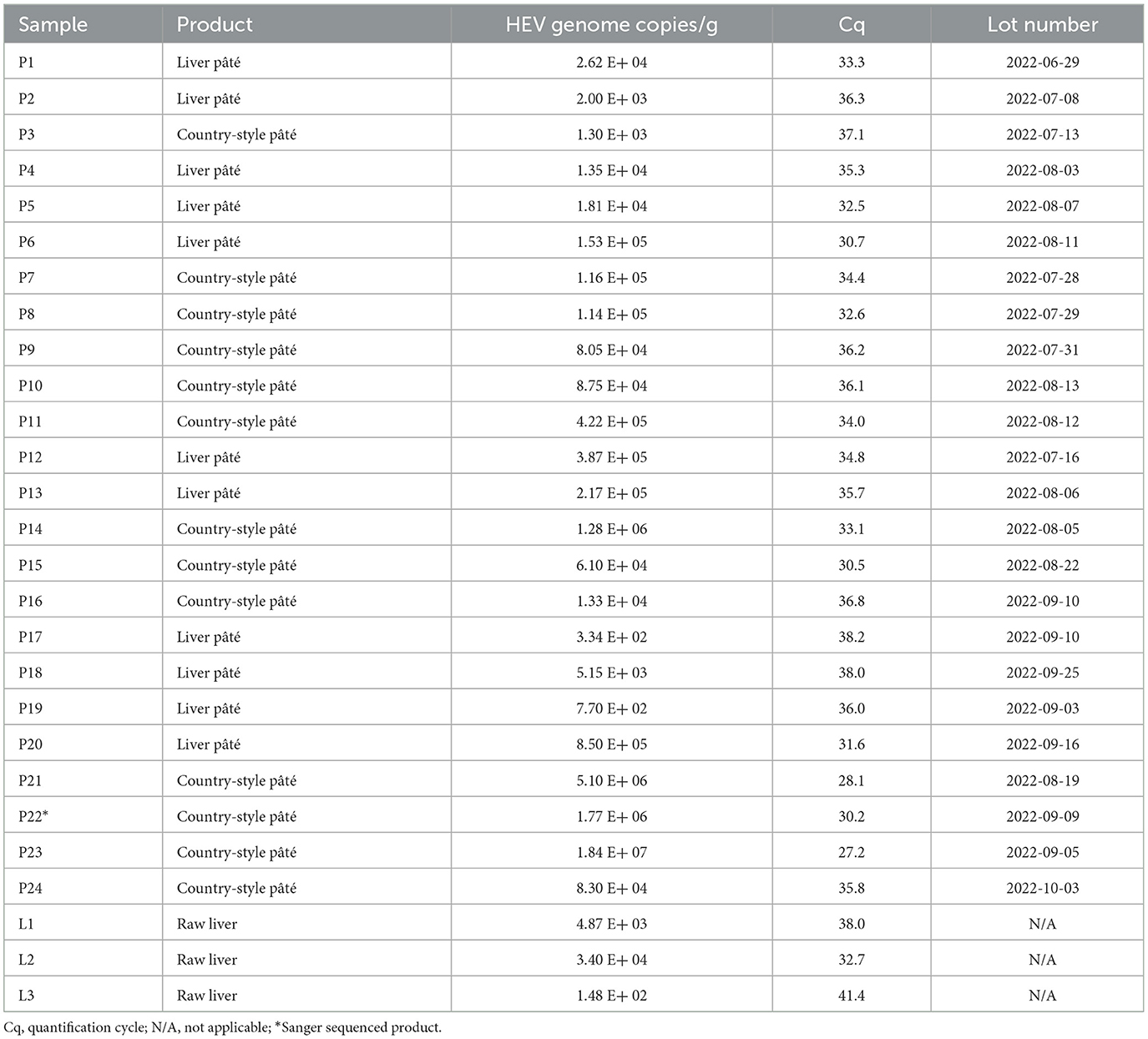
Table 3. HEV RNA detection in samples of pork liver pâté, country-style pâté, or raw liver.
3.2. HEV sequence analysis
A large proportion of the samples found positive by RT-qPCR then transcribed into cDNA and amplified did not show up as positive on an electrophoresis gel. Only one sample showed a PCR product of the expected size (348 nt) and was sent for Sanger sequencing (P22* in Table 3). The sequence was found to match HEV-3 from that in Tokyo, Japan with a sequence similarity of 95% (accession number: LC699232.1). The sequence of the HEV detected in our study appears to be HEV-3 and has been deposited in GenBank (accession number: OQ032868).
4. Discussion
No public data are currently available regarding the number of hepatitis E cases per year in Canada. However, the seroprevalence of HEV in Canadian blood donors is ~6% (Fearon et al., 2017). Moreover, the virus may circulate among animals and humans that could be infected by ingesting contaminated food products. Food safety is a major public health concern and foodborne viruses, including HEV, are not sufficiently traced in food products. In the present study, we investigated the occurrence of HEV in raw pork liver and pork liver pâté produced in the Canadian province of Quebec. We focused on pork products because swine are known hosts of HEV and because Quebec is the leading pork-producing province, accounting for 30% of Canadian production. The situation of HEV in pork needs to be monitored, and the most recent data for Quebec are now 9 years old. Pork (fresh and frozen) is also the third most consumed meat in the province of Quebec although consumption has been decreasing slightly since 2016. Quebec is the largest exporter of meat and pork products, with a stable share of nearly 40% of the value of total Canadian exports (Ministère de l'Agriculture, des Pêcheries et de l'Alimentation du Québec, 2022). Wildlife may also carry HEV (Weger et al., 2017; Lorusso et al., 2022; Takahashi et al., 2022) and sausages or terrines made from deer or others are sold in grocery stores and therefore should be monitored as well. In Canadian pig herds, HEV seroprevalence has been measured at 59.4% and in Quebec at 88.8% (Yoo et al., 2001). Previous prevalence studies conducted in other provinces have concluded similar or higher levels. Between 2011 and 2012, 25 of 283 livers tested (~9%) were reportedly positive or suspected positive for HEV contamination, mainly in the provinces of Ontario and Quebec (Wilhelm et al., 2014). In 2017, Health Canada declared an HEV prevalence of 47% in pork liver pâtés and 10.5% in raw pork livers in the province of Ontario (Mykytczuk et al., 2017). In the USA, the most significant study was conducted in California, which found HEV in 45% of pork livers and 12.6% of ground pork (Harrison et al., 2021). This is also one of the few studies to have found the virus in ground pork.
In the present study, the sampling of pork liver pâtés sold in grocery stores in Quebec City (QC, Canada) included different brands, recipes, and production lots to obtain as much sample diversity as possible. HEV RNA was detected in 29% of the pâtés and in 4% of the raw pork livers. As found in other studies, our results show that liver-based products are more likely to be tested positive for HEV than raw livers (Mykytczuk et al., 2017; Boxman et al., 2019; Pallerla et al., 2021). This is not surprising since many livers (30 to 35) are used to make a large mix that will be then divided into numerous portions of pâté. Our findings are broadly consistent with those in other countries around the world, particularly in Europe where most of the prevalence studies have been conducted. This suggests that sanitation and/or hog processing procedures in Canada and these other countries have a similar impact on the presence of HEV in pork products.
Only the presence of HEV RNA was demonstrated in the present study, which does not provide any indication of the potentially infectious status of virions in the sample. Nevertheless, a link has been previously established between the consumption of pork products and the transmission of HEV to humans (Yapa et al., 2016). The oral infectious dose of HEV is unknown in humans, but it has been estimated experimentally in pigs, where 105 HEV RNA copies are sufficient to induce a productive infection (Andraud et al., 2013). In this study, the quantity of HEV RNA found in the products reached up to 1.84 × 107 copies of HEV RNA per gram which is very high and 11 of the positive samples contain more than 105 copies of genome per gram. It is therefore possible that these HEV quantities are sufficient to induce an infection.
From the methodological point of view, as far as we have been able to determine, there is still (as of this writing) no standardized method, ISO, or other forms, for processing and analyzing complex samples containing pork liver or pork liver-based products for the presence of HEV. Many protocols are described in the literature and sometimes include similar steps, such as using Trizol®, to dissolve tissues and cells or chloroform to clarify the resulting solutions (Hennechart-Collette et al., 2019; Pallerla et al., 2021; Wang et al., 2021). This methodological diversity creates numerous possibilities for inconsistencies in findings and raises questions about the efficiency of HEV extraction from these food matrices. A straightforward method involving chemical and mechanical homogenization of the samples and a dilution step was performed in this study. The use of the Mengo virus or MNV-1 as control viruses in each method allows us to confirm the efficiency of the protocol. In the absence of a reliable cell culture model, detecting RNA is the only widely available way to confirm that a food sample contains HEV. Since pork-based recipes are the foods most likely to carry a risk of HEV transmission to humans, a standardized method should be developed for this product category. An alternative method for the molecular detection of HEV RNA, based on the integrity of viral capsid, should also be developed to evaluate HEV's possible infectivity (Fraisse et al., 2018).
The HEV RNA sequenced in this study matches genotype 3, frequently associated with HEV zoonotic transmission in industrialized countries (Treagus et al., 2021) and more particularly in North America, Europe, and Japan. In addition, only 1 of 24 positive samples yielded a PCR product that showed up clearly on the electrophoresis gel. HEV genomes could be fragmented during the viral recovery using Trizol®. Our results emphasize the need to develop a standardized method, especially regarding the viral recovery and sequencing methods of HEV in meat products.
Finally, this pilot study provides a glimpse of the potential for contamination with HEV of certain pork liver foods sold commercially in the province of Quebec. It also suggests that monitoring more samples over longer periods is warranted since this virus is now considered an emerging pathogen with potentially severe consequences for growing susceptible populations, such as immunocompromised patients (Damiris et al., 2022) and pregnant women and fetuses (Navaneethan et al., 2008). This research also highlights the continuous presence of HEV in pork liver-based products in Canada over the past 8 years, reinforcing the need for regular surveillance of its prevalence in food and hence, its epidemiology.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found at: https://www.ncbi.nlm.nih.gov/nuccore/OQ032868.1/.
Author contributions
EC, EJ, VG-B, and JJ: conceptualization. EC and MJ: methodology. EJ, VG-B, and JJ: validation, conceptualization, supervision, and project administration. EC: formal analysis, investigation, resources, and preparation and writing of the original draft. JJ: data curation and funding acquisition. EC, EJ, VG-B, NP, and JJ: writing, review, and editing. All authors contributed to the article and approved the submitted version.
Funding
This research was part of the VIROCONTROL research chair funded by the Ministère de l'Agriculture, des Pêcheries et de l'Alimentation (MAPAQ, PPIA08) and the Natural Sciences and Engineering Research Council of Canada (NSERC, RDCPJ 538872-19).
Acknowledgments
The authors would like to thank the company Olymel for providing the raw pork liver samples used in this study and Stephen Davids for proofreading the manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Andraud, M., Dumarest, M., Cariolet, R., Aylaj, B., Barnaud, E., Eono, F., et al. (2013). Direct contact and environmental contaminations are responsible for HEV transmission in pigs. Vet. Res. 44, 102. doi: 10.1186/1297-9716-44-102
Balayan, M. S., Andjaparidze, A. G., Savinskaya, S. S., Ketiladze, E. S., Braginsky, D. M., Savinov, A. P., et al. (1983). Evidence for a virus in non-A, non-B hepatitis transmitted via the fecal-oral route. Intervirology. 20, 23–31. doi: 10.1159/000149370
Boxman, I. L. A., Jansen, C. C. C., Hägele, G., Zwartkruis-Nahuis, A., Tijsma, A. S. L., Vennema, H., et al. (2019). Monitoring of pork liver and meat products on the Dutch market for the presence of HEV RNA. Int. J. Food Microbiol. 296, 58–64. doi: 10.1016/j.ijfoodmicro.2019.02.018
Damiris, K., Aghaie Meybodi, M., Niazi, M., and Pyrsopoulos, N. (2022). Hepatitis E in immunocompromised individuals. WJH. 14, 482–94. doi: 10.4254/wjh.v14.i3.482
Fearon, M. A., O'Brien, S. F., Delage, G., Scalia, V., Bernier, F., Bigham, M., et al. (2017). Hepatitis E in Canadian blood donors. Transfusion. 57, 1420–5. doi: 10.1111/trf.14089
Fraisse, A., Niveau, F., Hennechart-Collette, C., Coudray-Meunier, C., Martin-Latil, S., Perelle, S., et al. (2018). Discrimination of infectious and heat-treated norovirus by combining platinum compounds and real-time RT-PCR. Int. J. Food Microbiol. 269, 64–74. doi: 10.1016/j.ijfoodmicro.2018.01.015
Germer, J. J., Ankoudinova, I., Belousov, Y. S., Mahoney, W., Dong, C., Meng, J., et al. (2017). Hepatitis E virus (HEV) detection and quantification by a real-time reverse transcription-PCR assay calibrated to the world health organization standard for HEV RNA. Clin Microbiol. 55, 1478–87. doi: 10.1128/JCM.02334-16
Halac, U., Béland, K., Lapierre, P., Patey, N., Ward, P., Brassard, J., et al. (2012). Chronic hepatitis E infection in children with liver transplantation. Gut. 61, 597–603. doi: 10.1136/gutjnl-2011-300708
Harrison, L., and DiCaprio, E. (2018). Hepatitis E virus: an emerging foodborne pathogen. Front. Sust. Food Syst. 2, 14. doi: 10.3389/fsufs.2018.00014
Harrison, L., Ramos, T. D. M., Wu, X., and DiCaprio, E. (2021). Presence of hepatitis E virus in commercially available pork products. Int. J. Food Microbiol. 339, 109033. doi: 10.1016/j.ijfoodmicro.2020.109033
Hennechart-Collette, C., Fraisse, A., Guillier, L., Perelle, S., and Martin-latil, S. (2019). Evaluation of methods for elution of HEV particles in naturally contaminated sausage, figatellu and pig liver. Food Microbiol. 84, 103235. doi: 10.1016/j.fm.2019.05.019
Hewitt, J., Rivera-Aban, M., and Greening, G. E. (2009). Evaluation of murine norovirus as a surrogate for human norovirus and hepatitis A virus in heat inactivation studies. J. Appl. Microbiol. 107, 65–71. doi: 10.1111/j.1365-2672.2009.04179.x
Khuroo, M. S. (2011). Discovery of hepatitis E: the epidemic non-A, non-B hepatitis 30 years down the memory lane. Virus Res. 161, 3–14. doi: 10.1016/j.virusres.2011.02.007
Khuroo, M. S., Khuroo, M. S., and Khuroo, N. S. (2016). Transmission of hepatitis E virus in developing countries. Viruses. 22, 8. doi: 10.3390/v8090253
Lewis, H. C., Wichmann, O., and Duizer, E. (2010). Transmission routes and risk factors for autochthonous hepatitis E virus infection in Europe: a systematic review. Epidemiol Infect. 138, 145–66. doi: 10.1017/S0950268809990847
Li, P., Ji, Y., Li, Y., Ma, Z., and Pan, Q. (2022). Estimating the global prevalence of hepatitis E virus in swine and pork products. One Health. 14, 100362. doi: 10.1016/j.onehlt.2021.100362
Lorusso, P., Bonerba, E., Pandiscia, A., Mottola, A., Di Pinto, A., Piredda, R., et al. (2022). Occurrence of hepatitis E virus (HEV) in Calabrian wild boars. Int. J. Food Microbiol. 371, 109671. doi: 10.1016/j.ijfoodmicro.2022.109671
Martin-Latil, S., Hennechart-Collette, C., Delannoy, S., Guillier, L., Fach, P., Perelle, S., et al. (2016). Quantification of hepatitis E virus in naturally-contaminated pig liver products. Front Microbiol. 4, 83. doi: 10.3389/fmicb.2016.01183
Meng, X. J., Purcell, R. H., Halbur, P. G., Lehman, J. R., Webb, D. M., Tsareva, T. S., et al. (1997). A novel virus in swine is closely related to the human hepatitis E virus. Proc Natl Acad Sci USA. 94, 9860–5. doi: 10.1073/pnas.94.18.9860
Ministère de l'Agriculture des Pêcheries et de l'Alimentation du Québec (MAPAQ), ED_Portrait_Diagnostic_Industrie_Porcine_MAPAQ.pdf. (2022). Available online at: https://cdn-contenu.quebec.ca/cdn-contenu/adm/min/agriculture-pecheries-alimentation/agriculture/types-de-productions/ED_portrait_diagnostic_industrie_porcine_MAPAQ.pdf?1661970644 (accessed December 5, 2023).
Mykytczuk, O., Harlow, J., Bidawid, S., Corneau, N., and Nasheri, N. (2017). Prevalence and molecular characterization of the hepatitis E virus in retail pork products marketed in Canada. Food Environ Virol. 9, 208–18. doi: 10.1007/s12560-017-9281-9
Navaneethan, U., Mohajer, M. A., and Shata, M. T. (2008). Hepatitis E and pregnancy- understanding the pathogenesis. Liver Int. 28, 1190–9. doi: 10.1111/j.1478-3231.2008.01840.x
Pallerla, S. R., Schembecker, S., Meyer, C. G., Linh, L. T. K., Johne, R., Wedemeyer, H., et al. (2021). Hepatitis E virus genome detection in commercial pork livers and pork meat products in Germany. J Viral Hepatitis. 28, 196–204. doi: 10.1111/jvh.13396
Pavio, N., Meng, X. J., and Renou, C. (2010). Zoonotic hepatitis E: animal reservoirs and emerging risks. Vet Res. 22, 41. doi: 10.1051/vetres/2010018
Pavio, N., Merbah, T., and Thébault, A. (2014). Frequent hepatitis E virus contamination in food containing raw pork liver, France. Emerg Infect Dis. 20, 1925–7. doi: 10.3201/eid2011.140891
Pintó, R. M., Costafreda, M. I., and Bosch, A. (2009). Risk assessment in shellfish-borne outbreaks of hepatitis A. Appl. Environ. Microbiol. 75, 7350–5. doi: 10.1128/AEM.01177-09
Raji, Y. E., Toung, O. P., Taib, N. M., and Sekawi, Z. B. (2022). Hepatitis E virus: an emerging enigmatic and underestimated pathogen. Saudi J. Biol. Sci. 29, 499–512. doi: 10.1016/j.sjbs.2021.09.003
Smith, D. B., Simmonds, P., Jameel, S., Emerson, S. U., Harrison, T. J., Meng, X. J., et al. (2014). Consensus proposals for classification of the family Hepeviridae. J Gen Virol. 95, 2223–32. doi: 10.1099/vir.0.068429-0
Szabo, K., Trojnar, E., Anheyer-Behmenburg, H., Binder, A., Schotte, U., Ellerbroek, L., et al. (2015). Detection of hepatitis E virus RNA in raw sausages and liver sausages from retail in Germany using an optimized method. Int. J. Food Microbiol. 215, 149–56. doi: 10.1016/j.ijfoodmicro.2015.09.013
Takahashi, M., Nishizono, A., Kawakami, M., Fukui, E., Isogai, E., Matsuoka, H., et al. (2022). Identification of hepatitis E virus in wild sika deer in Japan. Virus Res. 308, 198645. doi: 10.1016/j.virusres.2021.198645
Tam, A. W., Smith, M. M., Guerra, M. E., Huang, C. C., Bradley, D. W., Fry, K. E., et al. (1991). Hepatitis E virus (HEV): molecular cloning and sequencing of the full-length viral genome. Virology. 185, 120–31. doi: 10.1016/0042-6822(91)90760-9
Treagus, S., Wright, C., Baker-Austin, C., Longdon, B., and Lowther, J. (2021). The foodborne transmission of hepatitis E virus to humans. Food Environ. Virol. 13, 127–45. doi: 10.1007/s12560-021-09461-5
Trudel-Ferland, M., Jubinville, E., and Jean, J. (2021). Persistence of hepatitis A virus RNA in water, on non-porous surfaces, and on blueberries. Front Microbiol. 12:618352. doi: 10.3389/fmicb.2021.618352
Wang, J., Li, N., Zhang, H., Li, F., Fanning, S., and Jiang, T. (2021). Detection of hepatitis E virus in the pig livers and retail pork samples collected in selected cities in China. Foodborne Pathog. Dis. 18, 97–103. doi: 10.1089/fpd.2020.2837
Webb, G. W., and Dalton, H. R. (2019). Hepatitis E: an underestimated emerging threat. Ther. Adv. Inf. 6, 204993611983716. doi: 10.1177/2049936119837162
Weger, S., Elkin, B., Lindsay, R., Bollinger, T., Crichton, V., Andonov, A., et al. (2017). Hepatitis E virus seroprevalence in free-ranging deer in Canada. Transbound Emerg Dis. 64, 1008–11. doi: 10.1111/tbed.12462
Wilhelm, B., Leblanc, D., Houde, A., Brassard, J., Gagné, M. J., Plante, D., et al. (2014). Survey of Canadian retail pork chops and pork livers for detection of hepatitis E virus, norovirus, and rotavirus using real time RT-PCR. Int. J. Food Microbiol. 185, 33–40. doi: 10.1016/j.ijfoodmicro.2014.05.006
World Health Organization. (2022). Hepatitis E. Available online at: https://www.who.int/news-room/fact-sheets/detail/hepatitis-e (accessed December 01, 2022).
Yapa, C. M., Furlong, C., Rosewell, A., Ward, K. A., Adamson, S., Shadbolt, C., et al. (2016). First reported outbreak of locally acquired hepatitis E virus infection in Australia. Med. J. Australia. 204, 274–274. doi: 10.5694/mja15.00955
Keywords: epidemiology, hepatitis E virus, pork liver products, RT-qPCR, zoonotic transmission, foodborne viruses
Citation: Chatonnat E, Julien M, Jubinville E, Goulet-Beaulieu V, Pavio N and Jean J (2023) High occurrence of hepatitis E virus in raw pork liver and pork liver pâté produced in the Canadian province of Quebec. Front. Sustain. Food Syst. 7:1163507. doi: 10.3389/fsufs.2023.1163507
Received: 10 February 2023; Accepted: 21 March 2023;
Published: 26 April 2023.
Edited by:
María Belén Pisano, Universidad Nacional de Córdoba, ArgentinaReviewed by:
Ilaria Di Bartolo, National Institute of Health (ISS), ItalyMaria Guadalupe Vizoso Pinto, CONICET Higher Institute of Biological Research (INSIBIO), Argentina
Annamaria Pandiscia, Dipartimento di Medicina Veterinaria Bari, Italy
Copyright © 2023 Chatonnat, Julien, Jubinville, Goulet-Beaulieu, Pavio and Jean. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Julie Jean, anVsaWUuamVhbkBmc2FhLnVsYXZhbC5jYQ==