
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
DATA REPORT article
Front. Sustain. Food Syst. , 25 October 2021
Sec. Crop Biology and Sustainability
Volume 5 - 2021 | https://doi.org/10.3389/fsufs.2021.735893
This article is part of the Research Topic Biostimulants as an Avenue of Abiotic Stress Tolerance Improvement in Crops View all 7 articles
 Ilara G. F. Budzinski1
Ilara G. F. Budzinski1 Paula O. Camargo1Raissa S. Rosa1Natália F. Calzado1
Paula O. Camargo1Raissa S. Rosa1Natália F. Calzado1 Suzana T. Ivamoto-Suzuki1,2
Suzana T. Ivamoto-Suzuki1,2 Douglas S. Domingues1*
Douglas S. Domingues1*Coffee (Coffea spp.) is one of the most important traded commodities in the international market [International Coffee Organization (ICO), 2018]. C. arabica, the only allotetraploid species in the genus, is the most planted one. Despite its economic importance, evaluation of physiological changes taking account molecular responses to biostimulants are still scarce. When plants recognize potential biotic/abiotic challenges, they often switch to a primed state of enhanced defense. This mechanism enables plants to respond robustly after exposure to stress (Aranega-Bou et al., 2014). The response of C. arabica plants to priming, in terms of transcriptional profiles, is a big gap in this area. Hexanoic acid (Hx) is a natural priming agent with proven efficiency in a wide range of host plants and pathogens (Llorens et al., 2016), including coffee pathogens. In this study we aimed to investigate the effect of Hx priming in C. arabica leaves transcriptome. We hypothesize if Hx application could modulate genes related to defense responses, being a potential eliciting agent in C. arabica. To test this effect, we applied Hx in roots of two Brazilian C. arabica cultivars with distinct breeding histories and contrasting resistance to the major disease in Arabica coffee, coffee leaf rust. While Catuai Vermelho is among the most used cultivars in Brazil, but it is susceptible to leaf rust, Obatã is a moderately resistant cultivar (Del Grossi et al., 2013). We performed transcriptome analysis of leaves. Reads were mapped to the C. arabica public genome and up to 94% of reads were mapped. Transcript expression level was quantified and differentially expressed genes (DEGs) were identified based on FPKM ratio and statistical analyses. A total of 57 and 63 DEGs were found in Catuai Vermelho and Obatã, respectively. Most DEGs correspond to upregulated genes in response to Hx, in both cultivars (86% Catuai Vermelho and 73% Obatã). Eight DEGs were found modulated in both cultivars, including ferredoxin-NADP reductase and phenylalanine ammonia-lyase. DEGs were functionally annotated through Blast2GO. Biological process and molecular function categorizations revealed that DEGs related to cellular, oxidation-reduction, organic substance and primary metabolic process, as well as transferase and ion binding activity might play a dominant role in the leaf response to priming. These data contribute to identify key genes differentially expressed in response to Hx as well as indicate pathways modulated by this eliciting agent.
• The species C. arabica is responsible for 60% of the world coffee production due to its fine flavor and aroma. Brazil is the largest producer and exporter of this commodity.
• Hexanoic acid (Hx) is a priming agent which leads plant to a physiological state that enables them to respond more rapidly and/or robustly to biotic or abiotic stress scenario.
• To date, there is no transcriptome analysis for C. arabica cultivars Catuai Vermlho and Obatã in response to the priming agent hexanoic acid (Hx) application.
• These data will contribute to identify key genes differentially expressed in response to Hx application and it can also indicates which pathways are modulated by Hx.
Five-month-old plants of C. arabica cv. Catuaí Vermelho IAC 144 and Obatã IAC 1669-20 (4–5 leaf pairs) were used in this study. Plants were selected based on size uniformity and were transferred to pots containing 3 L of aerated nutrient solution (ANS), adapted from Clark (1975) by de Carvalho et al. (2013). The hexanoic acid experiment was carried out as described in Silva et al. (2020), under controlled temperature (23 ± 2°C) and light/dark cycle (12/12 h, photosynthetically active photon flux density of ~400 μmol.m−2.s−1). The following treatments were assessed: (a) ANS (control); (b) ANS + hexanoic acid (Merck, final concentration 0.55 mM) for 48 h. Plants were grown in 3 to 6 plastic pots in which three pots received each treatment. The experiments were repeated 3 times to obtain biological replicates. The potted plants were grouped in “pools” (made of 9–18 plants), which were considered a biological replicate. Three biological replicates were used. The mature leaves of the middle third of the plants were collected within the 3rd hour of the light period and stored at −80°C to further analyses.
Total RNA was extracted from leaves pools using the RNeasy Plant kit (Qiagen, Hilden, North Rhine-Westphalia, Germany). Total RNA samples were purified using the RNeasy Minielute Cleanup kit (Qiagen, Hilden, North Rhine-Westphalia, Germany). The purity of RNA was determined using a NanoDrop ND-100 spectrophotometer (Thermo Scientific, San Jose, CA, USA) and concentrations were obtained using Qubit fluorimeter (Thermo Fisher Scientific, Wilmington, DE, USA). Poly(A) RNA sequencing library was prepared following Illumina's TruSeq-stranded-mRNA sample preparation protocol (Illumina Technologies, SanDiego, CA). Paired-ended sequencing (2 X 150 bp) was performed on Illumina's NovaSeq 6000 sequencing system at LC Sciences (Houston, TX, USA). Data were deposited into the European Nucleotide Archive (ENA), submission ERA6282544 and in Zenodo (doi: 10.5281/zenodo.5517785).
Adaptor contamination, low quality bases and undetermined bases were removed by using Cutadapt (Martin, 2011) and in house PERL scripts. Sequence quality was verified using FastQC (Andrews, 2010). HISAT2 (Kim et al., 2015) was used to map reads to the C. arabica genome (ftp://ftp.ncbi.nlm.nih.gov/genomes/all/GCF/003/713/225/GCF_003713225.1_Cara_1.0/.Mapped) reads were assembled using StringTie (Pertea et al., 2015). Table 1 shows the statistics of the transcriptome analysis. StringTie was also used to detect expression level for mRNAs by calculating FPKM. The differentially expressed genes (DEGs) were selected with log2 (fold change) >1 or log2 (fold change) < -1 and with statistical significance (p value < 0.05) by R package edgeR (Robinson et al., 2010). A second analysis was done on the differentially expressed mRNAs and only the ones with FPKM (ratio) ≥ 2 or FPKM (ratio) ≤ −2; coefficient of variation ≤ 30% and average FPKM ≥ 5. These genes are reported in Supplementary Table 1.
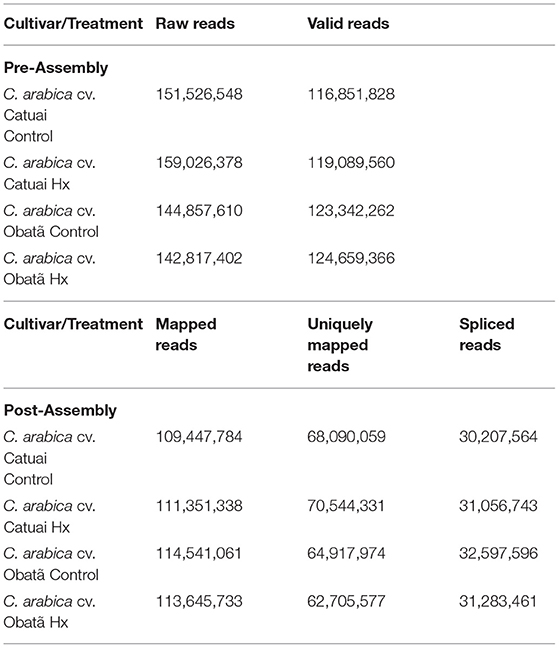
Table 1. Statistics of C. arabica cv. Catuai Vermelho and Obatã RNA-seq analysis.
Sequence annotation and functional analysis were done for the DEGs by using Blast2GO (Conesa et al., 2005), at the OmicsBox (Götz et al., 2008) platform. Sequences were annotated by blasting nucleotide sequences against the NCBI NR database (BLASTX, evalue ≤ 1.10−5). A total of 57 and 63 DEGs were found in Catuai Vermelho and Obatã, respectively. Most DEGs correspond to upregulated genes in response to Hx, in both cultivars (86% Catuai Vermelho and 73% Obatã). Eight DEGs were commonly found in the Catuai Vermelho and Obatã cultivars (AAA-ATPase, auxin-induced protein (two transcripts), ferredoxin-NADP reductase, premnaspirodiene oxygenase-like, glutathionyl-hydroquinone reductase, flavonol 3-O-glucosyltransferase and phenylalanine ammonia-lyase). DEGs were analyzed by Gene Ontology and were functionally assigned to the relevant terms. Biological process and molecular function categorizations revealed that DEGs related to cellular, oxidation-reduction, organic substance, and primary metabolic process, as well as transferase and ion binding activity might play a dominant role in the leaf response to priming. Most of the DEGs have a role in plant defense, corroborating to our hypothesis. These data contribute to identify key genes differentially expressed in response to Hx as well as indicate pathways modulated by this eliciting agent.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://www.ebi.ac.uk/ena, PRJEB39901, https://doi.org/10.5281/zenodo.5517786.
DD conceived the idea and acquired funding. PC, RR, NC, and SI-S conducted the experiment. IB performed analysis on the data. IB and DD wrote the manuscript. All authors contributed to the article and approved the submitted version.
This research was funded by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP), grant number #2016/10896-0. IB and NC were financed in part by the Brazilian Federal Agency for Support and Evaluation of Graduate Education (CAPES) – Finance Code 001. IB acknowledge the scholarship granted from the Brazilian Federal Agency for Support and Evaluation of Graduate Education (CAPES), in the scope of the Program CAPES-PrInt, process number 88887.310463/2018-00, International Cooperation Project number 88887.512173/2020-00. SI-S acknowledges FAPESP for providing a post-doctoral fellowship, process number #2017/01455-2. DD also acknowledges CNPq for a research productivity fellowship (process number #312823/2019-3).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fsufs.2021.735893/full#supplementary-material
Andrews, S. (2010). FastQC: A Quality Control Tool for High Throughput Sequence Data. Available online at: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed July 1, 2021).
Aranega-Bou, P., Leyva, M., Finiti, I., Garcia-Agustin, P., and Gonzalez-Bosch, C. (2014). Priming of plant resistance by natural compounds. Hexanoic acid as a model. Front. Plant Sci. 5:488. doi: 10.3389/fpls.2014.00488
Clark, R. B. (1975). Differential magnesium efficiency in corn inbreds: I. dry-matter yields and mineral element composition 1. Soil Sci. Soc. Am. J. 39, 488–491. doi: 10.2136/sssaj1975.03615995003900030032x
Conesa, A., Götz, S., García-Gómez, J. M., Terol, J., Talón, M., and Robles, M. (2005). Blast2GO: a universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 21, 3674–3676. doi: 10.1093/bioinformatics/bti610
de Carvalho, K., Bespalhok Filho, J. C., Dos Santos, T. B., de Souza, S. G. H., Vieira, L. G. E., Pereira, L. F. P., et al. (2013). Nitrogen starvation, salt and heat stress in coffee (Coffea arabica L.): identification and validation of new genes for qPCR normalization. Mol. Biotechnol. 53, 315–325. doi: 10.1007/s12033-012-9529-4
Del Grossi, L., Sera, T., Sera, G. H., Fonseca, I., Ito, D. S., Shigueoka, L. H., et al. (2013). Rust resistance in arabic coffee cultivars in Northern Paraná. Braz. Arch. Biol. Technol. 56, 27–33. doi: 10.1590/S1516-89132013000100004
Götz, S., Garcia-Gomez, J. M., Terol, J., Williams, T. D., Nagaraj, S. H., Nueda, M. J., et al. (2008). High-throughput functional annotation and data mining with the Blast2GO suite. Nucleic Acids Res. 36, 3420–3435. doi: 10.1093/nar/gkn176
International Coffee Organization (ICO) (2018). Statistics of Production. Available online at: http://www.ico.org/ (accessed July 1, 2021).
Kim, D., Langmead, B., and Salzberg, S. L. (2015). HISAT: a fast spliced aligner with low memory requirements. Nat. Methods 12, 357–360. doi: 10.1038/nmeth.3317
Llorens, E., Camañes, G., Lapeña, L., and García-Agustín, P. (2016). Priming by hexanoic acid induce activation of mevalonic and linolenic pathways and promotes the emission of plant volatiles. Front. Plant Sci. 7:495. doi: 10.3389/fpls.2016.00495
Martin, M. (2011). Cut adapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 17, 10–12. doi: 10.14806/ej.17.1.200
Pertea, M., Pertea, G. M., Antonescu, C. M., Chang, T.-C., Mendell, J. T., and Salzberg, S. L. (2015). StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 33, 290–295. doi: 10.1038/nbt.3122
Robinson, M., McCarthy, D., and Smyth, G. K. (2010). edgeR: a bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26, 139–140. doi: 10.1093/bioinformatics/btp616
Keywords: RNA-seq, hexanoic acid, priming, abiotic stress, Coffea
Citation: Budzinski IGF, Camargo PO, Rosa RS, Calzado NF, Ivamoto-Suzuki ST and Domingues DS (2021) Transcriptome Analyses of Leaves Reveal That Hexanoic Acid Priming Differentially Regulate Gene Expression in Contrasting Coffea arabica Cultivars. Front. Sustain. Food Syst. 5:735893. doi: 10.3389/fsufs.2021.735893
Received: 03 July 2021; Accepted: 27 September 2021;
Published: 25 October 2021.
Edited by:
Pushp Sheel Shukla, Dalhousie University, CanadaReviewed by:
Rakesh Upadhyay, United States Department of Agriculture (USDA), United StatesCopyright © 2021 Budzinski, Camargo, Rosa, Calzado, Ivamoto-Suzuki and Domingues. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Douglas S. Domingues, ZG91Z2xhcy5kb21pbmd1ZXNAdW5lc3AuYnI=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.