 Benito Cortés-Rivas
Benito Cortés-Rivas Víctor Hugo Monzón
Víctor Hugo Monzón Juliana Ordones Rego2
Juliana Ordones Rego2 José Neiva Mesquita-Neto
José Neiva Mesquita-Neto
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Sustain. Food Syst., 17 May 2023
Sec. Agroecology and Ecosystem Services
Volume 7 - 2023 | https://doi.org/10.3389/fsufs.2023.1142623
This article is part of the Research TopicCrop Pest Control and Pollination, volume IIView all 14 articles
Introduction: Blueberry is one of the most relevant buzz-pollinated crops worldwide, and Chile is the most important global producer of fresh blueberries during wintertime in the Northern Hemisphere. Non-buzzing bees, such as honey bees, may provide suboptimal services compared with bees capable of buzz pollination. The widely held contention that honey bees are inferior pollinators of blueberries drives the industry to place pressure on governments to allow bumblebee (Bombus terrestris) importation for pollination. However, the introduction of B. terrestris generates environmental problems in Chile by competing with and transmitting parasites to local bees. Despite some native Chilean bees being recently recognized as efficient pollen vectors of blueberry crops, no study has evidenced the influence of their visits on fruit yield. Therefore, we aimed to evaluate the native Chilean floral visitors' performance to improve the quantity and quality of highbush blueberry in comparison to the performance of managed visitors.
Methods: Per-visit pollination performance (fruit set and fruit quality) and visitation frequency were measured, and the performance of buzzing behavior by flower visitors was evaluated in four cultivars grown in five blueberry orchards located in southern Chile.
Results: We found that fruit set and weight were highly influenced by floral visitor taxon. Some native bee species can greatly improve the fruit set and fruit quality (greater weight) of the highbush blueberry cultivars. For instance, one single visit of C. occidentalis can increase fruit weight by a factor of 1.8 compared to an A. mellifera visit; however, visits of halictids and syrphids resulted in lower fruit sets than those of unvisited flowers. However, we found that the occurrence of sonication behavior alone was not a predictor of higher fruit set and fruit weight of highbush blueberry cultivars. Consequently, the taxonomic recognition of floral visitors, ideally to the species level, is still needed to distinguish the most efficient fruit yield promoters of blueberry.
Discussion: The conservation of the biotic pollinators, especially native pollinators, would improve blueberry fruit quality and is likely to improve overall crop productivity.
World blueberry production has more than doubled in the last 10 years, reaching 823,328 tons in 2019. Of this, 58% is produced in North America, with38% in the United States and 20% in Canada (FAO, 2020). Following the same trend, the area of Chilean blueberry cultivation has increased rapidly during the last 30 years (Retamales and Hancock, 2018). In 2021, Chile had 18,000 hectares planted with blueberry, exported 118,225 tons of fresh blueberry fruit, and was the third greatest provider of fresh blueberry fruit to the Northern Hemisphere during its winter (Velasco, 2021). However, there are several challenges to producing blueberry fruits of high and consistent quality (Retamales and Hancock, 2018). The number of fruits per plant and their fresh weight are two fundamental factors of blueberry plant yield (Hall et al., 2020). Large and consistent berries increase the market price (Gilbert et al., 2014).
High-quality fruit production by wild and commercial blueberries is highly dependent on the input of biotic pollinators (MacKenzie and Eickwort, 1996; Fulton et al., 2015). Studies in the native range of southern and northern highbush blueberries reveal that the most effective pollinators of blueberries are typically large bees that can extract pollen from anthers by vibrating their bodies during floral visits (Stubbs and Drummond, 1996; Javorek et al., 2002; Campbell et al., 2018). The vibrations produce an audible buzzing sound, which gives the name to this pollination syndrome: buzz pollination or floral sonication (Vallejo-Marín, 2019; Pritchard and Vallejo-Marín, 2020).
The predominant floral trait related to buzzing behavior is the presence of poricidal anthers (Buchmann, 1983; De Luca and Vallejo-Marin, 2013). For flowers with poricidal anther dehiscence, the only exit of pollen from the flower is via tiny openings at the apex of the anthers (Delaplane et al., 2000). This is the case for blueberry flowers and flowers of more than 20,000 species of native and cultivated flowering plants (Buchmann, 1983). Although most buzz-pollinated flowers are nectar-less (Vallejo-Marín et al., 2010), in some cases, such as with blueberry, flowers also produce floral nectar (Javorek et al., 2002), which implies that even a flower visitor capable of sonicating will not necessarily do so if it is looking for nectar. Furthermore, although hives of the honey bee (Apis mellifera) are widely managed for blueberry pollination throughout the world (Benjamin et al., 2014; FAO, 2020), these bees are unable to sonicate and tend to present lower per-visit pollen deposition rates than bees capable of vibrating flowers (Javorek et al., 2002; Rogers et al., 2013; Cortés-Rivas et al., 2023). Thus, honey bees and other non-buzzing bees may provide a suboptimal service compared with other bees capable of buzz pollination (Stubbs and Drummond, 1996; Javorek et al., 2002; Kim et al., 2005; Campbell et al., 2018; Nicholson and Ricketts, 2019; Cooley and Vallejo-Marín, 2021).
The widely held contention that honey bees are inferior pollinators of blueberries drives the industry to place pressure on governments to allow bumblebee (Bombus terrestris) importation for pollination. Managed beehives of B. terrestris achieve high yields due, partially, to their ability to sonicate flowers and be active during environmental conditions unfavorable to honey bees (De Luca et al., 2013; Howlett et al., 2019; Cortés-Rivas et al., 2023). However, the introduction of B. terrestris generates serious problems for the conservation of native pollinators since this species is highly invasive where it is introduced and can compete and transmit parasites to native bees (Matsumura et al., 2004; Dafni et al., 2010; Morales et al., 2013; Smith-Ramírez et al., 2018). As a result, strict laws prohibiting the movement of bumblebees are in place in parts of the USA (Winter et al., 2006), southern Africa, and Australasia (Goka, 2010; Inoue and Yokoyama, 2010) because their introduction could have undesired effects on native fauna and flora (Velthuis and Van Doorn, 2006; Goka, 2010; Inoue and Yokoyama, 2010).
However, the performance of native fauna to pollinate blueberry crops in Chile has recently been revealed, with some bee species having high conspecific pollen transference (Cortés-Rivas et al., 2023). These highly efficient native species would be a more sustainable alternative to B. terrestris importation. Despite this encouraging finding, no study has evidenced the influence of the visits of these native species on fruit yield. Therefore, distinguishing the pollinators associated with higher fruit quantity and quality among the local pool of visitor species is essential to reveal their relative importance for agricultural production (Javorek et al., 2002; Isaacs and Kirk, 2010; Nicholson and Ricketts, 2019). Thus, we aimed to evaluate the native Chilean floral visitors' ability to improve highbush blueberry fruit yield compared to that of managed visitors. We expected that, due to the capacity of some native bee taxa to perform sonication and transfer a high amount of conspecific pollen, their visits would result in higher fruit set and fruit weight in relation to A. mellifera and at least similar to B. terrestris, making them suitable alternative candidates to improve fruit yield of blueberry crops in Chile (Hypothesis 1). Because the occurrence of floral sonication is a predictor of higher pollen deposition on the stigmatic surface of blueberry flowers (Cortés-Rivas et al., 2023), we wanted to know if buzzing behavior was associated with higher fruit quantity and/or quality, so we compared blueberry fruit production between the subset of visits with floral sonication and the subset of visits without this behavior. We chose this approach to focus on the influence of the buzzing behavior itself rather than having to assume that a bee belonging to a taxon capable of vibrating does necessarily vibrate on every visit. We expected that visits to highbush blueberry flowers accompanied by buzzing behavior would be associated with higher fruit set, seed set, and fruit weight than visits without this behavior (Hypothesis 2).
Fieldwork was conducted in five highbush blueberry orchards located in southern Chile (Maule and Los Ríos Regions; Supplementary Figure S1; Table 1) between September and November in 2020 and 2021. The total area of cultivated blueberries per orchard ranged from 3.2 to 141 hectares of both organic and conventional farming. The most common growing cultivars were Legacy, Brigitta, Duke, Elliot, and Draper. Four of the five orchards were supplemented with colonies of managed exotic bees of Bombus terrestris and/or Apis mellifera (Table 1). The local temperature varied between 10°C and 37°C (average 20.2°C), and the wind varied between 0.16 and 8.85 km/h.
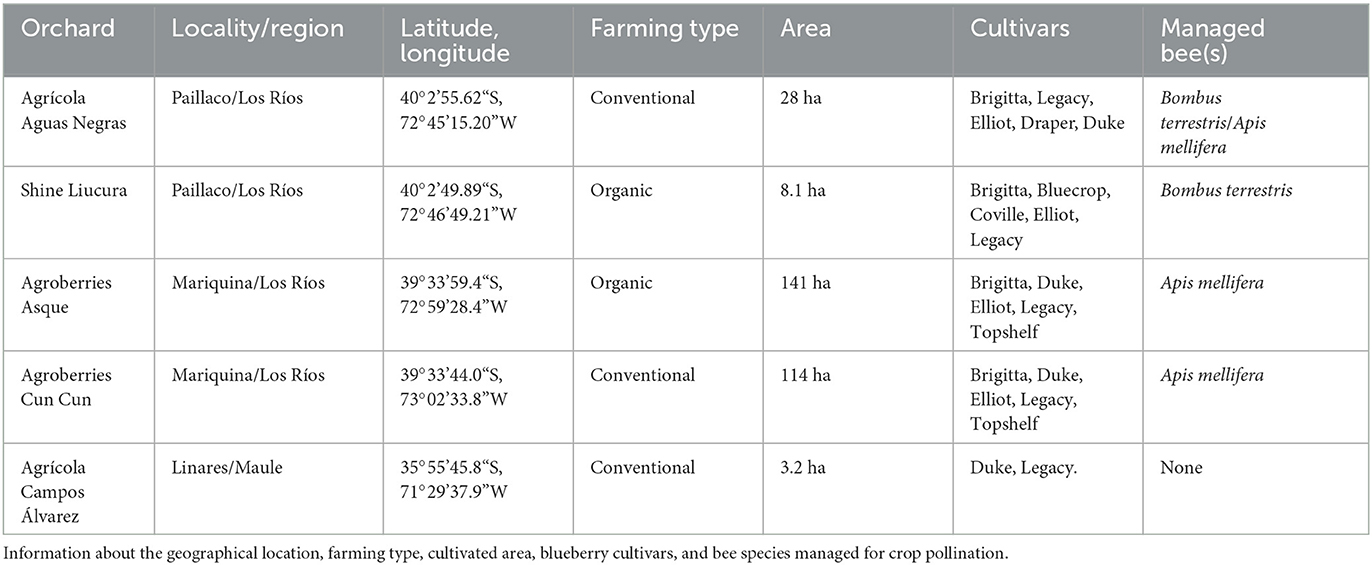
Table 1. Highbush blueberry orchards located in the southern region of Chile (Maule and Los Ríos Regions), where this study was conducted from September to November of 2020 and 2021.
We measured fruit set and seed set based on the single visit test (Ne'eman et al., 2010). To conduct this test, we randomly selected and labeled 10 individual plants per blueberry cultivar/orchard, maintaining a minimum distance of 10 m between each individual plant, as follows: (1) Agrícola Aguas Negras: Legacy, Elliot, Draper, Brigitta, and Duke; (2) Shine Liucura: Legacy, Elliot, and Brigitta; (3) Agroberries Asque: Brigitta, Duke, Elliot, and Legacy; (4) Agroberries Cun Cun: Brigitta, Duke, Elliot, and Legacy; (5) Agrícola Campos Álvarez: Duke and Legacy. For each labeled plant, we enclosed three clusters of flowers in the bud stage in tulle bags after removing all open flowers to ensure that only non-visited flowers remained. After the flower opening, we removed the bags and allowed a single visit to each cluster. We noted whether insects sonicated the flower during flower visits (based on our audible perception of the sound emitted by bees vibrating) and recorded information about individual plant identification, blueberry cultivar, orchard, date, and time. Immediately after an insect concluded its visit, we identified (or collected) it and marked the flower with colored wool yarn (N = 834 flowers). We also marked some non-visited flowers (N = 354 flowers) with wool yarn of a different color (control group) and re-bagged all of the flowers. We, then, killed the collected insects and stored them in individually labeled flasks. In the laboratory, we photographed and identified the insects to the lowest taxonomic level possible.
We kept the flowers bagged for approximately 70 (±3) days and then checked whether the flowers set fruit, and if they did, we immediately weighed each fruit using a precision balance (Dblue® model DBPDWS32S, Valparaíso, Chile; precision 0.001g). In the laboratory, we separated the seeds from the pulp and counted the number of viable seeds (according to Dogterom et al., 2000; Rogers et al., 2013).
We arranged the data according to visitor taxon and according to the occurrence of buzzing behavior during each visit. Data were not normally distributed according to visual inspection with the QQ plot and Shapiro–Wilk tests, so we used non-parametric tests.
We used generalized linear mixed models (GLMMs) to determine whether the fruit set (binomial distribution, link = “logit”) differed among floral visitor taxa and the control (non-visited flowers) (function “glmmTMB”, “glmmTMB” package, Brooks et al., 2017). We built another GLMM to determine whether fruit sets differed between visits with and without sonication, among study orchards, and blueberry cultivars. Individual plants (10 individual plants per blueberry cultivar/orchard) were included as the random factor for these models.
To analyze how fruit weight (response variable) was influenced by pollinator taxon, visits with and without sonication, study orchard identification, blueberry cultivar, and the interaction between pollinator taxon and blueberry cultivar, we used a GLMM with a gamma distribution using the glmmTMB function in the glmmTMB package (Magnusson et al., 2019). To analyze if the number of seeds per fruit (count response variable) was influenced by pollinator taxon, sonication, blueberry cultivar, and the interaction between pollinator taxon and blueberry cultivar, we used a GLMM with a zero-inflated negative binomial distribution (“nbinom1”) using the glmmTMB function in the glmmTMB package (Magnusson et al., 2019). We included the orchard name (as shown in Table 1) as a random factor for these models.
We assessed the fit of all models using the diagnostic plots of the “DHARMa” package (Hartig, 2020). We tabulated estimated marginal means for each visitor taxon using the R package “emmeans” (Lenth, 2019) and tested the significance of pairwise differences by the Tukey method. We specified the type II Wald chi-square (χ2) tests via the Anova() function in the car package (Fox et al., 2018). We performed all statistical analyses with R software version 4.0.3 (R Core Team, 2017).
During 1,056 h of sampling effort for the single-visit test, we recorded 927 visits of 18 insect species to flowers of highbush blueberry cultivars in the five studied orchards. Among the visiting species, 15 were native and three were exotic/managed. Buzzing behavior (sonication) occurred in 45.6% of all visits. Only bees were observed performing buzzing behavior, and among them, the native species were Cadeguala occidentalis (sonicated in 68.4% of visits) and Colletes cyanescens (sonicated in 50% of visits), and one managed species was Bombus terrestris (sonicated in 56.7% of visits). We pooled as “other native bees” (sonicated in 9.1% of visits) the native bee species that did not reach a minimum of 10 samples as follows: Corynura chloris, Corynura sp., Centris cineraria, Caenohalictus sp., and Ruizantheda proxima. Moreover, two native (B. dahlbomii and C. nigritulus) and two exotic (B. ruderatus and A. mellifera) bee species were not observed performing buzzing behavior. Similarly, the remaining floral visitor species (three flower flies, Syrphidae) were not observed performing buzzing behavior during visits to blueberry flowers. We pooled in “syrphids” all the flower flies because none of them reached the minimum of 10 samples (Figure 1).
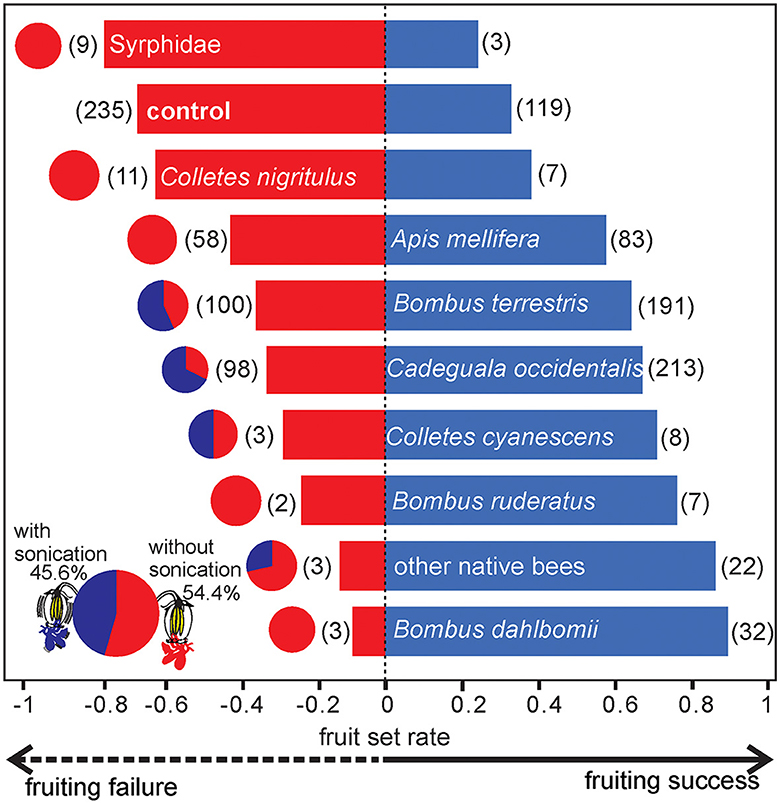
Figure 1. Proportion of fruit set failure (flowers that did not convert into fruits, red colored) and fruit set success (flowers that converted into fruits, blue colored) of single-visited flowers of highbush blueberry cultivars in five orchards in the southern region of Chile. The pie charts represent the proportion of visits with floral sonication (blue) and without (red) in relation to all visits observed of a visitor species. The number between brackets indicates the number of individuals observed (N) per species. Other native bees include bees that did not reach ten samples: Corynura chloris, Corynura sp., Centris cineraria, Caenohalictus sp., Ruizantheda proxima.
Overall, 60.4% of all visited flowers and 33.6% of unvisited flowers (bagged flowers, control) of highbush blueberry set fruit. However, the fruit set was influenced by the floral visitor taxon (glmmTMB: χ2 = 19.4, df = 10, p < 0.05, N = 1281). Visits of both native bees (especially B. dahlbomii, C. cyanescens, C. occidentalis, C. nigritulus, and “other native bees”) and exotic/managed bees (B. ruderatus, B. terrestris, and A. mellifera) produced proportionally more fruits than the control (Figure 1). On the other hand, visits by syrphids set fewer fruits than the control (Figure 1). However, none of these contrasts were statistically significant, except that those flowers visited by C. occidentalis and B. terrestris set more fruits than those visited by syrphids (Supplementary Table S2).
Similarly, the fruit set was also dependent on the type of blueberry cultivar (glmmTMB: χ2 = 12.89, df = 4, p < 0.01): 75.2% and 76.5% of all tested flowers of Elliott and Duke, respectively, featured fruit set, while 56.9%, 50%, and 43.6% of the flowers of Legacy, Draper, and Brigitta featured fruit sets, respectively. However, only the contrast between Draper and Elliott presented a significant difference (Odds ratio = 0.13, t-ratio = −2.838, p = 0.0374; Supplementary Table S3). However, the presence/absence of sonication behavior (glmmTMB: χ2 = 0.44, df = 1, p = 0.5074) and orchard ID (glmmTMB: χ2 = 3.35, df = 3, p = 0.3403) did not influence fruit set.
Fruit weight for single-visited flowers of highbush blueberry depended on floral visitor taxon (glmmTMB: χ2 = 99.5, df = 7, p < 0.001, N = 481). Flowers visited by C. occidentalis (Mean ± SD; 1.20 ± 0.87g, N = 213) set the heaviest fruits, followed by those visited by B. terrestris (0.86 ± 0.59g, N = 191) and A. mellifera (0.76 ± 0.78g, N = 83; Figure 2). However, there were no significant differences among these three bee species. Additionally, flowers visited by any of these species produced heavier fruits than unvisited flowers (control, 0.40 ± 0.34g, N = 119; Supplementary Table S4). On the other hand, flower visits of the native bees Corynura herbsti and Lasioglossum sp. resulted in lighter fruits than those of the control; visits of “other native bees” set fruits with no difference in weight from those of the control.
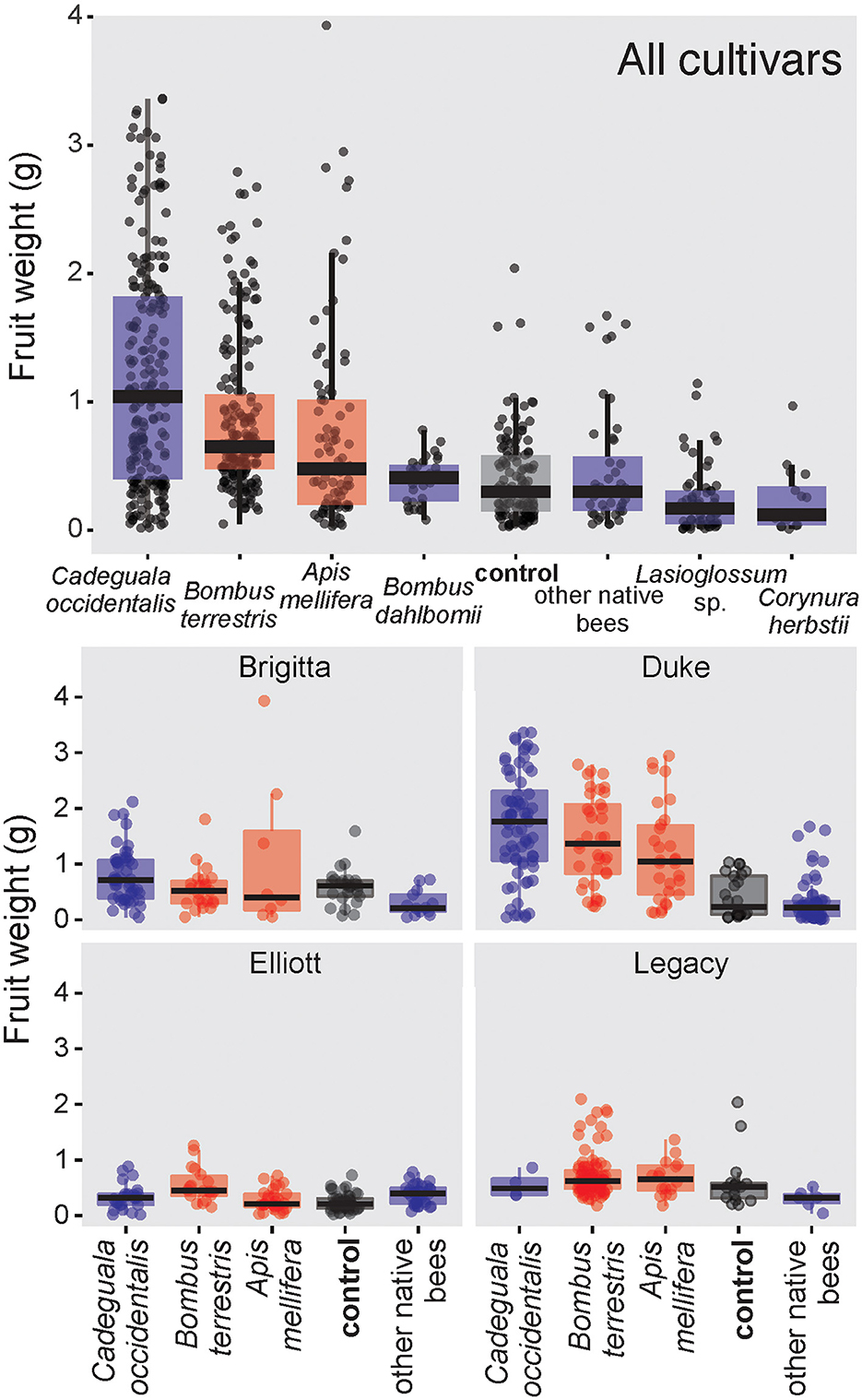
Figure 2. Boxplots comparing the effect of the pollinator taxon on the highbush blueberry fruit weight (grams) based on single-visit tests in five orchards in the southern region of Chile. The top graphic considers the effect of pollinator taxon on fruit weight of all blueberry cultivars together, and the four bottom graphics show the effect per highbush blueberry cultivar. The boxes are colored according to the geographic origin of the visiting insects: native (blue), exotic (red), and control (black). Other native bees include bees that did not reach ten samples: Corynura chloris, Corynura sp., Centris cineraria, Caenohalictus sp., Ruizantheda proxima, Colletes cyanescens, Colletes nigritulus.
Fruit weight was also influenced by the blueberry cultivar (glmmTMB: χ2 = 219.389, df = 4, P < 0.001). Elliott set the lightest fruits among all cultivars, followed by Brigitta with fruits approximately two times heavier than Elliott. The fruits of Brigitta did not differ from those of Legacy (Supplementary Table S5). Duke and Draper set the heaviest fruits with no difference between them (Supplementary Table S5). The fruits of Duke and Draper could be on average 4.3 times heavier than those from Elliott and almost double the weight of the fruits of the Legacy cultivar (Supplementary Table S5).
Fruit weight was also influenced by the interaction between pollinator taxon and blueberry cultivar (glmmTMB: χ2 = 62.3, df = 12, P < 0.001; Figure 2). The flowers of Brigitta visited by “other native bees” (composed mostly of small halictid bees) set lighter fruits than those visited by C. occidentalis and the unvisited flowers (control, Table 2; Figure 2). Comparisons among the other pollinator taxa and control were not significant for the Brigitta cultivar (Supplementary Table S6).
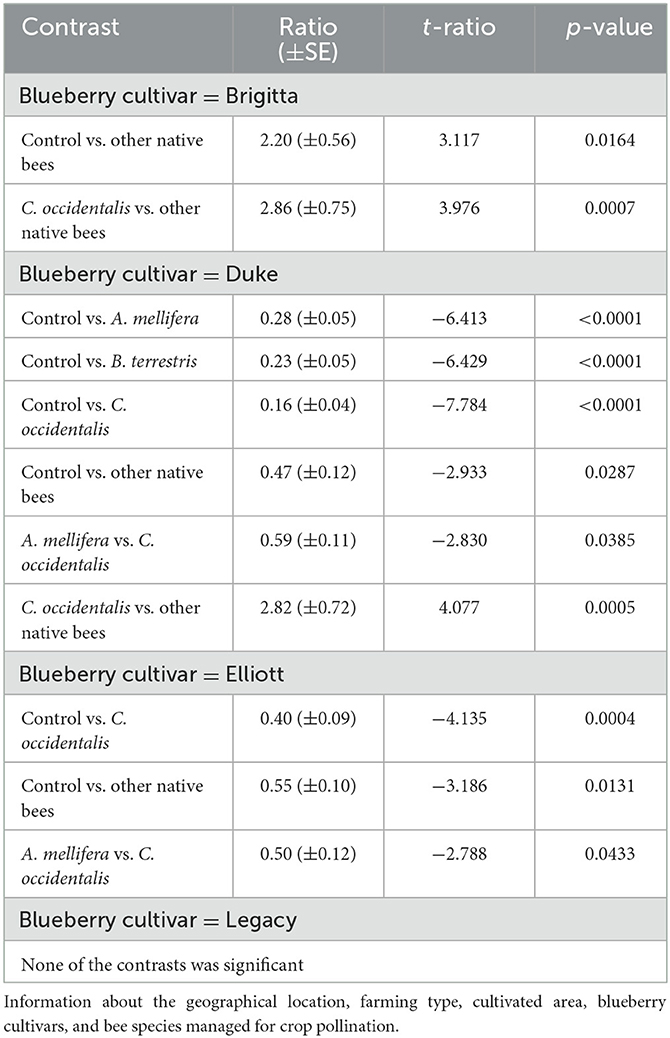
Table 2. Pairwise contrasting of the interaction between visitor taxon and highbush blueberry cultivar for fruit weight in five orchards of the southern region of Chile (gamma distribution, glmmTMB).
The flowers of the Duke cultivar visited by any bee species set heavier fruits than the control of unvisited flowers (Figure 2, Table 2). The floral visits of C. occidentalis produced the heaviest fruits of Duke with a significant difference from other bee species, except B. terrestris. The other comparisons were not statistically significant (see Figure 2; Supplementary Table S6). In the case of the Elliott cultivar, only pollination by C. occidentalis and “other native bees” resulted in heavier fruits than the control (Table 2); flowers visited by A. mellifera set lighter fruits than those of C. occidentalis (Figure 2). The other comparisons for Elliott were not significant (Table 2). Finally, the pollinator taxon did not influence fruit weight, and no contrasts were significant for the Legacy cultivar (Figure 2; Supplementary Table S6).
Fruit weight did not differ significantly between flowers visited with and without buzzing behavior (glmmTMB: χ2 = 0.90, df = 1, P = 0.3419).
Seed set varied according to pollinator taxon (glmmTMB: χ2 = 122.03, df = 4, P < 0.001, N = 595), highbush blueberry cultivar type (glmmTMB: χ2 = 127.10, df = 3, P < 0.001, N = 595), and the interaction between these two variables (glmmTMB: χ2 = 30.2, df = 12, P < 0.005, N = 595). Flowers pollinated by B. terrestris (Mean+SD: 21.7 ± 12.6) and C. occidentalis (10.3 ± 11.1 seeds/fruit) set fruits with more seeds than unvisited flowers (control, Figure 3), without a difference between them (Supplementary Table S7). Flowers pollinated by these two bees set fruits with more seeds than did flowers pollinated by A. mellifera (3.4 ± 5.3 seeds per fruit) and “other native bees” (3.4 ± 5.3). In addition, flowers visited by A. mellifera and “other native bees” set fruits with a similar number of seeds to those of the control (Supplementary Table S7); here, “other native bees” includes B. dahlbomii because it did not reach the minimum number of samples (N < 10).
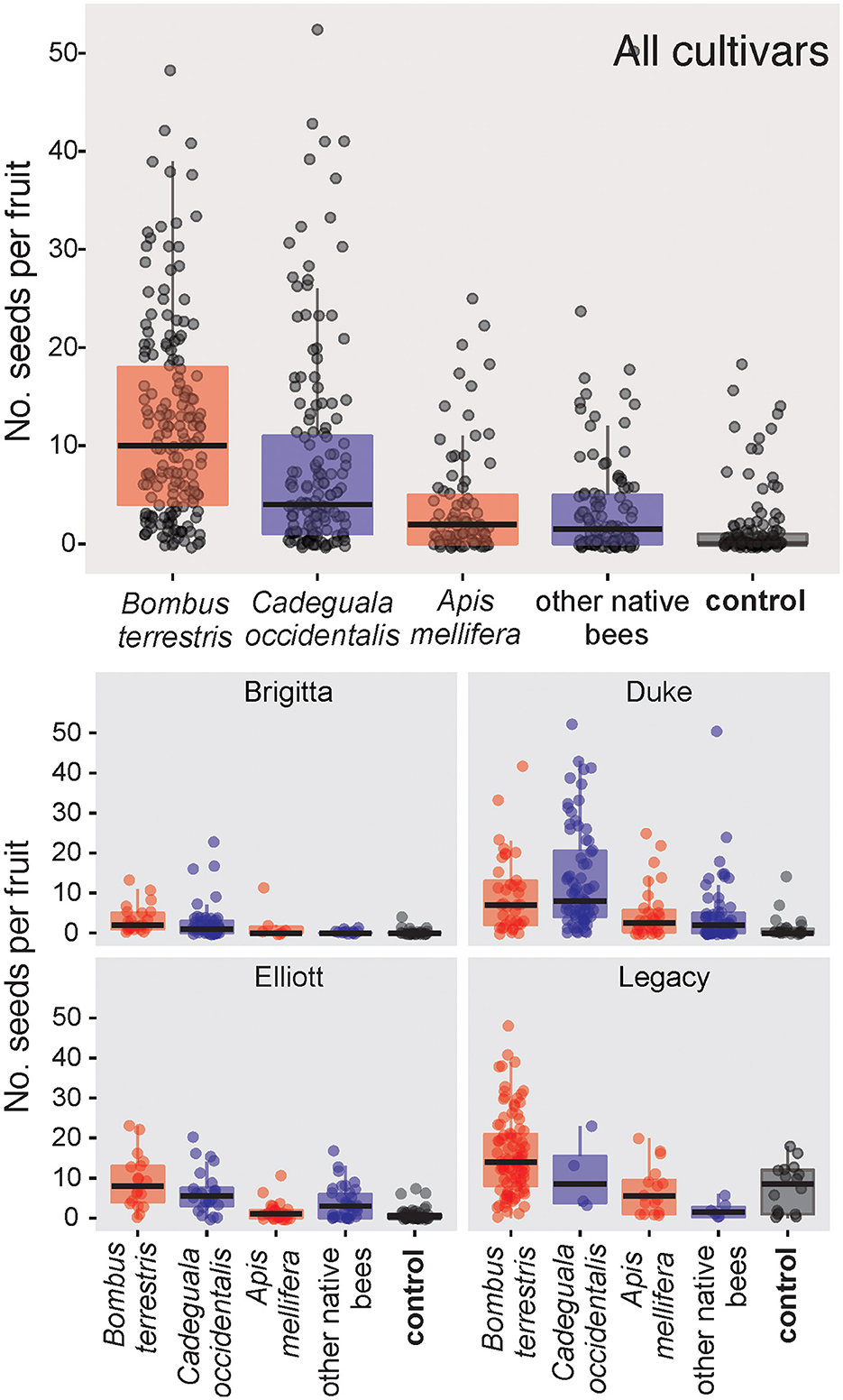
Figure 3. Boxplots comparing the effect of the pollinator taxon on the seed set of highbush blueberry cultivars based on single-visit tests in five orchards in the southern region of Chile. The top graphic considers the effect in all blueberry cultivars together, and the four bottom graphics show the effect per highbush blueberry cultivar. Boxes are colored according to the geographic origin of the visiting insects: native (blue), exotic (red), and control (black). “Other native bees” includes bees that did not reach ten samples: Bombus dahlbomii, Corynura chloris, Corynura sp., Centris cineraria, Caenohalictus sp., Ruizantheda proxima, Colletes cyanescens, Colletes nigritulus, Lasioglossum sp., Corynura herbsti.
Legacy was the cultivar that produced the greatest seed set per fruit among all the tested cultivars, while Brigitta produced the least (Supplementary Table S8). The remaining cultivars (Duke and Elliott) did not differ significantly for the seed set (Supplementary Table S8).
The interaction between the blueberry cultivar and pollinator taxon was also significant (glmmTMB: χ2 = 30.2, df = 12, P < 0.005, N = 595; Figure 3). Flowers visited by B. terrestris set more seeds than the control for all four cultivars (Table 3). However, flowers visited by C. occidentalis set more seeds than the control in three of the four cultivars (except Legacy, Table 3, Supplementary Table S9). Despite visits of B. terrestris resulting in a greater average seed set, there was no difference with C. occidentalis among the blueberry cultivars (Supplementary Table S9). On the other hand, “other native bees” only showed a positive effect compared to the control for the Elliott cultivar and presented a negative effect or no difference when compared to the control for the other cultivars.
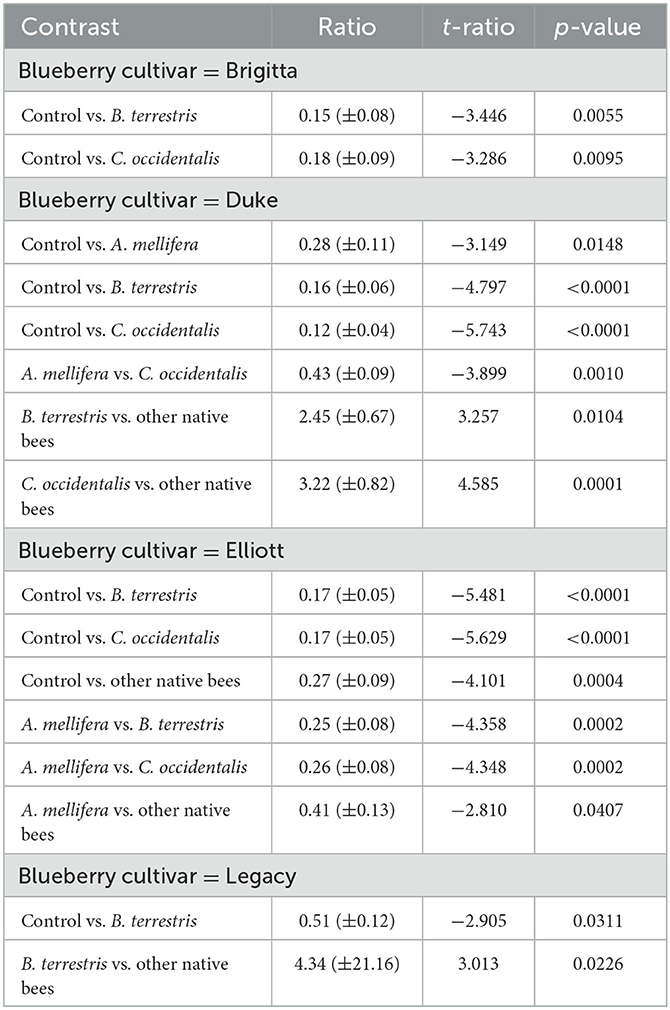
Table 3. Pairwise contrasting of the effect of the interaction between visitor taxon and highbush blueberry cultivar on seed set in five orchards of the southern region of Chile (nbinom1 distribution, glmmTMB).
All cultivars presented a strong positive correlation between seed set and fruit weight (Figure 4). However, the influence of seeds per fruit was stronger for Brigitta (R = 0.53, p < 0.001) and Duke (R = 0.79, p < 0.001), while for Elliot (R = 0.64, p < 0.001) and Legacy (R = 0.57, p < 0.001), it was weaker, especially for Legacy, which produced the largest seed set but without a proportional increase in fruit weight (Figure 4).
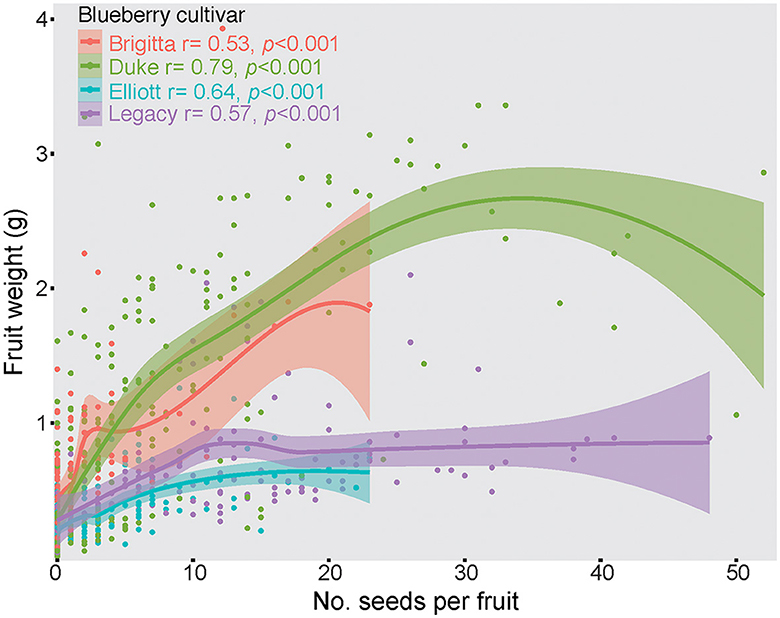
Figure 4. Non-linear correlation between the number of seeds per fruit and the fruit weight (grams) of four highbush blueberry cultivars in five orchards southern region of Chile.
Despite the occurrence of floral sonication being a predictor of higher pollen deposition on the stigmatic surface (see Cortés-Rivas et al., 2023), we found that the occurrence of this behavior alone was not a predictor of higher fruit set and fruit weight of highbush blueberry cultivars. Instead, fruit sets and fruit quality improvement are more dependent on the taxonomic identity of the visiting species. Among them, some native bees can greatly improve the fruit set and fruit quality (greater weight) of highbush blueberry cultivars. Therefore, the taxonomic identity of floral visitors, ideally at the species level, is still needed to distinguish the most efficient promoters of blueberry fruit yield.
As previously expected, the exclusion of biotic pollinators reduced the fruit set of highbush blueberry cultivars. However, the capacity of floral visitors to pollinate varied greatly, with only a subset of all floral visitors actually pollinating the flowers and promoting the fruit set. The flowers that were single-visited by native and managed bees, especially bumblebees (Bombus dahlbomii, B. ruderatus, and B. terrestris) and medium-sized solitary bees (C. cyanensis, C. occidentalis, and other native bees), set a higher number of fruits compared to the control of unvisited flowers, and are likely to increase fruit set. Among them, the giant Patagonian bumblebee (B. dahlbomii) achieved the highest proportion of fruit sets, with more than 90% of flowers visited setting fruits.
The high efficiency to set fruits of blueberry could be a new piece of evidence to enhance the functional role of B. dahlbomii as a relevant crop pollinator and highlights the urgent need to implement conservation strategies because of the delicate conservation status of this bumblebee (Morales et al., 2013; Smith-Ramírez et al., 2018; Henríquez-Piskulich et al., 2021). This bumblebee is classified as a threatened species by the IUCN Red List with declining populations (Morales et al., 2016), and its threat has been attributed to, among other factors, the introduction of B. terrestris hives for crop pollination, especially blueberries (Morales et al., 2013; Smith-Ramírez et al., 2018). Therefore, strategies to enhance crop pollination with this exotic species necessarily need to deal with the fragile conservation status of B. dahlbomii. These strategies could include integrative habitat management and more sustainable alternatives to replace the importation of B. terrestris colonies.
In addition to the effect on fruit set, fruit quality and seed set were strongly pollinator-dependent. Fruits from flowers visited by pollinators can be three times heavier and set up to 6.3 times more seeds than those from non-visited flowers. This is particularly relevant for the farmer since fruit quality (heavier and larger fruits) has direct consequences on market price (Gilbert et al., 2014; Retamales and Hancock, 2018; Hall et al., 2020). The ground-nesting solitary bee C. occidentalis achieved high performance as a pollinator of blueberry and is similar to the exotic and managed bumblebee (B. terrestris). Both of these species are sonicating bees. Surprisingly, we found that a single visit of C. occidentalis can increase fruit weight by 5.8 times over that of non-visited flowers and 1.8 times over those visited by A. mellifera. Cadeguala occidentalis, commonly found visiting other crops (Monzón et al., 2020; López-Aliste et al., 2021; Smith-Ramírez et al. in review), is widely distributed in Chile (GBIF Secretariat, 2021) and has known nesting behavior and biology (Torchio and Burwell, 1987; Montalva et al., 2011; López-Aliste et al., 2021). However, key aspects of the biology of C. occidentalis, especially its solitary behavior and ground-nesting, need to be taken into consideration to enhance blueberry pollination with this bee species. Grazing, for example, can destroy nest sites, remove forage plants, and alter the structure, diversity, and growth habits of the plant community (Black et al., 2011). Despite prescribed burns being beneficial for maintaining some ecosystems, they also significantly reduce ground-nesting bee populations (Black et al., 2011).
Despite the occurrence of buzzing behavior (or floral sonication) being related to higher conspecific pollen deposition on stigmas of highbush blueberry (Cortés-Rivas et al., 2023), we found that floral sonication did not have significant effects on fruit set and fruit weight. Instead, fruit weight was particularly dependent on pollinator taxon, even more than on the presence/absence of buzzing behavior. Buzzing behavior is widespread and occurs among many bee taxa, including bees that differ greatly in morphology and foraging behavior (Cardinal et al., 2018). For example, some small bees capable of performing buzzing behavior (Lasioglossum sp. and C. chloris) were less efficient pollinators of the studied highbush blueberry orchards, their visits only resulting in low or no fruit set. This must be, among other factors, because their body is not large enough to touch the stigmatic region while searching for nectar and pollen on the flower (Solís-Montero and Vallejo-Marín, 2017; Földesi et al., 2021). Therefore, bee body size relative to blueberry flower size and foraging behavior must be among the key co-factors affecting pollination efficiency (Solís-Montero and Vallejo-Marín, 2017; Morais et al., 2020; Földesi et al., 2021; Mesquita-Neto et al., 2021). The bees that were notably equal to or larger in size than the anther–stigma distance achieved greater performance at setting fruit with the highest weight (e.g., B. terrestris, B. dahlbomii, C. occidentalis, and Colletes spp.). However, we did not measure the body size of individual bees nor the anther–stigma distance of blueberry flowers, which could be a better predictor of fruit yield than the buzzing behavior alone and could be addressed by further studies. Therefore, our results cannot support our hypothesis that the buzzing behavior alone is a predictor of higher fruit set and weight. Instead, species identity remains a better predictor of the quantity and quality of blueberry fruits.
Like bees, some syrphids (flower flies) can generate vibrations of sufficient amplitude to extract pollen from buzz-pollinated flowers (Vallejo-Marín and Vallejo, 2021). However, we have not observed flies performing floral sonication in blueberry flowers. In fact, only one case of floral sonication by flower flies has been reported, involving a Mexican cactus fly (Volucella mexicana) in the flowers of a Solanum plant (Buchmann et al., 1978). The absence of this behavior is explained by the fact that the amount of pollen required by a flower fly is much less than that required by a bee (Larson et al., 2001). Syrphids usually do not feed their offspring with pollen, and their larvae consume other resources (Diaz et al., 2020). Pollen consumption is, then, only related to the diet of adult flies (Larson et al., 2001). In addition, there is very little information on the role of flower flies and other flies in the pollination and fruit production of blueberries in Chile or elsewhere. (Cook et al., 2020; Cortés-Rivas et al., 2023). Mann (2014) suggested that blowflies (Calliphoridae) may be effective pollinators due to their ability to sonicate, but no direct evidence of pollination efficiency was presented. To our knowledge, only the study by Cook et al. (2020) provides evidence that the presence of blowflies in greenhouses improves the pollination of blueberries. However, Cortés-Rivas et al. (2023) showed that flower flies were poor pollinators of blueberry plants, depositing few pollen grains on the stigmatic surface per visit. Our results showed that their visits resulted in a similar proportion of fruit compared with unvisited flowers. Although flies are capable of producing floral vibrations, our study provides evidence against the notion that flower flies are effective pollinators of highbush blueberry cultivars but are rather pollen or nectar thieves.
Buzz-pollinated plants and their flower visitors represent a tangible example of the importance of considering bee functional diversity in the pollination of cultivated species (Cooley and Vallejo-Marín, 2021). However, our results evidenced that taxonomic diversity is also a key component since the quality of the pollination provided also depends on the taxon of flower visitors. Consequently, the taxonomic recognition of species becomes indispensable to distinguishing the most efficient pollinators of blueberry (Cortés-Rivas et al., 2023). Therefore, by discriminating true fruit set promoters from nectar/pollen thieves and being aware of the value of bees to crop income, farmers could be encouraged to consider the pollination perspective in their crop management. This could result in the conservation of local wild bee species, thereby contributing to advances toward more sustainable and higher-yield agriculture (Isaacs and Kirk, 2010; Garibaldi et al., 2014; Nicholson and Ricketts, 2019).
Our results showed that some native Chilean pollinating bees (e.g., B. dahlbomii and C. occidentalis) can be much better fruit yield promoters than A. mellifera and equivalent to the exotic bumblebee. However, the potential to enhance blueberry fruit production alone does not justify the immediate abandonment of managed hives to pollinate blueberry cultivars. There are practical challenges associated with supplementing pollination with native pollinators (Cooley and Vallejo-Marín, 2021). One of them is increasing their abundance in crop fields (Isaacs and Kirk, 2010; Garibaldi et al., 2014; Henríquez-Piskulich et al., 2021), as they represented only 11.9% of all visits in the studied blueberry orchards (Cortés-Rivas et al., 2023). Strategies to increase native pollinators can include creating wildflower strips for pollinators and the restoration of hedgerows on farms, along with the conservation of nearby natural and semi-natural habitats (Morandin and Kremen, 2013; Potts et al., 2016; Sutter et al., 2018; Rodríguez et al., 2021). Such efforts would provide nesting habitats and floral resources for the most effective pollinators of blueberry and consequently, the overall native bee community within agricultural ecosystems since they not only visit crops but also depend on native habitats to nest, feed, and reproduce (Nicholson et al., 2017; Monzón and Ruz, 2018; Rodríguez et al., 2021). Second, attention must be taken to ensure that commercial colonies of native bees do not become a problematic resource in their native range (Mallinger et al., 2017). Despite that, we expect that our study will be one more piece of evidence to discourage the importation of B. terrestris hives for blueberry pollination in Chile while incentivizing the development and implementation of integrative habitat management strategies to conserve native bees and consequently enhance crop pollination.
To our knowledge, this is the first study evidencing the per-visit performance of native crop pollinators to enhance fruit quantity and quality in the Chilean environment. Here, we applied field experimental protocols and sampling methods to investigate blueberry pollination and evaluated the relative performance of managed bees and native bees in per-visit-based tests in commercial blueberry fields. Bees, however, vary in their foraging range (Greenleaf et al., 2007; Zurbuchen et al., 2010) and distribution, so examining the bees that visit blueberries over multiple regions and landscape scales can help reveal how these factors affect pollinators and their pollination services. Moreover, sampling bias must be another relevant factor, since a limited number of flowers set fruit. Considering that a plant is typically visited by multiple flower visitors and per visitor replicates are needed, replicates were missing for some flower visitor species.
In summary, our results indicate that visits of native bees can enhance fruit production of highbush blueberry crops in Chile. However, fruit set and fruit quality improvement are more dependent on the taxonomic identity of the visiting species rather than their sonication behavior alone. Some native Chilean bees, especially C. occidentalis and the giant Patagonian bumblebee (B. dahlbomii) were efficient fruit promoters of blueberry and performed better or similar to honeybees (A. mellifera) and the exotic bumblebee. We suppose that the conservation of these native pollinators would result in increased crop yields because their visits enhance blueberry fruit quality and are likely to improve overall crop productivity and sustainability. Therefore, instead of continuing to rent or pay for managed beehives, which represents a high cost for farmers (Isaacs and Kirk, 2010; Henríquez-Piskulich et al., 2021), the service provided by native pollinators should be considered, with priority given to the subset of the most successful pollinators.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
BC-R, JM-N, and VM contributed to the conception and design of the study. BC-R, JR, and JM-N carried out the experiment. JM-N organized the database and performed the data and statistical analysis. BC-R and JM-N wrote the first draft of the manuscript. JR and VM wrote sections of the manuscript. All authors contributed to the manuscript revision and read and approved the submitted version.
This study was supported by the ANID/Fondecyt Iniciación en Investigación under Grant No. 11190012, FIC GORE Maule under Grant No. BIP 40.019.177-0, and ANID/CONICYT FONDECYT Regular under Grant No. 1231212.
We thank the staff of Agrícola Aguas Negras S.A., Shine Liucura, Agroberries S.A., and Agrícola Campos Álvarez, especially Claudio Troncoso, André Didier, Gabriel Esteyes, and Freddy Campos, for their assistance during the fieldwork.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fsufs.2023.1142623/full#supplementary-material
Benjamin, E. F. R., Reilly, J., and Winfree, R. (2014). Pollinator body size mediates the scale at which land use drives crop pollination services. J. Appl. Ecol. 51, 440–449. doi: 10.1111/1365-2664.12198
Black, S. H., Shepherd, M., and Vaughan, M. (2011). Rangeland management for pollinators. Rangelands. 33, 9–13. doi: 10.2111/1551-501X-33.3.9
Brooks, M. E., Kristensen, K., Van Benthem, K. J., Magnusson, A., Berg, C. W., Nielsen, A., et al. (2017). glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 9, 378–400. doi: 10.32614/RJ-2017-066
Buchmann, S. L. (1983). Buzz Pollination in Angiosperms (New York, NY: Van Nostrand Reinhold Company), 73–113.
Buchmann, S. L., Jones, C. E., and Colin, L. J. (1978). Vibratile pollination of Solanum douglassii Dunal and S. Xanti Gray (Solanaceae) in Southern California. Wasmann J. Biol. 35, 1.
Campbell, J. W., Kimmel, C. B., Bammer, M., Stanley-Stahr, C., Daniels, J. C., and Ellis, J. D. (2018). Managed and wild bee flower visitors and their potential contribution to pollination services of low-chill highbush blueberry (Vaccinium corymbosum L.; Ericales: Ericaceae). J. Econ. Entomol. 111, 2011–2016. doi: 10.1093/jee/toy215
Cardinal, S., Buchmann, S. L., and Russell, A. L. (2018). The evolution of floral sonication, a pollen foraging behavior used by bees (Anthophila). Evolution. 72, 590–600. doi: 10.1111/evo.13446
Cook, D. F., Deyl, R. A., Mickan, B. S., and Howse, E. T. (2020). Yield of southern highbush blueberry (Vaccinium corymbosum) using the fly Calliphora albifrontalis (Diptera: Calliphoridae) as a pollinator. Austral Entomol. 59, 345–352. doi: 10.1111/aen.12455
Cooley, H., and Vallejo-Marín, M. (2021). Buzz-pollinated crops: a global review and meta-analysis of the effects of supplemental bee pollination in tomato. J. Econ. Entomol. 114, 505–519. doi: 10.1093/jee/toab009
Cortés-Rivas, B., Smith-Ramirez, C., Monzón, V. H., and Mesquita-Neto, J. N. (2023). Native bees with floral sonication behaviour can achieve high-performance pollination of highbush blueberry in Chile. Agric. For. Entomol. 25, 91–102. doi: 10.1111/afe.12533
Dafni, A., Kevan, P., Gross, C. L., and Goka, K. (2010). Bombus terrestris, pollinator, invasive and pest: An assessment of problems associated with its widespread introductions for commercial purposes. Appl. Entomol. Zool. 45, 101–113. doi: 10.1303/aez.2010.101
De Luca, P. A., Bussiere, L. F., Souto-Vilaros, D., Goulson, D., Mason, A. C., and Vallejo-Marín, M. (2013). Variability in bumblebee pollination buzzes affects the quantity of pollen released from flowers. Oecologia. 172, 805–816. doi: 10.1007/s00442-012-2535-1
De Luca, P. A., and Vallejo-Marin, M. (2013). What's the 'buzz' about? The ecology and evolutionary significance of buzz pollination. Curr. Opin. Plant Biol. 16, 429–435. doi: 10.1016/j.pbi.2013.05.002
Delaplane, S., Keith, D., and Mayer, F. (2000). Crop Pollination by Bees. New York, New York: CABI Publishing. p. 344. doi: 10.1079/9780851994482.0000
Diaz, B. M., Maza, N., Castresana, J. E., and Martínez, M. A. (2020). Los sírfidos como agentes de control biológico y polinización en horticultura. Buenos Aires: Ediciones INTA, Estación Experimental Agropecuaria Concordia. p. 1–9.
Dogterom, M. H., Winston, M. L., and Mukai, A. (2000). Effect of pollen load size and source (self, outcross) on seed and fruit production in highbush blueberry cv.‘Bluecrop'(Vaccinium corymbosum; Ericaceae). Am. J. Bot. 87, 1584–1591. doi: 10.2307/2656734
FAO (2020). Available online at: http://www.fao.org/faostat/en/#home (accessed December 15, 2020).
Földesi, R., Howlett, B. G., Grass, I., and Batáry, P. (2021). Larger pollinators deposit more pollen on stigmas across multiple plant species—A meta-analysis. J. Appl. Ecol. 58, 699–707. doi: 10.1111/1365-2664.13798
Fox, J., Weisberg, S., Price, B., Adler, D., Bates, D., Baud-Bovy, G., et al (2018). Package ‘Car'. R Foundation for Statistical Computing, Vienna, Austria. Available online at: https://r-forge.r-project.org/projects/car/ (accessed October 20, 2022).
Fulton, M., Jesson, L. K., Bobiwash, K., and Schoen, D. J. (2015). Mitigation of pollen limitation in the lowbush blueberry agroecosystem: effect of augmenting natural pollinators. Ecosphere. 6, 1–19. doi: 10.1890/ES15-00148.1
Garibaldi, L. A., Carvalheiro, L. G., Leonhardt, S. D., Aizen, M. A., Blaauw, B. R., Isaacs, R., et al. (2014). From research to action: enhancing crop yield through wild pollinators. Front. Ecol. Environ. 12, 439–447. doi: 10.1890/130330
GBIF Secretariat (2021). “Cadeguala occidentalis (Haliday, 1836),” in GBIF Backbone Taxonomy (Copenhagen: GBIF Secretariat).
Gilbert, J. L., Olmstead, J. W., Colquhoun, T. A., Levin, L. A., Clark, D. G., and Moskowitz, H. R. (2014). Consumer-assisted selection of blueberry fruit quality traits. HortScience. 49, 864–873. doi: 10.21273/HORTSCI.49.7.864
Goka, K. (2010). Introduction to the special feature for ecological risk assessment of introduced bumblebees: status of the European bumblebee, Bombus terrestris, in Japan as a beneficial pollinator and an invasive alien species. Appl. Entomol. Zool. 45, 1–6. doi: 10.1303/aez.2010.1
Greenleaf, S. S., Williams, N. M., Winfree, R., and Kremen, C. (2007). Bee foraging ranges and their relationship to body size. Oecologia. 153, 589–596. doi: 10.1007/s00442-007-0752-9
Hall, M. A., Jones, J., Rocchetti, M., Wright, D., and Rader, R. (2020). Bee visitation and fruit quality in berries under protected cropping vary along the length of polytunnels. J. Econ. Entomol. 113, 1337–1346. doi: 10.1093/jee/toaa037
Hartig, F. (2020). DHARMa: Residual Diagnostics for Hierarchical Regression Models. Available online at: http://florianhartig.github.io/DHARMa/ (accessed August 20, 2022).
Henríquez-Piskulich, P. A., Schapheer, C., Vereecken, N. J., and Villagra, C. (2021). Agroecological strategies to safeguard insect pollinators in biodiversity hotspots: Chile as a case study. Sustainability. 13, 6728. doi: 10.3390/su13126728
Howlett, B. G., Lankin-Vega, G. O., and Jesson, L. K. (2019). Bombus terrestris: a more efficient but less effective pollinator than Apis mellifera across surveyed white clover seed fields. N. Z. J. Crop Hortic. Sci. 47, 32–47. doi: 10.1080/01140671.2018.1466341
Inoue, M. N., and Yokoyama, J. (2010). Competition for flower resources and nest sites between Bombus terrestris (L.) and Japanese native bumblebees. Appl. Entomol. Zool. 45, 29–35. doi: 10.1303/aez.2010.29
Isaacs, R., and Kirk, A. K. (2010). Pollination services provided to small and large highbush blueberry fields by wild and managed bees. J. Appl. Ecol. 47, 841–849. doi: 10.1111/j.1365-2664.2010.01823.x
Javorek, S. K., Mackenzie, K. E., and Vander Kloet, S. P. (2002). Comparative pollination effectiveness among bees (Hymenoptera: Apoidea) on lowbush blueberry (Ericaceae: Vaccinium angustifolium). Ann. Entomol. Soc. Am. 95, 345–351. doi: 10.1603/0013-8746(2002)0950345:CPEABH2.0.CO;2
Kim, Y., Jo, Y., Lee, S., Lee, M., Yoon, H., Lee, M., et al. (2005). The comparison of pollinating effects between honey bees (Apis mellifera) and bumblebee (Bombus terrestris) on the Kiwifruit raised in greenhouse. Korean J. Apicult. 20: 47–52 doi: 10.17519/apiculture.2019.11.34.4.295
Larson, B. M. H., Kevan, P. G., and Inouye, D. W. (2001). Flies and flowers: taxonomic diversity of anthophiles and pollinators. Can. Entomol. 133, 439–465. doi: 10.4039/Ent133439-4
Lenth, R. (2019). Emmeans: Estimated marginal means, aka least-squares means. R Package Version 1.4.7 [Computer software]. Available online at: https://CRAN.R-project.org/package=emmeans (accessed October 21, 2022).
López-Aliste, M., Flores-Prado, L., Ruz, L., Sepúlveda, Y., Rodríguez, S., Saraiva, A. M., et al. (2021). Wild bees of Chile: a database on taxonomy, sociality, and ecology. Ecology. 102, e03377. doi: 10.1002/ecy.3377
MacKenzie, K. E., and Eickwort, G. C. (1996). Diversity and abundance of bees (Hymenoptera: Apoidea) foraging on highbush blueberry (Vaccinium corymbosum L.) in central New York. J. Kans. Entomol. Soc. 185–194.
Magnusson, A., Skaug, H., Berg, C., Kristensen, K., Maechler, M., van Bentham, K., et al (2019). glmmTMB. CRAN. Available online at: https://cran.r-project.org/web/packages/glmmTMB/index.html (accessed July, 2022).
Mallinger, R. E., Gaines-Day, H. R., and Gratton, C. (2017). Do managed bees have negative effects on wild bees?: a systematic review of the literature. PLoS ONE. 12, e0189268. doi: 10.1371/journal.pone.0189268
Mann, N. A. (2014). Intensive Berry Production Using Greenhouses, Substrates and Hyrdoponics. Is This This Way Forward. North Sydney, NSW, Australia: A Report for Nuffield Australia Farming Scholars:
Matsumura, C., Yokoyama, J., and Washitani, I. (2004). Invasion status and potential ecological impacts of an invasive alien bumblebee, Bombus terrestris L.(Hymenoptera: Apidae) naturalized in Southern Hokkaido, Japan. Global Env. Res. 8, 51–66.
Mesquita-Neto, J. N., Vieira, A. L. C., and Schlindwein, C. (2021). Minimum size threshold of visiting bees of a buzz-pollinated plant species: consequences for pollination efficiency. Am. J. Bot. 108, 1006–1015 doi: 10.1002/ajb2.1681
Montalva, J., Sepúlveda, Y., and Baeza, R. (2011). Cadeguala occidentalis (Haliday, 1836) (Hymenoptera: Colletidae: Diphaglossinae): Biología de nidificación y morfología de los estados inmaduros. Boletín de Biodiversidad de Chile. 5, 3–21.
Monzón, V., and Ruz, L. (2018). Estudio de Caso: Determinación del Servicio Ecosistémico de Polinización de Artrópodos Nativos en Agroecosistemas de la Localidad de Caleu, Til-Til. Santiago, Chile: Ministerio del Medio Ambiente, ONU Medio ambiente. Available online at: https://gefmontana.cl/wp-content/uploads/2020/05/Determinaci%C3%B3n-del-Servicio-de-Polinizaci%C3%B3n-de-Artr%C3%B3podos-Nativos-en-Agroecosistemas-de-la-Localidad-de-Caleu-Til-Til.pdf. (accessed August 15, 2022).
Monzón, V. H., Avendaño-Soto, P., Araujo, R. O., Garrido, R., and Mesquita-Neto, J. N. (2020). Avocado crops as a floral resource for native bees of Chile. Revista chilena de historia natural. 93, 1–7. doi: 10.1186/s40693-020-00092-x
Morais, J. M., Consolaro, H. N., Bergamini, L. L., and Ferrero, V. (2020). Patterns of pollen flow in monomorphic enantiostylous species: the importance of floral morphology and pollinators' size. Plant Syst. Evol. 306, 1–12. doi: 10.1007/s00606-020-01627-1
Morales, C., Montalva, J., Arbetman, M., Aizen, M. A., Smith-Ramírez, C., Vieli, L., et al (2016). Bombus dahlbomii. The IUCN Red List of Threatened Species 2016: e.T21215142A100240441. Available online at: https://dx.doi.org/10.2305/IUCN.UK.2016-3.RLTS.T21215142A100240441.en (accessed July 27, 2021).
Morales, C. L., Arbetman, M. P., Cameron, S. A., and Aizen, M. A. (2013). Rapid ecological replacement of a native bumble bee by invasive species. Front. Ecol. Environ. 11, 529–534. doi: 10.1890/120321
Morandin, L. A., and Kremen, C. (2013). Hedgerow restoration promotes pollinator populations and exports native bees to adjacent fields. Ecol. Appl. (2013) 23, 829–839. doi: 10.1890/12-1051.1
Ne'eman, G., Jürgens, A., Newstrom-Lloyd, L., Potts, S. G., and Dafni, A. (2010). A framework for comparing pollinator performance: effectiveness and efficiency. Biol. Rev. 85, 435–451. doi: 10.1111/j.1469-185X.2009.00108.x
Nicholson, C. C., Koh, I., Richardson, L. L., Beauchemin, A., and Ricketts, T. H. (2017). Farm and landscape factors interact to affect the supply of pollination services. Agric. Ecosyst. Environ. 250, 113–122. doi: 10.1016/j.agee.2017.08.030
Nicholson, C. C., and Ricketts, T. H. (2019). Wild pollinators improve production, uniformity, and timing of blueberry crops. Agric. Ecosyst. Environ. 272, 29–37. doi: 10.1016/j.agee.2018.10.018
Potts, S. G., Imperatriz-Fonseca, V., Ngo, H. T., Aizen, M. A., Biesmeijer, J. C., Breeze, T. D., et al. (2016). Safeguarding pollinators and their values to human well-being. Nature. 540, 220–229. doi: 10.1038/nature20588
Pritchard, D. J., and Vallejo-Marín, M. (2020). Buzz pollination. Curr. Biol. 30, R858–R860. doi: 10.1016/j.cub.2020.05.087
Retamales, J. B., and Hancock, J. F. (2018). Blueberries (Vol. 27). Cambridge, MA, U.S.A: CABI. doi: 10.1079/9781780647265.0000
Rodríguez, S., Pérez-Giraldo, L. C., Vergara, P. M., Carvajal, M. A., and Alaniz, A. J. (2021). Native bees in Mediterranean semi-arid agroecosystems: Unravelling the effects of biophysical habitat, floral resource, and honeybees. Agric. Ecosyst. Environ. 307, 107188. doi: 10.1016/j.agee.2020.107188
Rogers, S. R., Tarpy, D. R., and Burrack, H. J. (2013). Multiple criteria for evaluating pollinator performance in highbush blueberry (Ericales: Ericaceae) agroecosystems. Environ. Entomol. 42, 1201–1209. doi: 10.1603/EN12303
Smith-Ramírez, C., Vieli, L., Barahona-Segovia, R. M., Montalva, J., Cianferoni, F., Ruz, L., et al. (2018). Las razones de por qué Chile debe detener la importación del abejorro comercial Bombus terrestris (Linnaeus) y comenzar a controlarlo. Gayan. 82, 118–127. doi: 10.4067/S0717-65382018000200118
Solís-Montero, L., and Vallejo-Marín, M. (2017). Does the morphological fit between flowers and pollinators affect pollen deposition? An experimental test in a buzz-pollinated species with anther dimorphism. Ecol. Evol. 7, 2706–2715. doi: 10.1002/ece3.2897
Stubbs, C. S., and Drummond, F. A. (1996). Blueberry and cranberry (Vaccinium spp.) pollination: a comparison of managed and native bee foraging behavior. Int. Symposium Pollinat. 437, 341–344. doi: 10.17660/ActaHortic.1997.437.42
Sutter, L., Albrecht, M., and Jeanneret, P. (2018). Landscape greening and local creation of wildflower strips and hedgerows promote multiple ecosystem services. J. Appl. Ecol. 55, 612–620. doi: 10.1111/1365-2664.12977
Torchio, P. F., and Burwell, B. (1987). Notes on the biology of Cadeguala occidentalis (Hymenoptera: Colletidae), and a review of colletid pupae. Ann. Entomol. Soc. Am. 80, 781–789. doi: 10.1093/aesa/80.6.781
Vallejo-Marín, M. (2019). Buzz pollination: studying bee vibrations on flowers. New Phytol. 224, 1068–1074. doi: 10.1111/nph.15666
Vallejo-Marín, M., Da Silva, E. M., Sargent, R. D., and Barrett, S. C. (2010). Trait correlates and functional significance of heteranthery in flowering plants. New Phytol. 188, 418–425. doi: 10.1111/j.1469-8137.2010.03430.x
Vallejo-Marín, M., and Vallejo, G. C. (2021). Comparison of defence buzzes in hoverflies and buzz-pollinating bees. J. Zool. 313, 237–249. doi: 10.1111/jzo.12857
Velasco, J. (2021). Arándanos en Chile, una temporada compleja y diferente. Redagrícola, Perú S.A.C. 74, 56–57.
Velthuis, H. H., and Van Doorn, A. (2006). A century of advances in bumblebee domestication and the economic and environmental aspects of its commercialization for pollination. Apidologie. 37, 421–451. doi: 10.1051/apido:2006019
Winter, K., Adams, L., Thorp, R., Inouye, D., Day, L., Ascher, J., et al. (2006). Importation of Non-Native Bumble Bees into North America: Potential Consequences of Using Bombus terrestris and other Non-Native Bumble Bees for Greenhouse Crop Pollination in Canada, Mexico, and the United States.
Keywords: buzz pollination, buzz pollinated crops, Chile, ecosystem services, non-apis bees, Vaccinium corymbosum
Citation: Cortés-Rivas B, Monzón VH, Rego JO and Mesquita-Neto JN (2023) Pollination by native bees achieves high fruit quantity and quality of highbush blueberry: a sustainable alternative to managed pollinators. Front. Sustain. Food Syst. 7:1142623. doi: 10.3389/fsufs.2023.1142623
Received: 11 January 2023; Accepted: 17 April 2023;
Published: 17 May 2023.
Edited by:
Fang Ouyang, Chinese Academy of Sciences (CAS), ChinaReviewed by:
Hongying Cui, Shandong Academy of Agricultural Sciences, ChinaCopyright © 2023 Cortés-Rivas, Monzón, Rego and Mesquita-Neto. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: José Neiva Mesquita-Neto, am1lc3F1aXRhQHVjbS5jbA==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.