 Roberto A. Distel
Roberto A. Distel José I. Arroquy
José I. Arroquy Sebastián Lagrange3,4
Sebastián Lagrange3,4 Juan J. Villalba
Juan J. Villalba- 1Departamento de Agronomía, Universidad Nacional del Sur (UNS) - Centro de Recursos Naturales Renovables de la Zona Semiárida (CERZOS), UNS-CONICET, Bahía Blanca, Argentina
- 2Instituto Nacional de Tecnología Agropecuaria (INTA), Estación Experimental Agropecuaria Cesáreo Naredo, Casbas, Argentina
- 3Department of Wildland Resources, Quinney College of Natural Resources, Utah State University, Logan, UT, United States
- 4Estación Experimental Agropecuaria Bordenave, Instituto Nacional de Tecnología Agropecuaria (INTA), Bordenave, Argentina
Pasture-based production systems represent a significant sustainable supplier of animal source foods worldwide. For such systems, mounting evidence highlights the importance of plant diversity on the proper functioning of soils, plants and animals. A diversity of forages and biochemicals –primary and secondary compounds- at appropriate doses and sequences of ingestion, may lead to benefits to the animal and their environment that are greater than grazing monocultures and the isolated effects of single chemicals. Here we review the importance of plant and phytochemical diversity on animal nutrition, welfare, health, and environmental impact while exploring some novel ideas about pasture design and management based on the biochemical complexity of traditional and non-traditional forage sources. Such effort will require an integration and synthesis on the morphology, ecophysiology, and biochemistry of traditional and non-traditional forage species, as well as on the foraging behavior of livestock grazing diverse pasturelands. Thus, the challenge ahead entails selecting the “right” species combination, spatial aggregation, distribution and management of the forage resource such that productivity and stability of plant communities and ecological services provided by grazing are enhanced. We conclude that there is strong experimental support for replacing simple traditional agricultural pastures of reduced phytochemical diversity with multiple arrays of complementary forage species that enable ruminants to select a diet in benefit of their nutrition, health and welfare, whilst reducing the negative environmental impacts caused by livestock production systems.
Introduction
Numerous current studies highlight the importance of plant diversity for the proper functioning of soils, plants and herbivores (Eisenhauer et al., 2018; Hautier et al., 2018; Schaub et al., 2020). This is because plant diversity affects soil physical, chemical, and biological attributes, both indirectly through the promotion of biomass production, and directly through plant species, to soil attributes and functioning (Coleman et al., 2017). Plants feed and grow soil biota through their litter and root-derived organic inputs (i.e., root exudates, root necromass), which are increased by plant diversity (Zak et al., 2003). In addition, plant species differ in belowground structure and function, which influence soil biota and directly relate to soil functioning (McNally et al., 2015; Eisenhauer et al., 2017). Recent results have demonstrated the importance of soil biodiversity for soil functionality at the local level and across biomes (Delgado-Baquerizo et al., 2020).
With regards to primary productivity, many studies, and meta-analyses have shown that plant diversity increases the productivity and stability of plant communities (e.g., Hector et al., 1999; Tilman et al., 2001; Isbell et al., 2009, 2015, 2017; Polley et al., 2013; Prieto et al., 2015). The three primary mechanisms that have been proposed to explain the positive relationship between plant diversity and plant productivity are niche differentiation, positive interactions and selection effects (Hector et al., 2002). Niche differentiation arises due to species differences in morphological and physiological characteristics, which allow differential use of resources in space and time, thus increasing primary productivity when grown in mixtures. Positive interactions result from facilitation between species [e.g., associative nitrogen fixation from legumes to grasses (Kakraliya et al., 2018)], whereas selection effects result from an increased probability of the presence of species that perform the best in a certain growing environment (Valencia et al., 2018). Plant communities composed of different functional groups of species are also expected to exhibit greater temporal stability in yield, because they are more resistant or resilient to environmental or biological disturbances due to differences in tolerance among species (Cottingham et al., 2001; Polley et al., 2013). Note that plant communities composed of species functional groups with different responses to changes in environmental conditions become critical in the face of future climate change. Finally, plant diversity is strongly correlated with phytochemical diversity at the community level (Moore et al., 2014; Marzetz et al., 2017), which is a biological need for the ruminant animal's optimal expression of its potential functioning (Provenza et al., 2007).
Plant diversity, and the inherent phytochemical diversity in plant communities, are biologically important to the ruminant animal for several reasons. First, ruminants are generalist herbivores and they evolved experiencing a multidimensionality of orosensorial and post-ingestive stimuli that contribute to improved fitness (e.g., Rapport, 1980; Provenza et al., 2007; Beck and Gregorini, 2020). Dietary mixing is thought to benefit herbivores by allowing either a more balanced intake of nutrients (Westoby, 1978; Rapport, 1980; Provenza et al., 2003) or a diluted ingestion of toxins (Freeland and Janzen, 1974; Marsh et al., 2006). A corollary of dietary mixing theory is that it allows generalist herbivores to reach similar fitness in habitats with different forage and chemical compositions (Franzke et al., 2010). For instance, metabolic disorders caused by excessive nutrient intake from a single forage could be diluted by ingesting alternative sources that differ in nutrient concentration (Rutter, 2006). Phytochemical diversity may also help counteract toxicity caused by single plant secondary metabolites (PSMs), because nutrients attenuate the negative post-ingestive effects of certain toxins through an enhancement in detoxification and elimination pathways (Illius and Jessop, 1995), and because some PSMs form stable complexes with other PSMs in the gastrointestinal tract that attenuate toxicity (Villalba and Provenza, 2005; Copani et al., 2013). In addition, ingestion of diverse PSMs, at appropriate doses, could provide medicinal benefits (Cozier et al., 2006), improve product quality (Priolo and Vasta, 2007; Vasta et al., 2019), and reduce the negative environmental impacts from ruminants' (e.g., enteric methane and nitrous oxide emissions, and nitrate leaching; Mueller-Harvey et al., 2019; Clemensen et al., 2020; Lagrange et al., 2020). Finally, individuals differ in their need for nutrients and tolerance to toxins due to inherent morphological and physiological differences (Provenza et al., 2003). Therefore, a diversity of forage species allows for the expression of such individual variability and a better fulfillment of individual needs than a uniform diet designed to satisfy requirements for the average animal within a group.
Even though there is mounting experimental evidence on the importance of plant diversity for soil, plant and ruminant animal's functioning, and thus on farmer profitability (Schaub et al., 2020), current agricultural pasture systems are usually composed of either one or a few “conventional” or widely known plant species. There is a need for designing diverse agricultural pasture systems, to replace traditional ones of limited diversity. Nevertheless, devising diverse pasture-based grazing systems for improving ruminant production and welfare, while reducing environmental impacts, entails a big challenge. It requires an extensive set of work of analysis, integration and synthesis of knowledge on the morphology, physiology and biochemistry at the plant species and biochemical level, and on the interactions among plant species/genotypes/chemicals under grazing conditions. From this complexity, it may be possible to create pasture mixtures that on the one hand enhance primary productivity through complementary and positive interactions between species, and on the other, enhance animal health, welfare and the efficiency of nutrient use by ruminants.
This review is limited to a consideration on the importance of plant diversity for ruminants to perform at their potential, and some ideas on designing diverse pasturelands and chemical landscapes (i.e., chemoscapes). Our aim was to highlight the key role of forage diversity on ruminant production systems, given that there is emerging experimental evidence on the benefits of chemically and taxonomically diverse plant communities on animal fitness. Then, we engage in some basics of pasture design, such as selection of species composition, spatial arrangement and grazing management of forage mixtures. By this means, we expect to stimulate novel fundamental research and applied approaches aimed at the design of diverse pasturelands that enrich the ruminant's environment and enhance the system's efficiency while remaining productive for multiple years.
An Explanation for Diverse Diets in Ruminants
Dietary diversity is ubiquitous among mammalian herbivores. When allowed to select among alternative foods of different types and concentration of nutrients and PSMs, ruminant animals learn to select varied diets that meet their nutritional requirements and circumvent toxicity and nutritional disorders (Provenza, 1995; Villalba et al., 2002, 2004). Nutrient constraints and detoxification limitations have been proposed as alternative biological bases of varied diets. The “nutrient constraints” or “nutrient complementation” hypothesis argues that no one plant species can provide all nutrients in the proportion needed by herbivores and thus dietary mixing allows for a more balanced nutrient intake (Westoby, 1974, 1978; Rapport, 1980). The “detoxification limitation hypothesis” argues that the detoxification systems of animals are incapable of metabolizing high levels of PSMs present in a single plant species, and thus, PSMs ingested as a mixture are less toxic because they are less concentrated and potentially detoxified by different pathways (Freeland and Janzen, 1974). These hypotheses are not mutually exclusive, and both assume an underlying physiological and behavioral mechanism.
Transient food aversion has also been proposed as a non-mutually exclusive alternative to nutrient balancing and toxin dilution for explaining partial preference and diverse diets in ruminant animals (Provenza, 1996). The underlying mechanism of food aversion is the association between sensorial receptors (that respond to a food's taste, odor, texture, visual aspect) and visceral receptors (that respond to chemical and physical stimuli), which enable herbivores to learn through post-ingestive consequences (Provenza, 1995). Temporary food aversions develop when the same food is consumed too frequently or in excess, when the food is nutritionally imbalanced, restricted in nutrient content or when it contains toxins (Provenza, 1996). Through this mechanism, animals attempt to fulfill their metabolic requirements and achieve homeostasis (Villalba and Provenza, 2009). Note that, animals can develop temporary aversions even for nutritionally balanced diets, since animals satiate when the same food is eaten repeatedly or in excess (Provenza, 1996). It has been argued that hedonic and motivational incentives associated with foods, through experiences and expectations of rewards, are also determinants of feeding behavior (Ginane et al., 2015).
In summary, the transient food aversion hypothesis suggests that dietary diversity is based on the nutritional and toxicological disorders experienced by ruminants consuming nutritional unbalanced and/or potentially toxic feeds. Such disorders (e.g., acidosis, hyperammonemia, bloat, toxicity) commonly occur when herbivores are faced to single feeds. If alternative complementary foods are available, animals could circumvent this constrain through their diet selection. For instance, lambs increase their preference for a chemical buffer (bentonite; Villalba et al., 2006, sodium bicarbonate; Phy and Provenza, 1998), and dairy cows increase their intake of larger feed particles that stimulate saliva production (Kmicikewycz and Heinrichs, 2015) when experiencing ruminal acidosis. High intakes of readily degradable sources of nitrogen lead to increments in the concentration of ammonia in the peripheral circulation once the liver detoxification threshold is surpassed (Lobley and Milano, 1997). This increase causes reductions in food intake, mediated through aversive post-ingestive feedback, which occurs very quickly within a meal (Villalba and Provenza, 1997). Sheep fed a basal diet high in rumen-degradable protein and allowed to ingest a feed with condensed tannins (PSMs that bind to proteins and reduce their ruminal degradability) showed reduced rumen ammonia nitrogen and blood urea nitrogen, and a tendency to develop a preference for and intake of the tannin-containing feed (Fernández et al., 2012).
Pasture bloat occurs in fresh, high-protein forages, with high rate of particle breakdown that results in a rapid release of plant soluble proteins and disruption of chloroplasts, providing large quantities of gas and bacterial slime, which create a stable foam that prevents the animal eructation of fermentation gases (CO2 and CH4), and thus promote rumen distension (Majak et al., 2003). Sheep learn to avoid foods that cause rumen distension and to prefer foods that attenuate this effect (Villalba et al., 2009).
The aforementioned theories of partial preference focus on just one aspect of diet like presence of plant toxins, flavors or nutrients. A multifaceted theory that considers processes beyond feed properties like motivation to eat (e.g., Ginane et al., 2015) and interactions with other factors such as the animal's past experiences (e.g., Provenza, 1995), sequence of feed ingestion (e.g., Yearsley et al., 2006) and energetic costs of food acquisition and processing (e.g., Hobbs et al., 2003) is still lacking, although the transient food aversion hypothesis is a first approach into fulfilling this need.
Diverse Diets: Nutrients, Plant Secondary Metabolites, and Flavors
Ruminants grazing diverse chemoscapes are faced with the complex task of building a diet with appropriate proportions and concentrations of nutrients that satisfy their individual needs, while balancing the ingestion of potentially toxic (but also medicinal) PSMs (Provenza, 1995; Villalba et al., 2017). In the process, animals are exposed to a diverse array of flavors that influence their grazing behavior (Villalba et al., 2011). This section reviews the influence of nutrients, PSMs and flavors during grazing on some key aspects of animal nutrition, health, welfare, and environmental impact.
Diverse Diets: Nutrient Intake
A diversity of forages and biochemicals available in pasturelands may enhance the nutritional benefits that forages offer to ruminants because complementary relationships among multiple food resources in nature improves animal fitness (Tilman, 1982). This is because diverse diets offer ruminants a variety of biochemicals (nutrients and PSMs) which allow for associative effects and synergies with the potential to enhance the efficiency of nutrient utilization relative to single forage species in monocultures (Provenza et al., 2003; Waghorn and McNabb, 2003). Biodiversity in pasturelands may lead to positive associative effects among forages, which improve the nutrition (i.e., nitrogen retention, diet digestibility) and welfare of livestock (i.e., reductions in stress caused by single forages with unbalanced nutrient profiles and monotonous flavors).
Ruminants mix forage alternatives that lead to a balanced diet at greater levels of intake than for single species (Askar et al., 2006; Villalba et al., 2015). For instance, sheep and goats eating mixed diets in rangeland display daily dry matter intakes two or more times greater than reference intake values obtained with animals fed single forages of similar nutritive value (Agreil and Meuret, 2004; Meuret and Provenza, 2015). The consumption of different legumes with contrasting chemical composition (i.e., different content of non-fiber carbohydrates, fiber and proteins) and presence of PSMs leads to associative effects, like rumen protein degradability lower than the average of the individual forages. This reduces ammonia formation (and thus its toxic effects and nitrogen loses to the environment) while increasing the amount of dietary protein reaching the small intestine (Mueller-Harvey, 2006; Waghorn, 2008). This is supported by in vitro studies for a mixture of sainfoin (Onobrichis viciifolia) and cocksfoot (Dactylis glomerata) (Niderkorn et al., 2012) and by in vivo studies with beef cows grazing combinations of sainfoin, alfalfa (Medicago sativa) and birdsfoot trefoil (Lotus corniculatus) (Lagrange et al., 2020). Moreover, since individuals differ morphologically and physiologically, the possibility of free choice among food alternatives allows for the expression of individual nutritional needs (Provenza et al., 2007; Baraza et al., 2009).
Cattle fed the ingredients of a total mixed ration in a free-choice test were able to select a diet adequate to meet their individual needs, without compromising gain (kg/day) and food conversion efficiency relative to a mixed ration that prevented animals from selecting individual ingredients (Atwood et al., 2001; Moya et al., 2011). Moreover, individual animals varied considerably in their preferred ratio of protein to energy, which resulted in lower food cost/day and cost/kg gain compared to animals fed the total-mixed ration (Atwood et al., 2001). In another study, sheep offered a free choice of three legume species, differing in nutrient and PSMs content, showed enhanced intake and diet digestibility relative to feeding single species (Lagrange and Villalba, 2019). Goats fed a free choice of five shrub species selected a mixed diet of greater digestibility than controls fed the single shrub species (Egea et al., 2016).
Diverse Diets: Environmental Impact
The challenge ahead entails creating diverse pastures that enhance phytochemical richness and enable animals to practice selectivity, whilst also having positive effects on the environment. Forage combinations with a diversity of biochemical compositions may contribute to reductions in carbon and nitrogen footprints by ruminant animals, a positive attribute that adds value to livestock products beyond their nutritional quality (Rochfort et al., 2008; Patra and Saxena, 2010). It is recognized that a negative byproduct of ruminant production systems entails negative environmental impact (de Vries and de Boer, 2010; de Vries et al., 2015). The largest contributing source of greenhouse gas (GHG) emissions from beef cattle production is enteric methane (CH4), accounting for 56% (Rotz et al., 2019) to 63% (Beauchemin et al., 2010) of all GHG from beef industry and 39% of all GHG emissions from the livestock sector. Methane is a byproduct of the microbial fermentation of feeds in the rumen, which represents an energy loss to the animal that ranges between 2 and 12% of the gross energy consumed with the diet (Johnson and Johnson, 1995). Forages with high concentration of non-fibrous carbohydrates, that are readily fermented in the rumen (i.e., soluble carbohydrates plus pectin) and low proportion of structural carbohydrates (cellulose and hemicellulose), enhance the efficiency of nutrient use by cattle, yielding animal weight gains that are comparable to feeding high-grain rations (Chail et al., 2017; MacAdam, 2019). This chemical profile increases the proportion of potentially propionate-forming bacteria and decreases hydrogen production, which results in decreased CH4 emissions relative to forages with lower content of non-fibrous carbohydrates (Sun et al., 2015; Stewart et al., 2019). In vitro rumen fermentation of perennial ryegrass forages differing in the concentration of water-soluble carbohydrates showed lower acetate:propionate ratio and CH4 concentration in high than in low sugar ryegrass pastures (Rivero et al., 2020). Similarly, lambs fed fresh winter forage rape (Brassica napus) showed reduced CH4 yields compared with lambs fed ryegrass, a response which was attributed to higher concentration of non-structural carbohydrates and lower ruminal pH in the former than in the latter forage species (Sun et al., 2012, 2015, 2016).
Incorporation of species rich in bioactive PSMs in diverse pastures also reduces CH4 production in the rumen. For instance, legume species containing phenolic compounds (condensed tannins) like sainfoin have been shown to reduce methane emissions (Wang et al., 2015). A recent study showed lower numerical values of enteric methane emissions by heifers grazing combinations of legumes (alfalfa, birdsfoot trefoil, sainfoin) relative to controls grazing monocultures of the same species. Heifers offered the combinations showed the greatest body weight gains, implying reductions in the number of days to slaughter, which reduces methane emissions during the finishing process (Lagrange et al., 2020). Other legume species like Macrotyloma axillare also showed antimethanogenic potential, associated with a decrease in the relative abundance of methanogenic archaea and protozoa (Lima et al., 2018, 2020). Essential oils, of which terpenes are major compounds, have been demonstrated to reduce methane production during in vitro and in vivo studies (Cobellis et al., 2016).
Greenhouse gas emissions from livestock production systems also involve the production of the potent GHG nitrous oxide (N2O) (Rotz et al., 2019). High levels of ammonia in urine “hot spots” are sources of this gas produced during microbial nitrification and denitrification processes (Oenema et al., 2005; Huang et al., 2015). Strategies to reduce problems with excess of nitrogen, while maintaining high levels of animal productivity, entails the provision of high sugar and bioactive-containing forages that increase nitrogen retention and/or reduce the proportion of urinary nitrogen losses. For instance, nitrogen use efficiency is higher in high than in low sugar ryegrass pastures (Rivero et al., 2020). Polyphenols, like condensed tannins in sainfoin or birdsfoot trefoil bind to proteins protecting their degradation in the rumen (Scharenberg et al., 2007; Theodoridou et al., 2010, 2012), which alters the fate of the excreted nitrogen to greater fecal to urinary ratios (Mueller-Harvey, 2006). A shift in the route of nitrogen excretion from urine to feces means more stable nitrogen fractions in manure since nitrogen is mainly bound to organic compounds like neutral detergent and acid detergent insoluble nitrogen, which lessens the rate of nitrogen losses to the environment (Whitehead, 2000; Grosse Brinkhaus et al., 2016; Stewart et al., 2019). A diversity in the chemical structures of condensed tannins in sainfoin and birdsfoot trefoil (McAllister et al., 2005) may influence their capacities to bind proteins and microbial enzymes in the rumen (Mueller-Harvey et al., 2019), which could promote positive associative effects that attenuate protein degradability in the rumen, and thus the fate of nitrogen excretion. In support of this, heifers grazing a choice between strips of sainfoin and birdsfoot trefoil showed declines in urinary nitrogen and blood urea nitrogen relative to animals grazing an alfalfa monoculture (Lagrange et al., 2020). Moreover, this decline was even greater than reductions observed for the single tanniferous species grazed individually. This novel finding suggests a positive associative effect between condensed tannins on the reduction of ruminal protein degradation, attributed to the different chemical structure of condensed tannins in different legumes. Condensed tannins in birdsfoot trefoil have average molecular weight of 4,400 Da (McAllister et al., 2005), with a degree of polymerization in the range of 6 to 14 of predominantly procyanidin type subunits of oligomers and polymers (Jonker and Yu, 2017), while sainfoin's condensed tannins are basically constituted by prodelphinidin monomers of a mean molecular weight of 5,100 Da (McAllister et al., 2005), with polymer sizes that vary between 4 and 12 subunits (Jonker and Yu, 2017).
Another problem with excesses of urinary nitrogen deposited in beef production systems entails the eutrophication of watersheds by nitrates, produced by ammonia oxidation and then leached into ground water, streams and lakes (Whitehead, 2000). Based on experimental results involving diverse pastures, in combination with a whole-farm model, significant reductions in nitrogen leaching were predicted for a well-drained soil in the Waikato region of New Zealand when replacing traditional simple forage mixtures by complex forage mixtures in dairy farm systems (Romera et al., 2017). An integrated modeling assessment of intensive sheep and beef production systems for the Cantebury region of New Zealand also predicted reductions in nitrogen leaching by using complex forage mixtures (Vogeler et al., 2017). For instance, the inclusion of forbs in can reduce nitrogen leaching (Totty et al., 2013; Bryant et al., 2018). The reduction in nitrogen leaching is strongly associated with declines in urinary nitrogen concentration, which may reflect either a better nutritionally balanced diet or PSMs-protein binding that decreases the amount of rumen degradable protein (Waghorn, 2008; Mueller-Harvey et al., 2019) in complex forage mixtures. Furthermore, reduced urinary nitrogen concentration in cows that graze diverse pastures can lead to significantly decreased nitrous oxide emissions during the denitrification process (Di and Cameron, 2016).
Diverse Diets: Plant Secondary Metabolite Intake and Toxic Effects
Ruminants grazing diverse landscapes typically encounter plants that, in addition to nutrients, contain potentially toxic PSMs. Mammalian herbivores can ingest toxins up to a threshold level determined by their potential detoxification capacity (Freeland and Janzen, 1974; Dearing et al., 2000, 2005). The accomplishment of this potential is dependent upon nutrient availability, given that nutrients (carbohydrates, protein) are needed to fuel detoxification mechanisms (Illius and Jessop, 1995; Villalba and Provenza, 2005). Moreover, because different toxins may be metabolized through distinctive detoxification mechanisms, food intake is less compromised if a diversity of PSMs is consumed such that no single detoxification pathway is saturated in the process (Freeland and Janzen, 1974). For instance, lambs consume greater amounts of dry matter when they have available a choice among feeds containing different types of PSMs that are metabolized through different detoxification pathways (oxalates, condensed tannins, terpenes) than when only one feed with one PSM is available (Villalba et al., 2004). Likewise, PSMs can bind to each other forming stable molecular bonds in the gastrointestinal tract (e.g., alkaloid-condensed tannins or alkaloid-saponins) that are not absorbed and then excreted through feces, which neutralize their negative post-ingestive effects. Sheep fed foods with different alkaloids and either condensed tannins or saponins, ate more food than when offered only the foods with alkaloids (Lyman et al., 2008). Similarly, when cattle and sheep grazed first a forage high in tannins or saponins, they subsequently increased their grazing time on alkaloid-containing forages (Lyman et al., 2011; Owens et al., 2012). Complexation of condensed tannins with alkaloids was confirmed during in vitro studies (Villalba et al., 2016; Clemensen et al., 2018).
In Mediterranean ecosystems, sheep and goats increase total shrub intake when tanniferous shrubs were fed in combination with a shrub high in saponins, suggesting complementarity between tannin and saponins (Rogosic et al., 2006, 2007). It has also been shown that sheep consume more food with either condensed tannins or terpenes when the basal diet was of high- rather than low-nutritional quality, highlighting the importance of nutrients at enhancing the animals' detoxification and elimination capacities (Baraza et al., 2005).
Diverse Diets: Plant Secondary Metabolite Intake and Medicinal Effects
In present intensive animal production systems, feeding is almost exclusively based on plant primary metabolites (mainly carbohydrates and proteins). Improved forage species and rations are low in concentration and profiles of PSMs given their potential toxic effects (see previous section). However, PSMs at the appropriate dose could provide medicinal benefits (e.g., Moreno et al., 2010). In addition, there is evidence that herbivores learn about these benefits and potentially self-medicate (Engel, 2003; Hutchings et al., 2003). For instance, it has been shown that sheep can form multiple malaise-medicine associations and prefer specific medicines based on different negative physiological states like acidosis, tannin or oxalate toxicosis (Villalba et al., 2006).
Parasitic infections represent one of the main vectors that challenge ruminant health (Hoste et al., 2006, 2015), which force trade-offs between nutrition and parasitism in foraging decisions (Hutchings et al., 2000). In order to counteract the negative effects of parasitism herbivores have also evolved behavioral mechanisms to self-select medicinal foods at effective doses of PSMs (e.g., alkaloids, terpenes, phenols) that minimize toxicity (Glasser et al., 2009; Amit et al., 2013; Villalba et al., 2017). Such mechanism involves the association of a food's flavor with its post-ingestive medicinal effects (i.e., prophylactic self-medication; Juhnke et al., 2012), or the chronic consumption of small daily doses of medicinal PSMs with the animals' diet (i.e., a preventive or prophylactic “feed forward” mechanism; Glasser et al., 2009; Villalba et al., 2014).
Following the same logic described for nutrients, a diversity of medicinal PSMs from an array of varied forages with different mechanisms of antiparasitic action, may increase their effectiveness relative to single PSMs (Hoste et al., 2006). Moreover, since large quantities of PSMs are required to achieve meaningful antiparasitic doses in ruminants (Waghorn and McNabb, 2003), a diverse diet with multiple PSMs may allow animals to harvest the appropriate amounts and ratios of nutrients while consuming diverse antiparasitic PSMs with fewer single harmful side effects to the animal. Thus, complementarities among multiple bioactive molecules have the potential to enhance medicinal effects over single chemicals (Spelman et al., 2006). For instance, minor chemical compounds in plants may act as synergistic metabolites, producing greater overall efficacy than individual components (Hummelbrunner and Isman, 2001). Six chemical compounds from the medicinal plant Petiveria alliacea did not show acaricide activity against the common cattle tick (Rhipicephalus microplusticks) when tested individually in vitro. However, when the compounds were combined, some of the mixtures exhibited a synergistic increase in acaricidal activity, promoting high mortality rates (Arceo-Medina et al., 2016). Such complementarities are not always observed in livestock production systems. For instance, sheep infected with the gastrointestinal nematode Haemonchus contortus and offered a choice between feeds containing condensed tannins or saponins (both antiparasitic PSMs), displayed greater levels of infection than control sheep offered either tannins or saponins in single rations (Copani et al., 2013). As described above, tannins and saponins cross-react and bind in the gastrointestinal tract (Freeland et al., 1985), forming stable complexes that reduce the bioactivity of the single compounds. Thus, the nature of the relationship among PSMs needs to be considered on a case-by-case basis, in order to gain reliable information on whether the combination of compounds in a diverse diet yields synergistic, antagonistic or independent medicinal effects.
Diverse Diets: Flavor Variation and Animal Welfare
Large ruminants are generalist herbivores and they evolved selecting a diverse diet with different orosensorial experiences (Provenza et al., 2007; Villalba et al., 2015). It has also been argued that dietary diversity reduces oxidative and physiological stresses and improves the nutritional status and welfare of animals (Beck and Gregorini, 2020). Forage diversity provides animals with varied sensorial and post-ingestive experiences that increase the motivation to eat (Meuret and Bruchou, 1994; Villalba et al., 2010). Sheep fed the same ration but in a choice of different flavors consumed more total dry matter and tended to gain more weight than sheep exposed to the same ration but containing single flavors (Distel et al., 2007; Villalba et al., 2011). As for toxins or nutrients (see section Diverse Diets: Plant Secondary Metabolite Intake and Toxic Effects), herbivores grazing monocultures of single species or monotonous rations satiate on the orosensorial characteristics of single feeds (i.e., sensory-specific satiety) due to transient food aversions caused by flavors ingested too frequently or in excess, and satiety can be stressful (Provenza, 1996). However, if diverse options are available, animals continue responding to other orosensorial dimensions. This response has been attributed in part to the sensory properties of food, because an animal that stops eating one flavored food will often consume another food or the same food presented in a different flavor (Provenza, 1996; Atwood et al., 2001). Feeding to satiety decreases the responses of hypothalamic neurons to the sight and/or taste of a food on which the animal has been satiated, but leaves the responses of the same neurons to other foods on which the animal has not been satiated relatively unchanged (Rolls et al., 1986).
A diversity of flavors contributes to enhanced animal welfare because generalist herbivores exposed to diverse arrays of feeds have less likelihood of experiencing stressful situations, like frustration due to lack of food alternatives available to build a balanced diet (Villalba and Manteca, 2019), or satiety due to repeated or excessive orosensorial exposure to the same single feeds (Villalba et al., 2010; Catanese et al., 2012). Consistent with this notion, a diversity of food items offered to sheep early in life reduces plasma cortisol (a hormone involved in stress responses by mammals) levels (Villalba et al., 2012; Catanese et al., 2013), lymphocyte counts (Catanese et al., 2013) and stress-induced hyperthermia in open field tests (Villalba et al., 2012) relative to animals fed monotonous rations early in life.
Knowledge Gaps
Despite the emerging findings described above, key information is still lacking regarding the chemical characteristics of different forage constituents that may contribute to specific effects of forages on the animal's internal environment and on potential chemical interactions when multiple forages are ingested. For instance, it is still unknown whether some less-explored constituents of forages such as neutral detergent soluble fiber and other non-structural or non-fiber carbohydrates (Hall et al., 1999) vary in concentration or in composition in response to different biotic or abiotic factors, or on how these carbohydrates interact with other chemicals like PSMs. Breeding programs or managerial interventions (i.e., defoliation frequency), or ecological conditions (i.e., climate, elevation), may influence the concentration or composition of these compounds. In turn, some of these changes may contribute to enhance chemical associations among forages and thus impact some key variables like the efficiency of nitrogen utilization by ruminants. The same can be said about the concentration, composition and chemical affinities of some PSMs like phenolic compounds (Mueller-Harvey et al., 2019), information that will be key for promoting (i.e., through breeding programs or management) forage characteristics that enhance interspecies synergies. Finally, a broader database is needed regarding forage and chemical complementarities like those described by Lagrange et al. (2020) on the combined effect of two tanniferous legumes at reducing urinary nitrogen excretions. This broader knowledge base could be applied in different ecoregions under different environmental conditions and with potentially greater synergistic effects when different species or a broader number of combinations are assayed.
Ruminant Production and Phytochemical Diversity
Based on the beneficial impacts on animal nutrition, welfare and health, it is expected that there would be a positive effect of phytochemical diversity on animal production at the individual level. Diverse phytochemically rich pastures often provide a more nutritious diet compared with monocultures, leading to enhanced forage intake and animal nutrition. Higher per head milk production was observed when cows grazed on complex forage mixtures compared to simple forage mixtures (Totty et al., 2013; Jonker et al., 2019). Similarly, per head milk volume and composition (milk protein, milk fat) was greater for cows grazing on pasture mixtures (Lolium perenne, Trifolium repens, Plantago lanceolata) or spatially adjacent monocultures of the three forages than in L. perenne monoculture (Pembleton et al., 2016). The authors attributed treatment differences to improved nutrition and increase in forage intake of cows grazing on diverse pastures. Experimental evidence also indicates that forage species diversity increased food intake in sheep, especially at the latter phase of the meal (Wang et al., 2010; Feng et al., 2016). Improved sheep, goat and cattle performance has also been reported in mixed grass-legume swards relative to monocultures (Rutter, 2006; Chapman et al., 2007), and in heifers grazing combinations of three different legumes relative to legume monocultures (Lagrange et al., 2020). However, other authors have reported similar per head milk production when cows grazed simple vs. complex forage mixtures (Wedin et al., 1965; Sanderson et al., 2004; Soder et al., 2006). Likewise, increments in species richness of temperate pastures (three, five, or eight forage species) did not influence cow-calf performance (Tracy and Faulkner, 2006).
Forage allowance at plant species level may explain the aforementioned differential ruminant performance in response to pasture diversity. For instance, milk production per cow in ryegrass-clover mixtures was lower when the clover represented 25% total dry matter available than when it represented 50% or 75% of the total dry matter present (Harris et al., 1997). Another possible explanation for the variable ruminant performance in response to complex forage mixtures is that species identity, or chemical identity of PSMs, is more important than the complexity of the mixtures (Deak et al., 2007). As described before for medicinal effects (section Diverse Diets: Plant Secondary Metabolite Intake and Medicinal Effects), it is important to understand the nature of the species interactions on a case-by-case basis. This idea is represented in a study where interactions between legumes and grasses could be either complementary or non-complementary, depending on species identity. A combination of Trifolium repens and Schedonorus pratensis had a positive additive effect on forage digestibility, which was not observed for a combination between Medicago sativa and Phalaris arundinacea (Brink et al., 2015). Moreover, even within a single forage species, genotypic diversity can exert strong influences on herbivore performance (Kotowska et al., 2010). A large-scale metabolomics study in Lolium perenne allowed the identification of high- and low-sugar genotypes (Subbaraj et al., 2019) with implications for variable interactions among forages. Since sugar content is directly and positively related with metabolizable energy and protein capture and supply in ruminants (Richardson et al., 2003; Jacobs et al., 2009), it should be expected that there would be a greater positive additive effect on animal production of high than low-sugar genotypes when associated in mixtures with high-nitrogen legume species. Plant secondary metabolite concentration may also vary among genotypes, as high- and low-tannin content varieties have been identified within different legume species (Donnelly et al., 1971), with potential to impact complementarities and synergies among forages.
Finally, climatic conditions also influence the concentration of biochemicals in forages, which may also influence synergisms and complementarities among forages. Water deficit inhibits photosynthetic activity in plant tissues, owing to an imbalance between light capture and utilization (Reddy et al., 2004), a dysfunction that leads to the generation of reactive oxygen species. In turn, plants have mechanisms of reactive oxygen species detoxification, with some of them involving flavonoids and phenolic compounds like tannins, which are antioxidant (Gourlay and Constabel, 2019). Thus, at least some phenolic compounds in plants are expected to increase their growth under water deficits (Popović et al., 2016). Water stress also reduces the rate of plant maturation (Wilson and Ng, 1975), with increments in the concentration of non-structural carbohydrates in the cell contents of forages like birdsfoot trefoil, sainfoin, white clover (Trifolium repens) and perennial ryegrass (Lolium perenne) (Kuchenmeister et al., 2013). Temperature has a strong influence on plant growth, development and chemical composition (Buxton, 1996). Lignin synthetic enzyme activities increase in plants in response to increasing temperature (Buxton and Fales, 1994), while high proportions of non-structural carbohydrates are metabolized into structural carbohydrates (Deinum and Knoppers, 1979).
Collectively, a broader knowledge base on interactions among multiple forages and chemicals—from in vitro to field studies- will allow for making more accurate predictions about potential synergies in order to create the next generation of functional pasturelands (see section Designing Diverse Agricultural Pastures). Another knowledge gap that needs to be bridged entails a better understanding on the impacts of environmental factors on plant chemistry and their implications for interactions among forages consumed in diverse diets.
Designing Diverse Agricultural Pastures
This review argues that plant diversity has the potential to enhance ecological services in pasturelands such as animal nutrition, health and welfare, while reducing environmental impacts. Nevertheless, pasture diversity is not simply a numbers game of mixing and sowing as many forage species as possible (Sanderson et al., 2007). Kinds and amounts of different forages along with their spatial arrangement and use in time are key variables that need to be considered when designing diverse and multifunctional pasture communities. The first step in undertaking this endeavor entails selecting the forage species and numbers that satisfy specific system goals (e.g., forage production, biodiversity, animal production) (Hobbs and Morton, 1999; Sanderson et al., 2007). Then, managers should ask (1) whether the species selected can tolerate the expected environmental conditions (e.g., soil characteristics, climate, plant-plant competition), (2) if the species should be planted in mixes or in blocks (e.g., spatial arrangement, architecture of the landscape or chemoscape), and finally they should (3) develop an adaptive management plan (e.g., through grazing management) incorporating long-term monitoring (Figure 1). In what follows, we explore some ideas regarding the design of the next generation of multifunctional pasture communities under the context of the aforementioned questions and tasks, with the aim of stimulating new research on the gaps in knowledge identified during the process.
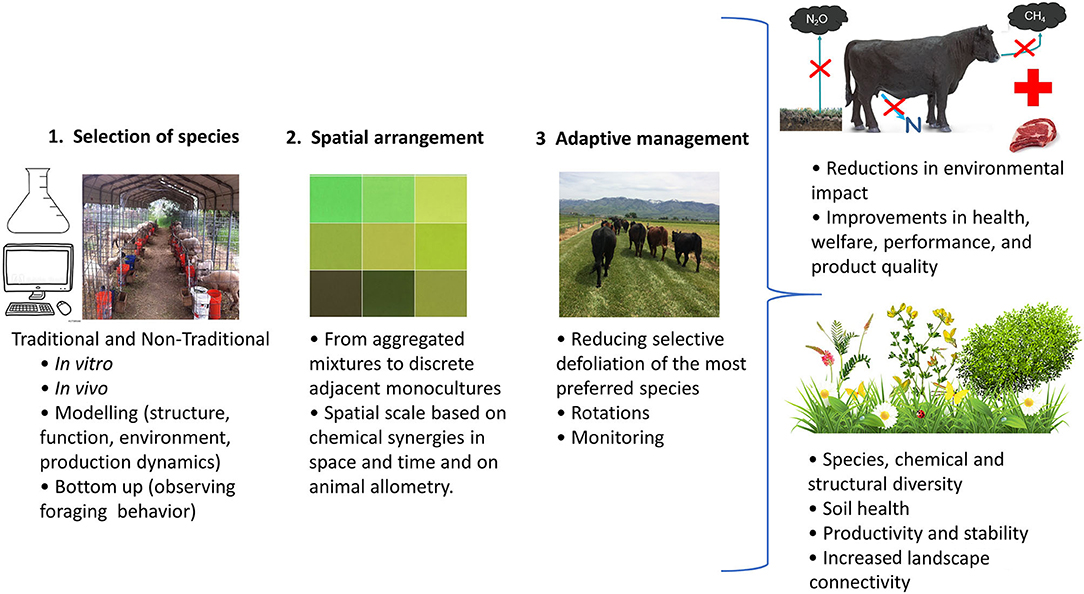
Figure 1. Flow diagram for the design, benefit, and management of diverse, multifunctional pasture communities (expanded from Hobbs and Morton, 1999; Sanderson et al., 2007).
Species Identity
Given our current state of knowledge, it can justifiably be argued that species identity (taxonomy at the level of species/cultivars) to design diverse agricultural pasture systems is critical for pasture productivity and stability as well as for animal performance and environmental impact. Although pasture productivity/ stability is beyond the scope of this review, we will briefly refer to essentials in selecting species to improve productivity and stability of agricultural pastures.
The benefit of plant diversity on productivity and stability is well-documented for natural communities (Lehman and Tilman, 2000; Isbell et al., 2009); however, it is equivocal for agricultural pasture systems (Jing et al., 2017). The most parsimonious possible explanation for the ambivalent responses in agricultural pastures is inadequate selection of species identity. Adequate selection of species represents a big challenge in designing diverse pastures (Tracy et al., 2018). This is because most diverse communities in pasturelands tend to become dominated by one or two species over time (Tracy and Sanderson, 2004; Sanderson et al., 2007; Skinner and Dell, 2016), making monitoring a key task for understanding the compositional “trajectory” of the designed pasture over seasons and years. This scenario suggests the convenience of the management of a few select species (e.g., grass/legume combinations) in order to improve resilience and other ecosystem functions (Tracy et al., 2018). Adequate species selection is also critical in the face of climate change, as it should consider the utilization of diverse mixtures that have the potential to be productive during more challenging climatic scenarios like the predicted increases in ambient temperature, drought and elevated concentration of atmospheric CO2. Diverse agricultural pastures, better adapted to changes in environmental conditions, will be those composed of species varying in tolerance, and thus responses to changes in different climatic parameters. Functionally diverse plant communities typically exhibit greater temporal stability of productivity, than do their counterparts composed of fewer functional groups (Yachi and Loreau, 1999). Thus, a high diversity of cultivated forage species, high intraspecific genetic diversity, and the use of species and variety mixtures have been proposed as a means to enhance productivity and resilience of grasslands in the Mediterranean and Nordic regions challenged by unstable and uncertain climatic conditions (Ergon et al., 2018). Identification of moderately diverse, site-specific grass-legume mixtures and greater use of complementary forage species such as C3 and C4 grasses in order to lengthen the grazing season and provide a buffer against weather variation have also been proposed as strategies to improve the resilience of pasturelands facing changes in climate (Tracy et al., 2018). From the animal perspective, detoxification pathways are thermogenic and PSMs uncouple mitochondrial oxidative phosphorylation, which also generates heat (Beale et al., 2018). Thus, increased ambient temperatures may cause selection of diets with lower content of PSMs (Beale et al., 2018) from pasturelands of declining crude protein content (Craine et al., 2016). Therefore, associations of forages that enhance the efficiency of protein use (i.e., through a balanced provision of non-fibrous carbohydrates or condensed tannins) may be needed in predicted warmer environments.
Besides productivity and stability in production, species identity selection needs to consider improvement in animal performance while reducing environmental impact. The fulfillment of this objective requires the adequate harnessing of phytochemical diversity of both primary and secondary metabolites. Like plant production responses to diverse forage mixtures, animal performance in response to complex mixtures has also been equivocal (see previous section) which was explained through different degrees of complementarities among species identity. Complementary arises not only due to nutrient-nutrient interactions (e.g., carbohydrates-protein) that better match nutrient intake with nutrient demands, but also due to nutrient-PSMs and PSMs-PSMs interactions, as described under section Diverse Diets: Nutrients, Plant Secondary Metabolites and Flavors. Therefore, adequate selection of species identity from the animal side requires a profound knowledge on the chemical profile of individual forage species and interactions among their elements, in the benefit of animal nutrition, welfare, health, productivity, and environmental impact.
Some efforts have been undertaken to explore the influence of plant-plant interactions on plant chemistry and foraging behavior. Concentrations of nitrogen and of an alkaloid (ergovaline) in endophyte-infected tall fescue (E+; Festuca arundinaceas Schreb) were observed to be greater when the plant grew adjacent to legumes than when it grew in monoculture. In contrast, no differences in saponins or condensed tannins concentrations were found when alfalfa or birdsfoot trefoil grew in monoculture or in mixtures (Clemensen et al., 2017). The chemical composition of E+ as influenced by growing next to legumes or not also modified foraging behavior by lambs (Friend et al., 2015). More research is needed on these types of interactions in order to broaden the knowledge base for an informed selection of species based on their chemical composition.
Modeling Approaches
According ecological theory, selection of forage species to create diverse agricultural pastures should be based mainly upon species productivity in the local environment as well as niche differentiation, positive interactions and differential tolerance to disturbance and stress among species. Accordingly, adequate selection of species to design diverse agricultural pastures is a process that requires a profound understanding of the structure and function of forage species and of their interactions. Because of the complexity of diverse agricultural pastures, functional-structural plant modeling represents an important tool to synthesize and integrate knowledge and to recognize research problems (Evers et al., 2019). This approach emerged from single species or growth forms, continuing with models that predicted the behavior of simple mixes considering each species separately, which represented a great complexity of inputs and outputs for highly diverse pastures (Moore et al., 1997). More recently, modeling efforts have focused on functional-structural approaches under the assumption that diverse pasture functioning can be explained by the mean value of biological attributes (i.e., functional traits) of its constituent forage groups (Jouven et al., 2006). Functional traits could be associated with production dynamics (Craine et al., 2002), environmental conditions (Diaz et al., 1998) and responses to defoliation (Louault et al., 2005). Future models should incorporate additional functional traits related to the chemical characteristics of the species, such as type and concentration of PSMs, water-soluble carbohydrates or rumen degradable protein, integrating the knowledge available on biological attributes with chemical dimensions in order to obtain more developed scenarios about the integrated benefits of diverse pastures.
In addition to functional-structural approaches, computational predictive methods have emerged in the field of novel drug discovery as time- and cost-efficient ways to explore potential chemical combinations that are successful to treat disease (Preuer et al., 2018). Drug combinations are investigated across various medical areas, such as cancer, viral disease, fungal, and bacterial infection using predictive methods that select novel synergistic drug combinations from training datasets with available information about investigational combinations (Bulusu et al., 2016; Preuer et al., 2018). A similar approach could be undertaken for pasture design based on chemical associations, with training datasets from nutrient-nutrient, nutrient-PSMs, PSMs-PSMs interactions gathered in multiple studies, in order to explore novel synergistic interactions among different forage species.
In vitro Approaches
In vitro studies have been traditionally used for screening the potential degradability and environmental impacts (i.e., through the production of CH4 and CO2) of single forage species (e.g., Tavendale et al., 2005; Roca-Fernández et al., 2020) and medicinal effects of bioactive-containing plants (e.g., Githiori et al., 2006) given their low cost, rapid turnaround and repeatable results. After careful assessment of the outputs obtained, the most promising candidate treatments are tested in vivo. Time-related gas production techniques have been extensively used to quantify the kinetics of ruminant feed fermentation (Groot et al., 1996). Gas production (e.g., CH4 and CO2) can be quantified and this variable is positively correlated with greater digestibility, greater energy content of the forage and potentially reduced fill effect (Blümmel et al., 2005). The technique also allows for the estimation of organic matter disappearance and fermentation efficiency (Blümmel et al., 1997). More recent studies explore in vitro gas production approaches using combinations of forage mixes relative to the single substrates. For instance, Aufrère et al. (2005) showed in an in vitro study that mixing sainfoin with alfalfa could be an efficient way to reduce the N solubility of pure alfalfa, a result that was then explored in vivo with positive results (Aufrère et al., 2013). Likewise, Niderkorn et al. (2011) tested grass-legume mixtures in vitro, showing that sainfoin can interact with different grasses to reduce the degradation of proteins and the production of CH4 with transitory negative effects on fiber digestion. Finally, but not less important, is that in this type of studies dietary adaptation can affect substrate digestion; therefore, it should be controlled in order to avoid wrong conclusions (Gordon et al., 2002).
Bottom Up Approaches
As for the discovery of new drugs through the observation of sick wild herbivores self-selecting plants in nature (Huffman, 2003), it may be possible to learn more about synergistic forage combinations by observing how ruminants select their diet from diverse pasturelands. This approach has been applied into the design of grass-legume mixtures at biomass availabilities that reflect the preferred proportion selected by the target animal in free-choice scenarios (Chapman et al., 2007). Sheep and cattle grazing perennial ryegrass –white clover pastures, consistently prefer clover over ryegrass in a 70:30 ratio (Rutter, 2006). Thus, plant species availability is planned based on such proportion learned from the animals' preference (Chapman et al., 2007).
In a recent study, lambs were offered all possible 2-way and a 3-way choices among sainfoin, birdsfoot trefoil and alfalfa. Animals selected these legumes in a 70:30 and 50:35:15 ratio for binary and trinary combinations, respectively (Lagrange and Villalba, 2019). Lambs showed highest preference for alfalfa, intermediate for sainfoin and lowest for birdsfoot trefoil. Subsequently, the in vitro ruminal degradability and gas production kinetics of different mixtures of the same legumes were assessed using the gas production technique. The proportions in the mixtures represented: (1) those selected by lambs in the previously described free-choice study (Lagrange and Villalba, 2019); (2) equal proportions (50:50 or 33:33:33 ratios for binary or trinary mixtures, respectively); and, (3) single legumes. The proportion selected by lambs exhibited greater gas production rates than equal parts mixtures (i.e., indifferent selection), and similar to alfalfa, the single forage that exhibited the greatest gas production rates (Lagrange et al., 2019). Thus, lambs built diverse diets that maintained fermentability values as high as pure alfalfa while ingesting a diverse diet with some bioactives (e.g., condensed tannins) with benefits to the internal and external environment such as reduced bloat and ammonia formation, as well as the described advantages related to dietary diversity and amelioration in sensory-specific satiety. More studies like this one may contribute to generate a knowledge base that allows for the construction of diverse and chemically functional pasturelands that enhance animal performance and welfare while reducing environmental impacts.
Species Spatial Arrangement
Forage species in diverse landscapes can be arranged in spatially aggregated mixtures or discrete adjacent monocultures. Both arrangements present advantages and disadvantages. Mixtures may allow for the expression of niche differentiation and plant positive interactions (Tilman et al., 2001; Isbell et al., 2009; Clemensen et al., 2017), but hinder the maintenance of stable pasture composition (Sanderson et al., 2007) (although see next section Grazing Management), and the application of species-specific management like fertilization and weed control. As food preference in herbivores is not random, time is lost while animals search for and handle preferred food items in a finely grained mix of forage species. These activities inevitably reduce harvest efficiency with declines in forage intake and increases in grazing times (Chapman et al., 2007). The potential advantages and disadvantages of mixtures become potential disadvantages and advantages, respectively, in spatially segregated and adjacent monocultures. Studies offering animals the choice of alternative forage species such as ryegrass and white clover growing side-by-side, rather than sown as a conventional intermingled mixture, have provided evidence that animal performance benefits from the patchy spatial arrangement (Nuthall et al., 2000; Cosgrove et al., 2001). When grass and clover are planted in strips, as opposed to homogeneous mixtures, intake of forage by sheep increased by 25% (265 g/sheep/d) and milk production by dairy cows increased by 11% (2.4 kg/cow/d) (Cosgrove et al., 2001). In contrast, per cow milk production in early lactation was similar between diverse forages mixture and spatial adjacent monocultures (Pembleton et al., 2016). These differences may be related to the foraging costs of handling and searching for preferred pasture species in mixtures (Thornley et al., 1994). It is advantageous for ruminants to forage on patches when the preferred vegetation is aggregated as handling and searching activities are lower than when plants are intermingled in a mix (Dumont et al., 2002). When searching costs are low, because preferred plant species are abundant and can be encountered frequently and/or the spatial scale of separation among species facilitate finding, total forage intake in mixtures and spatial adjacent monocultures should be similar (Dumont et al., 2002). For example, a critical spatial scale of separation of grass and clover of 12–36 cm prevents beef heifers incurring selection costs (Rutter et al., 2005). In addition, planting forages in strips overcomes many difficulties inherent in establishing and maintaining mixed pastures, and also mimics what happens naturally as different plant species aggregate in response to environmental conditions (Chapman et al., 2007).
Trade-offs between spatially aggregated mixtures or segregated and adjacent monocultures in plant species interactions and plant-animal interactions can be controlled to some extent by modifying the spatial separation between monocultures, from narrow to wider strips (Sharp et al., 2014). Narrow strips may facilitate forage species establishment and allow the expression of plant complementarity (in the use of plant-growing resources) and plant positive interactions, while reducing searching foraging cost and maintaining high daily forage intake. The relative scale for “narrow” or “wide” depends on the size of the ruminant. For instance, spatial foraging strategies by sheep and cattle differ, with larger-sized cattle exhibiting a much coarser-grained use of a diverse landscape, and lower levels of patch selectivity when patches become smaller than 10 m2 (Laca et al., 2010).
As part of the spatial arrangement of diverse agricultural pastures, it may be beneficial to establish monoculture strips of plant species containing specific bioactive compounds. Forb and shrub species contain PSMs with varied properties for the health and wellbeing of ruminants and positive effects to the environment (Vercoe et al., 2009; Monjardino et al., 2010). For instance, some shrubs and forbs have anthelmintic properties (Kotze et al., 2009), influence nitrogen metabolism with benefits to the animal and the environment (Patra, 2010), attenuate or prevent toxicity by molecular binding (Rogosic et al., 2006, 2007), represent a significant source of vitamins (Salem et al., 2010), have a positive effect on gut function (Vercoe et al., 2009) and counteract bloat caused by legumes like alfalfa by forming stable complexes with soluble protein in the rumen (McMahon et al., 2000). In addition, shrubs contribute to extend the grazing season and tolerate grazing during extended dry periods and in marginal soils, since they provide green and bioactive edible plant material where a ‘feed gap’ would otherwise exist (Emms et al., 2013). Woody species could also be included in diverse pasture systems as live fences, used in agroforestry for biodiversity conservation because they supply habitat for native species and increase connectivity in the landscape (Pulido-Santacruz and Renjifo, 2011). Finally, shrubs contribute to the structural diversity of the vegetation, which is significant for the maintenance of habitats for terrestrial wildlife species in agricultural landscapes (Sullivan and Sullivan, 2006). With appropriate training, ruminants can learn the use of species with bioactive compounds that cause positive post-ingestive consequences (Wallis et al., 2014).
Grazing Management
The mixture ryegrass-clover is one of the most commonly used in temperate environments around the world. However, clover presents limitations due to its low proportion in the pasture mix (typically < 0.20), its patchy spatial distribution and temporal variability (Chapman et al., 1996; Edwards et al., 1996; Fothergill et al., 1996). These limitations have been attributed to the metabolic costs of nitrogen fixation, interspecific competition, high preference by grazing animals and patchy dung and urine deposition, characteristics that are linked at least in part with grazing management of the mixture. In fact, grazing management and climatic conditions are the key factors that more strongly influence grazing system productivity (Tracy et al., 2018). Continuous selective grazing has been claimed as the main cause of reduced abundance or disappearance of preferred species in pasturelands (Parsons et al., 1991).
In contrast to continuous stocking, intensively adaptive management of rotational grazing, through reducing selective defoliation of the most preferred species, may contribute to stabilize the botanical composition of diverse agricultural pastures. In this type of grazing management, there is a tight control on stocking density, residency time and rest period in each grazed area, which contributes to the persistence of the different species in the plant community (Teague et al., 2011). These controls point to shorten the residency time to avoid grazing the regrowth, and to provide forage species with an adequate rest period for full physiological recovery. In addition, targeted animal rotations among different forages may allow for synergies among species as the sequence of forage ingestion influences intake and interactions among nutrients and PSMs with implications for animal health and performance (Mote et al., 2008; Lyman et al., 2011). Rotational grazing has been claimed to reduce per-animal production because it limits selective grazing (Briske et al., 2008); however, in diverse agricultural pastures composed by species that complement each other in primary and secondary metabolites, limitations in grazing selectivity, as discussed in section Diverse Diets: Nutrients, Plant Secondary Metabolites and Flavors, should not be expected to negatively impact animal production on a per animal basis.
Conclusions
Much of the research effort on diverse agricultural pastures has been limited to measure pasture and animal productive responses. Following a more holistic approach, our review contributes to provide new insights into pasture-base ruminant production systems through expanding the conception of the role of plant and phytochemical diversity on animal function and environmental impact. What emerged as critical consideration in the creation of diverse phytochemical pastures for proper animal function and environmental care is the incorporation of genotypes with primary and secondary bioactive metabolites that either by themselves or through their interactions improve nutrition, welfare and health of ruminants, whilst reducing negative environmental impact.
Given our current state of knowledge, it can justifiably be argued that plant diversity and the inherent phytochemical diversity represent a fundamental biological need for efficiency of nutrient use, animal performance, welfare and health (Figure 1). Thus, the final goal of diverse functional pasturelands is the design of more productive and stable plant communities with appropriate associations that promote synergies and complementarities among forage species that enhance these services. Management efforts in the design of diverse pasturelands up to the present have been limited to combination of grasses, legumes and forbs from “traditional” species that historically have been used in grazing systems. The challenge ahead entails the design of diverse agricultural pastures by selecting species identities from a wider array of forage options, exploring “non-traditional” forage species like cicer milkvetch (Astragalus cicer L.), small burnet (Sanguisorba minor), hairy vetch (Vicia villosa Roth), forage radish (Raphanus sativus L.), safflower (Carthamus tinctorius) (Meccage et al., 2019; Stewart et al., 2019; Roca-Fernández et al., 2020) and shrubs like saltbrush (Atriplex spp.) (Pearce et al., 2010), Leucaena leucocephala, Guazuma ulmifolia (Casanova-Lugo et al., 2014), or Tricomaria usillo, and Mimosa ephedroides (Egea et al., 2016). Even if these species represent a small component of the diet, the provision of plant bioactives or other nutrients (i.e., vitamins, minerals, aromas) to the internal environment may represent a significant contribution to the nutrition, welfare, and health of the animal.
A broader knowledge base regarding nutrient-nutrient, PSMs-PSMs and nutrient-PSMs interactions is needed to build the “puzzle” of species selection to be grazed not only in a spatial but also in a temporal scale, understanding potential synergisms and complementarities. The concept of developing a forage chain (Garcia et al., 2008; Cosentino et al., 2014) could be applied to the design of new multifunctional pasture communities. Forage chains provide forage at its peak of production and nutritive value to livestock over an extended period of time to take advantage of the natural growth-distribution differences existing among forage species and varieties. In this new scenario, emphasis should be given not only to biomass production and quality, but also on the presence of PSMs, flavors and nutrients that effectively complement a grazing cycle from rotations across forages at temporal scales shorter than days or seasons. Experimental evidence shows that the sequence of forage ingestion in grazing rotations that entail short periods (just hours) influence forage intake and preference in sheep and cattle (Mote et al., 2008; Lyman et al., 2011). Temporal rotations among diverse forage species are being applied successfully by sheepherders in France to create meal sequences during the day to stimulate intake and optimize use of forage diversity on rangelands (Meuret and Provenza, 2015).
A knowledge base of interactions among traditional and non-traditional species should be implemented by scaling up from in vitro tests to modeling and computational predictive methods, to controlled feeding trials (i.e., cut-and-carry approaches) (e.g., Lagrange and Villalba, 2019), and then to small scale and then large scale field trials (e.g., Lagrange et al., 2020; Figure 1). In addition to species identity, proportion and spatial arrangement of species need to be planned based on the available knowledge of the morphology, ecophysiology, biochemistry and preference of forage species under grazing conditions, as well as on the allometry of the animal species that will graze the forage mix. In the end, all these principles will allow for the design of functional and stable diverse pasturelands. Finally, but not less important, adaptive management of the forage resource through controlled grazing and monitoring would contribute to the persistence and productivity of the newly designed diverse agricultural pastures.
Author Contributions
RD and JV conceived the review and acted as leading authors. JA and SL brought in specific expertise and contributed to the writing of the manuscript. All authors contributed to the article and approved the submission version.
Funding
This review was supported by the United States Department of Agriculture, National Institute of Food and Agriculture (NIFA Award No. 2016-67019-25086), and the Utah Agricultural Experiment Station (Project No.1321), Utah State University, and approved as journal paper number 9379.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Agreil, C., and Meuret, M. (2004). An improved method for quantifying intake rate and ingestive behaviour of ruminants in diverse and variable habitats using direct observation. Small Rumin. Res. 54, 99–113. doi: 10.1016/j.smallrumres.2003.10.013
Amit, M., Cohen, I., Marcovics, A., Muklada, H., Glasser, T. A., Ungar, E. D., et al. (2013). Self-medication with tannin-rich browse in goats infected with gastro-intestinal nematodes. Vet. Parasitol. 198, 305–311. doi: 10.1016/j.vetpar.2013.09.019
Arceo-Medina, G. N., Rosado-Aguilar, J. A., Rodríguez-Vivas, R. I., and Borges-Argaez, R. (2016). Synergistic action of fatty acids, sulphides and stilbene against acaricide-resistant Rhipicephalus microplus ticks. Vet. Parasitol. 228, 121–125. doi: 10.1016/j.vetpar.2016.08.023
Askar, A. R., Guada, J. A., González, J. M., de Vega, A., and Castrillo, C. (2006). Diet selection by growing lambs offered whole barley and a protein supplement, free choice: effects on performance and digestion. Livest. Sci. 101, 81–93. doi: 10.1016/j.livprodsci.2005.09.012
Atwood, S. B., Provenza, F. D., Wiedmeier, R. D., and Banner, R. E. (2001). Changes in preferences of gestating heifers fed untreated or ammoniated straw in different flavors. J. Anim. Sci. 79, 3027–3033. doi: 10.2527/2001.79123027x
Aufrère, J., Dudilieu, M., Andueza, D., Poncet, C., and Baumont, R. (2013). Mixing sainfoin and lucerne to improve the feed value of legumes fed to sheep by the effect of condensed tannins. Animal 7, 82–92. doi: 10.1017/S1751731112001097
Aufrère, J., Dudilieu, M., Poncet, C., and Baumont, R. (2005). “Effect of condensed tannins in sainfoin on in vitro protein solubility of Lucerne,” in Grasslands–a global resource, eds F. P. O'Mara, R. J. Wilkins, L. 't Mannetje, D. K. Lovett, P. A. M. Rogers, and T. M. Boland (Dublin: University College Dublin), 248.
Baraza, E., Hádar, J. A., and Zamora, R. (2009). Consequences of plant-chemical diversity for domestic goat food preference in Mediterranean forests. Acta Oecol. 35, 117–127. doi: 10.1016/j.actao.2008.09.001
Baraza, E., Villalba, J. J., and Provenza, F. D. (2005). Nutritional context influences preferences of lambs for foods with plant secondary metabolites. Appl. Anim. Behav. Sci. 92, 293–305. doi: 10.1016/j.applanim.2004.11.010
Beale, P. K., Marsh, K. J., Foley, W. J., and Moore, B. D. (2018). A hot lunch for herbivores: physiological effects of elevated temperatures on mammalian feeding ecology. Biol. Rev. 93, 674–692. doi: 10.1111/brv.12364
Beauchemin, K. A., Henry Janzen, H., Little, S. M., McAllister, T. A., and McGinn, S. M. (2010). Life cycle assessment of greenhouse gas emissions from beef production in western Canada: a case study. Agric. Syst. 103, 371–379. doi: 10.1016/j.agsy.2010.03.008
Beck, M. R., and Gregorini, P. (2020). How dietary diversity enhances hedonic and eudaimonic well-being in grazing ruminants. Front. Vet. Sci.7:191. doi: 10.3389/fvets.2020.00191
Blümmel, M., Cone, J. W., Van Gelder, A. H., Nshalai, I., Umunna, N. N., Makkar, H. P. S., et al. (2005). Prediction of forage intake using in vitro gas production methods: comparison of multiphase fermentation kinetics measured in an automated gas test, and combined gas volume and substrate degradability measurements in a manual syringe system. Anim. Feed Sci. Technol. 123, 517–526. doi: 10.1016/j.anifeedsci.2005.04.040
Blümmel, M., Steingass, H., and Becker, K. (1997). The relationship between in vitro gas production, in vitro microbial biomass yield and 15N incorporation and its implications for the prediction of voluntary feed intake of roughages. Br. J. Nutr. 77, 911–921. doi: 10.1079/BJN19970089
Brink, G. E., Sanderson, M. A., and Casler, M. D. (2015). Grass and legume effects on nutritive value of complex forage mixtures. Crop Sci. 55, 1329–1337. doi: 10.2135/cropsci2014.09.0666
Briske, D. D., Derner, J. D., Brown, J. R., Fuhlendorf, S. D., Teague, W. R., Havstad, K. M., et al. (2008). Rotational grazing on rangelands: reconciliation of perception and experimental evidence. Rangeland Ecol. Manag. 61, 3–17. doi: 10.2111/06-159R.1
Bryant, R. H., Welten, B., Costall, D., Shorten, P. R., and Edwards, G. R. (2018). Milk yield and urinary-nitrogen excretion of dairy cows grazing forb pasture mixtures designed to reduce nitrogen leaching. Livest. Sci. 209, 46–53. doi: 10.1016/j.livsci.2018.01.009
Bulusu, K. C., Guha, R., Mason, D. J., Lewis, R. P., Muratov, E., Motamedi, Y. K., et al. (2016). Modelling of compound combination effects and applications to efficacy and toxicity: state-of-the-art, challenges and perspectives. Drug Discov. Today 21, 225–238. doi: 10.1016/j.drudis.2015.09.003
Buxton, D. R. (1996). Quality-related characteristics of forages as influenced by plant environment and agronomic factors. Anim. Feed Sci. Tech. 59, 37–49. doi: 10.1016/0377-8401(95)00885-3
Buxton, D. R., and Fales, S. L. (1994). “Plant environment and quality,” in Forage Quality, Evaluation, and Utilization, ed G. C. Fahey Jr. (Madison, WI: American Society of Agronomy), 155–199. doi: 10.2134/1994.foragequality.c4
Casanova-Lugo, F., Petit-Aldana, J., Solorio-Sánchez, F. J., Parsons, D., and Ramírez-Avilés, L. (2014). Forage yield and quality of Leucaena leucocephala and Guazuma ulmifolia in mixed and pure fodder banks systems in Yucatan, Mexico. Agroforest. Syst. 88, 29–39. doi: 10.1007/s10457-013-9652-7
Catanese, F., Distel, R. A., Provenza, F. D., and Villalba, J. J. (2012). Early experience with diverse foods increases intake of nonfamiliar flavors and feeds in sheep. J. Anim. Sci. 90, 2763–2773. doi: 10.2527/jas.2011-4703
Catanese, F., Obelar, M., Villalba, J. J., and Distel, R. A. (2013). The importance of diet choice on stress-related responses by lambs. Appl. Anim. Behav. Sci. 148, 37–45. doi: 10.1016/j.applanim.2013.07.005
Chail, A., Legako, J. F., Pitcher, L. R., Ward, R. E., Martini, S., and MacAdam, J. W. (2017). Consumer sensory evaluation and chemical composition of beef gluteus medius and triceps brachii steaks from cattle finished on forage or concentrate diets. J. Anim. Sci. 95, 1553–1564. doi: 10.2527/jas.2016.1150
Chapman, D. F., Parsons, A. J., Cosgrove, G. P., Barker, D. J., Marotti, D. M., Venning, K. J., et al. (2007). Impacts of spatial patterns in pasture on animal grazing behavior, intake, and performance. Crop Sci. 47, 399–415. doi: 10.2135/cropsci2006.01.0036
Chapman, D. F., Parsons, A. J., and Schwinning, S. (1996). “Management of clover in grazed pastures: Expectations, limitations and opportunities,” in White Clover: New Zealand's competitive edge, ed D. R. Woodfield (Palmerston North: Agron. Soc. N. Z. Spec. Pub. No. 11 Grassl. Res. and Practice Series No. 6), 55–64.
Clemensen, A. K., Provenza, F. D., Lee, S. T., Gardner, D. R., Rottinghaus, G. E., and Villalba, J. J. (2017). Plant secondary metabolites in alfalfa, birdsfoot trefoil, reed canarygrass, and tall fescue unaffected by two different nitrogen sources. Crop Sci. 57, 964–970. doi: 10.2135/cropsci2016.08.0680
Clemensen, A. K., Rottinghaus, G. E., Lee, S. T., Provenza, F. D., and Villalba, J. J. (2018). How planting configuration influences plant secondary metabolites and total N in tall fescue (Festuca arundinacea Schreb.), alfalfa (Medicago sativa L.) and birdsfoot trefoil (Lotus corniculatus L.): implications for grazing management. Grass Forage Sci.73, 94–100. doi: 10.1111/gfs.12298
Clemensen, A. K., Villalba, J. J., Rottinghaus, G. E., Lee, S. T., Provenza, F. D., and Reeve, J. R. (2020). Do plant secondary metabolite-containing forages influence soil processes in pasture systems? Agron. J. 112, 3744–3757. doi: 10.1002/agj2.20361
Cobellis, G., Trabalza-Marinucci, M., and Yu, Z. (2016). Critical evaluation of essential oils as rumen modifiers in ruminant nutrition: a review. Sci. Total Environ. 545, 556–568. doi: 10.1016/j.scitotenv.2015.12.103
Coleman, D. C., Callaham, M. A. Jr., and Crossley, D. A. Jr. (2017). Fundamentals of Soil Ecology, 3rd Edn. San Diego, CA: Academic Press.
Copani, G., Hall, J. O., Miller, J., Priolo, A., and Villalba, J. J. (2013). Plant secondary compounds as complementary resources: are they always complementary? Oecologia 172, 1041–1049. doi: 10.1007/s00442-012-2551-1
Cosentino, S. L., Gresta, F., and Testa, G. (2014). Forage chain arrangement for sustainable livestock systems in a Mediterranean area. Grass Forage Sci. 69, 625–634. doi: 10.1111/gfs.12085
Cosgrove, G. P., Parsons, A. J., Marotti, D. M., Rutter, S. M., and Chapman, D. F. (2001). Opportunities for enhancing the delivery of novel forage attributes. Proc. N. Z. Soc. Anim. Prod. 61, 16–19.
Cottingham, K. L., Brown, B. L., and Lennon, J. T. (2001). Biodiversity may regulate the temporal variability of ecological systems. Ecol. Lett. 4, 72–85. doi: 10.1046/j.1461-0248.2001.00189.x
Cozier, A., Clifford, M. N., and Ashihara, H. (2006). Plant Secondary Metabolites: Occurrence, Structure and Role in the Human Diet. Oxford: Blackwell Publishing. doi: 10.1002/9780470988558
Craine, J. M., Angerer, J. P., Elmore, A., and Fierer, N. (2016). Continental-scale patterns reveal potential for warming-induced shifts in cattle diet. PLoS ONE 11:e0161511. doi: 10.1371/journal.pone.0161511
Craine, J. M., Tilman, D. G., Wedin, D. A., Reich, P., Tjoelker, M., and Knops, J. (2002). Functional traits, productivity and effects on nitrogen cycling of 33 grassland species. Funct. Ecol. 16, 563–574. doi: 10.1046/j.1365-2435.2002.00660.x
de Vries, M., and de Boer, I. J. M. (2010). Comparing environmental impacts for livestock products: a review of life cycle assessments. Livest. Sci.128, 1–11. doi: 10.1016/j.livsci.2009.11.007
de Vries, M., van Middelaar, C. E., and de Boer, I. J. M. (2015). Comparing environmental impacts of beef production systems: a review of life cycle assessments. Livest. Sci. 178, 279–288. doi: 10.1016/j.livsci.2015.06.020
Deak, A., Hall, M. H., Sanderson, M. A., and Archibald, D. D. (2007). Production and nutritive value of grazed simple and complex forage mixtures. Agron. J. 99, 814–821. doi: 10.2134/agronj2006.0166
Dearing, M. D., Foley, W. J., and McLean, S. (2005). The influence of plant secondary metabolites on the nutritional ecology of herbivorous terrestrial vertebrates. Ann. Rev. Ecol. Evol. Syst. 36, 169–189. doi: 10.1146/annurev.ecolsys.36.102003.152617
Dearing, M. D., Mangione, A. M., and Karasov, W. H. (2000). Diet breadth of mammalian herbivores: nutrient vs. detoxification constraints. Oecologia 123, 397–405. doi: 10.1007/s004420051027
Deinum, B., and Knoppers, J. (1979). The growth of maize in the cool temperate climate of the Netherlands: effect of grain filling on production of dry matter and on chemical composition and nutritive value. NJAS-Wagen. J. Life Sci. 27, 116–130. doi: 10.18174/njas.v27i2.17059
Delgado-Baquerizo, M., Reich, P. B., Trivedi, C., Eldridge, D. J., Abades, S., Alfaro, F. D., et al. (2020). Multiple elements of soil biodiversity drive ecosystem functions across biomes. Nat. Ecol. Evol. 4, 210–220. doi: 10.1038/s41559-019-1084-y
Di, H. J., and Cameron, K. C. (2016). Inhibition of nitrification to mitigate nitrate leaching and nitrous oxide emissions in grazed grassland: a review. J. Soils Sediment. 16, 1401–1420. doi: 10.1007/s11368-016-1403-8
Diaz, S., Cabido, M., and Casanoves, F. (1998). Plant functional traits and environmental filters at a regional scale. J. Veg. Sci. 9, 113–122. doi: 10.2307/3237229
Distel, R. A., Rodríguez Iglesias, R. M., Arroquy, J. I., and Merino, J. (2007). A note on increased intake in lambs through diversity in food flavor. Appl. Anim. Behav. Sci. 105, 232–237. doi: 10.1016/j.applanim.2006.06.002
Donnelly, E. D., Anthony, W. B., and Langford, J. W. (1971). Nutritive relationships in low-and high-tannin Sericea lespedeza under grazing. Agron. J. 63, 749–751. doi: 10.2134/agronj1971.00021962006300050027x
Dumont, B., Carrère, P., and D'Hour, P. (2002). Foraging in patchy grasslands: diet selection by sheep and cattle is affected by the abundance and spatial distribution of preferred species. Anim. Res. 51, 367–381. doi: 10.1051/animres:2002033
Edwards, G. R., Parsons, A. J., Newman, J. A., and Wright, I. A. (1996). The spatial pattern of vegetation in cut and grazed grass/white clover pastures. Grass Forage Sci. 51, 219–231. doi: 10.1111/j.1365-2494.1996.tb02057.x
Egea, A. V., Allegretti, L. I., Lama, S. P., Grilli, D., Fucili, M., Guevara, J. C., et al. (2016). Diet mixing and condensed tannins help explain foraging preferences by Creole goats facing the physical and chemical diversity of native woody plants in the central Monte desert (Argentina). Anim. Feed Sci. Tech. 215, 47–57. doi: 10.1016/j.anifeedsci.2016.02.021
Eisenhauer, N., Hines, J., Isbell, F., van der Plas, F., Hobbie, S. E., Kazanski, C. E., et al. (2018). Plant diversity maintains multiple soil functions in future environments. Elife 7:e41228. doi: 10.7554/eLife.41228.020
Eisenhauer, N., Lanoue, A., Strecker, T., Scheu, S., Steinauer, K., Thakur, M. P., et al. (2017). Root biomass and exudates link plant diversity with soil bacterial and fungal biomass. Sci. Rep. 7:44641. doi: 10.1038/srep44641
Emms, J. E., Vercoe, P. E., Hughes, S., Jessop, P., Norman, H. C., Kilminster, T., et al. (2013). “Making decisions to identify forage shrub species for versatile grazing systems,” in Proceedings of the 22nd International Grassland Congress, eds D. L. Michalk, G. D. Millar, W. B. Badgery, and K. M. Broadfoot (Sydney: International Grasslands Organization), 1372–1373.
Engel, C. (2003). Wild Health: Lessons in Natural Wellness from the Animal Kingdom. Boston, MA: Houghton Mifflin Co.
Ergon, Å., Seddaiu, G., Korhonen, P., Virkajärvi, P., Bellocchi, G., Jørgensen, M., et al. (2018). How can forage production in Nordic and Mediterranean Europe adapt to the challenges and opportunities arising from climate change? Eur. J. Agron. 92, 97–106. doi: 10.1016/j.eja.2017.09.016
Evers, J. B., van der Werf, W., Stomph, T. J., Bastiaans, L., and Anten, N. P. R. (2019). Understanding and optimizing species mixtures using functional-structural plant modelling J. Exp. Bot. 70, 2381–2388. doi: 10.1093/jxb/ery288
Feng, C., Ding, S. W., Zhang, T. Y., Li, Z. Q., Wang, D. L., Wang, L., et al. (2016). High plant diversity stimulates foraging motivation in grazing herbivores. Basic Appl. Ecol. 17, 43–51. doi: 10.1016/j.baae.2015.09.004
Fernández, H. T., Catanese, F., Puthod, G., Distel, R. A., and Villalba, J. J. (2012). Depression of rumen ammonia and blood urea by quebracho tannin-containing supplements fed after high-nitrogen diets with no evidence of self-regulation of tannin intake by sheep. Small Rumin. Res. 105, 126–134. doi: 10.1016/j.smallrumres.2012.03.013
Fothergill, M., Davies, D. A., Morgan, C. T., and Jones, J. R. (1996). “White clover crashes,” in Legumes in Sustainable Farming Systems, ed D. Younie (Reading: British Grassland Society), 172–176.
Franzke, A., Unsicker, S. B., Specht, J., Koehler, G., and Weisser, W. W. (2010). Being a generalist herbivore in a diverse world: how do diets from different grasslands influence food plant selection and fitness of the grasshopper Chorthippus parallelus? Ecol. Entomol. 35,126–138. doi: 10.1111/j.1365-2311.2009.01168.x
Freeland, W. J., Calcott, P. H., and Anderson, L. R. (1985). Tannins and saponin: interaction in herbivore diets. Biochem. Syst. Ecol. 13, 189–193. doi: 10.1016/0305-1978(85)90078-X
Freeland, W. J., and Janzen, D. H. (1974). Strategies in herbivory by mammals: the role of plant secondary compounds. Am. Nat. 108, 269–289. doi: 10.1086/282907
Friend, M. A., Provenza, F. D., and Villalba, J. J. (2015). Preference by sheep for endophyte-infected tall fescue grown adjacent to or at a distance from alfalfa. Animal 9, 516–525. doi: 10.1017/S1751731114002559
Garcia, S. C., Fulkerson, W. J., and Brookes, S. U. (2008). Dry matter production, nutritive value, and efficiency of nutrient utilization of a complementary forage rotation compared to a grass pasture system. Grass Forage Sci. 63, 284–300. doi: 10.1111/j.1365-2494.2008.00636.x
Ginane, C., Bonnet, M., Baumont, R., and Revell, D. K. (2015). Feeding behaviour in ruminants: a consequence of interactions between a reward system and the regulation of metabolic homeostasis. Anim. Prod. Sci. 55, 247–260. doi: 10.1071/AN14481
Githiori, J. B., Athanasiadou, S., and Thamsborg, S. M. (2006). Use of plants in novel approaches for control of gastrointestinal helminths in livestock with emphasis on small ruminants. Vet. Parasitol. 139, 308–320. doi: 10.1016/j.vetpar.2006.04.021
Glasser, T. A., Ungar, E. D., Landau, S. Y., Perevolotsky, A., Muklada, H., and Walker, J. W. (2009). Breed and maternal effects on the intake of tannin-rich browse by juvenile domestic goats (Capra hircus). Appl. Anim. Behav. Sci.119, 71–77. doi: 10.1016/j.applanim.2009.02.028
Gordon, I. J., Pérez-Barbería, F. J., and Cuartas, P. (2002). The influence of adaptation of rumen microflora on in vitro digestion of different forages by sheep and reed deer. Can. J. Zool. 80, 1930–1937. doi: 10.1139/z02-179
Gourlay, G., and Constabel, C. P. (2019). Condensed tannins are inducible antioxidants and protect hybrid poplar against oxidative stress. Tree Physiol. 39, 345–355. doi: 10.1093/treephys/tpy143
Groot, J. C. J., Cone, J. W., Williams, B. A., Debersaques, F. M. A., and Lantinga, E. A. (1996). Multiphasic analysis of gas production kinetics for in vitro fermentation of ruminant feeds. Anim. Feed Sci. Technol. 64, 77–89. doi: 10.1016/S0377-8401(96)01012-7
Grosse Brinkhaus, A., Bee, G., Silacci, P., Kreuzer, M., and Dohme-Meier, F. (2016). Effect of exchanging Onobrychis viciifolia and Lotus corniculatus for Medicago sativa on ruminal fermentation and nitrogen turnover in dairy cows. J. Dairy Sci. 99, 4384–4397. doi: 10.3168/jds.2015-9911
Hall, M. B., Hoover, W. H., Jennings, J. P., and Webster, T. K. (1999). A method for partitioning neutral detergent-soluble carbohydrates. J. Sci. Food Agric. 79, 2079–2086. doi: 10.1002/(SICI)1097-0010(199912)79:15<2079::AID-JSFA502>3.0.CO;2-Z
Harris, S. L., Clark, D. A., Auldist, M. J., Waugh, C. D., and Laboyrie, P. G. (1997). Optimum white clover content for dairy pastures. Proc. N. Z. Grassl. Assoc. 59, 29–33. doi: 10.33584/jnzg.1997.59.2261
Hautier, Y., Isbell, F., Borer, E. T., Seabloom, E. W., Harpole, W. S., Lind, E. M., et al. (2018). Local loss and spatial homogenization of plant diversity reduce ecosystem multifunctionality. Nat. Ecol. Evol. 2, 50–56. doi: 10.1038/s41559-017-0395-0
Hector, A., Bazeley-White, E., Loreau, M., Otway, S., and Schmidt, B. (2002). Overyielding in grassland communities: testing the sampling effect hypothesis with replicated biodiversity experiments. Ecol. Lett. 5, 502–511. doi: 10.1046/j.1461-0248.2002.00337.x
Hector, A., Schmid, B., Beierkuhnlein, C., Caldeira, M. C., Diemer, M., Dimitrakopoulos, P. G., et al. (1999). Plant diversity and productivity experiments in European grasslands. Science 286, 1123–1127. doi: 10.1126/science.286.5442.1123
Hobbs, R. J., Gross, J. E., Shipley, L. A., Spalinger, D. E., and Wunder, B. A. (2003). Herbivore functional response in heterogeneous environments: a contest among models. Ecology 84, 666–681. doi: 10.1890/0012-9658(2003)084[0666:HFRIHE]2.0.CO;2
Hobbs, R. J., and Morton, S. R. (1999). Moving from descriptive to predictive ecology. Agroforest. Syst. 45, 43–55. doi: 10.1023/A:1006263503404
Hoste, H., Jackson, F., Athanasiadou, S., Thamsborg, S. M., and Hoskin, S. O. (2006). The effects of tannin-rich plants on parasitic nematodes in ruminants. Trends Parasitol. 22, 253–261. doi: 10.1016/j.pt.2006.04.004
Hoste, H., Torres-Acosta, J. F., Sandoval-Castro, C. A., Mueller-Harvey, I., Sotiraki, S., Louvandini, H., et al. (2015). Tannin containing legumes as a model for nutraceuticals against digestive parasites in livestock. Vet. Parasitol. 212, 5–17. doi: 10.1016/j.vetpar.2015.06.026
Huang, T., Gao, B., Hu, X. K., Lu, X., Well, R., Christie, P., et al. (2015). Ammonia-oxidation as an engine to generate nitrous oxide in an intensively managed calcareous Fluvo-aquic soil. Sci. Rep. 4:3950. doi: 10.1038/srep03950
Huffman, M. A. (2003). Animal self-medication and ethno-medicine: exploration and exploitation of the medicinal properties of plants. Proc. Nutr. Soc. 62, 371–381. doi: 10.1079/PNS2003257
Hummelbrunner, L. A., and Isman, M. B. (2001). Acute, sublethal, antifeedant, and synergistic effects of monoterpenoid essential oil compounds on the tobacco cutworm Spodoptera litura (Lep. Noctuidae). J. Agric. Food Chem. 49, 715–720. doi: 10.1021/jf000749t
Hutchings, M. R., Athanasiadou, S., Kyriazakis, I., and Gordon, I. J. (2003). Can animals use foraging behaviour to combat parasites? Proc. Nutr. Soc. 62, 361–370. doi: 10.1079/PNS2003243
Hutchings, M. R., Kyriazakis, I., Papachristou, T. G., Gordon, I. J., and Jackson, F. (2000). The herbivores' dilemma: trade-offs between nutrition and parasitism in foraging decisions. Oecologia 124, 242–251. doi: 10.1007/s004420000367
Illius, A. W., and Jessop, N. S. (1995). Modeling metabolic costs of allelochemical ingestion by foraging herbivores. J. Chem. Ecol. 21, 693–719. doi: 10.1007/BF02033456
Isbell, F., Adler, P. R., Eisenhauer, N., Fornara, D., Kimmel, K., Kremen, C., et al. (2017). Benefits of increasing plant diversity in sustainable agroecosystems. J. Ecol. 105, 871–879. doi: 10.1111/1365-2745.12789
Isbell, F., Craven, D., Connolly, J., Loreau, M., Schmid, B., Beierkuhnlein, C., et al. (2015). Biodiversity increases the resistance of ecosystem productivity to climate extremes. Nature 526, 574–577. doi: 10.1038/nature15374
Isbell, F. I., Polley, H. W., and Wilsey, B. J. (2009). Biodiversity, productivity and the temporal stability of productivity: Patterns and processes. Ecol. Lett. 12, 443–451. doi: 10.1111/j.1461-0248.2009.01299.x
Jacobs, J. L., Hill, J., and Jenkin, T. (2009). Effect of different grazing strategies on dry matter yields and nutritive characteristics of whole crop cereals. Anim. Prod. Sci. 49, 608–618. doi: 10.1071/EA08245
Jing, J., Søegaard, K., Cong, W. F., and Eriksen, J. (2017). Species diversity effects on productivity, persistence and quality of multispecies swards in a four-year experiment. PLoS ONE 12:e0169208. doi: 10.1371/journal.pone.0169208
Johnson, K. A., and Johnson, D. E. (1995). Methane emissions from cattle. J. Anim. Sci. 73, 2483–2492. doi: 10.2527/1995.7382483x
Jonker, A., Farrell, L., Scobie, D., Dynes, R., Edwards, G., Hague, H., et al. (2019). Methane and carbon dioxide emissions from lactating dairy cows grazing mature ryegrass/white clover or a diverse pasture comprising ryegrass, legumes and herbs. Anim. Prod. Sci. 59, 1063–1069. doi: 10.1071/AN18019
Jonker, A., and Yu, P. (2017). The occurrence, biosynthesis, and molecular structure of proanthocyanidins and their effects on legume forage protein precipitation, digestion and absorption in the ruminant digestive tract. Int. J. Mol. Sci. 18:1105. doi: 10.3390/ijms18051105
Jouven, M., Carrère, P., and Baumont, R. (2006). Model predicting dynamics of biomass, structure and digestibility of herbage in managed permanent pastures. 1. Model description. Grass Forage Sci. 61, 112–124. doi: 10.1111/j.1365-2494.2006.00515.x
Juhnke, J., Miller, J., Hall, J. O., Provenza, F. D., and Villalba, J. J. (2012). Preference for condensed tannins by sheep in response to challenge infection with Haemonchus contortus. Vet. Parasitol. 188, 104–114. doi: 10.1016/j.vetpar.2012.02.015
Kakraliya, S. K., Singh, U., Bohra, A., Choudhary, K. K., Kumar, S., Meena, R. S., et al. (2018). “Nitrogen and legumes: a meta-analysis,” in Legumes for Soil Health and Sustainable Management, eds R. S. Meena, A. Das, G.S. Yadav, and R. Lal (Singapore: Springer), 277–314. doi: 10.1007/978-981-13-0253-4_9
Kmicikewycz, A. D., and Heinrichs, A. J. (2015). Effect of corn silage particle size and supplemental hay on rumen pH and feed preference by dairy cows fed high-starch diets. J. Dairy Sci. 98, 373–385. doi: 10.3168/jds.2014-8103
Kotowska, A. M., Cahill, J. F., and Keddie, B. A. (2010). Plant genetic diversity yields increased plant productivity and herbivore performance. J. Ecol. 98, 237–245. doi: 10.1111/j.1365-2745.2009.01606.x
Kotze, A. C., O'Grady, J., Emms, J., Toovey, A. F., Hughes, S., Jessop, P., et al. (2009). Exploring the anthelmintic properties of Australian native shrubs with respect to their potential role in livestock grazing systems. Parasitology 136, 1065–1080. doi: 10.1017/S0031182009006386
Kuchenmeister, K., Kuchenmeister, F., Kayser, M., Wrage-Mönnig, N., and Isselstein, J. (2013). Influence of drought stress on nutritive value of perennial forage legumes. Int. J. Plant Prod. 7, 693–710.
Laca, E. A., Sokolow, S., Galli, J. R., and Cangiano, C. A. (2010). Allometry and spatial scales of foraging in mammalian herbivores. Ecol. Lett. 13, 311–320. doi: 10.1111/j.1461-0248.2009.01423.x
Lagrange, S., Beauchemin, K. A., MacAdam, J., and Villalba, J. J. (2020). Grazing diverse combinations of tanniferous and non-tanniferous legumes: implications for beef cattle performance and environmental impact. Sci. Total Environ. 746:140788. doi: 10.1016/j.scitotenv.2020.140788
Lagrange, S., Lobón, S., and Villalba, J. J. (2019). Gas production kinetics and in vitro degradability of tannin-containing legumes, alfalfa and their mixtures. Anim. Feed Sci. Tech. 253, 56–64. doi: 10.1016/j.anifeedsci.2019.05.008
Lagrange, S., and Villalba, J. J. (2019). Tannin-containing legumes and forage diversity influence foraging behavior, diet digestibility, and nitrogen excretion by lambs. J. Anim. Sci. 97, 3994–4009. doi: 10.1093/jas/skz246
Lehman, C., and Tilman, D. (2000). Biodiversity, stability, and productivity in competitive communities. Am. Nat. 156, 534–552. doi: 10.1086/303402
Lima, P. M. T., Filho, A. L. A., Issakowicz, J., Ieda, E. H., Corrêa, P. S., Mattos, W. T., et al. (2020). Methane emission, ruminal fermentation parameters and fatty acid profile of meat in Santa Inês lambs fed the legume macrotiloma. Anim. Prod. Sci. 60, 665–673. doi: 10.1071/AN19127
Lima, P. M. T., Moreira, G. D., Sakita, G. Z., Natel, A. S., Mattos, W. T., Gimenes, F. M. A., et al. (2018). Nutritional evaluation of the legume Macrotyloma axillare using in vitro and in vivo bioassays in sheep. J. Anim. Physiol. An. N. 102, e669–e676. doi: 10.1111/jpn.12810
Lobley, G. E., and Milano, G. D. (1997). Regulation of hepatic nitrogen metabolism in ruminants. Proc. Nutr. Soc. 56, 547–563. doi: 10.1079/PNS19970057
Louault, F., Pillar, V. D., Aufre‘re, J., Garnier, E., and Soussana, J. F. (2005). Plant traits and functional types in response to reduced disturbance in semi-natural grassland. J. Veg. Sci. 16, 151–160. doi: 10.1111/j.1654-1103.2005.tb02350.x
Lyman, T. D., Provenza, F. D., and Villalba, J. J. (2008). Sheep foraging behavior in response to interactions among alkaloids, tannins and saponins. J. Sci. Food Agric. 88, 824–831. doi: 10.1002/jsfa.3158
Lyman, T. D., Provenza, F. D., Villalba, J. J., and Wiedmeier, R. D. (2011). Cattle preferences differ when endophyte-infected tall fescue, birdsfoot trefoil, and alfalfa are grazed in difference sequences. J. Anim. Sci. 89, 1131–1137. doi: 10.2527/jas.2009-2741
MacAdam, J. W. (2019). “The elevated carbohydrate concentrations of high altitude-grown perennial legume forages,” in Annual Meetings Abstracts. ASA, CSSA, and SSSA (Madison, WI). Available online at: https://scisoc.confex.com/scisoc/2019am/meetingapp.cgi/Paper/119690 (accessed August 10, 2020).
Majak, W., McAllister, T. A., McCartney, D., Stanford, K., and Cheng, K. J. (2003). “Bloat in cattle.,” in Alberta Agric. Food Rural Dev. Information Packaging Centre (Edmonton, AB), 1–24.
Marsh, K. J., Wallis, I. R., Andrew, R. L., and Foley, W. J. (2006). The detoxification limitation hypothesis: where did it come from and where is it going? J. Chem. Ecol. 32, 1247–1266. doi: 10.1007/s10886-006-9082-3
Marzetz, V., Koussoroplis, A., Martin-Creuzburg, D., Striebel, M., and Wacker, A. (2017). Linking primary producer diversity and food quality effects on herbivores: a biochemical perspective. Sci. Rep. 7:11035. doi: 10.1038/s41598-017-11183-3
McAllister, T. A., Martinez, T., Bae, H. D., Muir, A. D., Yanke, L. J., and Jones, J. A. (2005). Characterization of condensed tannins purified from legume forages: chromophore production, protein precipitation, and inhibitory effects on cellulose digestion. J. Chem. Ecol. 31, 2049–2068. doi: 10.1007/s10886-005-6077-4
McMahon, L. R., McAllister, T. A., Berg, B. P., Majak, W., Acharya, S. N., Popp, J. D., et al. (2000). A review of the effects of forage condensed tannins on ruminal fermentation and bloat in grazing cattle. Can. J. Plant Sci. 80, 469–485. doi: 10.4141/P99-050
McNally, S. R., Laughlin, D. C., Rutledge, S., Dodd, M. B., Six, J., and Schipper, L. A. (2015). Root carbon inputs under moderately diverse sward and conventional ryegrass-clover pasture: implications for soil carbon sequestration. Plant Soil 392, 289–299. doi: 10.1007/s11104-015-2463-z
Meccage, E., Carr, P. M., Bourgault, M., McVay, K., and Boss, D. (2019). Potential of annual forages in the Northern Great Plains. Crops Soils 52, 18–22. doi: 10.2134/cs2019.52.0101
Meuret, M., and Bruchou, C. (1994). Modélisation de l'ingestion selon la diversité des choix alimentaires réalisés par la chèvre au pâturage sur parcours. Rencontres Rech. Rumin. 1, 225–228.
Meuret, M., and Provenza, F. (2015). How French shepherds create meal sequences to stimulate intake and optimise use of forage diversity on rangeland. Anim. Prod. Sci. 55, 309–318. doi: 10.1071/AN14415
Monjardino, M., Revell, D., and Pannell, D. J. (2010). The potential contribution of forage shrubs to economic returns and environmental management in Australian dryland agricultural systems. Agr. Syst. 103, 187–197. doi: 10.1016/j.agsy.2009.12.007
Moore, A. D., Donnelly, J. R., and Freer, M. (1997). GRAZPLAN: decision support system for Australian grazing enterprises. III. Pasture growth and soil moisture submodels, and the GrassGro DSS. Agr. Syst. 54, 535–582. doi: 10.1016/S0308-521X(97)00023-1
Moore, B. D., Andrew, R. L., Kühlheim, C., and Foley, W. J. (2014). Explaining intraspecific diversity in plant secondary metabolites in an ecological context. New Phytol. 201, 733–750. doi: 10.1111/nph.12526
Moreno, F. C., Gordon, I. J., Wright, A. D., Benvenutti, M. A., and Saumell, C. A. (2010). In vitro anthelmintic effect of plant extracts against infective larvae of ruminants gastrointestinal nematode parasites. Arch. Med. Vet. 42, 155–163. doi: 10.4067/S0301-732X2010000300006
Mote, T. E., Villalba, J. J., and Provenza, F. D. (2008). Sequence of food presentation influences intake of foods containing tannins and terpenes. Appl. Anim. Behav. Sci. 113, 57–68. doi: 10.1016/j.applanim.2007.10.003
Moya, D., Mazzenga, A., Holtshausen, L., Cozzi, G., González, L. A., Calsamiglia, S., et al. (2011). Feeding behavior and ruminal acidosis in beef cattle offered a total mixed ration or dietary components separately. J. Anim. Sci. 89, 520–530. doi: 10.2527/jas.2010-3045
Mueller-Harvey, I. (2006). Unravelling the conundrum of tannins in animal nutrition and health. J. Sci. Food Agric. 86, 2010–2037. doi: 10.1002/jsfa.2577
Mueller-Harvey, I., Bee, G., Dohme-Meier, F., Hoste, H., Karonen, M., Kölliker, R., et al. (2019). Benefits of condensed tannins in forage legumes fed to ruminants: importance of structure, concentration, and diet composition. Crop Sci. 59, 861–885. doi: 10.2135/cropsci2017.06.0369
Niderkorn, V., Baumont, R., Le Morvan, A., and Macheboeuf, D. (2011). Occurrence of associative effects between grasses and legumes in binary mixtures on in vitro rumen fermentation characteristics. J. Anim. Sci. 89, 1138–1145. doi: 10.2527/jas.2010-2819
Niderkorn, V., Mueller-Harvey, I., Le Morvan, A., and Aufrère, J. (2012). Synergistic effects of mixing cocksfoot and sainfoin on in vitro rumen fermentation. Role of condensed tannins. Anim. Feed Sci. Technol. 178, 48–56. doi: 10.1016/j.anifeedsci.2012.09.014
Nuthall, R., Rutter, S., and Rook, A. (2000). “Milk production by dairy cows grazing mixed swards or adjacent monocultures of grass and white clover,” in Proceedings 6th British Grassland Society Research Conference (Reading: Brit. Grassl. Soc.), 117–118.
Oenema, O., Wrage, N., Velthof, G. L., van Groenigen, J. W., Dolfing, J., and Kuikman, P. J. (2005). Trends in global nitrous oxide emissions from animal production systems. Nutr. Cycl. Agroecosys. 72, 51–65. doi: 10.1007/s10705-004-7354-2
Owens, J., Provenza, F. D., Wiedmeier, R. D., and Villalba, J. J. (2012). Supplementing endophyte infected tall fescue or reed canarygrass with alfalfa or birdsfoot trefoil increases forage intake and digestibility by sheep. J. Sci. Food Agr. 92, 987–992. doi: 10.1002/jsfa.4681
Parsons, A. J., Harvey, A., and Johnson, I. R. (1991). Plant/ animal interactions in continuously grazed mixtures: 2. The role of differences in the physiology of leaf expansion and of selective grazing on the performance and stability of species in a mixture. J. Appl. Ecol. 28, 635–658. doi: 10.2307/2404573
Patra, A. K. (2010). Aspects of nitrogen metabolism in sheep-fed mixed diets containing tree and shrub foliages. Brit. J. Nutr. 103, 1319–1330. doi: 10.1017/S0007114509993254
Patra, A. K., and Saxena, J. (2010). A new perspective on the use of plant secondary metabolites to inhibit methanogenesis in the rumen. Phytochemistry 71, 1198–1222. doi: 10.1016/j.phytochem.2010.05.010
Pearce, K. L., Norman, H. C., and Hopkins, D. L. (2010). The role of saltbush-based pasture systems for the production of high quality sheep and goat meat. Small Ruminant Res. 91, 29–38. doi: 10.1016/j.smallrumres.2009.10.018
Pembleton, K. G., Hills, J. L., Freeman, M. J., McLaren, D. K., French, M., and Rawnsley, R. P. (2016). More milk from forage: milk production, blood metabolites, and forage intake of dairy cows grazing pasture mixtures and spatially adjacent monocultures. J. Dairy Sci. 99, 3512–3528. doi: 10.3168/jds.2015-10542
Phy, T. S., and Provenza, F. D. (1998). Eating barley too frequently or in excess decreases lambs' preference for barley but sodium bicarbonate and lasalocid attenuate the response. J. Anim. Sci. 76, 1578–1583. doi: 10.2527/1998.7661578x
Polley, H. W., Isbell, F. I., and Wilsey, B. J. (2013). Plant functional traits improve diversity-based predictions of temporal stability of grassland productivity. Oikos 122, 1275–1282. doi: 10.1111/j.1600-0706.2013.00338.x
Popović, B. M., Štajner, D., Ždero-Pavlović, R., Tumbas-Šaponjac, V., Canadanović-Brunet, J., and Orlović, S. (2016). Water stress induces changes in polyphenol profile and antioxidant capacity in poplar plants (Populus spp.). Plant Physiol. Biochem. 105, 242–250. doi: 10.1016/j.plaphy.2016.04.036
Preuer, K., Lewis, R. P., Hochreiter, S., Bender, A., Bulusu, K. C., and Klambauer, G. (2018). DeepSynergy: predicting anti-cancer drug synergy with Deep Learning. Bioinformatics 34, 1538–1546. doi: 10.1093/bioinformatics/btx806
Prieto, I., Violle, C., Barre, P., Durand, J. L., Ghesquiere, M., and Litrico, I. (2015). Complementary effects of species and genetic diversity on productivity and stability of sown grasslands. Nat. Plants 1:15033. doi: 10.1038/nplants.2015.33
Priolo, Q., and Vasta, V. (2007). Effects of tannin-containing diets on small ruminant meat quality. Ital. J. Anim. Sci. 6(Suppl. 1), 527–530. doi: 10.4081/ijas.2007.1s.527
Provenza, F. D, Villalba, J. J., Dziba, L. E., Atwood, S. B., and Banner, R. E. (2003). Linking herbivore experience, varied diets, and plant biochemical diversity. Small Rumin. Res. 49, 257–274. doi: 10.1016/S0921-4488(03)00143-3
Provenza, F. D. (1995). Postingestive feedback as an elementary determinant of food preference and intake in ruminants. Rangeland Ecol. Manage. 48, 2–17. doi: 10.2307/4002498
Provenza, F. D. (1996). Acquired aversions as the basis for varied diets of ruminants foraging on rangelands. J. Anim. Sci. 74, 2010–2020. doi: 10.2527/1996.7482010x
Provenza, F. D., Villalba, J. J., Haskell, J., MacAdam, J. W., Griggs, T. C., and Wiedmeier, R. D. (2007). The value to herbivores of plant physical and chemical diversity in time and space. Crop Sci. 47, 382–398. doi: 10.2135/cropsci2006.02.0083
Pulido-Santacruz, P., and Renjifo, L. M. (2011). Live fences as tools for biodiversity conservation: a study case with birds and plants. Agroforest. Syst. 81, 15–30. doi: 10.1007/s10457-010-9331-x
Rapport, D. J. (1980). Optimal foraging for complementary resources. Am. Nat. 116, 324–346. doi: 10.1086/283631
Reddy, A., Chaitanya, K., and Vivekanandan, M. (2004). Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. J. Plant Physiol. 161, 1189–1202. doi: 10.1016/j.jplph.2004.01.013
Richardson, J. M., Wilkinson, R. G., and Sinclair, L. A. (2003). Synchrony of nutrient supply to the rumen and dietary energy source and their effects on the growth and metabolism of lambs. J. Anim. Sci. 81, 1332–1347. doi: 10.2527/2003.8151332x
Rivero, M. J., Keim, J. P., Balocchi, O. A., and Lee, M. R. F. (2020). In vitro fermentation patterns and methane output of perennial ryegrass differing in water-soluble carbohidrates and nitrogen concentration. Animals 10:1076. doi: 10.3390/ani10061076
Roca-Fernández, A. I., Dillard, S. L., and Soder, K. J. (2020). Ruminal fermentation and enteric methane production of legumes containing condensed tannins fed in continuous culture. J. Dairy Sci. 103, 7028–7038. doi: 10.3168/jds.2019-17627
Rochfort, S., Parker, A. J., and Dunshea, F. R. (2008). Plant bioactives for ruminant health and productivity. Phytochemistry 69, 299–322. doi: 10.1016/j.phytochem.2007.08.017
Rogosic, J., Estell, R. E., Skobic, D., Martinovic, A., and Maric, S. (2006). Role of species diversity and secondary compound complementarity on diet selection of Mediterranean shrubs by goats. J. Chem. Ecol. 32, 1279–1287. doi: 10.1007/s10886-006-9084-1
Rogosic, J., Estell, R. E., Skobic, D., and Stanic, S. (2007). Influence of secondary compound complementarity and species diversity on consumption of Mediterranean shrubs by sheep. Appl. Anim. Behav. Sci. 107, 58–65. doi: 10.1016/j.applanim.2006.09.013
Rolls, E. T., Murzi, E., Yaxley, S., Thorpe, S. J., and Simpson, S. J. (1986). Sensory-specific satiety: food-specific reduction in responsiveness of ventral forebrain neurons after feeding in the monkey. Brain Res. 368, 79–86. doi: 10.1016/0006-8993(86)91044-9
Romera, A. J., Doole, G. J., Beukes, P. C., Mason, N., and Mudge, P. L. (2017). The role and value of diverse sward mixtures in dairy farm systems of New Zealand: an exploratory assessment. Agr. Syst. 152, 18–26. doi: 10.1016/j.agsy.2016.12.004
Rotz, C. A., Asem-Hiablie, S., Place, S., and Thoma, G. (2019). Environmental footprints of beef cattle production in the United States. Agr. Syst. 169, 1–13. doi: 10.1016/j.agsy.2018.11.005
Rutter, S. M. (2006). Diet preference for grass and legumes in free-ranging domestic sheep and cattle: current theory and future application. Appl. Anim. Behav. Sci. 97, 17–35. doi: 10.1016/j.applanim.2005.11.016
Rutter, S. M., Cook, J. E., Young, K. L., and Champion, R. A. (2005). “Spatial scale of heterogeneity affects diet choice but not intake in beef cattle,” in Pastoral Systems in Marginal Environments, ed J. A. Milne (Wageningen: Wageningen Academic Publishers), 127.
Salem, H. B., Norman, H. C., Nefzaoui, A., Mayberry, D. E., Pearce, K. L., and Revell, D. K. (2010). Potential use of oldman saltbush (Atriplex nummularia Lindl.) in sheep and goat feeding. Small Rumin. Res. 91, 3–28. doi: 10.1016/j.smallrumres.2009.10.017
Sanderson, M. A., Goslee, S. C., Soder, K. J., Skinner, R. H., Tracy, B. F., and Deak, A. (2007). Plant species diversity, ecosystem function, and pasture management: a perspective. Can. J. Plant Sci. 87, 479–487. doi: 10.4141/P06-135
Sanderson, M. A., Soder, K. J., Brzezinski, N., Muller, L. D., Skinner, R. H., Wachendorf, M., et al. (2004). “Plant species diversity influences on forage production and performance of dairy cattle on pasture,” in Land use systems in grassland dominated regions, eds A. Lüscher, B. Jeangros, W. Kessler, O. Huguenin, M. Lobsiger, N. Millar, and D. Suter, (Luzern: Proceedings of the 20th General Meeting of the European Grassland Federation), 632–634.
Scharenberg, A., Arrigo, Y., Gutzwiller, A., Wyss, U., Hess, H. D., Kreuzer, M., et al. (2007). Effect of feeding dehydrated and ensiled tanniferous sainfoin (Onobrychis viciifolia) on nitrogen and mineral digestion and metabolism of lambs. Arch. Anim. Nutr. 61, 390–405. doi: 10.1080/17450390701565081
Schaub, S., Finger, R., Leiber, F., Probst, S., Kreuzer, M., Weigelt, A., et al. (2020). Plant diversity effects on forage quality, yield and revenues of semi-natural grasslands. Nat. Commun. 11:768. doi: 10.1038/s41467-020-14541-4
Sharp, J. M., Edwards, G. R., and Jeger, M. J. (2014). A spatially explicit population model of the effect of spatial scale of heterogeneity in grass-clover grazing systems. J. Agr. Sci. 152, 394–407. doi: 10.1017/S0021859613000154
Skinner, R. H., and Dell, C. J. (2016). Yield and soil carbon sequestration in grazed pastures sown with two or five forage species. Crop Sci. 56, 2035–2044. doi: 10.2135/cropsci2015.11.0711
Soder, K. J., Sanderson, M. A., Stack, J. L., and Muller, L. D. (2006). Intake and performance of lactating cows grazing diverse forage mixtures. J. Dairy Sci. 89, 2158–2167. doi: 10.3168/jds.S0022-0302(06)72286-X
Spelman, K., Duke, J. A., and Bogenschutz-Godwin, M. J. (2006). “The synergy principle at work with plants, pathogens, insects, herbivores, and humans,” in Natural Products from Plants, eds P. B. Kaufman, L. J. Cseke, S. Warber, J. A. Duke, and H. L. Brielmann (Boca Raton, FL: CRC Press), 475–501. doi: 10.1201/9781420004472.ch13
Stewart, E. K., Beauchemin, K. A., Dai, X., MacAdam, J. W., Christensen, R. G., and Villalba, J. J. (2019). Effect of tannin-containing hays on enteric methane emissions and nitrogen partitioning in beef cattle. J. Anim. Sci. 97, 3286–3299. doi: 10.1093/jas/skz206
Subbaraj, A. K., Huege, J., Fraser, K., Cao, M., Rasmussen, S., Faville, M., et al. (2019). A large-scale metabolomics study to harness chemical diversity and explore biochemical mechanisms in ryegrass. Commun Biol. 2:87. doi: 10.1038/s42003-019-0289-6
Sullivan, T. P., and Sullivan, D. S. (2006). Plant and small mammal diversity in orchard versus non-crop habitats. Agr. Ecosyst. Environ. 116, 235–243. doi: 10.1016/j.agee.2006.02.010
Sun, X. Z., Henderson, G., Cox, F., Molano, G., Harrison, S. J., Luo, D., et al. (2015). Lambs fed fresh winter forage rape (Brassica napus L.) emit less methane than those fed perennial ryegrass (Lolium perenne L.), and possible mechanisms behind the difference. PLoS ONE 10:e0119697. doi: 10.1371/journal.pone.0119697
Sun, X. Z., Pacheco, D., and Luo, D. W. (2016). Forage brassica: a feed to mitigate enteric methane emissions? Anim. Prod. Sci. 56, 451–456. doi: 10.1071/AN15516
Sun, X. Z., Waghorn, G. C., Hoskin, S. O., Harrison, S. J., Muetzel, S., and Pacheco, D. (2012). Methane emissions from sheep fed fresh brassicas (Brassica spp.) compared to perennial ryegrass (Lolium perenne). Anim. Feed Sci. Tech. 176, 107–116. doi: 10.1016/j.anifeedsci.2012.07.013
Tavendale, M. H., Meagher, L. P., Pacheco, D., Walker, N., Attwood, G. T., and Sivakumaran, S. (2005). Methane production from in vitro rumen incubations with Lotus pedunculatus and Medicago sativa, and effects of extractable condensed tannin fractions on methanogenesis. Anim. Feed Sci. Tech. 123, 403–419. doi: 10.1016/j.anifeedsci.2005.04.037
Teague, W. R., Dowhower, S. L., Baker, S., Haile, N., DeLaune, P. B., and Conover, D. M. (2011). Grazing management impacts on vegetation, soil biota and soil chemical, physical and hydrological properties in tall grass prairie. Agric. Ecosyst. Environ.141, 310–322. doi: 10.1016/j.agee.2011.03.009
Theodoridou, K., Aufrère, J., Andueza, D., Le Morvan, A., Picard, F., Pourrat, J., et al. (2012). Effects of condensed tannins in wrapped silage bales of sainfoin (Onobrychis viciifolia) on in vivo and in situ digestion in sheep. Animal 6, 245–253. doi: 10.1017/S1751731111001510
Theodoridou, K., Aufrère, J., Andueza, D., Pourrat, J., Le Morvan, A., Stringano, E., et al. (2010). Effects of condensed tannins in fresh sainfoin (Onobrychis viciifolia) on in vivo and in situ digestion in sheep. Anim. Feed Sci. Tech. 160, 23–38. doi: 10.1016/j.anifeedsci.2010.06.007
Thornley, J. H. M., Parsons, A. J., Newman, J. A., and Penning, P. D. (1994). A cost-benefit model of grazing intake and diet selection in a two species temperate grassland sward. Funct. Ecol. 8, 5–16. doi: 10.2307/2390105
Tilman, D. (1982). Resource Competition and Community Structure. Princeton, NJ: Princeton University Press. doi: 10.1515/9780691209654
Tilman, D., Reich, P. B., Knops, J., Wedin, D., Mielke, T., and Lehman, C. (2001). Diversity and productivity in a long-term grassland experiment. Science 294, 843–845. doi: 10.1126/science.1060391
Totty, V. K., Greenwood, S. L., Bryant, R. H., and Edwards, G. R. (2013). Nitrogen partitioning and milk production of dairy cows grazing simple and diverse pastures. J. Dairy Sci. 96, 141–149. doi: 10.3168/jds.2012-5504
Tracy, B. F., and Faulkner, D. B. (2006). Pasture and cattle responses in rotationally stocked grazing systems sown with differing levels of species richness. Crop Sci. 46, 2062–2068. doi: 10.2135/cropsci2005.12.0473
Tracy, B. F., Foster, J. L., Butler, T. J., Islam, M. A., Toledo, D., and Vendramini, J. M. (2018). Resilience in forage and grazinglands. Crop Sci. 58, 31–42. doi: 10.2135/cropsci2017.05.0317
Tracy, B. F., and Sanderson, M. A. (2004). Productivity and stability relationships in mowed pastures of varying species composition. Crop Sci. 44, 2180–2186. doi: 10.2135/cropsci2004.2180
Valencia, E., Gross, N., Quero, J. L., Carmona, C. P., Ochoa, V., Gozalo, B., et al. (2018). Cascading effects from plants to soil microorganisms explain how plant species richness and simulated climate change affect soil multifunctionality. Global Change Biol. 24, 5642–5654. doi: 10.1111/gcb.14440
Vasta, V., Daghio, M., Cappucci, A., Buccioni, A., Serra, A., Viti, C., et al. (2019). Invited review: Plant polyphenols and rumen microbiota responsible for fatty acid biohydrogenation, fiber digestion, and methane emission: experimental evidence and methodological approaches. J. Dairy Sci.102, 3781–3804. doi: 10.3168/jds.2018-14985
Vercoe, P. E., Durmic, Z., and Revell, D. K. (2009). “Rumen microbial ecology: helping to change landscapes,” in Nutritional and foraging ecology of sheep and goats, eds T. G. Papachristou, Z. M. Parissi, H. Ben Salem, and P. Morand-Fehr (Zaragoza : CIHEAM / FAO / NAGREF), 225–236.
Villalba, J. J., Bach, A., and Ipharraguerre, I. R. (2011). Feeding behavior and performance of lambs are influenced by flavor diversity. J. Anim. Sci. 89, 2571–2581. doi: 10.2527/jas.2010-3435
Villalba, J. J., Catanese, F., Provenza, F. D., and Distel, R. A. (2012). Relationships between early experience to dietary diversity, acceptance of novel flavors, and open field behavior in sheep. Physiol. Behav. 105, 181–187. doi: 10.1016/j.physbeh.2011.08.031
Villalba, J. J., Costes-Thiré, M., and Ginane, C. (2017). Phytochemicals in animal health: diet selection and trade-offs between costs and benefits. Proc. Nutr. Soc. 76, 113–121. doi: 10.1017/S0029665116000719
Villalba, J. J., and Manteca, X. (2019). A case for eustress in grazing animals. Front. Vet. Sci. 6:303. doi: 10.3389/fvets.2019.00303
Villalba, J. J., Miller, J., Ungar, E. D., Landau, S. Y., and Glendinning, J. (2014). Ruminant self-medication against gastrointestinal nematodes: evidence, mechanism, and origins. Parasite 21:31. doi: 10.1051/parasite/2014032
Villalba, J. J., and Provenza, F. D. (1997). Preference for flavoured foods by lambs conditioned with intraruminal administration of nitrogen. Br. J. Nutr. 78, 545–561. doi: 10.1079/BJN19970174
Villalba, J. J., and Provenza, F. D. (2005). Foraging in chemically diverse environments: Concentrations of energy and protein, and food alternatives influence ingestion of plant secondary metabolites by lambs. J. Chem. Ecol. 31, 123–138. doi: 10.1007/s10886-005-0979-z
Villalba, J. J., and Provenza, F. D. (2009). Learning and dietary choice in herbivores. Rangeland Ecol. Manag. 62, 399–406. doi: 10.2111/08-076.1
Villalba, J. J., Provenza, F. D., and Bryant, J. P. (2002). Consequences of the interaction between nutrients and plant secondary metabolites on herbivore selectivity: benefits or detriments for plants? Oikos 97, 282–292. doi: 10.1034/j.1600-0706.2002.970214.x
Villalba, J. J., Provenza, F. D., Catanese, F., and Distel, R. A. (2015). Understanding and manipulating diet choice in grazing animals. Anim. Prod. Sci. 55, 261–271. doi: 10.1071/AN14449
Villalba, J. J., Provenza, F. D., and GouDong, H. (2004). Experience influences diet mixing by herbivores: Implications for plant biochemical diversity. Oikos 107, 100–109. doi: 10.1111/j.0030-1299.2004.12983.x
Villalba, J. J., Provenza, F. D., and Manteca, X. (2010). Links between ruminants' food preference and their welfare. Animal 4, 1240–1247. doi: 10.1017/S1751731110000467
Villalba, J. J., Provenza, F. D., and Shaw, R. (2006). Sheep self-medicate when challenged with illness-inducing foods. Anim. Behav. 71, 1131–1139. doi: 10.1016/j.anbehav.2005.09.012
Villalba, J. J., Provenza, F. D., and Stott, R. (2009). Rumen distension and contraction influence feed preference by sheep. J. Anim. Sci. 87, 340–350. doi: 10.2527/jas.2008-1109
Villalba, J. J., Spackman, C., Goff, B., Klotz, J. L., Griggs, T., and MacAdam, J. W. (2016). Interaction between a tannin-containing legume and endophyte-infected tall fescue seed on lambs' feeding behavior and physiology. J. Anim. Sci. 94, 845–857. doi: 10.2527/jas.2015-9790
Vogeler, I., Vibart, R., and Cichota, R. (2017). Potential benefits of diverse pasture swards for sheep and beef farming. Agr. Syst. 154, 78–89. doi: 10.1016/j.agsy.2017.03.015
Waghorn, G. (2008). Beneficial and detrimental effects of dietary condensed tannins for sustainable sheep and goat production - Progress and challenges. Anim. Feed Sci. Tech. 147, 116–139. doi: 10.1016/j.anifeedsci.2007.09.013
Waghorn, G. C., and McNabb, W. C. (2003). Consequences of plant phenolic compounds for productivity and health of ruminants. Proc. Nutr. Soc. 62, 383–392. doi: 10.1079/PNS2003245
Wallis, R. H., Thomas, D. T., Speijers, E. J., Vercoe, P. E., and Revell, D. K. (2014). Short periods of prior exposure can increase the intake by sheep of a woody forage shrub, Rhagodia preissii. Small Rumin. Res. 121, 280–288. doi: 10.1016/j.smallrumres.2014.08.006
Wang, L., Wang, D. L., He, Z. B., Liu, G. F., and Hodgkinson, K. C. (2010). Mechanisms linking plant species richness to foraging of a large herbivore. J. Appl. Ecol. 47, 868–875. doi: 10.1111/j.1365-2664.2010.01837.x
Wang, Y., McAllister, T. A., and Acharya, S. (2015). Condensed tannins in sainfoin: composition, concentration, and effects on nutritive and feeding value of sainfoin forage. Crop Sci. 55, 13–22. doi: 10.2135/cropsci2014.07.0489
Wedin, W. F., Donker, J. D., and Marten, G. C. (1965). An evaluation of nitrogen fertilization in legume-grass and all-grass pasture. Agron. J. 58, 185–188. doi: 10.2134/agronj1965.00021962005700020015x
Westoby, M. (1974). An analysis of diet selection by large generalist herbivores. Am. Nat. 108, 290–304. doi: 10.1086/282908
Westoby, M. (1978). What are the biological bases of varied diets? Am. Nat. 112, 627–631. doi: 10.1086/283303
Whitehead, D. C. (2000). Nutrient Elements in Grassland: Soil-Plant-Animal Relationships. New York, NY: CABI. doi: 10.1079/9780851994376.0000
Wilson, J. R., and Ng, T. T. (1975). Influences of water stress on parameters associated with herbage quality of Panicum maximum var. trichoglume. Aust. J. Agric. Res. 26, 127–136. doi: 10.1071/AR9750127
Yachi, S., and Loreau, M. (1999). Biodiversity and ecosystem productivity in a fluctuating environment: the insurance hypothesis. Proc. Natl. Acad. Sci. U.S.A. 96, 1463–1468. doi: 10.1073/pnas.96.4.1463
Yearsley, J. M., Villalba, J. J., Gordon, I. J., Kyriazakis, I., Speakman, J. R., Tolkamp, B. J., et al. (2006). A theory of associating food types with their post-ingestive consequences. Am. Nat. 167, 705–716. doi: 10.1086/502805
Keywords: diverse pastures, phytochemical diversity, ruminants, environmental impacts, animal nutrition, animal health, animal welfare, animal production
Citation: Distel RA, Arroquy JI, Lagrange S and Villalba JJ (2020) Designing Diverse Agricultural Pastures for Improving Ruminant Production Systems. Front. Sustain. Food Syst. 4:596869. doi: 10.3389/fsufs.2020.596869
Received: 20 August 2020; Accepted: 12 October 2020;
Published: 30 October 2020.
Edited by:
Iain James Gordon, Australian National University, AustraliaReviewed by:
Lucinda Watt, Commonwealth Scientific and Industrial Research Organisation (CSIRO), AustraliaKarl Kunert, University of Pretoria, South Africa
Copyright © 2020 Distel, Arroquy, Lagrange and Villalba. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Roberto A. Distel, Y2VkaXN0ZWxAY3JpYmEuZWR1LmFy