 Eduardo Henrique Marcandalli Boleta
Eduardo Henrique Marcandalli Boleta Fernando Shintate Galindo
Fernando Shintate Galindo Arshad Jalal
Arshad Jalal José Mateus Kondo SantiniWilliam Lima RodriguesBruno Horschut de LimaOrivaldo ArfMarcelo Rinaldi da SilvaSalatiér Buzetti
José Mateus Kondo SantiniWilliam Lima RodriguesBruno Horschut de LimaOrivaldo ArfMarcelo Rinaldi da SilvaSalatiér Buzetti Marcelo Carvalho Minhoto Teixeira Filho*
Marcelo Carvalho Minhoto Teixeira Filho*- Department of Rural Engineering, Plant Health and Soils (DEFERS), São Paulo State University, São Paulo, Brazil
Agricultural practices that allow a productive increase in a sustainable manner are becoming increasingly necessary to feed an ever-growing global population. The inoculation with Azospirillum brasilense has the potential to reduce the use of synthetic mineral fertilizers with efficient capacity to promote plant growth and increase nutrition. Therefore, this research was developed to investigate the potential use of A. brasilense to increase the accumulation of macro- and micronutrients and its influence on grain yield, plant height, and spike population in two wheat cultivars (CD1104 and CD150), under irrigated conditions in the Brazilian Cerrado. The study was carried out in a Rhodic Hapludox under a no-tillage system. The experiment was designed in randomized blocks with six replications, arranged in a 2 × 2 factorial scheme: two cultivars (CD150 and CD1104) and two levels of inoculation (control and with A. brasilense). The inoculation with A. brasilense provided greater accumulation of micronutrients in the aerial part of the wheat. In the cultivar CD1104, B and Cu had an accumulation 27.7 and 57.4% higher compared with those of the control without bacteria. In the cultivar CD150, Azospirillum increased the accumulation of B, Fe, and Mn by 43.8, 49.9, and 22%, respectively, and reduced Cu by 21.9%, compared with those of the control. The cultivar CD150 has greater efficiency to accumulate N (+35.5 kg N ha−1) as compared with the cultivar CD1104. Interactions between inoculation and cultivars resulted in greater accumulation of S and K in the shoot of the wheat cultivar CD150, as well greater accumulation of Cu in CD1104. In growth assessments, inoculation or cultivars did not statistically influence wheat grain yield and spike population. Howevere, for plant height, the CD1104 genotype has 13.1% bigger plant height on average than that of the CD150 genotype. Inoculation can contribute more sustainably to wheat nutrition.
Introduction
Wheat (Triticum aestivum L.) stands and ranks second in world cereal production with volume of around 760 million tons (CONAB, 2019). It is an annual cycle plant, being considered a winter and spring cereal with great economic importance (Galindo et al., 2019a). The world average wheat consumption as food was 98.7 kg person−1 year−1 in 2019 (USDA – United States Department of Agriculture, 2020). The climatic extremes and food security predicted and estimated that the agriculture sector will face the challenge of expanding food production by 70% in the year 2050 to meet the increased demand without significant price impacts and shortages in the world (FAO, 2019; ONU, 2019).
Nitrogen (N) is one of the most necessary nutrients in high doses to meet the nutrient limitation and current wheat productivity demands (Teixeira Filho et al., 2012; Galindo et al., 2016). The complex nature of N dynamics in the soil and the use of non-ideal management practices in several crops (Abalos et al., 2014) lead to a large amount of N losses from leaching and volatilization that can affect water bodies and pollute the environment (Bindraban et al., 2015). Given its high cost in the production process and the need for agriculture that maintains ecosystems and biodiversity, the search for solutions that combine production increases and increasingly sustainable agricultural practices is necessary.
In this sense, inoculation and application of plant growth-promoting bacteria (PGPB) especially Azospirillum brasilense is an important strategy in cereal cultivation. Researches about A. brasilense report enhanced plant growth by a number of mechanisms, including but not limited to the synthesis and secretion of hormones that increase the root system (Pankievicz et al., 2015). Greater accumulation and availability of nutrients (Galindo et al., 2016; Rosa et al., 2020) and greater tolerance to stresses (such as drought, salinity), plant vigor (Forni et al., 2017), chlorophyll content, and stomatal conductance (Bulegon et al., 2017), as well as increases in grain productivity (Munareto, 2016; Galindo et al., 2019a), were also reported.
The triticulture countries like Brazil and United States have the potential to reduce N fertilizers by 30–40 kg ha−1 through inoculation with A. brasilense (Fukami et al., 2016, 2017). This is due to the capability of inoculation to develop a well-penetrated root system, thus exploring more soil volume and resulting in higher nutrient absorption (Galindo et al., 2018). Ardakani et al. (2011) reported a synergistic effect between inoculation and nutrient absorption, increasing the absorption efficiency of N, P, and K in wheat by inoculation with A. brasilense. Nevertheless, Teixeira Filho et al. (2017) observed that inoculation with A. brasilense strains AbV5 and AbV6 increased the concentrations of Ca, Mg, Mn, and Zn in wheat grains. The authors also concluded that inoculation is a low-cost, easy-to-apply, and pollutant-free technique that fits the desired sustainable context in reality.
The most frequent technique of inoculation is via seeds; however, spraying during sowing has also demonstrated similar possible results (Munareto, 2016). Several studies indicated positive impacts on crop productivity via seed inoculation (Teixeira Filho et al., 2017), as in the sowing furrow (Morais et al., 2016), or via the leaf (Ferreira et al., 2014; Munareto, 2016). Munareto (2016) highlighted that inoculation with A. brasilense via the leaf is an efficient strategy to increase productivity of wheat cultivars in the Brazilian Cerrado region. It should also be noted that the efficiency of inoculation via seeds can be affected by the use of chemicals during treatment, such as fungicides and insecticides, which reinforces the importance of researches and analysis to use supplementary forms of inoculation.
Inoculation with A. brasilense can take place in three ways as also described above (Teixeira Filho and Galindo, 2019); however, few studies have evaluated the effectiveness of application via the leaf, and using this method as an alternative to increase the uptake of N and other nutrients in wheat crop is more ecological and less costly, reducing the risks or incompatibilities with chemical seed treatments and enabling even greater productivity. Although one or more benefits in the literature have been found, increased accumulation of nutrients and increased productivity of wheat have not always been observed, highlighting the need for further studies. Based on the above discussion, this study was aimed to evaluate the efficiency of inoculation with A. brasilense via the leaf in wheat crop in an irrigated area, verifying its effect on biometrics, grain yield, and nutritional accumulation of wheat cultivars and studying the effect of PGBP on crop nutrient uptake.
Materials and Methods
Location of the Experimental Design
The study was carried out under field conditions at the Teaching, Research and Extension Farm of São Paulo State University (UNESP) in the city of Selvíria, state of Mato Grosso do Sul, Brazil (20°20′43″S; 51°24′7″W, at 355 m above sea level), in 2016. The soil of the experimental area is classified as Dystrophic Red Latosol, clay texture (EMBRAPA, 2018), and was classified as clayey Oxisol (Rhodic Hapludox) according to the Soil Survey Staff (2014). The experimental area was cultivated with annual crops (cereals and legumes) for more than 30 years, with the last 12 years using the no-till system (direct planting system). The last harvest sequence was corn before wheat sowing.
The climatic classification of the region is Aw-Köppen (Alvares et al., 2013) with an average annual rainfall of 1,232 mm and average annual temperature of 24.5°C (Hernandez et al., 1995). The maximum and minimum temperatures and relative humidity of the air and rain indexes were all verified during the study and summarized in Figure 1. The experimental area has a sprinkler irrigation system via a central pivot with 14 mm application layer for 72-h watering shifts.
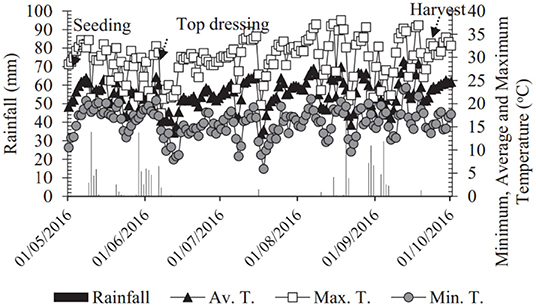
Figure 1. Rainfall and maximum, average, and minimum temperatures obtained from the weather station located at the Education and Research Farm of UNESP during wheat cultivation in the period May 2016 to October 2016.
Twenty soil samples were collected randomly before the beginning of the field trial and mixed well, forming a subsample, which was used to determine the chemical attributes according to Raij et al. (2001). The results are shown in Table 1.

Table 1. Soil chemical attributes in 0–0.20 m layer before the beginning of the wheat field trial.
Experimental Setup
Wheat cultivars CD150 and CD11104 (Coodetec®) are the most used wheat varieties in Latin America (especially in Brazil and Paraguay) with good performance aptitude for the experimental region. Wheat sowing was carried out on 16th May 2016 and harvest on 5th September 2016 (totaling 116 days cycle) with sowing density of 400 viable seeds m−2 (99% purity and 93% germination) in both varieties. The planting furrows were fertilized with formulated NPK (08-28-16) at the dose of 275 kg ha−1. Chemical seed treatment occurred 24 h before sowing, using fungicide and insecticide [thiophanate-methyl + pyraclostrobin fungicide (56 g + 6 g active ingredient—ai—per 100 kg seeds) and insecticide fipronil (62 g ai per 100 kg seeds)] in both cultivars.
The experiment was designed in completely randomized blocks with six replications, arranged in a 2 × 2 factorial scheme. The treatment combination was composed of two wheat cultivars (CD150 and CD1104) and inoculation with PGPB (control—without inoculation and inoculation with A. brasilense via the leaf). Each experimental plot was composed of 12 lines of 5 m in length spaced 0.17 m between the lines. The useful area of each plot was eight central lines with the exclusion of 0.5 m from each end. The size of each plot was 10.2 m2.
Plant emergence occurred 6 days after sowing, on 22nd May 2016. The inoculation with A. brasilense (strains Abv5 and Abv6, with a guarantee of 2 × 108 CFU ml−1) was carried out via leaf spray at 9 days after crop emergence (DAE). The spray was done in the early morning under a clear sky (no cloudiness) using a CO2-type pump sprayer with a full cone nozzle at a dose of 200 ml ha−1 of the commercial inoculant in a solution volume of 120 L ha−1. Nitrogen fertilization in coverage was carried out at 28 and 43 DAE, using 70 kg N ha−1 (applied as urea) in each stage. A total of 140 kg N ha−1 in coverage was applied and incorporated by central pivot irrigation on the same day.
In crop management, weed control was carried out in the total area by applying Pendimentalina (4 L ha−1) right after sowing, methyl methulfuron (3.3 g ai ha−1) at 20 DAE and 2,4-dichlorophenoxy (1.2 L ha−1) at 35 DAE. To prevent disease in the spike, trifloxystrobin + tebuconazole (0.1 L ha−1) was sprayed at 75 and 90 DAE, respectively. There was no need of insecticide application in the experimental area.
Assessments
The evaluation of nutritional accumulation in wheat shoot was carried out at the full flowering stage (50% wheat plants with fully expanded spike) by collecting the aerial part (aboveground) of the wheat plants (1 × 0.17 m) from the central lines of each experimental unit. The collected materials were placed in a forced air ventilation oven at 65°C until uniform mass (72 h) is obtained and weighed with an analytical precision balance (0.001 g) to obtain the dry mass of the plants. Afterwards, the samples were ground in a Wiley mill with a 1-mm (20 mesh) sieve, and the nutrients were determined as described by Malavolta et al. (1997). Using the concentrations of these nutrients and the dry matter of the collected plants, we calculated the accumulation of nutrients in wheat shoot and extrapolated the data into kg ha−1.
At the end of the wheat crop cycle, at the time of harvest, the following attributes were determined: (a) plant height: as being the distance (m) from the ground level to the apex in a cluster of 15 plants plot−1; (b) spike population: determined by the spikes counted in 0.51 m2 (0.17 × 3 m of the line) in the central rows of each plot; and (c) grain yield: by manual harvesting of all plants in the useful area of each plot [eight central lines, excluding 0.5 m at the ends being calculated after mechanical threshing and data obtained were corrected to 13% humidity (wet basis) and transformed into 60 kg bags per ha−1].
Statistical Analysis
The Shapiro–Wilk normality test was used to analyze the data set, followed by the variance and covariance test (F test, p < 0.05). The comparison of treatment means was performed by Tukey test (p < 0.05). Statistical calculations were performed using the Minitab v17.0 software and graphics were made using SigmaPlot v12.5 software.
Results
Interactions between wheat cultivars and inoculation with A. brasilense were observed in the accumulation of K in the shoot of the wheat crop (Supplementary Table 1; p < 0.05). Regarding the nutritional accumulation of primary macronutrients (N, P, and K), with the exception of P, positive responses were verified for accumulations of N and K (Figures 2A–C). The wheat genotype CD150 resulted in greater N accumulation in wheat shoot with an average difference of 33.1% in relation to the CD1104 genotype. Higher accumulation of K in the shoot was observed by analyzing the interaction between cultivars and inoculation with A. brasilense, where the control (without) resulted in greater accumulation of K by 36% in the genotype CD150 in relation to CD1104 (Table 2, Figure 2C).
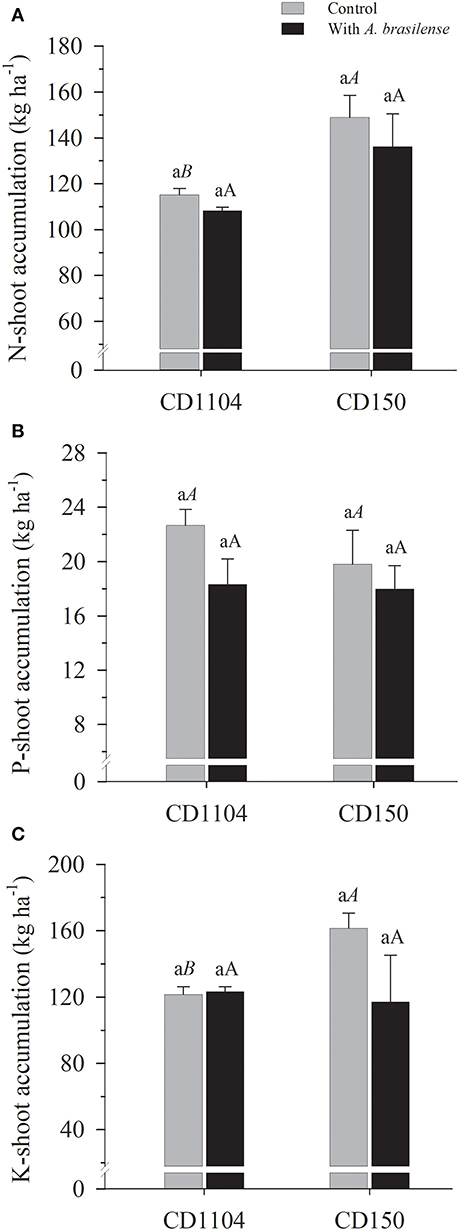
Figure 2. Effects of inoculation with Azospirillum brasilense on the shoot of the wheat cultivars CD1104 and CD150 in the nutritional accumulation of N (A), P (B), and K (C). Different letters indicate difference between means according to a Tukey test (p ≤ 0.05). Lowercase letters correspond to cultivars within each inoculation level, and uppercase letters correspond to inoculations within each cultivar level. Identical letters do not differ from each other according to a Tukey test (p ≤ 0.05). CV (%) = 17.9 (A), 16.2 (B), and 18.8 (C). Error bars indicate the standard error of the mean (n = 6 replications).
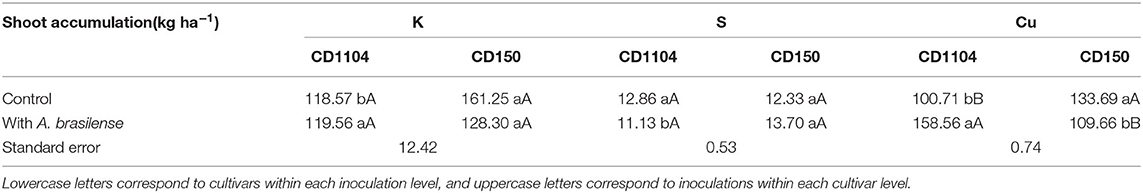
Table 2. Interaction between wheat cultivars and inoculation with Azospirillum brasilense for K, S, and Cu shoot accumulation.
Interactions between wheat cultivars and inoculation with A. brasilense were observed for the accumulation of S in the shoot (Supplementary Table 1; p < 0.01). Responses in the accumulation of secondary macronutrients such as Ca and Mg were observed to be not significant among both genotypes and inoculation with A. brasilense (Figures 3A,B; Supplementary Table 1). Significant responses in the accumulation of S were observed by the interaction between cultivars and inoculation. The inoculation with A. brasilense efficiently increased the accumulation of S in the wheat genotype CD150 in relation to genotype CD1104. This corresponds to a difference of 23.1% (Table 2, Figure 3C).
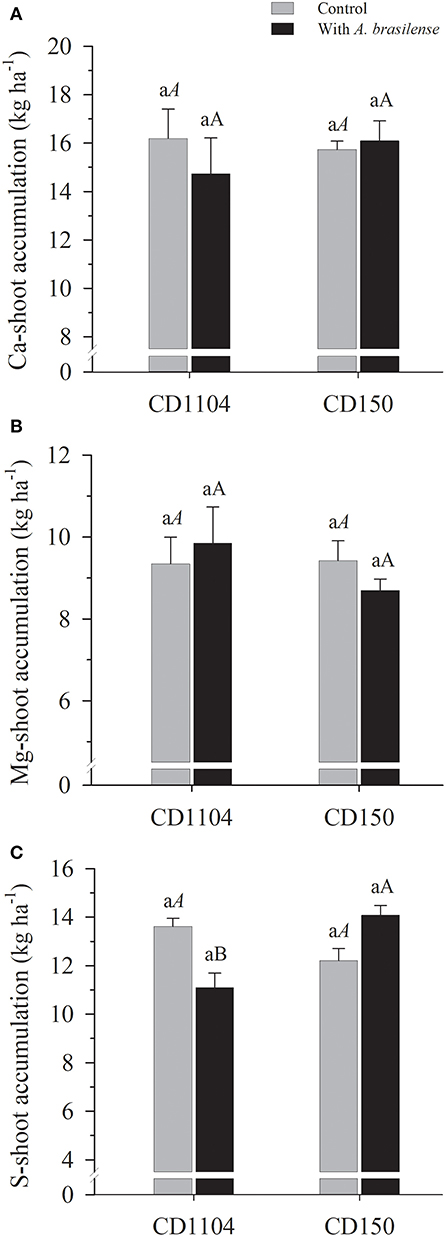
Figure 3. Effects of inoculation with A. brasilense on the shoot of the wheat cultivars CD1104 and CD150 in the nutritional accumulation of Ca (A), Mg (B), and S (C). Different letters indicate difference between means according to a Tukey test (p ≤ 0.05). Lowercase letters correspond to cultivars within each inoculation level, and uppercase letters correspond to inoculations within each cultivar level. Identical letters do not differ from each other according to a Tukey test (p ≤ 0.05). CV (%) = 20.1 (A), 18.2 (B), and 11.9 (C). Error bars indicate the standard error of the mean (n = 6 replications).
The inoculation with A. brasilense also influenced the accumulation of micronutrients in the wheat shoot (Supplementary Table 2; p < 0.01). With the exception of Zn, greater accumulation of B and Cu in response to inoculation with A. brasilense for the CD1104 genotype, in the order of 73.1 and 158.1%, respectively, was observed, in comparison with the control (without inoculation). Meanwhile, for CD150, A. brasilense increased the accumulation of B, Fe, and Mn in the order of 43.8, 49.9, and 22%, respectively, while reducing the accumulation of Cu in the order of 21.9%, for this genotype, in comparison with the control (Figures 4, 5A, Supplementary Table 2).
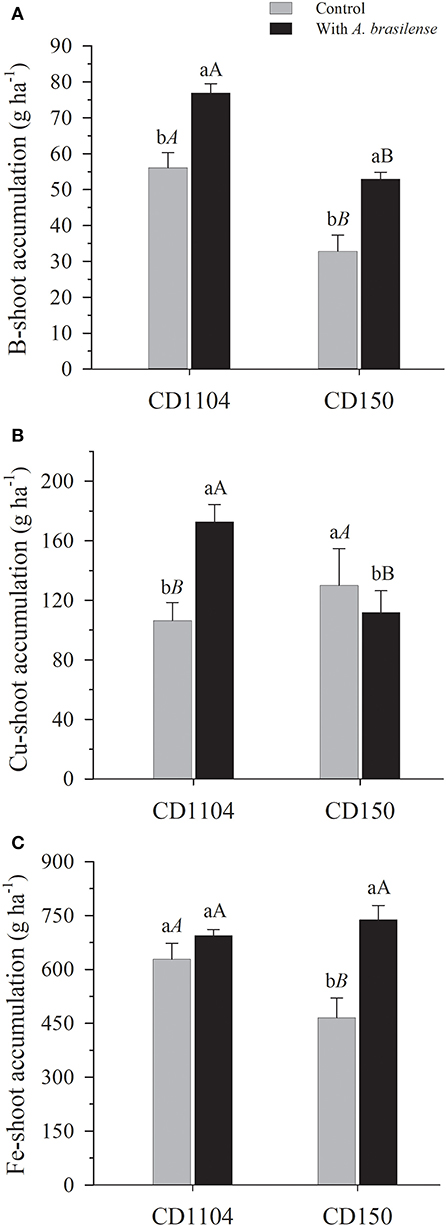
Figure 4. Effects of inoculation with A. brasilense on the shoot of the wheat cultivars CD1104 and CD150 in the nutritional accumulation of B (A), Cu (B), and Fe (C). Different letters indicate difference between means according to a Tukey test (p ≤ 0.05). Lowercase letters correspond to cultivars within each inoculation level, and uppercase letters correspond to inoculations within each cultivar level. Identical letters do not differ from each other according to a Tukey test (p ≤ 0.05). CV (%) = 8.7 (A), 9.3 (B), and 13.9 (C). Error bars indicate the standard error of the mean (n = 6 replications).
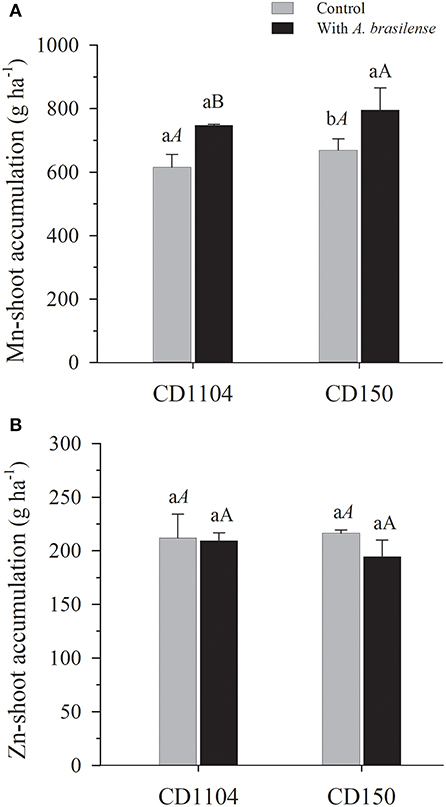
Figure 5. Effects of inoculation with A. brasilense on the shoot of the wheat cultivars CD1104 and CD150 in the nutritional accumulation of Mn (A) and Zn (B). Different letters indicate difference between means according to a Tukey test (p ≤ 0.05). Lowercase letters correspond to cultivars within each inoculation level, and uppercase letters correspond to inoculations within each cultivar level. Identical letters do not differ from each other according to a Tukey test (p ≤ 0.05). CV (%) = 9.8 (A) and 10.8 (B). Error bars indicate the standard error of the mean (n = 6 replications).
We also observed that the wheat cultivars differed on the accumulation of micronutrients in the shoot (Figures 4, 5, Supplementary Table 2). The cultivars influenced the accumulation of B and Mn in the shoot (Supplementary Table 2). The results of the CD1104 genotype demonstrated greater accumulation capacity of B with an average of 65.16 g accumulated B ha−1, whereas genotype CD150 resulted in the accumualtion of 41.37 g B ha−1 in the wheat shoot (Figure 4A), with a difference of 57.5% greater accumulation of B in the CD1104 genotype. Inverse behavior was observed for the accumualtion of Mn, where the CD150 genotype resulted in higher Mn accumulation (737.42 g Mn ha−1) with 13.3% greater accumulation than the CD1104 genotype (650.57 g Mn ha−1) (Figure 5A). With respect to Cu accumulation, inoculation with A. brasilense promoted the accumulation of Cu in the wheat genotype CD1104 by 57.5% in relation to the control (Figure 4B). It was also interesting that inoculation with A. brasilense decreased the accumulation of Cu in genotype CD150 by 18% in relation to its control. Interactions between cultivars and inoculation were observed for the accumulation of Cu in the wheat shoot (Supplementary Table 2, p < 0.01).
The wheat grain yield (Figure 6A) and spike population (Figure 6B) were not significantly influenced by wheat genotypes or by the A. brasilense inoculation in the present study. Analyzing the wheat genotypes, significant differences were observed in the height of the wheat genotypes (Figure 6C; Supplementary Table 3). The CD1104 genotype has 13.1% bigger plant height on average than the CD150 genotype (Figure 6C). This behavior was also visually characterized in the field which can be observed in the records of Supplementary Figure 1.
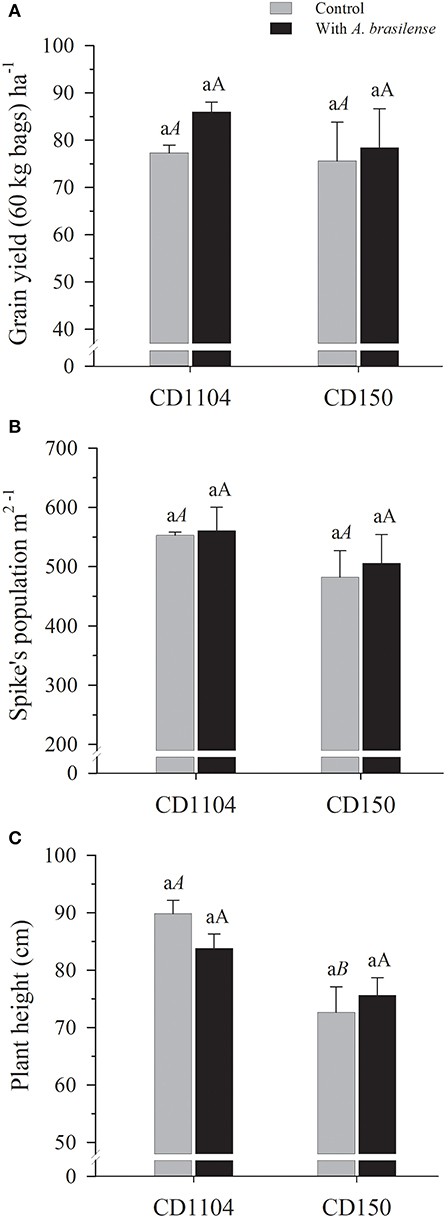
Figure 6. Grain yield (A), spike population (B), and plant height (C) of the wheat cultivars CD1104 and CD150 subjected to inoculation with A. brasilense. Different letters indicate difference between means according to a Tukey test (p ≤ 0.05). Lowercase letters correspond to cultivars within each inoculation level, and uppercase letters correspond to inoculations within each cultivar level. Identical letters do not differ from each other according to a Tukey test (p ≤ 0.05). CV (%) = 10.6 (A), 12.86 (B), 12.9 (C). Error bars indicate the standard error of the mean (n = 6 replications).
Discussion
Interestingly, the CD150 genotype, despite being a predecessor of CD1104 in the wheat cultivar released line (COODETEC, 2017), was reported to have a higher capacity of N uptake and input in the wheat shoot (Figure 2A). Being a constituent of amino acids, proteins, chlorophyll, and several secondary metabolites, N is the most interfering nutrient in the productivity of crops, especially non-atmospheric N2 fixers (Marschner, 2012). It is among the mineral nutrients that has been noted to have higher concentrations in plant tissues and grains and makes it the most required nutrient in wheat crop productivity (Galindo et al., 2019b). The average accumulation of nutrients in the shoot at the wheat flowering stage of both cultivars resulted in a decreasing trend of K > N > P > Ca > S > Mg. Phosphorous (P) has several important functions in plants, is a constituent element of the ATP molecule, and has a role in the supply and storage of energy in metabolic processes, such as the synthesis of organic compounds and the absorption of nutrients, in addition to being involved in the phosphorylation and electron transport reactions of photosynthesis (Hawkesford et al., 2012). Calcium (Ca) is an important enzyme activator having an indispensable role in the formation of pollen grains and growth of the pollen tube, and it is also a constituent of cell wall together with Mg. Being a central ion of the chlorophyll molecule, Mg is involved in the activation of Rubisco in the Calvin cycle (Taiz et al., 2017).
Potassium (K) is a non-structural component that participates in more than 60 enzymatic reactions in plants, such as playing an important function in the osmotic potential of cells, regulating stomata opening and closing, and helping in the transport of carbohydrates (Marschner, 2012). Teixeira Filho et al. (2017) evaluated A. brasilense strains (AbV5–AbV6) in wheat and observed its response in the higher contents of Ca and Mg exported by grains leading to high grain productivity, although accumulation of K was not influenced. The data obtained for K in the present study (Table 2, Figure 2C) demonstrates that the response patterns of this particular bacterium–PGPB association are not only dependent on the ideal conditions, such as temperature, viable CFUs, and PGPB affinity, for establishing inoculation, but also on physiological–molecular interrelationships, intrinsic to plant genetic material (cultivars). Thus, cultivars are able to respond in different ways to the same bacterial strain in the same environment (climate and soil) under the same experimental conditions.
Bacteria of the genus Azospirillum are free-living gram-negative species (Galindo et al., 2016), native to the soil, and its association with plants has a series of beneficial responses such as increasing the synthesis of photosynthetic pigments (chlorophyll) (Bulegon et al., 2017), increasing the content of ascorbic acid (Fasciglione et al., 2015), providing greater tolerance to stress (Forni et al., 2017) and greater accumulation and availability of nutrients (Hungria et al., 2010; Galindo et al., 2016), and finally, helping to achieve a higher grain yield (Munareto, 2016; Galindo et al., 2019a).
The accumulation of S followed a similar trend as that of K under cultivar–inoculation interaction (Figure 3B, Table 2). Involved in structural and metabolic functions, S has a synergistic relationship and a triple bond with N in the synthesis of proteins, in addition to being involved in the metabolism of carbohydrates and lipids and decarboxylation reactions (Astolfi, 2009). Similar results were found by Teixeira Filho et al. (2017), where the wheat inoculated with A. brasilense was observed with higher accumulation of S in the straw as compared to the non-inoculated. This is probably due to the greater development of the root system of the inoculated wheat, which allowed greater absorption of S-SO4− in the depth of 0.20–0.40 m, where normally we find higher sulfur contents in the soil.
Evaluating the performance of wheat cultivars in the accumulation of micronutrients, responses to boron were observed in both cultivars (Figure 4A; Supplementary Table 2). Boron participates in the development of pollen grains and pollen tube, promoting cell wall deposition. Therefore, it participates in cell division, increasing cell size and transporting carbohydrates in plants (Marschner, 2012). Manganese is an important enzyme activator, especially in redox reactions of the Krebs cycle, and acts as a co-factor in the the process of photosystem II (Hawkesford et al., 2012).
Regarding the greater absorption of micronutrients due to inoculation with A. brasilense (Figures 4, 5A), Teixeira Filho et al. (2017) also reported positive responses of A. brasilense in the nutrition of irrigated wheat in the Brazilian Cerrado. These findings demonstrated the importance and response of PGPBs not only in fixing atmospheric N2, but it is also involved in several other mechanisms such as production of sidophores, low molecular organic compounds that boost iron uptake from the soil, solubilizing and transporting it to plant (Banik et al., 2016), synthesis of abscisic acid (ABA) and zeatin, tolerance to pathogens (Tortora et al., 2011; Sahoo et al., 2013), production of kinetin (Patel and Saraf, 2017), and cytokinin that promotes growth.
According to Galindo et al. (2017), A. brasilense is a bacterium with high mobility in plants and assists in several metabolic processes of plant tissues to better use available water in the soil (Shakir et al., 2012); to increase soluble sugar, amino acid, and proline contents (Bano et al., 2013); and to absorb and assimilate nutrients (Galindo et al., 2017). It has also improved the assimilation of P (Rosa et al., 2020) by solubilization of non-available P in the soil rhizosphere (Rodriguez et al., 2004) to increase its availability.
Regarding the absorption and accumulation of copper builders in different cultivars (Figure 4B, Table 2), this behavior of A. brasilense in CD150 may be related to greater absorption of other cationic micronutrients such as B, Fe, and Mn, which were promoted with the inoculation with A. brasilense in the same cultivar, by 44, 49.9, and 21.9% (Figures 4A–C, 5A). The cationic micronutrients compete for the same sites of absorption in the cell of the plant root (Marschner, 2012), which may justify the accumulation in CD150. Furthermore, when analyzing the contents of these nutrients in the soil (Table 1), all micronutrients were at a high level for the cultivation of wheat and other cereals (Raij, 2011). These findings support the hypothesis of multiple researches triggered by A. brasilense in cereal which go far beyond the capacity of biological nitrogen fixation (Pankievicz et al., 2015) and synthesis of phytohormones (Bashan and De-Bashan, 2010). The inoculation with A. brasilense signals intrinsic absorption processes (Fukami et al., 2017) and solubilization (Rodriguez et al., 2004; Arzanesh et al., 2011; Banik et al., 2016), promoting better use of micronutrients (Galindo et al., 2019b), which, therefore, highlights the need for more specific studies to understand these mechanisms and metabolic pathways of plants.
Regarding the influence of A. brasilense, wheat genotypes and their effect on biometric evaluations (height and spike population) were studied at harvest time and resulted in taller plants (Figures 6B,C). This characteristic was clearly observed in the current experimental stand, which can be observed in Supplementary Figure 1. The attributes regarding plant height and size were consistently high in the technical data of each genetic material (COODETEC, 2017). However, regions with a high incidence of wind blow can damage the cultivation of CD1104, as taller wheat plants are more prone to lodging which may lead to more losses in the harvest (Nascimento et al., 2016).
In contrast to plant height, grain yield in the present study was not influenced by wheat genotypes or inoculation with A. brasilense during the harvest (Figure 6A). It was studied by Tabassum et al. (2017) and Ferreira et al. (2019) in an extensive review on PGPB-induced capacity of plant tolerance to stress, and they highlighted that inoculation with A. brasilense does not always increase productivity. The authors postulated that inoculation responses are most strongly expressed in plants in adverse situations such as water deficit (Shakir et al., 2012), pathogen attack, and plant–weed competition, and light, water, and nutrient deficiencies are among the other abiotic factors (Forni et al., 2017).
In the present study, the climatic situations were normal, and no adverse situations occurred, coinciding with the explanations proposed by Tabassum et al. (2017) and Ferreira et al. (2019) for the none increasing productivity (Figure 6). Climatic values can be consulted from the temperature and exclusion graphs during the experimental period (Figure 1). In addition, it is important to note that the experimental area has a central pivot irrigation system, grown with cereals (cereals and legumes for over 30 years), the last 12 years being under the no-tillage system with straw conservation and minimal soil turnover aiming at the conservation of the soil–plant–microorganism system.
Conclusions
The inoculation with A. brasilense increases the accumulation of B and Cu in the aerial part of wheat, by 27.7 and 57.4%, respectively, for the cultivar CD150. A. brasilense increased the accumulation of B, Fe, and Mn by 43.8, 49.9, and 22%, respectively, and reduced Cu by 21.9% in the cultivar CD1104.
The wheat cultivar CD150 has greater efficiency of N shoot accumulation of +35.5 kg N ha−1 in comparison with cultivar CD1104. However, the grain yield of the wheat cultivars was similar.
Interactions between A. brasilense inoculation and cultivars resulted in greater accumulation of S and K in the wheat shoot for cultivar CD150, as well as Cu shoot accumulation in the CD1104 genotype. A. brasilense has more sustainable contribution to wheat nutrition with no significant increase in crop productivity.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author Contributions
EB wrote the manuscript, with contributions from AJ and MT. FS, OF, and SB corrected and improved the manuscript. EB, WR, JS, and FS conducted the samplings and data collection. EB and BL did the analysis, with the support of SB, WR, and MS. All authors contributed to the article and approved the submitted version.
Funding
This research was funded by CNPq (National Council for Scientific and Technological Development), award numbers 305.299/2014-0 and 312359/2017-9.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fsufs.2020.607262/full#supplementary-material
References
Abalos, D., Jeffery, S., Sanz-Cobena, A., Guardia, G., and Vallejo, A. (2014). Meta-analysis of the effect of urease and nitrification inhibitors on crop productivity and nitrogen use efficiency. Agric. Ecosyst. Environ. 189, 136–144. doi: 10.1016/j.agee.2014.03.036
Alvares, C. A., Stape, J. L., Sentelhas, P. C., De Moraes, G., Leonardo, J., and Sparovek, G. (2013). Köppen's climate classification map for Brazil. Meteorologische Zeitschrift Stuttgart 22, 711–728. doi: 10.1127/0941-2948/2013/0507
Ardakani, M. R., Mazaheri, D., Mafakheri, S., and Moghaddam, A. (2011). Absorption efficiency of N, P, K through triple inoculation of wheat (Triticum aestivum L.) by Azospirillum brasilense, Streptomyces sp., Glomus intraradices and manure application. Physiol. Mol. Biol. Plants. 17, 181–192. doi: 10.1007/s12298-011-0065-7
Arzanesh, M., Alikhani, H., Khavazi, K., Rahimian, H., and Miransari, M. (2011). Wheat (Triticum aestivum L.) growth enhancement by Azospirillum sp. under drought stress. World J. Microbiol. Biotechnol. 27, 197–205. doi: 10.1007/s11274-010-0444-1
Astolfi, S. (2009). Sulfur in plants: an ecological perspective. Ann. Bot. 104:viii. doi: 10.1093/aob/mcp164
Banik, A., Mukhopadhaya, S. K., and Dangar, T. K. (2016). Characterization of N2-fixing plant growth promoting endophytic and epiphytic bacterial community of Indian cultivated and wild rice (Oryza spp.) genotypes. Planta 243, 799–812. doi: 10.1007/s00425-015-2444-8
Bano, Q., Ilyas, N., Bano, A., Zafar, N., Akram, A., and Hassan, F. (2013). Effect of Azospirillum inoculation on maize (Zea mays L.) under drought stress. Pak. J. Bot. 45:1320.
Bashan, Y., and De-Bashan, L. E. (2010). How the plant growth-promoting bacterium Azospirillum promotes plant growth - a critical assessment. Adv. Agron. 108, 77–136.
Bindraban, P. S., Dimkpa, C., Nagarajan, L., Roy, A., and Rabbinge, R. (2015). Revisiting fertilisers and fertilisation strategies for improved nutrient uptake by plants. Biol. Fertil. Soils 51, 897–911. doi: 10.1007/s00374-015-1039-7
Bulegon, L. G., Guimaraes, V. F., Klein, J., Battistus, A. G., Inagaki, A. M., Offemann, L. C., et al. (2017). Enzymatic activity, gas exchange and production of soybean co-inoculated with'Bradyrhizobium japonicum' and'Azospirillum brasilense. Aust J. Crop Sci. 11, 888–896. doi: 10.21475/ajcs.17.11.07.pne575
CONAB (2019). Acompanhamento da safra Brasileira de grãos (In portuguese), v. 6 - Safra 2018/19 - Décimo segundo levantamento, Brasília, 1–47, setembro 2019. Available online at: http://www.conab.gov.br (accessed March 30, 2020).
COODETEC (2017). Wheat cultivars, Product guide (In portuguese), 12–13 and 18–19. Available online at: http://unisagro.com/pastaarquivos/85Coodetec%202017%20-%20Guias%202017%20Guia%20Completo%20Centro.pdf (accessed August 14, 2020).
EMBRAPA (2018). Brazilian Agricultural Research Corporation. National Soil Research Center. Brazilian system of soil classification (In Portuguese). 3a Edn. Brasília, DF: EMBRAPA, 353p.
FAO (2019). Food and Agriculture Organization of the United Nations. Available online at: http://www.fao.org/search/pt/?cx=018170620143701104933%3Aqq82jsfba7w&q=population+growth&cof=FORID%3A9&siteurl=www.fao.org%2Fbrasil%2Fpt%2F&ref=www.google.com%2F&ss (accessed September 14, 2020).
Fasciglione, G., Casanovas, E., Quillehauquy, V., Yommi, A., Goñi, M., Roura, S., et al. (2015). Azospirillum inoculation effects on growth, product quality and storage life of lettuce plants grown under salt stress. Scientia Horticulturae. 195, 154–162. doi: 10.1016/j.scienta.2015.09.015
Ferreira, C. M. H., Soares, H. M. V. M., and Soares, E. V. (2019). Promising bacterial genera for agricultural practices: an insight on plant growth-promoting properties and microbial safety aspects. Sci. Total Environ. 682, 779–799. doi: 10.1016/j.scitotenv.2019.04.225
Ferreira, J. P., Andreotti, M., Arf, O., Kaneko, F. H., Nascimento, V., and Sabundjian, M. T. (2014). Inoculation with Azospirillum brasilense and nitrogen in topdressing in wheat in Cerrado region. Tecnologia e Ciência Agropecuária 8 27–32. (in Portuguese with abstract in English)
Forni, C., Duca, D., and Glick, B. R. (2017). Mechanisms of plant response to salt and drought stress and their alteration by rhizobacteria. Plant Soil 410, 335–356. doi: 10.1007/s11104-016-3007-x
Fukami, J., Nogueira, M. A., Araujo, R. S., and Hungria, M. (2016). Accessing inoculation methods of maize and wheat with Azospirillum brasilense. AMB Express 6, 3–16. doi: 10.1186/s13568-015-0171-y
Fukami, J., Ollero, F. J., Megías, M., and Hungria, M. (2017). Phytohormones and induction of plant-stress tolerance and defense genes by seed and foliar inoculation with Azospirillum brasilense cells and metabolites promote maize growth. AMB Express 7, 153–163. doi: 10.1186/s13568-017-0453-7
Galindo, F. S., Teixeira Filho, M. C. M., Buzetti, S., Rodrigues, W., Boleta, E. H. M., Santini, J., et al. (2018). Effects of boron (B) doses and forms on boron use efficiency of wheat. Aust. J. Crop Sci. 12, 1835–2707. doi: 10.21475/ajcs.18.12.09.PNE1382
Galindo, F. S., Teixeira Filho, M. C. M., Buzetti, S., Rodrigues, W. L., Santini, J. M. K., and Alves, C. J. (2019a). Nitrogen fertilization efficiency and wheat grain yield affected by nitrogen doses and sources associated with Azospirillum brasilense. Acta Agric. Scand. B Soil Plant Sci. 69, 606–617. doi: 10.1080/09064710.2019.1628293
Galindo, F. S., Teixeira Filho, M. C. M., Buzetti, S., Santin, J. M. K., Alves, C. J., and Ludkiewicz, M. G. Z. (2017). Wheat yield in the Cerrado as affected by nitrogen fertilization and inoculation with Azospirillum brasilense. Research Agropec. Bras. 52, 794–805. doi: 10.1590/S0100-204X2017000900012
Galindo, F. S., Teixeira Filho, M. C. M., Buzetti, S., Santini, J. M. K., Alves, C. J., Nogueira, L. M., et al. (2016). Corn yield and foliar diagnosis affected by nitrogen fertilization and inoculation with Azospirillum brasilense. Revista Brasileira de Ciência do Solo 40:e015036. doi: 10.1590/18069657rbcs20150364
Galindo, F. S., Teixeira Filho, M. C. M., Buzetti, S., Santini, J. M. K., Montanari, R., Freitas, L. A., et al. (2019b). Micronutrient accumulation with Azospirillum brasilense associated with nitrogen fertilization management in wheat. Commun. Soil Sci. Plant Anal. 50, 2429–2441. doi: 10.1080/00103624.2019.1667369
Hawkesford, M., Horst, W., Kichey, T., Lambers, H., Schjoerring, J., Møller, I. S., et al. (2012). “Functions of macronutrients,” in Marschner's Mineral Nutrition of Higher Plants, 3rd Edn, ed P. Marschner (Pergamon: Elsevier/Academic Press), 135–189. doi: 10.1016/B978-0-12-384905-2.00006-6
Hernandez, F. B. T., Lemos Filho, M. A., and Buzetti, S. (1995). Software hidrisa and the water balance of Ilha Solteira (In Portuguese). Ilha Solteira: UNESP/FEIS/DCSER, 34–45.
Hungria, M., Campo, R. J., Souza, E. M. S., and Pedrosa, F. O. (2010). Inoculation with selected strains of Azospirillum brasilense and A. lipoferum improves yields of maize and wheat in Brazil. Plant Soil. 331, 413–425. doi: 10.1007/s11104-009-0262-0
Malavolta, E., Vitti, G. C., and Oliveira, S. A. (1997). Evaluation of the Nutritional Status of Plants: Principles and Applications. (In Portuguese). 2nd Edn, 319. Piracicaba, São Paulo: Potafos.
Marschner, P. (2012). Marschner's Mineral Nutrition of Higher Plants, 651. 3rd Edn. New York, NY: Academic Press.
Morais, T. P., Brito, C. H., Brandao, A. M., and Rezende, W. S. (2016). Inoculation of corn with Azospirillum brasilense in the sowing furrow (In portuguese). Rev. Ciênc. Agron. 47, 290–298. doi: 10.5935/1806-6690.20160034
Munareto, J. D. (2016). Aspects physiological seed, wheat grain productivity applied nitrogen doses, inoculation and Azospirillum brasilense of foliar application. 55 f. Dissertação (Mestrado em Agronomia). Federal University of Santa Maria, Santa Maria, Brazil. Available online at: http://repositorio.ufsm.br/handle/1/5171 (accessed February 23, 2020).
Nascimento, V., Arf, O., Tarsitano, M. A. A., Garcia, N. F. S., Penteado, M. S., and Sabundjian, M. T. (2016). Economic analysis of rice and common bean production in succession to green manure crops and mecahnical soil decompression in Brazilian Savannah. Rev. Ceres. 63, 315–322. doi: 10.1590/0034-737X201663030006
ONU – United Nations Organization. (2019). World Population Prospects 2019: Highlights (ST/ESA/SER.A/423). New York, NY: Department of Economic and Social Affairs, Population Division.
Pankievicz, V. C. S., Amaral, F. P., Santos, K. F. D. N., Agtuca, B., Xu, Y., Schueller, M. J., et al. (2015). Robust biological nitrogen fixation in a model grass-bacterial association. Plant J. 81, 907–919. doi: 10.1111/tpj.12777
Patel, T., and Saraf, M. (2017). Biosynthesis of phytohormones from novel rhizobacterial isolates and their in vitro plant growth-promoting efficacy. J. Plant Interact. 12, 480–487. doi: 10.1080/17429145.2017.1392625
Raij, B, van, Andrade, J. C., Cantarella, H., and Quaggio, J. A. (2001). Chemical Analysis for Fertility Evaluation of Tropical Soils. (In Portuguese). Campinas, Brazil: IAC, 285.
Raij, B. van. (2011). Soil Fertility and Nutrient Management. Piracicaba: International Plant Nutrition Institute, 420.
Rodriguez, H., Gonzalez, T., Goire, I., and Bashan, Y. (2004). Gluconic acid production and phosphate solubilization on by the plant growth promoting bacterium Azospirillum spp. Naturwissenschaften 91, 552–555. doi: 10.1007/s00114-004-0566-0
Rosa, P. A. L., Mortinho, E. S., Jalal, A., Galindo, F. S., Buzetti, S., Fernandes, G. C., et al. (2020). Inoculation with growth-promoting bacteria associated with the reduction of phosphate fertilization in sugarcane. Front. Environ. Sci. 8, 1–32. doi: 10.3389/fenvs.2020.00032
Sahoo, R. K., Ansari, M. W., Dangar, T. K., Mohanty, S., and Tuteja, N. (2013). Phenotypic and molecular characterisation of efficient nitrogen-fixing Azotobacter strains from rice fields for crop improvement. Protoplasma 251, 511–523. doi: 10.1007/s00709-013-0547-2
Shakir, M. A., Asghari, B., and Arshad, M. (2012). Rhizosphere bacteria containing ACC-deaminase conferred drought tolerance in wheat grown under semi-arid climate. Soil Environ. 31, 108–112.
Tabassum, B., Khan, A., Tariq, M., Ramzan, M., Khan, M. S. I., Shahid, N., et al. (2017). Bottlenecks in commercialisation and future prospects of PGPR. Appl. Soil Ecol. 121, 102–117. doi: 10.1016/j.apsoil.2017.09.030
Taiz, L., Zeiger, E., Møller, I. M., and Murphy, A. (2017). Plant Physiology and Development, 6th Edn. Porto Alegre: Artmed.
Teixeira Filho, M. C. M., Buzetti, S., Garcia, C. M. P., Sá, M. E., Arf, O., and Maeda, A. S. (2012). Nitrogen doses and fungal application in wheat crop under no-tillage system. Cultura Agronomica 21, 31–41. doi: 10.1590/S0103-84782011000800013
Teixeira Filho, M. C. M., and Galindo, F. S. (2019). Inoculation of bacteria with a focus on biological nitrogen fixation and promotion of plant growth. Topics in Soil Science. (In Portuguese). Braz. Soil Sci. Soci. 11, 577–648.
Teixeira Filho, M. C. M., Galindo, F. S., Buzetti, S., and Santini, J. M. K. (2017). “Inoculation with Azospirillum brasilense improves nutrition and increases wheat yield in association with nitrogen fertilization,” in Wheat Improvement, Management and Utilization, eds R. Wanyera and J. Owuoche J. (IntechOpen).
Tortora, M. L., Díaz-Ricci, J. C., and Pedraza, R. O. (2011). Azospirillum brasilense siderophores with antifungal activity against Colletotrichum acutatum. Arch. Microbiol. 193, 275–286. doi: 10.1007/s00203-010-0672-7
USDA – United States Department of Agriculture (2020). United State Department of Agriculture. Foreign Agricultural Service / Global Market Analysis: International Production Assessment Division (IPAD). Available online at: https://apps.fas.usda.gov/psdonline/circulars/production.pdf (accessed February 25, 2020).
Keywords: cereal, no-tillage system, Triticum aestivum L., crop nutrition, plant-growth-promotion, sustainable-agriculture
Citation: Boleta EHM, Shintate Galindo F, Jalal A, Santini JMK, Rodrigues WL, Lima BHd, Arf O, Silva MRd, Buzetti S and Teixeira Filho MCM (2020) Inoculation With Growth-Promoting Bacteria Azospirillum brasilense and Its Effects on Productivity and Nutritional Accumulation of Wheat Cultivars. Front. Sustain. Food Syst. 4:607262. doi: 10.3389/fsufs.2020.607262
Received: 16 September 2020; Accepted: 23 November 2020;
Published: 23 December 2020.
Edited by:
Everlon Cid Rigobelo, Universidade Estadual Paulista, BrazilReviewed by:
Ernesto Garcia-Pineda, Universidad Michoacana de San Nicolás de Hidalgo, MexicoGustavo Moro, Universidade Estadual Paulista, Brazil
Copyright © 2020 Boleta, Shintate Galindo, Jalal, Santini, Rodrigues, Lima, Arf, Silva, Buzetti and Teixeira Filho. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marcelo Carvalho Minhoto Teixeira Filho, bWNtLnRlaXhlaXJhLWZpbGhvQHVuZXNwLmJy