 Annita Seok Kian Yong
Annita Seok Kian Yong Nur Suhailah Syed Mubarak1
Nur Suhailah Syed Mubarak1 Li-Chao Zhuo
Li-Chao Zhuo Yu-Hung Lin
Yu-Hung Lin Rossita Shapawi
Rossita Shapawi- 1Borneo Marine Research Institute, Universiti Malaysia Sabah, Kota Kinabalu, Malaysia
- 2Department of Aquaculture, National Pingtung University of Science and Technology, Neipu, Taiwan
Vegetable oil is progressively replacing fish oil (FO) in aquafeed production. However, the effect of oxidized vegetable oil is less reported. This study was done to investigate the effects of oxidized refined palm oil (RPO) on the growth performance, feed utilization, proximate and fatty acid profiles, apparent digestibility coefficients and oxidative stress of hybrid grouper juvenile (Epinephelus fuscoguttatus × E. lanceolatus). RPO was force-oxidized and introduced into diets at three oxidative levels to replace 50% of FO. Diets with FO and fresh RPO were served as controls. The peroxide value of the oxidized diets was measured as 14.73, 24.50, and 36.10 meq/kg for diets OxL, OxM, and OxH, respectively. These diets were fed to triplicate groups of hybrid grouper (initial body weight 9.91 ± 0.01 g) for 8 weeks. There was no significant difference in terms of growth performance, survival, feed utilization, and whole-body protein proximate composition in all treatments (p > 0.05). However, different dietary treatments affected the body fatty acids profile and fish-fed OxM diet achieved the highest body weight gain without significant difference (p > 0.05). In terms of hepaticsomatic index (HSI), fish-fed OxH showed a significantly lower value compared to other treatments. Similarly, the lowest apparent digestibility coefficients (ADC) of nutrients were observed in fish fed OxH diet. The oxidized diets caused significant lower blood cholesterol and high-density lipoprotein cholesterol (HDL) (p < 0.05). The oxidative stress in the liver as indicated by the significantly higher thiobarbituric acid-reactive substance (TBARS) value in fish fed all the oxidized diets while no significant changes on the muscle TBARS values. In conclusion, the oxidized RPO diets significantly affected the liver, the nutrient digestibility, and the blood total cholesterol and HDL. Furthermore, hybrid grouper can tolerate up to 20 meq/kg of oxidized RPO in the diet without compromising the growth.
Introduction
In aquafeed production, conventionally fish oil (FO) is used as the source of the main lipid [National Research Council (NRC), 2011]. However, over the last few decades, the limited production and the increased prices of FO had surged the effort to find alternative lipid sources to replace or reduce the use of FO in aquafeeds. There has been an increasing trend to use vegetable oil or edible oil extracted from plants such as soybean oil, corn oil, canola oil, and palm oil in aquafeed. Studies had shown that vegetable oils can replace FO partially or even up to 100% replacement (Peng et al., 2008; Mozanzadeh et al., 2016; Li et al., 2019; Gudid et al., 2020). The use of the vegetable oils is evident in maintaining a good growth performance, feed utilization, and feed digestibility, although in general the fatty acids profile of the fish is affected by the level and type of vegetable oils (Lin and Shiau, 2007; Peng et al., 2008; Shapawi et al., 2008; Ng et al., 2013; Mozanzadeh et al., 2016).
Palm oil is among the cheapest and largest production vegetable oil in the world. In recent years, palm oil products had been successfully used as a substitution for FO in diets for several marine fish species such as Japanese sea bass Lateolabrax japonicas (Gao et al., 2012); barramundi, Lates calcarifer (Wan Ahmad et al., 2013); humpback grouper, Cromileptis altivelis (Shapawi et al., 2008) and hybrid grouper Epinephelus fuscoguttatus × E. lanceolatus (Yong et al., 2019b; Gudid et al., 2020). Hybrid grouper is a cross breeding between brown-marbled grouper (E. fuscoguttatus, ♀) × giant grouper (E. lanceolatus, ♂). The hybrid grouper is an economically important aquaculture fish in the Southeast Asian region, Taiwan and China. The fish can accept a dietary lipid in the range of 12–16% for optimum growth (Yong et al., 2019a). Studies also showed that palm oil products can be used to replace 50–100% of FO without compromising the growth of hybrid grouper (Yong et al., 2019b; Gudid et al., 2020).
Lipid in fish feed is prone to oxidation when the feed is stored under aerobic and high temperature conditions. Dietary lipids are known to be susceptible to lipid peroxidation, which leads to the development of undesirable influences on the fish. Lower growth and survival, changes in whole-body composition, and liver damage were reported in several fish species when fed a diet with oxidized FO (Peng et al., 2008; Zhong et al., 2008; Chen et al., 2012; Gao et al., 2012; Long et al., 2022a). This is evidenced by the physiological changes in tested animals such as decreased antioxidant enzymes, changes in the transcriptome profiles, and increased byproducts of lipid peroxidation (Gao et al., 2012; Long et al., 2021, 2022a,b,c). With the increasing usage and different fatty acids profile of vegetable oils, there is a concern about the effect of oxidized vegetable oils on the performance of the fish (Liang et al., 2019). The effect of oxidized FO in aquaculture fish is well-reported, however, there is less information on the oxidized vegetable oils in aquaculture. The severe effects of oxidized vegetable oils on the animals can be different from FO due to the different fatty acid profiles (Liang et al., 2019). Therefore, the present study investigated the effects of oxidized palm oil diets on the growth, survival, feed efficiency, whole-body proximate composition, liver, and muscle fatty acids profile, apparent digestibility coefficient of nutrients, and blood serum as well as liver and muscle oxidative status of hybrid grouper juvenile.
Materials and Methods
Experimental Diets and Preparation
Five isoproteic (50%) and isolipidic (12%) experimental diets were formulated. Experimental diets were prepared by substituting 50% of FO with three levels of oxidized refined palm oil (RPO). Diets with 100% FO and 50% fresh RPO were served as control diets (Table 1). The RPO was force-oxidized by heating at 100°C under vigorous aeration (Mourente et al., 2002) and the three levels of oxidized RPO were achieved at every 48 h intervals. The degrees of oxidation was then determined according to the method of AOCS (1991). The different levels of oxidized oils were added to the test diets to replace 50% of the FO.
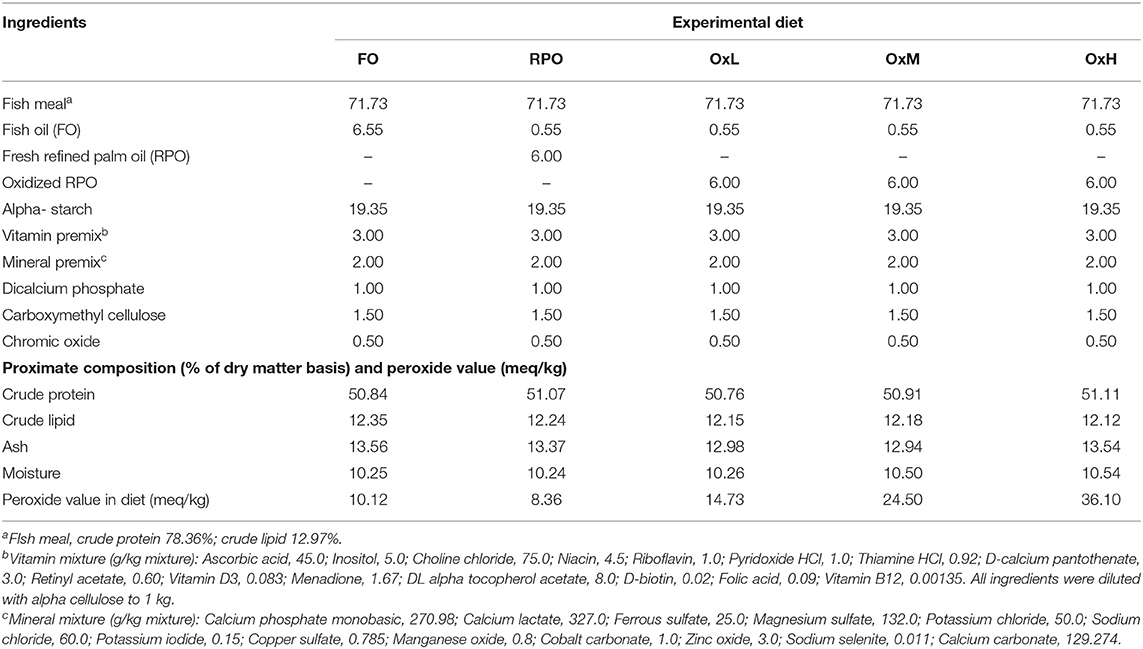
Table 1. Experimental diet formulation, proximate composition and peroxide value.
The experimental diets were prepared by mixing all the ingredients until a homogenous mixture, then 40% water was added to form moist dough before passing the dough through a kitchen meat mincer and then dried in the oven at 40°C until dry. All experimental diets were stored in air-tight plastic containers at 4°C until used. All diets were subjected to proximate analysis, peroxide value determination, and fatty acid analysis before use in the feeding trial. All diets contained ~50% crude protein and 12% crude lipid corresponded to the formulated protein and lipid content (Table 1). The peroxide value of the experimental diets FO, RPO, OxL, OxM, and OxH was estimated at 10.12, 8.36, 14.73, 24.50, and 36.10 meq/kg, respectively.
Feeding Trial and Sampling
The feeding trial was conducted in the hatchery of Borneo Marine Research Institute, Universiti Malaysia Sabah, Malaysia. The hybrid grouper juveniles were procured from a local hatchery and acclimatized for 1 week in two 800 L tanks upon arrival. The fish were then acclimatized to experimental conditions for another week while feeding with a commercial diet (Leong Hup Feedmill, 44 % crude protein, 8 % crude lipid) before the feeding trial.
The feeding trial was conducted in a triplicate tank for each dietary treatment. In each tank, 20 hybrid grouper juveniles with an average body weight (BW) 9.87 ± 0.3 g and total length (TL) 8.32 ± 0.05 cm were randomly distributed to each tank with a 150 L volume. The experimental system was a flow-through sea water system where the water flow rate to each tank was ~1 L/min. During the feeding trial, the fish were hand-fed to apparent satiation daily at 0800 and 1,500 h for 8 weeks. The feed intake and uneaten feed if any were recorded daily for estimation of feed consumption. The water salinity, temperature, pH and dissolved oxygen were recorded as 30–32 ppt, 29–31°C, 6.0–7.0, and 5.8–6.8 mg/L, respectively.
During the feeding trial, fish feces were collected to determine the apparent digestibility coefficient. Inert marker chromic oxide was added to the test diets for the apparent digestibility coefficient estimation. After 2 h of feeding, the bottom of the tank was cleaned by siphoning out the uneaten feed. Then fresh intact strands of feces were collected, rinsed with distilled water, and dried using filter paper before keeping it in a zip-lock bag and stored at −20°C until further analysis.
For growth monitoring, the BW of the fish was measured in bulk once every 2 weeks. Prior to each measurement, the fish were starved for 24 h. A commercial anesthetic (Transmore, Nika Trading) was used at ~200 ppm to anesthetize the fish (Noor et al., 2019) before taking the measurements to minimize the handling stress.
At the end of the feeding trial, the BW and TL of the fish were measured individually to estimate the growth performance. The final number of fish was recorded to calculate survival. Fish from each treatment was sampled for whole body proximate analysis (n = 3) and body indices estimation (n = 3). The muscle and liver from another fish were sacrificed for fatty acids (n = 3) and thiobarbituric acid-reactive substance (TBARS) analysis (n = 3). Blood samples were collected from the caudal vein using syringe (23 G) and quickly placed into vacutainer tubes for analysis.
Analysis Methods
The peroxide value analysis of the oils and experimental diets were conducted using the acetic acid-iso-octane method, AOCS Cd 8b-90 (AOCS, 1991). The proximate analysis included crude protein, ash, and moisture were analyzed following the method from AOAC (1999). While the crude lipid content was extracted using chloroform and methanol method (Folch et al., 1957). Transmethylation of fatty acids to fatty acids methyl esters was performed using sodium hydroxide (NaOH-methanol) and hydrogen chloride in methanol (HCl-methanol, Sigma) (Yoshikara and Satoh, 1989). The fatty acids methyl esters were analyzed using gas chromatography (Shimadzu GC-2010, Shimadzu Corporation, Kyoto, Japan) and separated using a capillary column (60 m × 0.25 mm ID; BPX70 column, SGE, Australia). The temperature program ranged from 80°C to 230°C (4°C min−1 from 80°C to 180°C and 2°C min−1 from 181°C to 230°C (Mohd Faudzi et al., 2018). The peaks obtained were identified by comparing the retention times of each peak with known standards Supelco™37 Component fatty acids methyl esters mix (Supelco Inc. Bellefonte, USA).
For the blood biochemical analysis, serum was obtained after centrifugation for 10 min at 30,000 rpm. The biochemical analysis was performed using commercial Roche reagent kits to determine cholesterol (cat. 03039773), triglycerides (cat. 20767107), and total protein (cat. 03183734) (Gudid et al., 2020). The TBRAS analysis was performed following the methods described by Uchiyama and Mihara (1978). For apparent digestibility coefficients (ADC), the feed and feces were digested with nitric and perchloric acids following the method of Furukawa and Tsukahara (1966). The apparent digestibility was calculated using the following formula:
ADC of nutrient = 100 × [1 – (% feces nutrient/% dietary nutrient) × (% dietary chromic oxide/feces chromic oxide)].
Statistical Analysis
The percentage data obtained in the study were arcsine-transformed before the statistical analysis. All data were analyzed using the software Statistical Package for the Social Science (SPSS 20.0). Normality of the data was analyzed using Shapiro–Wilk's test and homogeneity of variance was analyzed using Levene statistical test. One-way analysis of variance (ANOVA) and Tukey's multiple comparison tests were used to determine the significant differences among the means of the treatments at p < 0.05. Each of the data was the means of three replicates. For data on growth, feed efficiency, body composition, apparent digestibility co-efficient, blood biochemical, and TBRAS analysis, a polynomial contrast analysis was conducted. A second-order polynomial analysis of the percentage of body weight gain was conducted to determine the dietary oxidized oil that can be tolerated by hybrid grouper.
Results
Fatty Acid Composition of Experimental Diet
The fatty acids analysis showed that diets added with RPO contained higher levels of saturated and significantly lower levels of polyunsaturated fatty acids (PUFA) compared to the FO diet (Table 2). The addition of RPO also decreased the total n-3 PUFA and increased the total n-6 PUFA, due to the high percentage of fatty acid C18:2n6. Among the RPO diets, the addition of a higher level of oxidized RPO led to the increase of total monounsaturated fatty acids (MUFA) and decreased the total PUFA.
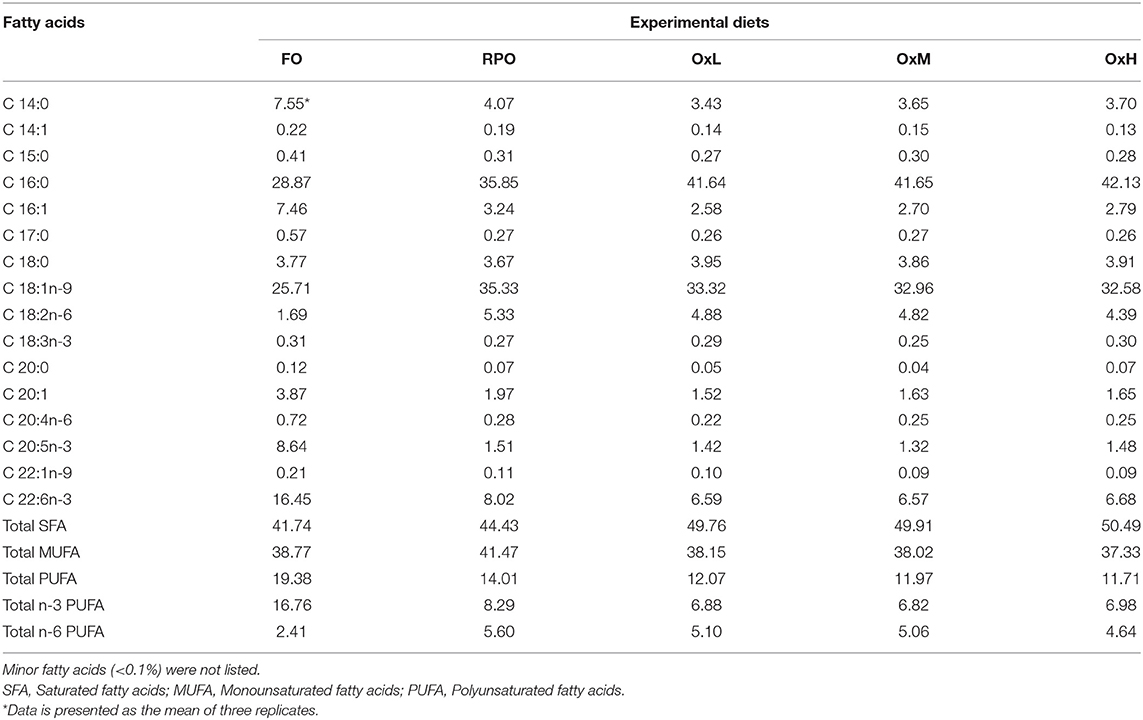
Table 2. Major fatty acid compositions (% of fatty acids/total fatty acids) of the experimental diets.
Growth Performance, Survival, and Feed Utilization
The effects of oxidized RPO on growth, survival, and feed utilization on the hybrid grouper juveniles are shown in Table 3. There was no significant difference in the growth performance, survival and feed utilization among the treatments (p > 0.05) and the polynomial contrast analysis did not show any linear or quadratic effects of the oxidized oil on the all the parameters observed. However, hybrid grouper juveniles fed with the OxM diet showed the highest weight gain, specific growth rate, and higher protein efficiency ratio while fish fed with the OxH diet resulted in the lowest growth and highest feed conversion ratio without significant difference (p > 0.05). Yet, we attempted to estimate a threshold level of dietary peroxide values using a second-order polynomial regression analysis based on the percentage of body weight gain. The model estimated 20 meq/kg as a value that can be acceptable by the hybrid grouper juvenile (Figure 1). A further increase in dietary oxidized levels decreased the growth of the fish.
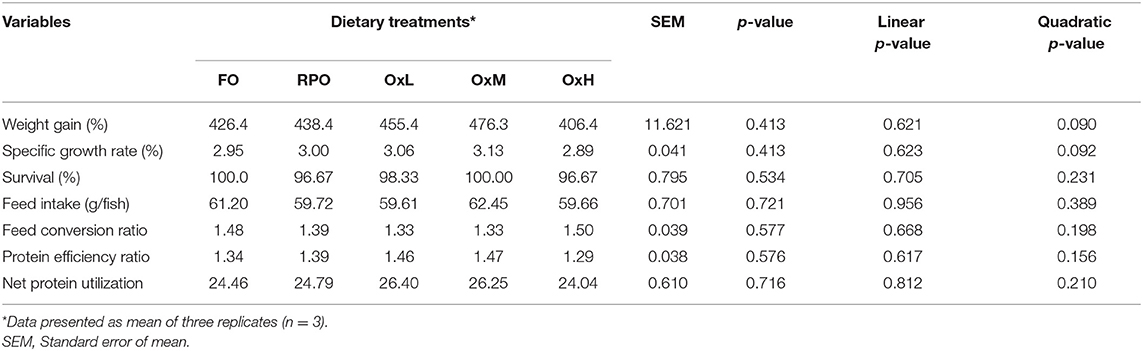
Table 3. Growth performances, survival and feed utilization of hybrid grouper juvenile fed control diets and treatment diets with oxidized RPO at the end of feeding trial.
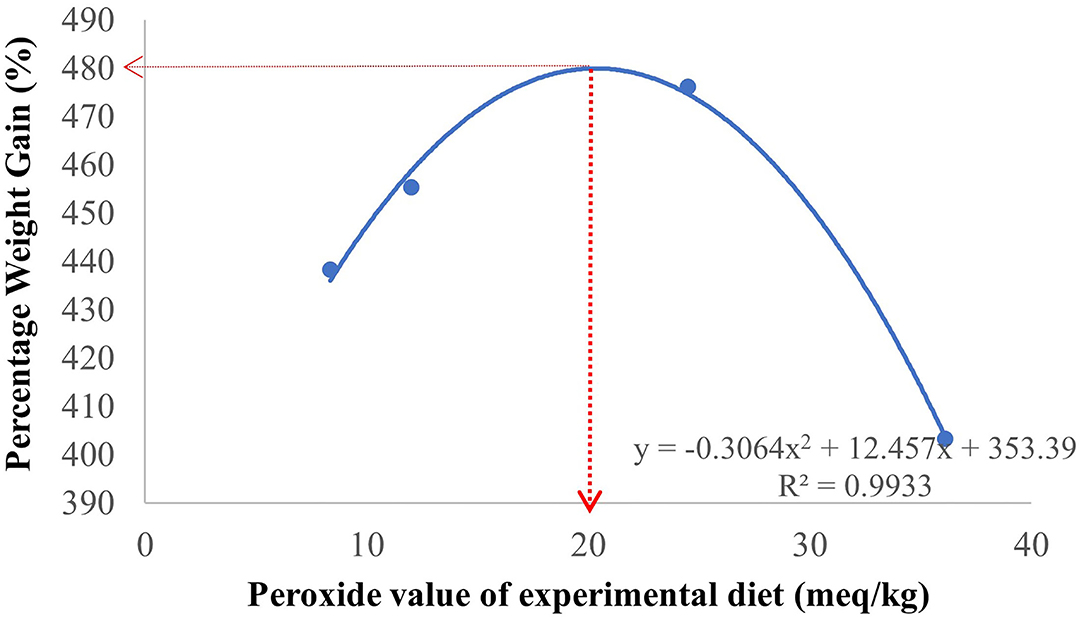
Figure 1. A second-order polynomial regression based on the percentage body weight gain (%) with increasing level of dietary peroxide values showed that 20 meq/kg is the estimated threshold peroxide value in the diet that can be tolerated by hybrid grouper juvenile.
Body Indices, Whole Body Proximate Composition, and Apparent Digestibility Coefficient (ADC)
The different degrees of oxidized RPO did not significantly affect the viscerosomatic index (VSI), intraperitoneal fat (IPF), and intesto-somatic index (ISI) of the fish (p > 0.05, Table 4). However, the increasing oxidized RPO levels inversely reduced the HSI and showed highly significant linear and quadratic effects on the HSI (p < 0.001).
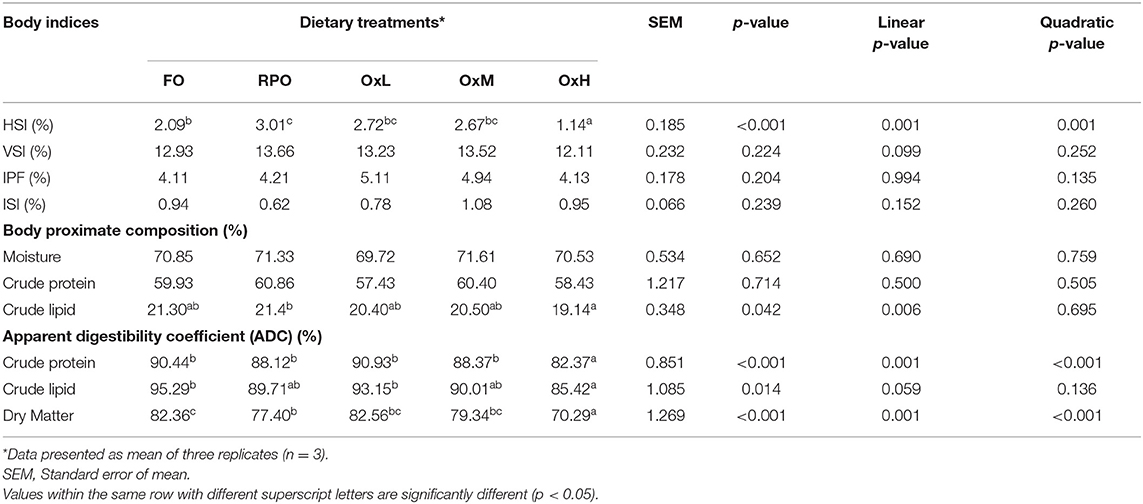
Table 4. The body indices, whole body proximate composition (dry weight basis), and apparent digestibility coefficient of hybrid grouper juvenile fed control and treatment diets with oxidized RPO at the end of feeding trial.
The body proximate analysis showed that there were no significant differences in the whole-body protein and moisture contents of the fish among the different treatments (p > 0.05). However, the increased level of oxidized RPO decreased the body lipid and significantly lower body lipid content was shown in fish fed OxH (p = 0.042) compared to other oxidized level diets. The polynomial contrast analysis showed a significant inverse linear effect of the oxidized RPO on the body lipid (p = 0.006).
In terms of ADC of nutrients, fish-fed OxH showed significantly lower values of ADC protein, lipid, and dry matter levels compared to other treatments (p < 0.05). The increasing oxidized RPO levels from OxL to OxH gradually decreased the ADC of protein (p < 0.001). Similarly, the increased levels of oxidized RPO in the diets decreased the ADC of lipid (p = 0.014) and the dry matter (p < 0.001) of the fish body. The oxidized RPO also showed significant linear and quadratic effects on the ADC of crude protein and dry matter (p < 0.001).
Fatty Acids Profile
The major fatty acids profile of the fish liver (Table 5) and muscle (Table 6) reflected the fatty acids profile of the respective experimental diets.
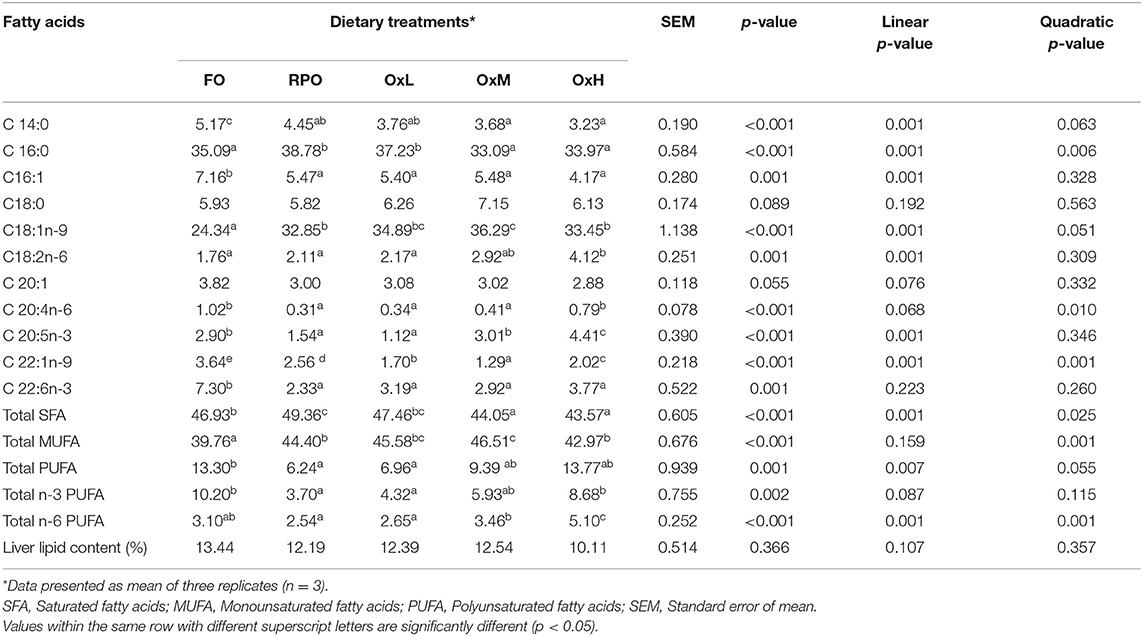
Table 5. Major fatty acid compositions (% fatty acids/ total fatty acids) of liver tissue of hybrid grouper juvenile fed control diets and treatment diets with oxidized RPO at the end of feeding trial.
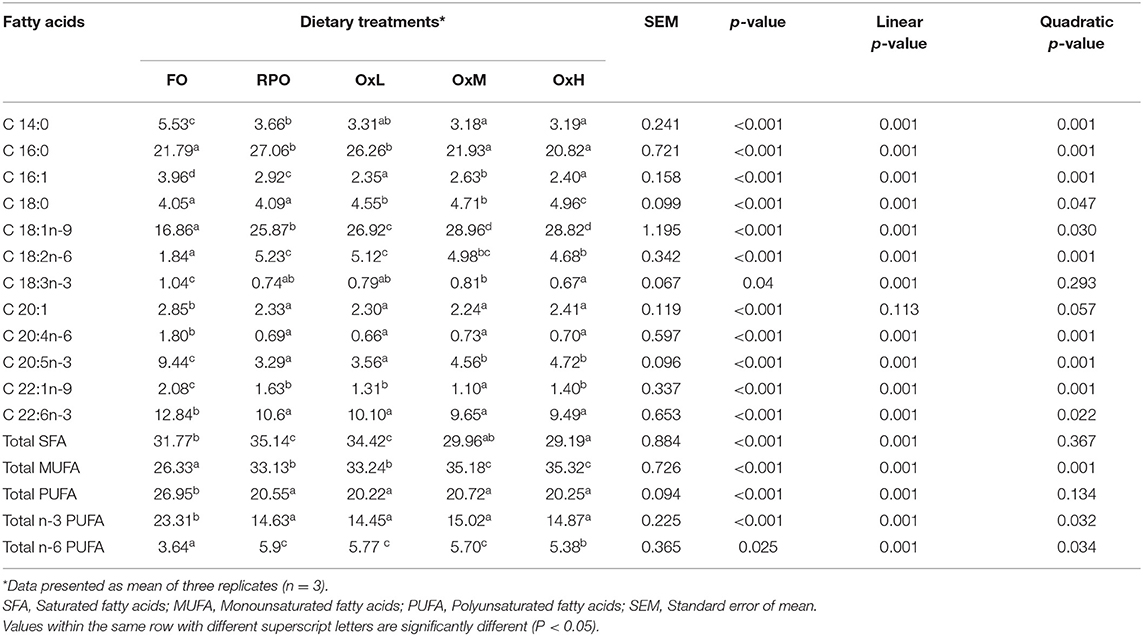
Table 6. Major fatty acid compositions (% fatty acids/ total fatty acids) of muscle tissue of hybrid grouper juvenile fed control diets and treatment diets with oxidized RPO at the end of the feeding trial.
The increased level of oxidized RPO in the diets resulted in a lower level of C16:0 fatty acid, total saturated fatty acids (SFA), and total MUFA in the liver (p < 0.05) among the RPO diets. While the total PUFA, especially the n-3 PUFA in the liver, increased with the oxidized RPO levels compared to those fed a fresh RPO diet (p < 0.05). The oxidized RPO diets linearly affected most of the fatty acids in the liver (p < 0.05). On the other hand, the fish fed oxidized diets showed a decreasing trend in the liver lipid content with the lowest liver lipid observed in fish fed OxH diet without significant difference among the treatments (p > 0.05).
Almost similar results can be observed in the muscle of the fish fed the experimental diets. However, in the muscle, the C18 fatty acid and total MUFA increased with the increasing level of oxidized RPO in diets. The SFA in the muscle decreased significantly with the increase of the oxidized RPO (p < 0.05). The total PUFA and total n-3 PUFA in the muscle of fish fed oxidized diets were maintained at an almost similar level with fish RPO, while the total n-6 PUFA showed decreasing level with the increased level of oxidized diets. The polynomial contrast analysis showed highly significant linear and quadratic effects on most of the fatty acids in the muscle (p < 0.05).
Blood Biochemical Constituents and Thiobarbituric Acid Reactive Substances (TBARS)
The blood biochemical constituents and TBARS in the liver and muscle of hybrid grouper juvenile at the end of the feeding trial are presented in Table 7. The total cholesterol and high-density lipoprotein (HDL) cholesterol were significantly decreased with the increasing level of dietary oxidized RPO levels in the diets (p < 0.05). The oxidized levels also showed significant linear and quadratic effects on the total cholesterol and HDL in the hybrid grouper (p < 0.05). However, no significant differences were observed in the triglycerides and total protein among the oxidized diets with the control diets except that fish-fed OxH showed higher triglycerides and lower protein (p > 0.05).
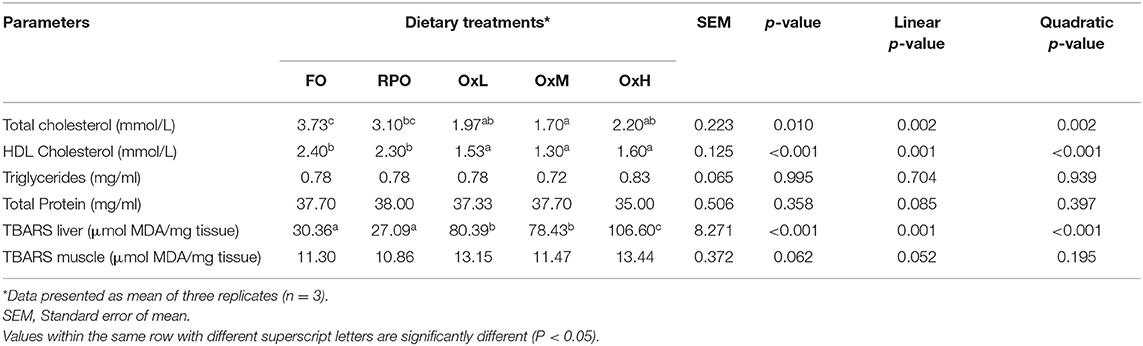
Table 7. Blood biochemical and TBARS in liver and muscle of hybrid grouper juvenile fed control diets and treatment diets with oxidized RPO at the end of the feeding trial.
The TBARS in the liver increased correspondingly to the increasing level of the oxidized degree of the diets and the TBARS levels were significantly higher in all oxidized diets compared to fish-fed FO and RPO diets (p < 0.05). Significant linear and quadratic effect of oxidized levels were also observed on the TBARS value in the liver (p < 0.001). However, no significant differences were observed in the TBARS value in the fish muscle among all treatments. Overall, fish-fed fresh RPO diets exhibited the lowest TBARS values among the treatments without significant difference (p > 0.05).
Discussion
In the present study, the oxidized RPO at different oxidation levels in the diets did not significantly affect the growth, feed utilization, survival, and overall body indices of the fish. However, the lowest growth was recorded in fish fed OxH diet with a peroxides value of 36.1 meq/kg. In a study testing oxidized FO in diets on a slightly bigger size of hybrid grouper (initial body weight 30 g) showed that feeding the fish with oxidized FO diets with peroxide value at more than 2.26 mmol/kg or 17.8 mg/kg malondialdehyde (MDA) significantly reduced the growth of the fish compared to the lower peroxide values diets (Long et al., 2021, 2022b). Besides, feeding the hybrid grouper with 9% of oxidized FO (FO oxidized level of 122 mmol/kg) decreased the mRNA expression of fraction growth factors in the muscle of the hybrid grouper (Yan et al., 2022). In channel catfish, Ictalurus punctatus, the inclusion of oxidized FO and soybean oil with a peroxide value of 350 meq/kg at 40 g/kg and 60 g/kg diet has significantly reduced the growth of the fish in an 8-week trial (Liang et al., 2019). The study also showed that at a similar inclusion level of 40 g/kg of oxidized oils, the oxidized FO caused more negative effects on the growth and oxidative stress than those fed oxidized soybean oil (Liang et al., 2019). Besides, a feeding trial on large yellow croaker, Larmichthys crocea reported that feeding the fish with oxidized FO diets with peroxides values from 28.3 to 104.2 meq/kg resulted in a significantly lower specific growth rate compared to the control group that fed fresh FO diet for 10 weeks (Wang et al., 2016). However, feeding largemouth bass Micropterus salmoides with oxidized FO diets with peroxides values from 132 to 555 meq/kg had led to a significant increase in body weight gain and specific growth rate compared with fish fed fresh FO at the end of the 12-week trial (p < 0.05) (Chen et al., 2012). These studies suggest that the growth responses of fish are markedly influenced by the type of oxidized oils (FO, vegetable oils) and the level of oxidized oil that can be tolerated by the fish is species specific. Based on the second-order polynomial regression analysis on the percentage of body weight gain and the dietary oxidized levels in the diets in the present study, 20 meq/kg is estimated to be the peroxide value of an oxidized diet with RPO that can be tolerated by hybrid grouper without comprising the growth. A further increase in oxidized RPO levels led to a reduction in growth performances.
The experimental diets were well-accepted by the hybrid grouper as the feed intake and other feed efficiency parameters were not significantly affected by the inclusion of fresh RPO and oxidized RPO. Long et al. (2021) reported that feed intake is also not affected by the different levels of oxidized FO diets (1.10–3.86 mg/kg MDA) in hybrid grouper, however, the FCR was significantly higher in fish fed the oxidized FO diets. A study on Japanese sea bass, L. japonicas also showed that the feed intake was not significantly affected by the oxidized FO diets (peroxide values 25.6–26.6 meq/kg) compared to fish fed FO and FO in combination with palm oil diets (peroxide value in diets 5.6–10.2 meq/kg) (Gao et al., 2012). However, the oxidized FO lowered the growth performance of the fish compared to fish-fed FO in combination with palm oil diets (Gao et al., 2012). While a study on largemouth bass (M. salmoides) showed that the feed intake was significantly higher in fish fed diet with a peroxide value 155 meq/kg and thus enhanced the growth of the fish compared to the control group that fed FO and other dietary treatments with higher peroxide values (Yin et al., 2019). The study also showed that the feed intake was higher in fish fed the all the oxidized diets compared to the fish fed FO (Yin et al., 2019). On the contrary, Atlantic salmon (Salmo salar) was shown to be able to discriminate against a diet with a higher oxidized diet with a TBRAS level 76 nmol/g compared to a diet with a TBARS level 34 nmol/g (Hamre et al., 2001).
The ADC of nutrients was significantly influenced by the oxidized diets. A decreasing trend of the ADC of protein, lipid and dry matter was observed with the increasing level of the oxidized RPO in the diets. A significant inverse linear effect of the oxidized RPO diets was noted on the ADC of protein and dry matter. The significantly lower ADC of nutrients had induced a lower growth performance in hybrid grouper fed the OxH diet. This finding is supported by a previous study on hybrid grouper that showed oxidized FO diets significantly induced oxidative stress in the intestine of hybrid grouper. The fish intestine is an important organ for digestion and absorption of nutrients (Le et al., 2019). The increasing level of oxidized FO in the diets (8.27–24.4 mg/kg MDA in diet) lowered the antioxidant enzyme activities while increasing the MDA level in the intestine (Long et al., 2022c). The oxidative stress was also clearly observed to disrupt the intestinal tissue structures (Long et al., 2022c). Thus, the oxidative stress and structure alteration observed in the intestine of hybrid grouper due to the inclusion of the oxidized FO (Long et al., 2022c) can lead to the reduction of digestion and absorption of nutrients in fish fed oxidized RPO diets and reduce the growth of hybrid grouper fed OxH in the present study.
The whole-body protein level of the hybrid grouper juvenile in the present study was not influenced by the experimental diets (p > 0.05). This is similar to the findings reported by Long et al. (2022b) and Yan et al. (2022) on hybrid grouper fed oxidized FO diets; the protein level in the muscle was not affected by the different levels of oxidized FO, and the feeding duration. A study of Chinese long snout catfish, L. longirostris also demonstrated that dietary oxidized FO diets did not affect the carcass composition of the fish (Dong et al., 2011). However, the whole-body lipid level was significantly lower in the hybrid grouper fed OxH diet in the present study (p < 0.05), and a significant decreasing linear effect of the body lipid was noted with the increasing level of oxidized RPO in the diets. This finding is also similar to the study by Long et al. (2022b) reported that the muscle lipid content in the hybrid grouper decreased with the increasing level of oxidized FO in the diets. Besides, the fatty acid profiles of experimental diets are well-reported to directly affect the fatty acid profiles of the fish (Li et al., 2019; Yong et al., 2019b; Yan et al., 2022). In the present study, the type of oils and the oxidized RPO in the diets significantly affected the fatty acids profile of the liver and muscle of hybrid grouper. This finding is similar to the liver and muscle fatty acid profiles of Japanese seabass L. japonicas fed different levels of palm oil and oxidized FO (Gao et al., 2012). In the present study, the RPO diets produced particularly higher levels of total MUFA and lower levels of total PUFA in both liver and muscle compared to fish fed control FM diet. On the other hand, the oxidized RPO diet decreased the total SFA in the liver and muscle compared to the RPO diet. Besides, the total PUFA level in the liver was higher in the higher oxidized diets compared to the RPO diet, while the total PUFA in the muscle was not significantly affected by the oxidized RPO diets. A slightly different fatty acid profile was reported in hybrid grouper fed oxidized FO, where higher total SFA and MUFA, and lower PUFA was reported in the muscle compared to fish fed a control diet (Yan et al., 2022). These findings showed that the alteration of fatty acid profiles in fish can be due to the different types and levels of oxidized oils and thus affect the quality and nutritional value of the fish for human consumption.
The liver is an important organ in metabolizing products coming from the digestive tract, and changes in the liver indicate the nutritional condition of the fish (Najdegerami et al., 2016). In the present study, fish fed OxH diet showed significantly lower HSI value or smaller liver (p < 0.05) and lower lipid content compared to dietary treatments OxL and OxM. A similar trend was reported in juvenile largemouth bass, M. salmoides where the HSI was lower in the fish fed diet with oxidized FO (peroxide value 277 meq/kg) compared to those fed fresh FO (Chen et al., 2012). On the contrary, oxidized FO diets did not lower the HSI of large yellow croaker, L. crocea (Wang et al., 2016). The study reported that the HSI of large yellow croakers increased with the increase of oxidized level (peroxides values 28.29 to 104.21 meq/kg) in the diets. While a study on yellow catfish Pelteobagrus fulvidraco showed that feeding the fish with oxidized FO diets (peroxides values 4.62–63.37 meq/kg) did not cause significant changes in the HSI (Zhang et al., 2021). These studies suggest that the changes in the HSI value or liver may be due to species specific in responses to the different level and type of oxidized oil.
Despite the differences of HSI changes in response to the oxidized diets in previous studies, oxidized oil is evidently causing liver oxidative stress in fish. In the present study, the oxidized RPO diets significantly increased the hepatic TBARS values in a dose-dependent manner with the peroxide value of the diets. TBARS are used as an indicator to evaluate tissue damage from dietary oxidative stress (Hamre et al., 2001). The higher value of TBARS indicates higher lipid peroxidation and oxidative stress. The TBARS of fish fed OxH diet was significantly higher than other treatments. This suggests that the hepatic lipid peroxidation of hybrid grouper was accelerated by the increasing level of oxidized RPO in diets. This hepatic oxidative stress is also obvious from the increasing levels of PUFA in a dose-dependent response to the dietary oxidized levels in the diets due to the antioxidant and anti-inflammatory properties of PUFA particularly n-3 PUFA to reduce oxidative stress (Oppedisano et al., 2020). In largemouth bass, M. salmoides the oxidized FO diets were reported to increase the liver catalase, superoxide dismutase, glutathione peroxidase, glutathione reductase, and glutathione-S-transferase activities (Chen et al., 2012). Besides, in large yellow croaker, L. crocea increasing levels of MDA content in the liver were observed in fish fed the oxidized FO diets (Wang et al., 2016). In yellow catfish, P. fulvidraco, oxidized FO diets caused oxidative stress in the liver, increased lipogenesis and reduced lipolysis (Zhang et al., 2021). These findings are also consistent with the previous study on hybrid grouper reported that oxidized FO significantly decreased the antioxidant enzymes such as superoxide dismutase, glutathione peroxidase and catalase, and vitamin E content in the liver and muscle with the increased level of oxidized FO in the diets (Long et al., 2021, 2022b). These studies also showed that oxidized FO diets led to higher liver lipid content and suggested that oxidized FO impaired liver lipid metabolism and morphological abnormalities (Long et al., 2021, 2022a). While the present findings showed that oxidized RPO diets did not significantly affect the lipid content in the liver and the TBARS values in the fish muscle. These varied results could also be due to the level and type of oxidized oils that were tested on the fish.
Blood biochemical parameters are an indicator of the nutritional status of fish (Tan et al., 2013). The total protein is an indicator of dietary protein status and is related to absorption in the animals (Wang et al., 2016). The present study, it showed that the total protein was not significantly affected by the oxidized diets. Similar to the total protein, triglycerides are not significantly influenced by the oxidized RPO or the oxidized level. However, the inclusion of oxidized RPO in the diets regardless of the oxidation level had markedly reduced the high-density lipoprotein cholesterol (HDL) and the total cholesterol content compared to the fish fed the control diets with FO and fresh RPO. This is in contrast with a previous study on hybrid grouper where an increase of oxidized FO did not alter the total cholesterol level but significantly increased the HDL in the serum of hybrid grouper (Long et al., 2021). The reduction of total cholesterol with the increased oxidized level of RPO oil could probably be due to the decrease of HDL in the fish and the presence of vitamin E in the palm oil (Ng et al., 2008). A study on Japanese flounder, Paralichtyhs olivaceus reported that the plasma total cholesterol increased when the fish were fed with an oxidized FO diet (Gao et al., 2014). However, lower total cholesterol was observed when the fish was fed with an oxidized FO diet supplemented with vitamin E and C (Gao et al., 2014). Replacement of FO with vegetable oil leads to a decrease in plasma cholesterol. It is attributed partially to the decreased content of cholesterol in vegetable oil compared to FO (Turchini et al., 2009). Kim et al. (2012) reported that the lower content of total cholesterol in fish fed with vegetable oil is likely related to the cholesterol lowering factor that is present in linseed and soybean oils. The various results showed that the effects of oxidized oil on the total cholesterol may be due to variation between species and probably due to the difference in heating treatment and type of lipid used (Ringseis and Eder, 2011).
Conclusion
The use of oxidized RPO in the diets caused oxidative stress in the hybrid grouper especially in fish fed the OxH diet demonstrated by a significantly lower value of ADC of nutrients, HSI, total cholesterol, and HDL and significantly high TBARS values in the liver. The present study also showed that the hybrid grouper can tolerate up to 20 meq/kg of RPO oxidized diet within a period of 8 weeks without compromising the growth.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics Statement
The animal study was reviewed and approved by Animal Ethics Committee, Centre for Research & Innovation, Universiti Malaysia Sabah.
Author Contributions
AY: conceptualization, interpretation of data, manuscript drafting, review, and editing. NS: investigation, collection, interpretation of data, and manuscript drafting. L-CZ: investigation, analysis, and interpretation of data. Y-HL: manuscript review and research methodology. RS: funding acquisition, project administration, and manuscript review. All authors contributed to the article and approved the submitted version.
Funding
The present study was supported by the Ministry of Higher Education Malaysia under Niche Research Grant Scheme (NRGS0004) in Universiti Malaysia Sabah.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
AOAC (1999). Official Methods of Analysis. Association of Official Analytical Chemists. Washington, DC: AOAC International.
AOCS (1991). Official Methods and Recommendation Practices of the American Oil Chemists Society. Champaign, IL: American Oil Chemists Society.
Chen, Y. J., Liu, Y. J., Yang, H. J., Yuan, Y., Liu, F. J., Tian, L. X., et al. (2012). Effect of dietary oxidized fish oil on growth performance, body composition, antioxidant defence mechanism and liver histology of juvenile largemouth bass, Micropetrus salmonides. Aquacult. Nutr. 18, 321–331. doi: 10.1111/j.1365-2095.2011.00900.x
Dong, X., Lei, W., Zhu, X., Han, D., Yang, Y., and Xie, S. (2011). Effects of dietary oxidized fish oil on growth performance and skin colour of Chinese longsnout catfish, Leiocassis longirostris. Aquacult. Nutr. 17, e861–8. doi: 10.1111/j.1365-2095.2011.00854.x
Folch, J., Lees, M., and Sloane, G. H. (1957). Simple method for isolation and purification of total lipids from animal tissues. J. Biol. Chem. 226, 497–507. doi: 10.1016/S0021-9258(18)64849-5
Furukawa, A., and Tsukahara, H. (1966). On the acid digestion method for the determination of chromic oxide as an index substance in the study of digestibility of fish feed. Bull. Jpn. Soc. Sci. Series 32, 502–506. doi: 10.2331/suisan.32.502
Gao, J., Koshio, S., Ishikawa, M., Yokoyama, S., and Mamauag, R. E. P. (2014). Interactive effects of vitamin C and E supplementation on growth performance, fatty acid composition and reduction of oxidative stress in juvenile Japanese flounder Paralichthys olivaceus fed dietary oxidized fish oil. Aquaculture 42, 84–90. doi: 10.1016/j.aquaculture.2013.11.031
Gao, J., Koshio, S., Ishikawa, M., Yokoyama, S., Ren, T., Komilus, C. F., et al. (2012). Effects of dietary palm oil supplements with oxidized and non-oxidized fish oil on growth performances and fatty acid compositions of juvenile Japanese sea bass, Lateolabrax japonicas. Aquaculture. 324–325, 97–103. doi: 10.1016/j.aquaculture.2011.10.031
Gudid, S. N., Lim, L.-S., Yong, A. S. K., Yanuhar, U., Mustafa, S., and Shapawi, R. (2020). Growth performance and organoleptic quality of hybrid grouper (Epinephelus fuscogutattus ♀ × Epinephelus lanceolatus ♂) fed palm-oil based diets at grow-out stage. Sains Malays. 49, 1567–1576. doi: 10.17576/jsm-2020-4907-09
Hamre, K., Kolås, K., Sandess, K., Julshamn, K., and Kiessling, A. (2001). Feed intake and absorption of lipid oxidation products in Atlantic salmon, Salmo salar fed diets coated with oxidized fish oil. Fish Physiol. Biochem. 25, 209–219. doi: 10.1023/A:1022257928437
Kim, D. K., Kim, K. D., Seo, J. Y., and Lee, S. M. (2012). Effects of dietary lipid source and level on growth performance, blood parameters and flesh quality of sub-adult oliveflounder, Paralichthys olivaceus. Asian Austral. J. Anim. Sci. 25, 869–879. doi: 10.5713/ajas.2011.11470
Le, H. T. M. D., Shao, X., Krogdahl, Å., Kortner, T. M., Lein, I., Kousoulaki, K., et al. (2019). Intestinal function of the stomachless fish, ballan wrasse (Labrus bergylta). Front. Mar. Sci. 6, 140. doi: 10.3389/fmars.2019.00140
Li, B., Wang, J., Huang, Y., Hao, T., Wang, S., Huang, B., et al. (2019). Effects of replacing fish oil with wheat germ oil on growth, fat deposition, serum biochemical indices and lipid metabolic enzyme of juvenile hybrid grouper (Epinephelus fuscoguttatus♀ × Epinephelus lanceolatus♂). Aquaculture 505, 54–62. doi: 10.1016/j.aquaculture.2019.02.037
Liang, G., Li, X., Yang, H., Huan, D., Xu, Z., Leng, X., et al. (2019). Dietary oxidized oils decreased growth, antioxidative capacity, and negatively affected skin color of channel catfish, Ictalurus punctatus. J. World Aquacult. Soc. 50, 692–706. doi: 10.1111/jwas.12556
Lin, Y. H., and Shiau, S. Y. (2007). Effects of dietary blend of fish oil with corn oil on growth and non-specific immune responses of grouper, Epinephelus malabaricus. Aquacult. Nutr. 13, 137–144. doi: 10.1111/j.1365-2095.2007.00458.x
Long, S., Dong, X., Liu, H., Yan, X., Tan, B., Zhang, S., et al. (2022a). Effect of dietary oxidized fish oil on liver function in hybrid grouper (♀ Epinephelus fuscoguttatus × ♂ Epinephelus lanceolatus). Aquacult. Rep. 22, 101000. doi: 10.1016/j.aqrep.2021.101000
Long, S., Dong, X., Liu, H., Yan, X., Tan, B., Zhang, S., et al. (2022a). Effect of dietary oxidized fish oil on liver function in hybrid grouper (♀ Epinephelus fuscoguttatus×♂ Epinephelus lanceolatus). Aquacult. Rep. 22, 101000. doi: 10.1016/j.aqrep.2021.101000
Long, S., Dong, X., Tan, B., Zhang, S., Chi, S., Yang, Q., et al. (2022b). The antioxidant ability, histology, proximate, amino acid and fatty acid compositions, and transcriptome analysis of muscle in juvenile hybrid grouper (♀ Epinephelus fuscoguttatus × ♂ Epinephelus lanceolatus) fed with oxidized fish oil. Aquaculture 547, 737510. doi: 10.1016/j.aquaculture.2021.737510
Long, S., Dong, X., Tan, B., Zhang, S., Xie, S., Yang, Q., et al. (2021). Growth performance, antioxidant ability, biochemical index in serum, liver histology and hepatic metabolomics analysis of juvenile hybrid grouper (♀ Epinephelus fuscoguttatus × ♂ Epinephelus lanceolatus) fed with oxidized fish oil. Aquaculture 545, 737261. doi: 10.1016/j.aquaculture.2021.737261
Long, S., Dong, X., Yan, X., Liu, H., Tan, B., Zhang, S., et al. (2022c). The effect of oxidized fish oil on antioxidant ability, histology and transcriptome in intestine of the juvenile hybrid grouper (Epinephelus fuscoguttatus♀ ×Epinephelus lanceolatus♂). Aquacult. Rep. 22, 100921. doi: 10.1016/j.aqrep.2021.100921
Mohd Faudzi, N., Yong, A. S. K., Shapawi, R., Shigeharu, S., Biswas, A., and Takii, K. (2018). Soy protein concentrate as an alternative in replacement of fish meal in the feeds of hybrid grouper, brown-marbled grouper (Epinephelus fuscoguttatus) x giant grouper (E. lanceolatus) juvenile. Aquacult. Res. 49, 431–441. doi: 10.1111/are.13474
Mourente, G., Diaz-Salvago, E., Bell, J. G., and Tocher, D. R. (2002). Increased activities of hepatic antioxidant defence enzymes in juvenile gilthead sea bream (Sparus aurata L.) fed dietary oxidized oil: attention by dietary vitamin E. Aquaculture 214, 343–361. doi: 10.1016/S0044-8486(02)00064-9
Mozanzadeh, M. T., Agh, N., Yavari, V., Marammazi, J. G., Mohammadian, T., and Gisbert, E. (2016). Partial or total replacement of dietary fish oil with alternative lipid sources in silvery-black porgy (Sparidentex hasta). Aquaculture 451, 232–240. doi: 10.1016/j.aquaculture.2015.09.022
Najdegerami, E. H., Bakhshi, F., and Lakani, F. B. (2016). Effects of biofloc on growth performance, digestive enzyme activities and liver histology of common carp, Cyprinus carpio L. fingerlings in zero-water exchange system. Fisher. Physiol. Biochem. 42, 457–465. doi: 10.1007/s10695-015-0151-9
National Research Council (NRC) (2011). Nutrient Requirements of Fish and Shrimp. Animal Nutrition Series. Washington, DC: The National Academic Press.
Ng, W. K., Chong, C. H., Wang, Y., and Romana, N. (2013). Effects of dietary fish and vegetable oils on the growth, tissue fatty acid composition, oxidative stability and vitamin E content of red hybrid tilapia and efficacy of using fish oil finishing diets. Aquaculture 372–375, 97–110. doi: 10.1016/j.aquaculture.2012.10.030
Ng, W.-K, Wang, Y., and Yuen, K.-H. (2008). Palm vitamin E for aquaculture feeds. J. Oil Palm Res. 1–7.
Noor, N. M., Cob, Z. C., Ghaffar, M. A., and Das, S. K. (2019). An evaluation of the effect of salinities on oxygen consumption and wellbeing in the hybrid grouper Epinephelus fuscoguttatus × E. lanceolatus. Turkish J. Fish. Aquat. Sci. 19, 1017–1023. doi: 10.4194/1303-2712-v19_12_04
Oppedisano, F., Macrì, R., Gliozzi, M., Musolino, V., Carresi, C., Maiuolo, J., et al. (2020). The anti-inflammatory and antioxidant properties of n-3 PUFAs: their role in cardiovascular protection. Biomedicines 8, 306. doi: 10.3390/biomedicines8090306
Peng, S., Chen, L., Qin, J. G., Hou, J., Yu, N., Long, Z., et al. (2008). Effects of replacement of dietary fish oil by soybean oil on growth performance and liver biochemical composition in juvenile black seabream, Acanthopagrus schlegeli. Aquaculture 276, 154–161. doi: 10.1016/j.aquaculture.2008.01.035
Ringseis, R., and Eder, K. (2011). Regulation of genes involved in lipid metabolism by dietary oxidized fat. Mol. Nutr. Food Res. 55, 109–121. doi: 10.1002/mnfr.201000424
Shapawi, R., Mustafa, S., and Ng, W. K. (2008). Effects of dietary fish oil replacement with vegetable oils on growth and tissue fatty acid composition of humpback grouper, Cromileptes altivelis (Valenciennes). Aquac. Res. 39, 315–323. doi: 10.1111/j.1365-2109.2007.01882.x
Tan, Q., Liu, Q., Chen, X., Wang, M., and Wu, Z. (2013). Growth performance, biochemical indices and hepatopancreatic function of grass carp, Ctenopharyngodon idellus, would be impaired by dietary rapeseed meal. Aquaculture 414, 199–195. doi: 10.1016/j.aquaculture.2013.07.036
Turchini, G. M., Torstensen, B. E., and Ng, W. K. (2009). Fish oil replacement in finfish nutrition. Rev. Aquacult. 1, 10–57. doi: 10.1111/j.1753-5131.2008.01001.x
Uchiyama, M., and Mihara, M. (1978). Determination of malonaldehyde precursor in tissues by thiobarbituric acid test. Anal. Biochem. 86, 271–278. doi: 10.1016/0003-2697(78)90342-1
Wan Ahmad, W. A., Stone, D. A. J., and Schuller, K. A. (2013). Dietary fish oil replacement with palm or poultry oil increases fillet oxidative stability and decreases liver glutathione peroxidise activity in barramundi, Lates calcarifer. Fish Physiol. Biochem. 39, 1631–1640. doi: 10.1007/s10695-013-9815-5
Wang, J., Xu, H., Zuo, R. T., Mai, K. S., Xu, W., and Ai, Q. H. (2016). Effects of oxidized dietary fish oil and high-dose vitamin E supplementation on growth performance, feed utilization and antioxidant defense enzyme activities of juvenile large yellow croaker, Larmichthys crocea. Br. J. Nutr. 115, 1531–1538. doi: 10.1017/S0007114516000398
Yan, X., Li, Z., Dong, X., Tan, B., Pan, S., Li, T., et al. (2022). Degradation of muscle quality in hybrid grouper (♀ Epinephelus fuscoguttatus × ♂ Epinephelus lanceolatus) due to oxidative damage caused by ingestion of oxidized fish oil. Front. Nutr. 9, 840535. doi: 10.3389/fnut.2022.840535
Yin, P., Xie, S., Huo, Y., Guo, T., Fang, H., Zhang, Y., et al. (2019). Effects of dietary oxidized fish oil on growth performance, antioxidant defense system, apoptosis and mitochondrial function of juvenile largemouth bass (Micropterus salmoides). Aquaculture 500, 347–358. doi: 10.1016/j.aquaculture.2018.09.009
Yong, A. S. K., Mohd Faudzi, N., Shapawi, R., and Shigeharu, S. (2019a). The optimum level of dietary protein and lipid for hybrid grouper, brown-marbled grouper (Epinephelus fuscoguttatus) x giant grouper (E. lanceolatus). J. Sust. Sci. Manag. 14, 1–15.
Yong, A. S. K., Syed Mubarak, N. S., and Shapawi, R. (2019b). The effects of partial replacement of fish oil with different vegetable oils on the growth, feed utilization and fatty acid profile of hybrid grouper juvenile (Epinephelus fuscoguttatus x Epinephelus lanceolatus). J. Oil Palm Res. 31, 110–121. doi: 10.21894/jopr.2019.0003
Zhang, D. G., Zhao, T., Hogstrand, C., Ye, H.-M., Xu, X.-J., and Luo, Z. (2021). Oxidized fish oils increased lipid deposition via oxidative stress-mediated mitochondrial dysfunction and the CREB1-Bcl2-Beclin1 pathway in the liver tissues and hepatocytes of yellow catfish. Food Chem. 360, 129814. doi: 10.1016/j.foodchem.2021.129814
Keywords: hybrid grouper, fish oil, oxidized palm oil, apparent digestibility coefficients, TBARS, liver
Citation: Yong ASK, Syed Mubarak NS, Zhuo L-C, Lin Y-H and Shapawi R (2022) Oxidized Palm Oil Diet Affects Fatty Acid Profiles, Apparent Digestibility Coefficients and Liver of Hybrid Grouper Juvenile (Epinephelus fuscoguttatus × Epinephelus lanceolatus). Front. Sustain. Food Syst. 6:837469. doi: 10.3389/fsufs.2022.837469
Received: 16 December 2021; Accepted: 16 May 2022;
Published: 20 June 2022.
Edited by:
Keitaro Kato, Kindai University, JapanReviewed by:
Janice Ragaza, Ateneo de Manila University, PhilippinesHüseyin Sevgili, Hüseyin Sevgili, Turkey
Copyright © 2022 Yong, Syed Mubarak, Zhuo, Lin and Shapawi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Annita Seok Kian Yong, YW5uaXRheUB1bXMuZWR1Lm15