 Nathaniel Mengers
Nathaniel Mengers Natasha Rouse
Natasha Rouse Kathryn A. Daltorio
Kathryn A. Daltorio- Biologically Inspired Robotics Laboratory, Mechanical and Aerospace Engineering, Case Western Reserve University, Cleveland, OH, United States
Stable Heteroclinic Channels (SHCs) are dynamical systems composed of connected saddle equilibria. This work demonstrates a control system that combines SHCs with movement primitives to enable swimming in a simulated six segment snake robot. We identify control system parameters for lateral undulation, where all joints oscillate with the same amplitude, and anguilliform swimming, where joint amplitudes increase linearly from the head to the tail. Swimming speed is improved by learning SHC movement primitive parameters. We also propose a method for adapting the gait amplitude and frequency with tactile sensor input to accommodate obstacles. Then, we evaluate the relationship between SHC movement primitive parameters and the resulting trajectories. The swimming speed and efficiency of SHC controllers for each gait are compared against a conventional serpenoid controller, which derives joint trajectories from sinusoids. Controllers are evaluated first in an unobstructed environment, then in straight passages of various widths, and finally in 65 randomly generated uneven channels. We find that the amplitudes of joint oscillations scale proportionally with the SHC controller parameters. Due to gait optimization, as well as adaptive amplitude and frequency in response to tactile input, the learned SHC control system exhibits an average 28.8% greater speed than a serpenoid controller that only adapts amplitude during contact. This research demonstrates that SHCs benefit from intuitive tuning like serpenoid control, while also effectively incorporating sensory information to generate smooth kinematic trajectories.
1 Introduction
Modeling neuroanatomical structures not only supports biological research, but advances robotics by inspiring frameworks for adaptive, flexible control. Motor primitives (MPs) describe fundamental units of actuation that are assembled to produce complex behavior (Giszter, 2015). MP models in robots offer modular (Paraschos et al., 2013), learnable (Kober and Peters, 2009) control with efficient sensor integration (Bonardi et al., 2012). Meanwhile, central pattern generators (CPGs) are neural circuits that produce rhythms without rhythmic input (Ijspeert, 2008) for locomotion and other functions (Sherrington, 1910; Grillner and Wallén, 2010; Cropper et al., 2004). Hierarchical MP and CPG combinations have been proposed for adaptable rhythms in biological (Guertin, 2013) and mechanical contexts (Moreno and Gomez, 2011). However, relationships between MP parameters and resulting behaviors are nonlinear, complicating modification and analysis (Rouse and Daltorio, 2021). We demonstrate that kinematic MPs incorporating Stable Heteroclinic Channels (SHCs), an emerging CPG model (Daltorio et al., 2013; Brecelj and Petrič, 2023a), enable transparent, learnable, and sensor-adaptable control in a simulated swimming snake robot. In obstacle-free and confined environments, we compare speed and efficiency for SHC controllers with a serpenoid controller, which prescribes joint angles via sinusoids. We further show that incorporating tactile sensing improves speed in confined spaces.
MPs have been used to both model animal behavior (Tresch and Bizzi, 1999; Fod et al., 2002; Tagliabue et al., 2015) and plan robot motion (Ijspeert et al., 2013). MPs arise when modular clusters of neurons and muscle cells reliably produce specific kinematic trajectories or force outputs (Giszter, 2015). Dynamic Movement Primitives (DMPs) are a popular framework for constructing kinematic MPs in robots (Paraschos et al., 2013; Hoffmann et al., 2009; Pastor et al., 2009; Schaal, 2006; Kober and Peters, 2009). They rely on sequentially arranged, weighted Gaussian distributions (kernels), where weights are learned from demonstration to produce arbitrary trajectories (Schaal, 2006; Pastor et al., 2009; Kong et al., 2023). Sensory inputs adapt the planned trajectory, allowing robots to interact with their environment Paraschos et al. (2013). Periodically activating kernels yields rhythmic behavior (Ijspeert et al., 2013). The mapping from learned parameters to the planned trajectory is nonlinear (Rouse and Daltorio, 2021). Thus, the controller behaves as a black box, making it difficult to discern how adjusting the weights may influence robot behavior.
Like MPs, CPGs are an important aspect of biological motor function, and have been modeled for robotic control. Biological CPGs can emerge from a pacemaker neuron that rhythmically self excites (Harris-Warrick, 2010; Marder and Eisen, 1984), or mutually inhibitory, modular neuron clusters (e.g., half-center oscillators) (Ijspeert, 2008; Grillner and Wallén, 2010). The peripheral nervous system can modulate rhythms by inhibiting or exciting CPG neurons (Andersson et al., 1978). In addition to modeling neural connections, biomimetic CPG simulate cellular processes using Hodgkin-Huxley (HH) (Simoni and DeWeerth, 2007) dynamics or approximations such as leaky integrators (Yu et al., 2014). More abstract models use limit cycles, or coupled oscillatory differential equations, to approximate neuron clusters without reproducing cellular dynamics (Wang Z. et al., 2017; Ijspeert et al., 2013). Couplings represent relationships between joints, as well as extensors and flexors for a single joint (Thandiackal et al., 2021). Biomimetic models offer insight into mechanisms for rhythmogenesis, while abstractions address neuron cluster interactions (Ijspeert, 2008). Modularity and sensor integration make both types of CPGs attractive tools for rhythmic control in robots (Hunt et al., 2017; Ijspeert et al., 2007).
Segmental robots based on snakes (Wang J. et al., 2017), salamanders (Ijspeert et al., 2007), and worms (Riddle et al., 2023) are useful for examining CPG controllers due to their modular construction and variety of motor patterns. Biomimetic CPGs have been demonstrated for controlling a biohybrid snake robot, where light responsive cells induce turning by stimulating neurons that control the anterior joint (Cheslet et al., 2024). Additionally, Norman-Tenazas (2021) algorithmically evaluates neuron configurations for CPGs with leaky-integrator dynamics, then applies the best performer to a snake robot with motor torque sensing for navigating peg arrays. More abstract models emphasizing CPG connectivity have been examined as well. For example, Thandiackal et al. (2021) applies Matsuoka oscillators to show that intrinsic joint oscillations, intersegmental coupling, and reflex chains sensitive to fluid forces can all drive undulation, though a combined approach is more robust. Meanwhile, Moreno and Gomez (2011) use motor primitives to adapt CPG amplitude and frequency. Combined with range sensors on the front and sides of the robot, the hierarchical structure enables the robot to avoid and climb over obstacles.
While abstract CPGs based on limit cycles have demonstrated effective control in segmented robots, some research suggests that CPGs constructed with stable heteroclinic channels (SHCs) are more responsive to sensory inputs (Horchler et al., 2015; Rabinovich et al., 2006; Shaw et al., 2015). SHCs consist of saddle equilibria connected cyclically such that the unstable manifold of one flows into the stable manifold of the next (Horchler et al., 2015). Perturbing the system away from equilibria with noise or sensor input causes state transitions, analogous to pushing a ball off of a hill Horchler et al. (2015). Perturbation intensity and timing influence residence times near equilibria (Horchler et al., 2015; Rouse et al., 2024). Ashwin and Postlethwaite (2016); Ashwin et al. (2011) describe heteroclinic cycle stability criteria with consideration to noise. Biological research on Aplysia shows that SHCs tuned for finite state machine-like transitions are responsive to sensor inputs and qualitatively reproduce in vivo feeding behaviors (Shaw et al., 2015). SHCs have also been applied in a simulated worm-like robot, with force sensors modulating transitions between expansion and contraction to improve locomotion through a pipe (Daltorio et al., 2013). Similarly, Brecelj and Petrič (2023b) use SHCs in a humanoid robot, where saddles correspond to stances (e.g., standing, raised hands) and forces on the grasper drive transitions. Recently, Rouse and Daltorio (2021) demonstrated that MPs based on SHCs instead of Gaussian kernels exhibit transparent control. Specifically, weights visually resemble the trajectory in a kinematic model.
Our research demonstrates movement primitives based on SHCs for control of a simulated snake robot, then assesses methods for adapting gaits by scaling and learning weight parameters, as well as incorporating sensory input. We validate the hypothesis that joint trajectories, which collectively constitute the gait, vary proportionally with SHC weight parameters. We exploit this characteristic to construct an anguilliform (eel-like) swimming gait from the lateral undulation gait. The anguilliform gait is characterized by smaller oscillations in anterior joints. We demonstrate that weights are learnable, enabling gait optimization with respect to swimming speed and efficiency. Finally, we show that gaits are enhanced by modifying the rate of progression between saddle points with sensory information. To our knowledge, this is the first work demonstrating correspondence between SHC movement primitive weights and the resulting gait in a nonlinear dynamical system.
To validate our controller, we implement both serpenoid and SHC control in a planar snake robot simulation and demonstrate that the controllers achieve comparable performance (i.e., efficiency and swimming speed) in quiescent fluid. We show that the anguiliform gait emerges by independently scaling the weight amplitudes for each joint, then characterize the relationship between weights and joint amplitudes (Figure 1). Separately, we apply gradient descent to optimize the SHC weights, improving the speed of the robot in an obstacle free environment. Then we integrate tactile sensing to increase the progression rate between saddle points during contact and assess the controller performance as the robot maneuvers through both straight and uneven passages. The efficiency and speed of the simulated robot with SHC control and compliant serpenoid control are compared. Our work serves as a basis for future controllers that adapt to fluid and obstacle interactions in a biologically plausible manner.
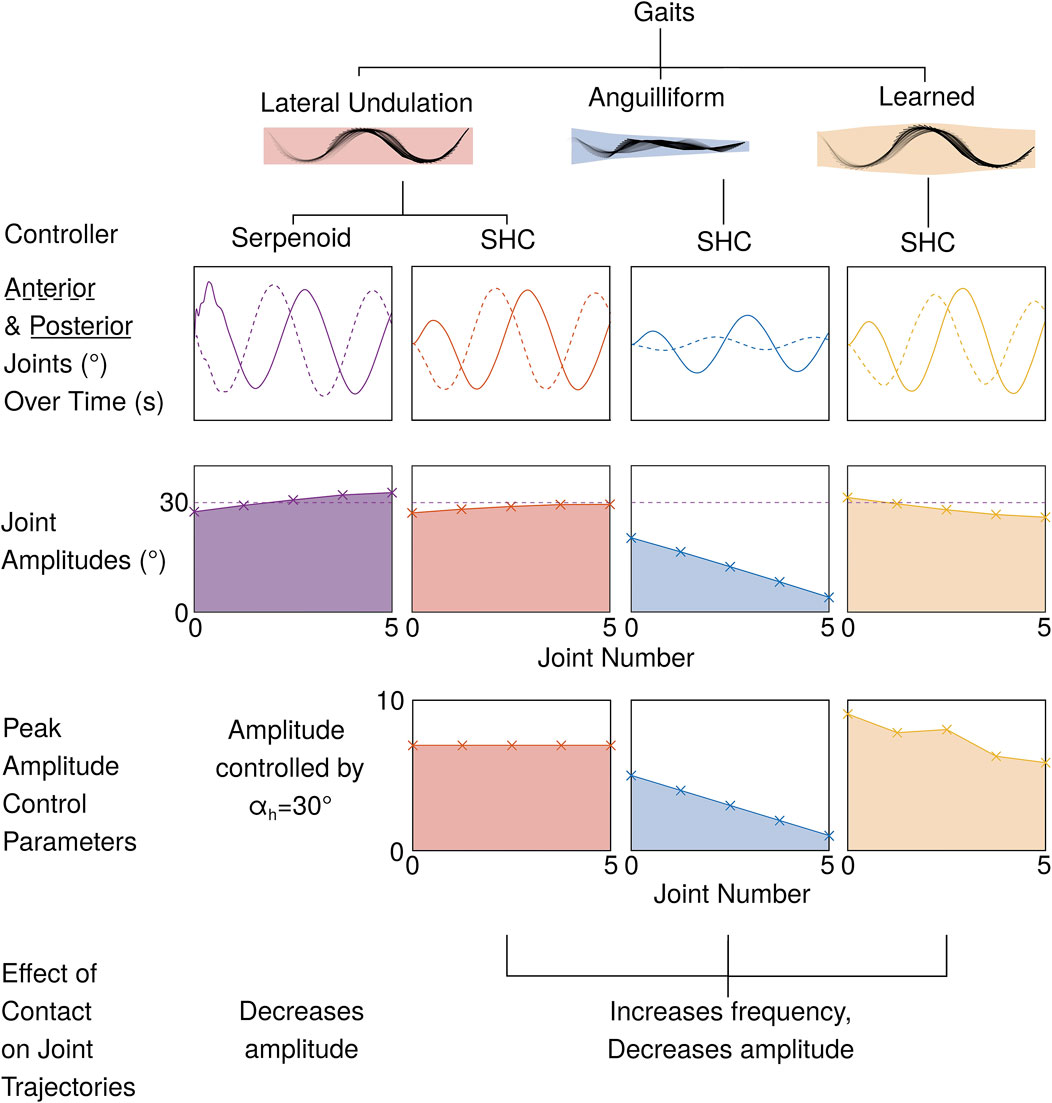
Figure 1. An SHC movement primitive controller, like a serpenoid controller, can realize a lateral undulation gait. With SHC movement primitives, joint amplitudes can be independently tuned by manually scaling weights, as shown by the anguilliform SHC controller. Alternatively, learning weights permits gait optimization to improve swimming speed (4 learned weights per joint; maximum weight for each joint is shown). In our SHC-based controllers, contact sensing produces a coupled change in the frequency and amplitude of the gait.
2 Methods
2.1 Modes of locomotion
Biological snakes use various gaits for different environmental conditions. In uncluttered terrestrial environments, snakes perform lateral undulation, where a transverse wave propagates from the head to the tail (Figure 2A). All points on the body trace the same path because ground forces prevent slipping in the transverse direction (Gray, 1946; Jayne, 2020). Aquatic lateral undulation, or anguilliform locomotion, is also characterized by a similar transverse wave, but the amplitude and wavelength increases from the head to the tail (Figure 2B). Different points trace distinct paths because there are no ground contact forces to facilitate nonholonomic constraints (Gray, 1946; Jayne, 2020). Instead, resistive and reactive fluid forces provide thrust (Piñeirua et al., 2015). Additionally, the muscle activation timing differs from terrestrial lateral undulation (Jayne, 2020).
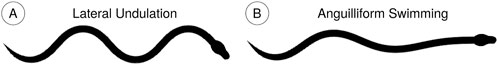
Figure 2. (A) During terrestrial lateral undulation, all joints oscillate with the same amplitude such that the lateral displacement remains the same at the front of the body as at the rear (B) In anguilliform locomotion, the lateral displacement is minimal near the head and increases posteriorally.
In loose or slippery substrates such as sand, snakes in the Caenophidia clade switch to a sidewinding gait (Mosauer, 1932). Sidewinding is characterized by anchoring some sections of the body on the ground, thrusting the unanchored sections to one side, then repeating, alternating the unanchored and anchored sections to produce lateral locomotion. In tunnel concertina locomotion, a snake will anchor itself against both sides of a narrow passage, extend forward by straightening its joints starting from the head, then anchor itself again (Gray, 1946). A variation of concertina locomotion is also used for arboreal locomotion during climbing or along thin branches (Astley and Jayne, 2007). In spaces too narrow for concertina locomotion, snakes may use a rectilinear gait where portions of the body are lifted off the ground and moved forward, akin to walking (Lissmann, 1950).
2.2 Control
2.2.1 Lateral undulation gait with serpenoid control
In early work parameterizing snake kinematics, Shigeo (1994) proposed the serpenoid curve, wherein sinusoids approximate the shape of an undulating snake. The desired relative joint angle
Hirose’s model has become popular for snake robots due to its simplicity and versatility. The serpenoid curve has been used for both terrestrial and aquatic lateral undulation (Shigeo, 1994; Ostrowski and Burdick, 1996; Sato et al., 2002; Kelasidi et al., 2014). It has also been adapted for sidewinding (Tesch et al., 2009), concertina (Chen and Roth, 2023), and rectilinear (Tesch et al., 2009) gaits. Additionally, several authors have augmented the serpenoid curve to support obstacle aided locomotion (Rollinson and Choset, 2013; Travers et al., 2018; Travers et al., 2015) and mitigate slipping (Dehghani and Mahjoob, 2009). In our work, we use it as a basis for comparison with our SHC controllers. We select gait parameters
To ensure locomotion along the
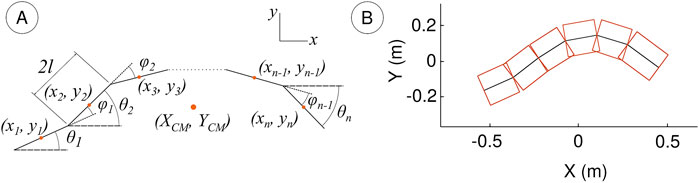
Figure 3. (A) Kinematic diagram of
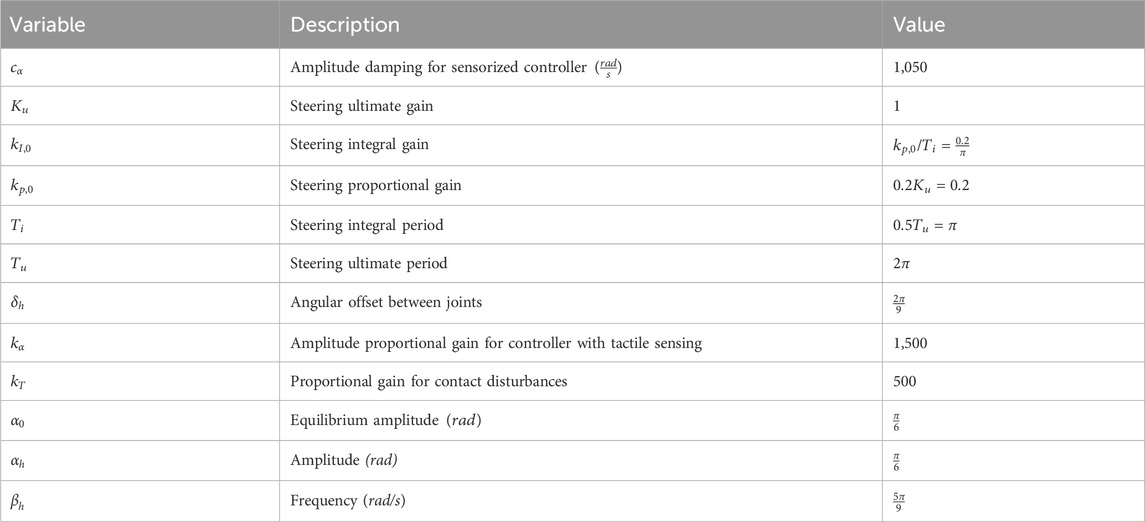
Table 1. Serpenoid controller parameters.
The actuator torques
2.2.2 Tactile sensing with serpenoid control
Trajectory planning is enhanced by considering the interactions between the robot and environmental obstacles. Travers et al. (2015) define shape compliant control, where serpenoid curve parameters vary due to obstacle contact. Specifically, their robot senses externally applied torques on each joint and adjusts the reference joint amplitudes using an admittance controller, enabling the robot to brace against obstacles (Equation 6).
In Travers et al. (2015), the admittance controller gains
We adopt a simplified shape controller based on Travers et al. (2015), where the robot narrows its gait by decreasing joint amplitudes during contact. Once contact ceases, the joint amplitudes return to the reference value
2.2.3 Lateral undulation gait with SHC control
2.2.3.1 SHC dynamics
To validate SHC-based MPs as a viable control method snake robots, we first emulate the lateral undulation gait employed by the serpenoid controller. SHCs encode rhythmic behavior as cyclic progression between saddle points or kernels. The kernel activity
With a phase offset of
Considering only the subformula
We refer to Rouse and Daltorio (2024) as a guideline for tuning the SHC MP controller. The study outlines how inhibition parameters impact kernel activation and trajectories as the controller traces a square. The growth rate
In addition to the excitatory and inhibitory components, the heteroclinic dynamics are augmented with Brownian noise via a time invariant Wiener process,
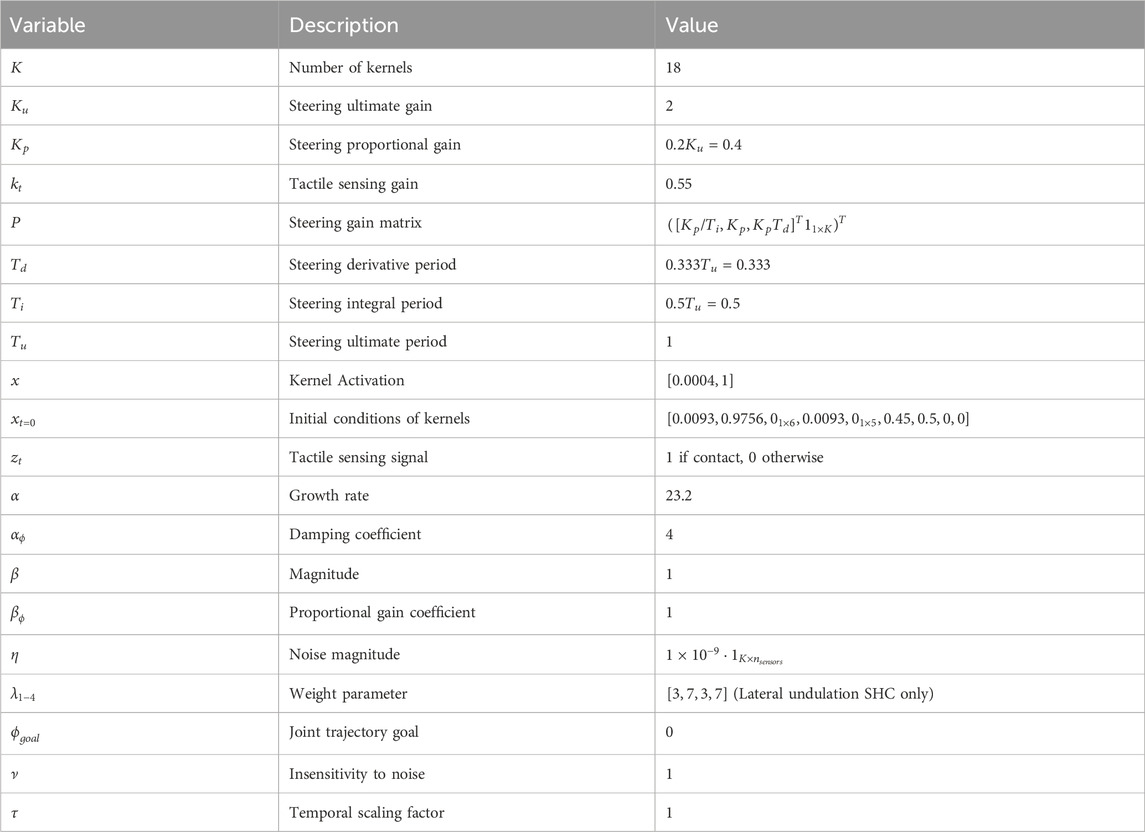
Table 2. SHC controller parameters.
2.2.3.2 Movement primitives with SHCs
To integrate SHCs with the MP framework, we define the desired joint accelerations
The first term constitutes a PD controller with gain parameters
The weight matrix
The weight submatrix
Once the desired joint accelerations, velocities, and positions are computed for a given time step, we evaluate the joint torques using the same methods as in Equation 5 in Section 2.2.1.
2.2.4 Anguilliform gait with SHC control
Replicating the anguilliform gait with SHC control provides an opportunity to assess the scalability of SHC movement primitive trajectories. In Rouse and Daltorio (2021), the authors demonstrate that weight parameters for SHC movement primitives visually represent and are proportional to the desired trajectory for a simple kinematic model. Inspired by their work, we examine how gaits can be adapted by scaling the weights. Specifically, we assess weight scaling as a means of switching from the terrestrial lateral undulation gait to anguilliform locomotion. Kelasidi et al. (2014) approximate anguilliform locomotion by linearly scaling the joint amplitudes in their serpenoid controller as in Equations 15, 16.
to implement an anguilliform gait with SHC control, we apply the scaling relationship in Equation 15 to the weight matrix (Equation 17). We expect that scaling the weights will proportionally change the amplitude of the trajectories for each joint.
2.2.5 SHC control with tactile sensing
CPGs, both in biological systems and robots, are most useful when they can be adapted by sensory information. Here, we establish a mechanism for incorporating tactile signals into SHC movement primitives, with the goal of improving speed of the simulated snake robot as it undulates through confined spaces. We introduce the term
2.2.6 Learned gait with SHC control
Robots, like animals, benefit from optimizing the speed and energy efficiency of locomotion. In (Rouse and Daltorio, 2021), the authors demonstrate that weights in SHC movement primitives can be learned to replicate a known trajectory for a purely kinematic system. Here, we extend their work, improving the robot’s performance by adjusting weights with consideration to the system dynamics. Beginning with the SHC controller for lateral undulation (see Section 2.2.5), we simulate swimming for
We establish a cost function (Equation 21) rewarding velocity, while penalizing poor efficiency. Note that the velocity is larger than the COT for lateral undulation such that this cost function prioritizes speed.
we numerically approximate the gradient of the cost function with respect to
2.3 Experimental design
2.3.1 Obstacle-free swimming
We first consider a simulated robot quiescent water with no obstacles. We adopt the framework proposed by Kelasidi et al. (2017), which addresses a submerged planar robot comprised of discrete linkages with elliptical cross sections. Fluid is viscous, incompressible, and irrotational in the reference frame of the robot. The authors develop equations of motion for the center of mass
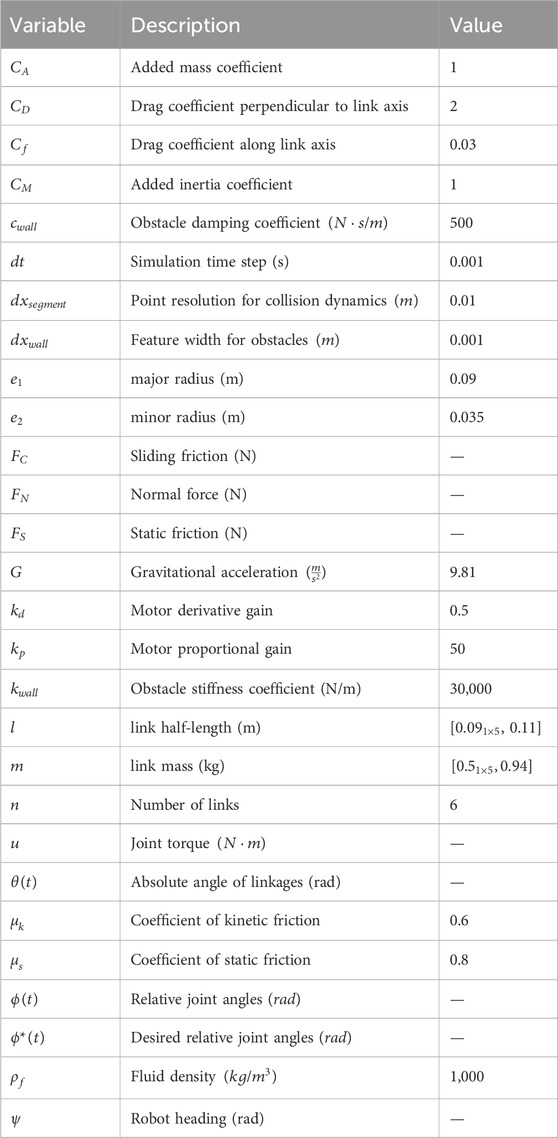
Table 3. Parameters for fluid dynamics, obstacle modeling, and kinematics.
The experiment is conducted for the serpenoid, lateral undulation SHC, anguilliform SHC, and learned SHC controllers. The simulation runs for 60 s, recording the robot’s position, joint torques, and joint velocities. Power consumption is approximated as
2.3.2 Straight channels
Simulating the robot in structured environments allows us to assess our method for integrating tactile information with SHC control. We examine straight passages ranging from
Obstacles are treated as points connected by flat surfaces with stiffness
2.3.3 Uneven channels
One of the most daunting aspects for exploratory robots is maintaining productive locomotion in complex terrain. We simulate the robot moving through channels with randomly generated feature (see examples in Figure 4). To establish consistent initial conditions, each channel begins with a
Figure 4. Examples of uneven channels with randomly generated features that impede the robot.
We simulate 60 s of swimming in each channel using the serpenoid and lateral undulation SHC controllers with and without tactile sensing. The learned SHC controller (+T) is also examined. Steering is disabled since the direction of the channel varies. We estimate the minimum number of channels required
3 Results
3.1 Obstacle-free swimming
3.1.1 Lateral undulation gait with SHC control
In quiescent fluid with no obstacles, the SHC controller successfully replicates the lateral undulation gait. The SHC controller produces similar joint oscillations to the serpenoid controller, but with a smoother transition from rest to steady state oscillation (Figure 5A). For both methods, the heading naturally oscillates during undulation. The steering controllers compensate by increasing or decreasing the desired joint trajectories, causing joint amplitudes to differ from the reference value of
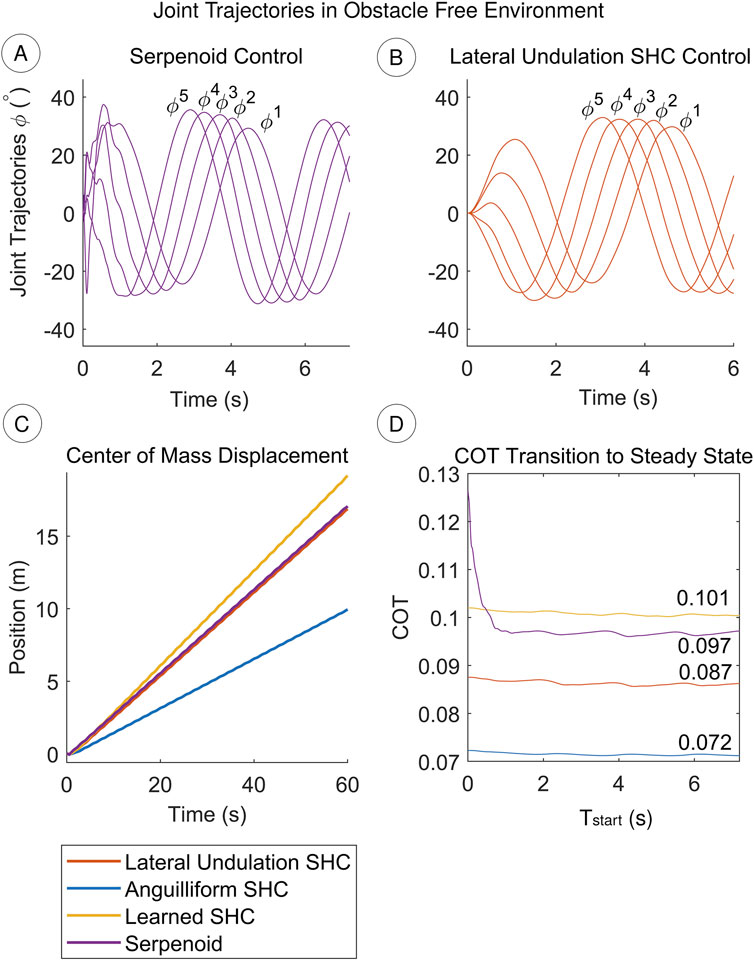
Figure 5. (A, B) The SHC controller produces comparable trajectories to the serpenoid controller (C) In obstacle-free conditions, the lateral undulation SHC performs similarly to the serpenoid controller. The anguilliform SHC controller swims slower because the joint oscillation amplitude is reduced, so thrust decreases. Learning SHC weights improves speed. Video is included in Supplementary Files (D) The COT reaches steady state performance after
Since the serpenoid and lateral undulation SHC controllers generate similar trajectories, they achieve comparable performance. The average velocity of the center of mass for both controllers is
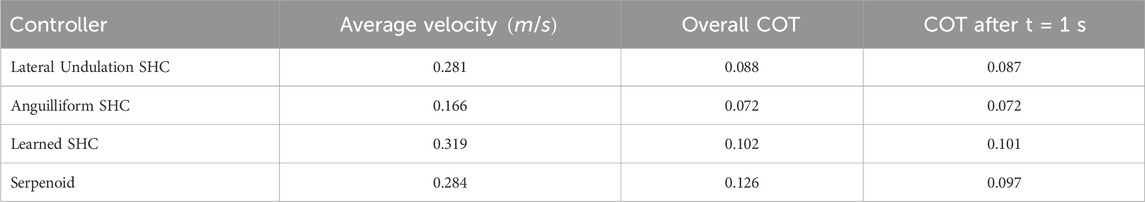
Table 4. Performance during obstacle-free swimming.
A benefit of SHC control over serpenoid control is apparent when assessing the overall COT including transient effects
3.1.2 Anguilliform gait with SHC control
By linearly scaling the SHC weights, we transform the lateral undulation gait to an anguilliform gait, with reduced anterior joint amplitudes. For example, we scale the weights for the anterior-most joint by
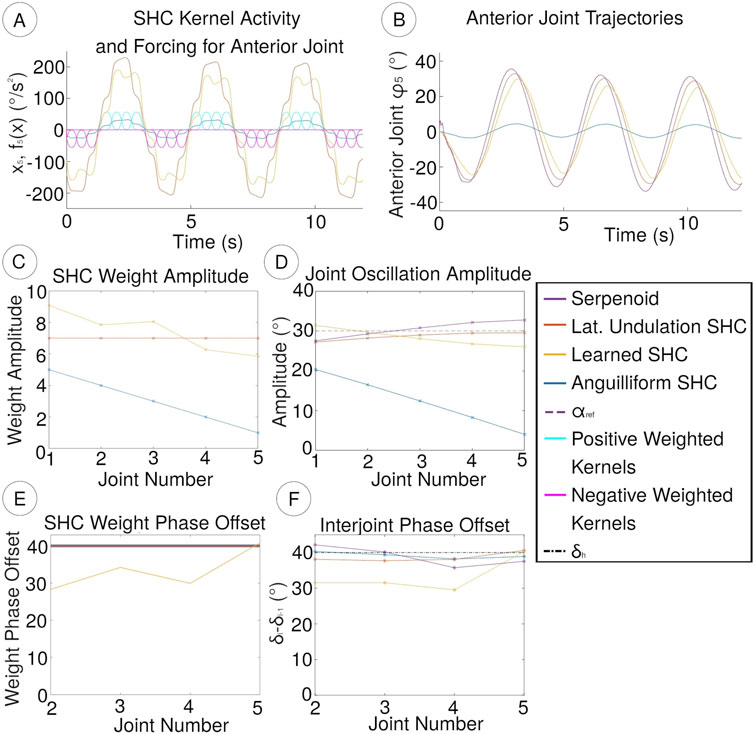
Figure 6. (A) Positively (cyan) and negatively (magenta) weighted SHC kernels
Scaling effects are more thoroughly characterized by considering the amplitudes of weights and
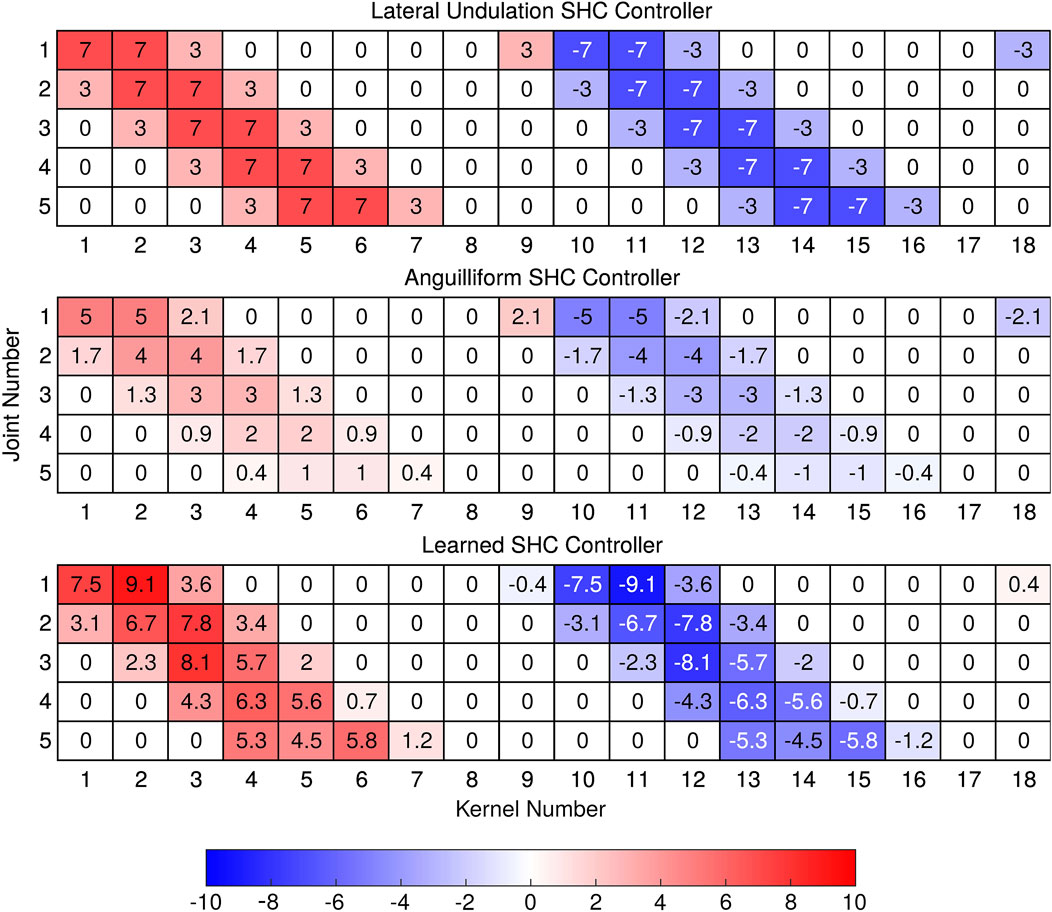
Figure 7. Weights for SHC controllers. Lateral undulation uses consistent weights across all joints, while the anguilliform weights decrease in magnitude from the tail (joint 1) to the head (joint 5). In the learned gait, peak weights also decrease towards the head, although the phase differences between joints also differ because
3.1.3 Learned gait with SHC control
Our gait optimization for SHC control modifies the weight parameters to enhance speed of locomotion, while minimally affecting energy efficiency. The learned SHC controller achieves a
The improved speed of locomotion in the learned SHC controller corresponds to changes in the controller weights and joint amplitudes. Relative to the lateral undulation SHC controller, the learned SHC controller possesses higher weight amplitudes for the posterior joints and lower weights for the anterior joints (7 (B)). The learned SHC weight amplitudes correspond loosely to the joint amplitudes (Figure 6D), which show a gradual decrease towards the head of the robot. Compared with anguilliform and lateral undulation SHC controllers, the learned controller shows weaker correspondence between weights and joint amplitudes because the distribution of weights, and thus acceleration patterns, may differ between joints.
When optimizing weight parameters for the learned SHC controller, the distribution of weights for each joint changes along with the peak amplitude. We compute the phases of weight distributions
Gait optimization produces weight phase offsets between joints
3.2 Straight channels
We assess our method for integrating sensory information in the SHC controller by examining the robot’s progress through straight, narrow passages. Video of the robot in a
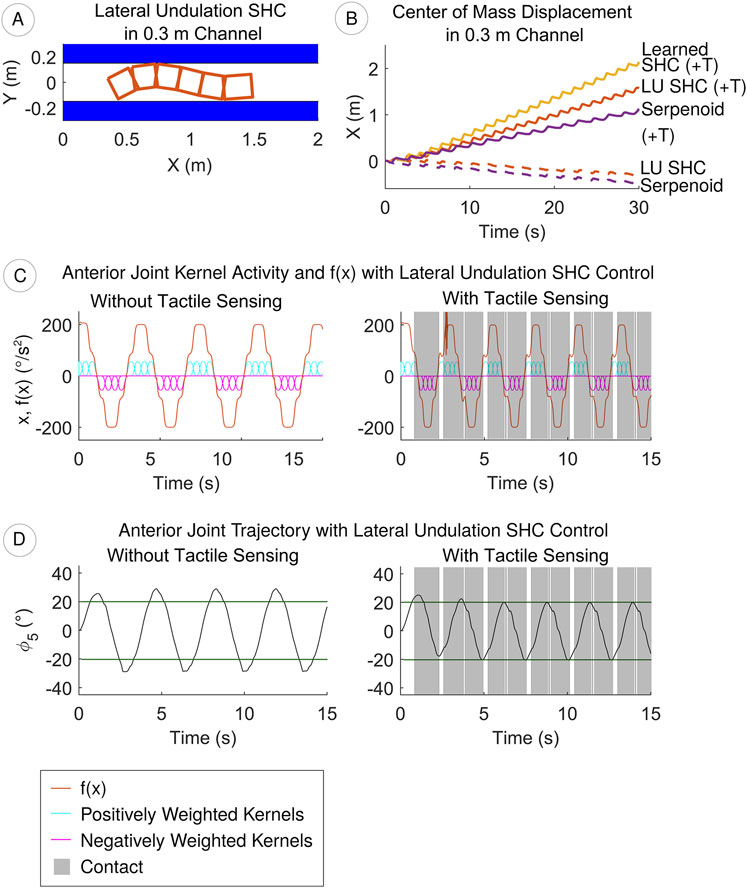
Figure 8. (A) Using lateral undulation SHC control with tactile sensing (+T), the robot moves through a 0.3 m channel (B) Tactile sensing improves the swimming speed through the channel, especially with learned SHC control (C) Contact detection increases the frequency of SHC kernels and
The relative performance of the controllers depends on the width of the crevice (Figure 9). In the narrowest channels all controllers yield backwards net locomotion, as the robot incidentally pushes backwards against the walls. As channel width increases, forward velocity generally increases before saturating when the channels are wide enough that wall contact becomes negligible. Controllers that incorporate tactile sensing achieve forward progress at lower channel widths and maintain higher average velocities than their unsensorized counterparts. Moreover, the SHC controllers with tactile sensing achieve higher average velocities than the serpenoid (+T) controller, suggesting that the frequency and amplitude modulation is more effective than amplitude reduction alone. Consistent with results for the unimpeded swimming study, the learned SHC (+T) controller yields the highest velocity of all controllers in most regimes.
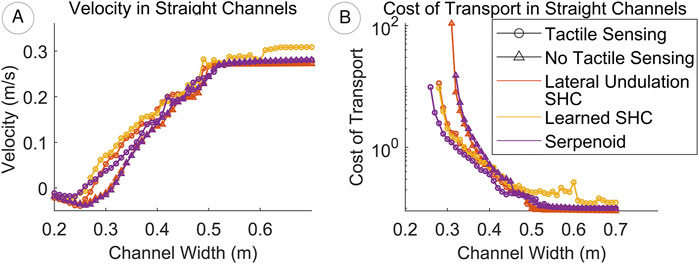
Figure 9. (A) Controllers yield backwards locomotion in the narrowest channels. Velocities increase until no wall contact occurs. The learned SHC controller (+T) permits faster swimming than both the lateral undulation SHC and serpenoid controllers (B) COT is defined only where the robot progresses forwards and decreases (i.e., locomotion becomes more efficient) as channel width increases. Tactile sensing reduces COT especially in narrow channels. While the learned gait generally improves speed, it sacrifices efficiency in some conditions.
The advantages of tactile sensing are similarly evident from COT trends for narrow crevices (Figure 9). For the same channels, COTs for controllers equipped with tactile sensors are consistently lower than for controllers without. The lateral undulation and learned SHC controllers (+T) achieve similar efficiency to the serpenoid controller (+T) in channels narrower than approximately
3.3 Uneven channels
In uneven channels, contact detection remains crucial to the performance of the robot. Without tactile sensing, the serpenoid and SHC controllers cannot adapt the body shape and become trapped when the robot contacts both sides of the channel (Figure 10A). With tactile sensing, the controllers reduce the amplitude of the joints, narrowing the gait and allowing the robot to pass obstacles (see video in Supplementary Files). However, the robot may still not progress consistently if it comes into contact with an obstacle head on, for example, the lateral undulation SHC (+T) controller in Figure 10A. In this case, the robot continually retries forward motion, colliding with the channel wall until a more suitable approach configuration occurs by chance and the robot overcomes the obstacle. As in prior experiments, the learned SHC allows the greatest forward progress when it is not trapped in the channel (Figure 10).
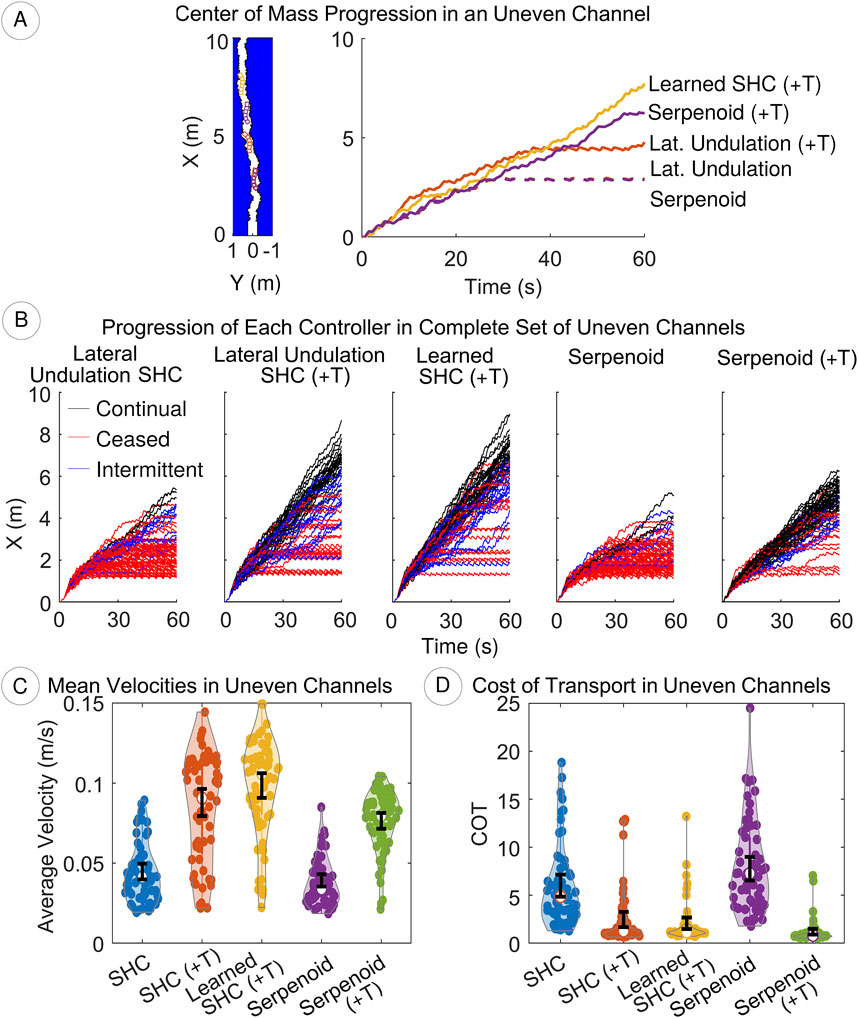
Figure 10. (A) Tactile sensing improves locomotion in uneven terrain, though many attempts may be needed if approach is sub-optimal (e.g., lateral undulation SHC (+T); see Supplementary Video) (B) We categorize progress for each trial as continual (black), ceased (red), or intermittent (blue). Lateral undulation SHC (+T) and learned SHC (+T) controllers progress faster, but are trapped more often than the serpenoid controller (+T) (C, D) Velocity and COT distributions (dots) are shown.
In the complete set of channels, the serpenoid and SHC controllers with tactile sensing achieve greater progress than those without. Of 65 obstacles, our analyses include 59 (serpenoid), 65 (serpenoid (+T)), 62 (lateral undulation SHC), 64 (lateral undulation SHC (+T)), and 62 (learned SHC (+T)) data points. In
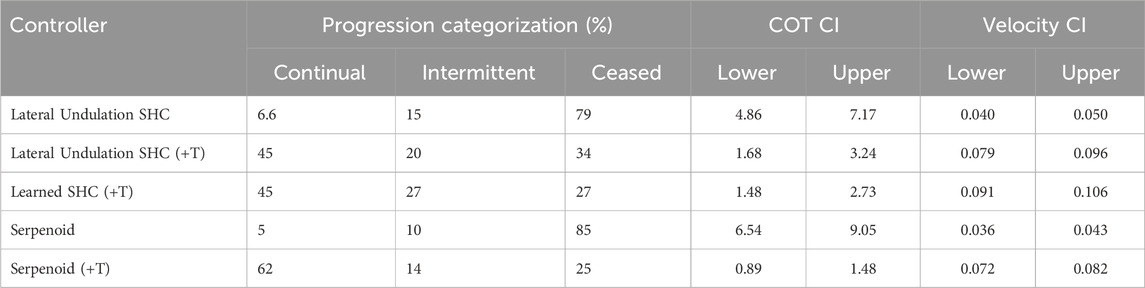
Table 5. Uneven channel performance.
Mean velocities are compared between controllers with a Kruskal–Wallis test, which indicates that one or more controllers differs
Incorporating tactile sensing improves efficiency, as well as speed. A Kruskal–Wallis test shows significant differences between the controllers
4 Discussion
In this work, we implement movement primitives based on stable heteroclinic channels to control a simulated aquatic snake robot. The SHC controller provides comparable performance to a conventional serpenoid controller. We demonstrate that MP weights relate proportionally to joint trajectories by linearly scaling the weights from the lateral undulation SHC controller to achieve an anguilliform gait. We optimize the SHC controller by modeling the system dynamics and iteratively tuning the weights, resulting in a
Our research serves as a practical demonstration of controlling a nonlinear dynamical system with SHC movement primitives and adapting its behavior by incorporationg sensory information. By showing that the weight parameters in SHC movement primitive relate proportionally to the resulting joint trajectories, we provide a pathway for robot operators to intuitively visualize, alter, and design gaits. In optimized controllers, the same visualizations may permit operators to heuristically predict emergent behavior without extensive testing. Thus, the SHC movement primitive controller offers a transparent approach to modeling CPGs, enabling faster development of bio-inspired systems. The abstract, visualizable approach allows researchers to address how CPG modules, represented by saddle equilibria, interact with sensory inputs and other modules to produce complex rhythmic motor patterns.
The present work demonstrates that SHCs are a promising control framework for replicating rhythmic behavior, and suggests that other locomotion patterns may be constructed by tuning weights and modulating progression between kernels. Future research may examine more advanced methods for incorporating sensory feedback to adapt individual joint trajectories, rather than regulating the entire network. Investigating weights and arrangements of saddle equilibria that enable other gaits, such as concertina locomotion would also be worthwhile. Transitioning between gaits may be achievable by expanding upon research by Rouse and Daltorio (2024), which explores the effects of sensor noise on decision making. Additionally, integration of more complex sensory information, such as vision, for localization and long term path planning would be beneficial. The effects of multisensory joint adaptation and gait transitions could be analyzed in environments with abrupt directional changes or multiple paths, such as S or T-shaped channels. With these improvements, movement primitives based on SHCs may contribute to research in biology by providing a convenient method for abstractly modeling neurological functions that support locomotion without detailed simulation of underlying cellular dynamics.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
NM: Conceptualization, Formal Analysis, Funding acquisition, Investigation, Methodology, Visualization, Writing–original draft, Writing–review and editing. NR: Writing–review and editing. KD: Funding acquisition, Project administration, Resources, Supervision, Writing–review and editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This research would not be possible without funding from Department of Defense (DoD) through the National Defense Science & Engineering Graduate (NDSEG) Fellowship Program. Research was also supported by the National Science Foundation’s CAREER award (2047330).
Acknowledgments
Additionally, we thank Dr. Hillel Chiel (Department of Biology, Case Western Reserve University) for reviewing and editing this work, as well as Dr. Zonghe Chua (Department of Electrical, Computer, and Systems Engineering, Case Western Reserve University) for advice on statistical analysis.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/felec.2025.1507644/full#supplementary-material
SUPPLEMENTARY VIDEO S1 | A comparison of swimming behavior for SHC movement primitive and serpenoid controllers in an unobstructed environment.
SUPPLEMENTARY VIDEO S2 | In a 0.3 m channel, tactile sensing improves performance for both SHC movement primitive and serpenoid controllers. SHC controllers undulate both at higher frequency and lower amplitude during contact, while the serpenoid controller only reduces amplitude.
SUPPLEMENTARY VIDEO S3 | Tactile sensor input improves the robot’s ability to navigate uneven terrain. However, the robot can still be stymied by obstacles. With sensing, the SHC controller tuned for lateral undulation becomes stuck (t∼35 s), but ultimately recovers.
SUPPLEMENTARY FIGURE S1 | When learning weight parameters via gradient descent for the SHC control system, the velocity and COT stabilize within about 150 iterations.
SUPPLEMENTARY FIGURE S2 | Confidence intervals on cost of transport for experiments in uneven channels are constructed by bootstrapping. 10,000 iterations is sufficient to ensure convergence of both upper and lower bounds for any of the tested controllers.
SUPPLEMENTARY FIGURE S3 | Bootstrapped confidence intervals for velocity in uneven channels converge within 10,000 iterations for all examined controllers.
References
Anastasiadis, A., Paez, L., Melo, K., Tytell, E. D., Ijspeert, A. J., and Mulleners, K. (2023). Identification of the trade-off between speed and efficiency in undulatory swimming using a bio-inspired robot. Sci. Rep. 13, 15032. doi:10.1038/s41598-023-41074-9
Andersson, O., Grillner, S., Lindquist, M., and Zomlefer, M. (1978). Peripheral control of the spinal pattern generators for locomotion in cat. Brain Res. 150, 625–630. doi:10.1016/0006-8993(78)90827-2
Ashwin, P., Karabacak, Ã., and Nowotny, T. (2011). Criteria for robustness of heteroclinic cycles in neural microcircuits. J. Math. Neurosci. 1, 13. doi:10.1186/2190-8567-1-13
Ashwin, P., and Postlethwaite, C. (2016). Quantifying noisy attractors: from heteroclinic to excitable networks. SIAM J. Appl. Dyn. Syst. 15, 1989–2016. doi:10.1137/16M1061813
Astley, H. C., and Jayne, B. C. (2007). Effects of perch diameter and incline on the kinematics, performance and modes of arboreal locomotion of corn snakes (Elaphe guttata). J. Exp. Biol. 210, 3862–3872. doi:10.1242/jeb.009050
Bonardi, S., Moeckel, R., Sproewitz, A., Vespignani, M., and Ijspeert, A. J. (2012). “Locomotion through reconfiguration based on motor primitives for roombots self-reconfigurable modular robots,” in ROBOTIK 2012; 7th German conference on robotics, 1–6.
Brecelj, T., and Petrič, T. (2023a). Stable heteroclinic channel networks for physical human-humanoid robot collaboration. Sensors 23, 1396. doi:10.3390/s23031396
Brecelj, T., and Petrič, T. (2023b). “Utilizing a phase state system for reliable physical assistance in human-humanoid robot collaboration,” in 2023 21st international conference on advanced robotics (ICAR), 258–263. doi:10.1109/ICAR58858.2023.10406553
Chen, S., and Roth, A. (2023). Gait design of a novel arboreal concertina locomotion for snake-like robots. ArXiv:2309.06000. doi:10.48550/arXiv.2309.06000
Cheslet, J., Beaubois, R., Khoyratee, F., Kohno, T., Ikeuchi, Y., and Levi, T. (2024). “Biomimetic snake locomotion using Central Pattern Generators network and bio-hybrid robot perspective,” in Isarob (Beppu, Japan).
Cropper, E. C., Evans, C. G., Hurwitz, I., Jing, J., Proekt, A., Romero, A., et al. (2004). Feeding neural networks in the mollusc Aplysia. Neurosignals 13, 70–86. doi:10.1159/000076159
Daltorio, K. A., Boxerbaum, A. S., Horchler, A. D., Shaw, K. M., Chiel, H. J., and Quinn, R. D. (2013). Efficient worm-like locomotion: slip and control of soft-bodied peristaltic robots. Bioinspiration and Biomimetics 8, 035003. doi:10.1088/1748-3182/8/3/035003
[Dataset] Rouse, N., Horchler, A., Chiel, H., and Daltorio, K. (2024). Sttable heteroclinic channels as a decision-making model: overcoming low signal-to-noise ratio with mutual inhibition.
Dehghani, M., and Mahjoob, M. J. (2009). “A modified serpenoid equation for snake robots,” in 2008 IEEE international conference on robotics and biomimetics, 1647–1652. doi:10.1109/ROBIO.2009.4913248
Fod, A., Matarić, M. J., and Jenkins, O. C. (2002). Automated derivation of primitives for movement classification. Aut. Robots 12, 39–54. doi:10.1023/A:1013254724861
Giszter, S. (2015). Motor primitives—new data and future questions. Curr. Opin. Neurobiol. 33, 156–165. doi:10.1016/j.conb.2015.04.004
Gray, J. (1946). The mechanism of locomotion in snakes. J. Exp. Biol. 23, 101–120. doi:10.1242/jeb.23.2.101
Grillner, S., and Wallén, P. (2010). “The lamprey locomotor central pattern generator,” in Handbook of brain microcircuits. Editors D. Shepherd, S. Grillner, and P. Gordon (Oxford University Press). doi:10.1093/med/9780195389883.003.0032
Guertin, P. A. (2013). Central pattern generator for locomotion: anatomical, physiological, and pathophysiological considerations. Front. Neurology 3, 183. doi:10.3389/fneur.2012.00183
Harris-Warrick, R. M. (2010). General principles of rhythmogenesis in central pattern generator networks. Prog. Brain Res. 187, 213–222. doi:10.1016/B978-0-444-53613-6.00014-9
Hoffmann, H., Pastor, P., Park, D.-H., and Schaal, S. (2009). “Biologically-inspired dynamical systems for movement generation: automatic real-time goal adaptation and obstacle avoidance,” in 2009 IEEE international conference on robotics and automation, 2587–2592. doi:10.1109/ROBOT.2009.5152423
Horchler, A. D., Daltorio, K. A., Chiel, H. J., and Quinn, R. D. (2015). Designing responsive pattern generators: stable heteroclinic channel cycles for modeling and control. Bioinspiration and Biomimetics 10, 026001. doi:10.1088/1748-3190/10/2/026001
Huang, Z., Kong, D., Ren, C., Li, S., and Ma, S. (2019). “Performance study of an underwater snake-like robot with a flexible caudal fin,” in 2019 IEEE international conference on mechatronics and automation (ICMA), 1–5. doi:10.1109/ICMA.2019.8816412
Hunt, A., Szczecinski, N., and Quinn, R. (2017). Development and training of a neural controller for hind leg walking in a dog robot. Front. Neurorobotics 11, 18. doi:10.3389/fnbot.2017.00018
Ijspeert, A. J. (2008). Central pattern generators for locomotion control in animals and robots: a review. Neural Netw. 21, 642–653. doi:10.1016/j.neunet.2008.03.014
Ijspeert, A. J., Crespi, A., Ryczko, D., and Cabelguen, J.-M. (2007). From swimming to walking with a salamander robot driven by a spinal cord model. Science 315, 1416–1420. doi:10.1126/science.1138353
Ijspeert, A. J., Nakanishi, J., Hoffmann, H., Pastor, P., and Schaal, S. (2013). Dynamical movement primitives: learning attractor models for motor behaviors. Neural Comput. 25, 328–373. doi:10.1162/NECO_a_00393
Jayne, B. C. (2020). What defines different modes of snake locomotion? Integr. Comp. Biol. 60, 156–170. doi:10.1093/icb/icaa017
Kelasidi, E., Jesmani, M., Pettersen, K. Y., and Gravdahl, J. T. (2018). Locomotion efficiency optimization of biologically inspired snake robots. Appl. Sci. 8, 80. doi:10.3390/app8010080
Kelasidi, E., Pettersen, K. Y., Gravdahl, J. T., and Liljebäck, P. (2014). “Modeling of underwater snake robots,” in 2014 IEEE international conference on robotics and automation (ICRA), 4540–4547. doi:10.1109/ICRA.2014.6907522
Kelasidi, E., Pettersen, K. Y., Gravdahl, J. T., Strømsøyen, S., and Sørensen, A. (2017). “Modeling and propulsion methods of underwater snake robots,” in 2017 IEEE conference on control technology and applications (CCTA), 819–826. doi:10.1109/CCTA.2017.8062561
Kober, J., and Peters, J. (2009). “Learning motor primitives for robotics,” in 2009 IEEE international conference on robotics and automation, 2112–2118. doi:10.1109/ROBOT.2009.5152577
Kong, L.-H., He, W., Chen, W.-S., Zhang, H., and Wang, Y.-N. (2023). Dynamic movement primitives based robot skills learning. Mach. Intell. Res. 20, 396–407. doi:10.1007/s11633-022-1346-z
Lissmann, H. W. (1950). Rectilinear locomotion in a snake (Boa occidentalis). J. Exp. Biol. 26, 368–379. doi:10.1242/jeb.26.4.368
Marder, E., and Eisen, J. S. (1984). Electrically coupled pacemaker neurons respond differently to same physiological inputs and neurotransmitters. J. Neurophysiology 51, 1362–1374. doi:10.1152/jn.1984.51.6.1362
Marques, F., Flores, P., Claro, J., and Lankarani, H. (2016). A survey and comparison of several friction force models for dynamic analysis of multibody mechanical systems. Nonlinear Dyn. 86, 1407–1443. doi:10.1007/s11071-016-2999-3
McCormack, A., and Godfrey, K. (1998). Rule-based autotuning based on frequency domain identification. IEEE Trans. Control Syst. Technol. 6, 43–61. doi:10.1109/87.654876
Moreno, R., and Gomez, J. (2011). “Central pattern generators and hormone inspired messages: a hybrid control strategy to implement motor primitives on chain type modular reconfigurable robots,” in 2011 IEEE international conference on robotics and automation, 1014–1019. doi:10.1109/ICRA.2011.5980149
Mosauer, W. (1932). On the locomotion of snakes. Science 76, 583–585. doi:10.1126/science.76.1982.583
Norman-Tenazas, R. (2021). Robust snake robot control via A spiking neuron central pattern generator. Baltimore, MD: Johns Hopkins University. Master’s thesis.
Ostrowski, J., and Burdick, J. (1996). Gait kinematics for a serpentine robot. In , Proceedings of IEEE international conference on robotics and automation. 1294–1299. doi:10.1109/ROBOT.1996.506885
Paraschos, A., Daniel, C., Peters, J. R., and Neumann, G. (2013). “Probabilistic movement primitives,” in Advances in neural information processing systems (San Diego, CA: Neural Information Processing Systems, Inc.), 26.
Pastor, P., Hoffmann, H., Asfour, T., and Schaal, S. (2009). “Learning and generalization of motor skills by learning from demonstration,” in 2009 IEEE international conference on robotics and automation, 763–768. doi:10.1109/ROBOT.2009.5152385
Piñeirua, M., Godoy-Diana, R., and Thiria, B. (2015). Resistive thrust production can be as crucial as added mass mechanisms for inertial undulatory swimmers. Phys. Rev. E 92, 021001. doi:10.1103/PhysRevE.92.021001
Rabinovich, M. I., Huerta, R., Varona, P., and Afraimovich, V. S. (2006). Generation and reshaping of sequences in neural systems. Biol. Cybern. 95, 519–536. doi:10.1007/s00422-006-0121-5
Riddle, S., Jackson, C., Daltorio, K. A., and Quinn, R. D. (2023). “A dynamic simulation of a compliant worm robot amenable to neural control,” in Biomimetic and biohybrid systems. Editors F. Meder, A. Hunt, L. Margheri, A. Mura, and B. Mazzolai (Cham: Springer Nature Switzerland), 338–352. doi:10.1007/978-3-031-38857-6_25
Rollinson, D., and Choset, H. (2013). “Gait-based compliant control for snake robots,” in 2013 IEEE international conference on robotics and automation, 5138–5143. doi:10.1109/ICRA.2013.6631311
Rouse, N., and Daltorio, K. (2024). Stable heteroclinic channel-based movement primitives: tuning trajectories using saddle parameters. Appl. Sci. 14, 2523. doi:10.3390/app14062523
Rouse, N. A., and Daltorio, K. A. (2021). Visualization of stable heteroclinic channel-based movement primitives. IEEE Robotics Automation Lett. 6, 2343–2348. doi:10.1109/LRA.2021.3061382
Sato, M., Fukaya, M., and Iwasaki, T. (2002). Serpentine locomotion with robotic snakes. IEEE Control Syst. Mag. 22, 64–81. doi:10.1109/37.980248
Schaal, S. (2006). “Dynamic movement primitives -A framework for motor control in humans and humanoid robotics,” in Adaptive motion of animals and machines. Editors H. Kimura, K. Tsuchiya, A. Ishiguro, and H. Witte (Tokyo: Springer), 261–280. doi:10.1007/4-431-31381-8_23
Shaw, K. M., Lyttle, D. N., Gill, J. P., Cullins, M. J., McManus, J. M., Lu, H., et al. (2015). The significance of dynamical architecture for adaptive responses to mechanical loads during rhythmic behavior. J. Comput. Neurosci. 38, 25–51. doi:10.1007/s10827-014-0519-3
Sherrington, C. S. (1910). Flexion-reflex of the limb, crossed extension-reflex, and reflex stepping and standing. J. Physiology 40, 28–121. doi:10.1113/jphysiol.1910.sp001362
Shigeo, H. (1994). Biologically inspired robots: snake-like locomotors and manipulators by Shigeo hirose oxford university press, oxford, 1993, 220pages, incl. Index (£40). Robotica 12, 282. doi:10.1017/S0263574700017264
Simoni, M. F., and DeWeerth, S. P. (2007). Sensory feedback in a half-center oscillator model. IEEE Trans. Biomed. Eng. 54, 193–204. doi:10.1109/TBME.2006.886868
Tagliabue, M., Ciancio, A. L., Brochier, T., Eskiizmirliler, S., and Maier, M. A. (2015). Differences between kinematic synergies and muscle synergies during two-digit grasping. Front. Hum. Neurosci. 9, 165. doi:10.3389/fnhum.2015.00165
Tesch, M., Lipkin, K., Brown, I., Hatton, R., Peck, A., Rembisz, J., et al. (2009). Parameterized and scripted gaits for modular snake robots. Adv. Robot. 23, 1131–1158. doi:10.1163/156855309X452566
Thandiackal, R., Melo, K., Paez, L., Herault, J., Kano, T., Akiyama, K., et al. (2021). Emergence of robust self-organized undulatory swimming based on local hydrodynamic force sensing. Sci. Robotics 6, eabf6354. doi:10.1126/scirobotics.abf6354
Travers, M., Gong, C., and Choset, H. (2015). “Shape-constrained whole-body adaptivity,” in 2015 IEEE international symposium on safety, security, and rescue robotics (SSRR), 1–6. doi:10.1109/SSRR.2015.7442945
Travers, M., Whitman, J., and Choset, H. (2018). Shape-based coordination in locomotion control. Int. J. Robotics Res. 37, 1253–1268. doi:10.1177/0278364918761569
Tresch, M. C., and Bizzi, E. (1999). Responses to spinal microstimulation in the chronically spinalized rat and their relationship to spinal systems activated by low threshold cutaneous stimulation. Exp. Brain Res. 129, 0401–0416. doi:10.1007/s002210050908
Wang, J., Ouyang, W., Gao, W., and Ren, Q. (2017a). “Locomotion control of a serpentine crawling robot inspired by central pattern generators,” in 2017 asia-pacific signal and information processing association annual summit and conference (APSIPA ASC), 414–419. doi:10.1109/APSIPA.2017.8282067
Wang, Z., Gao, Q., and Zhao, H. (2017b). CPG-inspired locomotion control for a snake robot basing on nonlinear oscillators. J. Intelligent and Robotic Syst. 85, 209–227. doi:10.1007/s10846-016-0373-9
Keywords: stable heteroclinic channels, snake robot, autonomy, central pattern generator, motor primitive, nonlinear control
Citation: Mengers N, Rouse N and Daltorio KA (2025) Stable heteroclinic channels for controlling a simulated aquatic serpentine robot in narrow crevices. Front. Electron. 6:1507644. doi: 10.3389/felec.2025.1507644
Received: 08 October 2024; Accepted: 31 January 2025;
Published: 24 February 2025.
Edited by:
Afaque Manzoor, Washington University in St. Louis, United StatesReviewed by:
Giovanni Bianchi, Polytechnic University of Milan, ItalyQiyuan Fu, Biorobotics Laboratory (BioRob), Switzerland
Jahan Zeb Gul, Air University, Pakistan
Copyright © 2025 Mengers, Rouse and Daltorio. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nathaniel Mengers, bm5tMjJAY2FzZS5lZHU=