 Siwajali Selemani
Siwajali Selemani Richard Madege
Richard Madege Yasinta Nzogela
Yasinta Nzogela- Department of Crop Science and Horticulture, Sokoine University of Agriculture, Morogoro, Tanzania
Understanding the impact of seed-borne fungi on seed quality is crucial for improving sunflower productivity. This study was conducted to investigate the incidence of seed-borne fungi in stored sunflower seeds under ambient conditions and their effects on seed viability and vigor. Certified seeds, quality-declared seeds (QDS), and farmer-saved seeds (FSS) were stored for 6 months, with monthly sampling to evaluate fungal infection percentage (FIP), germination percentage (G), and seedling vigor index (SVI). Key findings include the identification of fungal species: Aspergillus flavus, A. niger, Alternaria padwickii, A. zinniae, Botrytis cinerea, Curvularia lunata, Fusarium vericillioides, and Rhizopus arrihizus. The highest incidence of F. vericillioides (14.39%) and B. cinerea (14.28%) occurred in FSS, while the lowest incidence of A. zinniae (1.83%) was recorded in QDS. Total fungal infection was highest in FSS (73.11%) compared to QDS (66.94%) and certified seeds (67.06%). Seed quality parameters showed a clear trend: QDS exhibited the highest G (81.56%), with certified seeds following closely (81.33%). The highest SVI was recorded in certified seeds (96.76), with QDS showing a similar result (95.69). In contrast, FSS showed the lowest G% (60.67) and SVI (65.98). Seeds with minimal fungal infection consistently demonstrated higher G% and SVI. These results highlight the detrimental impact of fungal infections on seed vigor and viability. This study is the first in Tanzania to report and quantify the incidence of seed-borne fungi in stored sunflower seeds, providing critical insights for improving seed management practices and sunflower production.
1 Introduction
Sunflower (Helianthus annuus L.) is among the major oil seed crops grown globally due to its short growing season (FAO, 2017). Sunflower is grown widely for edible oil (Mgeni and Mpenda, 2021; Reuben and Meliyo, 2022), and the seeds are eaten by humans. The sunflower seed cake is used as animal feed (Konyali, 2017). The crop is produced on over 27 million hectares worldwide (Muradov et al., 2019). Tanzania is among the top 10 sunflower producers in the world, contributing approximately 2.4% of the world’s production and 34% of African production (Guney et al., 2017; Isinika and John, 2021). In Tanzania, sunflower crops are grown mainly by small-scale farmers (Bank of Tanzania, 2017). The central zone regions, which include Singida and Dodoma, are the major sunflower producers, contributing approximately 61% of the country’s total production (Tibamanya et al., 2022). Sunflower production improves smallholder farmers’ income, which in turn contributes to the country’s income generation (Sebyiga, 2020).
Despite being one of the top 10 major sunflower producers in the world, Tanzania can meet only approximately 30 to 45% of the internal demand for edible oil while importing approximately 55 to 70% (Balchin et al., 2018). Among the factors contributing to the low production of sunflowers is the unavailability of quality seeds to smallholder farmers linked to poor seed health due to high infection rates of seed-borne diseases caused by fungal pathogens (Lyanga, 2024; Tibamanya et al., 2022).
According to the Food and Agriculture Organization of the United Nations, seed health is among the important factors for successful cultivation and yield exploration of a crop (FAO, 2017). Studies show that seed-borne fungal infections cause approximately 9.7% of annual yield loss worldwide (Kumar et al., 2020). Seed-borne fungal infection reduces germination seedling vigor and causes variation in plant morphology after seed deterioration through seed abortion, shrinking, reduced size, and seed rot, as reported by Niaz and Dawar (2009) and Sserumaga et al. (2015).
Furthermore, the infection rate of stored seeds depends on environmental conditions such as high relative humidity, suitable temperature, and high moisture content level in the seed. The study of seed-borne pathogens is necessary to determine seed health and improve seed germination potential, which finally leads to an increase in crop production (Masomeh et al., 2012). Since smallholder farmers store their produced seeds for the next planting season, the seeds become infected by seed-borne fungi, including pathogenic fungi, during storage, which could reduce seed germination and vigor, as described by Gebeyaw (2020).
Therefore, seed health testing to detect seed-borne pathogens is an important step in the management of crop diseases. According to Abd-El-Aziz and El-Satar (2016), 10 fungi were identified from four sunflower seed genotypes. These were Alternaria alternata, Aspergillus flavus, A. niger, Aspergillus spp., Cladosporium herbarum, Fusarium equiseti, Paecilomyces variotii, Penicillium spp. and Rhizopus arrihizus. However, little is known about the status of fungal pathogens in stored sunflower seeds in Tanzania. The information on seed-borne fungal pathogens is necessary for local seed growers and smallholder farmers to understand the seed-borne fungal pathogens infecting stored sunflower seeds. Therefore, this study aimed to identify seed-borne mycoflora of stored sunflower seeds under ambient conditions in the country and their influence on seed viability and vigor.
2 Materials and methods
2.1 Collection of seed material
The sunflower seed samples, including quality-declared seeds (QDS), certified OPV, and farmer-saved seeds (FSS), were collected from QDS producers, the Agricultural Seed Agency (ASA), and farmers during the 2022–2023 growing season. Seeds were collected during the harvest season after the crops had reached full maturity (International Seed Testing Association (ISTA), 2022). After collection, the seeds were stored in well-ventilated areas by spreading them in thin layers on clean drying trays and turning them regularly to ensure even drying (International Seed Testing Association (ISTA), 2022). The seeds were allowed to air dry naturally until their moisture content reached 9%, which is optimal for storage and minimizes the risk of fungal infections and deterioration (FAO, 2018). The moisture content of the seeds was measured using the high-temperature oven method at 105°C for 24 h.
2.2 Seed preservation experiment
The storage experiment was conducted at the African Seed Health Lab within the Department of Crop Science and Horticulture at Sokoine University of Agriculture. A triplicated split-split-plot experiment in a randomized complete block design was established, consisting of three types of packaging materials (plastic containers, polypropylene bags, and sisal bags) as the main factor, three classes of sunflower seeds (QDS, certified seeds, and FSS) as the sub-plot factor, and storage period as the sub-sub plot factor, with six levels spanning from month one to month 6 (Genes and Nyomora, 2018). For each seed class, 1.5 kg was packaged in each packaging material. The plastic containers were securely closed with their plastic coverings, and the sisal and polypropylene bags were expertly sewn together using strings.
Subsequently, the seeds were placed in the laboratory for ambient preservation, each in its own packaging material for 6 months, from December 2022 to June 2023. Seed samples were collected monthly to evaluate the seed health, viability, and vigor of stored seeds. In accordance with International Seed Testing Association (ISTA) (2022) requirements, the sunflower seeds were tested for fungal infection, viability, and seedling vigor prior to storage to determine the seeds’ initial quality.
2.3 Data collection
2.3.1 Fungal assays
Two hundred seeds from each treatment were placed in Petri dishes monthly, with 10 seeds per dish, utilizing blotter papers adopted from Patil et al. (2018) and Nahar et al. (2005), owing to their effective approach in detecting a variety of seed-borne mycoflora in crops, including sunflower.
Prior to plating, the sunflower seeds were submerged in a 2% NaOCl₂ solution for 5 min, followed by 3 min of sterile water rinsing. The cultures were then maintained at room temperature for 7 days, with light and dark cycles. On the eighth day, the seeds were examined under a stereomicroscope to monitor the growth of the fungus. A compound microscope was then used to examine the fungal conidia and conidiomata on slides, and different magnifications were used to identify the spores and mycelia produced by each group of fungi. Fungal species were identified by Mathur and Kongsdal (2003). Isolation frequencies of fungal species were determined according to Kator et al., 2016 as follows Equations 1, 2:
2.3.2 Germination test
A total of 200 seeds from each respective sample were sown in four replications. The test was assessed by placing the seedlings in germination bowls filled with sterile sand soil. The soil was watered to keep it at the necessary moisture content. Germination was monitored every day for 10 days, and the Gs of the seedlings were determined as per he International Seed Testing Association (ISTA) (2022) formula (Equation 3):
2.4 Seedling vigor index
The SVI was determined according to Abdul-Baki and Anderson (1973). Briefly, after 10 days of germination tests, all seedlings with complete morphological parts were selected and regarded as vigorous seedlings and seedling dry weight was determined for 20 vigorous seedlings from which the SVI was determined using the formula (Equation 4):
2.5 Statistical data analysis
Analysis of Variance (ANOVA) was used to establish differences between treatments. The Tukey Honest Significance Difference test (HSD) was used for means separation at a significance threshold of 0.05. ‘The GenStat Discovery Statistical Package, 16th version’ was used for statistical analysis.
3 Results
3.1 Fungal infection and seed-borne fungal species
3.1.1 Impact of packaging, seed class, and storage duration on fungal infection, germination, and vigor of sunflower seeds
The study revealed that packaging material, seed class, and storage duration had significantly different (p < 0.001) effects on the prevalence of fungal infection in stored sunflower seeds. The results in Table 1 indicate that seeds stored in sisal bags had the highest fungal infection rate (7.56%), followed by polypropylene bags (6.91%), while plastic containers had the lowest rate (6.63%).
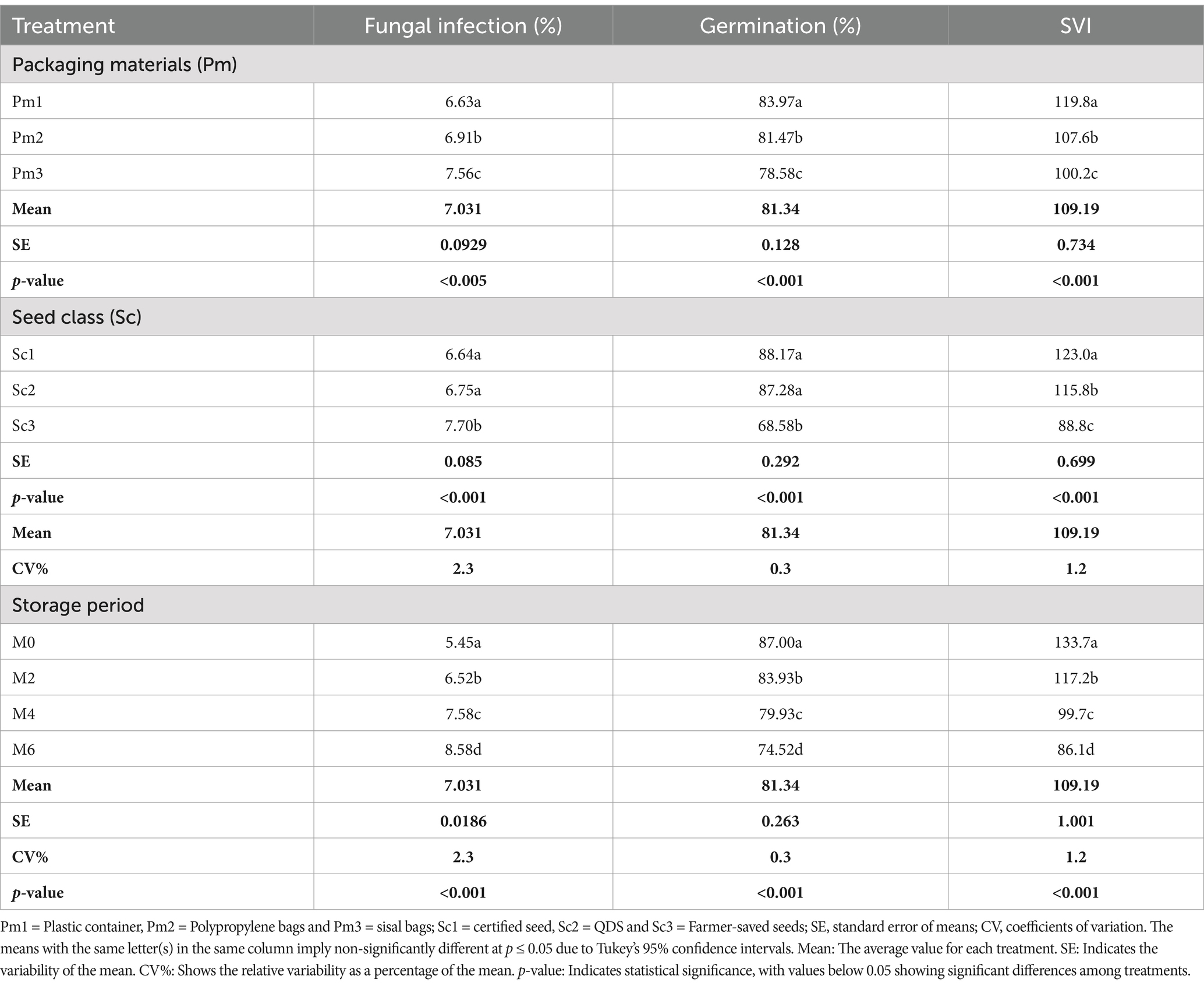
Table 1. The effects of packaging material, seed class, and storage period on fungal infection, seed germination, and SVI class.
Moreover, the results demonstrated that the effect of seed class on fungal infection was also significantly different (p < 0.001). FSS had the highest infection rate (7.70%), followed by QDS (6.75%), while certified seeds had the lowest rate (6.64%; Table 1). Moreover, the storage period also significantly influenced fungal infection (p < 0.00). The infection increased significantly with the duration of storage, starting from 5.45% at the control (Month 0) to 6.52, 7.58, and 8.58% at Month 2, Month 4, and Month 6 of storage, respectively (Table 1).
3.1.2 Effect of interaction between packaging material and seed class on fungal infection, seed germination, and SVI
In contrast to the previous section, which focused on the independent effects of packaging material and seed class, this analysis explored their combined influence on fungal seed-borne. The findings in Table 2 show that the interaction between packaging materials and seed class had no significant (p = 0.386) effects on fungal infection and seed germination but was significantly influenced by the SVI. The study elucidates that the interaction of plastic containers and certified seeds stored in plastic containers had the highest SVI (138.1). At the same time, the combination of sisal bags and the FSS had the lowest SVI (81.3), the same as that recorded in a combination of polypropylene bags and FSS (87.7).

Table 2. Fungal infection percentage (FIP), seed germination percentage (G), and SVI as influenced by the interaction between packaging material and seed class.
3.1.3 Occurrence of sunflower seed-borne fungal species
The study identified eight fungal species from the tested sunflower seed classes. These species were Aspergillus flavus, Aspergillus niger, Alternaria padwickii, Alternaria zinniae, Botrytis cinerea, Curvularia lunata, Fusarium vericillioides, and Rhizopus arrihizus. Among these fungi, Fusarium vericillioides had the highest incidence of 13.1%, followed by Botrytis cinerea at 12.5%, and the lowest incidence was observed with Alternaria zinniae at 1.2%. This information is presented in Figure 1A.
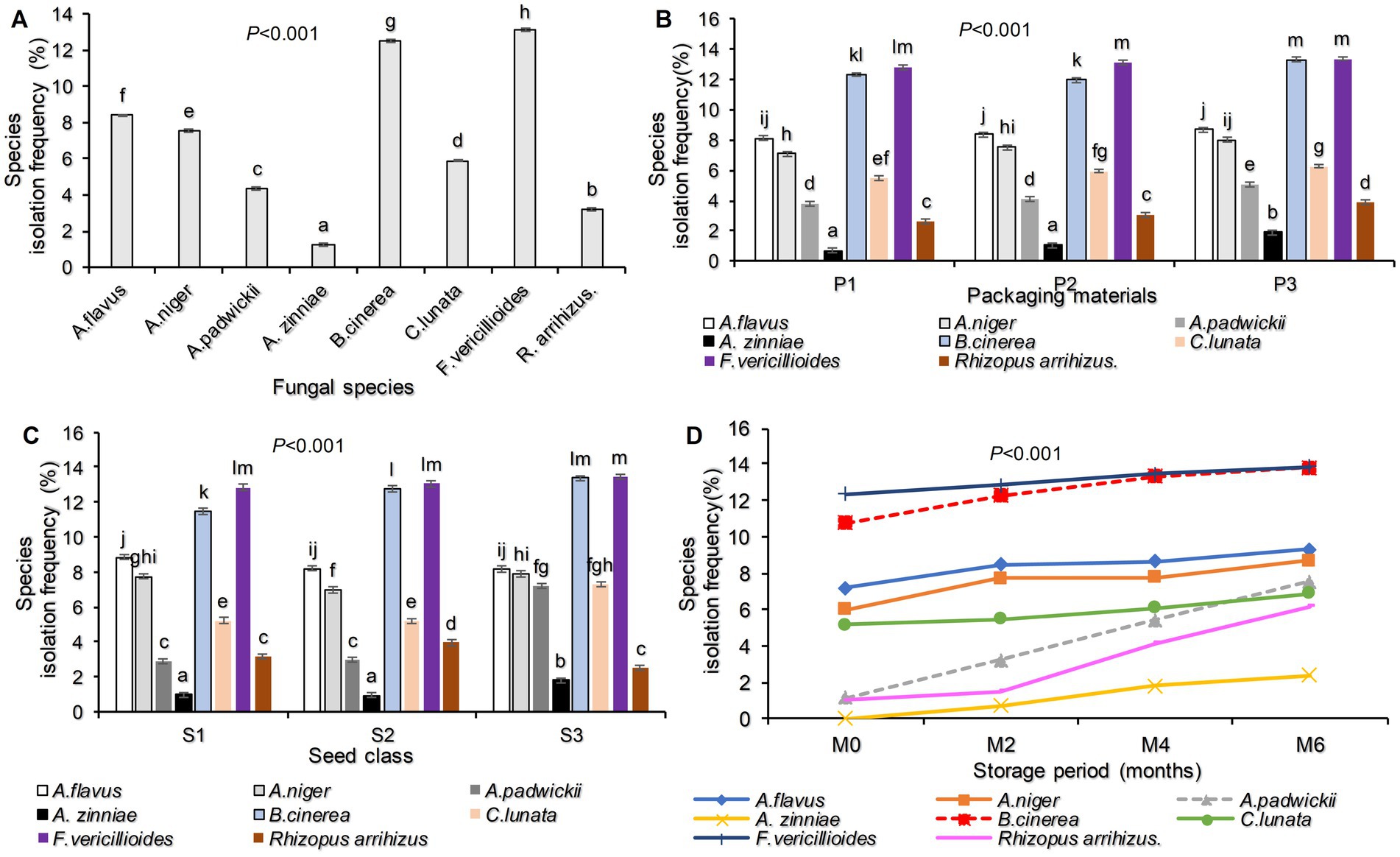
Figure 1. Fungal species isolation frequency as identified in stored seeds and their species (A); influenced by the interaction of fungal species and packaging material (B); influenced fungal species and seed class (C); influenced by fungal species and storage period (D) on stored sunflower seeds. Where P1 = Plastic container, P2 = Polypropylene bags and, P3 = sisal bags, S1 = certified seed, S2 = QDS, and S3 = Farm saved seeds.
Regarding the effects of packaging material and seed class, the isolation frequencies of fungal species were highly significantly different (p < 0.001; Figures 1B,C). Fusarium vericillioides had the highest isolation frequency of 13.38% for seeds stored in sisal bags, followed by Botrytis cinerea (13.29%) recorded in the same packaging materials. The lowest fungal isolation frequency was recorded for Alternaria zinnae (0.71%), followed by Rhizopus arrihizus (2.63%) in seeds packed in plastic containers (Figure 1B).
With respect to seed class, Fusarium vericillioides was also recorded as the most frequently isolated fungal species, with a 13.44% isolation frequency in farmer-saved seeds, followed by Botrytis cinerea (13.36%) in the same seed class. The lowest FIP was recorded with the Alternaria zinnae (0.93%) in QDS (Figure 1C). The isolation frequency of seed-borne fungal pathogens also varied with the longevity of the storage period. Figure 1D shows that fungal species isolation frequency increased with increasing storage time. It was observed that at the end of the 6th month storage period, Fusarium vericillioides had the highest isolation frequency of 13.85%, followed by Botrytis cinerea (13.78%), while Alternaria zinniae (2.41%) had the lowest frequency.
3.1.4 Fungal species isolation frequency as influenced by the interaction between seed class and packaging materials
The study established that the interaction between seed class and packaging materials significantly influenced the fungal species prevalence (p < 0.001; Figures 2A–C). The highest occurrence of.
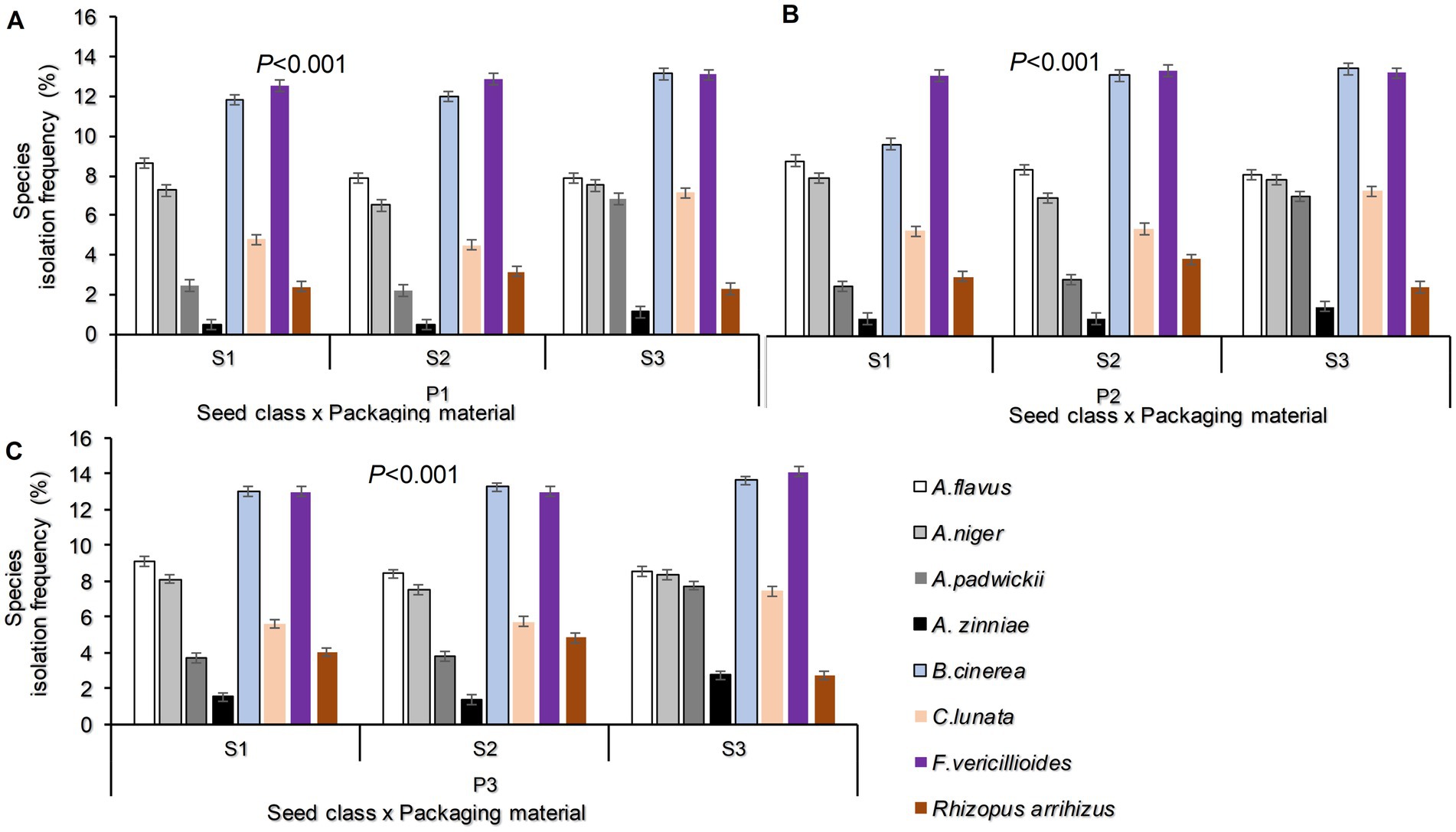
Figure 2. Fungal species isolation frequency as influenced by the interaction between Seed class and Packaging materials (A–C) Where S1 = certified seed, S2 = QDS and S3 = Farm saved seeds; P1 = Plastic container, P2 = Polypropylene bags and P3 = sisal bags.
A prevalence of 14.13% for Fusarium vericillioides was recorded in FSS stored in sisal bags, followed by Botrytis cinerea at 13.63%, which was also observed in the same seed class and packaging materials. The lowest prevalence of fungal species, Alternaria zinniae (0.5%), was found primarily in certified seeds and QDS stored in plastic containers.
3.1.5 Fungal species isolation frequency as influenced by the interaction between storage period and seed class
The incidence of pathogenic seed-borne fungal species was significantly affected by the interaction between storage periods and seed classes (p < 0.001; Figure 3). Consistently, the highest incidences of F. verticillioides and B. cinerea were observed in all sunflower seed classes, irrespective of their storage periods. Similarly, the incidences of A. zinnia were the lowest (Figure 3). In these results, it was noted that by the end of Month 6, Fusarium vericillioides recorded the highest infection percentage in FSS (14.39%), while Alternaria zinniae exhibited the lowest infection percentage (1.83%) in QDS (Figures 3A–C).
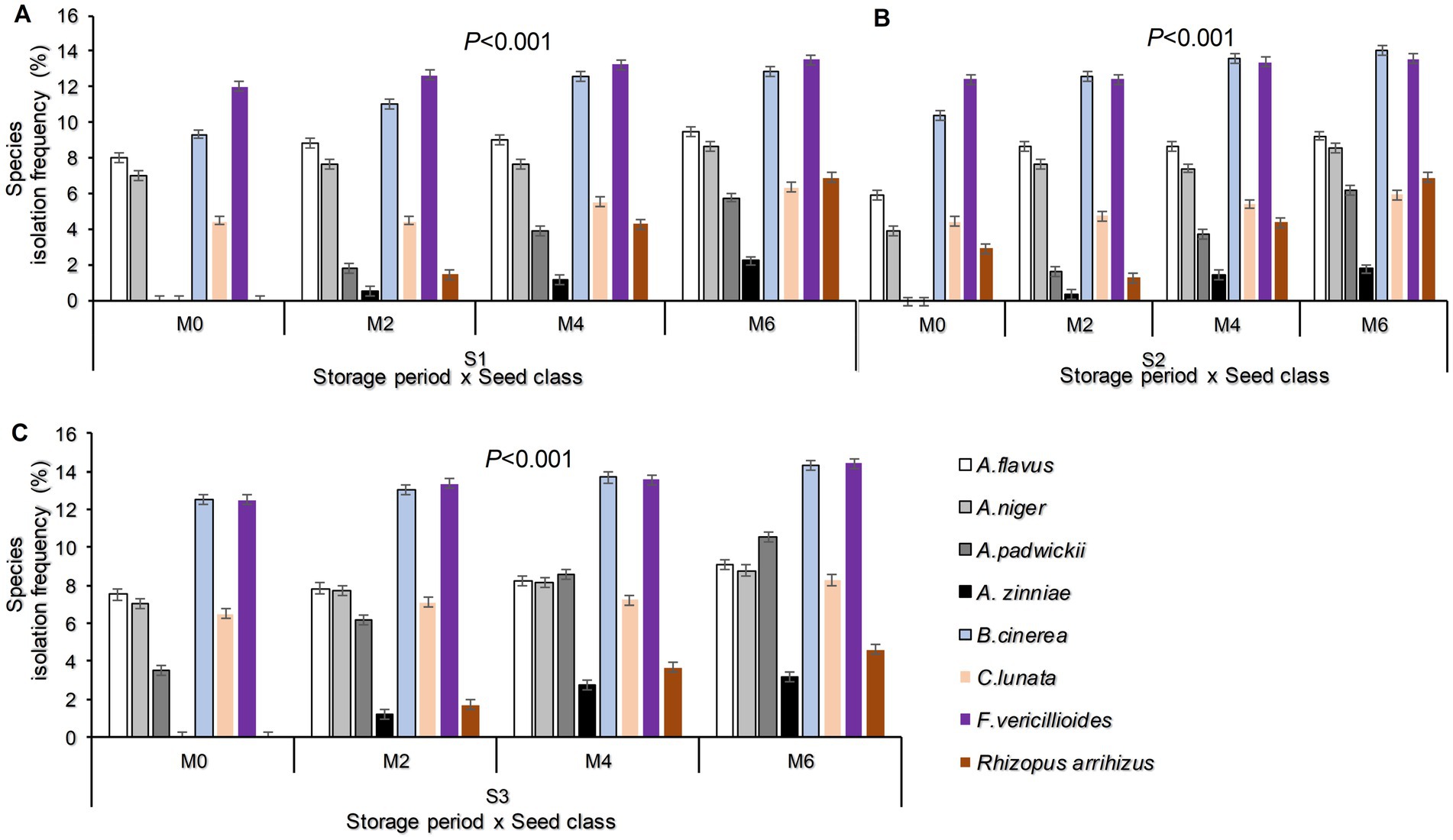
Figure 3. Isolation frequency of Fungal species as influenced by the interaction between storage period and seed class (A–C) Where M0, M2, M4, and M6 are Month 0 (Before storage), month 2, month 4, and month 6, respectively; S1 = Certified seed, S2 = QDS, and S3 = Farmer-saved seeds.
3.1.6 Total fungal infection incidence as influenced by the interaction of storage period and packaging materials, storage period, and seed class on stored sunflower seeds
The study revealed that the total fungal infection incidence was significantly influenced (p < 0.001) by the interaction between storage period and packaging materials, as well as storage period and seed class (Figure 4). Seeds stored in sisal bags had the highest total fungal infection (74.33%), followed by seeds in polypropylene bags (68.61%). Seeds packed in plastic containers had the lowest infection incidence (64.17%). In terms of seed class, FSS recorded the highest fungal infection incidence (73.11%), while Certified seeds had the lowest (67.06%), which was not significantly different from the infection incidence recorded with the QDS (66.94%; Figures 4A,B).
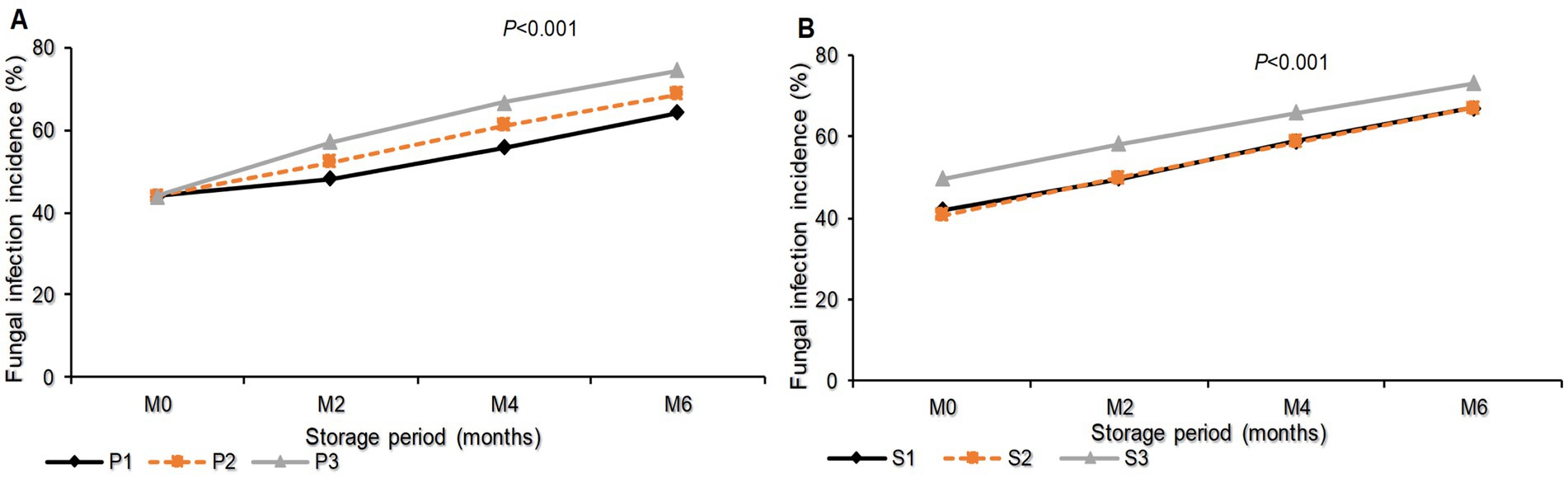
Figure 4. Total Fungal infection incidence as influenced by the interaction of Storage period and packaging materials (A) Storage period and seed class (B) on stored sunflower seeds. M0, M2, M4, and M6 are Month 0 (Before storage), month 2, month 4, and month 6, respectively; P1 = Plastic container, P2 = Polypropylene bags and P3 = sisal bags, S1 = certified seed, S2 = QDS and S3 = Farm saved seeds.
3.2 Influence on G (%) and SVI
From Table 1, the finding showed that seed G and SVI were highly significantly influenced (p < 0.001) by seed-borne fungi, packaging material, seed class, and storage period. The seeds stored in sisal bags recorded the smallest G and SVI, 78.58% and 100.2, respectively. This was followed by the seeds stored in polypropylene bags, while the plastic container recorded the highest G and SVI (83.97% and 119.8 respectively).
Additionally, the influence of seed class on G and SVI of stored sunflower seeds was highly significant (p < 0.001; Table 1). The lowest G and SVI of 68.58 and 88.8%, respectively, were followed by QDS. The maximum G and SVI of 88.17 and 123%, respectively, were recorded in certified seeds. In addition, the results show that G and SVI were significantly influenced (p < 0.001) by the storage period. As the storage period increased, both G (%) and SVI decreased significantly from 87, 83.93, 79.93, and 74.52% for G and 133.7, 117.2, 99.7 and 86.1 for the SVI (Table 1).
The G was not significantly influenced by the combination of packaging materials and seed class (p = 0.86), as shown in Table 2. However, the SVI was significantly affected by the interaction between packaging materials and seed class (p < 0.001). The highest SVI (138.1) was recorded with certified seeds packaged in plastic materials, while the smallest value (81.3) was noted in FSS packed in sisal bags (Table 2).
The interaction effect between packaging materials and storage period had significant effects (p < 0.001) on seed G (Figure 5A) and SVI (Figure 5B). Until the end of the storage period (month 6), the highest G (80.22%) and SVI (106.67) were recorded in seeds packed in a plastic container. This was followed by seeds packed in polypropylene bags with a seed G of 74.44% and an SVI of 85.76. The lowest G (68.89%) and SVI (66.0) were noted in seeds packed in sisal bags (Figures 5A,B).
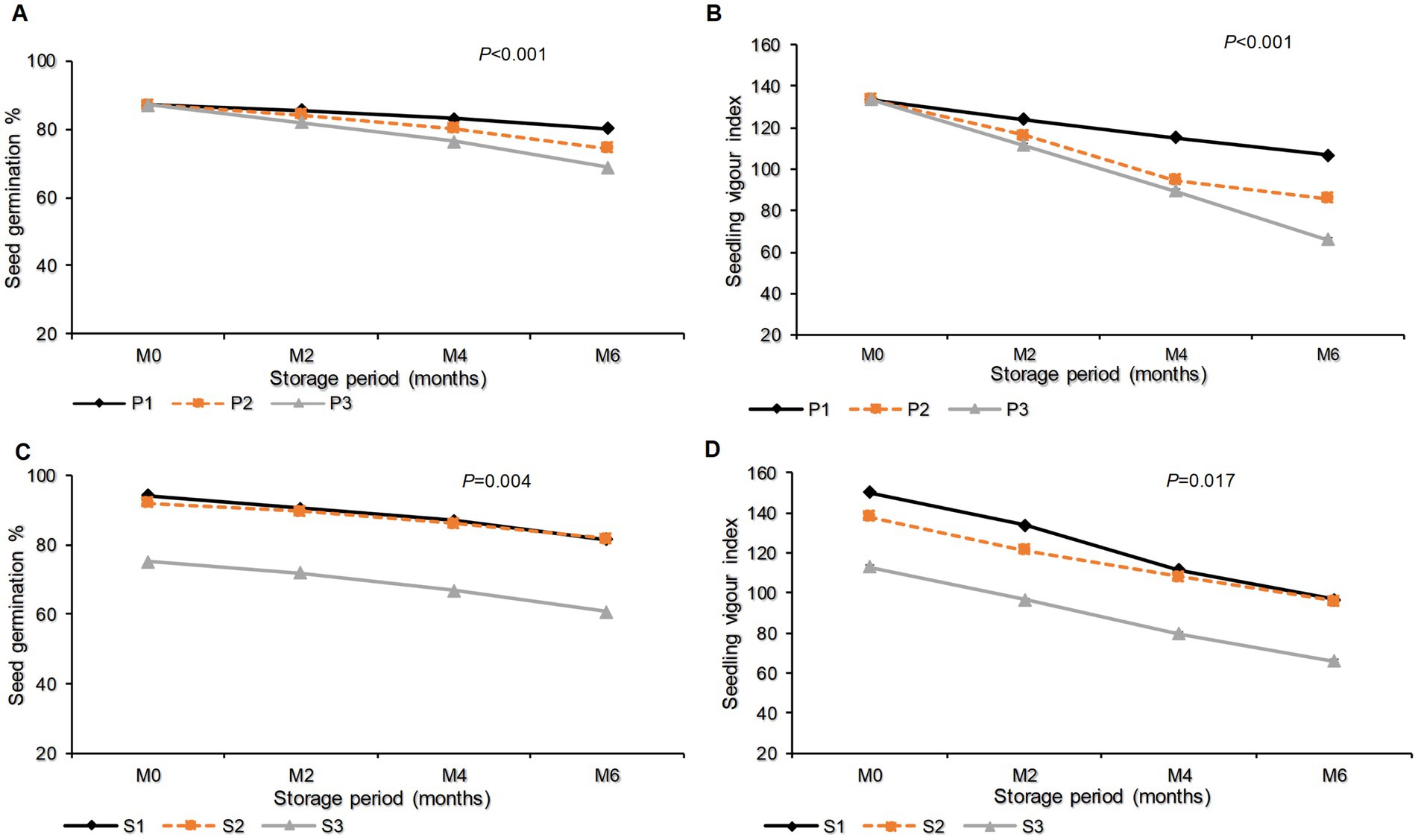
Figure 5. G and SVI as influenced by the interaction effect of packaging materials and storage period (A,B) and the interaction effect of storage period and seed class (C,D). Where M0, M2, M4, and M6 are Month 0 (Before storage), month 2, month 4, and month 6, respectively; P1 = Plastic container, P2 = Polypropylene bags and P3 = sisal bags, S1 = certified seed, S2 = QDS and S3 = Farm saved seeds.
Moreover, the effects of the interaction between seed class and storage period on G and SVI were significant (p = 0.004 and p = 0.017), respectively. In these results, it is clear that the QDS had the highest G (81.56%), which was not significantly different from the 81.33% recorded in certified seeds. The maximum SVI (96.76) was observed in certified seeds, which did not differ significantly from the 95.69 that was recorded in QDS. The minimum G (60.67%) and SVI (65.98) were recorded in FSS (Figures 5C,D).
4 Discussion
4.1 Fungal infection and seed-borne fungal species
This study provides evidence that the quality and health status of sunflower seeds depends on the type of storage material, seed class, and duration of storage. These results demonstrate that storing seeds in sisal bags is linked to high fungal infection rates. This could be attributed to their high permeability to moisture in the storage room, resulting in elevated seed moisture content. Consequently, this led to the rapid multiplication of fungi compared to less permeable packaging materials like plastic containers. The findings align with previous studies conducted by Shabana et al. (2015), Asha (2012), and Martín et al. (2022), who reported that highly permeable materials contribute to increased seed moisture content and thereby enhance the likelihood of fungal growth. Hasan et al. (2017) also found that seed moisture content significantly influenced seed viability and contamination by storage fungi.
The variations in infection rates among seed classes may be attributed to production practices. For instance, FSS is produced without using conventional practices for seed production, according to International Seed Testing Association (ISTA) (2022). This conclusion aligns with the reports by Dadlani and Yadava (2023); Selemani et al. (2023), FAO (2018), and Patharkar et al. (2013) highlight that seed quality during storage depends on factors such as seed class, initial seed quality, seed moisture content, temperature, and relative humidity. Additionally, other researchers noted varying percentages of fungal infection in stored seeds as the duration of storage increased, as observed by Wiewióra (2009) in spring barley, Kandhare (2018) in green-gram, Shabana et al. (2015) in maize, and Saeed et al. (2020) in cotton and wheat seeds.
The important fungal species recorded from stored seeds were Fusarium vericillioides, Botrytis cinerea, Aspergillus species, Alternaria spp., Curvularia lunata, and Rhizopus species. Among these, the dominant seed-borne pathogenic fungi found were Alternaria padwickii, Alternaria zinniae, Botrytis cinerea, Curvularia lunata, and Fusarium vericillioides. Also, three major saprophytic sunflower seed-borne fungi observed were Aspergillus flavus, A. niger, and Rhizopus arrihizus. All of these pathogenic and saprophytic seed-borne fungi were found to affect seed viability and seedling vigor. These findings were in harmony with those reported by Khalil et al. (2014) and El-Wakil (2014), who noted that the association of a large number of fungi with sunflower seeds included Alternaria species, Fusarium species, and Rhizoctonia species. These seed mycoflora are insignificant, as they contribute to seed deterioration, leading to considerable seed losses. This may be due to the secretion of mycotoxins and fungal secondary metabolites, which severely reduce seed quality and quantity. Selemani et al. (2023), Chau et al. (2019), and Martín et al. (2022), who stored seeds at different storage conditions and moisture contents, reported that lower storage temperatures between 5 and 20°C and seed moisture content of 6–8% observed less fungal infection compared to seeds stored at high temperature of 35°C with high moisture content.
The findings also align with the study conducted by Patil et al. (2018), which identified Alternaria alternata, Fusarium oxysporum, as well as three saprophytic fungi (Aspergillus flavus, A. niger, and Rhizopus stolonifer) as the major phytopathogenic fungi present in the sunflower varieties examined. These seed-borne fungi are known to adversely affect seed health and vigor. Previous studies by Ghoneem et al. (2014), Kandhare (2018), and Chandel and Kumar (2017) have documented that infected seeds serve as a means of disseminating various crop diseases to different regions. Factors such as high relative humidity, suitable temperature, and increased moisture content contribute to the establishment of pests and diseases within seeds. Infection by these fungi has been observed in all parts of the seeds, leading to external or internal damage, including seed rot, necrosis, and seedling diseases (Deshmukh and Kare, 2010). Numerous researchers have previously reported that these fungi species cause the deterioration of seeds, consequently reducing the viability and vigor of stored seeds (De Vitis et al., 2020; Martín et al., 2022; Gebeyaw, 2020; Chhabra and Singh, 2019; Mycock and Berjak, 2017; Pollard, 2018).
4.2 Influence on G and SVI
The detected seed-borne fungi were found to influence seed germination and SVI. Concerning seed class, G and SVI were recorded as the lowest in the FSS (68.58% and 88.8, respectively) in seed class, with the highest FIP followed by QDS, while the maximum was recorded in certified seeds (88.17% and 123, respectively), the seed class with the less fungal infection. This is similar to the findings of other researchers, Selemani et al. (2023) and Nehra et al. (2021), who reported that seeds with less fungal infection in storage recorded a high G compared to seeds with high fungal infection incidences.
The seed-borne fungal pathogens arising from seed storage were possibly the cause of reduced seed germination and seedling vigor. Fusarium species were reported to cause seedling rot, as most protuberant symptoms exhibited in sunflowers and other crops. In this situation, the findings of the current research become in line with earlier reports by other researchers, including Afzal et al. (2010) in sunflower, Singh et al. (2003) in pearl millet, Ahammed et al. (2006) in soybean, and Nagaraja and Krishnappa (2009) in niger crop. Aspergillus and Alternaria species are responsible for reducing seed quality, protein, and carbohydrate contents by damaging seedlings, causing root collar, seed rot, and damping off seedlings. These all together result in the reduction of crop yield qualitatively and quantitatively. This was further reported by Anjorin and Mohammed (2014), Masomeh et al. (2012), and Lambat et al. (2014).
Field fungi may weaken or kill embryos, while storage fungi gradually destroy the embryos of the seeds they infect (Martín et al., 2022). Furthermore, Botrytis cinerea has been identified as the cause of grey mold disease in sunflower crops, as found by Williamson et al. (2007). Seedlings grown from such infected seeds often exhibit reduced vigor (Gebeyaw, 2020). Seed-borne pathogenic fungi can hinder germination, kill seedlings, or impair plant growth by damaging the roots and vascular system, thereby disrupting the transport of water and nutrients. Similar effects have been reported in sunflower, common bean, and peanut crops by Hatim et al. (2022), Marcenaro and Valkonen (2016), and Aslam et al. (2015), respectively.
5 Conclusion and recommendation
Based on the current research, it can be deduced that the fungi transmitted through the seeds were discovered to have a detrimental impact on both the viability of the seeds in terms of germination and the vigor index of the resulting seedlings in sunflower crops. As the duration of storage increased, a decrease in the percentage of seed germination and the SVI was observed. Consequently, it is strongly advised to utilize high-quality seeds, including certified seeds and QDS, and employ appropriate storage materials to preserve seeds effectively for future sowing seasons. Such measures will help ensure optimal crop productivity and success. The remarkable limitation of this study is the short duration of storage. The study only considered a 6-month storage period. The long-term impact of fungal infections on seed viability and vigor might not have been fully captured. Future studies could extend the storage period beyond 6 months to assess the effects of fungal infection over a longer duration, such as a year or more, to better understand long-term seed quality.
Based on the findings of this research, potential directions for future studies include investigating the efficacy of various fungicides and biocontrol methods to reduce fungal infections during seed storage. Another avenue is exploring genetic resistance to fungal infections by identifying sunflower varieties with natural resistance to common seed-borne fungal pathogens. Additionally, future studies could focus on developing Integrated Seed Management Systems that combine optimal storage techniques, seed treatment methods, and fungal control strategies.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Author contributions
SS: Conceptualization, Formal analysis, Methodology, Writing – original draft. RM: Investigation, Supervision, Writing – review & editing. YN: Data curation, Supervision, Validation, Writing – review & editing.
Funding
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abd-El-Aziz, S. A., and El-Satar, A. (2016). Effect of storage durations on fungal infection, aflatoxin contamination, quality and yield of sunflower seeds. Zagazig J. Agricul. Res. 43, 1965–1975.
Abdul-Baki, A. A., and Anderson, J. D. (1973). Vigor determination in soybean by multiple criteria. Crop Sci. 13, 630–633. doi: 10.2135/cropsci1973.0011183X001300060013x
Afzal, R., Mughal, S. M., Munir, M., Sultana, K., Qureshi, R., Arshad, M., et al. (2010). Mycoflora associated with seeds of different sunflower cultivars and its management. Pak. J. Bot. 42, 435–445.
Ahammed, S. K., Anandam, R. J., Prasad Babu, G., Munikrishnaiah, M., and Gopal, K. (2006). Studies on seed mycoflora of soybean and its effect on seed and seedling quality characters. Legum. Res. 29, 186–190. doi: 10.18805/LR-225
Anjorin, S. T., and Mohammed, M. (2014). Effect of seed-borne fungi on germination and seedling vigor of watermelon (Citrullus lanatus thumb). African J. Plant Sci. 8, 232–236. doi: 10.5897/AJPS08.090
Asha, A. M. (2012). Effect of plant products and containers on storage potential of maize hybrid. M. Sc. (Agri.) Thesis. University of Agricultural Sciences, Dharwad. Karnataka, India.
Aslam, M. F., Irshad, G., Naz, F., Aslam, M. N., and Ahmed, R. (2015). Effect of seed-borne mycoflora on germination and fatty acid profile of peanuts. Pakistan J. Phytopathol. 27, 131–138. doi: 10.5555/20163084047
Balchin, Neil, Kweka, Josaphat, and Mseri, Maximiliano Mendez-Parra Honest, (2018). “ANSAF Tarif setting for the development of the edible oil sector” in Tanzania 4th annual agricultural policy conference (AAPC) 14Th -16th, February 2018. Pp. 1–12.
Bank of Tanzania (2017). Potentiality of sunflower sub-sector in Tanzania. Tanzania: Bank of Tanzania, Dar-Es-Salaam.
Chandel, S., and Kumar, V. (2017). Effect of plant extracts as pre-storage seed treatment on storage fungi, G and seedling vigor of pea (Pisum sativum). Indian J. Agric. Sci. 87, 1476–1481. doi: 10.56093/ijas.v87i11.75703
Chau, M. M., Chambers, T., Weisenberger, L., Keir, M., Kroessig, T. I., Wolkis, D., et al. (2019). Seed freeze sensitivity and ex-situ longevity of 295 species in the native Hawaiian flora. Am. J. Bot. 106, 1248–1270. doi: 10.1002/ajb2.1351
Chhabra, R., and Singh, T. (2019). Seed aging, storage and deterioration: an irresistible physiological phenomenon. Agric. Rev. 40, 234–238. doi: 10.18805/ag.R-1914
Dadlani, M., and Yadava, D. K. (2023). Seed science and technology: biology, Production, quality. Cham, Switzerland: Springer Nature, 430.
De Vitis, M., Hay, F. R., Dickie, J. B., Trivedi, C., Choi, J., and Fiegener, R. (2020). Seed storage: maintaining seed viability and vigor for restoration use. Restor. Ecol. 28, S249–S255. doi: 10.1111/rec.13174
Deshmukh, A. M., and Kare, M. A. (2010). Study of seed mycoflora of some oilseed crops. Bioinfolet 7, 295–297.
El-Wakil, D. A. (2014). Seed-bome fungi of sunflower (Helianthus annuus L.) and their impact on oil quality. IOSR J. Agricul. Vet. Sci. 6, 38–44.
FAO, (2017) FAOSTAT data production, Trade, Food Balance, Food Security. Food and Agriculture Organization of the United Nations (FAO).
FAO (2018). Seeds Toolkit-Module 6: Seed storage. Rome: Food and Agriculture Organization of United Nations, 13–79.
Gebeyaw, M. (2020). Review on: impact of seed-borne pathogens on seed quality. American J. Plant Biol. 5, 77–81. doi: 10.11648/j.ajpb.20200504.11
Genes, F., and Nyomora, A. M. S. (2018). Effect of storage time and temperature on germination ability of Escoecaria bussei. Tanzania J. Sci. 44, 123–133. doi: 10.4314/tjs.v44i1.12
Ghoneem, K. M., Ezzat, S. M., and El-Dadamony, N. M. (2014). Seedborne fungi of sunflower in Egypt with reference to pathogenic effects and their transmission. Plant Pathol. J. 13, 278–284. doi: 10.3923/ppj.2014.278.284
Guney, Y., Kallinterakis, V., and Komba, G. (2017). Herding in frontier markets: evidence from African stock exchanges. J. Int. Financ. Mark. Inst. Money 47, 152–175. doi: 10.1016/j.intfin.2016.11.001
Hasan, K., Sabagh, E. L., Celaleddin, B., and Islam, M. S. (2017). Seed quality of lentil (Lens culinaris L.) as affected by different containers and storage periods. Scientific. J. Crop. Sci. 6, 146–152. doi: 10.14196/SJCS.V6I1.2382
Hatim, S. H., Al-Salami, I., and Jabbar, M. K. (2022). Detection of seed-borne Fungi associated with three sunflower cultivars and their effect on seed germination. Int. J. Agricul. Statistical Sci. 18, 2041–2045. doi: 10.48165/ijass.2022.1801.2041
International Seed Testing Association (ISTA) (2022), International rules for seed testing; international seed testing association: Zürichstr. 50, CH-8303 Bassersdorf, Switzerland : International Seed Testing Association.
Isinika, A. C., and John, J. (2021). The political economy of sunflower in Tanzania: A case of Singida region : LAP Lambert Academic Publishing, 1–32.
Kandhare, A. S. (2018). Effect of storage containers on seed Mycoflora and seed health of green gram (Vigna radiate L.) and its cure with botanicals. J. Agricul. Res. Technol. 14:555912. doi: 10.19080/ARTOAJ.2018.14.555912
Kator, L., Ogo-Oluwa, A. T., and Kemi, A. B. (2016). Isolation and identification of seed borne fungi of common bean (Phaseolus vulgaries L.) from selected markets in Makurdi. International journal of. Agric. Sci. 2, 75–78. doi: 10.11648/j.ijaas.20160205.11
Khalil, A. A., Elwak-il, D. A., and Ghonim, M. I. (2014). Mycoflora association and contamination with aflatoxins in sunflower (Helianthus annuus L.) seeds. Int. J. Plant Soil sci. 3, 685–694. doi: 10.9734/IJPSS/2014/8571
Konyali, S. (2017). Sunflower production and agricultural policies in Turkey. Sosyal Bilimler Araştırma Dergisi 6, 11–19.
Kumar, R., Gupta, A., Srivastava, S., Devi, G., Singh, V. K., Goswami, S. K., et al. (2020). Diagnosis and detection of seed-borne fungal phytopathogens. Seed-Borne Diseases of Agricultural Crops: Detection, Diagnosis & Management. 107–142.
Lambat, A., Patil, S., Charjan, S., Gadewar, R., Babhulkar, V., and Lambat, P. (2014). Effect of storage on seed germination, seedling vigor and Mycoflora on cowpea. Int. J. Res. Biosci. Agricul. Technol. 2, 86–90.
Lyanga, T. M. (2024). Analysis of costs of inputs for sunflower production at Mkalama District in Tanzania. Pan-African J. Business Manag. 8, 37–57. doi: 10.61538/pajbm.v8i1.1523
Marcenaro, D., and Valkonen, J. P. T. (2016). Seedborne pathogenic Fungi in common bean (Phaseolus vulgaris cv. INTA Rojo) in Nicaragua. PLoS One 11:e0168662. doi: 10.1371/journal.pone.0168662
Martín, I., Gálvez, L., Guasch, L., and Palmero, D. (2022). Fungal pathogens and seed storage in the dry state. Plan. Theory 11:3167. doi: 10.3390/plants11223167
Masomeh, H., Hajihassani, A., and Khaghani, S. (2012). Incidence and distribution of seed-borne fungi associated with wheat in Markazi Province. Afr. J. Biotechnol. 11, 6290–6295. doi: 10.5897/AJB11.3838
Mathur, S. B., and Kongsdal, O. (2003). Common laboratory seed health testing methods for detecting fungi. Int. Seed Testing Association., 1–425.
Mgeni, C. P., and Mpenda, Z. T. (2021). Can sub-Saharan Africa become food self-sufficient? Analyzing the market demand for sunflower edible oil in Tanzania. Tanzania J. Agricul. Sci. 20, 42–53. doi: 10.4314/tjas.v20i1.5
Muradov, A., Hasanli, Y., and Hajiyev, N. (2019). World market Price of oil. Cham, Switzerland: Springer, Impacting factors and forecasting, 184.
Mycock, D. J., and Berjak, P. (2017). “The implications of seed-associated mycoflora during storage” in Seed development and germination. Eds. J. Derek Bewley and C. W. Black (New York: Routledge), 747–766.
Nagaraja, O., and Krishnappa, M. (2009). Seedborne mycoflora of Niger (Guizotia abyssinica cass.) and its effect on germination. Indian Phytopathol. 62, 513–517. doi: 10.1007/s42360-019-00047-2
Nahar, S., Mushtaq, M., and Hashmi, M. H. (2005). Seedborne mycoflora of sunflower. Eds. Shubha B.N, Pradeep K. Patil and Indu S.S (Duxford, UK: Helianthus annuus L.). Pak. J. Bot. 37, 451–457. doi: 10.33866/phytopathol.029.01.0303
Nehra, S., Gothwal, R. K., Varshney, A. K., Solanki, P. S., Chandra, S., Meena, P., et al. (2021). “Bio management of fusarium spp. associated with oil crops” in Microbiome stimulants for crops (Duxford, UK. Woodhead Publishing), 453–474.
Niaz, I., and Dawar, S. (2009). Detection of seed borne mycoflora in maize (Zea mays L.). Pak. J. Bot. 41, 443–451. doi: 10.5555/20093118623
Patharkar, S. P., Sontakke, N. R., and Hedawoo, G. B. (2013). Evaluation of seed mycoflora and G in Helianthus annuus L. Int. J. Innovations in Bio Sci. 3, 1–5.
Patil, A. C., Surpawanshi, A. P., Anbhule, K. A., Raner, R. B., and Hurule, S. S. (2018). Detection of sunflower seedborne mycoflora and their effect on seed and seedling parameters. Int. J. Curr. Microbiol. App. Sci. 6, 2509–2514.
Pollard, A. T. (2018). Seeds vs fungi: an enzymatic battle in the soil seedbank. Seed Sci. Res. 28, 197–214. doi: 10.1017/S0960258518000181
Reuben, F. M., and Meliyo, J. L. (2022). Unlocking opportunities in edible oil crop production and market dynamics to accelerate agricultural Investment in Kagera Region, Tanzania. Int. J. Environ. Agricul. Biotechnol. 7, 128–132. doi: 10.22161/ijeab.73.17
Saeed, M. F., Jamal, A., Ahmad, I., Ali, S., Shah, G. M., Husnain, S. K., et al. (2020). Storage conditions deteriorate cotton and wheat seeds quality: an assessment of farmers’ awareness in Pakistan. Agronomy 10, 1–19. doi: 10.3390/agronomy10091246
Sebyiga, B. (2020). Sunflower production and its potential for improving income of smallholder producers in the central agricultural zone of Tanzania: a case of villages in Kongwa and Singida rural districts. Local Admin. J. 13, 223–234. doi: 10.14456/laj.2020.19
Selemani, S., Madege, R., and Nzogela, Y. (2023). Influence of storage conditions on viability and vigour of sunflower (Helianthus annuus L.) seeds. Journal of current opinion. Crop Sci. 4, 141–153. doi: 10.62773/jcocs.v4i3.211
Shabana, Y. M., Ghazy, N. A., Tolba, S. A., and Fayzalla, E. A. (2015). Effect of storage condition and packaging material on incidence of storage Fungi and seed quality of maize grains. J. Plant Protection and Pathol. 6, 987–996. doi: 10.21608/jppp.2015.74662
Singh, S. D., Swami, S. D., and Rawal, P. (2003). Seed mycoflora of pearl millet (Pennisetum glaucum) and its control. Plant Dis. Res. 18, 115–118.
Sserumaga, J. P., Makumbi, D., Simyung, L., Njoroge, K., Muthomi, J. W., Chemining’wa, G. N., et al. (2015). Incidence and severity of potentially toxigenic aspergillus flavus in maize (Zea mays L.) from different major maize growing regions of Uganda. Afr. J. Agric. Res. 10, 1244–1250. doi: 10.5897/AJAR2015.9523
Tibamanya, F. Y., Henningsen, A., and Milanzi, M. A. (2022). Drivers of and barriers to adoption of improved sunflower varieties amongst smallholder farmers in Singida, Tanzania: a double-hurdle approach. Q Open 2:qoac008. doi: 10.1093/qopen/qoac008
Wiewióra, B. (2009). Long-time storage effect on the seed heath of spring barley grain. Plant Breeding Seed Sci. 59, 3–12. doi: 10.2478/v10129-009-0001-y
Keywords: packaging materials, seed-borne fungi, seedling vigor, sunflower, viability
Citation: Selemani S, Madege R and Nzogela Y (2025) Incidence of seed-borne fungi of stored sunflower seeds (Helianthus annuus) in Tanzania and their influence on seed viability and vigor. Front. Sustain. Food Syst. 9:1534929. doi: 10.3389/fsufs.2025.1534929
Edited by:
Pushp Sheel Shukla, Centre for Cellular and Molecular Platforms, IndiaReviewed by:
Zorica Nikolic, Institute of Field and Vegetable Crops, SerbiaNythri Tshering, Royal University of Bhutan, Bhutan
Copyright © 2025 Selemani, Madege and Nzogela. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Siwajali Selemani, c2VsZW1hbmlzaXdhamFsaUBnbWFpbC5jb20=