 M. Lucchetta
M. Lucchetta M. Y. A. Zuluaga
M. Y. A. Zuluaga S. Monterisi
S. Monterisi R. Fattorini
R. Fattorini F. Gaiotti
F. Gaiotti S. Cesco
S. Cesco Y. Pii
Y. Pii- 1Faculty of Agricultural, Environmental and Food Sciences, Free University of Bozen/Bolzano, Bolzano, Italy
- 2Council for Agricultural Research and Economics-Research Centre for Viticulture and Enology (CREA-VE), Conegliano, Italy
Healthy and fertile soils are the foundation of sustainable global agriculture, supporting both intensive and extensive crop cultivation, including orchards and vineyards across diverse agroecosystems. However, widespread use of synthetic fertilizers has degraded soil organic matter and overall quality, leading to significant environmental consequences and threats to ecosystem services. Challenges in accessing mineral fertilizers, along with growing concerns over environmental sustainability, have driven interest in alternative approaches such as compost, biofertilizers, biopesticides, and microbiome-based strategies to enhance soil fertility and reduce chemical inputs in agriculture. Compost derived from grapevine pruning waste shows promise in addressing agricultural challenges, but further research is needed to understand its effects on nutrient release dynamics and microbial activity. Inoculating compost with a fungal consortium presents an innovative strategy to enhance nutrient cycling and microbial interactions, addressing nutrient availability challenges. This study aimed to evaluate the impact of three fertilization methods on soil health: NPK (mineral fertilizer), PWC (pruning waste compost), and iPWC (pruning waste compost inoculated with fungal consortia). Conducted over 135 days in a controlled growth chamber, fifteen pots received equal macronutrient doses. Environmental parameters (humidity, temperature, light/dark cycles) were closely monitored. Throughout the experiment, release dynamics of key nutrients, soil enzyme activity, and microbial community responses were rigorously assessed. The results showed that compost inoculated with fungi significantly enhanced soil enzymes activities. Microbial community activity was higher in fungal and compost treatments, with greater functional diversity observed in the mineral fertilizer treatment. Compost released major minerals more slowly than chemical fertilizers, while enhancing the available fraction. These findings underscore compost’s potential, especially when supplemented with fungi, to improve soil health and promote sustainable agricultural practices and soil regeneration.
1 Introduction
Soil quality and fertility are essential for sustainable development, as highlighted by the Sustainable Development Goals (SDGs) (Lal et al., 2021). In agriculture, maintaining soil health is critical, especially given the challenges posed by soil degradation, which is often linked to the intensive use of mineral fertilizers and cropping practices (Scotti et al., 2015; Pahalvi et al., 2021). This issue is particularly pronounced in Europe, where viticulture is widespread (Eurostat, 2024). The intensive nature of grape production requires substantial inputs of fertilizers and phytosanitary products, contributing to soil acidification (Litskas et al., 2020), biodiversity loss, and soil fertility depletion (Karimi et al., 2020).
In response to these challenges, the European Union’s Farm to Fork strategy (Directorate-General for Agriculture and Rural Development, 2024) advocates for reducing chemical inputs and enhancing soil fertility. Organic amendments, such as compost, manure, and cover crops, have emerged as promising solutions in viticulture. These amendments can significantly improve soil structure, increase nutrient availability, and enhance grapevine resistance to pests and diseases (Diacono and Montemurro, 2011; Mondini et al., 2018; Lazcano et al., 2020; Romero et al., 2022). However, organic amendments can also lead to unintended consequences. For instance, a study by Fan et al. (2023) reported increased greenhouse gas emissions, primarily methane, when organic amendments replaced synthetic fertilizers. Similarly, Burgos et al. (2006) found that municipal waste compost can increase nitrate leaching, reducing nitrogen availability for plants. Additional studies have highlighted adverse effects such as soil immobilization due to high C/N ratios, leading to the depletion of readily available nitrogen for plant uptake (Schoenau and Davis, 2006; Moral et al., 2009).
Given these complexities, evaluating various organic amendments, including compost from grapevine pruning waste, presents a viable alternative. The application of pruning waste compost (PWC) adheres to circular economy principles, transforming waste into a valuable resource (Velasco-Muñoz et al., 2022), this practice can reduce chemical inputs and contribute to a more sustainable viticulture approach. Studies have shown that pruned grapevine wood (2 tons per hectare per year) can improve and maintain soil organic matter and properties without adverse effects over the long term (Morlat and Chaussod, 2008). Other researchers have observed that PWC enhances soil microbial activity (García-Orenes et al., 2016; Mondini et al., 2018), and improves grapevine yield and quality (Gaiotti et al., 2017). Economically, PWC is a renewable, cost-effective resource that promotes soil biological parameters (Karimi et al., 2020), making it a sustainable solution for vineyard nutrition management.
Recent studies have focused on the benefits of arbuscular mycorrhizal fungi (AMF) and multifunctional fungi such as Trichoderma in enhancing soil health (Balestrini et al., 2020; El Enshasy et al., 2020). AMF application enhances nutrient availability to plants by forming symbiotic relationships with roots, which is crucial for nutrient-deficient soils (Wipf et al., 2019). Additionally, T. harzianum enhances soil structure by promoting soil particle aggregation, thereby improving water and nutrient retention (O’Neill et al., 1996; Elad, 2000; Bernal-Vicente et al., 2015) and improving grape quality, yield, and plant nutrition (Pascale et al., 2017).
The incorporation of fungi into compost serves a dual purpose: it not only leverages the benefits of the fungi but also enhances the compost’s overall efficacy as a fertilizer. Compost can provide a conducive environment for fungal growth and activity, improving colonization rates compared to separate applications of dry or liquid inocula. This compost-mediated delivery mechanism may enhance the effectiveness of the inoculum by ensuring sustained contact with plant roots and fostering beneficial microbial interactions within the soil matrix.
For example, T. harzianum has been shown to induce resistance to abiotic stresses such as drought and salinity (Hidangmayum and Dwivedi, 2018). In Vitis vinifera, AMF and T. harzianum have demonstrated positive effects on antioxidant synthesis, shoot weight, and stress resilience. Hao et al. (2012) identified an increase in potted grapevine shoot fresh weight following treatment with T. harzianum, which also exhibited nematode-suppressing effects. Furthermore, the impacts of Glomus (Syn. Funneliformis) mosseae in potted vineyards, particularly under drought stress conditions of Cabernet Sauvignon, yielded encouraging findings regarding stomatal conductance and enhanced cytokinin production.
These findings suggest that inoculating compost with a consortium of these fungi may enhance its properties, leading to a more valuable and sustainable fertilizer for vineyards.
Despite the promising outlooks provided by previous studies, there is currently a paucity of literature directly evaluating the effects of pruning waste compost (PWC) on soil fertility parameters. Notably, no studies have investigated the potential of a fungi consortium inoculation to enhance PWC properties and further improve soil health, so far. Considering these observations, our study focused on comparing the effects of uninoculated pruning waste compost (PWC), inoculated PWC, and standard mineral fertilizer on soil fertility parameters, microbial community structure and function, and soil enzyme activities. In this context, our research was aimed at determining the nutrient release kinetics for the main macronutrients (N, P, and K) and assess the response of the microbial community to the treatments. Additionally, to further investigate the influence of the treatments on soil quality and fertility parameters, we assessed the activities of key enzymes involved in nutrient acquisition and cycling.
The preservation of soil quality and fertility is fundamental to achieving sustainable agricultural development, as highlighted by the Sustainable Development Goals (SDGs) (Lal et al., 2021). However, soil degradation remains a significant challenge, particularly in agriculture, where intensive cropping and the excessive use of mineral fertilizers have led to the depletion of essential soil properties (Scotti et al., 2015; Pahalvi et al., 2021). The reliance on synthetic fertilizers poses a significant challenge in Europe, particularly due to the region’s status as a global leader in wine production, with three of the largest wine-producing countries located in Europe. Viticulture is not only integral to the agricultural sector but also plays a vital role in the broader European economy (Eurostat, 2024). The intensive nature of vineyard management, driven by the high economic stakes, has led to increased fertilizer use, raising concerns about soil degradation, environmental sustainability, and the long-term viability of grape production in the region. The intensive nature of grape cultivation demands high inputs of fertilizers and phytosanitary products, often resulting in soil acidification, loss of biodiversity, and long-term fertility decline (Karimi et al., 2020; Litskas et al., 2020). Furthermore, in 2022, consumption of mineral fertilizers in the EU-27 experienced a significant decline, reaching levels not seen since 2009. This downturn was largely driven by the energy crisis that followed the Russian invasion of Ukraine, which had a profound impact on the European industry. Notably, consumption of nitrogen, phosphate and potash fertilizers decreased by 11, 16 and 15%, respectively (Fertilizers Europe, 2023). This raises concerns about the reliability of these fertilizers for viticultural production.
In response to these challenges, the European Union’s Farm to Fork strategy (Directorate-General for Agriculture and Rural Development, 2024) promotes reducing chemical inputs and enhancing soil fertility through sustainable practices. Organic amendments such as compost, manure, and cover crops have emerged as viable alternatives to synthetic fertilizers in viticulture. These amendments have been shown to improve soil structure, increase nutrient availability, and bolster grapevine resistance to pests and diseases (Diacono and Montemurro, 2011; Mondini et al., 2018; Lazcano et al., 2020; Romero et al., 2022). However, the use of organic amendments is not without potential drawbacks. For instance, some studies have reported increased greenhouse gas emissions, particularly methane, when organic amendments are used in place of synthetic fertilizers (Fan et al., 2023). Other studies have noted increased nitrate leaching (Burgos et al., 2006) and nitrogen immobilization due to high carbon-to-nitrogen (C/N) ratios, which may reduce the immediate availability of nitrogen for plant uptake (Schoenau and Davis, 2006; Moral et al., 2009). The use of green-waste compost has demonstrated significant benefits in various agricultural contexts, underscoring its potential for enhancing soil health and productivity in viticulture. Eldridge et al. (2018) conducted a long-term study in peri-urban vegetable production, comparing different compost treatments with conventional farming practices. The results revealed that compost treatments, even when supplemented with inorganic fertilizers, consistently outperformed conventional methods in terms of yield and economic benefits. Notably, a one-time application of green-waste compost significantly improved crop yields over 10 crops, highlighting the potential of compost to close yield gaps by improving soil quality. Such findings are relevant to soils of perennial crops, where improving soil structure and nutrient availability through compost application could similarly enhance grape yield and quality, while promoting sustainable soil management practices. In urban settings, where soil degradation and compaction are common, the application of municipal green-waste compost (MGWC) has been shown to improve soil properties and plant growth (Somerville et al., 2018). This research, focused on the establishment of deep-rooted woody plants, demonstrated that compost amendments enhanced soil physical characteristics such as bulk density and hydraulic conductivity. The implications of this for sustainable cultivation management are clear: compost can play a crucial role in restoring soil functions, particularly in compacted soils, by improving water infiltration, gas exchange, and root development. However, as the study also notes, site-specific soil conditions must be considered, as the benefits of compost may vary depending on factors such as nitrogen immobilization.
Despite these complexities, the use of grapevine pruning compost (PWC) presents a promising alternative, aligning with circular economy principles by transforming agricultural waste into a valuable resource (Velasco-Muñoz et al., 2022). Pruning waste compost exemplifies a practical application of circular economy principles within viticulture, as it allows for the reuse of organic materials that would otherwise contribute to agricultural waste. By converting vineyard residues into nutrient-rich compost, this process closes the nutrient loop, reducing dependency on synthetic fertilizers and minimizing environmental pollution. As discussed by De Corato (2020), the reuse of agricultural waste not only enhances soil quality but also contributes to broader sustainability goals, supporting ecosystem resilience and reducing waste disposal issues. The circular economy framework emphasizes the efficient use of resources by keeping materials in use for as long as possible, thus extending their lifecycle and value. Pruning waste compost fits seamlessly into this model by transforming biomass into valuable inputs for soil health, improving nutrient cycling, and enhancing plant growth. This approach contributes to resource conservation while fostering more sustainable production systems in agriculture. Moreover, adopting composting practices aligns with the European Union’s commitment to sustainable development and climate goals, reducing the carbon footprint of agricultural practices and supporting long-term soil fertility and crop productivity. Research has shown that PWC can enhance soil organic matter and improve soil properties without adverse long-term effects (Morlat and Chaussod, 2008; Torres et al., 2015; Işler et al., 2022). Additionally, pruning waste compost (PWC) has been associated with a significant increase in soil microbial activity, which plays a critical role in nutrient cycling and overall soil health. By enhancing microbial biomass and enzymatic processes, PWC fosters a more active and diverse soil microbiome, leading to improved nutrient availability for grapevines. This, in turn, can enhance root growth, water retention, and resistance to diseases, creating a more resilient agricultural system. Furthermore, studies have demonstrated that the use of PWC contributes to higher grapevine yields and improved grape quality, likely due to the steady release of nutrients and organic matter that promote balanced vine growth and optimal fruit development. The slow decomposition of organic matter in PWC ensures a continuous supply of nutrients, which can improve berry composition, including higher sugar content, better phenolic profiles, and improved organoleptic characteristics, making it a valuable input for high-quality wine production (García-Orenes et al., 2016; Gaiotti et al., 2017; Mondini et al., 2018). Economically, pruning waste compost (PWC) represents a renewable and cost-effective solution for sustainable vineyard management by turning an otherwise discarded by-product into a valuable resource. The reuse of pruning waste not only reduces the costs associated with waste disposal but also minimizes the need for synthetic fertilizers, thus lowering input costs for growers (Karimi et al., 2020).
Recent studies have further underscored the crucial role of beneficial fungi, such as arbuscular mycorrhizal fungi (AMF) and Trichoderma spp., in enhancing soil health and agricultural productivity. These fungi form symbiotic relationships with plant roots, facilitating greater nutrient uptake, particularly phosphorus, nitrogen, and micronutrients, and contributing to improved soil structure and plant resilience against biotic and abiotic stresses (Pascale et al., 2017; Balestrini et al., 2020; El Enshasy et al., 2020). AMF, for instance, create extensive hyphal networks that expand root surface area, enabling plants to access nutrients from a larger soil volume, which is particularly beneficial in nutrient-poor soils (Smith and Read, 2010).
The incorporation of beneficial fungi into compost, such as in the case of compost inoculated with AMF or Trichoderma spp., enhances the efficacy of compost as a biofertilizer. This fungal inoculation fosters beneficial microbial interactions, supports organic matter decomposition, and boosts soil organic carbon levels, thus improving overall soil fertility (Leifheit et al., 2014). Trichoderma harzianum, a well-known fungal species with biocontrol properties, not only suppresses pathogenic organisms but also induces systemic resistance in plants to various abiotic stresses such as drought, salinity, and temperature extremes, leading to improved plant growth, yield, and stress tolerance (Hao et al., 2012; Hidangmayum and Dwivedi, 2018). Furthermore, the application of T. harzianum has been shown to increase root biomass and enhance water use efficiency, which is particularly valuable in regions facing water scarcity (Hermosa et al., 2012).
Incorporating fungi into compost represents a promising strategy for improving nutrient cycling, promoting soil biodiversity, and enhancing the resilience of viticultural systems, aligning with the goals of sustainable and regenerative agriculture. This fungal-enriched compost can serve as a powerful tool to reduce the reliance on synthetic fertilizers, further contributing to soil health, carbon sequestration, and overall ecosystem balance.
Despite these promising findings, there is a notable gap in the literature regarding the combined use of fungal inoculation and pruning waste compost in soil for sustainable production in perennial crops. To address this gap, our study investigates the effects of uninoculated PWC, PWC inoculated with a fungal consortium, and standard mineral fertilizer on soil fertility parameters, microbial community structure, and soil enzyme activities. Specifically, we assess nutrient release kinetics for nitrogen (N), phosphorus (P), and potassium (K), as well as the microbial response to these treatments.
This research aims to provide new insights into the potential of pruning waste compost, particularly when combined with fungal inoculation, to enhance soil health and promote sustainable vineyard management.
2 Materials and methods
2.1 Experimental setup
The soil incubation experiment was conducted at the Free University of Bolzano within a climate-controlled chamber maintained at the following conditions: a 14-h light/10-h dark photoperiod, a temperature of 24°C during the day and 19°C at night, a light intensity of 250 μmol m–2 s–1, and 70% relative humidity.
To isolate the effects of pruned compost as a fertilizer and soil conditioner, an inert soil substitute was used as a control. The experimental soil substrate was a loamy-sandy fluvial deposit, the precise composition of which is detailed in Table 1. This material was obtained from a local river quarry (Fornaci Grigolin S.p.A., Nervesa della Battaglia, Italy; 45°48′56” N, 12°15′29″ E). The alluvial sand component, with an approximate grain size distribution of 0–4 mm, underwent a thorough washing and sorting process to ensure uniformity and remove impurities.
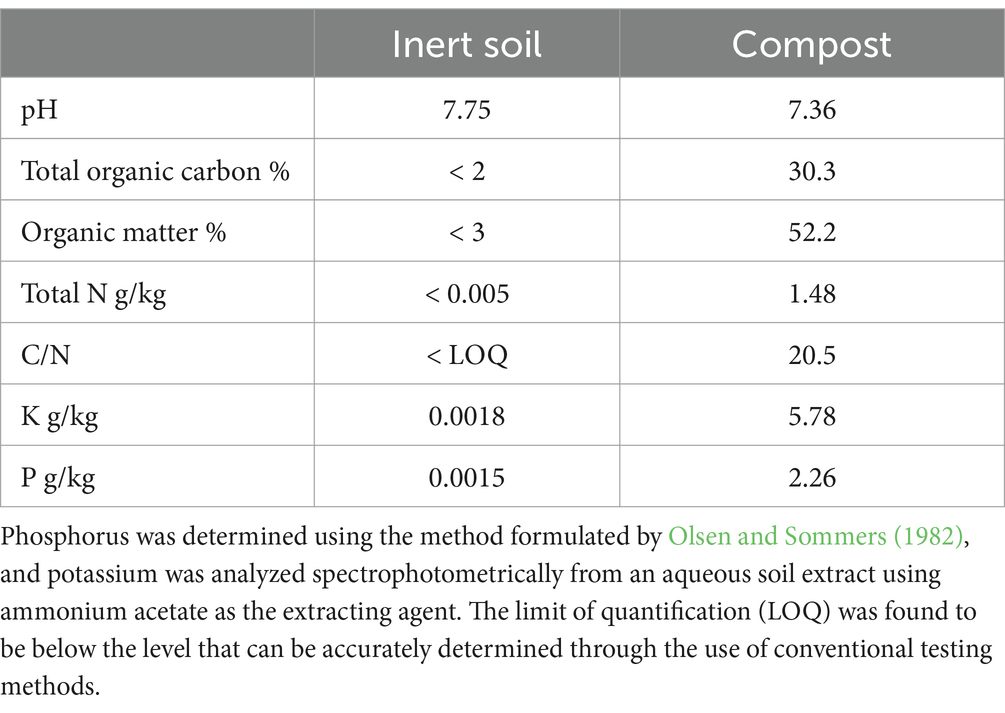
Table 1. Chemical characteristics of the compost and sandy soil utilized in the study, where total nitrogen and organic carbon were determined using the method of Dumas (1831), in accordance with the official chemical soil analysis protocol set by Ministerial Decree 248/99.
The compost was prepared from 100% pruning waste from a local vineyard. After pruning, the shoots were chopped and reduced to a size of 5–6 cm to increase the surface area/volume ratio. This bulk material was then placed in a heap and left under field conditions for about 10 months, with occasional turning to facilitate aeration. At the end of the period, the compost was collected, analysed by external laboratory (A.L.S. Life Science Italia, Zoppola, Italy) (results are shown in Table 1), parted in different boxes for the two treatments. The soil incubation experiment was carried out by setting up 15 pots (volume 5.5 L), five pots per treatment. Each treatment consisted in the application of either the mineral fertilizer (NPK), the pruning waste compost (PWC) or the inoculated pruning waste compost (iPWC). The iPWC was inoculated with a mixture of Rhizophagus irregularis and Funneliformis mosseae (80:20), at a concentration of approximately 300 propagules g−1, and Trichoderma harzianum at an approximate concentration of 1·109 cfu g−1. The fungal inoculants have been provided by Microgaia Biotech S. L. (Murcia, Spain). In order to have approximately the same supplementation of key nutrients with the different treatments, the nitrogen (N) concentration was equalized to standard supply in vineyards, considering 40 N units per hectare per year (Gatti et al., 2020). Based on N supplementation, defined amounts of phosphorus (P) and potassium (K) were added through the compost, and they were balanced in the mineral treatment accordingly. The treatments supplementations to pots were thus composed as follows:
• NPK: 3.5 kg sand +1.96 g Ca (NO₃)₂ + 0.56 g PK + 0.39 g K₂SO4.
• PWC: 3.5 kg sand +0.168 kg of Compost.
• IPWC: 3.5 kg sand +0.168 kg of Compost +0.067 g T. harzianum + 0.168 g R. irregularis and F. mosseae (80:20).
In order to simulate the growing season of a typical vineyard, soil moisture was maintained at 50% of the Water Holding Capacity and the experiment has been carried out for 135 days; soil samples have been collected every 15 days and stored for further analyses.
2.2 Community level physiological profiling
Five grams of soil samples were added to 45 mL of sterile 0.85% sterile NaCl physiological solution and shaken at 180 rpm for 1 h at room temperature. The resulting suspensions were serially diluted to 10−3 with sterile 0.85% NaCl solution. An aliquot (150 μL) of the resulting solution was added to the wells of Biolog Ecoplates® microplates. The microplates were incubated at 25°C for 7 days in the dark. The absorbance values were recorded every 24 h with a TECAN Infinite F200 plate reader at 590 nm (Ge et al., 2018). The metabolic activity developed by microorganisms in the carbon source wells results in formozan production, which develops a color change in the tetrazolium dye (Preston-Mafham et al., 2002). A greater change in the color intensity corresponds to a higher metabolic activity (Garland and Mills, 1991). The average well color development (AWCD) was determined as described by Zuluaga et al. (2021) using the equation AWCD = Σ Wi / n, where W is the adjusted absorbance from each well and n is the total number of wells. The functional diversity within the community was determined using Shannon’s diversity index (H), evenness (E), and substrate richness (R), as outlined by Ge et al. (2018). These calculations were derived from adjusted absorbance values obtained during the incubation phase of heightened metabolic activity, where a higher increase in the AWCD was observed. A threshold Wi value of 0.20 was assumed to indicate positive catabolism for each substrate.
2.3 Soil enzyme activity
Key enzymes involved in the P, carbon (C), sulfur (S), and N cycles, such as acid phosphatase, beta-glucosidase, arylsulfatase, protease, and urease, were analysed. Acid phosphatase activity was determined in 1.0 g of soil utilizing 50 mM p-nitrophenyl phosphate (pNPP) as a substrate (Tabatabai, 1994), with a modified universal buffer at pH 6.5. Spectrophotometry at 405 nm was employed for colorimetric analysis, and acid phosphatase activity was expressed as μg pNPP g−1 h−1. β-Glucosidase activity in soil was assessed by incubating 1.0 g of soil with PNG (p-nitrophenyl-β-dglucoside) at 50 mM, in a modified universal buffer at pH 6, as described by Tabatabai (1994). Absorbance was measured at 405 nm to quantify the β-glucosidase activity expressed μg PNG g−1 h−1. For arylsulfatase, 1.0 g of soil was mixed with a solution containing 50 mM p-nitrophenyl sulfate (pNS) in a modified universal buffer at pH 6. Absorbance of the reaction solution was measured at 420 nm and arilsulfatase activity was expressed as μg PNP g−1 h−1 (Tabatabai, 1994). Protease activity in soil was determined by incubating 1.0 g of soil with 2 mL of 1% casein at 50°C for 2 h. Thereafter, 1 mL of Lowry’s solution and 100 μL of 1 N Folin’s phenol reagent were added, and absorbance was measured at 750 nm. Proteolytic activity was expressed as micrograms of BSA equivalent per gram of dry soil. Urease activity was measured following the method of Kandeler and Gerber (1988). One gram of soil was incubated with 500 μL of 0.72 M urea at 37°C for 2 h. Subsequently, 6 mL of 1 M KCl was added, and the supernatant was collected. Addition of 1 mL of sodium salicylate and 400 μL of 0.1% sodium dichloroisocyanurate was followed by a second incubation of 30 min at room temperature in the dark. Absorbance was read at 690 nm, and urea activity was expressed as micrograms of NH4 per gram of soil per hour μg NH4 g−1 h−1.
2.4 Soil nutrient dynamic
To determine the dynamics of release of total N, available P, and exchangeable K, approximately 20 g of soil were sampled every 30 days throughout the whole experimental period. The samples were then analysed by a certified external laboratory through standard procedures (ALS Italia, Pordenone, Italy).
2.5 Statistical analysis
Data were subjected to statistical analysis using XLSTAT software version 2022.4.1 (Addinsoft, New York, USA). Differences between treatments were assessed using one-way Analysis of Variance, Tukey’s test was used to test the differences within the means, (p < 0.05). Principal Component Analysis (PCA) was applied using the 13 different soil measurements of the trial with Pearson correlation analysis. JASP version 0.18.1.0 for Windows (JASP Team, Amsterdam, Netherlands, 2023) and SigmaPlot version 14.0 for Windows (SigmaPlot 14.0, Systat Software, Inc., San Jose, California, 2023) were used for graph editing. R software (version 4.3.1) was also used to graphically represent data, by exploiting the following packages: agricolae, ggplot2.
3 Results
3.1 Community level physiological profiling
Substrate utilization was calculated by measuring the average wells color development (AWCD) values after 96 h of incubation. This specific time was selected since the substrate conversion rate of tetrazolium dye reached stable values for all indexes. Expecting significant differences in a short time frame, the first two points were sampled at a 15-day interval to assess the response of the microbial community to the change in environmental conditions.
The estimation of the metabolic activity of the microbial community highlighted significant differences among the treatments (i.e., NPK, PWC and iPWC) applied to soils (Figure 1). The NPK soil samples showed basal metabolic activities that significantly changed only at 15 days after the application (p < 0,001) as compared to the initial values. Afterwards, the AWCD for NPK samples progressively decreased, reaching values close to zero (Figure 1). On the other hand, PWC and iPWC-amended soils showed a remarkably different behavior when compared to NPK samples (Figure 1). Indeed, 15 days after the application of treatments, both PWC and iPWC soil samples showed a steep increase in the AWCD reaching approximately 10-fold higher levels as compared to NPK. This initial peak of metabolic activities was afterward followed by a gradual decrease. At the end of the experimental period, the metabolic activities of the microbial communities in the three soil samples were comparable and independent from the treatment applied. Interestingly, except for the last sampling point, the activity detected in the iPWC samples was always significantly higher with respect to PWC samples (Figure 1).
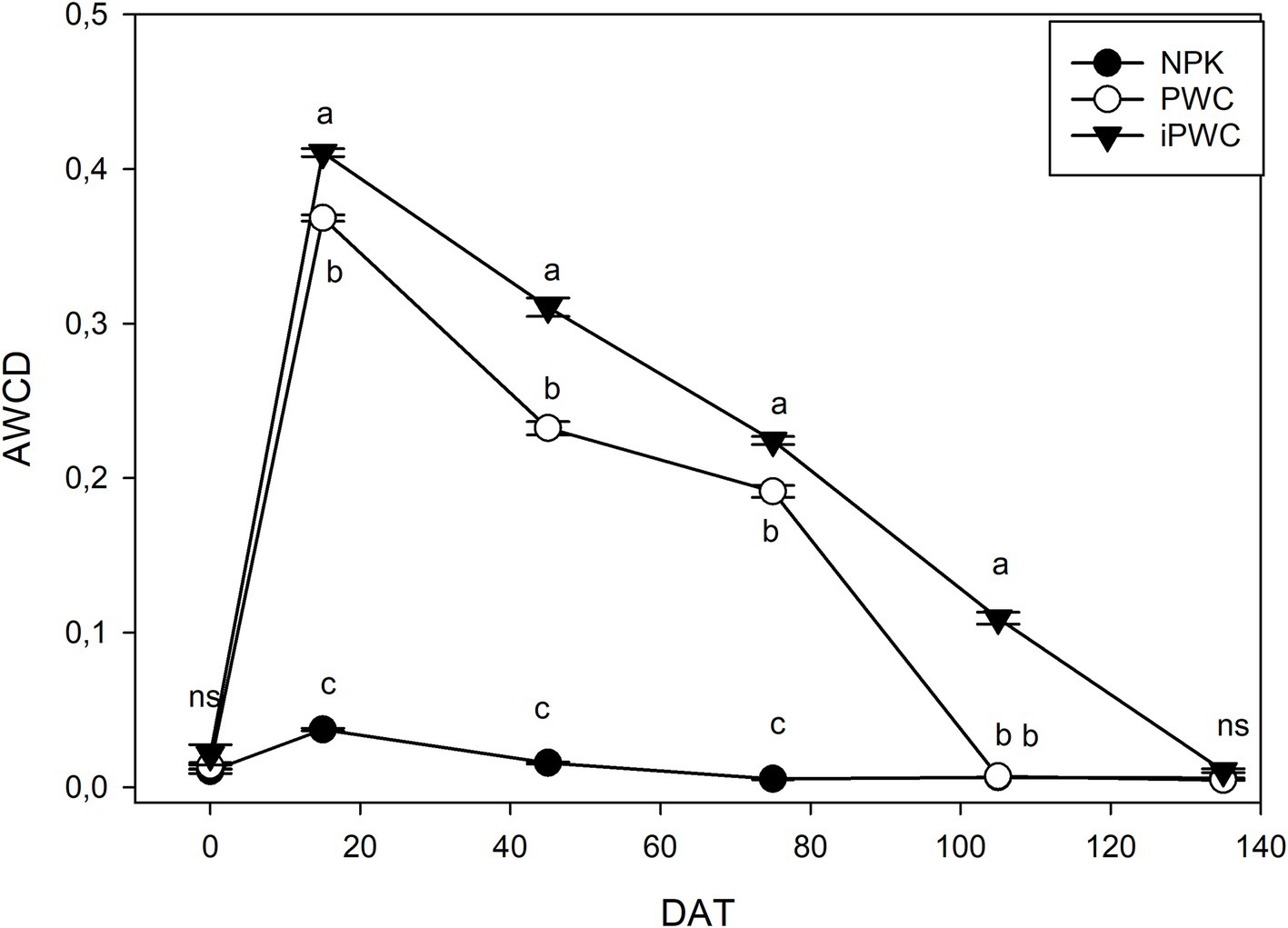
Figure 1. Average wells color development (AWCD) values. The average well color development (AWCD) was determined in soil samples subjected to three different treatments (NPK, PWC, and iPWC) at various days after treatment application (DAT). Data are presented as mean values ± SE (n = 5). Different letters indicate statistical differences as determined by one-way Analysis of Variance with Tukey Post-Hoc test (p < 0.05) (see Supplementary Table S2). Note that these data correspond to the 96-h time point from the CLPP analysis.
To gain further insight into the metabolic characteristics of the microbial communities associated with the different treatments, AWCD values related to classes of compounds were determined. Based on the overall AWCD, metabolic activity remained active for at least 75 days following fertilizer treatment. Specifically, at 15 days after application, the iPWC-fertilized soil exhibited significantly higher metabolic activity, primarily associated with polymer consumption, as compared with PWC (p < 0.01) and NPK (p < 0.001) (Figure 2). Notably, the specific polymers involved were polysorbate 40/80, glycogen, and α-cyclodextrin. Furthermore, both PWC and iPWC treatments showed a significantly higher degradation rates for amino acids (e.g., L-alanine, L-phenylalanine, L-serine, L-asparagine, L-pyroglutamic acid, L-threonine, and L-proline) and amines (e.g., putrescine and phenylethylamine) compared to NPK for at least 75 days after treatment (Figure 2 and Supplementary Table S3). The utilization rates of carboxylic acids were higher in compost treatments, albeit the inoculated samples showed significantly higher values during the experimental period. Conversely, the consumption rates of phenolic compounds and carbohydrates (D-xylose, N-acetylglucosamine) were comparatively slower in PWC and iPWC soil samples related to the above-mentioned substrates, albeit significantly higher than those observed in NPK-treated soil samples (p < 0.05).
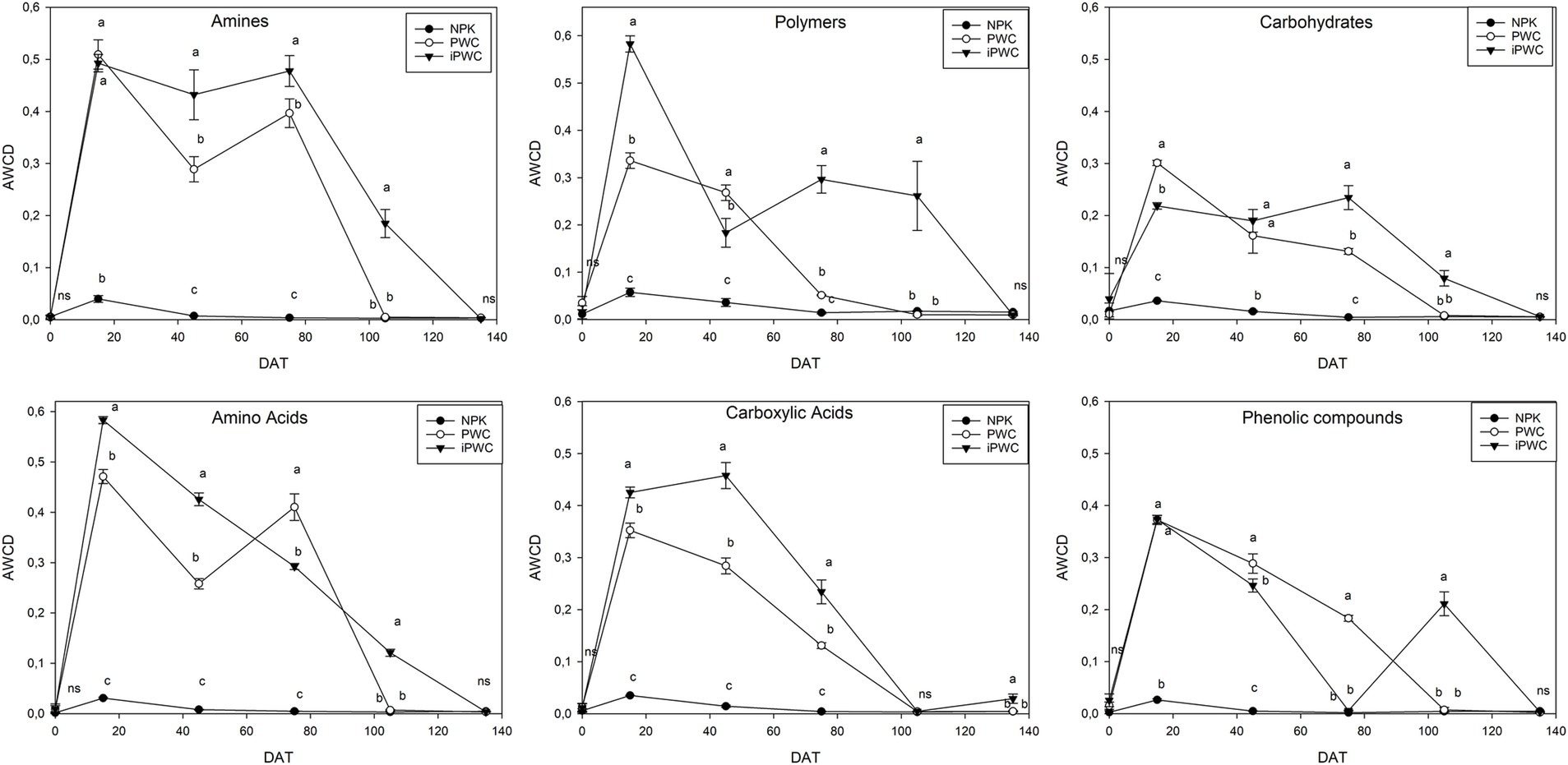
Figure 2. Average wells color development (AWCD) values for separated carbon sources. Amines, polymers, carbohydrates, amino acids, carboxylic acids, and phenolic compounds were quantified at 595 nm in soils subjected to three different treatments: mineral fertilizer (NPK), pruning waste compost (PWC), and inoculated pruning waste compost (iPWC). These values represent the average well color development, indicating the activity of the microbial community in response to the substrates of the organic compounds. A one-way analysis of variance (ANOVA) with Tukey’s post hoc test (p < 0.05) was conducted to compare the effects of the three treatments on the response variables. Different letters indicate statistically significant differences based on the ANOVA results. Five replicates were used for each treatment. The bars for each symbol represent the standard deviation (SD). DAT stands for the days after the application of treatments.
Metabolic functional diversity of microbial communities associated to the different fertilization treatments has been estimated by using diversity indices (Table 2 and Supplementary Table S1), as for instance Richness (R), Shannon diversity index (H′), Shannon evenness index (E), throughout the whole experimental period.
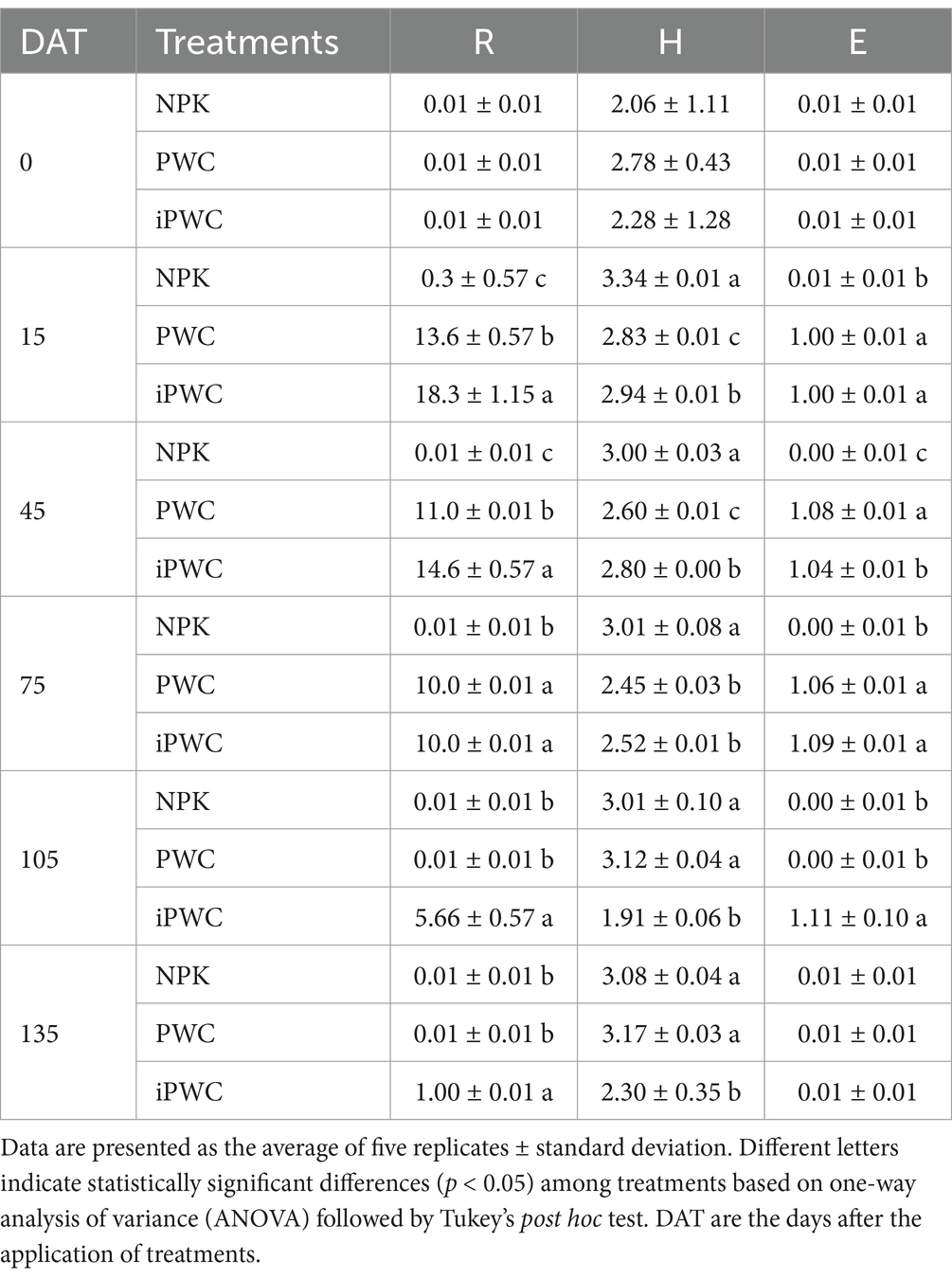
Table 2. The microbial community indices Richness (R), Shannon functional diversity (H), and Shannon evenness (E) were measured at seven time points (0 days, 15 days, 45 days, 75 days, 105 days, and 135 days) to assess the temporal changes in the microbial community in the PWC, iPWC, and NPK treatments.
Concerning the richness (R) parameter, expressed as the number of substrates utilized by the microbial community, no significant differences were observed at the beginning of the experiment (Table 2). In the following time points, R values of NPK-treated soil samples remained almost constant and close to zero. Nevertheless, R parameter calculated for PWC and iPWC-supplemented soil samples showed significantly higher values compared to NPK samples, starting from 15 days after the treatments (Table 2). Interestingly, at 105 days after treatments, R parameter of PWC-treated soil samples decreased back to basal values, whereas that of iPWC samples was maintained significantly higher until the end of the experiment (Table 2). Shannon functional diversity values (H) was significantly higher in NPK treatment as compared to PWC and iPWC, except for the initial sampling point. Despite the notable metabolic activity observed in both compost treatments in contrast to mineral fertilizer application, a discernible linear trend in functional diversity was absent. This observation underscores the complexity of the relationship between metabolic activity (measured by AWCD) and Shannon diversity index (H) within this experimental context. With regard to the evenness of the microbial communities, significant differences were observed at 45 and 75 days after treatment, showing the highest impact on compost-treated (PWC and iPWC) samples, independently from the inoculation with AMF. These results suggest that the mineral treatment supported a less evenly distributed community of microbial species compared to the compost treatments.
3.2 Soil enzyme activity
Soil enzymes play a crucial role in various biogeochemical processes, including nutrient cycling, organic matter decomposition, and plant-microbe interactions. Their activities can provide valuable insights into the overall health and functioning of soil ecosystems.
Enzyme activity was found to positively respond to both PWC and iPWC treatments (Figure 3) Specifically, PWC and iPWC fertilizers significantly stimulated arylsulfatase (Ars) activity throughout the whole experimental period with respect to NPK (p < 0.05), whose activity remained low and stable. On the other hand, the Ars activity in PWC-treated samples increased linearly (Figure 3). Differently, in iPWC-amended soil samples, Ars activity initially showed a moderate increase, reaching a peak at 40 days characterized by significantly higher values as compared to both NPK and PWC (p < 0.05). After a decrease at day 105, Ars activity in iPWC-amended soil samples recovered but remained lower than those determined in PWC-supplemented samples (Figure 3). Concerning the β-glucosidase (β-Glu) activity, steady trend was observed in the three treatments. The only exception was represented by the peak of activity detected at 60 days after application of NPK fertilizer (Figure 3). Overall, the stimulation of β-Glu activity was significantly higher in the samples amended with compost with respect to those supplemented with NPK fertilizers (Figure 2 and Supplementary Table S4), thereby confirming the low effect of this latter on the C-cycle related enzymes. All treatment applications positively stimulated Acid Phosphatase (AcP) activity, exhibiting a continual increase throughout the experiment. While analysis of variance confirmed significantly higher activity for iPWC compared to both PWC and NPK (p < 0.05), it’s noteworthy that NPK displayed the highest percentage increase in AcP activity (108%), followed by PWC (92%) and iPWC (79%). All three treatments demonstrated similar temporal trends in Protease activity (Ps), exhibiting a consistent increase throughout the experimental period. Notably, NPK consistently displayed the lowest activity values throughout the experiment (p < 0.05). In contrast, both PWC and iPWC treatments showcased significantly higher activity levels (p < 0.05). Interestingly, despite lowest absolute activity, NPK exhibited the greatest percentage increase from baseline (+485%), followed by PWC (+397%) and iPWC (+298%). Urease (Ur) activity in NPK and PWC treatments remained statistically unchanged (p > 0.05) throughout the initial 60 days after the treatments, exhibiting similar dynamics. Nonetheless, PWC-amended soil samples subsequently showed significant peak in activity at a later stage (p < 0.05), followed by a decrease. Notably, iPWC treatment demonstrated a distinctly different pattern, featuring a significantly higher and consistently rising activity rate (p < 0.05). Despite being it initially low, the Ur activity in iPWC-treated soil samples showed a 700% increase during the last 75 days of treatment (Figure 3).
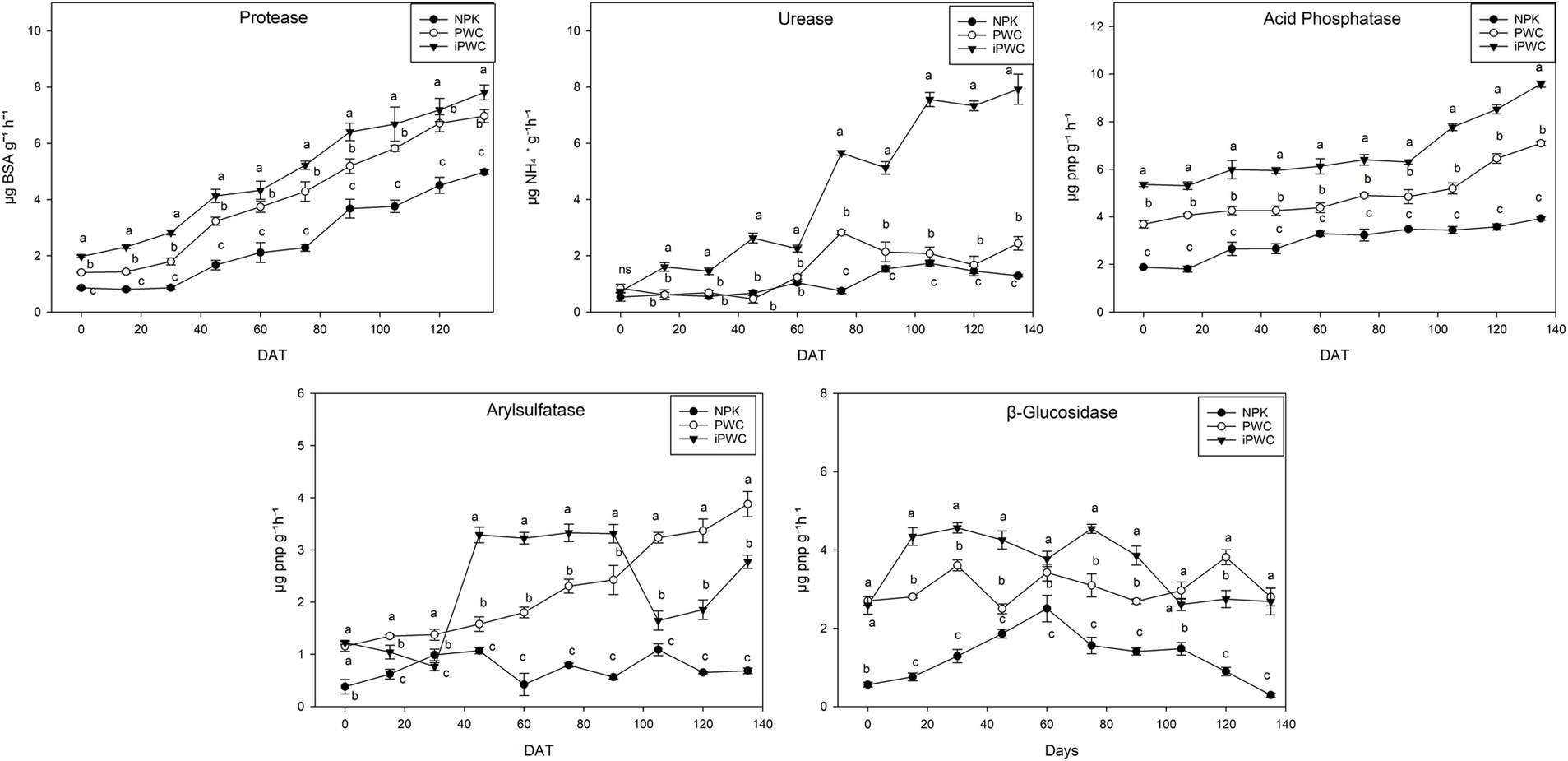
Figure 3. Soil enzymes activity measured over the trial. Protease (Ps), Urease (Ur), Acid Phosphatase (AcP), Arylsulfatase (Ars), and β-glucosidase (β-Glu) activities were measured. These values represent the average well color development, indicating the activity of the microbial community in response to the substrates of the organic compounds. A one-way analysis of variance (ANOVA) with Tukey’s post hoc test (p < 0.05) was conducted to compare the effects of three treatments: mineral fertilizer (NPK), pruning waste compost (PWC), and inoculated pruning waste compost (iPWC) on the response variables. Different letters indicate statistically significant differences based on the ANOVA results. Five replicates were used for each treatment. The bars for each symbol represent the standard deviation (SD). Data are presented as the arithmetic mean ± standard error. DAT means days after treatment application.
3.3 Soil nutrient dynamic
In Figure 4, the results of macronutrients analyses are shown. Total N concentration in the soil samples was not affected by both the treatments and the incubation time, except for the final sampling. At this point, both iPWC and PWC-amended soil samples exhibited significantly higher levels of total N as compared to NPK (p < 0.05). However, despite the lack of significant differences along the period monitored, the different treatments showed a different kinetic of N release. In fact, NPK treatment showed a rapid initial increase, peaking at 30 days before experiencing a substantial decline of approximately 100% at 60 days. Conversely, PWC and iPWC treatment demonstrated sustained increases of total N concentration throughout the trial, with final values 275 and 114% higher than their initial levels, respectively.
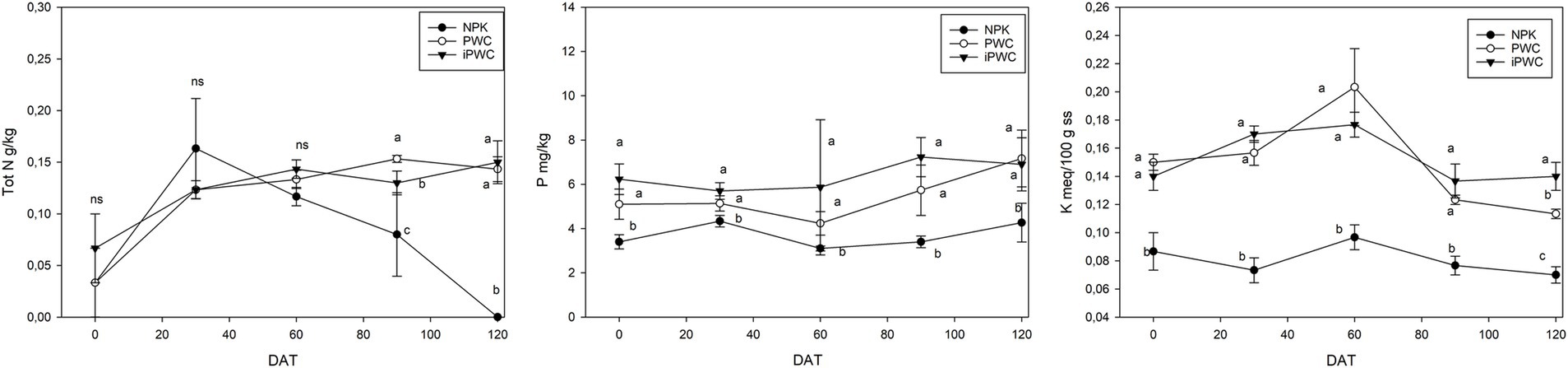
Figure 4. Soil nutrient concentrations. From left to right, total Nitrogen (Tot N), Olsen available Phosphorus (P), and exchangeable Potassium (K) were measured in the three different treatments—mineral fertilizer (NPK), pruning waste compost (PWC), and inoculated pruning waste compost (iPWC)—over the 135 days of experiment. Values represent means ± standard error (SE) with five replicates (N = 5). Different letters indicate statistically significant differences (p < 0.05) based on one-way analysis of variance (ANOVA) followed by Tukey’s HSD post-hoc test.
Phosphorus (Olsen-P) availability was generally low, as expected based on the chosen soil type. Whilst the statistical analysis did not reveal significant differences between treatments, the collected data indicated potentially higher average p values associated with compost amendments, particularly iPWC. Overall, P availability remained relatively stable across the treatments, highlighting the potential benefits of iPWC for maintaining slightly higher P levels in this soil type.
Levels of exchangeable K were measured at 100 meq per 100 g of dry matter soil. Although these values may not provide conclusive results about the impact on soil fertility, they present a promising perspective for plant nutrition management, especially considering the specific soil type and applied doses. Interestingly, Analysis of Variance revealed a significant difference from the beginning to the end of the experiment between compost treatments and NPK at equal doses (p < 0.05). Notably, exchangeable K was consistently higher in soils amended with compost, independently of the inoculation with AMF. Further analysis identified a trend where K availability peaked for all treatments at 60 days, followed by a subsequent decline that was more pronounced in PWC and iPWC compared to NPK.
3.4 Principal component analysis on soil measurements
For a more detailed understanding of the dataset, a Principal Component Analysis (PCA) was performed to explore latent relationships among the various variables (Figure 5). The variables considered include those pertaining to the microbial community activity, the substrate consumption, enzymatic activity, and macronutrients under examination. We correlated these variables with sampling times to simultaneously consider time and treatment factors. The PCA results were interpreted with various significance tests.
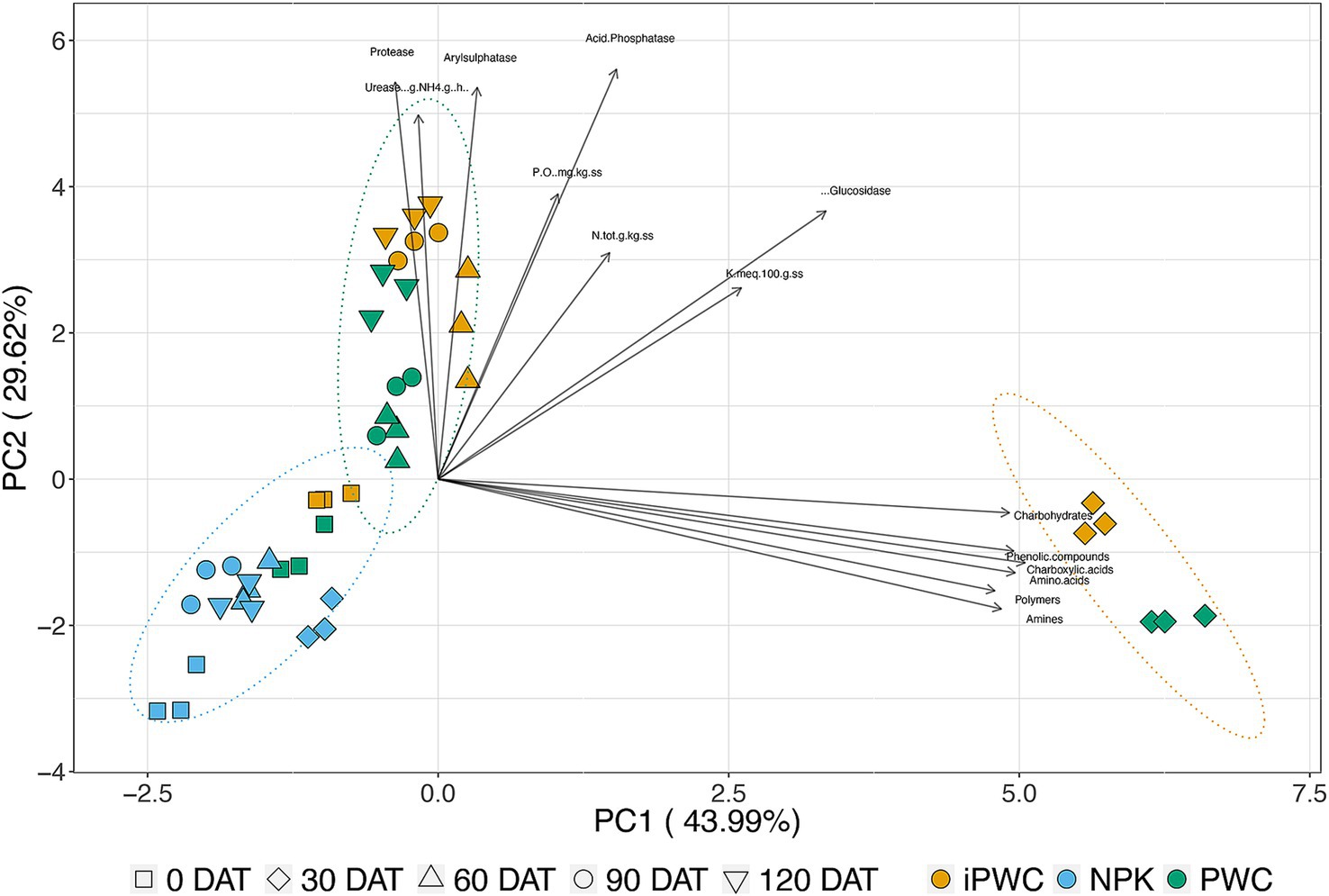
Figure 5. Principal component analysis. PCA was conducted on the substrate utilization rate (amines, amino acids, carbohydrates, carboxylic acids, phenolic compounds, and polymers), enzyme activity (urease, protease, arylsulfatase, acid phosphatase, and beta-glucosidase), and nutrient concentrations (total nitrogen, phosphorus, and potassium) in soil treated with specific fertilizers. Different colors represent different treatments. Symbols denote the sampling days. Ellipses indicate 95% confidence intervals.
The Bartlett sphericity test revealed that at least one of the correlations between variables significantly deviates from zero, indicating meaningful relationships among the variables. Additionally, the correlation matrix values demonstrate notable levels of association among the variables, further supporting the robustness of the relationships observed. While the Kaiser-Meyer-Olkin (KMO) sampling adequacy test confirmed the sample’s suitability for analysis (KMO = 0.786). The first two factors collectively explained 73.61% of the variance, with PC1 primarily reflecting substrate metabolic activity and PC2 being more influenced by enzymatic activities. Macronutrients had a lower weight within the PCA. The visual interpretation of PCA was facilitated through a Biplot in Figure 5, which illustrates both variables and observations in the same space. Interestingly, observations for all treatments at the first sampling (0 DAT) are clustred together in the negative quadrant of the scaterplot. In the following sampling times, a variable-dependent evolution concerning PWC and iPWC has been observed. Indeed, at 30 days after treatment, a clustering primarily driven by metabolic substrate consumption was obtained. However, at 60, 90, and 120 DAT, the variables more prominently influencing observations are the enzymes Urease, Arylsulfatase, and Protease. This evolution is further elucidated by examining the correlation between variables and the principal component axes, as defined by the cosine squared values. Notably, on axis 1, a stronger contribution is observed from the values associated with substrate consumption by microorganisms and betaglucosidase. Conversely, axis 2 demonstrates a greater contribution from other enzymes, namely Urease, Protease, Arylsulfatase, and Acid Phosphatase. Finally, the macronutrients—total nitrogen, potassium, and phosphorus—demonstrate a more significant contribution to Factors 4 (F4), 3 (F3), and 5 (F5), respectively, as identified in our principal component analysis (PCA). These factors represent the underlying patterns extracted from the data, with each factor corresponding to a specific combination of variables that account for the variance observed in the dataset.
4 Discussion
Soil degradation and loss of biodiversity in agricultural systems are primarily the result of decades of intensive cropping (i.e., mineral fertilizers and phytosanitary products), to achieve higher productivity levels (Scotti et al., 2015; Pahalvi et al., 2021). Nevertheless, the protection and preservation of soil quality and fertility parameters are a key point for sustainable development (Lal et al., 2021). In this context, different agricultural sectors, viticulture among them, have identified organic amendments as promising tools to ameliorate soil structure, increase nutrient availability and strengthen grapevine resistance to pests and diseases (Diacono and Montemurro, 2011; Mondini et al., 2018; Lazcano et al., 2020; Romero et al., 2022). In the present research, the role of pruning waste compost, either non inoculated or inoculated with AMF, in promoting soil quality has been investigated, thereby considering different fertility parameters.
The assessment of the functional diversity of complex microbial communities, such as those of soil, obtained by monitoring the catabolic activity of microbes, represents an interesting approach to unravel ecosystems functioning according with alterations in the structure and diversity of their microbiomes (Ros et al., 2008; Si et al., 2018). In our experimental model, these determinations reflected the response of microbial communities to changes in management (Lundquist et al., 1999; Aislabie and Deslippe, 2013). The results of our study demonstrate that the application of compost to soil significantly increases the metabolic activity of soil microbial community. This is evident from the significant differences registered in AWCD values among the different treatments. PWC and iPWC-amended soil samples displayed an increasing trend in the metabolic activities, which reached the highest values at 15 days after treatments, thereby showing a gradual decline afterwards. On the contrary, the NPK-treated soil samples displayed a constant, basal metabolic activity after the treatment application. Indeed, the substrate used for the experiment was chosen to be very poor in nutrients, featuring in fact low concentrations of total organic carbon and organic matter. As also observed by other authors, the application of organic amendments, like compost in the present research, has been demonstrated to provide a supplement that is much richer in readily available sources of nutrients to boost the metabolic activities of the microbial community (Li et al., 2018), supporting the observation of enhanced AWCD values in PWC and iPWC-amended soil samples.
On the other hand, consistent with findings from other studies (Okur et al., 2009; Zhong et al., 2010), soil samples treated with mineral fertilizers exhibited lower values of Average Well Color Development (AWCD), likely due to the limited provision of organic matter by these treatments. This suggests that microbial communities in mineral-fertilized soils may experience carbon limitation, as these treatments do not provide sufficient organic substrates required for microbial metabolism. In contrast, the higher AWCD values observed in PWC and iPWC treatments indicate that the organic matter supplied by compost stimulates microbial metabolic activity. These results suggest that the lack of carbon inputs in mineral fertilizer treatments restricts microbial growth and activity, whereas compost treatments provide the necessary organic carbon to support a more dynamic microbial community. This apparent carbon limitation in mineral-fertilized soils aligns with the overall reduced microbial function observed in these samples, further highlighting the importance of organic matter in maintaining soil microbial health.
To date, few studies have investigated the differences in C metabolism for bacteria belonging to different ecological groups (Wang et al., 2022). Indeed, the ability of microbes to use specific substrates as C source can influence their growth and their ecological competence (Goldfarb et al., 2011; Saifuddin et al., 2019; Muscarella et al., 2020). On the other side, also the ability of microbes to release specific C metabolistes could be of paramount importance in the mediation of inter-species communications within microbial communities (Zengler and Zaramela, 2018; Niehaus et al., 2019). The analysis of the specific substrate utilization reproduces the general trend observed in the AWCD values. In particular, amines and amino acids resulted the most used substrate by the microbial communities of PWC and iPWC-supplemented soil samples, whereas carbohydrates showed a lower consumption rate. Nevertheless, albeit reaching slightly lower values, the consumption rate of carboxylic acids showed a profile which followed those of amines and amino acids, suggesting that also this class of compounds might sustain the metabolic activity of soil microbes. This has also been observed in previous studies in which the application of organic amendments favoured a microbial community that preferentially degrades nitrogenous organic compounds rather than carbohydrates (Carrera et al., 2007; Ros et al., 2008; Bernard et al., 2012). In particular, species belonging to the phylum of Proteobacteria have been shown to be primarily involved in the mineralization of N-bearing organic substrates (Aguilar-Paredes et al., 2023). Other experimental evidence demonstrated that carboxylic acids and amino acids are the most used sources when the microbial community of a soil is subjected to an abiotic stress (Jin et al., 2015). Nevertheless, these metabolic preferences have also be shown to be variable depending on different factors, as for instance soil chemical and physical characteristics, as well as environmental conditions (Zheng et al., 2004). As described by Sandle (2016), our results indicate that certain microbes in our experiment exhibit distinct substrate preferences for energy production. Specifically, we observed a notable increase in the utilization of amino acids over carbohydrates, suggesting a preference for amino acid catabolism as an energy source.
The microbial community functional diversity indices revealed intriguing patterns during samplings at 15-, 45-, and 75-days post-fertilizer application. This suggests that soil microorganisms with a greater capacity for substrate degradation were notably more metabolically active during the initial 15 to 60 days following the commencement of the experiment. Interestingly, the Shannon diversity (H) exhibited significantly higher values in the mineral treatment compared to compost application. Similar results were found in previous studies (Jezierska-Tys et al., 2020; Liu et al., 2022). This counterintuitive finding may be explained by the fact that mineral fertilizers like NPK supply primarily inorganic nutrients without substantial organic matter. As a result, microbial communities may be more evenly distributed due to the limited availability of organic carbon, with a greater number of species coexisting and competing for these limited resources. In contrast, compost treatments provide a rich source of organic matter, which could lead to the rapid growth of specific microbial populations that dominate the microbial community, thereby reducing evenness and overall functional diversity. Furthermore, mineral fertilizers may induce a more heterogeneous environment, where opportunistic species that specialize in stress-tolerant conditions thrive, further contributing to higher functional diversity in these soils. However, these findings stand in contrast to the prevailing trend observed in other studies, which consistently report greater microbial diversity in soils from organic farming systems compared to conventional ones (Raupp et al., 2004; Zhong et al., 2010; Sradnick et al., 2013). This could be explained by the compost selecting for microbial communities that are more specialized in decomposing organic matter, thus leading to a decrease in the range of available carbon sources and less competition among microbial species.
As highlighted by Qiao et al. (2022), soil quality is closely related to its capacity to sustain agricultural productivity. A key method for assessing soil quality involves measuring the activity of soil hydrolytic enzymes (Gil-Sotres et al., 2005; Das and Varma, 2011). In nutrient-poor soils, the application of mineral fertilizers resulted in lower enzymatic activity, consistent with previous findings (Li et al., 2008; Iovieno et al., 2009; Zhang et al., 2015). Conversely, compost application, whether inoculated or not, significantly enhanced enzymatic activities. High levels of β-glucosidase (βGlu) and arylsulfatase (ArS) activity were particularly associated with compost amendments (Adetunji et al., 2017; Tang et al., 2020).
Nitrogen mineralization plays a critical role in soil fertility, correlating directly with plant development through the supply of essential nitrogen. Proteases, which cleave peptide bonds to generate free amino acids, serve as key indicators of soil quality (Ali et al., 2019). Our findings show that compost applications, regardless of inoculation, improved protease activity, corroborating earlier studies that linked organic amendments to elevated protease levels (Chang et al., 2007; Saha et al., 2008).
A significant increase in urease and acid phosphatase activities was observed with iPWC compared to PWC, suggesting that the inoculation with Trichoderma harzianum, Funneliformis mosseae, and Rhizophagus irregularis had an impact. While arbuscular mycorrhizal fungi (AMF) typically form symbiotic relationships with plant hosts (Veresoglou et al., 2012; Berruti et al., 2016), studies indicate that arbuscular mycorrhizal fungi (AMF) can also engage with bacterial communities (Hildebrandt et al., 2006; Hempel et al., 2007), which might explain their metabolic activity in our plant-free system. Nevertheless, there remains a lack of direct evidence to confirm AMF presence or activity in this study. It is possible that non-mycorrhizal fungi, like Trichoderma, drove the observed increase in enzyme activity (Ali et al., 2011; Souza et al., 2016).
Urease, which catalyzes urea hydrolysis, showed higher activity in iPWC-treated soils, possibly due to AMF’s preference for NH₄+ as a nitrogen source (Tanaka and Yano, 2005; Hodge et al., 2010). Despite the inoculation, previous pieces of research have also shown that compost application alone enhances urease activity (Garcı́a et al., 1994; Pascual et al., 1999; Chakrabarti et al., 2000; Chang et al., 2007), a trend corroborated by the presented data.
Acid phosphatase, an enzyme linked to phosphorus (P) solubilization, showed increased activity in inoculated treatments, potentially due to the role of AMF in enhancing P availability (Joner and Johansen, 2000; Wu et al., 2021). Similarly, Trichoderma harzianum has been associated with increased phosphatase activity and organic matter decomposition (Szczałba et al., 2019). While these enzymes are important for nutrient cycling, it remains speculative to attribute these increases solely to AMF activity, given the absence of a plant host and direct measurements of AMF persistence in the system.
In terms of nutrient dynamics, nitrogen availability was consistent across all treatments at the start of the experiment. However, NPK-treated soils showed a peak in nitrogen content at 30 days, which rapidly declined, likely due to nitrogen leaching (Quemada et al., 2013; Zhao et al., 2022). In contrast, compost-treated soils maintained a steady nitrogen release over time, suggesting a slower mineralization process that reduced the risk of leaching (Wei et al., 2021). These results underscore the advantage of organic amendments in providing a more sustained nitrogen supply compared to mineral fertilizers.
Phosphorus availability did not vary significantly between treatments, although compost-treated soils showed a trend toward higher bioavailability, in line with previous findings (Francioli et al., 2016). The lack of a marked response in P availability contrasts with other studies (Calleja-Cervantes et al., 2015; Maltais-Landry et al., 2015; Boutchich et al., 2018), possibly due to the shorter duration of this experiment or the specific compost composition used.
Potassium (K), essential for plant water management (Hue and Silva, 2000; Pandey and Mahiwal, 2020), remained stable throughout the study, though compost treatments exhibited significantly higher exchangeable K values than mineral fertilisers treated soils. This is consistent with research showing that organic amendments increase K availability (Najafi-Ghiri et al., 2018), this is likely due to the higher potassium content in the compost (see Table 1). The slow-release nature of the compost is thought to have contributed to the maintenance of potassium levels over time.
While enzyme activities increased with compost application, the contrasting results from community-level physiological profiling (CLPP) and enzymatic assays suggest different substrate utilization patterns. Compost-treated soils showed higher enzyme activities, indicating increased nutrient cycling, but lower average well-color development (AWCD) values, reflecting lower microbial functional diversity. This discrepancy might be explained by the specific substrate range targeted by enzymes versus the broader spectrum of carbon sources available in the CLPP analysis (Ros et al., 2008). Additionally, the Ecoplates technique, being culture-dependent, may underestimate the complexity of soil microbial communities (Nannipieri et al., 2002).
The inoculation with fungi significantly enhanced enzyme activity, but its impact on nutrient availability was less clear. While inoculated compost improved microbial activity, nutrient release patterns were not markedly different from compost alone. This raises questions about the direct contribution of fungal inoculation to nutrient mineralization, which remains an area for further research (Amora-Lazcano et al., 1998; Cavagnaro et al., 2007; Veresoglou et al., 2012).
The principal component analysis (PCA) further highlighted the distinct impacts of the treatments on soil characteristics. NPK-treated soils showed minimal change over time, while compost-treated soils, especially those with fungal inoculation, demonstrated significant shifts in microbial and enzymatic activity. This supports the hypothesis that compost, particularly when inoculated, has a more pronounced effect on soil biological indices and nutrient cycling, consistent with the gradual mineralization processes described by Diacono and Montemurro (2011).
Overall, the results of our study demonstrated that the addition of a fungal consortium to pruning waste compost enhanced enzymatic activities and microbial functionality compared to non-inoculated compost and mineral fertilizer. However, some limitations should be acknowledged. The study was conducted in a controlled growth chamber, which does not fully account for environmental factors that influence soil dynamics in field conditions. Additionally, the absence of plant–soil interactions limits our understanding of the direct effects of compost and microbial inoculants on plant performance and nutrient uptake. Variations in compost composition and microbial inoculation efficiency could also contribute to differences in nutrient availability and microbial responses. Moreover, discrepancies between CLPP and enzyme activity data highlight the need for more comprehensive methods to assess microbial function.
5 Conclusion
Grapevine pruning compost represents a critical advancement in the application of circular economy principles within sustainable agriculture. Transforming vineyard waste into a valuable organic amendment aligns with the European Green Deal’s objectives, particularly regarding the promotion of resource efficiency, biodiversity, and climate resilience. The findings of this study underscore the beneficial effects of pruning waste compost on key soil quality parameters. Enhanced microbial activity, coupled with increased enzymatic functions, has been shown to improve nutrient cycling, particularly with regard to sustained nitrogen availability, stable phosphorus levels, and elevated exchangeable potassium. These improvements contribute to soil fertility and structure, fostering a healthier soil ecosystem.
Despite these promising results, further research is essential to understand how these enhancements in soil quality translate into plant growth and productivity under real-world conditions. Future studies should prioritize field or pot experiments with grapevine plants to assess the broader agronomic impacts. Additionally, understanding the compost’s performance under extreme environmental conditions—such as droughts or heatwaves—will be crucial in determining its suitability in the context of climate change.
Moreover, the role of fungal inoculation in biogeochemical nutrient cycles requires further investigation, particularly in understanding how the selected fungi enhance nutrient solubilization and availability. Given the microbial interactions observed in this study, additional research is needed to refine its application. By demonstrating the effectiveness of pruning waste compost and fungal inoculation in improving soil microbial health and nutrient cycling, this research offers valuable insights for sustainable food systems. The findings highlight the potential of organic amendments to reduce environmental impact, maintain soil fertility, and support regenerative agricultural practices. In addition, optimizing composting techniques to maximize nutrient content and minimize potential contamination will ensure high-quality organic inputs. Moreover, implementing long-term soil monitoring programs will allow growers to track improvements in soil biodiversity, carbon sequestration, and nutrient availability, supporting more sustainable vineyard management. Overall, pruning waste compost presents a promising strategy for sustainable viticulture and soil management. While some uncertainties remain, its integration into agricultural systems could significantly improve soil health, decrease dependence on synthetic fertilizers, and contribute to long-term sustainability in viticulture.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding authors.
Author contributions
ML: Conceptualization, Data curation, Formal analysis, Investigation, Visualization, Writing – original draft, Writing – review & editing, Methodology, Validation. MZ: Data curation, Formal analysis, Investigation, Methodology, Supervision, Validation, Visualization, Writing – review & editing. SM: Formal analysis, Investigation, Methodology, Visualization, Writing – review & editing. RF: Data curation, Formal analysis, Investigation, Methodology, Writing – review & editing. FG: Conceptualization, Funding acquisition, Investigation, Methodology, Resources, Supervision, Validation, Visualization, Writing – review & editing. SC: Resources, Validation, Visualization, Writing – review & editing. YP: Conceptualization, Data curation, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This work was financially supported by the Free University of Bolzano through the project COMPETITIVE and by the PRIN project 2022SCAN2Z EMERGE financed by Italian Ministry of University and Research. This work was supported by the Open Access Publishing Fund of the Free University of Bozen-Bolzano.
Acknowledgments
The authors are grateful to Council for Agricultural Research and Economics-Research Centre for Viticulture and Enology (CREA-VE) for financially supporting Marco Lucchetta.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fsufs.2025.1504761/full#supplementary-material
References
Adetunji, A. T., Lewu, F. B., Mulidzi, R., and Ncube, B. (2017). The biological activities of β-glucosidase, phosphatase and urease as soil quality indicators: a review. J. Soil Sci. Plant Nutr. 17, 794–807. doi: 10.4067/S0718-95162017000300018
Aguilar-Paredes, A., Valdés, G., Araneda, N., Valdebenito, E., Hansen, F., and Nuti, M. (2023). Microbial Community in the Composting Process and its Positive Impact on the soil biota in sustainable agriculture. Agronomy 13:542. doi: 10.3390/agronomy13020542
Aislabie, J., and Deslippe, J. R. (2013). “Soil microbes and their contribution to soil services” in Ecosystem services in New Zealand. ed. J. R. Dymond (Lincoln, New Zealand: Manaaki Press).
Ali, A., Hanif, S., Bajwa, R., and Jabeen, R. (2011). Assessing enzyme activity of Trichoderma species. Int. J. Biol. Biotechnol. 8, 377–381.
Ali, J., Sharma, D. C., Bano, A., Gupta, A., Sharma, S., Bajpai, P., et al. (2019). “Exploiting microbial enzymes for augmenting crop production” in Enzymes in food biotechnology. ed. M. Kuddus (London, Academic Press), 503–519.
Amora-Lazcano, E., Vázquez, M. M., and Azcón, R. (1998). Response of nitrogen-transforming microorganisms to arbuscular mycorrhizal fungi. Biol. Fertil. Soils 27, 65–70. doi: 10.1007/s003740050401
Balestrini, R., Brunetti, C., Chitarra, W., and Nerva, L. (2020). Photosynthetic traits and nitrogen uptake in crops: which is the role of Arbuscular Mycorrhizal Fungi? Plan. Theory 9:1105. doi: 10.3390/plants9091105
Bernal-Vicente, A., Pascual, J. A., Tittarelli, F., Hernández, J. A., and Diaz-Vivancos, P. (2015). Trichoderma harzianum T-78 supplementation of compost stimulates the antioxidant defence system in melon plants. J. Sci. Food Agric. 95, 2208–2214. doi: 10.1002/jsfa.6936
Bernard, E., Larkin, R. P., Tavantzis, S., Erich, M. S., Alyokhin, A., Sewell, G., et al. (2012). Compost, rapeseed rotation, and biocontrol agents significantly impact soil microbial communities in organic and conventional potato production systems. Appl. Soil Ecol. 52, 29–41. doi: 10.1016/j.apsoil.2011.10.002
Berruti, A., Lumini, E., Balestrini, R., and Bianciotto, V. (2016). Arbuscular Mycorrhizal Fungi as natural biofertilizers: Let’s benefit from past successes. Front. Microbiol. 6:1559. doi: 10.3389/fmicb.2015.01559
Boutchich, G. E. K., Tahiri, S., Krati, M. E., Kabil, E. M., Lhadi, E. K., Mahi, M., et al. (2018). Sandy soil modification by bio-composts for wheat production. Waste Biomass Valoriz. 9, 1129–1139. doi: 10.1007/s12649-017-9847-7
Burgos, P., Madejón, E., and Cabrera, F. (2006). Nitrogen mineralization and nitrate leaching of a sandy soil amended with different organic wastes. Waste Manag. Res. 24, 175–182. doi: 10.1177/0734242X06062876
Calleja-Cervantes, M. E., Fernández-González, A. J., Irigoyen, I., Fernández-López, M., Aparicio-Tejo, P. M., and Menéndez, S. (2015). Thirteen years of continued application of composted organic wastes in a vineyard modify soil quality characteristics. Soil Biol. Biochem. 90, 241–254. doi: 10.1016/j.soilbio.2015.07.002
Carrera, L. M., Buyer, J. S., Vinyard, B., Abdul-Baki, A. A., Sikora, L. J., and Teasdale, J. R. (2007). Effects of cover crops, compost, and manure amendments on soil microbial community structure in tomato production systems. Appl. Soil Ecol. 37, 247–255. doi: 10.1016/j.apsoil.2007.08.003
Cavagnaro, T. R., Jackson, L. E., Scow, K. M., and Hristova, K. R. (2007). Effects of arbuscular mycorrhizas on ammonia oxidizing bacteria in an organic farm soil. Microb. Ecol. 54, 618–626. doi: 10.1007/s00248-007-9212-7
Chakrabarti, K., Sarkar, B., Chakraborty, A., Banik, P., and Bagchi, D. K. (2000). Organic recycling for soil quality conservation in a sub-tropical plateau region. J. Agron. Crop Sci. 184, 137–142. doi: 10.1046/j.1439-037x.2000.00352.x
Chang, E. H., Chung, R. S., and Tsai, Y. H. (2007). Effect of different application rates of organic fertilizer on soil enzyme activity and microbial population. Soil Sci. Plant Nutr. 53, 132–140. doi: 10.1111/j.1747-0765.2007.00122.x
Das, S. K., and Varma, A. (2011). “Role of enzymes in maintaining soil health” in Soil enzymology. eds. G. Shukla and A. Varma (Berlin, Heidelberg: Springer), 25–42.
De Corato, U. (2020). Agricultural waste recycling in horticultural intensive farming systems by on-farm composting and compost-based tea application improves soil quality and plant health: A review under the perspective of a circular economy. Sci. Total Environ. 738:139840. doi: 10.1016/j.scitotenv.2020.139840
Diacono, M., and Montemurro, F. (2011). “Long-term effects of organic amendments on soil fertility” in Sustainable Agriculture Volume 2. eds. E. Lichtfouse, M. Hamelin, M. Navarrete, and P. Debaeke (Springer Netherlands: Dordrecht), 761–786.
Directorate-General for Agriculture and Rural Development (2024). Sustainable use of nutrients. Available at: https://agriculture.ec.europa.eu/sustainability/environmental-sustainability/low-input-farming/nutrients_en (Accessed May 6, 2024).
El Enshasy, H. A., Ambehabati, K. K., El Baz, A. F., Ramchuran, S. O., Sayyed, R. Z., Amalin, D., et al. (2020). “Trichoderma: biocontrol agents for promoting plant growth and soil health” in Agriculturally important Fungi for sustainable agriculture: Volume 2: Functional annotation for crop protection. eds. A. N. Yadav, S. Mishra, D. Kour, N. Yadav, and A. Kumar (Cham: Springer International Publishing), 239–259.
Elad, Y. (2000). Biological control of foliar pathogens by means of Trichoderma harzianum and potential modes of action. Crop Prot. 19, 709–714. doi: 10.1016/S0261-2194(00)00094-6
Eldridge, S. M., Yin Chan, K., Donovan, N. J., Saleh, F., Orr, L., and Barchia, I. (2018). Agronomic and economic benefits of green-waste compost for peri-urban vegetable production: implications for food security. Nutr. Cycl. Agroecosyst. 111, 155–173. doi: 10.1007/s10705-018-9931-9
Eurostat (2024). Vineyards in the EU - statistics. Available at: https://ec.europa.eu/eurostat/statistics-explained/index.php?title=Vineyards_in_the_EU_-_statistics (Accessed May 6, 2024).
Fan, X., Chen, X., Chen, T., Liu, X., Song, Y., Tan, S., et al. (2023). Effects of substituting synthetic nitrogen with organic amendments on crop yield, net greenhouse gas emissions and carbon footprint: A global meta-analysis. Field Crop Res. 301:109035. doi: 10.1016/j.fcr.2023.109035
Fertilizers Europe (2023). Forecast of food, farming & fertilizer use in the European Union 2022–2032. Brussel: Fertilizers Europe.
Francioli, D., Schulz, E., Lentendu, G., Wubet, T., Buscot, F., and Reitz, T. (2016). Mineral vs. organic amendments: microbial community structure, activity and abundance of agriculturally relevant microbes are driven by long-term fertilization strategies. Front. Microbiol. 7:1446. doi: 10.3389/fmicb.2016.01446
Gaiotti, F., Marcuzzo, P., Belfiore, N., Lovat, L., Fornasier, F., and Tomasi, D. (2017). Influence of compost addition on soil properties, root growth and vine performances of Vitis vinifera cv cabernet sauvignon. Sci. Hortic. 225, 88–95. doi: 10.1016/j.scienta.2017.06.052
Garcı́a, C., Hernández, T., Costa, F., and Ceccanti, B. (1994). Biochemical parameters in soils regenerated by the addition of organic wastes. Waste Manag. Res. 12, 457–466. doi: 10.1006/wmre.1994.1035
García-Orenes, F., Roldán, A., Morugán-Coronado, A., Linares, C., Cerdà, A., and Caravaca, F. (2016). Organic fertilization in traditional Mediterranean grapevine orchards mediates changes in soil microbial community structure and enhances soil fertility. Land Degrad. Dev. 27, 1622–1628. doi: 10.1002/ldr.2496
Garland, J. L., and Mills, A. L. (1991). Classification and characterization of heterotrophic microbial communities on the basis of patterns of community-level sole-carbon-source utilization. Appl. Environ. Microbiol. 57, 2351–2359. doi: 10.1128/aem.57.8.2351-2359.1991
Gatti, M., Schippa, M., Garavani, A., Squeri, C., Frioni, T., Dosso, P., et al. (2020). High potential of variable rate fertilization combined with a controlled released nitrogen form at affecting cv. Barbera vines behavior. Eur. J. Agron. 112, 239–259. doi: 10.1016/j.eja.2019.125949
Ge, Z., Du, H., Gao, Y., and Qiu, W. (2018). Analysis on metabolic functions of stored Rice microbial communities by BIOLOG ECO microplates. Front. Microbiol. 9:1375. doi: 10.3389/fmicb.2018.01375
Gil-Sotres, F., Trasar-Cepeda, C., Leirós, M. C., and Seoane, S. (2005). Different approaches to evaluating soil quality using biochemical properties. Soil Biol. Biochem. 37, 877–887. doi: 10.1016/j.soilbio.2004.10.003
Goldfarb, K. C., Karaoz, U., Hanson, C. A., Santee, C. A., Bradford, M. A., Treseder, K. K., et al. (2011). Differential growth responses of soil bacterial taxa to carbon substrates of varying chemical recalcitrance. Front. Microbiol. 2:94. doi: 10.3389/fmicb.2011.00094
Hao, Z., Fayolle, L., van Tuinen, D., Chatagnier, O., Li, X., Gianinazzi, S., et al. (2012). Local and systemic mycorrhiza-induced protection against the ectoparasitic nematode Xiphinema index involves priming of defence gene responses in grapevine. J. Exp. Bot. 63, 3657–3672. doi: 10.1093/jxb/ers046
Hempel, S., Renker, C., and Buscot, F. (2007). Differences in the species composition of arbuscular mycorrhizal fungi in spore, root and soil communities in a grassland ecosystem. Environ. Microbiol. 9, 1930–1938. doi: 10.1111/j.1462-2920.2007.01309.x
Hermosa, R., Viterbo, A., Chet, I., and Monte, E. (2012). Plant-beneficial effects of Trichoderma and of its genes. Microbiology 158, 17–25. doi: 10.1099/mic.0.052274-0
Hidangmayum, A., and Dwivedi, P. (2018). Plant responses to Trichoderma spp. and their tolerance to abiotic stresses: A review. J. Pharmacogn. Phytochem. 7, 758–766. Available at: https://www.phytojournal.com/archives/2018.v7.i1.2575/plant-responses-to-trichoderma-spp-and-their-tolerance-to-abiotic-stresses-a-review (Accessed May 6, 2024).
Hildebrandt, U., Ouziad, F., Marner, F.-J., and Bothe, H. (2006). The bacterium Paenibacillus validus stimulates growth of the arbuscular mycorrhizal fungus Glomus intraradices up to the formation of fertile spores. FEMS Microbiol. Lett. 254, 258–267. doi: 10.1111/j.1574-6968.2005.00027.x
Hodge, A., Helgason, T., and Fitter, A. H. (2010). Nutritional ecology of arbuscular mycorrhizal fungi. Fungal Ecol. 3, 267–273. doi: 10.1016/j.funeco.2010.02.002
Hue, N. V., and Silva, J. A. (2000). “Organic soil amendments for sustainable agriculture” in Plant nutrient Management in Hawaii’s soils. eds. J. A. Silva and R. S. Uchida (Manoa, Hawaii: University of Hawaii at Manoa), 133–144.
Iovieno, P., Morra, L., Leone, A., Pagano, L., and Alfani, A. (2009). Effect of organic and mineral fertilizers on soil respiration and enzyme activities of two Mediterranean horticultural soils. Biol. Fertil. Soils 45, 555–561. doi: 10.1007/s00374-009-0365-z
Işler, N., İlay, R., and Kavdir, Y. (2022). Temporal variations in soil aggregation following olive pomace and vineyard pruning waste compost applications on clay, loam, and sandy loam soils. Environ. Monit. Assess. 194:418. doi: 10.1007/s10661-022-10093-w
Jezierska-Tys, S., Wesołowska, S., Gałązka, A., Joniec, J., Bednarz, J., and Cierpiała, R. (2020). Biological activity and functional diversity in soil in different cultivation systems. Int. J. Environ. Sci. Technol. 17, 4189–4204. doi: 10.1007/s13762-020-02762-5
Jin, Z., Lei, J., Li, S., and Xu, X. (2015). Metabolic characteristics of microbial communities of Aeolian sandy soils induced by saline water drip irrigation in shelter forests. Eur. J. Soil Sci. 66, 476–484. doi: 10.1111/ejss.12230
Joner, E. J., and Johansen, A. (2000). Phosphatase activity of external hyphae of two arbuscular mycorrhizal fungi. Mycol. Res. 104, 81–86. doi: 10.1017/S0953756299001240
Kandeler, E., and Gerber, H. (1988). Short-term assay of soil urease activity using colorimetric determination of ammonium. Biol. Fertil. Soils 6, 68–72. doi: 10.1007/BF00257924
Karimi, B., Cahurel, J.-Y., Gontier, L., Charlier, L., Chovelon, M., Mahé, H., et al. (2020). A meta-analysis of the ecotoxicological impact of viticultural practices on soil biodiversity. Environ. Chem. Lett. 18, 1947–1966. doi: 10.1007/s10311-020-01050-5
Lal, R., Bouma, J., Brevik, E., Dawson, L., Field, D. J., Glaser, B., et al. (2021). Soils and sustainable development goals of the United Nations: an International Union of Soil Sciences perspective. Geoderma Reg. 25:e00398. doi: 10.1016/j.geodrs.2021.e00398
Lazcano, C., Decock, C., and Wilson, S. G. (2020). Defining and managing for healthy vineyard soils, intersections with the concept of terroir. Front. Environ. Sci. 8:68. doi: 10.3389/fenvs.2020.00068
Leifheit, E. F., Veresoglou, S. D., Lehmann, A., Morris, E. K., and Rillig, M. C. (2014). Multiple factors influence the role of arbuscular mycorrhizal fungi in soil aggregation—a meta-analysis. Plant Soil 374, 523–537. doi: 10.1007/s11104-013-1899-2
Li, L., Xu, M., Ali, M. E., Zhang, W., Duan, Y., and Li, D. (2018). Factors affecting soil microbial biomass and functional diversity with the application of organic amendments in three contrasting cropland soils during a field experiment. PLoS One 13:e0203812. doi: 10.1371/journal.pone.0203812
Li, J., Zhao, B., Li, X., Jiang, R., and So, H. B. (2008). Effects of long-term combined application of organic and mineral fertilizers on microbial biomass, soil enzyme activities and soil fertility. Agric. Sci. China 7, 336–343. doi: 10.1016/S1671-2927(08)60074-7
Litskas, V., Mandoulaki, A., Vogiatzakis, I. N., Tzortzakis, N., and Stavrinides, M. (2020). Sustainable viticulture: first determination of the environmental footprint of grapes. Sustain. For. 12:8812. doi: 10.3390/su12218812
Liu, X., Shi, Y., Kong, L., Tong, L., Cao, H., Zhou, H., et al. (2022). Long-term application of bio-compost increased soil microbial community diversity and altered its composition and network. Microorganisms 10:462. doi: 10.3390/microorganisms10020462
Lundquist, E. J., Scow, K. M., Jackson, L. E., Uesugi, S. L., and Johnson, C. R. (1999). Rapid response of soil microbial communities from conventional, low input, and organic farming systems to a wet/dry cycle. Soil Biol. Biochem. 31, 1661–1675. doi: 10.1016/S0038-0717(99)00080-2
Maltais-Landry, G., Scow, K., Brennan, E., and Vitousek, P. (2015). Long-term effects of compost and cover crops on soil phosphorus in two California agroecosystems. Soil Sci. Soc. Am. J. 79, 688–697. doi: 10.2136/sssaj2014.09.0369
Mondini, C., Fornasier, F., Sinicco, T., Sivilotti, P., Gaiotti, F., and Mosetti, D. (2018). Organic amendment effectively recovers soil functionality in degraded vineyards. Eur. J. Agron. 101, 210–221. doi: 10.1016/j.eja.2018.10.002
Moral, R., Paredes, C., Bustamante, M. A., Marhuenda-Egea, F., and Bernal, M. P. (2009). Utilisation of manure composts by high-value crops: safety and environmental challenges. Bioresour. Technol. 100, 5454–5460. doi: 10.1016/j.biortech.2008.12.007
Morlat, R., and Chaussod, R. (2008). Long-term additions of organic amendments in a Loire Valley vineyard. I. Effects on properties of a calcareous Sandy soil. Am. J. Enol. Vitic. 59, 353–363. doi: 10.5344/ajev.2008.59.4.353
Muscarella, M. E., Howey, X. M., and Lennon, J. T. (2020). Trait-based approach to bacterial growth efficiency. Environ. Microbiol. 22, 3494–3504. doi: 10.1111/1462-2920.15120
Najafi-Ghiri, M., Niksirat, S. H., Soleimanpour, L., and Nowzari, S. (2018). Comparison of different organic amendments on potassium release from two fine-textured soils. Org. Agric. 8, 129–140. doi: 10.1007/s13165-017-0179-5
Nannipieri, P., Kandeler, E., and Ruggiero, P. (2002). “Enzyme activities and microbiological and biochemical processes in soil” in Enzymes in the environment: Activity, ecology, and applications. eds. R. Burns and R. Dick (New York: CRC Press).
Niehaus, L., Boland, I., Liu, M., Chen, K., Fu, D., Henckel, C., et al. (2019). Microbial coexistence through chemical-mediated interactions - nature communications. Nat. Commun. 10:2052. doi: 10.1038/s41467-019-10062-x
O’Neill, T. M., Elad, Y., Shtienberg, D., and Cohen, A. (1996). Control of grapevine Grey Mould with Trichoderma harzianum T39. Biocontrol Sci. Tech. 6, 139–146. doi: 10.1080/09583159650039340
Okur, N., Altindșli, A., Çengel, M., Göçmez, S., and Kayikçioğlu, H. (2009). Microbial biomass and enzyme activity in vineyard soils under organic and conventional farming systems. Turk. J. Agric. For. 33, 413–423. doi: 10.3906/tar-0806-23
Olsen, S. R., and Sommers, L. E. (1982). “Phosphorus”, in Methods of Soil Analysis. ed. A. L. Page (Wiley), 403–430.
Pahalvi, H. N., Rafiya, L., Rashid, S., Nisar, B., and Kamili, A. N. (2021). “Chemical fertilizers and their impact on soil health,” in Microbiota and biofertilizers, Vol 2: Ecofriendly tools for reclamation of degraded soil environs, eds. G. H. Dar, Bhat, R. A., and Mehmood, M. A., and K. R. Hakeem (Cham: Springer International Publishing), 1–20.
Pascale, A., Vinale, F., Manganiello, G., Nigro, M., Lanzuise, S., Ruocco, M., et al. (2017). Trichoderma and its secondary metabolites improve yield and quality of grapes. Crop Prot. 92, 176–181. doi: 10.1016/j.cropro.2016.11.010
Pascual, J. A., García, C., and Hernandez, T. (1999). Lasting microbiological and biochemical effects of the addition of municipal solid waste to an arid soil. Biol. Fertil. Soils 30, 1–6. doi: 10.1007/s003740050579
Preston-Mafham, J., Boddy, L., and Randerson, P. F. (2002). Analysis of microbial community functional diversity using sole-carbon-source utilisation profiles – a critique. FEMS Microbiol. Ecol. 42, 1–14. doi: 10.1111/j.1574-6941.2002.tb00990.x
Qiao, L., Wang, X., Smith, P., Fan, J., Lu, Y., Emmett, B., et al. (2022). Soil quality both increases crop production and improves resilience to climate change. Nat. Clim. Chang. 12, 574–580. doi: 10.1038/s41558-022-01376-8
Quemada, M., Baranski, M., Nobel-de Lange, M. N. J., Vallejo, A., and Cooper, J. M. (2013). Meta-analysis of strategies to control nitrate leaching in irrigated agricultural systems and their effects on crop yield. Agric. Ecosyst. Environ. 174, 1–10. doi: 10.1016/j.agee.2013.04.018
Raupp, J., Niehus, A., and Oltmanns, M. (2004). Die Diversität der Boden-Mikroflora ist bei Rottemistdüngung höher als bei Mineraldüngung. Mitt. Ges. Pflanzenbauwiss. 16, 149–150.
Romero, P., Navarro, J. M., and Ordaz, P. B. (2022). Towards a sustainable viticulture: the combination of deficit irrigation strategies and agroecological practices in Mediterranean vineyards. A review and update. Agric. Water Manag. 259:107216. doi: 10.1016/j.agwat.2021.107216
Ros, M., Goberna, M., Pascual, J. A., Klammer, S., and Insam, H. (2008). 16S rDNA analysis reveals low microbial diversity in community level physiological profile assays. J. Microbiol. Methods 72, 221–226. doi: 10.1016/j.mimet.2008.01.003
Saha, S., Mina, B. L., Gopinath, K. A., Kundu, S., and Gupta, H. S. (2008). Organic amendments affect biochemical properties of a subtemperate soil of the Indian Himalayas. Nutr. Cycl. Agroecosyst. 80, 233–242. doi: 10.1007/s10705-007-9139-x
Saifuddin, M., Bhatnagar, J. M., Segrè, D., and Finzi, A. C. (2019). Microbial carbon use efficiency predicted from genome-scale metabolic models. Nat. Commun. 10:3568. doi: 10.1038/s41467-019-11488-z
Sandle, T. (2016). “22.5.8 nutrients,” in pharmaceutical microbiology” in Essentials for quality assurance and quality control (Cambridge, UK: Woodhead Publishing).
Schoenau, J. J., and Davis, J. G. (2006). Optimizing soil and plant responses to land-applied manure nutrients in the Great Plains of North America. Can. J. Soil Sci. 86, 587–595. doi: 10.4141/S05-115
Scotti, R., Bonanomi, G., Scelza, R., Zoina, A., and Rao, M. A. (2015). Organic amendments as sustainable tool to recovery fertility in intensive agricultural systems. J. Soil Sci. Plant Nutr. 15, 333–352. doi: 10.4067/S0718-95162015005000031
Si, P., Shao, W., Yu, H., Yang, X., Gao, D., Qiao, X., et al. (2018). Rhizosphere microenvironments of eight common deciduous fruit trees were shaped by microbes in northern China. Front. Microbiol. 9:3147. doi: 10.3389/fmicb.2018.03147
Smith, S. E., and Read, D. J. (2010). “Arbuscular Mycorrhizas” in Mycorrhizal Symbiosis. ed. S. E. Smith (San Diego, US: Academic Press), 145–189.
Somerville, P. D., May, P. B., and Livesley, S. J. (2018). Effects of deep tillage and municipal green waste compost amendments on soil properties and tree growth in compacted urban soils. J. Environ. Manag. 227, 365–374. doi: 10.1016/j.jenvman.2018.09.004
Souza, A. A., Leitão, V. O., Ramada, M. H., Mehdad, A., Georg, R. D. C., Ulhôa, C. J., et al. (2016). Trichoderma harzianum produces a new thermally stable acid phosphatase, with potential for biotechnological application. PLoS One 11:e0150455. doi: 10.1371/journal.pone.0150455
Sradnick, A., Murugan, R., Oltmanns, M., Raupp, J., and Joergensen, R. G. (2013). Changes in functional diversity of the soil microbial community in a heterogeneous sandy soil after long-term fertilization with cattle manure and mineral fertilizer. Appl. Soil Ecol. 63, 23–28. doi: 10.1016/j.apsoil.2012.09.011
Szczałba, M., Kopta, T., Gąstoł, M., and Sękara, A. (2019). Comprehensive insight into arbuscular mycorrhizal fungi, Trichoderma spp. and plant multilevel interactions with emphasis on biostimulation of horticultural crops. J. Appl. Microbiol. 127, 630–647. doi: 10.1111/jam.14247
Tabatabai, M. A. (1994). “Soil Enzymes,” in Methods of soil analysis. eds. R. W. Weaver, S. Angle, P. Bottomley, D. Bezdicek, S. Smith, and A. Hedges, et al. (John Wiley & Sons, Ltd), 775–833.
Tanaka, Y., and Yano, K. (2005). Nitrogen delivery to maize via mycorrhizal hyphae depends on the form of N supplied. Plant Cell Environ. 28, 1247–1254. doi: 10.1111/j.1365-3040.2005.01360.x
Tang, J., Zhang, L., Zhang, J., Ren, L., Zhou, Y., Zheng, Y., et al. (2020). Physicochemical features, metal availability and enzyme activity in heavy metal-polluted soil remediated by biochar and compost. Sci. Total Environ. 701:134751. doi: 10.1016/j.scitotenv.2019.134751
Torres, I. F., Bastida, F., Hernández, T., and García, C. (2015). The effects of fresh and stabilized pruning wastes on the biomass, structure and activity of the soil microbial community in a semiarid climate. Appl. Soil Ecol. 89, 1–9. doi: 10.1016/j.apsoil.2014.12.009
Velasco-Muñoz, J. F., Aznar-Sánchez, J. A., López-Felices, B., and Román-Sánchez, I. M. (2022). Circular economy in agriculture an analysis of the state of research based on the life cycle. Sustain. Prod. Consum. 34, 257–270. doi: 10.1016/j.spc.2022.09.017
Veresoglou, S. D., Chen, B., and Rillig, M. C. (2012). Arbuscular mycorrhiza and soil nitrogen cycling. Soil Biol. Biochem. 46, 53–62. doi: 10.1016/j.soilbio.2011.11.018
Wang, Y., Wilhelm, R. C., Swenson, T. L., Silver, A., Andeer, P. F., Golini, A., et al. (2022). Substrate utilization and Competitive interactions among soil Bacteria vary with life-history strategies. Front. Microbiol. 13:914472. doi: 10.3389/fmicb.2022.914472
Wei, Z., Hoffland, E., Zhuang, M., Hellegers, P., and Cui, Z. (2021). Organic inputs to reduce nitrogen export via leaching and runoff: A global meta-analysis. Environ. Pollut. 291:118176. doi: 10.1016/j.envpol.2021.118176
Wipf, D., Krajinski, F., van Tuinen, D., Recorbet, G., and Courty, P.-E. (2019). Trading on the arbuscular mycorrhiza market: from arbuscules to common mycorrhizal networks. New Phytol. 223, 1127–1142. doi: 10.1111/nph.15775
Wu, F., Li, Z., Lin, Y., and Zhang, L. (2021). Effects of Funneliformis mosseae on the utilization of organic phosphorus in Camellia oleifera Abel. Can. J. Microbiol. 67, 349–357. doi: 10.1139/cjm-2020-0227
Zengler, K., and Zaramela, L. S. (2018). The social network of microorganisms — how auxotrophies shape complex communities - nature reviews microbiology. Nat. Rev. Microbiol. 16, 383–390. doi: 10.1038/s41579-018-0004-5
Zhang, X., Davidson, E. A., Mauzerall, D. L., Searchinger, T. D., Dumas, P., and Shen, Y. (2015). Managing nitrogen for sustainable development. Nature 528, 51–59. doi: 10.1038/nature15743
Zhao, J., Pullens, J. W. M., Sørensen, P., Blicher-Mathiesen, G., Olesen, J. E., and Børgesen, C. D. (2022). Agronomic and environmental factors influencing the marginal increase in nitrate leaching by adding extra mineral nitrogen fertilizer. Agric. Ecosyst. Environ. 327:107808. doi: 10.1016/j.agee.2021.107808
Zheng, H., Yun, O. Z., and Fang, Z. G. (2004). Application of BIOLOG to study on soil microbial community functional diversity. Acta Pedol. Sin. 41, 456–461. Available at: http://pedologica.issas.ac.cn/trxben/article/abstract/20040320 (Accessed May 3, 2024).
Zhong, W., Gu, T., Wang, W., Zhang, B., Lin, X., Huang, Q., et al. (2010). The effects of mineral fertilizer and organic manure on soil microbial community and diversity. Plant Soil 326, 511–522. doi: 10.1007/s11104-009-9988-y
Keywords: soil fertility, soil enzymes, mineral nutrients cycling, microbial communities, pruning waste compost
Citation: Lucchetta M, Zuluaga MYA, Monterisi S, Fattorini R, Gaiotti F, Cesco S and Pii Y (2025) Innovative approaches to soil health: evaluating pruning waste compost, inoculated compost, and mineral fertilizer in a controlled growth chamber for sustainable vineyard practices. Front. Sustain. Food Syst. 9:1504761. doi: 10.3389/fsufs.2025.1504761
Edited by:
Jay Prakash Verma, Banaras Hindu University, IndiaReviewed by:
Gowardhan Kumar Chouhan, Banaras Hindu University, IndiaYerlan Issakov, Abai University, Kazakhstan
Copyright © 2025 Lucchetta, Zuluaga, Monterisi, Fattorini, Gaiotti, Cesco and Pii. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: M. Lucchetta, bWFyY28ubHVjY2hldHRhQHN0dWRlbnQudW5pYnouaXQ=; Y. Pii, eW91cnkucGlpQHVuaWJ6Lml0
†ORCID: M. Lucchetta, orcid.org/0009-0006-2094-6751