 Abhay K. Pandey
Abhay K. Pandey Mahesh K. Samota
Mahesh K. Samota Abhishek Kumar
Abhishek Kumar Ana Sanches Silva
Ana Sanches Silva Nawal K. Dubey
Nawal K. Dubey
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Sustain. Food Syst., 05 May 2023
Sec. Agro-Food Safety
Volume 7 - 2023 | https://doi.org/10.3389/fsufs.2023.1162595
This article is part of the Research TopicMycotoxins and Their Management in Sustainable Food SystemsView all 5 articles
Mycotoxins are toxic secondary metabolites produced by fungi when they colonies the foodstuffs. These are potent toxins having severe health consequences in people, being mutagenic, teratogenic, and carcinogenic. In agricultural commodities, the contamination of mycotoxins is more prevalent. Several fungi can produce mycotoxins on agricultural products during harvest or in postharvest, and they have significant adverse effects on both animal and human beings. The most prevalent mycotoxins found in food commodities are aflatoxins and ochratoxins produced by Aspergillus species, ochratoxins and patulin produced by Penicillium, as well as fumonisins, deoxynivalenol, and zearalenone produced by Fusarium species. Worldwide, fumonisins, patulin, aflatoxins, and ochratoxins, among others, are responsible for numerous acute and chronic diseases in people and domestic animals. In food commodities, mycotoxins have been quantified and detected using various analytical methods. Besides, mycotoxins occurrence in food commodities were decontaminated through many potential approaches, such as physical, chemical, and biological methods. This review summarizes the findings of 30 years of research into mycotoxins in major commercial food crops including wheat, maize, sorghum, pearl millet, peanut, oat, pulses, barley, oilseeds, rice, and fruits and fruit juices. We also discuss the detection methods of major mycotoxins, available decontamination strategies along with their disadvantages and knowledge gaps. It is anticipated that data from meticulous studies on mycotoxins in food commodities will help in the development of safer food and in setting priorities for future research.
An estimated 30 to 50% of food commodities are lost during pre-harvest or post-harvest globally, which does not only threaten global food security but wastes 1.47–1.96 Gha of arable land, 0.75–1.25 trillion cubic meters of water and 1 to 1.5% of global energy (Fox and Fimeche, 2013). The pre-harvest and postharvest losses in food commodities may occur due to attack by several biotic and abiotic factors. In storage system, fungal bio-deterioration of stored food commodities is a chronic problem in tropical hot and humid climates. Harvested food grains can be contaminated by different genera of fungi such as Aspergillus, Alternaria, Fusarium, Cladosporium, Penicillium, Mucor, and Rhizopus (Mateus et al., 2021). Under specific conditions, some fungi can produce toxic metabolites called mycotoxins, leading to food contamination. The major toxin-producing moulds include genera of Aspergillus, Fusarium, Penicillium, and Alternaria (Khodaei et al., 2021; Pandey et al., 2022).
Human and livestock health is adversely affected by consumption of mycotoxins contaminated food products, which affect the marketability of food commodities and raise food safety concerns (Mateus et al., 2021). It is estimated that more than five billion people are exposed to mycotoxins daily by unknown pathways and consume contaminated foods every day (Khodaei et al., 2021). Taking mycotoxins through food can lead to intoxication, known as mycotoxicosis (Tanaka et al., 2007). Mycotoxicosis occurs when a mycotoxin leads to acute or chronic toxicity involving hepatotoxicity, cytotoxicity, teratogenicity, neurotoxicity, mutation, and carcinogenicity. At cellular levels, mycotoxins inhibit DNA and RNA synthesis by interacting with nucleic acids (Smith et al., 2007). In the past years, several reviews published on occurrence of mycotoxins in various food commodities (Tola et al., 2016; Lee and Ryu, 2017; Khodaei et al., 2021). However, these are either crop/country specific or discusses only occurrence or management strategies. In this review, the findings from research data collected over the past 30 years are discussed, including mycotoxins in major commercially important food crops like wheat, maize, sorghum, pearl-millets, peanut, oat, pulses, barley, oilseeds, rice, and fruits. Detection methods for major mycotoxins and available decontamination strategies are also discussed along with their limitations and knowledge gaps.
The mycotoxins produced by fungi are majorly classified as aflatoxins (AFs), ochratoxin A (OTA), patulin (PT), sterigmatocystin (STC), trichothecenes (TCTs) fumonisins (FBs), deoxynivalenol (DON), zearalenone (ZEA), alternariol (AOH), tenuazonic acid, and alternariol monomethyl ether. Table 1 provides information about some important mycotoxins, food commodities they are contaminating and their toxic effects. Aflatoxin is an enormously toxic mycotoxin and is produced worldwide mainly by A. flavus and A. parasiticus (Pandey et al., 2016). It includes four major types, such as B1, B2, G1, and G2. It has been determined that A. flavus produces B toxins, the most common of which is B1, which is carcinogenic and genotoxic (Payne and Brown, 1998; Abbas et al., 2008). Compared to other crops, maize and cotton have higher levels of AFs produced by A. flavus (Hell et al., 2000); nevertheless, in groundnut (peanut) AF derived from A. parasiticus is common (Kaaya et al., 2006). Another mycotoxin, ZEA is also known as F-2 mycotoxin. Many species of Fusarium produce DON, T2, and HT-2 toxins, ZEA, and diacetoxyscirpenol (DAS), toxic chemicals of considerable concern to livestock and poultry producers (Kuiper-Goodman et al., 1987). A number of food products can be affected by these toxins as demonstrated in Table 1. In pigs, especially, it can cause infertility, abortion, and other breeding problems (Kuiper-Goodman et al., 1987).
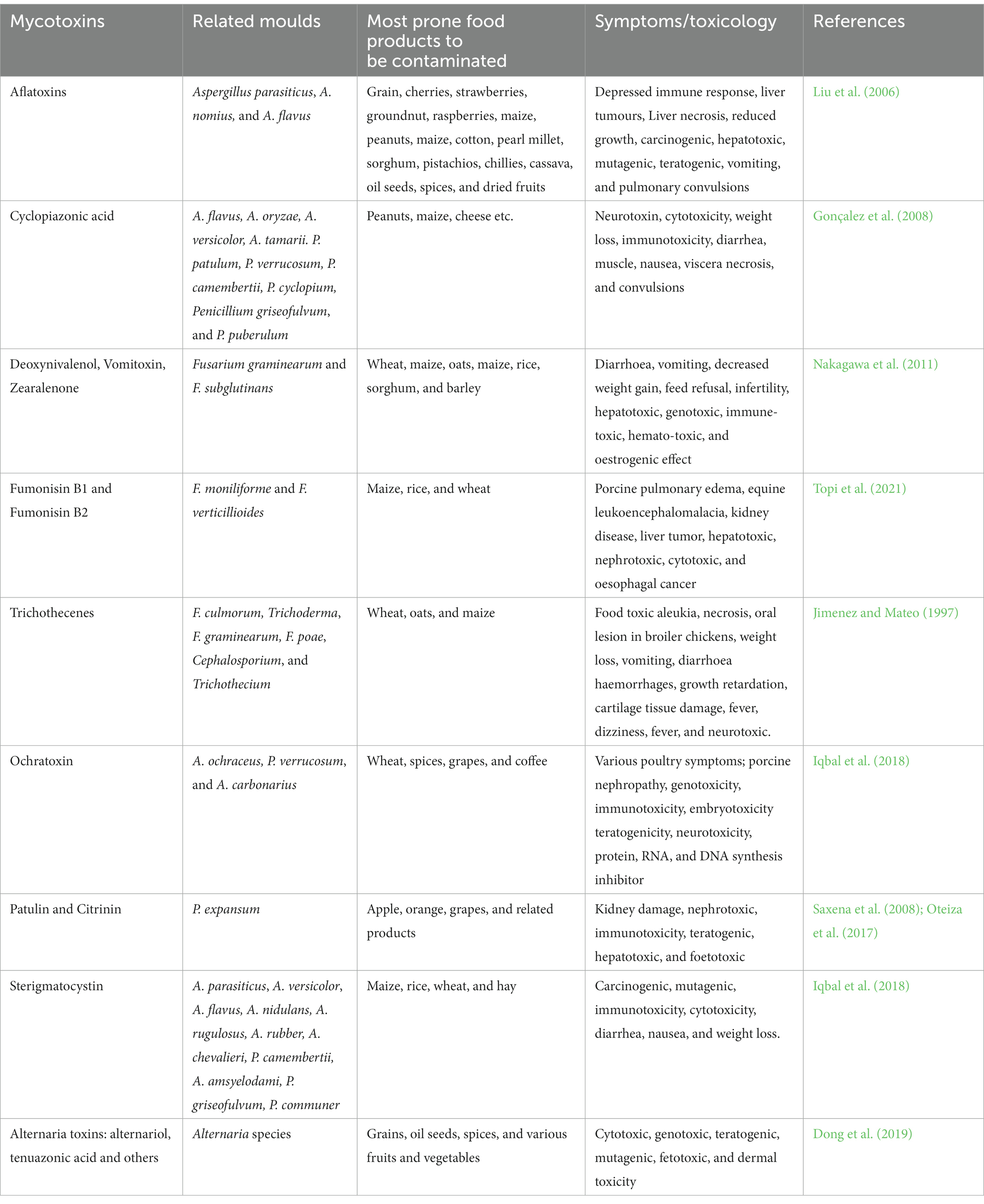
Table 1. List of important mycotoxins, most prone food commodities to be contaminated by mycotoxins and their principal toxic effects.
Fumonisins are mycotoxins produced by Fusarium and are composed of FB1 and FB2. Fusarium verticillioides mainly attack maize, wheat, and many other grains, produces FB1, which is the most prevalent member of this family of toxin-producing Fusarium moulds. There is also evidence that F. verticillioides and F. moniliforme produce FB2, which is structurally similar to FB1 (Polišenská et al., 2020). Compared with FB1, FB2 is more cytotoxic and inhibits acylsphingosine transferase. Maize and other commodities are also contaminated by FB2. There have been reports of 15 different FBs to date, although most have not been shown to occur in nature (Polišenská et al., 2020). As far as ochratoxins are concerned, three types of ochratoxins are present in food commodities, namely OTA, OTB, and OTC. OTA is the most prevalent mycotoxin found in foodstuffs, produced by Penicillium verrucosum and species of Aspergillus such as A. carbonarius and A. ochraceus (Al-Anati and Petzinger, 2006). Consumption of OTA-contaminated food products, such as grains, coffee, pork products, grapes, and grape products, can result in human acquaintance (Richard et al., 1999). Besides, Aspergillus, Byssochlamys, and Penicillium produce PT, which is typically found in rotten apples. It has been shown that PT possesses antimicrobial activity against some microorganisms. Health studies have caused several countries to regulate PT’s quantity in commodities as a result of health risks.
In order to prevent their detrimental effects on humans, mycotoxins are regulated by maximum permissible levels in food (Claeys et al., 2020). The contents of mycotoxins in food commodities have been restricted in several countries (Puel et al., 2010). Additionally, a number of national and international agencies, including the Food and Agriculture Organization, World Health Organization, EU Commission, and Codex Alimentarius Commission, have established regulations regarding different types of mycotoxins present in different foods aimed at protecting consumers (Adeyeye, 2016). Most countries have no specific limits on specific foods or food products, but all food products are subject to general regulations. There are, however, some countries in Europe and the USA that have legislated dietary limits for specific foods. Among mycotoxins, AFs have potent genotoxic, carcinogenic, and immunosuppressive effects on people. Therefore, in most food commodities, government agencies have established maximum levels of aflatoxins, including AFB1 (Bhat and Reddy, 2017). The maximum limits (μg/kg) of important fungal mycotoxins in major food commodities that were legislated are summarized in Table 2.
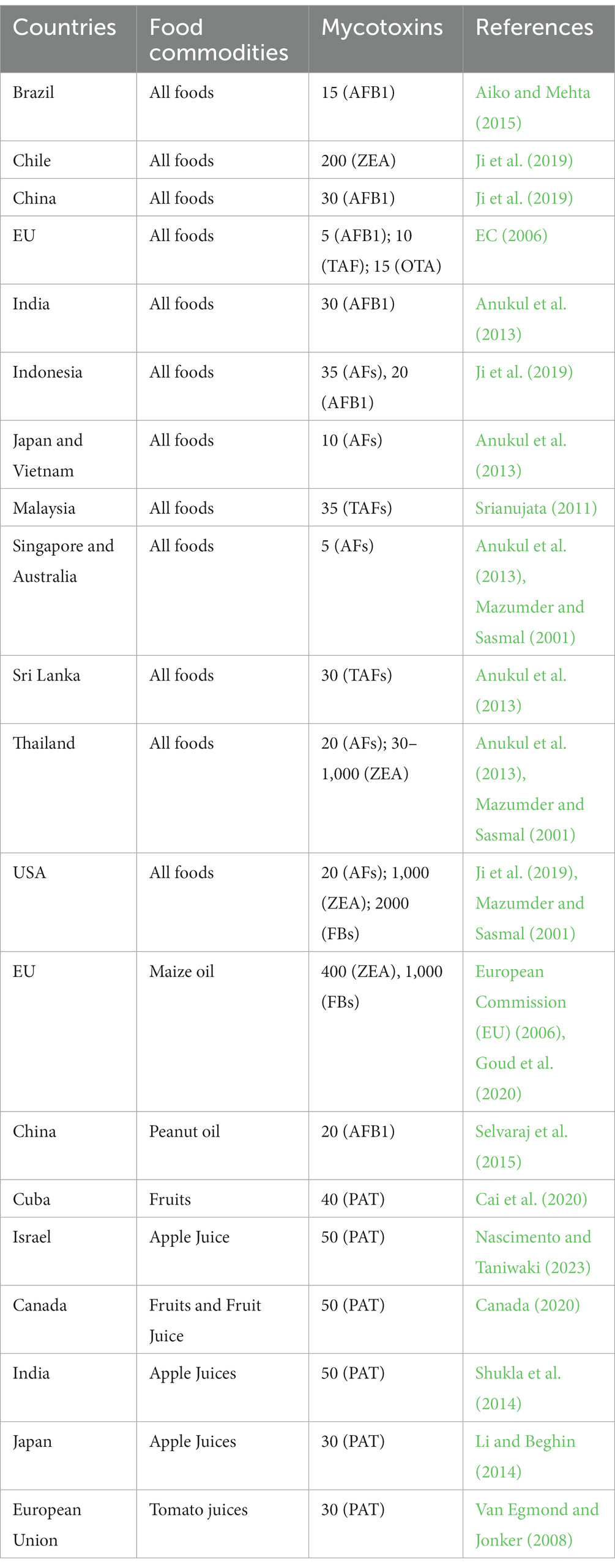
Table 2. The maximum limits (μg/kg) established for major mycotoxins in some countries/regions for all food commodities.
Mycotoxins are toxic and poisonous, can occur even with the very small quantities in food commodities, and consumption of such food products causes several health risks. Therefore, there is a need to analysis and quantification of mycotoxins by sensitive and accurate methods (Le et al., 2021) so that they can reduce before consumptions. For the quantification and detection of mycotoxins in foodstuffs, several analytical methods have been adopted (Berthiller et al., 2017). The most significant ones are discussed in this review, and their advantages and disadvantages are summarized in Table 3.
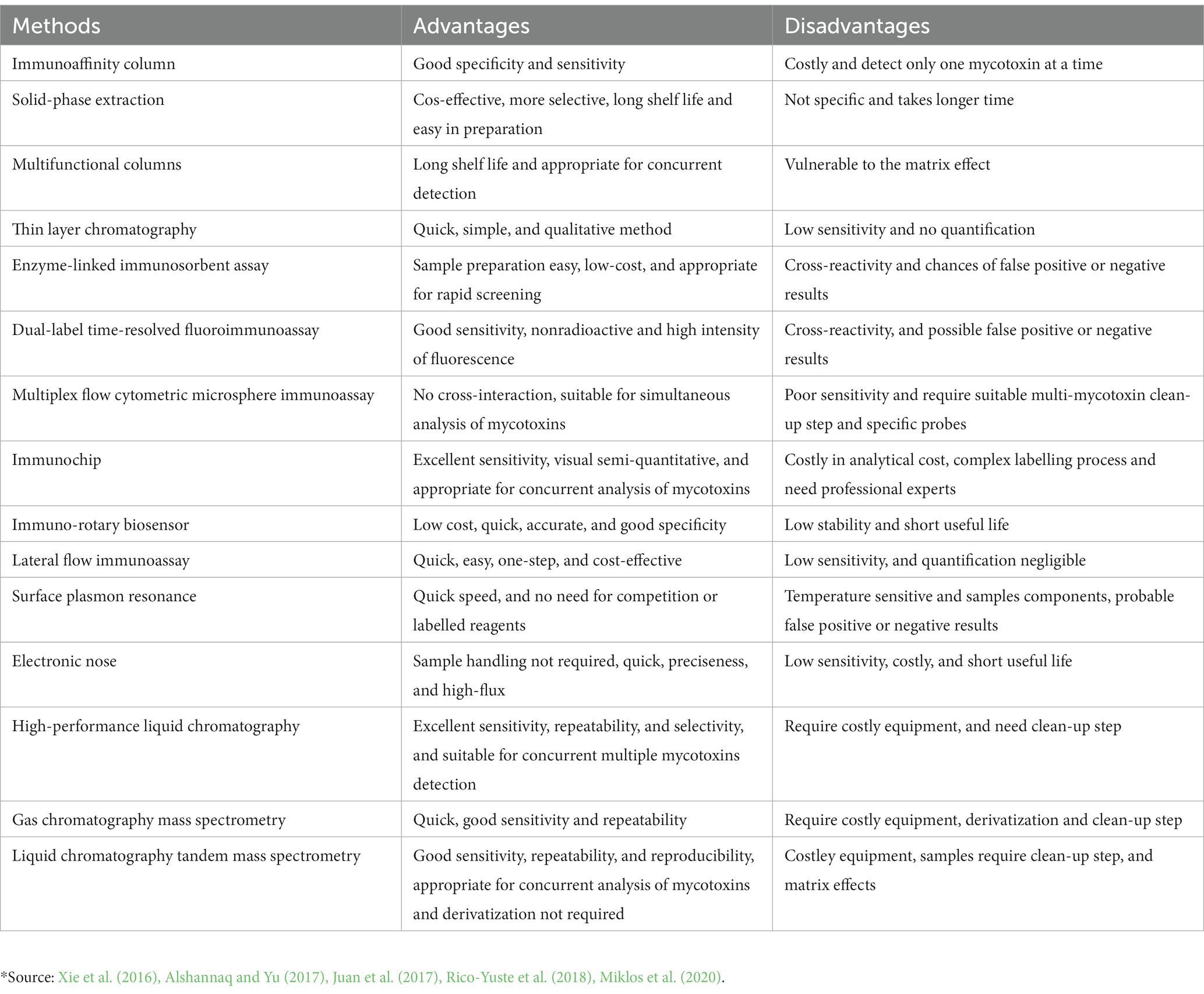
Table 3. Application, advantages, and disadvantages of mycotoxin detection methods.*
Extraction and purification of mycotoxins from foods samples using appropriate solvents is the first step of preparation of samples. QuEChERS (Quick, Easy, Cheap, Effective, Rugged, and Safe) is the method used for the detection of mycotoxins in different matrices. In this method, toxin is firstly extracted with acetonitrile-water, then added inorganic salts for induction of liquid–liquid partitioning, and mycotoxins are moved to the organic phase (Gonzalez-Jartin et al., 2019). QuEChERS has been used in berries-derived jam and juice for the analysis of mycotoxins such as AFs, OTA, etc. (Juan et al., 2017). Liquid – Liquid extraction (LLE) method is based on the different solubility of toxins in the organic phase and aqueous phase. LLE has been also applied for the analysis of AFs and OTA (Andrade et al., 2013; Turner et al., 2015). Liquid–solid extraction (SLE) is the method based on weighing of homogenized sample in extraction solvent followed by agitating it in a shaker. This has been used for the extraction of mycotoxins associated with cereals (Rubert et al., 2012; Xie et al., 2016). Pressurized Liquid Extraction (PLE) or Accelerated Solvent Extraction (ASE), is the method performed at increased temperature (100–180°C) and pressure (1,500–2,000 psi) in a vessel that improved extraction of mycotoxins. This method was used to detect mycotoxins in tomato samples (Rico-Yuste et al., 2018; Miklos et al., 2020). In another study, supercritical Fluid Extraction (SFE) is used for the extraction of non-polar organic molecules such as ZEA detection in maize flour (Zougagh and Ríos, 2008). The advantage of the SFE method is that it eliminates and minimized the use of organic solvents by supercritical CO2 (Xie et al., 2016). Extraction is required to release the mycotoxins from the matrix and eliminate the effect of matrix and substances which can interfere with the detection of mycotoxins.
The next step is the purification of the extract that increases the specificity, sensitivity, and accuracy of the quantification. The most common two methods used for mycotoxins purifications are solid-phase extraction (SPE) and immunoaffinity columns (IAC) (Alshannaq and Yu, 2017). The SPE is a rapid and efficient method and involves the absorbents (octadecylsilyl (C18), amino-propyl (NH2), multi-walled carbon nanotubes (MWCNTs), and silica gel), which are packed in cartridges and absorbs the mycotoxins (Wang M. et al., 2016; Huertas-Perez et al., 2017; Jiang et al., 2018). SPE has been used for the purification of type A TCTs in rice, maize, and wheat (Dong et al., 2015) and ZEA in maize (Han et al., 2017). In the case of IAC, for selective mycotoxins detection, specific monoclonal antibodies are used, and the target mycotoxin is bound by specific antibodies on the column. The mycotoxins are eluted from the column with pure acetonitrile or methanol (Liu et al., 2018). IAC was used in the analysis of OTA, ZEA, and AFs in wheat bran (Irakli et al., 2017), OTA, AFs, and Fusarium toxins in maize (Lattanzio et al., 2007) and cereals (Lattanzio et al., 2014).
Since the discovery of the mycotoxins, many different methods have been used to analyze the mycotoxins but the most common methods for mycotoxin analysis and detection include chromatographic techniques, immunoassays methods, or rapid strip screening tests (Zhang et al., 2018; Janik et al., 2021; Le et al., 2021). Methods belonging to the chromatographic group are aimed at quantitatively determine mycotoxins because of their potential to accurately detect, identify and quantify multiple toxins (Agriopoulou et al., 2020), and involve different types of chromatography: Liquid chromatography–tandem mass spectrometry (LC–MS/MS) or gas chromatography-tandem mass spectrometry (GC–MS/MS). Thin-layer chromatography (TLC) and high-performance liquid chromatography (HPLC) coupled to diode array, fluorescence, and UV-detectors (Zhang et al., 2018; Janik et al., 2021).
TLC is an effective method for qualitative and quantitative detection and analysis of many mycotoxins due to low costs, simplicity, and UV light fluorescent spots (Janik et al., 2021). But has low sensitivity and poor accuracy, which makes quantification very demanding (Singh and Mehta, 2020). The reliable separation capability of liquid chromatography (LC) coupled with the sensitivity of tandem mass spectrometry has positioned it as an effective analytical technique for the simultaneous detection and quantification of mycotoxins in a range of matrices (Adebo et al., 2018; Gbashi et al., 2019). The LC technique have been developed to overcome the constraints of the TLC method and it is applied for the detection of non-volatiles, high polarity, and thermally labile mycotoxins (Singh and Mehta, 2020; Yang et al., 2020). HPLC with different adsorbents and detectors viz. fluorescent (FLD) and UV visible (UV) depends on the presence of chromophore used for mycotoxin analysis. Sometimes directly detected in HPLC-FLD using natural fluorescence (e.g., AFs and OTA) in rice (Zinedine et al., 2007).
The MS/MS combination with LC (LC–MS/MS) provides a good analytical tool with high sensitivity, selectivity, and reliability (Malachova et al., 2018; Pascale et al., 2019). LC–MS/MS method has been effectively used for the detection of mycotoxins in a mixture of spices and herbs (El Darra et al., 2019). LC–MS/MS-based multi methods have gained growing relevance, as they allow rapid detection or even quantification of multiple-mycotoxins in several food groups and feedstuffs (Zhang K. et al., 2017). In order to quantification of different mycotoxins (AFB1, B2, G1, G2, OTA, DON, ZEA, T-2 toxin, and HT-2 toxin) for grain legumes, LC–MS/MS was used (Kunz et al., 2020). Tebele et al. (2020) were detected 12 mycotoxins in wheat, maize, and sorghum, including α-zearalenol (α-ZEL), FB3, FB1, tenuazonic acid (TeA), OTB, DON, OTA, 3-acetyldeoxynivalenol, sterigmatocystin (STG), 15-acetyldeoxynivalenol, cyclopiazonic acid (CPA), and aflatoxin B2 using liquid chromatography–tandem mass spectrometry.
When the rapid detection of mycotoxin is required, immunological tools are used such as lateral flow immunoassay (LFIA; Wolf and Schweigert, 2018; Lattanzio et al., 2019) and enzyme-linked immunosorbent assay (ELISA; Hendrickson et al., 2018). Detection depends on explicit monoclonal and/or polyclonal antibodies developed against these toxins (Thanushree et al., 2019) Because of simplicity and cheapness coupled to sensitivity and selectivity, immunoassays are preferably employed for the first level screening and survey studies on mycotoxin contamination. ELISA-based kits are commercially available for all regulated mycotoxins and provide the most-used analytical tool for assuring food safety through the food chain (Pereira et al., 2014). Besides, immunochemical-based tests in diverse formats are continuously developed to provide rapid, portable, and easy to operate systems (Zhang et al., 2015). Among these, the immunochromatographic test (ICT) technology plays the lead role and has been widely applied for the visual yes/no detection of mycotoxins and for their semi-quantification (Majdinasaba et al., 2015).
In recent years, biosensors such as electrochemical, optical, and piezoelectric have also been used to identify mycotoxins in foodstuffs (Younis et al., 2020). Biosensors have been classified as surface acoustic wave and optical-based on the operational principle. Multi-mycotoxins have been analysed in a single step by immunochemical biosensors (Magan, 2006). Surface Plasmon Resonance (SPR) and nanoparticle-based biosensor techniques are based on changes in refractive indices and called ‘label-free’ tools. SPR technique involves optical real-time detection of multiple analytes. A new fluorescence polarization method using near-infrared fluorescence sensors has also been developed that showed a great potential in fluorescence detection and measurement. Through this method, an antibody-binding fluorescence-labelled PT derivative exhibits an increase in fluorescence polarization emissions (Melinte et al., 2022). This method can detect PT in food commodities ranges from 6 to 102 μg/L (Melinte et al., 2022). This technique relied on the unique properties of crystals or quartz. Zhang W. et al. (2017) have developed a nano-sensor that discriminates mycotoxins selectively using manganese-doped ZnS quantum dots. Researchers have been focusing on restoring biosensor activity after use in recent years. For example, Soldatkin et al. (2017) pursued inhibitory action of PT, and developed a conduct metric urease-based biosensor. Because of its high PT sensitivity, strong selectivity, and high signal repeatability, this biosensor can be used to assess PT concentrations in apple juice beyond 50 g/L. As a technique to enhance biosensor signal transmission, lanthanide-doped rare earth-doped up-conversion nanoparticles (UCNPs) have attracted much attention (Loo et al., 2019). It offers several advantages over conventional down-conversion luminescent devices to use NIR-to-visible up-conversion nanoparticles (UCNPs). This technology has a low autofluorescence background which improves the signal-to-noise ratio, low toxicity, high photostability, tunable wavelength, high Stokes offsets, and deep tissue infiltration. Mycotoxins are detected in food using biosensors linked to transducers that use enzymes, aptamers, and antibodies as tools for recognition. Due to their remarkable ability to recognize mycotoxins at low dosages and modify their absorption properties, aptamers are becoming increasingly popular. Food industry applications can use them for on-line control of mycotoxins including PT. The main challenges of biosensors are the limited stability of the bio-recognition component (which affects the long-term storage stability of the biosensors), poor selectivity, particularly in enzyme inhibition-based biosensors, and the high cost of antibodies compared to synthetic recognition elements (Burcu Aydın et al., 2020).
Another emerging field for the detection of mycotoxins is nanotechnology. For detection of multiple mycotoxins, the analyte is labeled with probes such as nano-silver, graphene, and magnetic nanoparticles (Zinedine et al., 2006). The electronic nose is a rapid method based on aromas and ordour of the food. Fungal contamination of foods formed mycotoxins implies volatile by-products and characterized by GC–MS and correlated with fungal activity (Yao et al., 2015). In addition, recently non-destructive tools such as machine vision systems have emerged for the detection of toxins and identification of fungal contamination in food materials (Vithu and Moses, 2016). The combination of spectroscopy and NIR hyper-spectral imaging works in the wavelength range between 700 and 2,500 nm to differentiate between beneficial and fungal contaminated samples and detects AFs and OTA in foods (Senthilkumar et al., 2016).
In addition to the standard methods described above, several other methods have been developed such as electronic nose, fluorescent polarization, aggregation-induced emission, and molecularly imprinted polymers. The electronic nose (e-nose) was based on the chemical changes in the volatile organic compounds (VOCs) caused by the fungal species and observed the correlation between mycotoxin concentration and VOCs in food commodities (Ottoboni et al., 2018). The e-nose has been used for the detection of FBs and AFs in maize (Ottoboni et al., 2018), DON in durum wheat (Lippolis et al., 2014), and wheat bran (Lippolis et al., 2018). Fluorescent Polarization (FP) has been used in the detection of mycotoxins in various foodstuffs, such as ZEA in maize (Zhang K. et al., 2017), AFB1 in maize (Zhang et al., 2018), and OTA in rice (Huang X. et al., 2020). The Aggregation-Induced Emission (AIE) dye-based apta sensor has been successfully developed for OTA detection in wine and coffee (Zhu et al., 2019) and AFB1 in peanut oil and broad bean sauce (Xia et al., 2018). During molecular imprinting, cross-linked polymers are formed between functional monomer and template as mycotoxins (Krska et al., 2005). Molecularly Imprinted Polymers (MIPs) have been developed for the detection of AFB1 in wheat (Guo et al., 2019) and ZEA in cereals (Huang Z. et al., 2020).
There are various mycotoxigenic fungi that cause contamination of agricultural commodities, which are important food and feed sources. The rapid evolution of agricultural technology has contaminated food products directly like pulses, cereals, vegetables, and fruits with mycotoxins. Tropical regions seem to have a greater problem than temperate regions, but no place in the world can be considered mycotoxin-free. This may be related to the transportation of various foodstuffs from one country to another. In particular, following foodstuffs have been discovered to contain mycotoxins naturally. The detection levels of mycotoxins in various food commodities are also documented in Table 4.
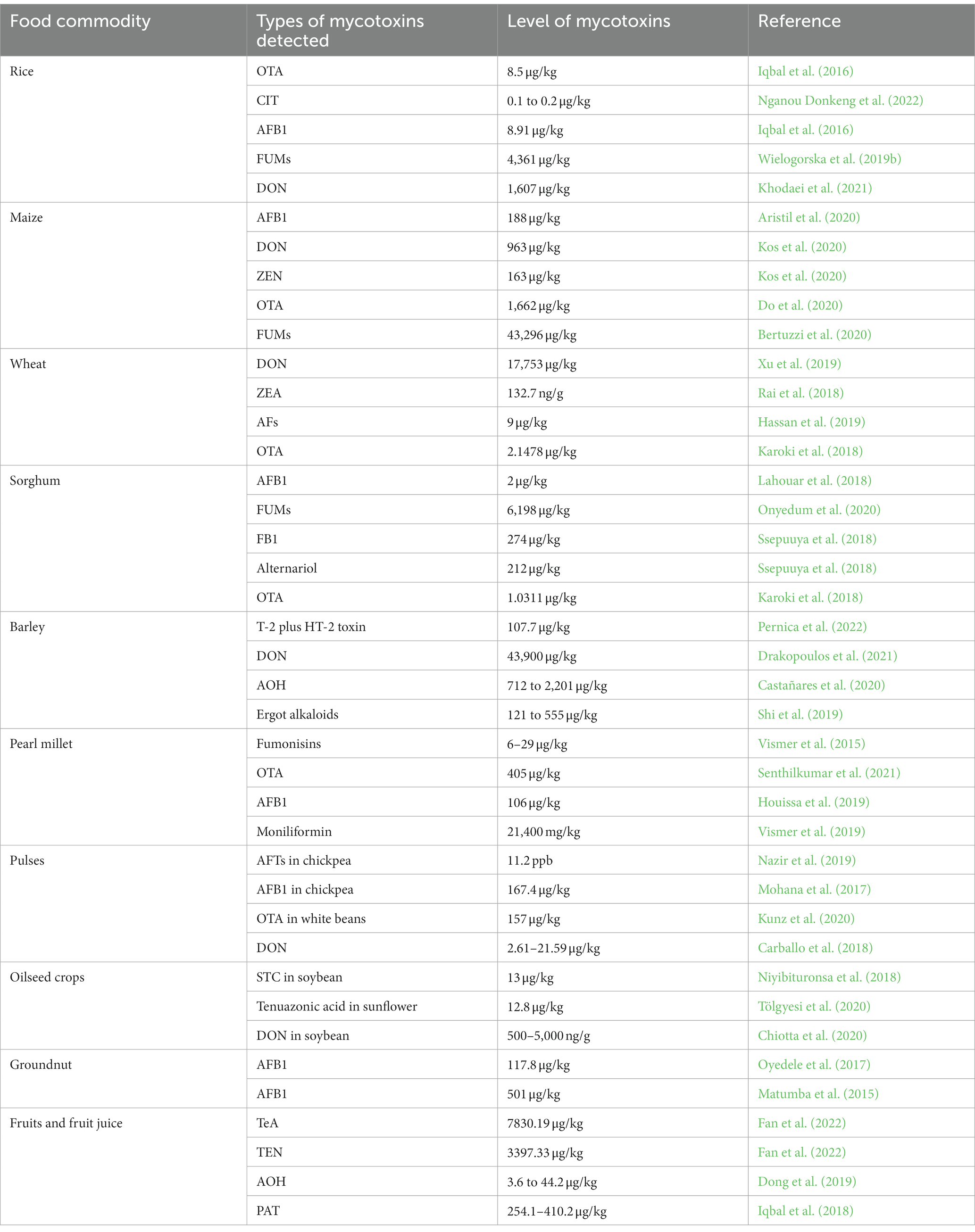
Table 4. Detection levels of mycotoxins in major food commodities.
Rice (paddy) is one of the most important food crops worldwide. Mycotoxin contaminations are greatly influenced the production of rice by the climate and storage conditions. Mycotoxigenic fungi in rice are believed to be species of Aspergillus, Fusarium, and Penicillium. As a result of these fungal infections, grains and glumes discolor, lose viability, quality, and become contaminated with toxins. In rice, presence of AFs, OTA, cyclopiazonic acid, FBs, TCT, ZEA, DON, CT, gliotoxin, and STC have been reported by several investigators (Gummert et al., 2009; Reddy et al., 2009a; Siruguri et al., 2012). Date back, Jayaraman and Kalyanasundaram (1990) found that AFB1 content of parboiled rice bran and rice bran samples was up to 35%. According to a study conducted in 1990, rice samples contained more AFs (184 to 2,830 μg/kg) than wheat and maize samples (Pande et al., 1990). In 36 samples of de-husked brown rice, Liu et al. (2006) found AFB1, AFB2, AFG1, and AFG2 which levels ranged from 0.99 to 3.87 μg/kg. Researchers in India have found 67.8% AFB1-positive rice and paddy samples out of the total samples examined, with toxin levels ranged from 0.5 to 38.5 μg/kg (Reddy et al., 2009a). Further, samples collected from six districts in Punjab by Siruguri et al. (2012) revealed that PAU 201 variety of stored rice had AFB1 levels below 15 μg/kg.
In Nigeria, out of 196 samples that were collected, AFs were detected in 97 samples (AFs levels ranged between 24 and 1,164 μg/kg), OTA in 56 samples (20 and 1,642 μg/kg), and ZEA in 93 samples (24 and 1,169 μg/kg; Hussaini et al., 2009a). Later, Iqbal et al. (2016) investigated that out of 62 samples analysed in Pakistan, 37% of the samples contained OTA in the range of 0.6 to 25.4 μg/kg and AFB1 in the range of 0.04 to 21.3 μg/kg. Likewise, Majeed et al. (2013) also found contamination of 50% of rice samples examined with OTA (0.06 to 15 μg/kg) and AFs (0.05 to 24 μg/kg). Other studies have found multiple mycotoxins in Turkish, Nigerian, Indian, and Chinese rice (Reddy et al., 2009b; Aydin et al., 2011; Makun et al., 2011). For examples, Makun et al. (2011) found that 14% of the samples were positive for FB1 with a level of mean concentration 0.2 μg/kg. A study conducted by Majeed et al. (2018) reported 23 mycotoxins in 180 rice samples with AFs, FBs, and ZEA were prevalent toxins reported in 20–56% of the samples, whereas OTA, HT-2 toxins and DON were reported in few samples. The concentration of these toxins was ranged in between 0.61 and 22.98 μg/kg.
In pre-and post-harvest conditions, corn is commonly colonized by spoilage fungi, whose relative abundance depends on both biotic and abiotic stresses that affect the mycotoxin production rate. ZEA, FBs and TCT were the most prevalent mycotoxins contaminating maize in Mediterranean countries (Jestoi, 2008), whereas AFs were common in tropical and subtropical regions (Muriuki and Siboe, 1995). Maize samples from the Mediterranean basin have been reported to contain different types of mycotoxins including FBs and AFs (Marin et al., 2012). A study conducted in Italy over 1995–1999 revealed that two maize samples had AFs levels of as much as 109 to 158 μg AFB1/kg, with the mean range of 0.3 to 4.10 μg AFB1/kg seeds (Pietri et al., 2004). Samples from Turkey also showed high levels of AFs in the range of 120.3 to 133 μg/kg (Giray et al., 2009). Besides, a study conducted in Syria reported AFs levels >20 μg/kg (Majid, 2007).
Maize sold in West Africa also had high levels of AFs contamination, ranging from 0.4to 490 μg/g in Ghana, 0.2 to 120 μg/g in Benin, and 0.7 to 110 μg/g in Togo (James et al., 2007). Four out of every five maize samples from the Southern Guinea Savanna exceeded the AFs level of 20 μg/g recommended by international authorities. Out of 190 maize samples that were analysed for contamination of multiple mycotoxins in India, 69 (34.8%) samples were contaminated majorly with AFs (Janardhana et al., 1999). There was also a high level of AFs detected in stored maize grains (43%), and most of the contaminated samples had levels greater than 20 μg/kg. The samples from Egypt included 30 μg/kg of TAFs (Abdelhamid, 1990), from Southern Guinea contained 77 μg/kg (Hell et al., 2003), and the samples collected in Croatia had AFB1 ranged between 224 and 614 μg/kg levels during 1996 to 1997 (Jurjevic et al., 2002). However, Croatia had the highest percentage of OTA positive samples (25%) with more OTA concentration, i.e., 31.7 μg/kg (Segvic et al., 2009).
The FBs synthesized by Fusarium are the most common contaminants, with contamination incidence often near 100% despite the presence of these two mycotoxins. FBs were found in 100% of maize samples grown in Turkey, with a mean level of 8,240 μg/kg of toxin production (Oruc et al., 2006). It has been found that 54 French maize samples imported into the UK between 2004 and 2007 contained higher than 10 μg/kg levels of FBs contamination (Scudamore and Satel, 2009). According to a study of Pietri et al. (2004), samples contaminated with Fusarium-derived ZEA over 200 μg/kg increased to 53.8%, with a mean contamination of 453 μg/kg and a sample showed much higher level, i.e., 2,531 μg/kg ZEA. The dominant mycoflora of 12 maize samples collected from affected households were Fusarium, which produced FB1 at levels between 0.25 and 64.7 mg/kg (Bhatt et al., 1997). Additionally, FB1 was detected in 25 rain-affected maize samples in the range of 00.04 to 65 mg/kg and 89% of normal maize samples contained AFB1 in the range of 0.38 to 109 μg/kg. In Albania, Topi et al. (2021) analysed 45 maize samples and found that 78% of the samples were majorly contaminated with FB1 and FB2 with concentration ranging from 59.9 to 16,970 μg/kg. They also found that 31% of the maize samples exceeded the EU maximum permitted level of FBs, i.e., 4,000 μg/kg.
During transport and storage, wheat is also affected by a wide species of fungi. Fusarenon X-glucoside (FUXGlc), a Fusarium mycotoxin reported for the first time in wheat grains induced by Fusarium species (Nakagawa et al., 2011). They also reported the levels of OTA and DON were, respectively, 12 and 53 μg/kg in wheat. The T-2 and AFB1 toxins were also detected in wheat flour by Badiale-Furlong et al. (2003). An additional study found that FB1 levels ranged from 0.5 to 3.9 μg/kg of wheat and between 0.6 and 2.3 μg/kg of flour (Birck et al., 2003). There were 74% (24.91%) of wheat samples contaminated with DON from the southern region, and levels of toxins ranged from 603.2 to 850.4 μg/kg (Mallmann et al., 2003). Sulyok et al. (2006) determined the detection limits of DON in wheat samples which ranged from 0.03 to 220 μg/kg. It was reported that Fusarium species, particularly F. gramanearum on wheat kernels in five Kenyan districts produced ZEA and DON (Muthomi and Mutitu, 2003).
It has been shown that samples of wheat and wheat flour purchased from resident Egyptian markets contain Fusarium mycotoxins (Aziz et al., 1997). In five wheat samples, DON was detected at levels from 103 to 287 μg/kg, and in one flour sample and one bread sample, DON was detected at 188 and 170 μg/kg, respectively. A total of 10 wheat samples were found to contain 28 to 42 μg/kg range of ZEN levels and four samples of bread and flour contained 95 μg/kg and 34 μg/kg, respectively. There are certain Fusarium species like F. graminearum, F. avenaceum, and F. culmorum that are predominant in Fusarium head blight infected wheat and produce mycotoxin (Bottalico and Perrone, 2002). Besides, 41 wheat samples harvested and consumed in few regions of Turkey showed contamination of Aspergillus and A. parasiticus as well as production of AFs including B1, B2, G1, and G2 in the range of 10.6 to 643.5 μg/kg (Giray et al., 2007). In total, 59% of the samples were positive for TAFs, such as B1, B2, G1, and G2 with respective percent of 42, 12, 37, and 12%. Besides, some emerging mycotoxins have also been reported in wheat samples. For instance, F. tricinctum was found to be produced various ENNs such as ENN A, A1, B, and B1 in durum wheat (n = 260; at levels 55 to 596 μg/kg) and wheat (n = 470; at levels 47 to 142 μg/kg; Orlando et al., 2019). More recently, Topi et al. (2021) found that 23% of the wheat samples (n = 71) were found to be often contaminated with Fusarium toxin, i.e., DON. In their study, the EU maximum permitted level (1,250 μg/kg) was exceeded on one wheat sample which showed 1916 μg/kg DON concentration.
In certain conditions, other cereals like barley (Hordeum vulgare L.), sorghum (Sorghum vulgare L.), oats (Avena sativa L.), and millets (Pennisetum glaucum L.) are also found to be associated with mycotoxigenic fungi during transportation, storage, and even at the field. There is evidence that these grains serve as a suitable substrate for the development of AFs. Eighty-four (72%) of the 116 samples of oat and barley collected from eastern Canada were associated with DON with levels ranged 8 to 9 mg/kg (Campbell et al., 2000). There were 34% of 73 oat samples that contained DON, and 34% of the 53 barley samples, and 15% of the 26 oat samples that contained nivalenol. Samples of barley contaminated with DON had a maximum level of 15.8 mg/kg (Abramson et al., 1998). The concentration of FB1 in sorghum ranged from 0.14 to 7.8 μg/kg (Bhatt et al., 1997). In another study, FB1 (0.07–8 μg/kg) was found to occur naturally in Indian sorghum along with AFB1 (5 to 125 μg/kg) (Shetty and Bhatt, 1997). Besides, ZEA was also found in grains with moisture contents of 20–22% (Jurjevic et al., 2007). In Nigeria, Hussaini et al. (2009b) reported AFs in several sorghum samples. It was also reported that 92 μg/kg of moniliformin and 414.6 μg/kg of beauvericin were found in pearl millet collected from few regions of Africa and Asia (Wilson et al., 2006). Orlando et al. (2019) analysed 282 spring barley, 172 tricale and 56 winter barley samples from the French harvest of 2012 to 2014 and found that samples were associated with F. avenaceum and F. Poae. The ENNs contents produced by both Fusarium species varied between years but were constantly highest on triticale (levels 131 to 1,218 μg/kg) and spring barley (levels 199 to 1,316 μg/kg). Polišenská et al. (2020) also found frequent occurrence of ENNs, beauvericin, and nivalenol in hulless barley and HT-2 and T-2 toxins, beauvericin and ENNB in hulless oats which significantly reduced the quality of oats and barley and caused health risk. The DON and ZEA contents in barley (DON: 1250 μg/kg, ZEA: 100 μg/kg) and oats (DON: 1750 μg/kg, ZEA: 100 μg/kg) were far from being exceeded the limit fixed by EU [Regulation 1881/2006; European Commission (EU), 2006]. Likewise, Edwards (2007) reported another mycotoxin, i.e., DON in barley (1,416 μg/kg) and oat 282 (μg/kg) samples. Correspondingly, Schöneberg et al. (2016) found that frequency of occurrence of DON was higher in barley (57%) compared to oats (45%) with a much higher level of DON in barley (4,860 μg/kg) compared to oats (1,328 μg/kg). However, researchers have found lower content of ZEA (<10 μg/kg) in barley (Gil-Serna et al., 2013) and oat (Schöneberg et al., 2016).
Pulses provide dietary protein and other essential nutrients. However, improper storage and handling can lead to mycotoxins contamination. It has been reported that Aspergillus species produced 333 to 10,416 μg/kg of AFB1 in pulses and oilseeds crops (Begum and Samajpati, 2000). In Ontario, Canada, Tseng and Tu (1997) investigated the presence of FB1 in adzuki bean (Vigna angularis) and green gram (Vigna radiata) samples but not in the healthy samples. By HPLC, it was determined that the adzuki and mungbean samples contained respective 261 and 230 μg/g of FB1. Thus, a future study of mycotoxin contamination of various foods including legumes needs to be undertaken. The results of another study showed that out of 66 isolates of A. flavus from mustard seeds, 24 produced AFs (0.25 to 22 μg/ml), with eight isolates producing extremely high levels of AFs and the remaining 16 isolates producing very low amounts of AFs.
Mycotoxins are not produced by all fungi that infect food crops. In this regard, Ahmad and Sinha (2002) found that out of 34 isolates of F. moniliforme, only 13 isolates were able to produce ZEA (range 1.2 to 4.0 μg/ml) and of 12 isolates of P. citrinum, only four produced CT at levels ranging from 1.0 to 3.0 μg/ml (Ahmad and Sinha, 2002). South Africa’s cowpea cultivars contained 0.12 and 0.61 μg/g FB1, while cultivars from Benin were free of FBs (Kritzinger et al., 2002). A total FB concentration of 0.8 to 25.30 μg/g was determined in the pulses, and the highest level of FB1 was detected at 16.86 μg/g (Kritzinger et al., 2002). Two types of fungi, A. parasiticus isolated from beans seeds and F. moniliforme isolated from soybeans, produced significant concentrations of AFs (196.58 μg/kg) and FBs (198 mg/kg), respectively, as demonstrated by Embaby et al. (2013). In a recent study, 81 cowpea samples analysed in Nigeria were positive for AFs with concentrations ranged between 84 and 209 μg/kg (Afolabi et al., 2019). In addition, beauvericin was also detected in few samples.
Peanuts are considered the second most important legume worldwide after beans. Unfortunately, lack of storage technologies causes mycotoxins, especially AFs, to be released (Kaaya et al., 2006). Researchers have discovered AFs types B and G as well as cyclopiazonic acid (CPA) in peanuts grown in Argentina (Formosa province) by A. flavus (Pildain et al., 2004). There were 10 to 346 μg/kg of AFs in peanut cake samples, and OTA was below the limit of quantification (2 μg/kg) (Ediage et al., 2011). Researchers from Córdoba, Argentina, reported the presence of black species of Aspergillus in peanuts that produce OTA. A 0.5 μg/kg to 170 μg/g range of OTA was found in 32% of the seed samples analysed by Magnoli et al. (2006). OTA-producing isolates were found in 43 of 47 Aspergillus section Nigri isolates (27%) studied and overall, A. carbonarius presented in higher amounts (57%) as OTA producing fungi (Magnoli et al., 2006). As a result of a subsequent study, they discovered that 104 (32) of 322 isolates of Aspergillus section Nigri produce OTA, and that the quantities were between 2 μg/kg and 24 μg/ml (Magnoli et al., 2007).
Several peanut samples from India were found to contain levels of AFs >30 μg/kg in the study of Kishore et al. (2002). A disquieting AF amount of 852 μg/kg and ZEA of 98.1 to 847.3 μg/g were found in samples analysed in India. There was also evidence of mycotoxins in Brazilian groundnut seeds from sowing to harvest. In this context, Gonçalez et al. (2008) found that the levels of AFs (4.20 to 198.8 μg/kg) and CPA (260 to 600 μg/kg) were higher in 32% of peanut samples studied. The most common toxic compounds are those produced by Aspergillus species. In terms of mycotoxins considered by regulations, AFs exceeded the USDA maximum limit of 20 μg/kg in 90% of samples studied by Ezekiel et al. (2012). Similarly, groundnuts from Ethiopia had AF levels that ranged from 15 μg/kg to 11,900 μg/kg (Chala et al., 2013). In Nigeria, the content of TAFs in groundnut cakes, roasted groundnut, and boiled groundnut were respective 11.15, 4.50, and 1.51 μg/kg, suggesting that groundnut cake had the highest incidence and concentrations of TAFs (Adefolalu et al., 2021). They also found that contents of AFs increased with prolonged storage time.
During and after harvest, raw and processed horticulture products may contain mycotoxins. In addition to cereals, fruits and fruit juice may also pose a potential risk to toxin contamination. Aspergillus and Penicillium are two of the most recognized species worldwide that produce toxins in fruits. The dried fruits of vines (e.g., raisins, sultanas) contain higher OTA levels worldwide (Palumbo et al., 2011). According to reports, grapes contain potential amounts of AFs (0.3%), OTA (6.0%), PT (0.5%), and TCT (1.2%; Serra et al., 2005). Sixty samples of retail dried vine fruits collected from the United Kingdom revealed presence of OTA and AFs above 53.7 μg/kg (MacDonald et al., 1999). The majority of the reports indicated that most contaminated samples had an OTA level over 2 μg/kg, with maximum values exceeding 100 μg/kg (Magnoli et al., 2004; Aksoy et al., 2007). Chulze et al. (2006) found higher OTA levels in grape juice than allowed, i.e., <10 μg/kg (Chulze et al., 2006). In pear inoculated with fungus P. expansum, PAT was found to form and to diffuse in the apparently sound flesh, in a concentration exceeding the accepted European limits (50 μg/kg; Laidou et al., 2001). The occurrence of PAT (up to 80.5 mg/kg) was recorded in 89 percent of 351 rotten apple samples collected throughout Portugal, and A. carbonarius and T. roseum produced OTA and TCT in grapes before harvest time (Martins et al., 2002). Likewise, Mogensen et al. (2010) also found contamination of grapes with FB2 (171–7,841 μg) and FB4 (14–1,157 μg) produced by A. niger strains.
Besides, Jimenez and Mateo (1997) reported the occurrence of FB1, ZEA, DON, TCTs, T-2 and HT-2 toxins. In another study conducted in Egypt, OTA was present in all fig samples (60 to 20 μg/kg), apricot samples (50 μg/kg to 110 μg/kg), and plum samples (210 to 280 μg/kg) (Zohri and Abdel-Gawad, 1993). However, no sample was found to exceed the safe level of 50 μg PT/I. Based on the findings of Beretta et al. (2000), apple derivatives tend to represent a PT intake below the tolerable level of 0.4 μg/kg bw/day. The different apples tested by Tangni et al. (2003) showed PT at 79, 86, and 43%. A study conducted by Hussain et al. (2020) found more PT content in mango (110.9 μg/kg) than that of orange (6.3 μg/kg). The study also revealed that PT content was higher in mango fruits than in processed products like juice, jam, and pulp.
Besides, out of 70 mango fruits and 77 orange-based products analysed for occurrence of mycotoxins, they found that 29 mango samples (21.8%) and one orange sample (0.7%) exceeded regulatory limit (50 μg/kg) of PT in Pakistan. These observations are also in accordance with the study conducted by Iqbal et al. (2018) who found high level of PT contents in fruits and fruit juice with a maximum of 1,100 μg/kg in a red globe grapes sample. The higher PT content in apple juice was also reported in USA (2,700 μg/kg, Harris et al., 2009), Tunisia (889 μg/kg, Zouaoui et al., 2015), Turkey (1,416 μg/kg, Icli, 2019), and India (845 μg/kg, Saxena et al., 2008). On the contrary, in Argentina Oteiza et al. (2017) also found lower level of PT content in mango samples (6,415 μg/kg) than that of fruit juice samples (7,339 to 19,662 μg/kg). Studies conducted in Malaysia also did not find PAT in mango juice (Abu-Bakar et al., 2014; Lee et al., 2014). Apple juice analysed from Saudi Arabia attained PT levels in the range of 5 to 50 μg/kg (Al-Hazmi, 2010). Likewise, lower PT contents were reported in various fruits juice (apple, orange, grapes) analysed in Greece (10.54–5.57 μg/kg, Moukas et al., 2008) and South Korea (30.9 μg/kg, Cho et al., 2010).
In recent years, decontamination of mycotoxins in food commodities became crucial due to their toxic effects on people and animal health. Worldwide, numerous chemicals, physical, botanical, and biological strategies for detoxifying mycotoxins from contaminated foodstuffs have been explored (Table 5). However, among the many methods established, only a limited number of decontamination techniques found a practical application are reviewed below. The ideal characteristics of mycotoxins’ detoxification processes applied to food are summarized in Figures 1, 2. Novel technologies include the use of essential oils, natural extracts, irradiation, ozone treatment, pulsed light, cold plasma, and microbiological methods.
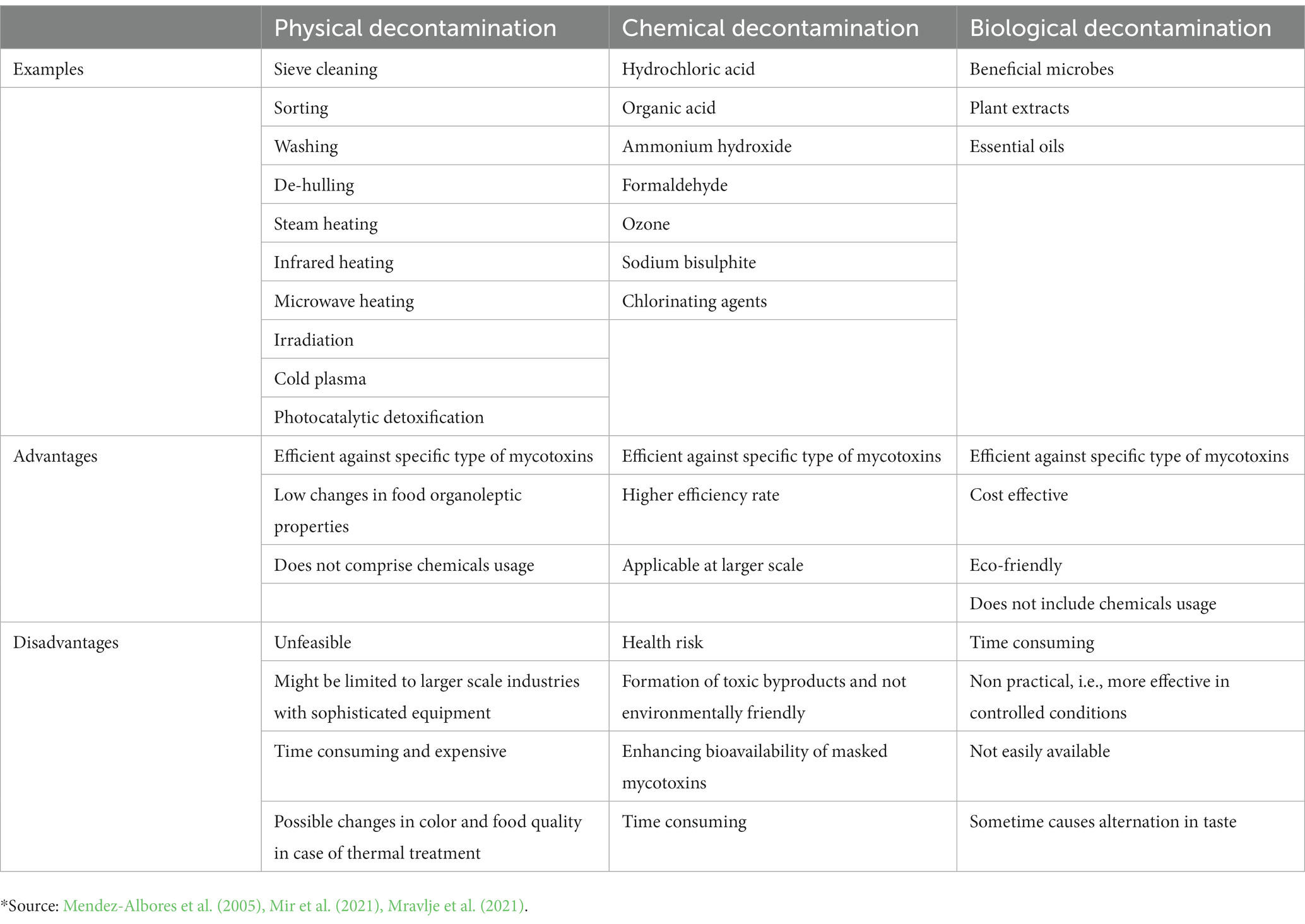
Table 5. Mycotoxins decontamination approaches in food commodities, their advantages, and disadvantages.*
Figure 1. Image showing major types of mycotoxins in food commodities and their various decontamination strategies.

Figure 2. Ideal characteristics of mycotoxins’ detoxification processes applied to food commodities.
The best approach to avoid contamination of food commodities by mycotoxins is to prevent this contamination rather than using detoxification or decontamination procedures. In order to prevent contamination, proper agricultural practices are required. However, in many cases fungal growth cannot be avoided as consequently leads to the formation of mycotoxins. Two of the conventional techniques to reduce mycotoxins levels are cleaning and milling or dehulling. Milling or dehulling is particularly applied to cereal grains and allows to remove the outer layer of the grains, which concentrated the higher mycotoxins level. Although these strategies allow to reduce contamination, if the contamination affects the milled grains, it cannot be controlled just with the milling process. The mechanism of detoxification of AFB1 by citric acid leads to the formation of the beta-keto acid structure, catalysed by an acid, followed by hydrolysis of the lactone ring with formation of AFD1, which has lower mutagenic activity (Mendez-Albores et al., 2005). The use of lactic acid also results into the conversion of AFB1 into AFD1.
Other methods include the use of chemicals such as hydrogen peroxide and ozonated water, and propionic acid. The ozone treatment is effective in the elimination of mycotoxins and it does not leave toxic residues. Besides, it is environmentally friendly and minimally affects the quality of foods. However, it can originate the oxidation of lipids, degradation of bioactive compounds and it has high costs associated (Mir et al., 2021). A study carried out by Trombete et al. (2017) applied ozone at a concentration of 60 mg/L for 300 min in wheat and allowed to reduce DON and AFs at a reduction rate of 64 and 48%, respectively. Savi et al. (2014, 2015) applied ozone gas at a concentration of 60 μmol/mol for 180 min to wheat samples and could effectively reduce mycotoxins contamination: 94.6% reduction rate in the case of AFB1, 84.5% for AFB2, 80% for AFG1, 81% for AFG2 and 100% for DON. However, these chemical methods present the drawback of leaving residues in foods and some have still limited effect, therefore the need for newer decontamination methods has boosted novel technologies. Additionally, their effectiveness is not broad spectrum and depends on the dose and target mycotoxigenic fungal isolates, and results have been found inconsistent when applied in the field/ or industry. Besides, more in vivo studies are required on the activity, bioavailability, and toxicity.
One of the technologies used to decontaminate food commodities from mycotoxins is irradiation, which uses ionizing energy. Irradiation is effective and a non-thermal method, however, can originate lipids or vitamins oxidation, off-flavors or changes in foods’ color (Mir et al., 2021). Gamma irradiation has been applied to different cereals (e.g., maize, wheat, barley, rice) to reduce mainly AFB1 (Aziz et al., 2004; Aquino et al., 2005; Mohamed et al., 2015) but also AFB2 (Aquino et al., 2005), ZEA (Aziz et al., 2004), OTA (Aziz et al., 2004), and FB1 (Aziz et al., 2007) in doses ranging from 0.5 to 10 kGy. The degradation of mycotoxins depends on the dose of gamma irradiation, but also on the moisture level of samples. Mehrez et al. (2016) reported that the degradation of mycotoxins in a cereals sample irradiated with 8 kGy and having a moisture content of 16% was significantly higher than when the sample have a moisture content of 11%. However, sometimes it may not kill all targeted mycotoxin-producing fungi. The environment where objects are being irradiated can be very harmful – standing there could lead to injury and mutation of people’s cells (Mir et al., 2021).
The pulsed light technology is innovative and promising. Moreover, it does not leave residues in food commodities, and it is cost-effective, but only decontaminates the foods surface and reduces the germination of seeds (Mir et al., 2021). This treatment is influenced by factors related with food (type of food matrix, thickness, composition), mycotoxins (type of mycotoxins) and pulse light (power of pulses, number of pulses, distance between lamp and light). Wang B. et al. (2016) has reported a reduction of 75% of AFB1 and 39% AFB2 in rice treated with 0.52 J/cm2/pulse for 80s. Moreover, Chen et al. (2019) reported a reduction of 35.5% of DON in barley treated with 180 pulses for 60s. Pulsed light has also been used in combination with citric acid to degrade AFs in peanuts (groundnuts; Abuagela et al., 2019). The technology has a low penetration power, which is one of its main disadvantages. It is therefore initially proposed to use pulse light technology for surface decontamination of solid food commodities, unpackaged or packaged in UV-transparent materials. It is crucial to consider food surface topography in these applications.
Cold plasma is a novel and non-thermal technology in which reactive species (e.g., O, O3, OH, NO, NO2) generated by cold plasma degrade mycotoxins and convert them in other less toxic compounds. This procedure presents several advantages such as the maintenance of the quality of foods (Mir et al., 2021). Wielogorska et al. (2019a) reduced 65% of AFB1 and 64% FB1 in maize after application of cold plasma (10 min, 20 Hz). According to Mravlje et al. (2021), the frequency of Alternaria and Epicoccum fungi (in common and tartarian buckwheat, respectively) were significantly reduced after treatment with cold plasma for 90 and 120 s. One study found a twenty-minute direct high voltage atmospheric cold plasma treatment of 100 μg of DON in 1 ml of aqueous suspension reduced DON structure by more than 99% and improved Caco-2 cell viability by over 80%. The same treatment on 100 μg of powdered DON toxin, however, only reduced DON levels by 33% and resulted in a 15% increase in cell viability (Ott et al., 2021). A study by Chen et al. (2022) showed that after 8 min of exposure at 50 kV in air atmosphere, 83.99% of the DON solid standard was degraded in wheat samples. After degradation, there were no noticeable effects on wheat quality other than a slight decrease in the whiteness of wheat powder. Therefore, cold plasma may be a promising strategy for mycotoxins decontaminant in food commodities. These technologies, however, have several limitations, such as lipid oxidation, protein oxidation, food discoloration, and changes in organoleptic properties, which limit their application in food industries (Olatunde et al., 2021).
Biological methods are environmentally friendly and do not leave toxic residues, but it is difficult to select nontoxigenic bio-competitive microorganisms and they require longer period of time for the detoxification (Mir et al., 2021).
Different microbials (e.g., bacteria, moulds, yeasts) and enzymes can be used to inactivate or degrade mycotoxins, allowing obtaining fewer toxic compounds. Some of the microorganisms that have already been used to decontaminate foods include lactic acid bacteria, Bacillus licheniforms, B. subtilis, and Saccharomyces cerevisiae. The microorganisms used for mycotoxin detoxification processes can be applied at any stage (pre or postharvest) and must show to be efficient and cost effective. Ansari et al. (2015) has used kefir-grains to reduce the contamination of pistachio with AFB1 in 96.8% while Farzaneh et al. (2012) used Bacillus subtilis UTBSP1 to obtain the reduction of AFB1 in the same matrix with an effectiveness of 95%. However, further investigations are needed to determine the safety and mechanisms of beneficial microbes against mycotoxin-producing fungi.
Mycoflora incursion on agriculturally important food commodities has been managed with several synthetic fungicides. To date, physical and chemical methods to reduce or remove mycotoxins from food have not proven to be satisfactory or useful. These toxins have negative effects on the food chain and can thus be eliminated with botanical-based detoxification (Gurney et al., 2014). In addition to the controversy surrounding synthetic preservatives, users have become more interested in natural food protection for improving the food quality and shelf life and protecting them from biodegradation by mycotoxigenic microbes (Pandey et al., 2016). In the academic and industrial communities, aromatic and medicinal plants are being studied for their antifungal effects against mycotoxigenic fungi, because of their eco-friendly and safety concern. Natural extracts are constituted of bioactive compounds (e.g., phenolics, alkaloids, terpenes) with anti-mycotoxigenic activity. Sultana et al. (2015) reported that the extract from neem leaves allowed reducing the contamination of cereals by AFs during storage. However, this extract presented a strong aroma which restricted its use. Ponzilacqua et al. (2019) reported the in vitro mycotoxins decontamination effect of Brazilian medicinal herbs extracts. These authors reported a time dependent AFB1 reduction by araçá, sweet passion fruit, oregano, and rosemary but no degradation on OTA or ZEA by any of the tested extracts were reported. The highest AFB1 reduction (approximately 60%) was found for rosemary extract after 48 h, followed by araçá and oregano extracts. Iram et al. (2016) compared the capacity of Cassia fistula and Ocimum basilicum aqueous extracts to decontaminate pistachio by AFB1 and AFB2. These authors reported that O. basilicum leaves extract was able to degrade 90.4% of AFB1 and 88.6% of AFB2, while the extracts of O. basilicum twigs, C. fistula leaves and twig were less efficient. Due to the search for more “natural” additives and agents, natural extracts from different parts of the plants (e.g., leaves, fruits, roots), are very promising for mycotoxins decontamination of foods. However natural extracts present high variability in their composition due to the influence of factors such as the selected cultivar, part of the plant used and edaphoclimatic conditions (Mateus et al., 2021). Therefore, there is the need of establishing guidelines for the standardization of these extracts in order to better control their effectiveness, in particular as anti-mycotoxigenic agents.
In addition, EO-based detoxification represents a promising substitute to eliminate mycotoxins and improve food and feed quality. Essential oils are produced by secondary metabolism of higher plants and are complex mixtures of volatile organic molecules. Hydro-distillation, low-pressure distillation, and high-pressure distillation are all methods of obtaining these essential oils. Essential oils are composed largely of terpenes and terpenoids followed by aromatic and aliphatic compounds. There are numerous structural variations of terpenes, which include monoterpenes, sesquiterpenes, diterpenes, hemiterpenes, triterpenes, and tetraterpenes. In this article, EOs that have been used against mycotoxigenic fungi or to reduce mycotoxin levels including AFs, ZEA, OTA, and FBs in food products are discussed.
Many researchers have suggested the use of EOs to reduce the growth of fungi and against the production of AFs by A. flavus and A. parasiticus (Maraqa et al., 2007; El-Nagerabi et al., 2012). Many food-borne fungi produce AFs that are inhibited by EOs and flavonoids (Alpsoy, 2010). Azadirachta indica seeds extract completely inhibited AFs production level in maize at concentrations of 500 and 1,000 mg/kg, while Morinda lucida seeds extract inhibited AFs production level at 1000 mg/kg concentration (Bankole, 1997). Iranian medicinal plant derived EOs also act as novel antioxidants and AFs inhibitors in food systems. As an example, Satureja hortensis and its active ingredients were found to act as effective inhibitors of AFs produced by A. parasiticus. The IC50 values for carvacrol and thymol for AFB1 and AFG1 were 0.50 and 0.06 mM, respectively. Carvacrol and thymol were found to be the potent constituents of S. hortensis and they may be used to reduce AFs contamination in food commodities (Razzaghi-Abyaneh et al., 2007). The EOs from Adansonia species has also inhibited the levels of TAFs and AFB1 secretion by A. flavus (47.2 to 95.7%; 28.1 to 89.7%) and A. parasiticus (42.7 to 93.3%; 25.9 to 80.2%; El-Nagerabi et al., 2013). NKD-208 isolates of AF-producing A. flavus were strongly inhibited by the Callistemon lanceolatus EO (Shukla et al., 2012). The EO of Zataria multiflora at 150 mg/kg reduced AFs production level up to 99.4% (Gandomi et al., 2009). Similarly, El-Nagerabi et al. (2012) also found that Nigella sativa EO caused potential reduction of AFB1 level by inhibiting the growth of A. flavus and A. parasiticus. Another EO, i.e., Ageratum conyzoides EO was found to inhibit AFs production level at 2.0 μl/ml produced by A. parasiticus (Ab2242) and at 1.5 μl/ml by A. flavus (La3228; Adjou et al., 2012). Marjoram and clary sage EOs significantly reduce the growth of A. parasiticus (Gomori et al., 2013). The 10 μl dose of A. conyzoides EO was more effective than Origanum vulgare in inhibition of AFs in soybeans (Esper et al., 2014). EOs derived from Cymbopogon martini, Foeniculum vulgare, and Trachyspermum ammi were also found to reduce various types of toxin production by A. niger and A. flavus at respective dose of 0.5 and 0.75 μl/ml (Gemeda et al., 2014). Similarly, EOs from Artemisia nilagirica (Sonker et al., 2014b) and Cymbopogon citratus (Sonker et al., 2014a) were found to inhibit AFs and OTA production levels in grapes at respective dosage 0.8 μl/ml and 1.6 μl/ml. Similar dose of EO from L. alba has been found to control AFB1 production level in green gram (Pandey et al., 2016). Likewise, in various food commodities, the level of AFs inhibition has been reported due to Rosmarinus officinalis and Trachyspermum copticum (450 mg/kg) EOs by Rasooli et al. (2008), Thymus eriocalyx and TX-porlock EOs (250 mg/kg) by Rasooli and Owlia (2005), oil derived from Turmeric leaves (95.3 and 100% inhibition) by Sindhu et al. (2011), and Ocimum gratissimum EO by Prakash et al. (2011).
The search of literature revealed that little work has been done on efficiency of EOs on reduction of ZEA levels in food commodities. EOs of palmarosa, cinnamon, clove, lemongrass, and oregano have been found effective in reduction of ZEA and DON levels. ZEA was found to be produced by F. graminearum in naturally contaminated maize grains, while in control sets levels of both toxins determined >500 mg/kg (Marin et al., 2004). Likewise, another study also found that clove and palmarosa EOs proved as better grains’ protector for maize grains as both EOs reduced ZEA and DON production levels as well as growth rate of F. graminearum under variable environmental conditions (Velluti et al., 2004).
EOs are also found potential in detoxification of FBs. Researchers have found that EOs from cinnamon, lemongrass, palmarose, clove, and oregano inhibited FB1 production level in maize grains (Velluti et al., 2003). Lopez et al. (2004) evaluated Aloysiatriphylla, A. polystachya, Origanum vulgare, and Mentha piperita EOs and found that among these EOs, O. vulgare EO significantly reduced FB1 level produced by F. verticillioides, whereas EO from A. triphyla was found to increase the mycotoxin levels at lower doses. Relatively, both O. vulgare and A. triphylla EOs at 250 and 500 μl/ml had efficient efficacy against FB-producing F. verticillioides (Lopez et al., 2004). In another study, cinnamon oil was also found to inhibit FB production and mycelia growth of F. culmorum and F. graminearum at 500 μg/g doses (Hope et al., 2005). Besides, the EO of Zingiber officinale had the inhibitory effect on FB1 and FB2 production at respective doses 4,000 and 2,000 μg/ml (Yamamoto-Ribeiro et al., 2013).
In the study of Xing et al. (2014), the EOs from Litsea cubeba, cinnamon, spearmint, citral, anise, clove, camphor, and eucalyptus inhibited levels of FB1 alongwith F. proliferatum growth. Among these EOs, they reported cinnamon as a most effective EO followed by citral, eugenol, eucalyptus, anise and camphor. As Cinnamon EO was found most potential, it reduced FB1 level from 15.03 to 0.89 μg/ml at a concentration of 280 μg/ml. In addition to EOs, their active constituents have also been found potential in reduction of mycotoxin level. For example, menthol, limonene, thymol, and menthone with thymol was the most active in reduction of FB1 level and its biosynthesis by F. Verticillioides at 75 mg/kg (Dambolena et al., 2008).
From time-to-time investigations were carried out to control OTAs contamination and related fungi using plant EOs. Both oregano and mint EOs were able to inhibit OTA production levels by A. ochraceus (Basílico and Basílico, 1999; Soliman and Badeaa, 2002). The EOs from Clove leaf, bay leaf, and cinnamon at 50 ppm caused reduction of OTA levels in wheat substrate and a complete inhibition of this toxin was found at 500 ppm doses (Cairns and Magan, 2003). Murthy et al. (2009) found that the EO from Plectranthus amboinicus was completely reduced OTA levels in foods as well as toxigenic strains of A. ochraceus at 500 mg/kg. Furthermore, the use of 100 mg/kg of this EOs in food samples such as maize, groundnut, and poultry feed inhibited the growth of A. ochraceus (Murthy et al., 2009). The EO from A. framomumdanielli also possessed ochratoxigenic activity in cocoa bean as it decreased OTA contents from 500 to 2,000 mg/kg (Aroyeun et al., 2009). Likewise, 0.10% of basil EOs reduced OTA production level from 135 to 98 μg/ml in study of Mohamed et al. (2012). Besides, EOs from both Artemisia nilagirica (Sonker et al., 2014b) and Cymbopogon citratus (Sonker et al., 2014a) were found to be completely reduced the levels of OTA in table grapes at respective doses 0.8 and 1.6 μl/ml.
The examples given here are just a few out of the many EOs that have been used to reduce mycotoxin contamination in food commodities, many of which cannot be covered in this short review. Thus, these EOs can be used for the detoxification and reduction of AFs, FBs, ZEA, and OTA levels as well as related fungi in food commodities. Food commodities can be protected from microbial contamination by encapsulating these potential EOs either as nanoemulsions or nanoparticles or packaging films. Furthermore, encapsulation enhances the oxidative stability, thermostability, shelf life, and biological activity of EOs. Additionally, it can be helpful in controlling the volatility and release properties of essential oils. However, more in vivo studies are required on the efficacy at larger scale storage conditions, bioavailability, and toxicity. Further studies are also needed to determine the safety and mechanisms of these EOs depending on mycotoxin-producing fungi as well as their potential health concern.
Increasing population requires the need to stockpile bulky foodstuffs for use in near future. But improper storage of such foodstuffs leads to the colonization by mycotoxigenic fungi. This results in contamination of food commodities by multiple mycotoxins, and this is a global concern. In this review, we describe mycotoxins in various food commodities, including their prevalence, toxicity, regulation, and detoxification. Mycotoxin detoxification in foods is a significant challenge due to the complexity of few food matrixes like fruit juice, edible oils and the relatively low content of mycotoxin. Researchers worldwide have used a variety of detection methods with varying levels of sensitivity. Sample preparation techniques, mycotoxin characteristics, food matrix type, and detection methods determine the lower limit of detection. The use of chromatographic techniques for the analysis of mycotoxin in rice, wheat, oil-seed crops, peanuts, etc. has been described in various papers (Majeed et al., 2018). Aptamer and surface plasmon resonance are novel detection methods with high affinities and specificities and low detection limits, but only in a few food commodities like vegetable oils (Abdolmaleki et al., 2021). This shows that these methods have many potential applications for the future. In future research, their application to the detection of mycotoxins in other commodities will be important.
To eliminate or decrease levels of mycotoxin in food commodities below consumption levels, chemical, physical, and biological methods were used. During decontamination, food commodities’ organoleptic properties and nutritional value should not be affected, and toxic degradation products should not be produced. Decontaminating mycotoxins efficiently poses a challenge in this regard. On the hand, controlling mycotoxigenic fungi with synthetic chemicals may cause a serious concern both at anthropogenic and ecological level. In this regard, eco-friendly mycotoxin management using plant’s extract and EOs would be safer to the user and the environment. Use of such methodologies could be cost-effective and can be one of the aims of sustainable agriculture. Lastly, before application, safety issues of botanicals should be fully addressed in food commodities. Therefore, more research is required in order to standardize the quality of the natural extracts, evaluate their safety, and to conclude about the most effective moment of application as well as the most effective concentration for different food commodities.
AP: original draft preparation. MKS: wrote detection techniques from significant inputs from AP, AK, AS, and NKD: review and edit. All authors contributed to the article and approved the submitted version.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abbas, H. K., Zablotowicz, R. M., and Bruns, H. A. (2008). Modelling the colonization of maize by toxigenic and non-toxigenic Aspergillus flavus strains: implications for biological control. World Mycotoxin J. 1, 333–340. doi: 10.3920/WMJ2008.x036
Abdelhamid, A. A. (1990). Occurrence of some mycotoxins (aflatoxins, ochratoxin A, citrinin, zearalenone and vomitoxin) in various Egyptian feeds. Arch. Anim. Nutr. 40, 647–664.
Abdolmaleki, K., Khedri, S., Alizadeh, L., and Javanmardi, F. (2021). The mycotoxins in edible oils: an overview of prevalence, concentration, toxicity, detection and decontamination techniques. Trends Food Sci. Technol. 115, 500–511. doi: 10.1016/j.tifs.2021.06.057
Abramson, D., Clear, R. M., Usleber, E., Gessler, R., Nowicki, T. W., and Martlbauer, E. (1998). Fusarium species and 8 keto-trichothecene mycotoxins in Manitoba barley. Cereal Chem. 75, 137–141.
Abuagela, M. O., Iqdiam, B. M., Mostafa, H., Marshall, S. M., Yagiz, Y., Marshall, M. R., et al. (2019). Combined effects of citric acid and pulsed light treatments to degrade B-aflatoxins in peanut. Food Bioprod. Process. 117, 396–403. doi: 10.1016/j.fbp.2019.08.011
Abu-Bakar, N. B., Makahleh, A., and Saad, B. (2014). Vortex-assisted liquid-liquid microextraction coupled with high performance liquid chromatography for the determination of furfurals and patulin in fruit juices. Talanta 120, 47–54. doi: 10.1016/j.talanta.2013.11.081
Adebo, O. A., Njobeh, P. B., Adebiyi, J. A., and Kayitesi, E. (2018). Coinfluence of fermentation time and temperature on physicochemical properties, bioactive components and microstructure of ting (a Southern African food) from whole grain sorghum. Food Biosci. 25, 118–127. doi: 10.1016/j.fbio.2018.08.007
Adefolalu, F. S., Apeh, D. O., Salubuyi, S. B., Galadima, M., Agbo, A. O., Makun, H. A., et al. (2021). Quantitative appraisal of total aflatoxin in ready-to-eat groundnut in north-Central Nigeria. J. Chem. Risk 12, 25–31. doi: 10.22034/JCHR.2021.1911495.1196
Adeyeye, S. A. (2016). Fungal mycotoxins in foods: A review. Cogent Food Agric. 2:1213127. doi: 10.1080/23311932.2016.1213127
Adjou, E. S., Dahouenon-Ahoussi, E., Degnon, R., Soumanou, M. M., and Sohounhloue, D. C. K. (2012). Investigations on bioactivity of essential oil of Ageratum conyzoides L., from Benin against the growth of fungi and aflatoxin production. Int. J. Pharma Sci. Rev. Res. 13, 143–148.
Afolabi, C. G., Ezekiel, C. N., Ogunbiyi, A. E., Oluwadairo, O. J., Sulyok, M., and Krska, R. (2019). Fungi and mycotoxins in cowpea (Vigna unguiculata L) on Nigerian markets. Food Addit. Contam. Part B Surv. 13, 52–58. doi: 10.1080/19393210.2019.1690590
Agriopoulou, S., Stamatelopoulou, E., and Varzakas, T. (2020). Advances in analysis and detection of major mycotoxins in foods. Foods 9:518. doi: 10.3390/foods9040518
Ahmad, M. S., and Sinha, K. K. (2002). Mycotoxin contamination of mustard seeds during storage. Indian Phytopath. 55, 299–302.
Aiko, V., and Mehta, A. (2015). Occurrence, detection and detoxification of mycotoxins. J. Biosci. 40, 943–954. doi: 10.1007/s12038-015-9569-6
Aksoy, U. R., Eltem, K. B., Meyvaci, A. A., and Karabat, S. (2007). Five-year survey of Ochratoxin A in processed sultanas from Turkey. Food Addit. Contam. 24, 292–296. doi: 10.1080/02652030601039021
Al-Anati, L., and Petzinger, E. (2006). Immunotoxic activity of ochratoxin A. J. Vet. Pharmacol. Ther. 29, 79–90. doi: 10.1111/j.1365-2885.2006.00718.x
Al-Hazmi, N. (2010). Determination of patulin and ochratoxin A using HPLC in apple juice samples in Saudi Arabia. Saudi J. Biol. Sci. 17, 353–359. doi: 10.1016/j.sjbs.2010.06.006
Alpsoy, L. (2010). Inhibitory effect of essential oil on aflatoxin activities. Afr. J. Biotechnol. 17, 2474–2481.
Alshannaq, A., and Yu, J. H. (2017). Occurrence, toxicity, and analysis of major mycotoxins in food. Int. J. Environ. Res. Public Health 14:632. doi: 10.3390/ijerph14060632
Andrade, P. D., Gomes da Silva, J. L., and Caldas, E. D. (2013). Simultaneous analysis of aflatoxins B1, B2, G1, G2, M1 and ochratoxin A in breast milk by high-performance liquid chromatography/fluorescence after liquid-liquid extraction with low temperature purification (LLE-LTP). J. Chromatogr. A 1304, 61–68. doi: 10.1016/j.chroma.2013.06.049
Ansari, F., Khodaiyan, F., Rezaei, K., and Rahmani, A. (2015). Modelling of aflatoxin G1 reduction by kefir grain using response surface methodology. J. Environ. Health Sci. Eng. 13:40. doi: 10.1186/s40201-015-0190-2
Anukul, N., Vangnai, K., and Mahakarnchanakul, W. (2013). Significance of regulation limits in mycotoxin contamination in Asia and risk management programs at the national level. J. Food Drug Anal. 21, 227–241. doi: 10.1016/j.jfda.2013.07.009
Aquino, S., Ferreira, F., Ribeiro, D. H. B., Correa, B., Greiner, R., and Villavicencio, A. L. C. H. (2005). Evaluation of viability of Aspergillus flavus and aflatoxins degradation in irradiated samples of maize. Braz. J. Microbiol. 36, 352–356. doi: 10.1590/S1517-83822005000400009
Aristil, J., Venturini, G., Maddalena, G., Toffolatti, S. L., and Spada, A. (2020). Fungal contamination and aflatoxin content of maize, moringa and peanut foods from rural subsistence farms in South Haiti. J. Stored Prod. Res. 85:101550. doi: 10.1016/j.jspr.2019.101550
Aroyeun, S. O., Adegoke, G. O., Varga, J., and Teren, J. (2009). Reduction of aflatoxin B1 and ochratoxin A in cocoa beans infected with Aspergillus via Ergosterol value. World Rev. Sci. Technol. Sustain. Dev. 6, 75–89. doi: 10.1504/WRSTSD.2009.022459
Aydin, A., Aksu, H., and Gunsen, U. (2011). Mycotoxin levels and incidence of mould in Turkish rice. Environ. Monit. Assess. 178, 271–280. doi: 10.1007/s10661-010-1688-9
Aziz, N. H., Attia, E. S. A., and Farag, S. A. (1997). Effect of gamma-irradiation on the natural occurrence of Fusarium mycotoxins in wheat, flour and bread. Food Nahrung 41, 34–37. doi: 10.1002/food.19970410109
Aziz, N. H., Ferial, M., Shahin, A. A., and Roushy, S. M. (2007). Control of Fusarium moulds and fumonisin B1 in seeds by gamma-irradiation. Food Control 18, 1337–1342. doi: 10.1016/j.foodcont.2005.12.013
Aziz, N. H., Moussa, L. A., and Far, F. M. (2004). Reduction of fungi and mycotoxins formation in seeds by gamma-radiation. J. Food Saf. 24, 109–127. doi: 10.1111/j.1745-4565.2004.tb00379.x
Badiale-Furlong, E., Dors, G. C., Oliveira, M., Dos, S., de Souza, M. M., and Kuhn, R. C. (2003). “Avaliacaodaqualidade de farinha de trigoeprodutos de panificaçãocomercializadas no Rio Grande do Sul” in Simpósio de Ciências de Alimentos e Saude (Anais: Florianópolis-SC/UFSC), 1–4.
Bankole, S. A. (1997). Effect of essential oils from two Nigerian medicinal plants (Azadirachta indica and Morinda lucida) on growth and aflatoxin B1 production in maize grain by a toxigenic Aspergillus flavus. Lett. Appl. Microbiol. 24, 190–192.
Basílico, M. Z., and Basílico, J. C. (1999). Inhibitory effects of some spice essential oils on aspergillus ochraceus NRRL 3174 growth and ochratoxin A production. Lett. Appl. Microbiol. 29, 238–241. doi: 10.1046/j.1365-2672.1999.00621.x
Begum, H., and Samajpati, N. (2000). Mycotoxin production on rice, pulses and oilseeds. Naturwissenschaften 87, 275–277. doi: 10.1007/s001140050720
Beretta, B., Gaiaschib, A., Galli, C. L., and Restani, P. (2000). Patulin in apple-based foods: occurrence and safety evaluation. Food Addit. Contam. 17, 399–406. doi: 10.1080/026520300404815
Berthiller, F., Cramer, B., Iha, M. H., Krska, R., Lattanzio, V. M. T., MacDonald, S., et al. (2017). Developments in mycotoxin analysis: an update for 2016-2017. World Mycotoxin J. 11, 5–32. doi: 10.3920/WMJ2017.2250
Bertuzzi, T., Giorni, P., Rastelli, S., Vaccino, P., Lanzanova, C., and Locatelli, S. (2020). Co-occurrence of moniliformin and regulated Fusarium toxins in maize and wheat grown in Italy. Molecules 25, 1–13. doi: 10.3390/molecules25102440
Bhat, R., and Reddy, K. R. N. (2017). Challenges and issues concerning mycotoxins contamination in oil seeds and their edible oils: updates from last decade. Food Chem. 215, 425–437. doi: 10.1016/j.foodchem.2016.07.161
Bhatt, R. V., Shetty, P. H., Amruth, R. P., and Sudershan, R. V. (1997). A foodborne disease outbreak due to the consumption of moldy sorghum and maize containing fumonisin mycotoxins. Clin. Toxicol. 35, 249–255.
Birck, N. M. M., Lorini, I., and Scussel, V. M. (2003). “Sanitary conditions and mycotoxins in wheat grains (Triticum aestivum) and flour (common and special) through milling processing” in IV Congreso Latino americano De Mycotoxicologia (LaHabana, Cuba: Anais)
Bottalico, A., and Perrone, G. (2002). Toxigenic Fusarium species and mycotoxins associated with head blight in small-grain cereals in Europe. Eur. J. Plant Pathol. 108, 611–624. doi: 10.1023/A:1020635214971
Burcu Aydın, E., Aydın, M., and Kemal Sezgintürk, M. (2020). Biosensors and the evaluation of food contaminant biosensors in terms of their performance criteria. Int. J. Environ. Anal. Chem. 100, 602–622. doi: 10.1080/03067319.2019.1672675
Cai, Y., McLaughlin, M., and Zhang, K. (2020). Advancing the FDA/office of regulatory affairs mycotoxin program: new analytical method approaches to addressing needs and challenges. J. AOAC Int. 103, 705–709. doi: 10.1093/jaocint/qsz007
Cairns, V., and Magan, N. (2003). “Impact of essential oils on growth and ochratoxin A production by Penicillium verrucosum and Aspergillus ochraceus on a wheat-based substrate.” in Advances in stored product protection. Proceedings of the 8th International Working Conference on Stored Product Protection, York, UK, 22–26 July 2002. Edited by Credland, P.F., Armitage, D.M., Bell, C.H., Cogan, P.M., Highley, E., 479–485.
Campbell, H., Choo, T. M., Vigier, B., and Underhill, L. (2000). Mycotoxins in barley and oat samples from Eastern Canada. Can. J. Plant Sci. 80, 977–980. doi: 10.4141/P00-042
Canada, H. (2020). Health Canada’s maximum levels for chemical contaminants in foods (Ontario: Health Canada Ottawa).
Carballo, D., Font, G., Ferrer, E., and Berrada, H. (2018). Evaluation of mycotoxin residues on ready-to-eat food by chromatographic methods coupled to mass spectrometry in tandem. Toxins 10:243. doi: 10.3390/toxins10060243
Castañares, E., Pavicich, M. A., Dinolfo, M. I., Moreyra, F., Stenglein, S. A., and Patriarca, A. (2020). Natural occurrence of Alternaria mycotoxins in malting barley grains in the main producing region of Argentina. J. Sci. Food Agric. 100, 1004–1011. doi: 10.1002/jsfa.10101
Chala, A., Mohammed, A., Ayalew, A., and Skinnes, H. (2013). Natural occurrence of aflatoxins in groundnut (Arachis hypogaeaL.) from eastern Ethiopia. Food Control 30, 2602–2605. doi: 10.1016/j.foodcont.2012.08.023
Chen, D., Chen, P., Cheng, Y., Peng, P., Liu, J., Ma, Y., et al. (2019). Deoxynivalenol decontamination in raw and germinating barley treated by plasma-activated water and intense pulsed light. Food Bioprocess Technol. 12, 246–254. doi: 10.1007/s11947-018-2206-2
Chen, X., Qiu, Y., Zhang, J., Guo, Y., Ding, Y., and Lyu, F. (2022). Degradation efficiency and products of deoxynivalenol treated by cold plasma and its application in wheat. Food Control 136:108874. doi: 10.1016/j.foodcont.2022.108874
Chiotta, M. L., Fumero, M. V., Cendoya, E., Palazzini, J. M., Alaniz-Zanon, M. S., Ramirez, M. L., et al. (2020). Toxigenic fungal species and natural occurrence of mycotoxins in crops harvested in Argentina. Rev. Argent. Microbiol. 52, 339–347. doi: 10.1016/j.ram.2020.06.002
Cho, M. S., Kim, K., Seo, E., Kassim, N., Mtenga, A. B., Shim, W. B., et al. (2010). Occurrence of patulin in various fruit juices from South Korea: an exposure assessment. Food Sci. Biotechnol. 19, 1–5. doi: 10.1007/s10068-010-0001-6
Chulze, S. N., Magnoli, C. E., and Dalcero, A. M. (2006). Occurrence of Ochratoxin A in wine and ochratoxigenic mycoflora in grape and dried vine fruits in South America. Int. J. Food Microbiol. 111, S5–S9. doi: 10.1016/j.ijfoodmicro.2006.02.006
Claeys, L., Romano, C., De Ruyck, K., Wilson, H., Fervers, B., Korenjak, M., et al. (2020). Mycotoxin exposure and human cancer risk: A systematic review of epidemiological studies. Compr. Rev. Food Sci. Food Saf. 19, 1449–1464. doi: 10.1111/1541-4337.12567
Dambolena, J. S., López, A. G., Cánepa, M. C., Theumer, M. G., Zygadlo, J. A., and Rubinstein, H. R. (2008). Inhibitory effect of cyclic terpenes (limonene, menthol, menthone and thymol) on Fusarium verticillioides MRC 826 growth and fumonisin B1 biosynthesis. Toxicon 51, 37–44. doi: 10.1016/j.toxicon.2007.07.005
Do, T. H., Tran, S. C., Le, C. D., Nguyen, H. B. T., Le, P. T. T., Le, H. H. T., et al. (2020). Dietary exposure and health risk characterization of aflatoxin B1, ochratoxin A, fumonisin B1, and zearalenone in food from different provinces in Northern Vietnam. Food Control 112:107108. doi: 10.1016/j.foodcont.2020.107108
Dong, M., Si, W., Jiang, K., Nie, D., Wu, Y., Zhao, Z., et al. (2015). Multi-walled carbon nanotubes as solid-phase extraction sorbents for simultaneous determination of type A trichothecenes in maize, wheat and rice by ultra-high performance liquid chromatography-tandem mass spectrometry. J. Chromatogr. A 1423, 177–182. doi: 10.1016/j.chroma.2015.10.068
Dong, H., Xian, Y., Xiao, K., Wu, Y., Zhu, L., and He, J. (2019). Development and comparison of single-step solid phase extraction and QuEChERS clean-up for the analysis of 7 mycotoxins in fruits and vegetables during storage by UHPLC-MS/MS. Food Chem. 274, 471–479. doi: 10.1016/j.foodchem.2018.09.035
Drakopoulos, D., Sulyok, M., Krska, R., Logrieco, A. F., and Vogelgsang, S. (2021). Raised concerns about the safety of barley grains and straw: A Swiss survey reveals a high diversity of mycotoxins and other fungal metabolites. Food Control 125:107919. doi: 10.1016/j.foodcont.2021.107919
EC (2006). Commission regulation (EC) no 1881/2006 of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs. Off J Eur Union.
Ediage, E. N., Di Mavungu, J. D., Monbaliu, S., Peteghem, C. V., and De Saeger, S. (2011). A validated multianalyte LC–MS/MS method for quantification of 25 mycotoxins in cassava flour, peanut cake and maize samples. J. Agric. Food Chem. 59, 5173–5180. doi: 10.1021/jf2009364
Edwards, S.G. (2007). Investigation of Fusarium mycotoxins in UK barley and oat production. Project Report No.415, AHDB Cereals & Oilseeds: Stoneleigh, UK.
El Darra, N., Gambacorta, L., and Solfrizzo, M. (2019). Multi mycotoxins occurrence in spices and herbs commercialized in Lebanon. Food Control 95, 63–70. doi: 10.1016/j.foodcont.2018.07.033
El-Nagerabi, S. A. F., Al-Bahry, S. N., Elshafie, A. E., and AlHilali, S. (2012). Effect of Hibiscus sabdariffa extract and Nigella sativa oil on the growth and aflatoxin B1 production of Aspergillus flavus and Aspergillus parasiticus strains. Food Control 25, 59–63. doi: 10.1016/j.foodcont.2011.09.033
El-Nagerabi, S. A. E., Elshafie, A. E., AlKhanjari, S. S., Al-Bahry, S. N., and Elamin, M. R. (2013). The potential of baobab (Adansonia digitata L.) extracts as biocontrol on the growth and aflatoxin production by Aspergillus flavus and A. parasiticus. J. Food Res. 2, 93–103. doi: 10.5539/jfr.v2n3p93
Embaby, E. M., Reda, M., Abdel-Wahhab, M. A., Omara, H., and Mokabel, A. M. (2013). Occurrence of toxigenic fungi and mycotoxins in some legume seeds. J. Agric. Technol. 9, 151–164.
Esper, R. H., Gonçalez, E., Marques, M. O. M., Felicio, R. C., and Felicio, J. D. (2014). Potential of essential oils for protection of grains contaminated by aflatoxin produced by Aspergillus flavus. Front. Microbiol. 5:269. doi: 10.3389/fmicb.2014.00269
European Commission (EU) (2006). European Union Commission regulation no. 1881/2006 setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Union 364, 5–24.
Ezekiel, C. N., Sulyok, M., Warth, B., Odebode, A. C., and Krska, R. (2012). Natural occurrence of mycotoxins in peanut cake from Nigeria. Food Control 27, 338–342. doi: 10.1016/j.foodcont.2012.04.010
Fan, Y., Liu, F., He, W., Qin, Q., Hu, D., Wu, A., et al. (2022). Screening of multi-mycotoxins in fruits by ultra-performance liquid chromatography coupled to ion mobility quadrupole time-of-flight mass spectrometry. Food Chem. 368:130858. doi: 10.1016/j.foodchem.2021.130858
Farzaneh, M., Shi, Z. Q., Ghassempour, A., Sedaghat, N., Ahmadzadeh, M., Mirabolfathy, M., et al. (2012). Aflatoxin B1 degradation by Bacillus subtilis UTBSP1 isolated from pistachio nuts of Iran. Food Control 23, 100–106. doi: 10.1016/j.foodcont.2011.06.018
Gandomi, H., Misaghi, A., Basti, A. A., Bokaei, S., Khosravi, A., Abbasifar, A., et al. (2009). Effect of Zataria multiflora Boiss. essential oil on growth and aflatoxin formation by Aspergillus flavus in culture media and cheese. Food Chem. Toxicol. 47, 2397–2400. doi: 10.1016/j.fct.2009.05.024
Gbashi, S., Madala, N. E., De Saeger, S., De Boevre, M., and Njobeh, P. B. (2019). Numerical optimization of temperature-time degradation of multiple mycotoxins. Food Chem. Toxicol. 125, 289–304. doi: 10.1016/j.fct.2019.01.009
Gemeda, N., Woldeamanuel, Y., Asrat, D., and Debella, A. (2014). Effect of Cymbopogon martinii, Foeniculum vulgare, and Trachyspermum ammi essential oils on the growth and mycotoxins production by Aspergillus species. Int. J. Food Sci. 874135, 1–9.
Gil-Serna, J., Mateo, E. M., Gonzalez-Jaen, M. T., Jimenez, M., Vazquez, C., and Patino, B. (2013). Contamination of barley seeds with Fusarium species and their toxins in Spain: an integrated approach. Food Addit. Contam. Part A 30, 372–380. doi: 10.1080/19440049.2012.743040
Giray, B., Atasayar, S., and Sahin, G. (2009). Determination of ochratoxin A and total aflatoxin levels in corn samples from Turkey by enzyme-linked immunosorbent assay. Mycotoxin Res. 25, 113–116. doi: 10.1007/s12550-009-0016-0
Giray, B., Girgin, G., Engin, A. B., Aydın, S., and Sahin, G. (2007). Aflatoxin levels in wheat samples consumed in some regions of Turkey. Food Control 18, 23–29. doi: 10.1016/j.foodcont.2005.08.002
Gomori, C., Nacsa-Farkas, E., Kerekes, E. B., Kocsube, S., Vagvolgyi, C., and Krisch, J. (2013). Evaluation of five essential oils for the control of food spoilage and mycotoxin producing fungi. Acta Biol. Szegediensis 57, 113–116.
Gonçalez, E., Nogueira, J. H. C., Fonseca, H., Felicio, J. D., Pino, F. A., and Correa, B. (2008). Mycobiota and mycotoxins in Brazilian peanut kernels from sowing to harvest. Int. J. Food Microbiol. 123, 184–190. doi: 10.1016/j.ijfoodmicro.2008.01.012
Gonzalez-Jartin, J. M., Alfonso, A., Rodriguez, I., Sainz, M. J., Vieytes, M. R., and Botana, L. M. (2019). A QuEChERS based extraction procedure coupled to UPLC-MS/MS detection for mycotoxins analysis in beer. Food Chem. 275, 703–710. doi: 10.1016/j.foodchem.2018.09.162
Goud, K. Y., Reddy, K. K., Satyanarayana, M., Kummari, S., and Gobi, K. V. (2020). A review on recent developments in optical and electrochemical aptamer-based assays for mycotoxins using advanced nanomaterials. Microchim. Acta 187:29. doi: 10.1007/s00604-019-4034-0
Gummert, M., Balingbing, C. B., Barry, G., and Estevez, L. A. (2009). Management options, technologies and strategies for minimized mycotoxin contamination of rice. World Mycotoxin J. 2, 151–159. doi: 10.3920/WMJ2008.1131
Guo, P., Yang, W., Hu, H., Wang, Y., and Li, P. (2019). Rapid detection of aflatoxin B1 by dummy template molecularly imprinted polymer capped CdTe quantum dots. Anal. Bioanal. Chem. 411, 2607–2617. doi: 10.1007/s00216-019-01708-2
Gurney, S. M. R., Scott, K. S., Kacinko, S. L., Presley, B. C., and Logan, B. K. (2014). Pharmacology, toxicology, and adverse effects of synthetic cannabinoid drugs. Forensic Sci. Rev. 26:53.
Han, Z., Jiang, K., Fan, Z., Diana Di Mavungu, J., Dong, M., Guo, W., et al. (2017). Multi-walled carbon nanotubes-based magnetic solid-phase extraction for the determination of zearalenone and its derivatives in maize by ultra-high performance liquid chromatography-tandem mass spectrometry. Food Control 79, 177–184. doi: 10.1016/j.foodcont.2017.03.044
Harris, K. L., Bobe, G., and Bourquin, L. D. (2009). Patulin surveillance in apple cider and juice marketed in Michigan. J. Food Prot. 72, 1255–1261. doi: 10.4315/0362-028X-72.6.1255
Hassan, Z. U., Al Thani, R., Balmas, V., Migheli, Q., and Jaoua, S. (2019). Prevalence of Fusarium fungi and their toxins in marketed feed. Food Control 104, 224–230. doi: 10.1016/j.foodcont.2019.04.045
Hell, K., Cardwell, K. F., and Poehling, H. M. (2003). Distribution of fungal species and aflatoxin contamination in stored maize in four agroecological zones in Benin, West-Africa. J. Phytopathol. 151, 690–698. doi: 10.1046/j.1439-0434.2003.00792.x
Hell, K., Setamou, M., Cardwell, K. F., and Poehling, H. M. (2000). The influence of storage practices on aflatoxin contamination in maize in four agroecological zones in Benin, West Africa. J. Stored Prod. Res. 36, 365–382. doi: 10.1016/S0022-474X(99)00056-9
Hendrickson, O. D., Chertovich, J. O., Zherdev, A. V., Sveshnikov, P. G., and Dzantiev, B. B. (2018). Ultrasensitive magnetic ELISA of zearalenone with pre-concentration and chemiluminescent detection. Food Control 84, 330–338. doi: 10.1016/j.foodcont.2017.08.008
Hope, R., Cairns-Fuller, V., Aldred, D., and Magan, N. (2005). Use of antioxidants and essential oils for controlling mycotoxins in grain. BCPC Crop Sci. Technol. 5, 429–436.
Houissa, H., Lasram, S., Sulyok, M., Šarkanj, B., Fontana, A., Strub, C., et al. (2019). Multimycotoxin LC-MS/MS analysis in pearl millet (Pennisetum glaucum) from Tunisia. Food Control 106:106738. doi: 10.1016/j.foodcont.2019.106738
Huang, Z., He, J., Li, H., Zhang, M., Wang, H., Zhang, Y., et al. (2020). Synthesis and application of magnetic-surfaced pseudo molecularly imprinted polymers for zearalenone pretreatment in cereal samples. Food Chem. 308:125696. doi: 10.1016/j.foodchem.2019.125696
Huang, X., Tang, X., Jallow, A., Qi, X., Zhang, W., Jiang, J., et al. (2020). Development of an ultrasensitive and rapid fluorescence polarization immunoassay for Ochratoxin A in Rice. Toxins 12:682. doi: 10.3390/toxins12110682
Huertas-Perez, J. F., Arroyo-Manzanares, N., Garcia-Campana, A. M., and Gamiz-Gracia, L. (2017). Solid phase extraction as sample treatment for the determination of Ochratoxin A in foods: a review. Crit. Rev. Food Sci. Nutr. 57, 3405–3420. doi: 10.1080/10408398.2015.1126548
Hussain, S., Asi, M. R., Iqbal, M., Khalid, N., Wajih-ul-Hassan, S., and Ariño, A. (2020). Patulin mycotoxin in mango and orange fruits, juices, pulps, and jams marketed in Pakistan. Toxins 12:52. doi: 10.3390/toxins12010052
Hussaini, A. M., Timothy, A. G., Olufunmilayo, H. A., Akanya, H., Ezekiel, A. S., and Ogbadu, G. H. (2009a). Fungi and some mycotoxins contaminating rice (Oryza sativa) in Niger State, Nigeria. Afr. J. Biotechnol. 6, 099–108.
Hussaini, A. M., Timothy, A. G., Olufunmilayo, H. A., Ezekiel, A. S., and Godwin, H. O. (2009b). Fungi and some mycotoxins found in mouldy sorghum in Niger state, Nigeria. World J. Agric. Sci. 5, 05–17.
Icli, N. (2019). Occurrence of patulin and 5-hydroxymethylfurfural in apple sour, which is a traditional product of Kastamonu, Turkey. Food Addit. Contam. Part A 36, 952–963. doi: 10.1080/19440049.2019.1605207
Iqbal, S. Z., Asi, M. R., Hanif, U., Zuber, M., and Jinap, S. (2016). The presence of aflatoxins and ochratoxin A in rice and rice products; and evaluation of dietary intake. Food Chem. 210, 135–140. doi: 10.1016/j.foodchem.2016.04.104
Iqbal, S. Z., Malik, S., Asi, M. R., Selamat, J., and Malik, N. (2018). Natural occurrence of patulin in different fruits, juices and smoothies and evaluation of dietary intake in Punjab, Pakistan. Food Control 84, 370–374. doi: 10.1016/j.foodcont.2017.08.024
Irakli, M. N., Skendi, A., and Papageorgiou, M. D. (2017). HPLC-DAD-FLD method for simultaneous determination of mycotoxins in wheat bran. J. Chromatogr. Sci. 55, 690–696. doi: 10.1093/chromsci/bmx022
Iram, W., Anjum, T., Iqbal, M., Ghaffar, A., Abbas, M., and Khan, A. M. (2016). Structural analysis and biological toxicity of aflatoxins B1 and B2 degradation products following detoxification by Ocimum basilicum and Cassia fistula aqueous extracts. Front. Microbiol. 7, 1–18. doi: 10.3389/fmicb.2016.01105
James, B., Adda, C., Cardwell, K., Annang, D., Hell, K., Korie, S., et al. (2007). Public information campaign on aflatoxin contamination of maize grains in market stores in Benin, Ghana and Togo. Food Addit. Contam. 24, 1283–1291. doi: 10.1080/02652030701416558
Janardhana, G. R., Raveesha, K. A., and Shetty, H. S. (1999). Mycotoxin contamination of maize grains grown in Karnataka (India). Food Chem. Toxicol. 37, 863–868. doi: 10.1016/S0278-6915(99)00067-8
Janik, E., Niemcewicz, M., Podogrocki, M., Ceremuga, M., Gorniak, L., Stela, M., et al. (2021). The existing methods and novel approaches in mycotoxins detection. Molecules 26:3981. doi: 10.3390/molecules26133981
Jayaraman, P., and Kalyanasundaram, I. (1990). Natural occurrence of toxigenic fungi and mycotoxins in rice bran. Mycopathologia 110, 81–85. doi: 10.1007/BF00446995
Jestoi, M. (2008). Emerging fusarium-mycotoxins fusaproliferin, beauvericin, enniatins, and Moniliformin-A review. Crit. Rev. Food Sci. Nutr. 48, 21–49. doi: 10.1080/10408390601062021
Ji, F., He, D., Olaniran, A., Mokoena, M., Xu, J., and Shi, J. (2019). Occurrence, toxicity, production and detection of Fusarium mycotoxin: A review. Food Prod. Proc. Nutr. 1:6. doi: 10.1186/s43014-019-0007-2
Jiang, D., Wei, D., Wang, L., Ma, S., Du, Y., and Wang, M. (2018). Multi walled carbon nanotube for one-step cleanup of 21 mycotoxins in corn and wheat prior to ultraperformance liquid chromatography-tandem mass spectrometry analysis. Toxins 10:409. doi: 10.3390/toxins10100409
Jimenez, M., and Mateo, R. (1997). Determination of mycotoxins produced by Fusarium isolates from banana fruits by capillary gas chromatography and high-performance liquid chromatography. J. Chromatogr. A 778, 363–372. doi: 10.1016/S0021-9673(97)00328-2
Juan, C., Mañes, J., Font, G., and Juan-Garcia, A. (2017). Determination of mycotoxins in fruit berry by-products using QuEChERS extraction method. LWT 86, 344–351. doi: 10.1016/j.lwt.2017.08.020
Jurjevic, J., Solfrizzo, M., Cvjetkovic, B., De Girolamo, A., and Visconti, A. (2002). Occurrence of Beauvericin in corn from Croatia. Food Technol. Biotechnol. 40, 91–94.
Jurjevic, Z., Wilson, J. P., Wilson, D. M., and Casper, H. H. (2007). Changes in fungi and mycotoxins in pearl millet under controlled storage conditions. Mycopathologia 164, 229–239. doi: 10.1007/s11046-007-9042-7
Kaaya, A. N., Eigel, W., and Harris, C. (2006). Peanut aflatoxin levels on farms and in markets of Uganda. Peanut Sci. 33, 68–75. doi: 10.3146/0095-3679(2006)33[68:PALOFA]2.0.CO;2
Karoki, P. K., Njue, W. M., Swaleh, S., Njoroge, E. K., and Kathurima, C. W. (2018). Determination of Ochratoxin A in selected cereal grains retailed in Nairobi County, Kenya. J. Food Res. 7:79. doi: 10.5539/jfr.v7n5p79
Khodaei, D., Javanmardi, F., and Khaneghah, A. M. (2021). The global overview of the occurrence of mycotoxins in cereals: a three-year survey. Curr. Opin. Food Sci. 39, 36–42. doi: 10.1016/j.cofs.2020.12.012
Kishore, G. K., Pande, S., Manjula, K., Rao, J. N., and Thomas, D. (2002). Occurrence of mycotoxins and toxigenic fungi in groundnut (Arachis hypogaea L.) seeds in Andhra Pradesh, India. Plant Pathol. J. 18, 204–209. doi: 10.5423/PPJ.2002.18.4.204
Kos, J., JanićHajnal, E., Malachová, A., Steiner, D., Stranska, M., Krska, R., et al. (2020). Mycotoxins in maize harvested in republic of Serbia in the period 2012–2015. Part 1: regulated mycotoxins and its derivatives. Food Chem. 312:126034. doi: 10.1016/j.foodchem.2019.126034
Kritzinger, Q., Aveling, T. A., Marasas, W. F., Rheeder, J. P., Van Der Westhuizen, L., and Shephard, G. S. (2002). Mycoflora and fumonisin mycotoxins associated with cowpea (Vigna unguiculata (L.) Walp) seeds. J. Agric. Food Chem. 51, 2188–2192. doi: 10.1021/jf026121v
Krska, R., Welzig, E., Berthiller, F., Molinelli, A., and Mizaikoff, B. (2005). Advances in the analysis of mycotoxins and its quality assurance. Food Addit. Contam. 22, 345–353. doi: 10.1080/02652030500070192
Kuiper-Goodman, T., Scott, P. M., and Watanabe, H. (1987). Risk assessment of the mycotoxin Zearalenone. Regul. Toxicol. Pharmacol. 7, 253–306. doi: 10.1016/0273-2300(87)90037-7
Kunz, B. M., Wanko, F., Kemmlein, S., Bahlmann, A., Rohn, S., and Maul, R. (2020). Development of a rapid multi-mycotoxin LC-MS/MS stable isotope dilution analysis for grain legumes and its application on 66 market samples. Food Control 109:106949. doi: 10.1016/j.foodcont.2019.106949
Lahouar, A., Jedidi, I., Sanchis, V., and Saïd, S. (2018). Aflatoxin B1, ochratoxin A and zearalenone in sorghum grains marketed in Tunisia. Food Addi. Contam. Part B Surv. 11, 103–110. doi: 10.1080/19393210.2018.1433239
Laidou, I. A., Thanassoulopoulos, C. C., and Liakopoulou-Kyriakides, M. (2001). Diffusion of patulin in the flesh of pears inoculated with four post-harvest pathogens. J. Phytopathol. 149, 457–461. doi: 10.1046/j.1439-0434.2001.00663.x
Lattanzio, V. M. T., Ciasca, B., Powers, S., and Visconti, A. (2014). Improved method for the simultaneous determination of aflatoxins, ochratoxin A and Fusarium toxins in cereals and derived products by liquid chromatography–tandem mass spectrometry after multi-toxinimmuno affinity clean up. J. Chromatogr. A 1354, 139–143. doi: 10.1016/j.chroma.2014.05.069
Lattanzio, V. M., Solfrizzo, M., Powers, S., and Visconti, A. (2007). Simultaneous determination of aflatoxins, ochratoxin A and Fusarium toxins in maize by liquid chromatography/tandem mass spectrometry after multitoxin immunoaffinity cleanup. Rapid Commun. Mass Spectrom. 21, 3253–3261. doi: 10.1002/rcm.3210
Lattanzio, V. M. T., Von Holst, C., Lippolis, V., De Girolamo, A., Logrieco, A. F., Mol, H. G. J., et al. (2019). Evaluation of mycotoxin screening tests in a verification study involving first time users. Toxins 11:129. doi: 10.3390/toxins11020129
Le, V. T., Vasseghian, Y., Dragoi, E. N., Moradi, M., and Mousavi Khaneghah, A. (2021). A review on graphene-based electrochemical sensor for mycotoxins detection. Food Chem. Toxicol. 148:111931. doi: 10.1016/j.fct.2020.111931
Lee, H. J., and Ryu, D. (2017). Worldwide occurrence of mycotoxins in cereals and cereal-derived food products: public health perspectives of their co-occurrence. J. Agric. Food Chem. 65, 7034–7051. doi: 10.1021/acs.jafc.6b04847
Lee, T. P., Sakai, R., Abdul Manaf, N., MohdRodhi, A., and Saad, B. (2014). High performance liquid chromatography method for the determination of patulin and 5-hydroxymethylfurfural in fruit juices marketed in Malaysia. Food Control 38, 142–149. doi: 10.1016/j.foodcont.2013.10.018
Li, Y., and Beghin, J. C. (2014). Protectionism indices for non-tariff measures: an application to maximum residue levels. Food Policy 45, 57–68. doi: 10.1016/j.foodpol.2013.12.005
Lippolis, V., Cervellieri, S., Damascelli, A., Pascale, M., Di Gioia, A., Longobardi, F., et al. (2018). Rapid prediction of deoxynivalenol contamination in wheat bran by MOS-based electronic nose and characterization of the relevant pattern of volatile compounds. J. Sci. Food Agric. 98, 4955–4962. doi: 10.1002/jsfa.9028
Lippolis, V., Pascale, M., Cervellieri, S., Damascelli, A., and Visconti, A. (2014). Screening of deoxynivalenol contamination in durum wheat by MOS-based electronic nose and identification of the relevant pattern of volatile compounds. Food Control 37, 263–271. doi: 10.1016/j.foodcont.2013.09.048
Liu, Z., Gao, J., and Yu, J. (2006). Aflatoxins in stored maize and rice grains in Liaoning province, China. J. Stored Prod. Res. 42, 468–479. doi: 10.1016/j.jspr.2005.09.003
Liu, X., Liu, X., Huang, P., Wei, F., Ying, G., Lu, J., et al. (2018). Regeneration and reuse of immunoaffinity column for highly efficient clean-up and economic detection of ochratoxin a in malt and ginger. Toxins 10:462. doi: 10.3390/toxins10110462
Loo, J. F. C., Chien, Y. H., Yin, F., Kong, S. K., Ho, H. P., and Yong, K. T. (2019). Upconversion and downconversion nanoparticles for biophotonics and nanomedicine. Coord. Chem. Rev. 400:213042. doi: 10.1016/j.ccr.2019.213042
Lopez, A. G., Theumer, M. G., Zygadlo, J. A., and Rubinstein, H. R. (2004). Aromatic plants essential oils activity on Fusarium verticillioides Fumonisin B1 production in corn grain. Mycopathologia 158, 343–349. doi: 10.1007/s11046-005-3969-3
MacDonald, S., Wilson, P., Barnes, K., Damant, A., Massey, R., Mortby, E., et al. (1999). Ochratoxin A in dried vine fruit: method development and survey. Food Addit. Contam. 16, 253–260. doi: 10.1080/026520399284019
Magan, N. (2006). Mycotoxins in Europe: prevention and early detection strategies. Mycopathologia 162, 245–253. doi: 10.1007/s11046-006-0057-2
Magnoli, C., Astoreca, A., Ponsone, L., Combina, M., Palacio, G., Rosa, C. A. R., et al. (2004). Survey of mycoflora and ochratoxin A in dried vine fruits from Argentina markets. Lett. Appl. Microbiol. 39, 326–331. doi: 10.1111/j.1472-765X.2004.01583.x
Magnoli, C., Astoreca, A., Ponsone, M. L., Fernández-Juri, M. G., Barberis, G., and Dalcero, A. M. (2007). Ochratoxin A and Aspergillus section Nigri in peanut seeds at different months of storage in Córdoba, Argentina. Int. J. Food Microbiol. 119, 213–218. doi: 10.1016/j.ijfoodmicro.2007.07.056
Magnoli, C., Astoreca, A., Ponsone, L., Fernández-Juri, M. G., Chiacchiera, S., and Dalcero, A. (2006). Ochratoxin A and the occurrence of ochratoxin A-producing black aspergilli in stored peanut seeds from Córdoba, Argentina. J. Sci. Food Agric. 86, 2369–2373. doi: 10.1002/jsfa.2625
Majdinasaba, M., Sheikh-Zeinoddin, M., Soleimanian-Zad, S., Li, P., Zhang, Q., Li, X., et al. (2015). Ultrasensitive and quantitative gold nanoparticle-based immunochromatographic assay for detection of ochratoxin A in agro-products. J. Chromatogr. B 974, 147–154. doi: 10.1016/j.jchromb.2014.10.034
Majeed, S., Boevre, M. D., Saeger, S. D., Rauf, W., Tawab, A., Habib, F., et al. (2018). Multiple mycotoxins in rice: occurrence and health risk assessment in children and adults of Punjab, Pakistan. Toxins 10:77. doi: 10.3390/toxins10020077
Majeed, S., Iqbal, M., Asi, M. R., and Iqbal, S. Z. (2013). Aflatoxins and ochratoxin A contamination in rice, corn and corn products from Punjab, Pakistan. J. Cereal Sci. 58, 446–450. doi: 10.1016/j.jcs.2013.09.007
Majid, I. (2007). Studying the mycotoxins found in wheat and maize under conditions of storage. Ph.D. Dissertation. Damascus University, Syria.
Makun, H. A., Dutton, M. F., Njobeh, P. B., Mwanza, M., and Kabiru, A. Y. (2011). Natural multi occurrence of mycotoxins in rice from Niger state, Nigeria. Mycotoxin Res. 27, 97–104. doi: 10.1007/s12550-010-0080-5
Malachova, A., Stranska, M., Vaclavikova, M., Elliott, C. T., Black, C., Meneely, J., et al. (2018). Advanced LC–MS-based methods to study the co-occurrence and metabolization of multiple mycotoxins in cereals and cereal-based food. Anal. Bioanal. Chem. 410, 801–825. doi: 10.1007/s00216-017-0750-7
Mallmann, C.A., Dilkin, M., Mürmann, L., Dilkin, P., and Almeida, C.A.A. (2003). Avaliação da contaminação por deoxivalenol em trigo utilizado na alimentação humana. In: Congresso Brasileiro de Farmácia. Anais.
Maraqa, A., Alsharoa, N. F., Farah, H., Albjeirami, W. M., Shakya, A. K., and Sallal, A. J. (2007). Effect of Nigella sativa extract and oil on aflatoxin production by Aspergillus flavus. Turk. J. Biol. 31, 155–159.
Marin, S., Ramos, A. J., Ho, G. C., and Sanchis, V. (2012). Reduction of mycotoxins and toxigenic fungi in the Mediterranean basin maize chain. Phytopathol. Mediterr. 51, 93–118.
Marin, S., Velluti, A., Ramos, A. J., and Sanchis, V. (2004). Effect of essential oils on zearalenone and deoxynivalenol production by Fusarium graminearum in non-sterilized maize grain. Food Microbiol. 21, 313–318. doi: 10.1016/j.fm.2003.08.002
Martins, M. L., Gimeno, A., Martins, H. M., and Bernardo, F. (2002). Co-occurrence of patulin and citrinin in Portuguese apples with rotten spots. Food Addit. Contam. 19, 568–574. doi: 10.1080/02652030210121320
Mateus, A. R. S., Barros, S., Pena, A., and Sanches Silva, A. (2021). Mycotoxins in pistachios (Pistacia vera L.): methods for determination, occurrence, decontamination. Toxins 13:682. doi: 10.3390/toxins13100682
Matumba, L., Van Poucke, C., Monjerezi, M., NjumbeEdiage, E., and De Saeger, S. (2015). Concentrating aflatoxins on the domestic market through groundnut export: A focus on Malawian groundnut value and supply chain. Food Control 51, 236–239. doi: 10.1016/j.foodcont.2014.11.035
Mehrez, A., Maatouk, I., Romero-González, R., Amara, A. B., Kraiem, M., Frenich, A. G., et al. (2016). Assessment of ochratoxin A stability following gamma irradiation: experimental approaches for feed detoxification perspectives. World Mycotoxin J. 9, 289–298. doi: 10.3920/WMJ2013.1652
Melinte, G., Hosu, O., Cristea, C., and Marrazza, G. (2022). DNA sensing technology a useful food scanning tool. Trends Analytic. Chem. 154:116679. doi: 10.1016/j.trac.2022.116679
Mendez-Albores, A., Arambula-Villa, G., Loarca-Pina, M. G. F., Castano-Tostado, E., and Moreno-Martínez, E. (2005). Safety and efficacy evaluation of aqueous citric acid to degrade B-aflatoxins in maize. Food Chem. Toxicol. 43, 233–238. doi: 10.1016/j.fct.2004.09.009
Miklos, G., Angeli, C., Ambrus, A., Nagy, A., Kardos, V., Zentai, A., et al. (2020). Detection of aflatoxins in different matrices and food-chain positions. Front. Microbiol. 11:1916. doi: 10.3389/fmicb.2020.01916
Mir, S. A., Dar, B. N., Shah, M. A., Sofi, S. A., Hamdani, A. M., Oliveira, C. A. F., et al. (2021). Application of new technologies in decontamination of mycotoxins in cereal grains: challenges, and perspectives. Food Chem. Toxicol. 148:111976. doi: 10.1016/j.fct.2021.111976
Mogensen, J. M., Thrane, J. C., and Nielsen, K. F. (2010). Production of fumonisin B2 and B4 by Aspergillus niger on grapes and raisins. J. Agric. Food Chem. 58, 954–958. doi: 10.1021/jf903116q
Mohamed, S. R., Deabes, M. M., Abdel-Motaleb, N. M., Mohamed, S. S., Badr, A. N., Shaban, E. M., et al. (2012). Antifungal activity of basil and mustard essential oils against spoilage toxigenic fungi in Egyptian pan bread and its economic evaluation. J. Appl. Sci. Res. 8, 5536–5542.
Mohamed, N. F., El-Dine, R. S. S., Kotb, M. A. M., and Saber, A. (2015). Assessing the possible effect of gamma irradiation on the reduction of aflatoxin B1, and on the moisture content in some cereal grains. Am. J. Biomed. Sci. 7, 33–39.
Mohana, D. C., Thippeswamy, S., Abhishek, R. U., Shobha, B., and Mamatha, M. G. (2017). Studies on seed-borne mycoflora and aflatoxin B1 contaminations in food-based seed samples: molecular detection of mycotoxigenic Aspergillus flavus and their management. Int. Food Res. J. 24, 422–427.
Moukas, A., Panagiotopoulou, V., and Markaki, P. (2008). Determination of patulin in fruit juices using HPLCDAD and GCMSD techniques. Food Chem. 109, 860–867. doi: 10.1016/j.foodchem.2008.01.015
Mravlje, J., Regvar, M., Staric, P., Mozetic, M., and Vogel-Mikus, K. (2021). Cold plasma affects germination and fungal community structure of buckwheat seeds. Plan. Theory 10:851. doi: 10.3390/plants10050851
Muriuki, G. K., and Siboe, G. M. (1995). Maize flour contaminated with toxigenic fungi and mycotoxins in Kenya. Afr. J. Health Sci. 2, 236–241.
Murthy, P. S., Ramalakshmi, K., and Srinivas, P. (2009). Fungitoxic activity of Indian borage (Plectranthus amboinicus) volatiles. Food Chem. 114, 1014–1018. doi: 10.1016/j.foodchem.2008.10.064
Muthomi,, and Mutitu, (2003). Occurrence of mycotoxin producing Fusarium species and other fungi on wheat kernels harvested in selected districts of Kenya. Afri. Crop Sci. Conf. Proc. 6, 335–339.
Nakagawa, H., Ohmichi, K., Sakamoto, S., Sago, Y., Kushiro, M., Nagashima, H., et al. (2011). Detection of a new Fusarium masked mycotoxin in wheat grain by high-resolution LC–Orbitrap™ MS. Food Addit. Contam: Part A 28, 1447–1456. doi: 10.1080/19440049.2011.597434
Nascimento, M. S., and Taniwaki, M. H. (2023). “Common and natural occurrence of pathogens, including fungi, leading to primary and secondary product contamination” in Present knowledge in food safety. eds. M. E. Knowles, L. E. Anelich, A. R. Boobis, and B. Popping (Greece: Elsevier), 330–356.
Nazir, A., Kalim, I., Sajjad, M., Usman, M., and Iqbal, M. (2019). Prevalence of aflatoxin contamination in pulses and spices in different regions of Punjab. Chem. Int. 5, 274–280.
Nganou Donkeng, N., SokamteTegang, A., TchindaSonwa, E., NtsamoBeumo, T. M., Nodem Sohanang, F. S., Douanla Nodem, N. F., et al. (2022). Fungal diversity and occurrence of Aflatoxin B1, Citrinine, and Ochratoxin A in rice of Cameroon. J. Food Proc. Preserve. 46:e16429. doi: 10.1111/jfpp.16429
Niyibituronsa, M., Onyango, A. N., Gaidashova, S., Imathiu, S. M., Uwizerwa, M., Wanjuki, I., et al. (2018). Evaluation of mycotoxin content in soybean (Glycine max L.) grown in Rwanda. Afr. J. Food Agric. Nutr. Dev. 18, 13808–13824. doi: 10.18697/ajfand.83.17710
Olatunde, O. O., Shiekh, K. A., and Benjakul, S. (2021). Pros and cons of cold plasma technology as an alternative non-thermal processing technology in seafood industry. Trends Food Sci. Technol. 111, 617–627. doi: 10.1016/j.tifs.2021.03.026
Onyedum, S. C., Adefolalu, F. S., Muhammad, H. L., Apeh, D. O., Agada, M. S., Imienwanrin, M. R., et al. (2020). Occurrence of major mycotoxins and their dietary exposure in north-Central Nigeria staples. Sci. African 7:e00188. doi: 10.1016/j.sciaf.2019.e00188
Orlando, B., Grignon, G., Vitry, C., Kashefifard, K., and Valade, R. (2019). Fusarium species and enniatin mycotoxins in wheat, durum wheat, triticale and barley harvested in France. Mycotoxin Res. 35, 369–380. doi: 10.1007/s12550-019-00363-x
Oruc, H. H., Cengiz, M., and Kalkanli, O. (2006). Comparison of aflatoxin and fumonisin levels in maize grown in Turkey and imported from the USA. Anim. Feed Sci. Technol. 128, 337–341. doi: 10.1016/j.anifeedsci.2006.02.008
Oteiza, J. M., Khaneghah, A. M., Campagnollo, F. B., Granato, D., Mahmoudi, M. R., Sant’Ana, A. S., et al. (2017). Influence of production on the presence of patulin and ochratoxin A in fruit juices and wines of Argentina. LWT Food Sci. Technol. 80, 200–207. doi: 10.1016/j.lwt.2017.02.025
Ott, L. C., Appleton, H. J., Shi, H., Keener, K., and Mellata, M. (2021). High voltage atmospheric cold plasma treatment inactivates Aspergillus flavus spores and deoxynivalenol toxin. Food Microbiol. 95:103669. doi: 10.1016/j.fm.2020.103669
Ottoboni, M., Pinotti, L., Tretola, M., Giromini, C., Fusi, E., Rebucci, R., et al. (2018). Combining E-nose and lateral flow immunoassays (LFIAs) for rapid occurrence/co-occurrence aflatoxin and fumonisin detection in maize. Toxins 10:416. doi: 10.3390/toxins10100416
Oyedele, O. A., Ezekiel, C. N., Sulyok, M., Adetunji, M. C., Warth, B., Atanda, O. O., et al. (2017). Mycotoxin risk assessment for consumers of groundnut in domestic markets in Nigeria. Int. J. Food Microbiol. 251, 24–32. doi: 10.1016/j.ijfoodmicro.2017.03.020
Palumbo, J. D., O’Keeffe, T. L., Vasquez, S. J., and Mahoney, N. E. (2011). Isolation and identification of ochratoxin A-producing Aspergillus section Nigri strains from California raisins. Lett. Appl. Microbiol. 52, 330–336. doi: 10.1111/j.1472-765X.2011.03004.x
Pande, N., Saxena, J., and Pandey, H. (1990). Natural occurrence of mycotoxins in some cereals. Mycoses 33, 126–128. doi: 10.1111/myc.1990.33.3.126
Pandey, A. K., Samota, M. K., and Silva, A. S. (2022). Mycotoxins along the tea supply chain: a dark side of an ancient and high valued aromatic beverage. Crit. Rev. Food Sci. Nutr. doi: 10.1080/10408398.2022.2061908
Pandey, A. K., Sonker, N., and Singh, P. (2016). Efficacy of some essential oils against Aspergillus flavus with special reference to Lippia alba oil an inhibitor of fungal proliferation and aflatoxin B1 production in green gram seeds during storage. J. Food Sci. Technol. 81, M928–M934. doi: 10.1111/1750-3841.13254
Pascale, M., De Girolamo, A., Lippolis, V., Stroka, J., Mol, H. G. J., and Lattanzio, V. M. T. (2019). Performance evaluation of LC-MS methods for multimycotoxin determination. J. AOAC Int. 102, 1708–1720. doi: 10.5740/jaoacint.19-0068
Payne, G. A., and Brown, M. P. (1998). Genetics and physiology of aflatoxin biosynthesis. Annu. Rev. Phytopathol. 36, 329–362. doi: 10.1146/annurev.phyto.36.1.329
Pereira, V. L., Fernandes, J. O., and Cunha, S. C. (2014). Mycotoxins in cereals and related foodstuffs: A review on occurrence and recent methods of analysis. Trends Food Sci. Technol. 36, 96–136. doi: 10.1016/j.tifs.2014.01.005
Pernica, M., Kyralová, B., Svoboda, Z., Boško, R., Brožková, I., Česlová, L., et al. (2022). Levels of T-2 toxin and its metabolites, and the occurrence of Fusarium fungi in spring barley in the Czech Republic. Food Microbiol. 102:103875. doi: 10.1016/j.fm.2021.103875
Pietri, A., Bertuzzi, T., Pallaroni, L., and Piva, G. (2004). Occurance of mycotoxins and ergosterol in maize harvested over 5 years in the northern Italy. Food Addit. Contam. 21, 479–487. doi: 10.1080/02652030410001662020
Pildain, M. B., Vaamonde, G., and Cabral, D. (2004). Analysis of population structure of Aspergillus flavus from peanut based on vegetative compatibility, geographic origin, mycotoxin and sclerotia production. Int. J. Food Microbiol. 93, 31–40. doi: 10.1016/j.ijfoodmicro.2003.10.007
Polišenská, I., Jirsa, O., Vaculová, K., Pospíchalová, M., Wawroszova, S., and Frydrych, J. (2020). Fusarium mycotoxins in two hulless oat and barley cultivars used for food purposes. Foods 9:1037. doi: 10.3390/foods9081037
Ponzilacqua, B., Rottinghaus, G. E., Landers, B. R., and Oliveira, C. A. F. (2019). Effects of medicinal herb and Brazilian traditional plant extracts on in vitro mycotoxin decontamination. Food Control 100, 24–27. doi: 10.1016/j.foodcont.2019.01.009
Prakash, B., Shukla, S., Singh, P., Mishra, P. K., Dubey, N. K., and Kharwar, R. N. (2011). Efficacy of chemically characterized Ocimum gratissimum L. essential oil as an antioxidant and a safe plant-based antimicrobial against fungal and aflatoxin B1 contamination of spices. Food Res. Int. 44, 385–390. doi: 10.1016/j.foodres.2010.10.002
Puel, O., Galtier, P., and Oswald, I. P. (2010). Biosynthesis and toxicological effects of patulin. Toxins 2, 613–631. doi: 10.3390/toxins2040613
Rai, A., Dixit, S., Singh, S. P., Gautam, N. K., Das, M., and Tripathi, A. (2018). Presence of zearalenone in cereal grains and its exposure risk assessment in Indian population. J. Food Sci. 83, 3126–3133. doi: 10.1111/1750-3841.14404
Rasooli, I., Fakoor, M. H., Yadegarinia, D., Gachkar, L., Allameh, A., and Rezaei, M. B. (2008). Anti mycotoxigenic characteristics of Rosmarinus officinalis and Trachyspermum copticum L. essential oils. Int. J. Food Microbiol. 122, 135–139. doi: 10.1016/j.ijfoodmicro.2007.11.048
Rasooli, I., and Owlia, P. (2005). Chemoprevention by thyme oils of Aspergillus parasiticus growth and aflatoxin production. Phytochemistry 66, 2851–2856. doi: 10.1016/j.phytochem.2005.09.029
Razzaghi-Abyaneh, M., Yoshinari, T., Shams-Ghahfarokhi, M., Rezaee, M. B., Nagasawa, H., and Sakuda, S. (2007). Dillapiol and apiolas specific inhibitors for the biosynthesis of aflatoxin G1 in Aspergillus parasiticus. Biosci. Biotechnol. Biochem. 71, 2329–2332. doi: 10.1271/bbb.70264
Reddy, K. R. N., Reddy, C. S., and Muralidharan, K. (2009a). Potential of botanicals and biocontrol agents on growth and aflatoxin production by Aspergillus flavus infecting rice grains. Food Control. 20, 173–178. doi: 10.1016/j.foodcont.2008.03.009
Reddy, K. R., Reddy, C. S., and Muralidharan, K. (2009b). Detection of Aspergillus spp. and aflatoxin B1 in rice in India. Food Microbiol. 26, 27–31. doi: 10.1016/j.fm.2008.07.013
Richard, J. L., Plattner, R. D., Mary, J., and Liska, S. L. (1999). The occurrence of Ochratoxin A in dust collected from a problem home hold. Mycopathologia 146, 99–103. doi: 10.1023/A:1007056627296
Rico-Yuste, A., Gomez-Arribas, L. N., Perez-Conde, M. C., Urraca, J. L., and Moreno-Bondi, M. C. (2018). Rapid determination of Alternaria mycotoxins in tomato samples by pressurized liquid extraction coupled to liquid chromatography with fluorescence detection. Food Addit. Contam. Part A 35, 2175–2182. doi: 10.1080/19440049.2018.1512759
Rubert, J., Dzuman, Z., Vaclavikova, M., Zachariasova, M., Soler, C., and Hajslova, J. (2012). Analysis of mycotoxins in barley using ultra high liquid chromatography high resolution mass spectrometry: comparison of efficiency and efficacy of different extraction procedures. Talanta 99, 712–719. doi: 10.1016/j.talanta.2012.07.010
Savi, G. D., Piacentini, K. C., Bittencourt, K. O., and Scussel, V. M. (2014). Ozone treatment efficiency on Fusarium graminearum and deoxynivalenol degradation and its effects on whole wheat grains (Triticum aestivum L.) quality and germination. J. Stored Prod. Res. 59, 245–253. doi: 10.1016/j.jspr.2014.03.008
Savi, G. D., Piacentini, K. C., and Scussel, V. M. (2015). Ozone treatment efficiency in Aspergillus and Penicillium growth inhibition and mycotoxin degradation of stored wheat grains (Triticum aestivum L.). J. Food Process. Preserv. 39, 940–948. doi: 10.1111/jfpp.12307
Saxena, N., Dwivedi, P. D., Ansari, K. M., and Das, M. (2008). Patulin in apple juices: incidence and likely intake in an Indian population. Food Addit. Contam. 1, 140–146. doi: 10.1080/02652030802378848
Schöneberg, T., Martin, C., Wettstein, F. E., Bucheli, T. D., Mascher, F., Bertossa, M., et al. (2016). Fusarium and mycotoxin spectra in Swiss barley are affected by various cropping techniques. Food Addit. Contam. Part A 33, 1608–1619. doi: 10.1080/19440049.2016.1219071
Scudamore, K. A., and Satel, S. (2009). Occurrence of Fusarium mycotoxins in maize imported into the UK, 2004–2007. Food Addit. Contam. Part A 26, 363–371. doi: 10.1080/02652030802406219
Segvic, M., Cvetnic, Z., Pepeljnjak, S., and Kosalec, I. (2009). Co-occurrence of aflatoxins, ochratoxin A, fumonisins, and zearalenone in cereals and feed, determined by competitive direct enzyme-linked immunosorbent assay and thin- layer chromatography. Arch. Ind. Hyg. Toxicol. 60, 427–434. doi: 10.2478/10004-1254-60-2009-1975
Selvaraj, J. N., Wang, Y., Zhou, L., Zhao, Y., Xing, F., Dai, X., et al. (2015). Recent mycotoxin survey data and advanced mycotoxin detection techniques reported from China: A review. Food Addit. Contam. Part A. 32, 440–452. doi: 10.1080/19440049.2015.1010185
Senthilkumar, T., Jayas, D. S., White, N. D. G., Fields, P. G., and Grafenhan, T. (2016). Nearinfrared (NIR) hyperspectral imaging: theory and applications to detect fungal infection andmycotoxin contamination in food products. Indian J. Entomol. 78:91. doi: 10.5958/0974-8172.2016.00029.8
Senthilkumar, R. P., Natarajan, A., and Kavitha, R. (2021). Co-occurrence of mycotoxins in pearl millet. Pharma Innov. J. 10, 253–255.
Serra, R., Bragab, A., and Venancio, A. (2005). Mycotoxin-producing and other fungi isolated from grapes for wine production, with particular emphasis on ochratoxin A. Res. Microbiol. 156, 515–521. doi: 10.1016/j.resmic.2004.12.005
Shetty, P. H., and Bhatt, R. V. (1997). Natural occurrence of fumonisin B1 and its co-occurrence with aflatoxin B1 in Indian sorghum, maize, and poultry feeds. J. Agric. Food Chem. 45, 2170–2173. doi: 10.1021/jf960607s
Shi, H., Schwab, W., Liu, N., and Yu, P. (2019). Major ergot alkaloids in naturally contaminated cool-season barley grain grown under a cold climate condition in western Canada, explored with near-infrared (NIR) and fourier transform mid-infrared (ATR-FT/MIR) spectroscopy. Food Control 102, 221–230. doi: 10.1016/j.foodcont.2019.03.025
Shukla, S., Shankar, R., and Singh, S. P. (2014). Food safety regulatory model in India. Food Control 37, 401–413. doi: 10.1016/j.foodcont.2013.08.015
Shukla, R., Singh, P., Prakash, B., and Dubey, N. K. (2012). Antifungal, aflatoxin inhibition and antioxidant activity of Callistemon lanceolatus (Sm.) sweet essential oil and its major component 1,8-cineole against fungal isolates from chickpea. Food Control 25, 27–33. doi: 10.1016/j.foodcont.2011.10.010
Sindhu, S., Chempakam, B., Leela, N. K., and Suseela Bhai, R. (2011). Chemoprevention by essential oil of turmeric leaves (Curcuma longa L.) on the growth of Aspergillus flavus and aflatoxin production. Food Chem. Toxicol. 49, 1188–1192. doi: 10.1016/j.fct.2011.02.014
Singh, J., and Mehta, A. (2020). Rapid and sensitive detection of mycotoxins by advanced and emerging analytical methods: A review. Food Sci. Nutr. 8, 2183–2204. doi: 10.1002/fsn3.1474
Siruguri, V., Kumar, U. P., Raghu, P., Rao, V. V. M., Sesikeran, B., Toteja, G. S., et al. (2012). Aflatoxin contamination in stored rice variety PAU 201 collected from Punjab, India. Ind. J. Med. Res. 136, 89–97.
Smith, C. A., Woloshuk, C. P., Robertson, D., and Payne, G. A. (2007). Silencing of the aflatoxin gene cluster in a diploid strain of Aspergillus flavus is suppressed by ectopic aflR expression. Genetics 176, 2077–2086. doi: 10.1534/genetics.107.073460
Soldatkin, O. O., Stepurska, K., Arkhypova, V., Soldatkin, A., El’Skaya, A., Lagarde, F., et al. (2017). Conductometric enzyme biosensor for patulin determination. Sensors Actuators B Chem. 239, 1010–1015. doi: 10.1016/j.snb.2016.08.121
Soliman, K. M., and Badeaa, R. I. (2002). Effect of oil extracted from some medical plants on different mycotoxigenic fungi. Food Chem. Toxicol. 4, 1669–1675. doi: 10.1016/s0278-6915(02)00120-5
Sonker, N., Pandey, A. K., and Singh, P. (2014b). Efficiency of Artemisia nilagirica (Clarke) Pamp. essential oil as a mycotoxicant against postharvest mycobiota of table grapes. J. Sci. Food Agric. 95, 1932–1939. doi: 10.1002/jsfa.6901
Sonker, N., Pandey, A. K., Singh, P., and Tripathi, N. N. (2014a). Assessment of Cymbopogon citratus (DC.) Stapf essential oil as herbal preservatives based on antifungal, antiaflatoxin and antiochratoxin activities and in vivo efficacy during storage. J. Food Sci. 79, 628–634. doi: 10.1111/1750-3841.12390
Srianujata, S. (2011). “Regulatory update and control measures for prevention and reduction of mycotoxins contamination in foods and feeds.” in Proceedings of FFTC-KU 2011 Conference: International Seminar ON Risk Assessment and Risk Management of Mycotoxins for Food Safety in Asia. Kasetsart University, Bangkok, Thailand.
Ssepuuya, G., Van Poucke, C., Ediage, E. N., Mulholland, C., Tritscher, A., Verger, P., et al. (2018). Mycotoxin contamination of sorghum and its contribution to human dietary exposure in four sub-Saharan countries. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 35, 1384–1393. doi: 10.1080/19440049.2018.1461253
Sultana, B., Naseer, R., and Nigam, P. (2015). Utilization of agro-wastes to inhibit aflatoxins synthesis by Aspergillus parasiticus: a biotreatment of three cereals for safe long-term storage. Bioresour. Technol. 197, 443–450. doi: 10.1016/j.biortech.2015.08.113
Sulyok, M., Berthiller, F., Krska, R., and Schuhmacher, R. (2006). Development and validation of a liquid chromatography/tandem mass spectrometric method for the determination of 39 mycotoxins in wheat and maize. Rapid Commun. Mass Spectrom. 20, 2649–2659. doi: 10.1002/rcm.2640
Tanaka, K., Sago, Y., Zheng, Y., Nakagawa, H., and Kushiro, M. (2007). Mycotoxins in rice. Int. J. Food Microbiol. 119, 59–66. doi: 10.1016/j.ijfoodmicro.2007.08.002
Tangni, E. K., Theys, R., Mignolet, E., Maudoux, M., Michelet, J. Y., and Larondelle, Y. (2003). Patulin in domestic and imported apple-based drinks in Belgium: occurrence and exposure assessment. Food Addit. Contam. 20, 482–489. doi: 10.1080/0265203031000093204
Tebele, S. M., Gbashi, S., Adebo, O., Changwa, R., Naidu, K., and Njobeh, P. B. (2020). Quantification of multimycotoxin in cereals (maize, maize porridge, sorghum and wheat) from Limpopo province of South Africa. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 37, 1922–1938. doi: 10.1080/19440049.2020.1808715
Thanushree, M. P., Sailendri, D., Yoha, K. S., Moses, J. A., and Anandharamakrishnan, C. (2019). Mycotoxin contamination in food: an exposition on spices. Trends Food Sci. Technol. 93, 69–80. doi: 10.1016/j.tifs.2019.08.010
Tola, M., Kebede, B., and Yildiz, F. (2016). Occurrence, importance and control of mycotoxins: A review. Cog. Food Agric. 2:1191103. doi: 10.1080/23311932.2016.1191103
Tölgyesi, Á., Kozma, L., and Sharma, V. K. (2020). Determination of Alternaria toxins in sunflower oil by liquid chromatography isotope dilution tandem mass spectrometry. Molecules 25:1685. doi: 10.3390/molecules25071685
Topi, D., Babic, J., Pavsic-Vrtac, K., Tavcar-Kalcher, G., and Jakovac-Strajn, B. (2021). Incidence of Fusarium mycotoxins in wheat and maize from Albania. Molecules 26:172. doi: 10.3390/molecules26010172
Trombete, F. M., Porto, Y. D., Freitas-Silva, O., Pereira, R. V., Direito, G. M., Saldanha, T., et al. (2017). Efficacy of ozone treatment on mycotoxins and fungal reduction in artificially contaminated soft wheat grains. J. Food Process. Preserv. 41:e12927. doi: 10.1111/jfpp.12927
Tseng, T. C., and Tu, J. C. (1997). Mycoflora and mycotoxins in adzuki and mung beans produced in Ontario, Canada. Microbios 90, 87–95.
Turner, N. W., Bramhmbhatt, H., Szabo-Vezse, M., Poma, A., Coker, R., and Piletsky, S. A. (2015). Analytical methods for determination of mycotoxins: an update (2009-2014). Anal. Chim. Acta 901, 12–33. doi: 10.1016/j.aca.2015.10.013
Van Egmond, H. P., and Jonker, M. A. (2008). “Regulations and limits for mycotoxins in fruits and vegetables” in Mycotoxins in fruits and vegetables. eds. R. Barkai-Golan and N. Paster (San Diego, CA: Elsevier), 45–74.
Velluti, A., Sanchis, V., Ramos, A. J., Egido, J., and Mari, S. (2003). Inhibitory effect of cinnamon, clove, lemongrass, oregano and palmarose essential oils on growth and fumonisin B1 production by Fusarium proliferatum in maize grain. Int. J. Food Microbiol. 89, 145–154. doi: 10.1016/S0168-1605(03)00116-8
Velluti, A., Sanchis, V., Ramos, A. J., Turon, C., and Marín, S. (2004). Impact of essential oils on growth rate, zearalenone and deoxynivalenol production by Fusarium graminearum under different temperature and water activity conditions in maize grain. J. Appl. Microbiol. 96, 716–724. doi: 10.1111/j.1365-2672.2004.02212.x
Vismer, H. F., Shephard, G. S., Rheeder, J. P., van der Westhuizen, L., and Bandyopadhyay, R. (2015). Relative severity of fumonisin contamination of cereal crops in West Africa. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 32, 1952–1958. doi: 10.1080/19440049.2015.1084654
Vismer, H. F., Shephard, G. S., van der Westhuizen, L., Mngqawa, P., Bushula-Njah, V., and Leslie, J. F. (2019). Mycotoxins produced by Fusarium proliferatum and F. pseudonygamai on maize, sorghum and pearl millet grains in vitro. Int. J. Food Microbiol. 296, 31–36. doi: 10.1016/j.ijfoodmicro.2019.02.016
Vithu, P., and Moses, J. A. (2016). Machine vision system for food grain quality evaluation: A review. Trends Food Sci. Technol. 56, 13–20. doi: 10.1016/j.tifs.2016.07.011
Wang, M., Jiang, N., Xian, H., Wei, D., Shi, L., and Feng, X. (2016). A single-step solid phase extraction for the simultaneous determination of 8 mycotoxins in fruits by ultra-high performance liquid chromatography tandem mass spectrometry. J. Chromatogr. A 1429, 22–29. doi: 10.1016/j.chroma.2015.12.004
Wang, B., Mahoney, N. E., Pan, Z., Khir, R., Wu, B., Ma, H., et al. (2016). Effectiveness of pulsed light treatment for degradation and detoxification of aflatoxin B1 and B2 in rough rice and rice bran. Food Control 59, 461–467. doi: 10.1016/j.foodcont.2015.06.030
Wielogorska, E., Ahmed, Y., Meneely, J., Graham, W. G., Elliott, C. T., and Gilmore, B. F. (2019a). A holistic study to understand the detoxification of mycotoxins in maize and impact on its molecular integrity using cold atmospheric plasma treatment. Food Chem. 301:125281. doi: 10.1016/j.foodchem.2019.125281
Wielogorska, E., Mooney, M., Eskola, M., Ezekiel, C. N., Stranska, M., Krska, R., et al. (2019b). Occurrence and human-health impacts of mycotoxins in Somalia. J. Agric. Food Chem. 67, 2052–2060. doi: 10.1021/acs.jafc.8b05141
Wilson, J. P., Jurjevic, Z., Hanna, W. W., Wilson, D. M., Potter, T. L., and Coy, A. E. (2006). Host-specific variation in infection by toxigenic fungi and contamination by mycotoxins in pearl millet and corn. Mycopathologia 161, 101–107. doi: 10.1007/s11046-005-0170-7
Wolf, K., and Schweigert, J. F. (2018). Mycotoxin analysis: a focus on rapid methods; Partnership for Aflatoxin Control in Africa: Addis Ababa, Ethiopia, 2018. J. Chromatogr. A 1304, 61–68.
Xia, X., Wang, H., Yang, H., Deng, S., Deng, R., Dong, Y., et al. (2018). Dual-terminal stemmed aptamer beacon for label-free detection of aflatoxin B1 in broad bean paste and peanut oil via aggregation-induced emission. J. Agric. Food Chem. 66, 12431–12438. doi: 10.1021/acs.jafc.8b05217
Xie, L., Chen, M., and Ying, Y. (2016). Development of methods for determination of aflatoxins. Crit. Rev. Food Sci. Nutr. 56, 2642–2664. doi: 10.1080/10408398.2014.907234
Xing, F., Hua, H., Selvaraj, J. N., Yuan, Y., Zhao, Y., Zhou, L., et al. (2014). Degradation of fumonisin B1 by cinnamon essential oil. Food Control 38, 37–40. doi: 10.1016/j.foodcont.2013.09.045
Xu, W., Han, X., and Li, F. (2019). Co-occurrence of multi-mycotoxins in wheat grains harvested in Anhui province, China. Food Control 96, 180–185. doi: 10.1016/j.foodcont.2018.09.006
Yamamoto-Ribeiro, M. M., Grespan, R., Kohiyama, C. Y., Ferreira, F. D., Mossini, S. A., Silva, E. L., et al. (2013). Effect of Zingiber officinale essential oil on Fusarium verticillioides and fumonisin production. Food Chem. 141, 3147–3152. doi: 10.1016/j.foodchem.2013.05.144
Yang, Y., Li, G., Wu, D., Liu, J., Li, X., Luo, P., et al. (2020). Recent advances on toxicity and determination methods of mycotoxins in foodstuffs. Trends Food Sci. Technol. 96, 233–252. doi: 10.1016/j.tifs.2019.12.021
Yao, H., Hruska, Z., and DiMavungu, J. D. (2015). Developments in detection and determination of aflatoxins. World Mycotoxin J. 8, 181–191. doi: 10.3920/WMJ2014.1797
Younis, M., Younis, A., and Xia, X. H. (2020). “Use of biosensors for mycotoxins analysis in food stuff” in Nanobiosensors: From design to applications. eds. A. Wu and W. S. Khan (Weinheim, Germany: Wiley-VCH Verlag GmbH & Co. KGaA), 171–201.
Zhang, L., Dou, X. W., Zhang, C., Logrieco, A. F., and Yang, M. H. (2018). A review of current methods for analysis of mycotoxins in herbal medicines. Toxins 10:65. doi: 10.3390/toxins10020065
Zhang, W., Han, Y., Chen, X., Luo, X., Wang, J., Yue, T., et al. (2017). Surface molecularly imprinted polymer capped Mn-doped ZnS quantum dots as a phosphorescent nanosensor for detecting patulin in apple juice. Food Chem. 232, 145–154. doi: 10.1016/j.foodchem.2017.03.156
Zhang, K., Schaab, M. R., Southwood, G., Tor, E. R., Aston, L. S., Song, W., et al. (2017). A collaborative study: determination of mycotoxins in corn, peanut butter, and wheat flour using stable isotope dilution assay (SIDA) and liquid chromatography-tandem mass spectrometry (LC-MS/MS). J. Agric. Food Chem. 65, 7138–7152. doi: 10.1021/acs.jafc.6b04872
Zhang, Z., Tang, X., Wang, D., Zhang, Q., Li, P., and Ding, X. (2015). Rapid on-site sensing aflatoxin B1 in food and feed via a chromatographic time-resolved fluoroimmunoassay. PLoS One 10:e0123266. doi: 10.1371/journal.pone.0145988
Zhu, Y., Xia, X., Deng, S., Yan, B., Dong, Y., Zhang, K., et al. (2019). Label-free fluorescent aptasensing of mycotoxins via aggregation-induced emission dye. Dyes Pigments 170:107572. doi: 10.1016/j.dyepig.2019.107572
Zinedine, A., Brera, C., Elakhdari, S., Catano, C., Debegnach, F., Angelini, S., et al. (2006). Natural occurrence of mycotoxins in cereals and spices commercialized in Morocco. Food Control 17, 868–874. doi: 10.1016/j.foodcont.2005.06.001
Zinedine, A., Soriano, J. M., Juan, C., Mojemmi, B., Moltó, J. C., Bouklouze, A., et al. (2007). Incidence of ochratoxin A in rice and dried fruits from Rabat and Salé area, Morocco. Food Addit. Contam. 24, 285–291. doi: 10.1080/02652030600967230
Zohri, A. A., and Abdel-Gawad, K. M. (1993). Survey of mycoflora and mycotoxins of some dried fruits in Egypt. J. Basic Microbiol. 33:279. doi: 10.1002/jobm.3620330413
Zouaoui, N., Sbaii, N., Bacha, H., and Abid-Essefi, S. (2015). Occurrence of patulin in various fruit juice marketed in Tunisia. Food Control 51, 356–360. doi: 10.1016/j.foodcont.2014.09.048
Keywords: ochratoxin, food commodities, aflatoxins, fumonisins, physical methods, essential oils
Citation: Pandey AK, Samota MK, Kumar A, Silva AS and Dubey NK (2023) Fungal mycotoxins in food commodities: present status and future concerns. Front. Sustain. Food Syst. 7:1162595. doi: 10.3389/fsufs.2023.1162595
Edited by:
Limbikani Matumba, Lilongwe University of Agriculture and Natural Resources, MalawiReviewed by:
K. S. Yoha, Entrepreneurship and Management - Thanjavur, IndiaCopyright © 2023 Pandey, Samota, Kumar, Silva and Dubey. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Abhay K. Pandey, YWJoYXlrdW1hcnBhbmRleS5rdUBnbWFpbC5jb20=; Mahesh K. Samota, bWFoZXNoLmlhcmkxQGdtYWlsLmNvbQ==; Ana Sanches Silva, YW5hdGVyZXNzQGdtYWlsLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.