 Rocío Gonzalez-Soltero
Rocío Gonzalez-Soltero Mariangela Tabone
Mariangela Tabone Mar Larrosa
Mar Larrosa Maria Bailen
Maria Bailen Carlo Bressa
Carlo Bressa- 1Masmicrobiota Group, Madrid, Spain
- 2Faculty of Biomedical and Health Sciences, Universidad Europea de Madrid, Madrid, Spain
- 3Department of Nutrition and Food Science, Faculty of Pharmacy, Universidad Complutense de Madrid, Madrid, Spain
- 4Department of Preventive Medicine, Public Health and Microbiology, Faculty of Medicine, Universidad Autónoma de Madrid, Madrid, Spain
- 5Faculty of Experimental Sciences, Universidad Francisco de Vitoria, Madrid, Spain
Background: The VDR gene is identified as a crucial host factor, influencing the gut microbiota. The current research focuses on an observational study that compares gut microbiota composition among individuals with different VDR gene TaqI polymorphisms in a Caucasian Spanish population. This study aims to elucidate the interplay between genetic variations in the VDR gene and the gut microbial composition.
Methods: 87 healthy participants (57 men, 30 women), aged 18 to 48 years, were examined. Anthropometric measures, body composition, and dietary habits were assessed. VDR gene polymorphism TaqI rs731236 was determined using TaqMan assays. The V3 and V4 regions of the 16S rRNA gene were sequenced to study bacterial composition, which was analyzed using QIIME2, DADA2 plugin, and PICRUSt2. Statistical analyses included tests for normal distribution, alpha/beta diversity, ADONIS, LEfSe, and DESeq2, with established significance thresholds.
Results: No significant differences in body composition or dietary habits were observed based on VDR genotypes. Dietary intake analysis revealed no variations in energy, macronutrients, or fiber among the different VDR genotypes. Fecal microbiota analysis indicated significant differences in alpha diversity as measured by Faith’s Phylogenetic Diversity index. Differential abundance analysis identified taxonomic disparities, notably in the genera Parabacteroides and Butyricimonas.
Conclusion: Overall, this study suggests potential associations between genetic variations in the VDR gene and the composition and function of gut microbiota.
1 Introduction
The Vitamin D receptor (VDR) is a conserved nuclear receptor controlling the primary physiological actions vitamin D’s, active form, 1α,25-dihydroxyvitamin D3 (1,25(OH)2D3) (1). It is part of the ligand-responsive transcription factors in the nuclear hormone receptor superfamily (2). This receptor is prominently expressed in multiple tissues, including the intestines, adipose tissue, liver, and immune cells, with higher expression observed in small intestine and colon epithelial cells (3, 4). It plays a key role in regulating metabolic and immune functions, serving as an essential regulator of intestinal cell proliferation and differentiation, barrier function, intestinal immunity and host-microbial interactions (5).
The VDR gene exhibits an extensive array of allelic variants within the VDR locus, encompassing over 900 variants. Among these, the most extensively researched polymorphisms include ApaI (rs7975232), BsmI (rs1544410), TaqI (rs731236), and FokI (rs10735810) (6). Regarding the BsmI–ApaI–TaqI polymorphism, the findings remain controversial. Although there is a tendency for the Bat haplotype (C allele for TaqI) to exhibit higher levels of mRNA expression compared to the baT haplotype (T allele for TaqI), the results have been inconsistent. Nevertheless, there appears to be a general trend indicating that the Bat haplotype demonstrates a more favorable response than the baT haplotype (7). The TaqI restriction site polymorphism (T/C) has been linked with health status in various studies. The T allele of the TaqI polymorphism has been associated with the risk of suffering post-transplant diabetes mellitus (DM) (8), and osteoarthritis in the knee (9). Links have been established between TaqI polymorphisms and susceptibility to Crohn’s disease, multiple sclerosis or primary hyperparathyroidism, renal carcinoma, and the risk of metastases in breast cancer (10). However, controversial results have been observed between TaqI polymorphisms and prostate cancer, DM type 2 and nephrolithiasis (10). Additionally, VDR has been proposed as a potential key player in the pathogenesis of obesity, through the discovery of a strong association between the VDR TaqI ‘T’ allele and obesity in a Greek population (11). A study in Iranian population also found a significant association between the TaqI ‘T’ allele and obesity, contributing to an increment in the BMI of 3 kg/m2 per risk allele (12), whereas in Asian Indians the tt (CC) genotype of the TaqI polymorphism was associated with ≥5% weight loss after lifestyle interventions (13). However, no significant relationships were observed between TaqI genotypes when analyzed as a risk score in a Mendelian Randomization study in 42,024 Caucasians. A significant causal effect of the vitamin D metabolism-risk score on obesity was not found (14). The combined effects of obesity and VDR gene polymorphisms on susceptibility to type 2 diabetes mellitus (T2DM) have been explored. However, the specific relationship between TaqI polymorphism and T2DM remains unclear (15). Overall, the association between TaqI VDR polymorphism and obesity or T2DM is still under investigation, and further research is required to fully understand the relationship. However, these data suggest that lifestyle interventions could reveal one or several modifiable factors associated with the TT genotype that may be responsible for the effect.
The human VDR gene is the first to be identified in a genome-wide association analysis, involving two cohorts, totaling 1812 individuals, as a critical host factor that shapes the gut microbiota at the genetic level (16). In mice lacking the VDR gene, significant shifts in the microbiota relative to control mice have been observed. These changes encompass reduced levels of bacteria belonging to the Firmicutes phylum, increased levels from the Bacteroidota and Proteobacteria phyla (17), modifications in various bacterial genera, including Eubacterium, Bacteroides, and Salmonella (18), and shifts in critical pathways within the intestinal microbiota. These alterations may have implications for detoxification, infection, cancer, and other diseases (19). The absence of VDR leads to dysbiosis, indicating a critical role of VDR in shaping gut microbiota communities (20).
In this work we have conducted an observational study to compare the gut microbiota composition among individuals with different VDR gene TaqI (rs731236) polymorphisms, in a Caucasian Spanish population. By analyzing genetic variations in the VDR gene and their potential impact on the gut microbial composition, we aim to shed light on the interplay between these factors and contribute to the development of personalized medical approaches.
2 Materials and methods
2.1 Ethics approval and consent to participate
The study was an observational study conducted in accordance with the Declaration of Helsinki, and the protocol received approval from the Research Ethics Committee of the Community of Madrid (CEIm-R; Ref: 47/560280.9/18). All participants provided written informed consent.
2.2 Participant characteristics
The study included a total of 87 healthy participants, comprising 57 men and 30 women, aged 18 to 48 years. Inclusion criteria included healthy Caucasian men and women aged 18–50 years with a body mass index (BMI) of 18.5–25 kg/m2 and exclusion criteria any kind of pathology (current or within 6 months prior to the study), previous gastrointestinal surgery, antibiotic intake 3 months before the study, smoking, use of prebiotics, probiotics, or nutritional complements, being vegetarian or vegan diets, and pregnancy or lactation.
2.3 Anthropometry and body composition
Height and weight were measured with a tallimeter (Asimed T2, Barcelona, Spain) and a balance scale (Ano Sayol SL, Barcelona, Spain), respectively. Body mass index (BMI) was calculated as weight (kg) divided by height (m2). Body composition, including estimated visceral adipose tissue (VAT), body fat percentage (BFP), and body fat mass (BFM), was evaluated on the day of stool sample collection using dual-energy X-ray absorptiometry (DEXA; Hologic DEXA scan, Hologic Inc., Barcelona, Spain).
2.4 Dietary habits
A self-reported food frequency questionnaire (FFQ), validated for the Spanish adult population, was used to gather information on the frequency and quantity of food consumption. The questionnaire includes 93 items encompassing various foods and food groups. Participants reported their usual intake of different foods and beverages categorizing their consumption frequency within predefined intervals (never or less than once a month, 1–3 times per month, 1 time per week, 2–4 times per week, 5–6 times per week, 1 time per day, 2–3 times per day, 4–5 times per day, 6 or more times per day). The data collected from the questionnaire was input into the DietSource 3.0 software (Novartis, Barcelona, Spain), a dietary analysis program to calculate the intake of macronutrients, fiber and total energy (21).
2.5 Sample collection
Participants were provided with the Fe-Col® Fecal Sample Collection Kit (Alpha Laboratories, Hampshire, United Kingdom), along with an insulated bag and ice blocks to preserve the samples until delivery to the laboratory. Stool samples were stored at −80°C until extraction.
2.6 DNA extraction
DNA from humans and bacteria was extracted from 100 mg of stool sample using the commercial E.Z.N.A.® Stool DNA Kit (Omega Biotek, Norcross, GA) and a bead-beating homogenizer (Bullet Blender Storm, Next Advance, NY). The concentration and purity of DNA were assessed using the Quant-iT PicoGreen dsDNA Assay Kit (ThermoFisher Scientific, Waltham, MA) and an FP-8300 spectrofluorimeter (Jasco, Tokyo, Japan). Bacterial DNA was utilized for microbiota analysis, while human DNA was used for VDR genotyping.
2.7 VDR genotyping
Applied Biosystems TaqMan® SNP Genotyping Assays (Applied Biosystems, Foster City, CA, Country; assay ID: C_2404008_10) were used for allelic discrimination analysis of the TaqI VDR (rs731236) gene polymorphism. The StepOnePlus Real-Time PCR system (ThermoFisher Scientific) was employed following a protocol that included denaturation at 95°C for 10 min, then 50 cycles of denaturation at 92°C for 15 s, annealing/extension at 60°C for 1 min, and a final extension step of 30 s at 60°C. Fluorescence analysis was performed with Allelic Discrimination 7,500 software v.2.0.2. After genotyping, participants were classified based on the VDR genotype: TT, TC and CC for subsequent analyses. The allele and genotypes frequencies were calculated using the SNPStat program (22). Experiments were conducted three times to solve incongruences in the genotype assignment. In cases where conflict arise, the Allelic Discrimination 7,500 software v.2.0.2 provided by Thermofisher was used.
2.8 Sequencing and bioinformatics
The hypervariable V3 and V4 regions of the 16S rRNA gene were amplified using the primer pair 5’-TCGTCGGCAGCGTCAGATGTGTATAAGAGACAG-3′ and 5’-GTCTCGTGGGCTCGG AGATGTGTATAAGAGACAG-3′. The 459 bp amplicon was visualized in a 0.8% agarose gel stained with ethidium bromide, and bands were cut and purified using the MinElute Gel Extraction Kit (Qiagen, Hilden, Germany). The DNA amplicons were sequenced on a MiSeq Illumina platform (Illumina, San Diego, CA). The sequencing outputs were analyzed using the Quantitative Insights into Microbial Ecology (QIIME2) program, version V2023.2 (23). The 16 s rRNA paired reads were imported in QIIME2 and processed with the DADA2 plugin (24), with the maximum expected error threshold set to 2.0 for both forward and reverse reads. Taxonomy assignments were conducted using the classify-sklearn method (25) and a customized classifier based on the SILVA reference database (26, 27). To build the customized reference database, sequences, aligned with our primers (forward primer sequence: CCTACGGGNGGCWGCAG, reverse primer sequence: GACTACHVGGGTA TCTAATCC) were extracted from the SILVA 138.1 database clustered at 99% identity. The classifier was trained using our tailored reference reads and SILVA 7-levels for reference taxonomy, including species probabilities (weights) likely observed in human stool1 (28, 29). Diversity analyses were performed through QIIME 2’s q2-diversity plugin. Beta-diversity was assessed by calculating the Bray-Curtis, Jaccard, unweighted and weighted Unifrac distance metrics. For alpha-diversity, observed features (ASVs), Evenness, Shannon and Faith’s Phylogenetic Diversity indices were calculated. Kyoto Encyclopedia of Genes and Genomes (KEGG) ortholog abundances predictions were obtained with the Phylogenetic Investigation of Communities by Reconstruction of Unobserved States (PICRUSt2) software (30) using default “max parsimony” method for hidden-state prediction and a Nearest Sequenced Taxon Index (NSTI) value of 2.0.
2.9 Statistical analysis
Statistical analysis was conducted using QIIME2 v2023.2, SPSS software v26.0 (SPSS, Chicago, IL) and the R statistical package v4.1.1 incorporating the packages: Phyloseq (31) and microbiomeMarker (32). The Shapiro–Wilk test was used to assess the normal distribution of variables; non-parametric tests were performed when a normal distribution was not observed. Differences between groups in terms of alpha-diversity were assessed using the Kruskal–Wallis test (33). A significance threshold of adjusted p value = 0.05 was applied in all calculations. For beta-diversity, the differences between groups were assessed using the ADONIS permutation-based statistical test adjusted for sex (34). A significance threshold of p value = 0.05 was used in all calculations. Linear discriminant analysis coupled with effect size (LEfSe v1.0) was employed to identify bacterial-associated pathways that exhibited differential representation between groups, utilizing default settings. Initially, the Kruskal-Wallis rank test was applied to discern specific differences among groups. Subsequently, the Wilcoxon rank test was conducted, utilizing sex as subgroups, to assess the consistency of differences identified in the preceding step. Significance was set at p < 0.05. To determine whether the allele frequencies were in Hardy–Weinberg equilibrium for the VDR genotypes we employed the Chi-square (χ2) test. The determination of differentially abundant bacterial taxa between VDR gene TaqI polymorphisms was performed using analysis of composition of microbiomes (ANCOM-BC) (35) and DESeq2 (36) adjusting for sex. DESeq2 applies a negative binomial generalized linear model for each taxon count, estimating log-fold changes between two classes and using a Wald test on this value for significance testing. ANCOM-BC addresses variability in microbiota samples by assuming that the observed sample represent an unknown fraction of the ecosystem’s unit volume, with varying sampling fractions among samples. This method includes sample-specific offsets in a linear regression framework, estimated from observed data, serving as bias corrections. Both DESeq2 and LEfSe analyses were conducted on prefiltered data. The raw dataset was prefiltered to remove rare taxa occurring in less than 20% of the samples. ANCOM-BC being more conservative and precise identifies only a small number of ASVs as significant, potentially reducing sensitivity. Therefore, DESeq2 was also utilized to enhance sensitivity (37). The p-values from the DESeq2 test were adjusted for multiple testing using the Benjamini-Hochberg false discovery rate (FDR) procedure, with results deemed significant at FDR <0.05, in line with DESeq2’s default settings. The p-values from the ANCOM-BC test were adjusted using the Holm–Bonferroni method, with results considered significant at alpha <0.05, as per ANCOMBC’s default settings.
3 Results
3.1 Subjects, genotypes and allelic frequencies
Eighty-seven subjects were recruited for this study comprising 57 men and 30 women. Genotyping results for the rs731236 (TaqI) polymorphism were obtained and are presented in Table 1. Compared to the 1,000 Genomes Project reference the calculated allele frequencies (T = 0.61 and C = 0.39) matched those expected for a European population sample. When examining genotypes, TT homozygotes were found at a frequency of 0.43, while TC heterozygotes and CC homozygotes had frequencies of 0.38 and 0.20, respectively, in accordance with Hardy–Weinberg equilibrium (p = 0.071; Table 1).
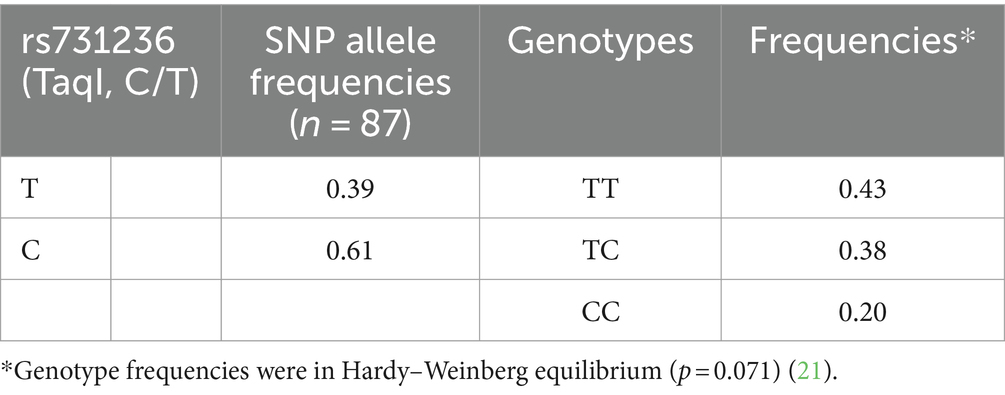
Table 1. SNP allele and genotype frequencies.
Furthermore, we grouped the carriers for the common allele (TT) for VDR and compared their frequency with carriers of the rare allele (CC and TC) across all the parameters of this study (body composition, dietary habits, and microbiota). Additionally, carriers of the recessive alleles (CC) were grouped and compared against carriers of the dominant allele (TT and TC) following the approach previously used by Vasilopoulos (11).
3.2 Body composition and dietary habits
In this study, we explored the potential impacts of the polymorphisms on body composition and dietary habits. The aim to control for dietary habits was to ensure that observed differences are not attributed to various dietary patterns, especially given the significant influence of diet on the microbiota. After categorizing participants based on their VDR genotypes (see Supplementary Table S1), no significant differences in body composition parameters were observed between the groups (genotypic or dominant and recessive allele grouping).
Furthermore, we utilized a Food Frequency Questionnaire (FFQ) to record and analyze the participants’ dietary intake, including energy, macronutrients, and fiber. Subsequent comparisons of various VDR genotypes (Supplementary Table S2) revealed no statistically significant variations in the intake levels of macronutrients (such as carbohydrates, protein, fat, protein/carbohydrate ratio, and protein/fat ratio), fiber, and total energy.
3.3 Fecal microbiota
The average number of reads per sample was 167,839. In the study of alpha diversity indices among VDR genotypes, no significant differences were observed in measures of richness or evenness except for Faith’s Phylogenetic Diversity index. This index, which accounts for both species richness and phylogenetic distance among species present in a community, showed significant differences (p = 0.013; Figure 1). The results for observed features (p = 0.250), Shannon entropy (p = 0.138) or Pielou evenness (p = 0.320) indicated no significant variations. Similarly, for β-diversity (Supplementary Figure S1), no significant differences were detected across the following distance metrics: Bray–Curtis (R2 = 0.02 p = 0.633), Jaccard (R2 = 0.02 p = 0.093), unweighted Unifrac (R2 = 0.03 p = 0.127) and weighted Unifrac (R2 = 0.02 p = 0.497).
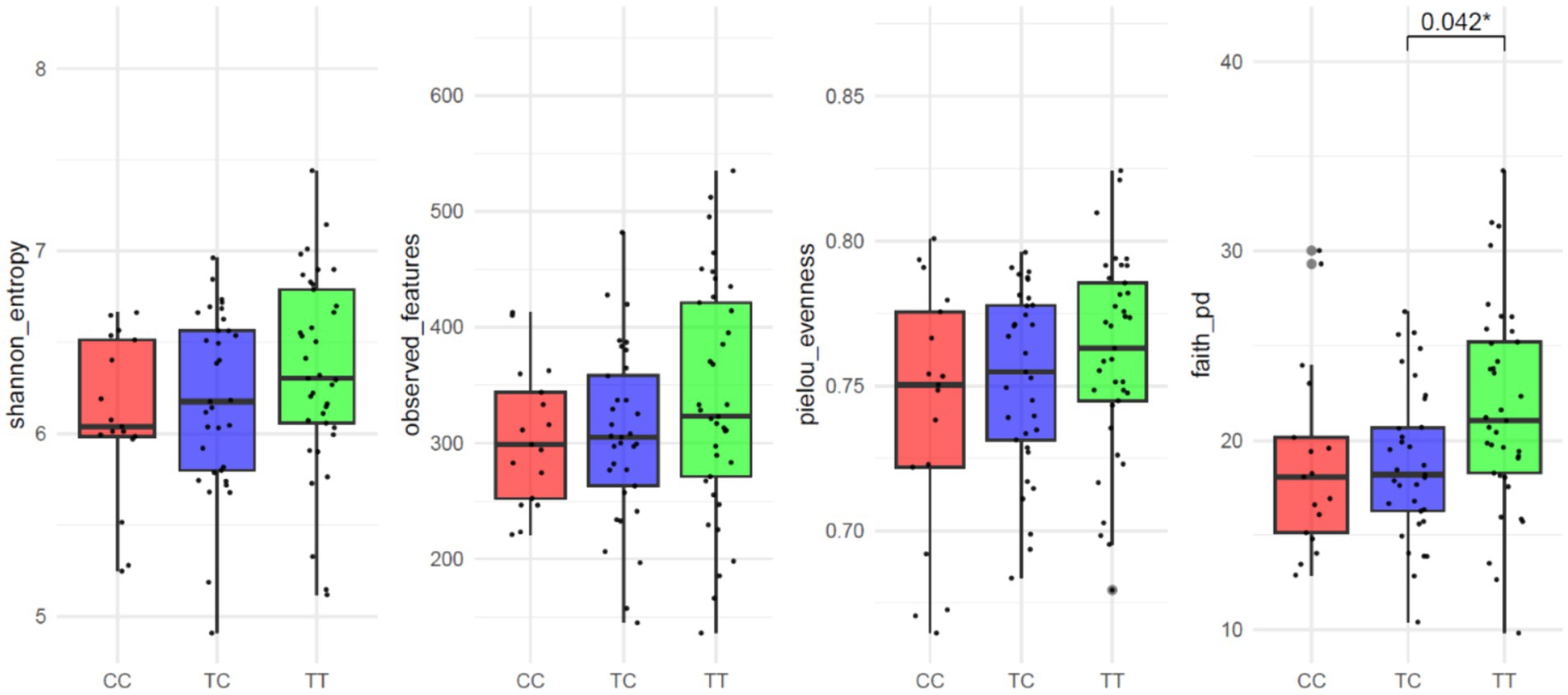
Figure 1. Alpha-diversity parameter of VDR polymorphisms: Shannon entropy, Observed features, Pielou evenness and Faith’s Phylogenetic Diversity.
When analyzing results were according to dominant and recessive allelic grouping (CC vs. TC_TT and TT vs. TC_CC) no significant differences between groups were observed for alpha diversity parameters except for Faith’s Phylogenetic Diversity common allele TT vs. rare allele TC_CC grouping (p = 0.006; CC vs. TC_TT p = 0.167; Supplementary Figure S1). Other alpha diversity parameters showed no significant variations: observed features (TT vs. TC_CC p = 0.105; CC vs. TC_TT p = 0.380), Shannon entropy (TT vs. TC_CC p = 0.063; CC vs. TC_TT p = 0.164), or Pielou evenness (TT vs. TC_CC p = 0.144; CC vs. TC_TT p = 0.234).
Additionally, no significant differences were found in β-diversity parameters except for unweighted Unifrac distance metric in the TT vs. TC_CC grouping (Figure 2; Supplementary Figures S2, S3): Jaccard (TT vs. TC_CC: R2 = 0.01 p = 0.059; CC vs. TC_TT: R2 = 0.01 p = 0.585), Bray–Curtis (TT vs. TC_CC: R2 = 0.01 p = 0.181; CC vs. TC_TT: R2 = 0.01 p = 0.997), unweighted Unifrac (TT vs. TC_CC: R2 = 0.02 p = 0.026; CC vs. TC_TT: R2 = 0.01 p = 0.743), and weighted Unifrac (TT vs. TC_CC: R2 = 0.01 p = 0.572; CC vs. TC_TT: R2 = 0.01 p = 0.709) distance metrics.
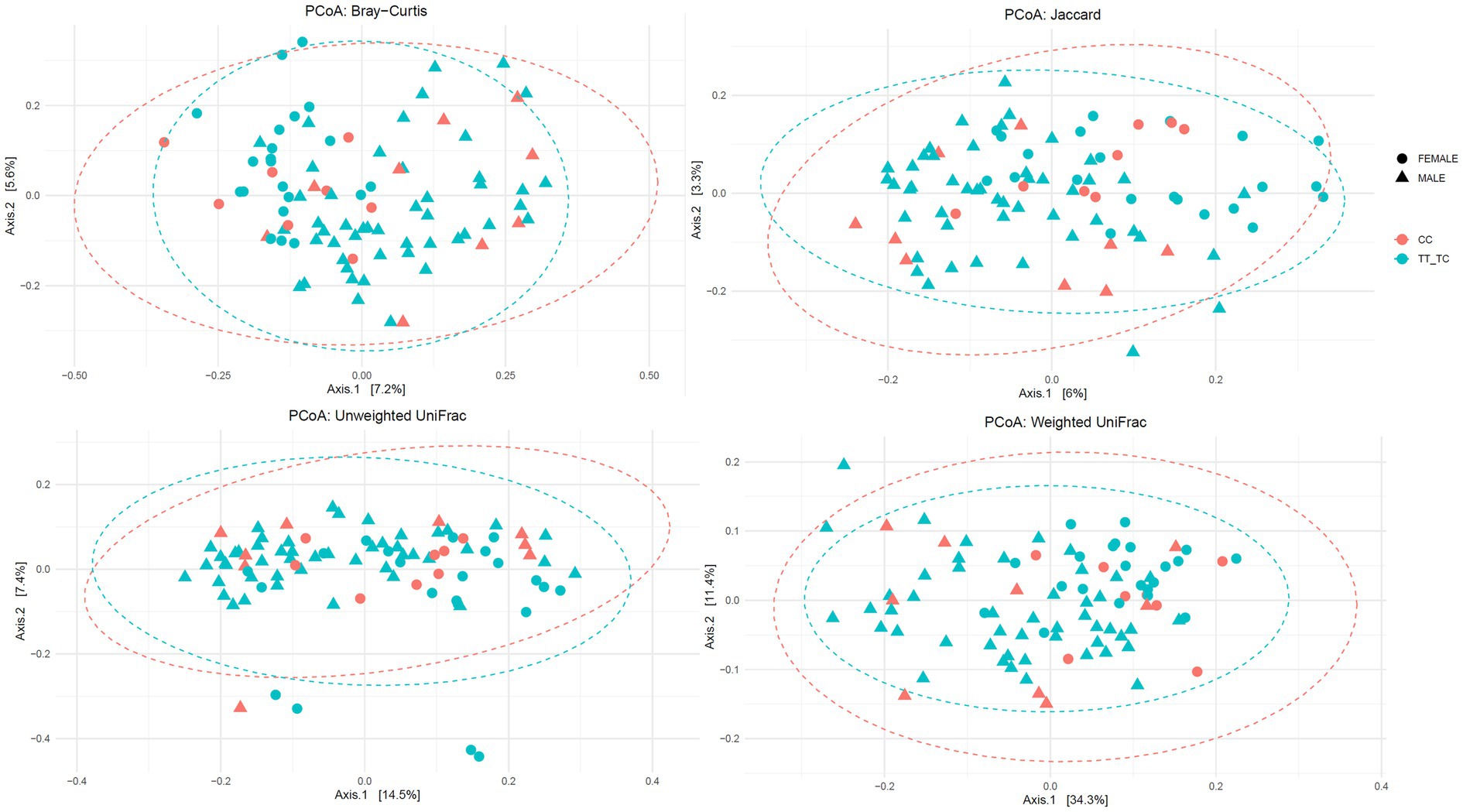
Figure 2. Principal coordinates analysis plots of Bray-Curtis, Jaccard, unweighted and weighted Unifrac distance metrics for VDR polymorphisms according to common versus rare allelic grouping (TT vs. TC_CC).
3.4 Differential abundance analysis
This study further assessed disparities in microbiota communities using DESeq2 methodology on three distinct polymorphisms, TT, CC, and TC, as depicted in Figures 3, 4. Notably, the CC polymorphism on taxa within the Parabacteroides and Butyricimonas genera, both constituents of the Bacteroidota phylum, and the Victivallis genus, showed a decrease when compared to TC and TT polymorphisms. In contrast, genera from Firmicutes phylum, inlcuding Ruminococcus gauvreauii group, Holdemanella, Catenibacterium, and the Christensenellaceae_R-7 group, were increased in CC polymorphism, as illustrated in Figure 3. Noteworthily, no significant disparities in microbiota were observed between TC and TT polymorphisms.
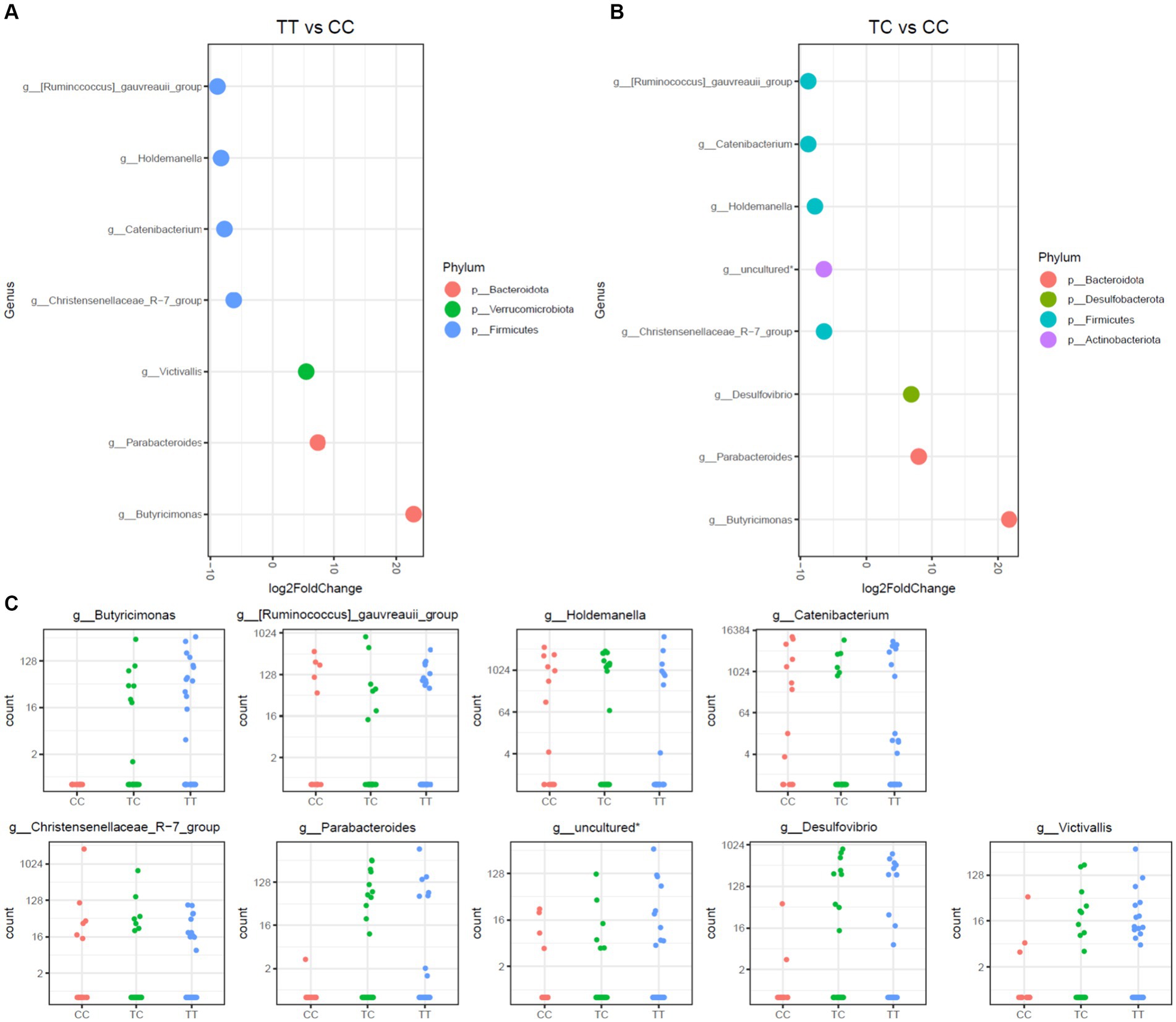
Figure 3. Differential abundance analysis of VDR polymorphisms (TT, TC, CC) by DESeq2 adjusted by sex. (A). Log fold changes for all significant genera between TT and CC. (B). Log fold changes for all significant genera between TC and CC. (C). Normalized abundances of genera identified by differential abundance analysis. Boxplots represent normalized count abundances of individual genera in each group (CC, TC, TT). *Belonging to order_Coribacteriales_family_uncultured_genus_uncultured.
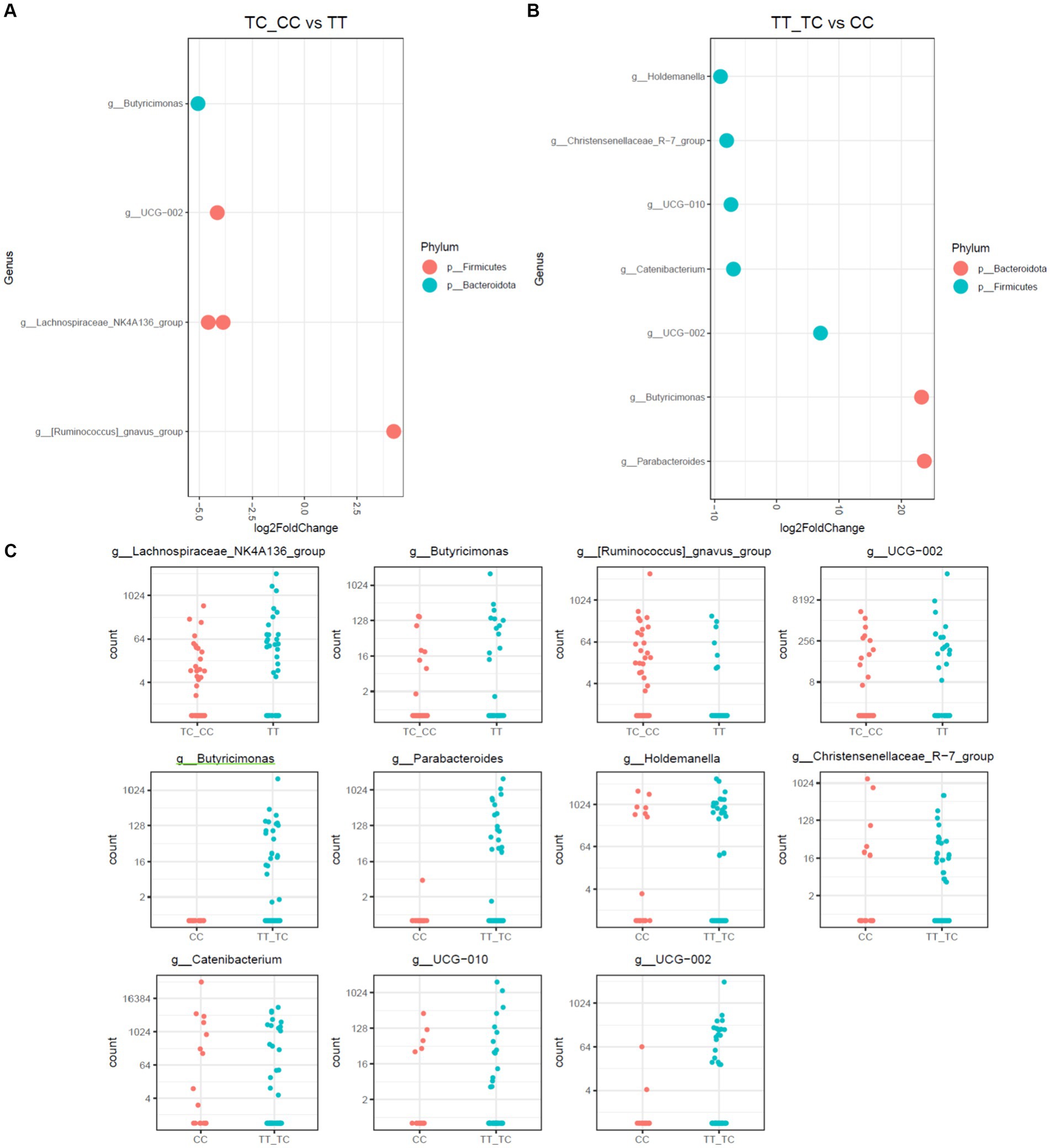
Figure 4. Differential abundance analysis of VDR polymorphisms in allelic grouping by DESeq2 adjusted by sex results for DESeq2 adjusted by sex. (A) Log fold changes for all significant genera between TC_CC and TT. (B) Log fold changes for all significant genera between TT_TC and CC. (C) Normalized abundances of genera identified by differential abundance analysis. Boxplots represent normalized count abundances of individual genera in each group.
When allelic grouping was employed for comparison of microbiota communities [carriers for the common allele (TT) vs. rare allele (CC and TC) and carriers for the recessive allele (CC) vs. dominant allele (TT and TC)], similar outcomes were discerned (Figure 4). The microbiota associated with the CC allelic variant also showed a decrease in Parabacteroides and Butyricimonas, alongside with an increase in the taxa Holdemanella, Catenibacterium, and the Christensenellaceae_R-7 group. Additionally, in the comparison of the TC_CC allelic variants, a reduction of Butyricimonas and a rise of the Ruminococcus gauvreauii group were observed.
ANCOM-BC, adjusted for sex, was employed to investigate dissimilarities in microbial communities. Notably, only a single Amplicon Sequence Variant (ASV) affiliated with Butyricimonas virosa (B. virosa) exhibited enrichment in the microbiota of the TT_TC group when compared with the CC group, as illustrated in Figure 5. No other taxonomic differences were identified when performing comparisons among genotyping groups (TT, TC, CC) or when comparing allelic groups TC_CC and TT.

Figure 5. Differential abundance analysis by ANCOM BC adjusted by sex. Log fold change of differentially abundant ASV corresponding to Butyricimonas virosa when comparing TT_TC to CC.
3.5 Predicted functional metagenome by PICRUSt
PICRUSt was employed to infer the functional capacities of microbial communities by projecting functional genes linked to various taxa (Figure 6). No significant differences in metabolic pathways were observed when comparing the microbiota from TT and TC polymorphisms. However, LEfSe analysis revealed the enrichment of several pathways within the CC polymorphisms. These pathways included nivalenol biosynthesis (PW 7013), ubiquinol-8 biosynthesis (early decarboxylation; PWY-6708), ubiquinol-10 biosynthesis (early decarboxylation; PWY-5857), ubiquinol-9 biosynthesis (early decarboxylation; PWY-5856), ubiquinol-7 biosynthesis (early decarboxylation; PWY-5855), and the superpathway ubiquinol 8 (UBISYN-PWY) within microbial communities associated with CC polymorphisms when compared to TC. In addition, nivalenol biosynthesis (PW 7013) was the only pathway that exhibited upregulation in CC when contrasted with TT_TC. Furthermore, the superpathway of (Kdo)2-lipid A biosynthesis (KDD-NAGLIPASYN-PWY) displayed downregulation in the CC group when compared with TT, as well as in the TC_CC group when compared to TT.
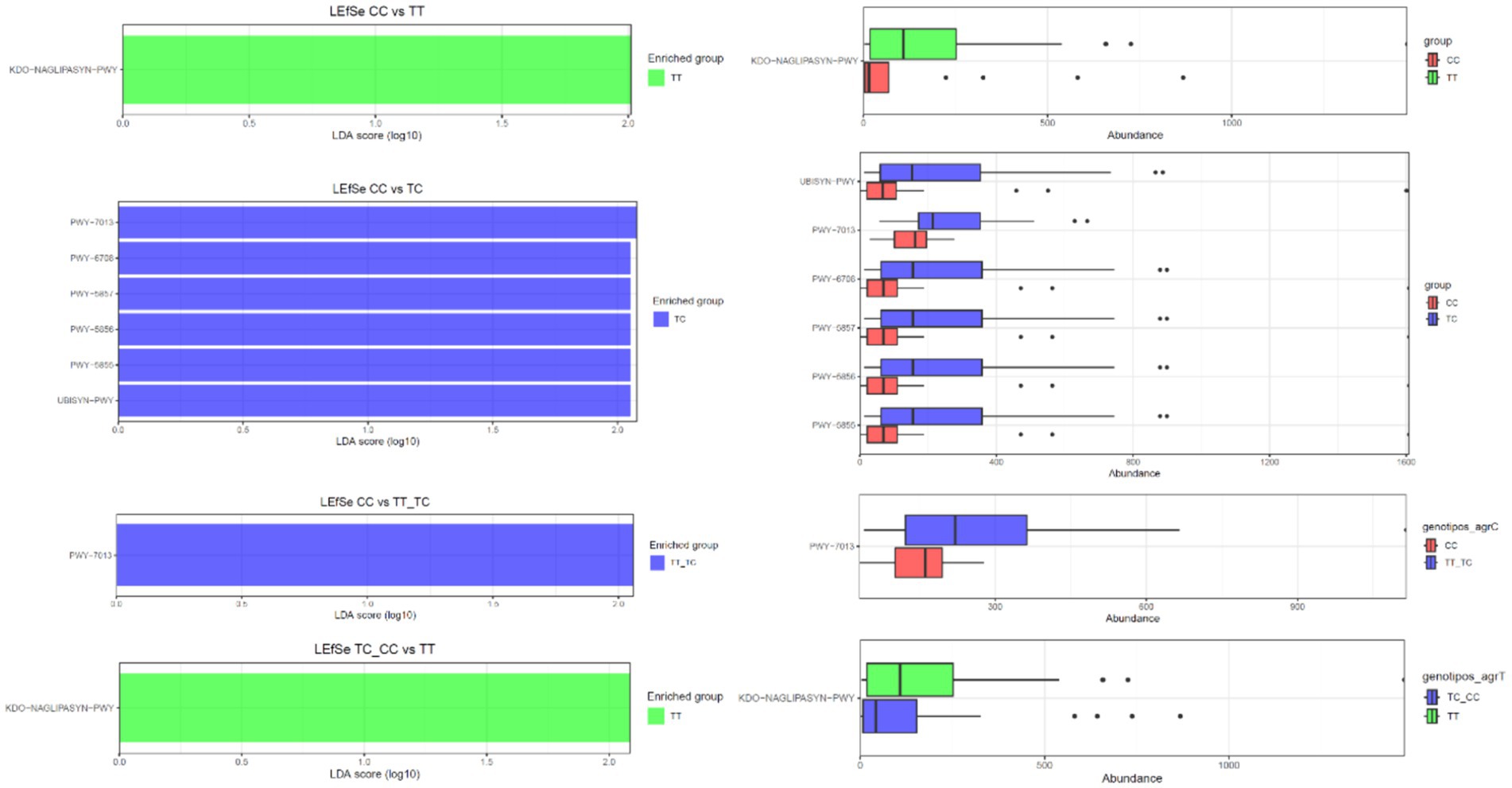
Figure 6. Predicted functional composition of metagenomes based on 16S rRNA gene sequencing data. LEfSe based on the PICRUSTt2 dataset revealed differentially enriched metabolic pathways associated with VDR polymorphisms (TT, TC, CC, TT_TC, TC_CC).
4 Discussion
The genetic variations associated with TaqI rs731236 have been linked to diverse responses in obesity, immune regulation, and inflammatory pathways, suggesting a potential influence on the composition and function of the gut microbiota. This study aimed to characterize the bacterial communities in individuals with different genotypes of the VDR gene genotypes (TaqI rs731236), employing the 16S rRNA gene study approach. We investigated microbial diversity and abundance across groups (CC, TT, TC), to explore potential associations between the TaqI polymorphism alleles C/T and genotypes groups with specific microbial profiles.
Analyses of alpha-diversity and beta-diversity reveal that among all the studied indices, only those considering phylogenetic diversity (Unweighted UniFrac, Faith PD) are significantly different among populations with different genotypes for the TaqI polymorphism of the VDR gene. This indicates that the observed differences among genotypes are primarily due to variations in the presence of taxonomically distinct bacterial species rather than abundance differences. Different phylogenetic lineages can play specific roles in the functionality of the microbiota (38). Therefore, changes in phylogenetic diversity may impact the metabolic and biological functions of the microbial community. The phylogenetic diversity was found to be higher in individuals with the TT genotype, possibly indicating a distinct and/or enhanced functionality of the microbiota in this group. The TT genotype/T allele is linked to a higher abundance of the Parabacteroides and Butyricimonas genera, recognized for their roles in degradation of complex carbohydrates and production of short-chain fatty acids. The Parabacteroides genus, a commensal member of the gut microbiota, has previously been associated as being influenced by the VDR gene (16). Species within Parabacteroides act as immune system regulators and possess anti-inflammatory properties (39). Some members of this genus play a crucial role in the digestion of complex carbohydrates, producing short-chain fatty acids (SCFAs) such as acetate, propionate, and butyrate, thereby providing benefits to the host. Hence, modifications in the population of this genus have been associated to host’s health/disease status, with some species suggested as next-generation probiotics (39). The Butyricimonas genus, as its name suggests, is associated with butyric acid production, offering several benefits, including energy provision to colonocytes, immune system modulation by providing anti-inflammatory properties, improved nutrient absorption, and hepatic lipid metabolism regulation (40). B. virosa has been proposed to have therapeutic potential for preventing high-fat diet (HFD)-induced diabetes and metabolic disorders in mice. Both live and heat-killed B. virosa improved body weight, serum glucose level, insulin resistance, and liver steatosis in HFD-induced obese mice (41). Nevertheless, the TT genotype has also been associated to increased insulin resistance and chronic low-grade inflammation (42). This could be explained by the fact that microbiota that produces a high amount of short-chain fatty acids is also associated with microbiota dysbiosis, obesity and cardiometabolic risk. However, the mentioned study does not conduct an analysis of the microbiota, and currently, this is the first time in the literature that the TT genotype/T allele is associated with the presence of Butyricimonas in the microbiota (43). Further research in this area is needed.
Finally, the Victivallis genus, overrepresented in the TT genotype/ T allele, is a sugar fermenter and acetate producer (44). Victivallis, known for anaerobic sugar fermentation, alongside genera like Parabacteroides and Butyricimonas, known for complex carbohydrate degradation and butyrate production, respectively, are also significant components of the gut microbiota. Victivallis has been associated with decreased adiposity, hepatic steatosis, and diabetes in animal models (45, 46), and is less abundant in hypercholesterolemic patients (47), suggesting potential positive effects on glucose and fat metabolism in individuals with the TT genotype/ T allele.
Conversely, groups less represented in the TT genotype included the R. gauvreauii group, Catenibacterium, Holdemanella, and the Christensenellaceae_R-7 group. The R. gauvreauii group is a bile-resistant acetate acid producer using sucrose and sugar alcohols as carbon sources (48). R. gauvreauii, stands out for its inability to metabolize resistant starches but proficient growth on sugar alcohols (49). This unique metabolic behavior may have implications for substrate utilization within the gut. It has been observed that R. gauvreauii increases in gut microbiota of obese men post exercise and is positively associated with respiratory capacity and negatively with visceral fat (50). Catenibacterium, a member of the Christensenellaceae family, has been linked to leanness and lower BMI, suggesting roles in energy metabolism and dietary fiber digestion. Its species produce acetic, lactic, butyric, and isobutyric acids from sugars (51). However, its health impacts are not fully unclear. While reduced Catenibacterium abundance has been linked to obesity (52), conflicting evidence showed its enrichment in obesity cases (53). High Catenibacterium levels have also been connected to lower trimethylamine oxide (TMAO) levels, a cardiovascular disease-related metabolite (54). Holdemanella, a relatively unknown genus, has been investigated for potential roles in fermentation and anti-inflammatory properties, particularly in colitis contexts. It is involved in carbohydrate metabolism and associated with health benefits due to its anti-inflammatory properties (55) and regulatory effects on glucose metabolism (56). However, links to pathological states such as thyroid cancer have been observed (57). Different Holdemanella taxa have been positively associated with android-type obesity in men and negatively in women (58). Lastly, the Christensenellaceae_R7 group, more prevalent in the CC genotype, is a heritable microbiota component (59) associated with metabolic health in children (60) and adults (61). It has also been linked to lower visceral fat and lean mass in several studies (62–64).
In the comparative analysis of metabolic profiles between genotypes CC and TT, the TT group showed an overexpression of the KDD-NAGLIPASYN-PWY pathway. This pathway is involved in the biosynthesis of lipid A, a key component of lipopolysaccharides (LPS) present in Gram-negative bacteria’s the outer membrane. This suggests a higher presence of Gram-negative bacteria or a specific immunological response in the TT group (16). When analyzing metabolic differences between genotypes CC and TC, pathways related to coenzyme Q (CoQ) biosynthesis, particularly the UBISYN-PWY pathway, were associated with the TC group. Ubiquinol-8, essential for the electron transport chain and cellular energy production, was significantly overexpressed in the TC group, indicating potential implications in metabolism and inflammation (65). In the comparison of CC vs. TT_TC genotypes, CoQ biosynthesis pathways (PWY-6708, PWY-5857, and PWY-5856) were also prominent in the TC group, emphasizing the importance of CoQ biosynthesis in this context. Furthermore, the TT_TC group showed overexpression of the PWY-7013 pathway (nivalenol biosynthesis and propane 1,2 diol degradation) indicating an increased degradation of this compound, potentially affecting energy metabolism or substrate utilization. In the TC_CC vs. TT comparison, pathways associated with menaquinone biosynthesis (PWY-5853) were also overexpressed in the TC group. Menaquinones, types of vitamin K produced by certain bacteria, are involved in electron transport. These observed differences may reflect alterations in the presence or metabolic activity of vitamin K-producing bacteria.
The results reveal substantial differences in metabolic pathways among the studied groups (CC, TT, TC, TT_TC), emphasizing the significance of lipid A and CoQ biosynthesis, as along with changes in the degradation of specific compounds and the biosynthesis of menaquinones. These findings suggest potential associations with specific VDR gene profiles.
Polymorphisms in the VDR gene can lead to significant differences in gut microbiota composition and diversity through several key mechanisms. The VDR gene encodes the vitamin D receptor, which regulates the immune system by influencing the expression of antimicrobial peptides that maintain the gut barrier and control pathogenic bacteria (66). VDR polymorphisms can alter immune responses, affecting gut inflammation levels and creating environments that favor certain microbial communities. Additionally, the VDR gene is expressed in gut epithelial cells, where it regulates cell proliferation, differentiation, and apoptosis processes vital for maintaining gut barrier integrity (5). Disruptions in VDR activity can influence which microbial species thrive, as evidenced by VDR knockout mice exhibiting distinct microbial profiles (20, 67). Furthermore, VDR polymorphisms may affect vitamin D status, impacting nutrient absorption and metabolism, which in turn alters the gut environment and influences microbial growth (68).
The VDR gene TaqI polymorphism can directly affect the composition and functionality of the gut microbiome due to the critical role that vitamin D signaling plays in immune regulation and gut health. Changes in microbiota due to differences in the TaqI genotypes could be related to different gene transcription associated with the genotypes. However, there are inconsistent results regarding which genotypes produce higher levels of mRNA expression, although there appears to be a general trend indicating that the C allele demonstrates a more favorable response than the T allele (7).
The TT genotype of the TaqI polymorphism may be associated with differential modulation in vitamin D levels or VDR activity which could favor an environment that benefits certain bacterial genera. This could be an environment that promotes the production or utilization of SCFAs, thereby benefiting the proliferation of bacteria involved in butyrate production and other SCFAs such as Butyricimonas and Parabacteroides. VDR is also involved in the regulation of bile acid metabolism. Thus, alterations in VDR signaling due to the TT genotype could modify bile acid composition in the gut, facilitating the growth of genera like Parabacteroides, with capability to metabolize bile acids. Moreover, for the TT genotype, there could be differential modulation of cytokines and other immunomodulatory molecules, creating an intestinal environment that favors the proliferation of specific bacteria such as Butyricimonas and Parabacteroides.
Overall, VDR gene polymorphisms can lead to altered vitamin D signaling, which impacts immune regulation, gut barrier integrity, and the gut environment, ultimately influencing the composition and diversity of the gut microbiome. These changes can have significant consequences for host health, including impacts on immune function, inflammation, nutrient absorption, and susceptibility to various diseases.
A limitation of the present study is that although we used a well-defined sample of healthy participants, the study had a relatively small sample size, which may limit the generalizability of the findings to larger populations. However, studying the association between genetic variants and the microbiota in healthy volunteers facilitates the examination of the direct influence of genetic variants on microbiota composition without confounding pathological factors. Moreover, in our study potential confounding factors, such as sex, dietary habits and smoking has been considered.
5 Conclusion
Our research work suggests that polymorphisms in the VDR gene may be linked to specific microbiota genera compositions and functionalities. This study highlights the importance of considering genetic variations, like VDR gene polymorphisms, to grasp the intricate interplay between host genetics and gut microbiota, which may have implications for personalized nutrition and health interventions. Further research is essential to unravel the precise mechanisms by which VDR gene polymorphisms influence gut microbiota composition and function, thereby setting the stage for future investigations and potential therapeutic approaches.
Knowledge of how VDR polymorphisms impact gut microbiota can contribute to the development of personalized medical approaches. Individuals with certain VDR gene variants might benefit from tailored probiotic or dietary interventions to optimize gut health.
Data availability statement
The data presented in the study are deposited in the NCBI BioProject repository, accession number PRJNA1152925.
Ethics statement
The studies involving humans were approved by Research Ethics Committee of the Community of Madrid. The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
Author contributions
RS: Conceptualization, Formal analysis, Funding acquisition, Investigation, Methodology, Validation, Writing – original draft, Writing – review & editing. MT: Investigation, Methodology, Writing – review & editing. ML: Funding acquisition, Validation, Writing – original draft, Writing – review & editing. MB: Formal analysis, Methodology, Software, Validation, Writing – original draft, Writing – review & editing. CB: Formal analysis, Methodology, Software, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This research was supported by the Ramón y Cajal program (grant 2012_11910), AGL2016-77288-R project of the Ministry of Economy and Competitiveness, Spain, and the Universidad Europea de Madrid grants 2018/UEM4 and 2022/UEM28.
Acknowledgments
We acknowledge Maria Gregoria Montalvo and Catalina Santiago for the technical help and assistance for the genotyping experiments. OpenAI’s ChatGPT-4 have been used to assist in improving the clarity and fluency of the English language in this manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnut.2024.1423472/full#supplementary-material
Footnotes
References
1. Newmark, H, Dantoft, W, and Ghazal, P. Evolutionary origin of the interferon–immune metabolic Axis: the sterol–vitamin D link. Front Immunol. (2017) 8:62. doi: 10.3389/fimmu.2017.00062
2. Pike, JW, and Meyer, MB. The vitamin D receptor: new paradigms for the regulation of gene expression by 1,25-Dihydroxyvitamin D3. Endocrinol Metab Clin N Am. (2010) 39:255–69. doi: 10.1016/j.ecl.2010.02.007
3. Sun, J . Dietary vitamin D, vitamin D receptor, and microbiome. Curr Opin Clin Nutr Metab Care. (2018) 21:471–4. doi: 10.1097/MCO.0000000000000516
4. Uhlén, M, Fagerberg, L, Hallström, BM, Lindskog, C, Oksvold, P, Mardinoglu, A, et al. Tissue-based map of the human proteome. Science (1979). (2015) 347:1260419. doi: 10.1126/science.1260419
5. Bakke, D, and Sun, J. Ancient nuclear receptor VDR with new functions: microbiome and inflammation. Inflamm Bowel Dis. (2018) 24:1149–54. doi: 10.1093/ibd/izy092
6. Usategui-Martín, R, De Luis-Román, DA, Fernández-Gómez, JM, Ruiz-Mambrilla, M, and Pérez-Castrillón, JL. Vitamin D receptor (VDR) gene polymorphisms modify the response to vitamin D supplementation: a systematic review and Meta-analysis. Nutrients. (2022) 14:360. doi: 10.3390/nu14020360
7. Uitterlinden, AG, Fang, Y, Van Meurs, JBJ, Pols, HAP, and Van Leeuwen, JPTM. Genetics and biology of vitamin D receptor polymorphisms. Gene. (2004) 338:143–56. doi: 10.1016/j.gene.2004.05.014
8. Numakura, K, Satoh, S, Tsuchiya, N, Horikawa, Y, Inoue, T, Kakinuma, H, et al. Incidence and risk factors of clinical characteristics, tacrolimus pharmacokinetics, and related genomic polymorphisms for Posttransplant diabetes mellitus in the early stage of renal transplant recipients. Transplant Proc. (2005) 37:1865–7. doi: 10.1016/j.transproceed.2005.02.086
9. Keen, RW, Hart, DJ, Lanchbury, JS, and Spector, TD. Association of early osteoarthritis of THE knee with a Taq I polymorphism of the vitamin D receptor gene. Arthritis Rheum. (1997) 40:1444–9. doi: 10.1002/art.1780400812
10. Valdivielso, JM, and Fernandez, E. Vitamin D receptor polymorphisms and diseases. Clin Chim Acta. (2006) 371:1–12. doi: 10.1016/j.cca.2006.02.016
11. Vasilopoulos, Y, Sarafidou, T, Kotsa, K, Papadimitriou, M, Goutzelas, Y, Stamatis, C, et al. VDR TaqI is associated with obesity in the Greek population. Gene. (2013) 512:237–9. doi: 10.1016/j.gene.2012.10.044
12. Rashidi, F, and Ostadsharif, M. Association of VDR gene ApaI polymorphism with obesity in Iranian population. Biomedica. (2021) 41:651–9. doi: 10.7705/biomedica.5898
13. Gulati, S, Misra, A, Tiwari, R, Sharma, M, Pandey, RM, and Upadhyay, AD. The influence of polymorphisms of fat mass and obesity (FTO, rs9939609) and vitamin D receptor (VDR, BsmI, TaqI, ApaI, FokI) genes on weight loss by diet and exercise interventions in non-diabetic overweight/obese Asian Indians in North India. Eur J Clin Nutr. (2020) 74:604–12. doi: 10.1038/s41430-020-0560-4
14. Vimaleswaran, KS, Berry, DJ, Lu, C, Tikkanen, E, Pilz, S, Hiraki, LT, et al. Causal relationship between obesity and vitamin D status: Bi-directional Mendelian randomization analysis of multiple cohorts. PLoS Med. (2013) 10:e1001383. doi: 10.1371/journal.pmed.1001383
15. Mezza, T, Muscogiuri, G, Sorice, GP, Prioletta, A, Salomone, E, Pontecorvi, A, et al. Vitamin D deficiency: a new risk factor for type 2 diabetes. Ann Nutr Metab. (2012) 61:337–48. doi: 10.1159/000342771
16. Wang, J, Thingholm, LB, Skiecevičienė, J, Rausch, P, Kummen, M, Hov, JR, et al. Genome-wide association analysis identifies variation in vitamin D receptor and other host factors influencing the gut microbiota. Nat Genet. (2016) 48:1396–406. doi: 10.1038/ng.3695
17. Ooi, JH, Li, Y, Rogers, CJ, and Cantorna, MT. Vitamin D regulates the gut microbiome and protects mice from dextran sodium sulfate–induced colitis. J Nutr. (2013) 143:1679–86. doi: 10.3945/jn.113.180794
18. Chen, J, Waddell, A, Lin, YD, and Cantorna, MT. Dysbiosis caused by vitamin D receptor deficiency confers colonization resistance to Citrobacter rodentium through modulation of innate lymphoid cells. Mucosal Immunol. (2015) 8:618–26. doi: 10.1038/mi.2014.94
19. Jin, D, Wu, S, Zhang, Y, Guo, LR, Xia, Y, Dong, H, et al. Lack of vitamin D receptor causes Dysbiosis and changes the functions of the murine intestinal microbiome. Clin Ther. (2015) 37:996–1009.e7. doi: 10.1016/j.clinthera.2015.04.004
20. Wu, S, Zhang, Y, Guo, LR, Xia, Y, Zhou, D, Petrof, EO, et al. Intestinal epithelial vitamin D receptor deletion leads to defective autophagy in colitis. Gut. (2015) 64:1082–94. doi: 10.1136/gutjnl-2014-307436
21. Jesús, Vioque López, and Quiles Izquierdo, J. Encuesta de Nutrición y Salud de la Comunidad Valenciana. Departamento Salud Pública, Historia de la Ciencia y Ginecología-Universidad Miguel Hernández. (2003).
22. Solé, X, Guinó, E, Valls, J, Iniesta, R, and Moreno, V. SNPStats: a web tool for the analysis of association studies. Bioinformatics. (2006) 22:1928–9. doi: 10.1093/bioinformatics/btl268
23. Bolyen, E, Rideout, JR, Dillon, MR, Bokulich, NA, Abnet, CC, Al-Ghalith, GA, et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat Biotechnol. (2019) 37:852–7. doi: 10.1038/s41587-019-0252-6
24. Callahan, BJ, McMurdie, PJ, Rosen, MJ, Han, AW, Johnson, AJA, and Holmes, SP. DADA2: high-resolution sample inference from Illumina amplicon data. Nat Methods. (2016) 13:581–3. doi: 10.1038/nmeth.3869
25. Pedregosa Fabianpedregosa, F, Michel, V, Grisel Oliviergrisel, O, Blondel, M, Prettenhofer, P, Weiss, R, et al. Scikit-learn: machine learning in Python Gaël Varoquaux Bertrand Thirion Vincent Dubourg Alexandre Passos Pedregosa, Varoquaux, Gramfort Et Al. J Mach Learn Res. (2011) 12:2825–30. doi: 10.48550/arXiv.1201.0490
26. Quast, C, Pruesse, E, Yilmaz, P, Gerken, J, Schweer, T, Yarza, P, et al. The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res. (2012) 41:D590–6. doi: 10.1093/nar/gks1219
27. Yilmaz, P, Parfrey, LW, Yarza, P, Gerken, J, Pruesse, E, Quast, C, et al. The SILVA and “all-species living tree project (LTP)” taxonomic frameworks. Nucleic Acids Res. (2014) 42:D643–8. doi: 10.1093/nar/gkt1209
28. Bokulich, NA, Kaehler, BD, Rideout, JR, Dillon, M, Bolyen, E, Knight, R, et al. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome. (2018) 6:90. doi: 10.1186/s40168-018-0470-z
29. Kaehler, BD, Bokulich, NA, McDonald, D, Knight, R, Caporaso, JG, and Huttley, GA. Species abundance information improves sequence taxonomy classification accuracy. Nat Commun. (2019) 10:4643. doi: 10.1038/s41467-019-12669-6
30. Douglas, GM, Maffei, VJ, Zaneveld, JR, Yurgel, SN, Brown, JR, Taylor, CM, et al. PICRUSt2 for prediction of metagenome functions. Nat Biotechnol. (2020) 38:685–8. doi: 10.1038/s41587-020-0548-6
31. McMurdie, PJ, and Holmes, S. Phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS One. (2013) 8:e61217. doi: 10.1371/journal.pone.0061217
32. Cao, Y, Dong, Q, Wang, D, Zhang, P, Liu, Y, and Niu, C. microbiomeMarker: an R/Bioconductor package for microbiome marker identification and visualization. Marschall T, editor. Bioinformatics. (2022) 38:4027–9. doi: 10.1093/bioinformatics/btac438
33. Kruskal, WH, and Wallis, WA. Use of ranks in one-criterion variance analysis. J Am Stat Assoc. (1952) 47:583–621. doi: 10.1080/01621459.1952.10483441
34. Anders, S, McCarthy, DJ, Chen, Y, Okoniewski, M, Smyth, GK, Huber, W, et al. Count-based differential expression analysis of RNA sequencing data using R and Bioconductor. Nat Protoc. (2013) 8:1765–86. doi: 10.1038/nprot.2013.099
35. Lin, H, and Das, PS. Analysis of compositions of microbiomes with bias correction. Nat Commun. (2020) 11:3514. doi: 10.1038/s41467-020-17041-7
36. Love, MI, Huber, W, and Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. (2014) 15:550. doi: 10.1186/s13059-014-0550-8
37. Calgaro, M, Romualdi, C, Waldron, L, Risso, D, and Vitulo, N. Assessment of statistical methods from single cell, bulk RNA-seq, and metagenomics applied to microbiome data. Genome Biol. (2020) 21:191. doi: 10.1186/s13059-020-02104-1
38. Davenport, ER, Sanders, JG, Song, SJ, Amato, KR, Clark, AG, and Knight, R. The human microbiome in evolution. BMC Biol. (2017) 15:127. doi: 10.1186/s12915-017-0454-7
39. Cui, Y, Zhang, L, Wang, X, Yi, Y, Shan, Y, Liu, B, et al. Roles of intestinal Parabacteroides in human health and diseases. FEMS Microbiol Lett. (2022) 369:fnac072. doi: 10.1093/femsle/fnac072
40. den Besten, G, van Eunen, K, Groen, AK, Venema, K, Reijngoud, DJ, and Bakker, BM. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J Lipid Res. (2013) 54:2325–40. doi: 10.1194/jlr.R036012
41. de la Cuesta-Zuluaga, J, Mueller, N, Álvarez-Quintero, R, Velásquez-Mejía, E, Sierra, J, Corrales-Agudelo, V, et al. Higher fecal short-chain fatty acid levels are associated with gut microbiome Dysbiosis, obesity, hypertension and Cardiometabolic disease risk factors. Nutrients. (2018) 11:51. doi: 10.3390/nu11010051
42. Bohdanowicz-Pawlak, A, and Lwow, F. Differences in chronic low-grade inflammation and metabolic disturbances between VDR genotypes in an ethnically homogenous postmenopausal female population from Poland. Nutrients. (2023) 15:2737. doi: 10.3390/nu15122737
43. Lee, H, An, J, Kim, J, Choi, D, Song, Y, Lee, CK, et al. A novel bacterium, Butyricimonas virosa, preventing HFD-induced diabetes and metabolic disorders in mice via GLP-1 receptor. Front Microbiol. (2022) 13:858192. doi: 10.3389/fmicb.2022.858192
44. Zoetendal, EG, Plugge, CM, Akkermans, ADL, and de Vos, WM. Victivallis vadensis gen. Nov., sp. nov., a sugar-fermenting anaerobe from human faeces. Int J Syst Evol Microbiol. (2003) 53:211–5. doi: 10.1099/ijs.0.02362-0
45. Rodriguez, J, Hiel, S, Neyrinck, AM, Le Roy, T, Pötgens, SA, Leyrolle, Q, et al. Discovery of the gut microbial signature driving the efficacy of prebiotic intervention in obese patients. Gut. (2020) 69:1975–87. doi: 10.1136/gutjnl-2019-319726
46. Li, H, Fang, Q, Nie, Q, Hu, J, Yang, C, Huang, T, et al. Hypoglycemic and Hypolipidemic mechanism of tea polysaccharides on type 2 diabetic rats via gut microbiota and metabolism alteration. J Agric Food Chem. (2020) 68:10015–28. doi: 10.1021/acs.jafc.0c01968
47. Morales, C, Rojas, G, Rebolledo, C, Rojas-Herrera, M, Arias-Carrasco, R, Cuadros-Orellana, S, et al. Characterization of microbial communities from gut microbiota of hypercholesterolemic and control subjects. Front Cell Infect Microbiol. (2022) 12:943609. doi: 10.3389/fcimb.2022.943609
48. Molinero, N, Conti, E, Walker, AW, Margolles, A, Duncan, SH, and Delgado, S. Survival strategies and metabolic interactions between Ruminococcus gauvreauii andRuminococcoides bili, isolated from human bile. Microbiol Spectr. (2022) 10:e02776–21. doi: 10.1128/spectrum.02776-21
49. Molinero, N, Conti, E, Sánchez, B, Walker, AW, Margolles, A, Duncan, SH, et al. Ruminococcoides bili gen. Nov., sp. nov., a bile-resistant bacterium from human bile with autolytic behavior. Int J Syst Evol Microbiol. (2021) 71:004960. doi: 10.1099/ijsem.0.004960
50. Verheggen, RJHM, Konstanti, P, Smidt, H, Hermus, ARMM, Thijssen, DHJ, and Hopman, MTE. Eight-week exercise training in humans with obesity: marked improvements in insulin sensitivity and modest changes in gut microbiome. Obesity (Silver Spring). (2021) 29:1615–24. doi: 10.1002/oby.23252
51. Rainey, FA . Catenibacterium In: WB Whitman , editor. Bergey’s manual of systematics of Archaea and Bacteria. 1st ed. Hoboken, New Jersey: Wiley (2015). 1–2.
52. Pinart, M, Dötsch, A, Schlicht, K, Laudes, M, Bouwman, J, Forslund, SK, et al. Gut microbiome composition in obese and non-obese persons: a systematic review and Meta-analysis. Nutrients. (2021) 14:12. doi: 10.3390/nu14010012
53. Martínez-Cuesta, MC, del Campo, R, Garriga-García, M, Peláez, C, and Requena, T. Taxonomic characterization and short-chain fatty acids production of the obese microbiota. Front Cell Infect Microbiol. (2021) 11:598093. doi: 10.3389/fcimb.2021.598093
54. James, KL, Gertz, ER, Cervantes, E, Bonnel, EL, Stephensen, CB, Kable, ME, et al. Diet, fecal microbiome, and trimethylamine N-oxide in a cohort of metabolically healthy United States adults. Nutrients. (2022) 14:1376. doi: 10.3390/nu14071376
55. Pujo, J, Petitfils, C, Le Faouder, P, Eeckhaut, V, Payros, G, Maurel, S, et al. Bacteria-derived long chain fatty acid exhibits anti-inflammatory properties in colitis. Gut. (2021) 70:1088–97. doi: 10.1136/gutjnl-2020-321173
56. Romaní-Pérez, M, López-Almela, I, Bullich-Vilarrubias, C, Rueda-Ruzafa, L, Gómez Del Pulgar, EM, Benítez-Páez, A, et al. Holdemanella biformis improves glucose tolerance and regulates GLP-1 signaling in obese mice. FASEB J. (2021) 35:e21734. doi: 10.1096/fj.202100126R
57. Hou, T, Wang, Q, Dai, H, Hou, Y, Zheng, J, Wang, T, et al. Interactive association between gut microbiota and thyroid cancer. Endocrinology. (2023) 165:bqad184. doi: 10.1210/endocr/bqad184
58. Min, Y, Ma, X, Sankaran, K, Ru, Y, Chen, L, Baiocchi, M, et al. Sex-specific association between gut microbiome and fat distribution. Nat Commun. (2019) 10:2408. doi: 10.1038/s41467-019-10440-5
59. Goodrich, JK, Waters, JL, Poole, AC, Sutter, JL, Koren, O, Blekhman, R, et al. Human genetics shape the gut microbiome. Cell. (2014) 159:789–99. doi: 10.1016/j.cell.2014.09.053
60. Alcazar, M, Escribano, J, Ferré, N, Closa-Monasterolo, R, Selma-Royo, M, Feliu, A, et al. Gut microbiota is associated with metabolic health in children with obesity. Clin Nutr. (2022) 41:1680–8. doi: 10.1016/j.clnu.2022.06.007
61. He, Y, Wu, W, Wu, S, Zheng, HM, Li, P, Sheng, HF, et al. Linking gut microbiota, metabolic syndrome and economic status based on a population-level analysis. Microbiome. (2018) 6:172. doi: 10.1186/s40168-018-0557-6
62. Beaumont, M, Goodrich, JK, Jackson, MA, Yet, I, Davenport, ER, Vieira-Silva, S, et al. Heritable components of the human fecal microbiome are associated with visceral fat. Genome Biol. (2016) 17:189. doi: 10.1186/s13059-016-1052-7
63. Oki, K, Toyama, M, Banno, T, Chonan, O, Benno, Y, and Watanabe, K. Comprehensive analysis of the fecal microbiota of healthy Japanese adults reveals a new bacterial lineage associated with a phenotype characterized by a high frequency of bowel movements and a lean body type. BMC Microbiol. (2016) 16:284. doi: 10.1186/s12866-016-0898-x
64. Tavella, T, Rampelli, S, Guidarelli, G, Bazzocchi, A, Gasperini, C, Pujos-Guillot, E, et al. Elevated gut microbiome abundance of Christensenellaceae, Porphyromonadaceae and Rikenellaceae is associated with reduced visceral adipose tissue and healthier metabolic profile in Italian elderly. Gut Microbes. (2021) 13:1–19. doi: 10.1080/19490976.2021.1880221
65. Pallotti, F, Bergamini, C, Lamperti, C, and Fato, R. The roles of coenzyme Q in disease: direct and indirect involvement in cellular functions. Int J Mol Sci. (2021) 23:128. doi: 10.3390/ijms23010128
66. Clark, A, and Mach, N. Role of vitamin D in the hygiene hypothesis: the interplay between vitamin D, vitamin D receptors, gut microbiota, and immune response. Front Immunol. (2016) 7:27. doi: 10.3389/fimmu.2016.00627
67. Shang, M, and Sun, J. Vitamin D/VDR, probiotics, and gastrointestinal diseases. Curr Med Chem. (2017) 24:876–87. doi: 10.2174/0929867323666161202150008
Keywords: VDR, microbiota, TaqI (rs731236), microbiome, polymorphism
Citation: Gonzalez-Soltero R, Tabone M, Larrosa M, Bailen M and Bressa C (2024) VDR gene TaqI (rs731236) polymorphism affects gut microbiota diversity and composition in a Caucasian population. Front. Nutr. 11:1423472. doi: 10.3389/fnut.2024.1423472
Edited by:
Clara G. De Los Reyes-Gavilan, Spanish National Research Council (CSIC), SpainReviewed by:
Sonia Roman, University of Guadalajara, MexicoLukasz Laczmanski, Polish Academy of Sciences, Poland
Copyright © 2024 Gonzalez-Soltero, Tabone, Larrosa, Bailen and Bressa. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Maria Bailen, bWFyaWEuYmFpbGVuQHVhbS5lcw==