 Roy H. Behnke
Roy H. Behnke- Department of Anthropology, University College London, London, United Kingdom
This essay examines three central components of extensive livestock production—herd composition, grazing/pasture management, and rangeland tenure. In all of these areas, fenced, and open-range forms of migratory pastoralism face a number of shared problems. Set aside the presumption that either one of these systems is technically or institutionally more advanced than the other, and it turns out that each has lessons for the other. 1. For a variety of reasons, including climate change, we can look forward to a future world with less grass, which presents a challenge for livestock producers reliant on grass feeding livestock. With little delay and minimal scientific support, East African pastoralists are already adjusting to a new woody world by diversifying the species composition of their herds to include more browsers—camels and goats. There is a potential lesson here for commercial ranchers who have traded the stability of mixed herds for the profitability of keeping sheep or cattle alone. 2. Migratory rangeland systems distribute livestock very differently than fenced, rotational systems of livestock, and pasture management. Whereas, migratory herds exploit environmental heterogeneity, fenced ranching attempts to suppress it. Emerging archaeological evidence is demonstrating that pastoralists have amplified rangeland heterogeneity for millennia; ecological research shows that this heterogeneity sustains both plant and wildlife biodiversity at the landscape scale; and new approaches to ranch management are appropriating aspects of migratory herding for use on fenced ranches. A rapprochement between the environmental sciences, ranching, and open-range migratory pastoralism has occurred and merits wider policy recognition. 3. In contemporary Africa, indigenous tenure regimes that sustain open rangelands are eroding under pressure from market penetration and state encapsulation. At the same time in the American West, there are emerging novel land tenure instruments that replicate some of the most important functional characteristics of tenure arrangements in pastoral Africa. After many false starts, it appears that some aspects of American ranching do provide an appropriate model for the preservation of the open-range migratory systems that they were once supposed to supplant. “Development” policy needs to reflect upon this inversion of roles and its implications for accommodating diversity.
Introduction
This article is built around the distinction between the Neo-Anthropocene and the Paleo-Anthropocene (Erlandson and Braje, 2013; Foley et al., 2013). The Neo-Anthropocene came into existence around 1800, with the birth of industrial capitalism. Many of the people contributing to this book represent extensive livestock systems that were invented in response to the Neo-Anthropocene. As the organizers of this collection clearly recognize, observers of these systems are now trying to think themselves out of some of the boxes that they find themselves in1. Social anthropology in the twentieth century, and archaeology in the twenty-first century, offer an insight into alternative kinds of extensive livestock production systems, those of the Paleo-Anthropocene. These systems were remarkably resilient for nearly 10,000 years. What do they tell us about how to survive?
I will argue that these pre-industrial systems of livestock production do not offer us literal models of how to reconstruct industrial livestock production; the Neo-Anthropocene has rendered them inoperative. Especially for those of us accustomed to the comforts of consumer capitalism, they are good to think, not good to live. But we can identify the principal characteristics of these systems, identify the ways in which they differ from commercial ranching, and explore the implications of these differences. Broadly speaking there are three differences—their economy, the nature of their political organization and the source of energy that powered these societies. These were self-provisioning and redistributive economies in the main. Households produced for themselves and surpluses tended to circulate locally, often in the interests of maintaining local political institutions. Local-level politics was important because rural communities were frequently autonomous and sovereign, and their political integrity was essential to their control of vital natural resources. Finally, the power that drove these economies and societies was human labor and animal energy. Everywhere on the globe these features of Paleo-Anthropocene livestock production are eroding with expanding state power, commodification, privatization, and the substitution of industrial inputs for human labor and animal energy. It's called development, and there's no going back, but it is possible that pastoral systems that remind us of our past may provide a useful platform for thinking creatively about our future. This paper examines that possibility.
To begin we must level the playing field. By this I mean that we must, from the perspective of the Neo-Anthropocene, establish some basis for respecting Paleo-Anthropocene forms of extensive livestock production. Table 1 does this, albeit crudely. It shows that in some sense African open-range livestock production in the twentieth century was more productive—often by several multiples—than contemporaneous forms of commercial ranching operating under comparable ecological conditions. The qualifier “in some sense” is important here. Table 1 engages in the denominator game; its results are startling because it expresses output per unit land area. If output is expressed per head of livestock or per unit of human labor the results are very different and in conformity with conventional expectations—commercial ranching wins every time. For example, referencing data on African and Australia ranching in the 1970s, Jahnke estimated that “Labour productivity [of open-range pastoralism] is in the order of $50 [USD] per man instead of over a thousand or thousands in ranching. Labour productivity in pastoral systems is therefore very low, or to put it the other way around, pastoral systems are labour-intensive; they have a high employment capacity at low levels of renumeration” (Jahnke, 1982, p. 87). Cattle weights are a convenient measure of per capita livestock output and these indicate that ranching is also more productive per animal than pastoralism. Across sub-Saharan Africa as a whole, Otte and Chilonda (2002) estimated that semi-arid pastoral cattle weighed on average 61–66% of semi-arid ranched cattle, which is broadly in agreement with available country-level data. For example, communal (open range) Nguni cattle weighed 66–70% of commercial breeds raised on private ranches in South Africa (Strydom, 2008; Strydom et al., 2008). In Botswana in the 1970s, cattle kept at unfenced boreholes weighed 75% (Rennie et al., 1977) to 81–82% (Animal Production Research Unit, 1979) compared to cattle kept on freehold ranches. According to Cossins (1985) pastoral Borana cattle kept in Ethiopia weighed 54–87% of Borana cattle kept on Kenya ranches in comparable environments.

Table 1. Relative productivity of commercial ranching and open-range pastoralism under comparable ecological conditions.
I conclude that both commercial ranching and open-range pastoralism are productive, but in different ways. To understand why they are so different we can begin by examining the factors of production that underpin each system. The regions of semi-arid Africa inhabited by indigenous African pastoralists are relatively densely settled compared to areas dominated by Euro-American forms of commercial ranching predicated on the extirpation of indigenous populations. As a consequence, in pastoral Africa land is valuable whereas labor is abundant and cheap, relative to commercial ranching areas. As a result of their relatively high population levels, African pastoralists need to squeeze every bit of value out of the natural resources that they control, and they have abundant supplies of labor to devote to this effort. Table 1 suggests that they are successful—on their terms. Commercial ranchers operate under very different conditions and with very different results. Labor is expensive because ranchers must compete for it against other sectors of the economies of industrialized nations. To do this they commonly replace human labor with hydrocarbons and a variety of industrial inputs the manufacture of which is ultimately based on the consumption of hydrocarbons: “In a broad sense, the intensive use of chemical inputs and fossil energy can be viewed as substitution of petroleum and natural gas for ecological functions and labor” (Tomich et al., 2011, p. 199)2.
Is this beginning to sound at least a little bit interesting—a productive form of extensive livestock production predicated on minimal hydrocarbon consumption and offering abundant job opportunities? Alas, there are snags. The abundant job opportunities come with low wages and difficult working conditions. High levels of output are also predicted on the self-provisioning aspects—home production for local consumption—of these partially self-sufficing economies. Self-provisioning means that consumers are permanently on hand to use a wide variety of livestock goods and services—meat and dairy products certainly, but also—depending on the society and environmental setting—dung, urine, transport and traction, blood, bones, hides, and hair—almost everything an animal has to offer. Some part of the high output of pastoral herds must be attributed to the wide spectrum of both live-animal and terminal products that are harvested from them. With market exposure much of this complexity falls away as managers focus their attention on marketable commodities and abandon the production of goods and services that are now superfluous or may even interfere with efficient commodity production3. This narrowing process has been repeated time and again in twentieth century pastoral Africa and I suggest that it is a near universal concomitant of increased market involvement. To the extent that capitalism is part of our future, the self-provisioning, broad-spectrum productivity of pastoral herds is probably not a relevant model, except for devotees of self-sufficiency who wish to disengage from market-based consumerism.
More relevant are the production systems and husbandry practices used by open-range pastoralists. At least three aspects of pastoral husbandry should be of interest to other types of contemporary extensive livestock producers. These are:
• The maintenance of herds composed of multiple livestock species, and in particular, the frequent mixing of large- and small-bodied browsers and grazers—cattle, camels, sheep, and goats;
• The way open-range producers exploit and amplify environmental heterogeneity through their herding practices.
• The institutional arrangements pertaining to resource control/access/ownership that facilitate the free movement of livestock at the landscape scale.
This paper will examine each of these possibilities.
The Mixed Species Composition of Pastoral Herds
We can begin with an abiding challenge of our time—global environmental and climate change.
There is robust evidence of shrub encroachment on a global scale in arid environments and on a regional scale in semi-arid areas including parts of the western USA, Australia, Africa, and South America (Van Auken, 2000, 2009; Andela et al., 2013; O'Connor et al., 2014; Stevens et al., 2017). In addition to local land use and ecological variables—e.g., grazing intensity, fire regime, soil and vegetation characteristics—global climate change, and in particular the frequency of large precipitation events (Schwinning and Sala, 2004), and enhanced levels of atmospheric carbon dioxide, are almost certainly implicated in these trends: “…. It is likely that shrub encroachment will be augmented in the future, even if other factors known to promote this land-cover change (e.g., grazing) are reduced” (Maestre et al., 2012, p. 3,065).
As elsewhere, bush encroachment is widespread in East Africa. East Africa has also experienced a three- to four-decade-long trend of declining rainfall in pastoral areas of eastern and southern Ethiopia, Somalia, and parts of semi-arid Kenya (Pricope et al., 2013; López-Carr et al., 2014). It is instructive to consider how livestock producers in these areas have coped with the dual challenges of aridification and bush encroachment.
One component of their response has been an increase the proportion of both large and small-bodied browsing species—camels and goats—in pastoral herds. This response has built on the pre-existing species diversity of East African herds. Pastoralists kept multiple species in order to exploit diverse grazing environments and to provide a variety of products for direct household consumption. Cattle were valued for the volume of milk they produced, for instance, while camels were valued for their capacity to give at least some milk in a drought. Small stock were kept for routine domestic meat consumption or for sale to meet small expenses. Because goats could rapidly multiply, pastoralists often used goats to rebuild their herds in the recovery years following a drought.
As late as the 1960s, however, herding societies in East Africa could be divided roughly into two groups based on the species composition of their herds—the cattle specialists vs. those who kept herds with a more diverse mix of species. Archetypal cattle keepers included the Maasai of Kenya/Tanzania (Jacobs, 1975; Galaty, 1982), the Borana of Ethiopia/Kenya (Dahl, 1979a,b), and the Pokot of Uganda/Kenya (Österle, 2008). Alternatively, the Turkana of Kenya (Coughenour et al., 1985) and the Karamajong of Uganda (Dyson-Hudson and Dyson-Hudson, 1969) exemplified the mixed species option, keeping as many as five types of livestock—cattle, camels, sheep, goats and donkeys—and selling, slaughtering, bleeding and milking all five species. Broadly speaking, the cattle specialists occupied the better-watered grasslands of the region, while mixed herds were located in more the arid areas and tended to rely for their forage on a combination of grazing and browsing.
Declining precipitation, increased frequency of drought, bush encroachment and reduced grassland cover and species diversity have in recent decades undermined the viability of cattle pastoralism and increased the attractiveness of livestock species that are recognized by pastoralists as better adapted to their changing environment (Megersa et al., 2013, 2014). The diversification of the herds kept by what were once iconic cattle-owning peoples is now well-documented.
• At one Maasai Group Ranch in Laikipia, Kenya, in 1980 there were no camels; by 2015 at the same ranch 10% of all households owned camels (Volpato and King, 2019). In all of Mukogodo Division of Laikipia District in Kenya in 1983 there were 254 camels; by 1998 there were 3,500—a more than 1,200% increase. Over the same period, the traditional cattle-dominated herds of the Mukogodo Maasai were being replaced by sheep and goat-dominated herds (Huho et al., 2011).
• In East Pokot, Kenya, the estimated maximum number of camels in the 1980s was 3,500 head; by 2011 the number had increased more than twofold to 9,600 (Bollig, 1992, 2016). Long-time observers of this area agree that, “… small stock numbers have hugely increased” (Bollig, 2016; Vehrs, 2016). In a paper entitled “From Cattle to Goats,” Österle argues that the East Pokot cattle herd oscillated around 100,000 head from 1920 to 2005, while the goat herd increased more than fivefold over the same period (Österle, 2008).
• Since the 1980s, the Ethiopian Borana of Yabello and Dire districts have doubled the contribution of sheep and goats to the composition of their livestock holdings (Cossins and Upton, 1987; Megersa et al., 2013) and involvement in camel keeping has expanded from 6% of households in 1980 to 40% in 2011 (Megersa et al., 2013). Wako et al. (2017) also report the expansion of camel husbandry in Yabello District. In northern Kenya, the Borana of Isiolo County have recently diversified their livestock holdings such that more than 40% of households now own camels (Kagunyu and Wanjohi, 2014). In nearby Marsabit County, by 2012 camels were being kept at higher altitudes by people who rarely kept camels in 2000 (Watson et al., 2016).
National statistics on livestock numbers in Kenya confirm the generality of these trends. Since the 1970s, the national cattle herd has more than doubled growing by 113%, but camel numbers have expanded by 574%, goat numbers by 483%, and sheep by 381% (FAO STAT). Though less clear-cut, global trends reflect developments in Kenya. Since 1980, global sheep numbers have been largely stable (up by 9%) and cattle numbers have expanded modestly (+23%) while goat populations have more than doubled (+123%), led by Africa (+200%) (FAOSTAT4). FAO data on world-wide camel populations are incomplete and unreliable, but a comprehensive review suggests that:
Between 1961 and 2018, the world camel population was multiplied by 2.75, a higher value than equines (1.06), sheep (1.21), cattle (1.58), small camelids (1.72) and buffalo (2.33). Only the growth of goat population appears higher (3.00). Such development testifies to the impact of climatic changes marked by widespread desertification of vast stretches of land in the world and of the renewed interest in the camel within this new global climatic context. It also highlights the growing interest for camel products (Faye, 2020).
Estell et al. (2012) correctly anticipate a future “world with less grass”: “Grasslands are in decline (a trend expected to continue) for a number of reasons (e.g., competing land uses, urban sprawl, and invasive species), though two dominant factors are conversion to cropland and woody plant encroachment” (Estell et al., 2012, p. 553). They have recommended a range of science-based responses including genetics and selection, detoxification, dietary supplementation, and behavioral modification to enhance the ability of livestock to consume shrubs.
The East African pastoralist response documented here has been more immediate—a shift to goat and camel production—and there is field evidence that these simple adaptations have helped people of modest means to rapidly ameliorate the negative impacts of global climate and environmental change:
… diversification of livestock species was associated with shorter periods of food deficit, better dietary intake and lower magnitude of household food insecurity…… Generally, livestock diversification significantly affects off-take and consequently improves access to food. Thus, multiple species herding does not only offer food products but also more ample choices for off-take, which can be liquidated in times of shortage and can smooth consumption (Megersa et al., 2013).
East African pastoralists have also been able to convince regional consumers to adjust their buying habits. Evidence of this flexibility is provided by the emergence of a new livestock commodity—commercially sold camel milk—first reported in 1990 (Herren, 1990) and now established in urban markets in Somalia, Ethiopia, and Kenya (Akweya et al., 2012; Anderson et al., 2012; Abdullahi et al., 2013; Noor et al., 2013; Elhadi et al., 2015; Gebremichael and Girmay, 2019).
In a recent keynote address to the Australian Rangeland Society, Walker (2019) noted the prevalence of mixed grazers and browsers on African savannas and the first glimmers of interest in Australia for domestic mixed-species goat husbandry. Huntsinger et al. note that American ranchers have “traded the stability of mixed herds for the efficiency of uniform production, with most ranchers relying on cattle alone” (Huntsinger et al., 2010, p. 17). Eldridge et al. have questioned the distorting effects of “a single land use: pastoralism involving grass feeding livestock” (Eldridge et al., 2011, p. 720) and the coupling of degradation with bush encroachment in scientific assessments of ecosystem structure and functioning.
With little delay and minimal scientific support, East African pastoralists are already adjusting to a new woody world that ranchers in advanced economies and their scientific compatriots have now noticed but have taken few practical steps to accommodate.
Fenced Rotation or Open-Range Migration
This section explores a possibility that would have been considered preposterous a decade or so ago: Migratory systems of production do a better job of distributing livestock over space and time than fenced, rotational systems of livestock and pasture management.
Multiple factors, including fundamental advances in theoretical and applied ecology, lend credence to this possibility, but declining scientific confidence in the utility of rotational grazing systems has also contributed. Between 1948 and 2003 roughly two out of every five articles in the Journal of Range Management—the preeminent journal of range science in North America—were about fenced “rotational” grazing systems (Brown and Kothmann, 2009). Reflecting this enthusiasm, for the last 50 years international development agencies have promoted fenced grazing schemes as a modern substitute for migratory livestock- keeping in pastoral Africa. These efforts met with limited success. Occasionally pastoralists did adopt fencing and deferred grazing, not necessarily because they thought it improved forage output or animal performance, but because it was subsidized or officially enforced, saved herding labor, established privileged (and sometimes private) access to collectively owned resources, or simply locked down their property rights. More commonly, donor-funded rotational grazing schemes collapsed whenever foreign personnel, money or enforcement were withdrawn (Sandford, 1981, 1983).
Interpreted at the time as irrational conservatism, pastoral reluctance to adopt rotational grazing makes sense in terms of the most systematic metanalysis yet conducted of the performance of these systems: “[S]ubjected to as rigorous a testing regime as any hypothesis in the rangeland profession,” rotational grazing systems have been found to “convey few, if any, consistent benefits” and it is likely that “… a continuation of costly grazing experiments adhering to conventional research protocols will yield little additional information” (Briske et al., 2008, p. 11; see also Heady, 1961; O'Reagain and Turner, 1992; Holechek et al., 2001; Bailey and Brown, 2011; Hawkins, 2017). Despite the decades of negative or mixed results in the works cited above, the debate about the efficacy of rotational systems in semi-arid rangelands grinds on without resolution (Teague et al., 2013; Briske et al., 2014). The safest conclusion may be that the advantages of rotational systems are either modest and difficult to detect, or so contingent upon local circumstance or skilled management as to make them difficult to replicate. Irrespective of the ultimate outcome of the debate, at this late date rotational grazing seems unlikely to produce any dramatic breakthroughs5.
Two recurrent features of migratory pastoralism set it off from the management practices associated with fenced grazing systems: the exploitation and potential amplification of environmental heterogeneity and the practice of herding/shepherding. These are discussed below, followed by an examination of the benefits and liabilities of fenced sedentary livestock systems vs. open-range migratory pastoralism.
Intrinsic Heterogeneity
Migration does not arise without functional environmental heterogeneity. If resources are constantly available, evenly distributed, or highly concentrated it makes little sense to engage in movement at the landscape scale—i.e., to migrate. A wide variety of environmental gradients encourage migration, including:
• Differences in elevation create vertical zonality in temperature, precipitation, vegetation, and the seasonal calendar, which all support transhumance—regular up-slope-down-slope movement to access resources and avoid extreme weather in mountainous areas (e.g., Barth, 1959).
• Across the temperate grasslands of the Eurasian steppes, changes in latitude create north-south horizontal zonality. As in mountainous areas, herds move both to access resources and avoid extremes of weather, but the movements might take place on a vast continental scale—north in spring tracking the green-up of the vegetation, south as winter approaches to avoid the worst of the snow and cold (Khazanov, 1994).
• Especially in the semi-arid tropics, precipitation gradients provoke movement. Lower rainfall areas provide high quality grazing in seasons when plants are growing, forage is relatively abundant, and herds need a nutritional boost to support reproduction. High rainfall areas provide abundant, low-quality forage in seasons when plants in lower rainfall areas are senescent, forage is scarce, and animals cannot afford to be selective. Migration occurs as herds shift from reproduction to survival by moving between areas of low and high plant biomass (Behnke et al., 2020).
• Topographically complex landscapes can support grazing habitats that are situated in close proximity to one another but differ markedly in their soils, drainage and vegetation characteristics. Because they are responding to micro-variations in their environments, herds may not migrate great distances (Scoones, 1995).
Despite obvious differences in scale, localized movements within a catena and long-distance migrations “are in reality exactly equivalent” (Bell, 1971, p. 92)—wildlife or livestock move along environmental gradients to access asynchronous pulses of resource abundance and escape temporary periods of localized scarcity. Following this strategy, heterogeneous environments can support larger migrant populations than similar but fragmented environments that are exploited by separate sedentary populations (Behnke and Scoones, 1993; Boone and Hobbs, 2004; Boone, 2005). Prior to the introduction of firearms and large-scale commercial fishing, the natural world provided evidence of the fecundity of the migratory strategy in the form of massive concentrations of animal biomass: the North American bison (Bison bison, Epp and Dyck, 2002) and passenger pigeons (Ectopistes migratorius, Schorger, 1955), the saiga antelope (Saiga tatarica) of the Asian steppes (Yagodin and Amirov, 2014), herds of migratory African herbivores (Venter et al., 2017), migratory fish stocks (Rosenberg et al., 2005)6, and global whale populations (Roman and Palumbi, 2003).
The capacity of migration to also support large livestock populations has long been recognized (Behnke and Scoones, 1993) but frowned upon in conventional ecological theory as a cause of overgrazing and environmental degradation (Illius and O'Connor, 1999). Supported by recent advances in pastoral archaeology and ungulate ecology, this negative assessment is now subject to qualification.
Engineered Heterogeneity—Natural Lawns, Anthropogenic Glades, and Semi-Natural Landscapes
The impact of migratory livestock on rangeland resources is complex and defies simple characterization. In tropical environments characterized by extremely low and variable rainfall, droughts may be frequent enough to hold livestock populations in check and minimize the impact of their grazing on pastures (Ellis and Swift, 1988), a hypothesis confirmed by meta-analyses based on decades of field studies (von Wehrden et al., 2012; Engler and von Wehrden, 2018). Extending the non-equilibrium model of ecosystem dynamics beyond the semi-arid tropics, research now suggests that extreme cold may buffer vegetation from herbivore impacts in some temperate and arctic rangelands (Begzsuren et al., 2004; Kerven, 2004; Sternberg, 2012).
There nonetheless remain many pastoral environments in which livestock do affect their grazing resources. Some sense of current research on these more “equilibrial” grazing systems is revealed by examining a phenomenon of increasing analytical significance—nutrient hot-spots—concentrations of soil, vegetation, and herbivore fertility in the form of grazing lawns or glades scattered across rangeland landscapes.
The concept of grazing lawns developed out of work on grazing successions on the Serengeti savannah in East Africa (Gwynne and Bell, 1968; Bell, 1971). Grazing successions referred to the regular sequence in which different ungulate species occupied an area. Generally, large-bodied bulk feeders moved in first, opening up the sward by removing coarser, more mature vegetation, and were followed by smaller-bodied more selective graziers who took advantage of the shorter, less mature, and more nutritious forage that had been exposed by the bulk feeders. Based on differences in anatomy, physiology, and dietary requirements, “The relationships between [herbivore] species in such a grazing succession can thus be seen to be facilitative rather than competitive,” which explained in part both the large migratory populations of Serengeti ungulates and their propensity to form herds (Gwynne and Bell, 1968, p. 393).
In a series of seminal publications, McNaughton (1979, 1984) expanded Bell's concept of facilitation between grazing animals to include the facilitation of plant productivity by grazing, which created grazing lawns. The enhanced productivity of the lawns was achieved through compensatory plant regrowth in response to grazing and—over time—the development of grazing-tolerant grass species and plant communities with higher nutrient quality and productivity than vegetation in ungrazed areas (McNaughton op.cit.). Humans were not viewed as a dominant force in shaping these landscapes. The shifting mosaic of vegetation in the Serengeti was instead attributed, as McNaughton phrased it in the title of a 1983 paper, to “composite environmental factors and contingency” driving the distribution of large migratory herds (McNaughton, 1983, p. 291).
The effects of fertilization by nutrient recycling from dung and urine—the latrine effect—was initially characterized as “so well-known that it warrants little additional comment,” at least from a biological perspective (McNaughton, 1979, p. 36), and through the 1990s investigations of the impact of soil fertilization on vegetation and herbivore behavior were indeed “relatively limited” (Augustine et al., 2003). Significantly, those studies that did exist tended to be conducted by researchers interested in the environmental impacts of pastoralists and pastoral livestock (Reid and Ellis, 1995; Young et al., 1995; Turner, 1998a,b).
After 2000 this literature expanded rapidly and focused on nutritional hotspots termed “grazing glades”—grass-dominated “islands of high fertility and high plant biomass” in wooded savannah environments (Muchiru et al., 2009, p. 322). Like grazing lawns, these grazing glades attracted high concentrations of both wild and domestic graziers (Young et al., 1995; Augustine et al., 2003; Muchiru et al., 2008; van der Waal et al., 2011; Porensky et al., 2013a). Unlike the lawns, however, the glades were situated on the dung left behind in abandoned livestock kraals and hence had a clear pastoral origin. The elevated levels of contemporary grazing, defecation and fertilization that researchers observed were perpetuating a legacy of past human occupation. The glades were also “creating a relatively permanent community that increases ecosystem heterogeneity” (Young et al., 1995, p. 97) through the redistribution of nutrients from peripheral bushland sites to the glades (Augustine et al., 2003) contributing to the heterogeneity (Muchiru et al., 2008; van der Waal et al., 2011; Porensky and Veblen, 2012) and biodiversity (Donihue et al., 2013; Porensky et al., 2013a,b) of both wildlife and nutrient-poor savanna vegetation.
From the perspective of archaeology, Marshall et al. could declare by 2018 that “the processes creating these glades are well-understood” although “the full time-depth of their creation and effects on African savannahs are as yet unexplained” (Marshall et al., 2018, p. 387). A “virtual fluorescence of archaeological research in traditionally pastoral nomadic regions” (Honeychurch and Makarewicz, 2016, p. 342) has in the last two decades equipped archaeologists to answer these outstanding questions and the answers are unequivocal: “herders have had a role in structuring and diversifying African savannah ecosystems for up to three millennia” and:
Pastoral Neolithic and Iron Age sites in diverse Kenyan savannahs demonstrate the spatial influences of niche construction by pastoralists on soil nutrients and savannah heterogeneity, on timescales that range from five centuries to three millennia (Marshall et al., 2018, p. 389).
The pastoral exploitation and amplification of environmental heterogeneity—and the capacity of archaeology to document these processes—is not confined to Africa. The Eurasian rangelands comprise the world's largest contiguous area of grazing (Babaev and Orlovsky, 1985; Mirzabaev et al., 2016), comprising 25% of the world's total rangelands and over 6% of the total world land area (FAOSTAT “permanent pastures”). The impact of pastoral livestock in pre-historic times across Eurasia is indicated by the spread of plant species with endozoochoric (ingested) seeds dispersed by herded animals, concentrations of plants with defenses against grazing, and—as in East Africa—grazing-mediated “hot spots” that contain nutritionally-rich vegetation (Spengler, 2014; Ventresca Miller et al., 2020). Six to eight millennia ago on the Tibetan plateau, pastoralists—in conjunction with Holocene climatic fluctuations and fire—transformed forests into alpine meadows suitable for herding (Miehe et al., 2009; Schlütz and Lehmkuhl, 2009). In southwestern Turkey within the last 600–700 years, pre-modern mobile pastoralists:
[D]id not merely exploit agriculturally marginal land; they… transformed this territory into a productive herding landscape through the construction of infrastructure, altering vegetation patterns and water availability, and sheltering themselves and their animals with locally available materials (Hammer, 2014, p. 285).
In the Middle East as in East Africa, ancient pastoralists left a permanent mark on the land by creating what Hammer has called landscape anchors—“geographic foci that structured the spatial organization of local landscapes” (Hammer, 2014, p. 269).
Herding
The previous discussion documented the results of herd movement: Viewed in the medium term across seasons and years, herds track environmental variability; viewed in the long term across decades and centuries, they reinforce it. This section examines herding in the short term—moment by moment, day by day—to better understand how it achieves these outcomes. Putting aside a host of complicating factors, migratory livestock move like migratory wildlife to wherever they can find the most favorable conditions at any time. Unlike migratory wildlife, however, domesticated ungulates are accompanied by humans and animal priorities are subject to abridgement or refinement in light of human judgement and social, economic, and political considerations. Very briefly, the following case studies illustrate these human-livestock interactions.
The Nenets reindeer herders of the Russian Arctic (Dwyer and Istomin, 2008), and the Wodaabe Fulani cattle pastoralists of the African Sahel (Krätli, 2008; Krätli and Schareika, 2010) keep different livestock species in very different environments. Both groups are atypical in their unusually high level of herding skills, which bring into sharp focus a recurrent challenge facing livestock keepers on the open range—the need to reconcile human and livestock priorities and decision-making (Stammler, 2005; Istomin and Dwyer, 2010; Stépanoff, 2012). As the following discussion shows, half-wild Arctic reindeer (Rangifer tarandus) exemplify the capacity of the animals to influence the grazing agenda; conversely, the Fulani ability to refine the behavior of their exceptionally docile cattle illustrates the contribution of the herder.
Reindeer exhibit a radical degree of animal agency because they are only semi-domesticated. The most common long-distance migratory pattern for reindeer—both domestic and wild—is to follow plant growth “advancing north with the greening of spring pastures and retreating south as plants senesce in autumn ….between lichen-rich winter pastures in a forest zone and herbaceous summer pastures at windy locations on the coast, where insect harassment is reduced, or at high altitude” (Stammler in Behnke et al., 2011, p. 159). Because wild and domesticated reindeer are biologically similar and occupy the same grazing ranges at the same time, domesticated reindeer have the capacity to abandon their owners and join wild herds. This vulnerability makes controlling reindeer movement a paramount concern for Nenets pastoralists. Effective movement control has two elements—rounding up animals to keep the herd together and controlling the speed at which the assembled herd moves in a desired direction and away from dangerous terrain, predators, pests, and other herds (Dwyer and Istomin, 2008). The two processes—gathering the herd and moving it forward—are interdependent. Success in holding the herd together rests on knowing when the herd should move to new pastures, and the reindeer themselves play a central role in making this determination:
[Reindeer are] very sensitive to even the slightest change to the environment. Thus, when making movement decisions, the herders, rather than constantly assessing an incalculable number of environmental factors and moving accordingly, generally attune their actions to environmental variability by responding to changes in reindeer behaviour alone…. [M]ovement is made according to (albeit not solely) the degree of effort that is required by the herders to keep his animals under control on this pasture. The herders move when reindeer no longer want to stay on the pasture (Dwyer and Istomin, 2008, p. 530).
“[R]eindeer pastoralism rests on successful deciphering of herd behavior by the herders” (Paine, 1994, p. 31), but domesticated reindeer must also accommodate human needs. Migratory routes are adjusted to reflect administrative boundaries or other institutional restrictions, to accommodate marketing or the resupply of herders, to permit herders to engage in non-pastoral activities such as hunting or fishing, or simply to give the humans a rest. Reindeer herding is a reciprocal relationship, a process of “day-to-day symbiotic domesticity” (Stammler in Behnke et al., 2011, p. 164).
In common with the Nenets and most other migratory pastoralists, the Wodaabe of West Africa follow a seasonal migratory cycle. They graze sand dunes early in the rains to exploit the ephemeral vegetation that emerges quickly following rain, moving as the dry season progresses onto clay plains with heavier soil that retains moisture and supports plant growth for a longer period. However, in the very low rainfall areas inhabited by the Wodaabe, seasonal regularity is complicated by the erratic spatial and temporal distribution of rainfall from year to year. By moving opportunistically in response to the unpredictable distribution of rainfall, the Wodaabe prolong the time their herds can graze on fresh vegetation before facing the hardships of the dry season:
Wodaabe herders do not consider scattered rainfall as a constraint…. For them it is a naturally provided mechanism by which they can control the availability of fodder resources according to the stage of growth in which they have attained best nutritive value. If rainfall were equally distributed in time and space, grass would develop beyond the state of optimal nutritive value everywhere at the same time and herders could exploit it only for a rather short period. The scattered nature of rain brings about a sequential series of beginnings of the vegetative cycle within one pastoral zone that herders can systematically exploit…. (Schareika, 2001, p. 73).
This is not a risk-averse way to make a living. In their pursuit of the highest quality pastures on the margins of an unpredictable environment, the Wodaabe are—in a manner roughly analogous to a professional gambler—embracing the benefits that come from “living off” uncertainty (Krätli and Schareika, 2010).
The Wodaabe control their herds through breeding and socialization, to a level not possible among the Nenets with their half-wild animals. Over generations of cattle and humans, the Wodaabe structure their herds around matrilineal cattle lineages, know the genealogy of each animal, carefully regulate mating, and cull underperforming animals by selling them. The object of this breeding programme is to select animals capable of the high levels of mobility and selective feeding that will enable them to harvest the best forage in their environment. Anatomically, these are large animals capable of migrating long distances to reach the best pastures, and animals with a slender head and small muzzle, which enables them to eat the short, nutritious vegetation without ingesting soil. Social traits are also important. Bororo cattle are put at ease by the presence of humans and are loyal to their owners in particular, reducing their stress when they are handled:
[H]uman-driven tasks are performed by these cattle in virtually complete absence of coercion. The cattle bred by the Wodaabe know nothing of enclosures, follow their herder of their own accord (rather than requiring to be herded from the rear) and it is common, in the bush, to see entire herds controlled by one or two young children waving only a twig (Krätli, 2008, p. 25).
This “persuasive management style” (Krätli, 2008, p. 26) permits the Wodaabe—who know their pastures intimately—to guide their cattle to “maximize opportunities for selective feeding” (Krätli and Schareika, 2010, p. 612).
The Wodaabe are aware that cattle eat more (that is the herders' goal) when they like what they feed on. Therefore, the herders are always seeking to stimulate their animals' appetite by leading them to fodder that, in their experience, the herd will particularly appreciate (the herders talk about favoured fodder with reference to ‘tastiness' and to how much the animals look ‘at ease' when feeding on it). They prefer certain species for these characteristics and target them consistently. Moreover, they enhance feeding performance by avoiding half-dry grass during the rainy season, or pasture soiled or malodorous from cattle droppings (Krätli and Schareika, 2010, p. 611).
Managing for or Against Heterogeneity
There is considerable variety in the way both migratory pastoralism and fenced ranching are practiced, which makes it difficult to rigorously compare them. The preceding account nonetheless suggests that these two forms of extensive livestock production distribute livestock very differently across a rangeland environment.
Underlying these differences are contrasting responses to environmental heterogeneity. Migratory pastoralists respond to variability—both temporal and spatial—by moving to seize opportunities and avoid problems. By seeking to exploit environmental heterogeneity they can—in certain circumstances—amplify it, a pattern that is well-documented in the archaeological record and by the ecological research reviewed in this paper. The Nenets and Wodaabe case studies illustrate how these results are produced by migration. At the landscape scale, it is unlikely that different seasonal pastures will be exposed to uniform levels of grazing. Types of pasture that remain attractive for a long period of time but cover a small area may be exposed to more grazing pressure than extensive pastures that are useful for a short period. Landscape attributes that have little to do with forage—insect pests, predators, slope, aspect, water sources in semi-arid environments or protection from snow or cold in temperate environments—may also produce uneven levels of pasture use, as herds congregate or avoid areas for reasons other than forage availability. Any differences in grazing pressure will be compounded by seasonality. Operating in unpredictable environments, pastoral herds are unlikely to use exactly the same pasture patch from year to year, but they do tend to use the same kind of pasture every year and do so at the same point in the seasonal cycle of plant development. In such migratory systems different vegetation communities are repeatedly subject to stress at the same point in their growth cycle, and the level of stress is potentially high:
“Migratory species, by avoiding seasons of resource scarcity or heightened mortality risk, may be able to sustain much larger populations than otherwise similar resident species. Indeed, migrants are often far more abundant than their closest resident relatives…and the community and ecosystems impacts are therefore bound to be of greater magnitude” (Holdo et al., 2011, p. 134).
Holdo et al. are referring in this quotation to wild migratory species, but their conclusions also apply to domesticated migrants. For semi-domesticated reindeer, the similarities between wild and pastoral herd movement are self-evident. The parallels are real but less obvious in the case of the intensively domesticated Wodaabe cattle. All the efforts of Wodaabe herders are directed at obtaining the best forage for their cattle. Subject to constraints like pest infestations, difficult terrain, or predator risk, freely distributing wild ungulates have the same objective. The art of Wodaabe herding is to facilitate a domestic analog for a natural process whereby herbivores pursue and attempt to match ephemeral resource distributions. When the opportunity arises, pastoral herds not only target the best pastures but exploit them selectively for the most attractive species within them, not necessarily “managing” but potentially affecting their forage base.
To the extent that they may unintentionally exacerbate resource heterogeneity, migratory systems are antithetical to the objectives of formal systems of grazing rotation. There is “little doubt that grazing systems result in better distribution of livestock and more uniform utilization of the range” (Stoddart et al., 1975, p. 297). Uniformity is promoted by a constant design feature of all fenced grazing schemes, whether they are based on rotation, deferred-rotation, rest-rotation, or short-duration. Unlike seasonal migrations, these schemes subject part of a ranch to stress or resting, and then reverse the process. No section is grazed or rested year after year in the same season. In this way, the intrinsic differences between paddocks within a grazing rotation are minimized by subjecting all of them to roughly equivalent levels of grazing pressure and compensatory relief, while rotating the periods of relief and stress annually or through the seasonal calendar: “Rotation of season of use on ranges unquestionably has advantages. Plants vary greatly in their season of palatability. Under rotation grazing, different plants will be grazed at one season then another resulting in all being more equally utilized” (Stoddart et al., 1975, p. 297). On degraded pastures, short-duration rotational systems have the added appeal of compelling livestock to graze a paddock unselectively, thereby consuming both preferred and less preferred vegetation and allowing preferred forage species to recover (Teague et al., 2008; Crawford et al., 2019).
In sum, animal decision-making is constrained in all rotational grazing systems, by fences at the paddock level and, in short-duration systems, by high stocking rates within paddocks. The assumption behind these systems is that humans need to take charge since they understand and can optimize forage-grazier interactions. The prolonged controversy over the efficacy of rotational grazing suggests, however, that this assumption may be premature. By contrast, in the absence of fences, herding in migratory systems is a consensual inter-species relationship in which humans have real but limited coercive powers. Since they are not sole decision-makers, humans do not need to perfectly understand the myriad interacting variables that drive these complex systems. They let—or are forced to let—their animals do some of the thinking for them, and different migratory patterns emerge out of the interplay between humans, livestock, and the changing environment. Rotational grazing systems are designed; migratory systems evolve.
In practical terms, with respect to livestock movement and distributions, the differences between migratory and rotational range management are considerable. If migratory grazing promotes heterogeneity, rotational systems suppresses it:
“Prevailing rangeland management practices emphasizing even distribution of livestock use have decreased both temporal and spatial heterogeneity” (Derner et al., 2009, p. 111). Most management activities in domestic grazing systems promote uniform grazing distribution. At the landscape scale, herding, water development, and fencing are used to manipulate animal distribution, and may play a larger role in transforming native grazing systems than the substitution of domestic grazers for wild ones (Adler et al., 2001).
Two lines of research—both offering improved livestock production in combination with rangeland conservation—offer some indication of what it might mean to reverse the trend to uniformity by extending pastoral management practices into areas not currently occupied by traditional pastoral societies.
With respect to conservation, the management of grassland birds is instructive. Since 1966, bird species dependent upon grassland habitats have been the most rapidly declining category of birds in the United States (Fuhlendorf et al., 2012). While multiple factors are involved, these declines “were simultaneous with nationwide improvements in rangeland condition and rangeland health, as our profession has [conventionally] defined these terms” (Fuhlendorf et al., 2012, p. 583). Grassland bird species prefer rangeland vegetation of variable density, height, and species composition (Figure 1), and the bird species most in decline have been those dependent on either very tall or very short vegetation and bare ground, pastures in “poor condition” from the perspective of uniform pasture management.
Traditional emphasis on homogeneous use of vegetation (i.e., “management to the middle”) at the pasture scale has resulted in the lack of suitable habitat for grassland birds at the extremes of the vegetation structure gradient in semiarid rangelands (Derner et al., 2009, p. 116).
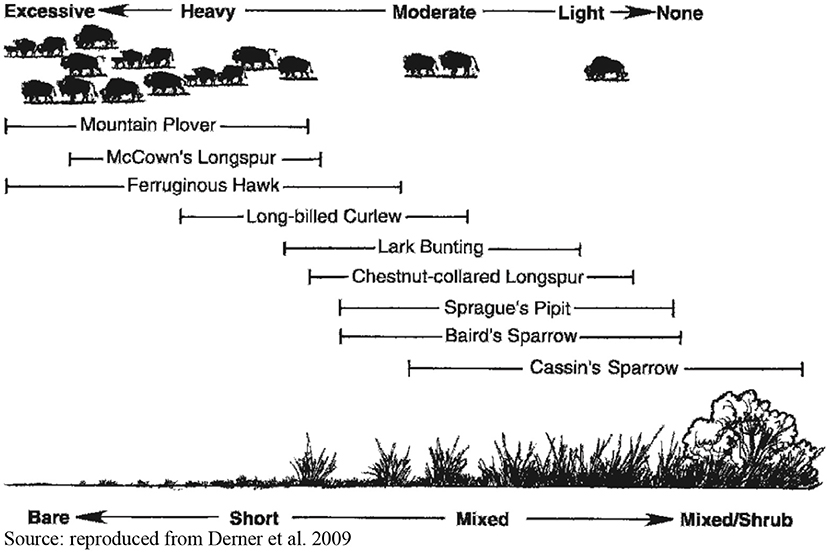
Figure 1. Responses of grassland birds in shortgrass steppe to a vegetation structure gradient. Source: reproduced from Derner et al. (2009).
In tallgrass prairie, vegetation stands at different heights can be created by a combination of burning and the free movement of cattle or bison that are either attracted to or avoid patches at various stages of post-fire regeneration. By promoting spatial discontinuities in grazing pressure, a patchwork of burned, unburned, and recovering areas creates a shifting vegetation mosaic that does not significantly depress and can increase cattle weight gain (Fuhlendorf and Engle, 2004; Limb et al., 2011), stabilizes primary and secondary productivity over time (Allred et al., 2014; McGranahan et al., 2016) and increases wildlife biodiversity (Fuhlendorf et al., 2006, 2010, 2017). As the variable buffalo numbers depicted at the top of Figure 1 suggest, there is no proper rangeland stocking rate from the perspective of avian conservation, which requires a heterogeneous environment to meet the requirements of different bird species.
Consistent with a focus on conservation, work on “pyric herbivory” at spatial scales that enable migratory grazing has primarily taken place in conservation areas with herds of bison, not cattle, and not with herded cattle. In the published material on North America reviewed for this article, there was no indication that the advocates of pyric herbivory were aware of the similarities between it and migratory livestock husbandry7.
For the proponents of pyric herbivory, the “uniformist” argument for rangeland use was exemplified by practices advocated in the standard textbooks of mainstream North American range management at the end of the last century (such as Stoddart et al., 1975; Holechek et al., 2001). For the proponents of an adaptive response to resource heterogeneity (Fynn, 2012; Fynn et al., 2016), the critique of uniform range use is directed at the “conceptual and theoretical flaws” underpinning a more recent manifestation of homogenous rangeland exploitation—short duration rotational grazing schemes (Fynn, 2012, p. 324). It is argued that the rigidity of these systems imposes artificial constraints that interfere with the free circulation of grazers in a typically pastoral migratory pattern, between “high-quality resources, to enable population growth, and reserve or buffer resources, to sustain the population after favored resources have been depleted, or are no longer accessible” (Owen-Smith and Novellie, 1982, p. 768). With minimal awareness of their similarity to pastoral migratory practices, recommendations for improved management are modeled on wild herbivore research and aim to create “similar heterogeneity in commercial rangelands … as well as … smaller conservation areas” (Grant et al., 2019, p. 7). With more ambition, others see the exploitation and enhancement of functional environmental heterogeneity as a basis for the management of broad landscapes and for wildlife-livestock co-existence on African savannahs (Fynn et al., 2016, 2019).
There are, in sum, multiple technical reasons to be optimistic about the future of migratory pastoralism and about the possibility that it can and should contribute to the improved design of all kinds of extensive livestock production. As reviewed in this paper, this optimism is grounded in progress in at least two distinct areas of scientific research:
• Advances in pastoral archaeology have recalibrated our understanding of the pristine, transforming apparently “natural” rangelands into “working wilderness” (Sayre, 2005) that has been shaped on an evolutionary timescale by both domesticated and wild herbivores.
• With respect to rangeland science, pyric herbivory emphasizes the use of engineered disturbance to enhance heterogeneity and build resilience, while the strategic management of functional heterogeneity recognizes mobility as an effective and sustainable use of rangeland resources. These approaches challenge both the uniformity-enhancing practices of mainstream range management in the last century and the restrictive practices of rotational grazing schemes. These approaches also constitute a significant rapprochement between migratory pastoralism and rangeland science and provide an unintentional legitimation of existing migratory practices.
Rangeland Tenure—Preserving Scale
As Huntsinger observed, ‘The name, “rangeland” implies a land for ranging' but “The extent and inherent flexibility of pastoral systems clashes with the increasingly fragmented landscapes and hardening borders of today's world” (Huntsinger, 2016, p. 316). Without large areas of open rangeland, migratory livestock production is a practical impossibility. This section draws on the example of indigenous pastoral land tenure systems in Africa to identify modern property arrangements that could meet the needs of today's migratory ranchers.
The commodification of rangelands in the Americas, Australia, and southern Africa occurred several centuries ago with European colonization and the spread of the “Euro-American ranching complex” (Strickon, 1965). Well into the middle of the twentieth century, however, much of the rangelands of pastoral Africa and Asia had escaped commodification. State socialism and pastoralist collectivization held the line in Soviet Central Asia (Kerven et al., 2020) and Mongolia (Sneath, 2003), as did the Chinese Communist Party in China's western provinces (Banks, 2003). In sub-Saharan Africa, the limited administrative reach of newly-created nation states left marginal pastoral areas to their own devices, and many if not most African pastoralists gained access to natural resources by being members of indigenous political communities rather than citizens of nation states (Cunnison, 1966; Dyson-Hudson, 1966; Hoben, 1988; Bassett, 1993; Turton, 1994).
Indigenous land tenure regimes provided the institutional framework that sustained Africa's open rangelands and associated migratory systems of livestock production, but they have not been robust in the face of market penetration and the expanding power of national administrations. In recent decades, the pastoral conception of land rights as an entitlement defined by group affiliation has been effectively challenged by a commercial concept of rangeland as a privately-owned and tradeable commodity secured by legal title backed by the power of the state (Behnke, 2018). These legal changes have been accompanied by rangeland fragmentation, reduced pastoral mobility, and increased environmental degradation. It is unclear to what extent these changes have been brought about by the erosion of indigenous land tenure systems or by other developments that have occurred simultaneously—increased population pressure, the growth of small towns in pastoral areas, land conversion for agriculture, mining and industry, the attractions of urban services and wage employment, and the expansion of protected conservation areas (Liao et al., 2020; Lind et al., 2020).
Just as the rangeland sciences are on the cusp of understanding the value of livestock production conducted at a landscape scale, it would appear that many African pastoralists are losing their capacity to do so. Contemporary Africa, therefore, provides few obvious lessons for the design of successful pastoral tenure systems, but the ethnographic record does contain numerous examples of indigenous tenure systems that, until recently, did operate at landscape scales. If these historical systems are being rendered obsolete, the “design principles” that they exemplify may nonetheless indicate the functional characteristics required of any successful modern replacement.
At least three recurrent features of traditional African pastoral tenures have a functional significance—their communal ideology, the collective benefits they confer, and their exploitation of the partible nature of property rights. This section argues that recent innovations in pastoral tenure in ranching areas of southwestern USA are pioneering approaches to rangeland tenure that duplicate in a legalistic and heavily bureaucratized institutional setting these functional attributes of historical African tenure systems.
Commodification and Conservation
In The Great Transformation Karl Polanyi famously declared that land and labor were fictitious commodities (Polanyi, 1944). By this he meant that land and labor were not items produced for the purpose of buying and selling. Labor was a monetarized version of our fellow human beings, land was commodified nature, and both humans and nature existed independent of market relations.
The significance of Polanyi's argument for an understanding of global capitalism may be debated, but its relevance to rangelands is unequivocal. In the community-secured property systems that once dominated pastoral Africa, group viability was paramount because it was the quasi-sovereign community that collectively defended a territory. Group membership through descent and shared political purpose, not purchase or written title, granted access to landed resources conceived of as a shared and inalienable patrimony (Behnke, 2018). State incorporation and market penetration have effectively challenged this concept of the value of land. Even without legal recognition, vernacular, land markets now flourish (Chimhowu and Woodhouse, 2006), and enclosure is increasingly an African “default mode of development” (Woodhouse, 2003). The commodification of rangeland is particularly problematic for pastoralists. With market penetration, rangelands that are naturally heterogeneous become commercially heterogeneous. As areas or resources acquire monetary value, it becomes profitable to excise the valuable bits that warrant “development,” leaving behind a fragmented, shrunken, residual rangeland environment for those productive activities such as pastoralism that are less susceptible to investment and intensification. Frequently, neither land use zoning nor the reservation of land by treaty have halted this process (Huntsinger and Hopkinson, 1996; Plieninger et al., 2012; Tyler et al., 2020).
In the modern world, at least one ideologically motivated form of land holding—the setting aside of protected areas for nature conservation—has the public recognition and political status to reliably enforce the old pastoral vision of land as a shared, inter-generational patrimony. If history provides any indication of the future, large-scale migratory pastoralism must make peace with nature conservation if it is to prosper8.
Collective Benefit
A communal ethic may have sustained indigenous pastoral tenure regimes, but it does not explain their emergence. Also relevant are mundane considerations of gain and loss that can be conveniently summarized under the heading of “collective benefit.”
The concept of collective benefit is exemplified by work conducted by Wade (1987) on community-level institutions in a semi-arid region of South India. Some of the villages in Wade's study area were “corporate” in that the villagers had created public institutions to collectively manage two of the productive resources upon which their livelihoods depended—irrigation water and livestock. Other villages in the same region made no such effort. Wade attributed these contrasting institutional outcomes to different incentive levels for joint action. In irrigated areas, corporate villages were located toward the tail-end of canal systems where water was scarce and farmers were strongly motivated to regulate its use, which was only possible if they acted together. At the head of canal systems where water was plentiful, individual farmers could act on their own to meet their water needs and collective institutions did not emerge. In rainfed farming areas, corporate villages were situated on rich, water-retentive soils that attracted large numbers of outside livestock that threatened the welfare of village farmers. In these villages, the control of stock numbers depended on a coordinated response, which local farmers undertook. Villages with poor soils attracted fewer livestock and took no such action. Wade concluded that the villagers in his study area were “likely to follow joint rules and arrangements only to achieve intensely felt needs that could not be met by individual action.” Collective action was contingent on what Wade called “collective benefits,” benefits that individuals acting alone could not obtain: “The opportunities for avoiding losses or boosting income by collective action will be taken only if the losses or gains are large” (Wade, 1987, p. 230).
Migratory African pastoralists have long engaged in collective action to secure important benefits that were unattainable for the isolated individual. Only group solidarity could provide individual security of tenure in unadministered, competitive, and potentially violent political environments and only collective territories were large enough to provide access to dispersed resources in variable natural environments. These collective benefits are now being undermined, politically by state incorporation and technologically by inputs that reduce the need for herd mobility, such as feed supplementation and the artificial provision of stock water. If collective action is to re-emerge, new sources of collective benefit are required (Behnke, 2008, 2018).
Partible Rights
That property is a bundle of rights and that different parties can own different parts of the bundle is not unusual. One party may own surface rights but not mineral rights, rights of possession but not disposal (as in legal trusts), temporary occupancy rights (as in time-sharing arrangements), or rights of access and transit over land that they do not otherwise control (Van de Laar, 1990):
More than one party can claim sole ownership interest in the same resource. One party may own the right to till the land, while another, perhaps the state, may own an easement to traverse or otherwise use the land for specific purposes. It is not the resource itself which is owed; it is a bundle, or a portion, of rights to use a resource that is owned (Alchian and Demsetz, 1973).
In migratory systems, mobile producers with a temporary interest in using an area routinely confront permanent occupants and other mobile producers. The land tenure systems that legitimate these encounters must represent the interests of both primary and peripheral rights holders with a multitude of different agendas but with important intersecting collective interests. For African pastoralists, the partibility of use rights has been an important mechanism for adjudicating the needs of these diverse categories of users. Different groups or individuals are entitled to use different resources in the same area, or to share in the use of a resource by using it for a limited period of time, by exploiting some aspects of its productivity but not others, or by using it only under certain circumstances. Migratory herders might, for example, have grazing rights in areas where they had limited rights to stock water; or be entitled to graze natural pastures with (or without) additional rights to graze the harvest residues on local farms, engage (or not) in cultivation themselves, or develop new water points, etc. Because of their diverse origins and interests, the participants in these systems of coordinated resource use frequently do not constitute homogeneous, readily identifiable corporate groups or occupy a clearly demarcated territory (Behnke, 1994; Turner, 1999; Fernandez-Gimenez, 2002).
For this, and a variety of other reasons, in pastoral Africa, indigenous systems of collective resource use deviated in fundamental ways from the classical model of the successful common property system (Wade, 1987; Ostrom, 1990). This conclusion has policy implications for efforts to encourage cooperative systems of pastoral resource use. In Africa, these efforts have often attempted—with mixed success—to foster the creation of socially and geographically bounded common property regimes. Indigenous pastoral systems never conformed to this model and their distinctive organization suggests that securing flexible access to extensive rangelands requires an alternative approach.
Conservation Easements and Grass Banks
Half a century ago international development agencies mistakenly promoted American ranching as a template for the reform of “backward” African and Asian pastoralism (Sandford, 1983; Behnke and Kerven, 1994). Decades later, some American ranchers might finally fulfill their potential as a positive role model. Faced with bureaucratic and legalistic restrictions of the kind that have overwhelmed many pastoral communities, these ranchers are developing cooperative forms of rangeland tenure with the potential to maintain some of the functional attributes of the open-range. To do this they have adopted new legal instruments that replicate the functionality of indigenous African tenures—an emphasis on collective benefit, exploitation of the partibility of property rights, and the recognition of wider social and conservation interests that restrict the free play of market forces.
With respect to collective benefit, some contemporary American ranchers have organized themselves to appropriate or defend a wide variety of “intensely felt needs that could not be met by individual action” (Wade, 1987, p. 230).
• In Texas, where most ranches are on private land, an increasingly profitable source of additional ranch income is commercial hunting. Game animals move around, however, showing little respect for property lines. This means that landowners who have invested in raising game animals cannot depend on harvesting them or selling the rights to harvest them, which encourages underinvestment in protecting wildlife and over-harvesting. In response, Texas ranchers and state officials have brought landowners together to form wildlife management associations that promote and regulate the hunting of a common pool resource—an itinerant game population (Huntsinger et al., 2014).
• From about 1900 until the 1980s the US Forest Service suppressed fires on the land it managed and “the regular fires that graziers used to keep land open for grazing were criminalized and halted” (Huntsinger et al., 2010, p. 23). When they were finally adopted, policies to encourage controlled burning did not bring relief. The intermingling of public and private lands and the involvement of multiple federal and state agencies each with its own procedures made coordinated action difficult; often little burning actually took place. Since conflicting regulations and bureaucratic inertia were the problem, more regulations and more bureaucracy were unlikely to be the solution. Coalitions that brought together government agencies, environmental groups, ranchers, and scientists have been more successful (Sayre, 2005).
• In the western USA, there are more than 5,000 migratory ranchers who move seasonally, usually between lowlands that they lease or own privately and higher altitude summer ranges that are publicly owned and managed by federal government agencies. Permits to graze public land are economically valuable, providing about 47% of income on ranches that rely principally on livestock production for their income (Huntsinger et al., 2010). These tenure rights have proved to be insecure because management agencies have imposed new restrictions on grazing and animal numbers. As a result, between 1980 and 2005, the amount of forage consumed on US Forest Service land declined by nearly 40 percent and the number of pastoralists declined by nearly 64 percent (Huntsinger et al., 2010). Restrictions to grazing rights on public land are an important reason why ranchers go out of business.
• Loss of rangeland to suburban sprawl, “ranchettes” and second homes is especially acute around metropolitan areas or touristic sites but is a threat to almost all ranching areas in the American West. The resulting parcelization complicates environmental management and is associated with habitat degradation and loss of biodiversity and ecosystem services (Havstad et al., 2009; Gutwein and Goldstein, 2013). Deteriorating conditions on the peri-urban edge and the increasing discrepancy between the income available from a ranch vs. the ranch's sale value for development also promote what has been characterized as an “impermanence syndrome” when ranchers conclude that selling for non-agricultural development is inevitable and stop investing in their operations (Berry and Plaut, 1978, cited in Liffmann et al., 2000, p. 363). These individual decisions often have wider social impacts:
“A single ranch-owner's decision may spell the fate of many thousands of acres. Landowner decisions affect more than their own property, as nearby properties are also influenced through the fragmentation of land use, weakening of the agricultural infrastructure, changing land values, and the creation of new growth nodes in previously undeveloped areas” (Johnson, 1998, cited in Liffmann et al., 2000 p. 363).
Because of the scale of ranch properties and the tightly integrated character of ranching communities, the loss of individual ranchers is not just an individual problem, but a collective one as well, which suggests that any solution may also need to be collective.
Of the collective challenges enumerated above, it is the last two—insecurity of tenure on public land and loss of private rangeland to alternative uses—that constitute the most geographically widespread threats to ranching in the American West. These two threats have also elicited what is arguably the most creative response. Part of this response has been an organized effort to dispel the hostility between conservationists and ranchers by making conservationists aware of the environmental benefits of grazing and, conversely, by convincing ranchers that conservation does not necessarily entail more regulations that interfere with their ability to run a business9. Also important has been a new legal device—the conservation easement—that has grown exponentially in popularity in the United States since the 1980s (Kay, 2016).
Conservation easements are voluntary legal agreements that recognize private ownership but limit the way private land can be used. Easements—like traditional pastoral tenure systems—rest on the notion that property is a bundle of rights that are divisible. In the case of conservation easements, property owners are typically paid by a government agency or non-profit land trust to relinquish the right to sell their land for subdivision or non-agricultural development. Private owners are then compensated for the reduced commercial value of their land while retaining the right to privately own, manage, sell or bequeath it (Liffmann et al., 2000). Conservation easements are attractive to ranchers who want to continue ranching but also want to realize some of the commercial development value of their ranch. The arrangement is attractive to conservation interests because easements are permanent, cheaper to acquire than outright land purchase, and are managed by their owners rather than hired employees (Brunson and Huntsinger, 2008).
Land owners who agree to conservation easements are rewarded financially through savings on their taxes and by payments that compensate for the opportunity cost of forgone development rights. In at least one case, the Malpai Borderlands Group, US ranchers have also traded their easements not for money but for grazing rights on the land of the non-profit organization that holds their easement (Sayre, 2005). The amount of grazing acquired through these “grass banking” agreements is based on the cost of leasing grazing land equivalent in value to the monetary compensation it has replaced. The Malpai easement contracts also contain clauses that void an easement agreement if ranchers lose access to state and federal grazing land through no fault of their own, something that ranchers insisted upon because the viability of their livestock operations depended on such access. Perhaps unexpectedly, the altered but conditional conservation status of private land has also had a beneficial impact on securing access to government land:
Malpai's easements have strengthened relations between ranchers and agencies, because the agencies recognize the benefit of preventing development of private lands to the conservation of adjacent public lands. In effect, the clause holds both the ranchers and the agencies to a higher standard of cooperation and effective management, as the former seek to maintain their leases and the latter seek to prevent the clause from being exercised (Huntsinger et al., 2014, citing Sayre, 2005; Rissman and Sayre, 2012).
In sum, conservationists and ranchers in the American West have pioneered the development for rangelands of a newly popular legal instrument—the conservation easement. These easements monetarize conservation values and recognize individual property rights and, hence, are compatible with market-based capitalism. But they also restrict the ability of individuals to alienate property in which a wider community has a permanent interest.
An additional feature of the Malpai programme—grass banking—provided an institutional framework for sharing privately owned rangeland. Grass banking involves the bartering of forage for conservation benefits. In Malpai these exchanges involved the trading of grazing privileges for conservation easements, but a wide range of other conservation activities can also be exchanged for forage, including the protection of endangered species, burning to reduce bush encroachment or control invasive plants, or grazing exclusion to rehabilitate degraded pastures. Enthusiasm for grass banking was based on a perceived win-win situation:
In theory, conservationists “win” because treatments, such as prescribed fire, that should improve overall health of an ecosystem, are implemented. Ranchers “win” because the grassbank provides forage to them, often at a discounted rate, so they don't suffer any economic harm as a result of the treatments which can require them to vacate their regular grazing pastures. Finally, local communities whom value “working landscapes” “win” because it is assumed that ranchers can remain in business while restoration treatments occur, thereby helping sustain the local economy and reduce the risk of subdivision (Gripne, 2005, p. 6).
By the early 2000s in the western US, more than twenty grass banking projects existed and more were being planned, but it is unclear how many new projects have been started since that time or how many of the original projects have survived up to the present. US tax laws required grass banks run by non-profit organizations to operate on a quid pro quo basis “where the economic value of the conservation benefit equals or exceeds the value of the forage' that was traded in return” (Gripne, 2005, p. 134). Many smaller grass banks did not have sufficient funding, personnel, or scale to meet this demand and hence were not economically sustainable, and a pilot effort by ranchers to form a collective grass bank to trade grazing rights among themselves failed to attract sufficient funding and collapsed (Gripne, 2005). Underlying these difficulties may have been a difference of opinion between conservationists and ranchers as to the purpose of grass banks. Conservations viewed grass banks as an innovative conservation tool; there are suggestions that ranchers saw them as a practical arrangement for managing drought and forage shortfalls on their home properties.
From an international perspective, the limited success of grass banking in the US may be less significant than its legalized status and formal organization. In both Australia and among newly privatized freeholders in Kenya, pastoralists have invented informal workarounds that allow them to share the grazing on their private holdings (McAllister et al., 2006; Mwangi, 2007; Lesorogol, 2010). Adjusted to meet local conditions, grass banking may provide a mechanism for strengthening these arrangements by legally recognizing shared grazing rights on private rangeland. Taken in combination, grass banks and conservation easements address what Fernandez-Gimenez (2002) has called the “paradox of pastoral tenure”—the simultaneous need for tenure security and for flexible access to extensive, erratically productive rangeland resources.
This essay began with the assertion that pre-industrial pastoralism had something to teach commercial ranchers. We close with the observation that there is a significant convergence between some of the features of traditional pastoral tenure systems, conservation easements, and grass banks. There is also evidence that commodification and individualization of property rights are taking place spontaneously in what were formerly open range areas in the developing world (Behnke, 2008; Bassett, 2009; Schareika et al., 2020), and that we may need to look for ways to accommodate this reality. It is, therefore, encouraging that some African conservationists are interested in developing “mobility-based livestock and wildlife management strategies” that exploit environmental heterogeneity and are based on institutional invocations that bear a strong resemblance to grass banks and conservation easements (Fynn et al., 2016, p. 390)10. American ranchers may at last have some development ideas that genuinely meet the needs of their Asian and African counterparts.
Conclusion
The organizers of this collection of papers asked the contributors to reimagine or redesign “monotonic pastoralism,” which they characterized as “pastoralism with the single objective of maximizing animal production and/or profit [that] has transformed landscapes, diminishing biodiversity, reducing water and air quality, accelerating loss of soil and plant biomass, and displacing indigenous animals and people” (Gregorini, 2019). This essay examined three central components of extensive livestock production—herd composition, grazing/pasture management, and rangeland tenure. In all of these areas, neither fenced nor open-range forms of extensive pastoralism are so dysfunctional as to constitute monotonic pastoralism, but they do face a number of shared problems. Set aside the presumption that either one of these systems of extensive production is technically or institutionally more advanced than the other, and it turns out that each has lessons for the other. Is it so farfetched to look back (with renewed respect) to the future?
Author Contributions
The author confirms being the sole contributor of this work and has approved it for publication.
Conflict of Interest
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
I thank Sarah Robinson who commented upon an earlier draft of this paper and Nathan Sayre who alerted me to published material on grass banks. Carol Kerven read, critiqued and edited every draft, and amendments suggested by the editor and peer reviewers substantially improved the paper.
Footnotes
1. ^A paper that focuses largely on intensive systems of livestock production, concludes that “In short, there is little or no scientific consensus on the sustainability of trajectories of various livestock production systems” (Tomich et al., 2011, p. 204).
2. ^Range science has a long history of encouraging these substitutions. Early North American range science promoted the systematic extermination of organisms that interfered with the maximization of livestock output and the minimalization of labour costs. Predator control achieved these objectives by allowing livestock to roam freely without supervision and without being eaten by anything other than humans. Fencing, it was argued, reinforced the benefits of predator control by eliminating herding and driving, which saved labour and also encouraged livestock to scatter more evenly, thereby diminishing their impact on pastures and increasing their productivity (Sayre, 2017, citing early twentieth century US government-sponsored range research).
3. ^As the term is used in this paper, a commodity is a good or service that can be readily exchanged in a market because it is broadly equivalent to other items of the same type.
4. ^Statistics Division, Food and Agriculture Organization of the United Nations Rome, Italy.
5. ^The environmental impact of the fences themselves is also an issue. While noting that the ecological effects are complex and varied, a recent systematic literature review concluded that “Scientists have begun to consider the ecological merits of conservation fence removal … and we recommend the expansion of fence removal programs to other fence types. Over time, the restoration of large tracts of fenceless land will benefit ecosystems and the services they deliver” (McInturff et al., 2020, p. 982).
6. ^“Although there are about 30000 species of teleost fish [the taxonomic category containing 96% of all fish species], only a small fraction of them are currently known to be migratory. However, these few species are the dominant marine species in terms of biomass and numbers, and most of the world's fish catches are based on them” (Bauer et al., 2011, p. 74).
7. ^The similarity between pastoral and wildlife migration is noted in a paper advocating pyric herbivory for the restoration of abandoned cropland in Kazakhstan (Brinkert et al., 2016) and by Fynn et al. (2019) in a consideration of the role of mobility in the exploitation of functional environmental heterogeneity by both wild and domesticated ungulates.
8. ^As long as commercial development can be held at bay, state ownership is another non-capitalistic, non-commodified form of land ownership. However, given the growing public awareness of environmental issues, continued pastoral access to public land may also rest on successfully addressing the concerns of conservationists, as discussed later in this paper with respect to state-owned land in the western United States.
9. ^Research documents the environmental benefits of ranching relative to alternative forms of land use in the American West. Maestas et al. (2003) examined the comparative levels of biodiversity on exurban developments, ranches, and nature reserves in a Colorado watershed. They concluded that “Reserves are often assumed to protect biodiversity, but our results suggest that reserves were somewhat ecologically degraded. Ranches can be more effective than reserves at maintaining native biotic communities in some instances, suggesting that the conversion of ranchland to exurban development has negative consequences” and that “efforts to protect the natural heritage of the Rocky Mountain West may require less reliance on nature reserves and greater focus on private lands” (Maestas et al., 2003, p. 1432–1433).
10. ^“[T]o enhance functional heterogeneity within PAs [protected areas] and to forestall the confinement of wildlife to ‘less functional isolated protected patches' with the growth of human populations, Fynn et al. advocate that”: [C]ommunities could be given grazing concessions within non-sensitive parts of PAs. Communities could benefit from grazing concessions within PAs by greater adaptive foraging options for livestock across larger landscapes (as do the wild herbivores), access to forage reserves during the dry season and greater ability to move livestock away from crop fields during the cropping season (Fynn et al., 2016, p. 390, 393).
References
Abdullahi, A., Mohammed, S., and Eid, A. (2013). “Town camels and milk villages: the growth of camel milk marketing in the Somali Region of Ethiopia,” in Pastoralism and Development in Africa, eds A. Catley, J. Lind, and I. Scoones (London: Routledge), 142–150. doi: 10.4324/9780203105979-20
Adler, P., Raff, D., and Lauenroth, W. (2001). The effect of grazing on the spatial heterogeneity of vegetation. Oecologia 128, 465–479. doi: 10.1007/s004420100737
Akweya, B. A., Gitao, C. G., and Okoth, M. W. (2012). The acceptability of camel milk and milk products from north eastern province in some urban areas of Kenya. Afr. J. Food Sci. 6, 465–473.
Alchian, A. A., and Demsetz, H. (1973). The property right paradigm. J. Econ. History 33, 16–27. doi: 10.1017/S0022050700076403
Allred, B. W., Scasta, J. D., Hovick, T. J., Fuhlendorf, S. D., and Hamilton, R. G. (2014). Spatial heterogeneity stabilizes livestock productivity in a changing climate. Agric. Ecosyst. Environ. 193, 37–41. doi: 10.1016/j.agee.2014.04.020
Andela, N., Liu, Y. Y., Van Dijk, A. I. J. M., De Jeu, R. A. M., and McVicar, T. R. (2013). Global changes in dryland vegetation dynamics (1988-2008) assessed by satellite remote sensing: comparing a new passive microwave vegetation density record with reflective greenness data. Biogeosciences 10, 6657–6676. doi: 10.5194/bg-10-6657-2013
Anderson, D. M., Elliott, H., Kochore, H. H., and Lochery, E. (2012). Camel herders, middlewomen, and urban milk bars: the commodification of camel milk in Kenya. J. Eastern Afr. Stud. 6, 383–404. doi: 10.1080/17531055.2012.696886
Animal Production and Research Unit (1979). Livestock and Range Research in Botswana. Ministry of Agriculture. Gaborone: Government Printer.
Augustine, D. J., McNaughton, S. J., and Frank, D. A. (2003). Feedbacks between soil nutrients and large herbivores in a managed savanna ecosystem. Ecol. Appl. 13, 1325–1337. doi: 10.1890/02-5283
Babaev, A. G., and Orlovsky, N. S. Natural conditions of deserts in the USSR. Adv. Desert Arid Land Technol. Dev. (1985) 4, 1–37.
Bailey, D. W., and Brown, J. R. (2011). Rotational grazing systems and livestock grazing behavior in shrub-dominated semi-arid and arid rangelands. Rangeland Ecol. Manage. 64, 1–9. doi: 10.2111/REM-D-09-00184.1
Banks, T. (2003). Property rights reform in rangeland China: dilemmas on the road to the household ranch. World Dev. 31, 2129–2142. doi: 10.1016/j.worlddev.2003.06.010
Barrett, J. C. (1992). The Economic Role of Cattle in Communal Farming Systems in Zimbabwe. Pastoral Development Network Paper 32c. London: Overseas Development Institute.
Barth, F. (1959). The land use pattern of migratory tribes of South Persia. Norsk Geografisk Tidsskrift 17, 1–11. doi: 10.1080/00291955908551758
Bassett, T. J. (1993). “Land use conflicts in pastoral development in northern Côte d'Ivoire,” in Land in African Agrarian Systems, eds T. J. Bassett and D. E. Crummey (Madison, WI: The University of Wisconsin Press), 131–154.
Bassett, T. J. (2009). Mobile pastoralism on the brink of land privatization in Northern Côte d'Ivoire. Geoforum 40, 756–766. doi: 10.1016/j.geoforum.2009.04.005
Bauer, S., Nolet, B. A., Giske, J., Chapman, J. W., Åkesson, S., Hedenström, A., et al. (2011). “Cues and decision rules in animal migration,” in Animal Migration: A Synthesis, eds E. J., Milner-Gulland, J. Fryxell, and A. R. E. Sinclair (Oxford: Oxford University Press), 68–87. doi: 10.1093/acprof:oso/9780199568994.003.0006
Begzsuren, S., Ellis, J. E., Ojima, D. S., Coughenour, M. B., and Chuluun, T. (2004). Livestock responses to droughts and severe winter weather in the Gobi Three Beauty National Park, Mongolia. J. Arid Environ. 59, 785–796. doi: 10.1016/j.jaridenv.2004.02.001
Behnke, R. (1994). Natural resource management in pastoral Africa. Dev. Policy Rev. 12, 5–28. doi: 10.1111/j.1467-7679.1994.tb00053.x
Behnke, R. (2008). “The drivers of fragmentation in arid and semi-arid landscapes,” in Fragmentation in Semi-Arid and Arid Landscapes. Consequences for Human and Natural Systems, eds K. A. Galvin, R. S. Reid, R. H. Jr. Behnke, and N. T. Hobbs (Dordrecht: Springer), 305–340. doi: 10.1007/978-1-4020-4906-4_13
Behnke, R. (2018). Open access and the sovereign commons: a political ecology of pastoral land tenure. Land Use Policy 76, 708–718. doi: 10.1016/j.landusepol.2018.02.054
Behnke, R., Fernandez-Gimenez, M. E., Turner, M. D., and Stammler, F. (2011). “Pastoral migration. Mobile systems of livestock husbandry,” in Animal Migration–A Synthesis, eds. E. J. Milner-Gulland, J. Fryxell, and A. R. E. Sinclair (Oxford: Oxford University Press), 144–252. doi: 10.1093/acprof:oso/9780199568994.003.0010
Behnke, R., and Kerven, C. (1994). “Redesigning for risk: tracking and buffering environmental variability in Africa's rangelands,” in Natural Resource Perspectives, Vol. 1 (London: Overseas Development Institute).
Behnke, R., Young, H., Sulieman, H. M., Robinson, S., and Idris, A. E. (2020). The seasonal imperative: environmental drivers of livestock mobility in East Darfur, Sudan. Land Use Policy 99:105014. doi: 10.1016/j.landusepol.2020.105014
Behnke, R.H., and Scoones, I. (1993). “Rethinking range ecology: implications for rangeland management in Africa,” in Range Ecology at Disequilibrium, eds R. Behnke, I. Scoones, and C. Kerven (London: Overseas Development Institute), 1–30.
Bell, R. H. (1971). A grazing ecosystem in the Serengeti. Sci. Am. 225, 86–93. doi: 10.1038/scientificamerican0771-86
Berry, D., and Plaut, T. (1978). Retaining agricultural activities under urban pressures: a review of land use conflicts and policies. Policy Sci. 9, 153–178. doi: 10.1007/BF00143740
Bollig, M. (2016). Adaptive cycles in the savannah: pastoral specialization and diversification in northern Kenya. J. Eastern Afr. Stud. 10, 21–44. doi: 10.1080/17531055.2016.1141568
Boone, R. B. (2005). Quantifying changes in vegetation in shrinking grazing areas in Africa. Conservat. Soc. 3:150.
Boone, R. B., and Hobbs, N. T. (2004). Lines around fragments: effects of fencing on large herbivores. Afr. J. Range For. Sci. 21, 147–158. doi: 10.2989/10220110409485847
Brinkert, A., Hölzel, N., Sidorova, T. V., and Kamp, J. (2016). Spontaneous steppe restoration on abandoned cropland in Kazakhstan: grazing affects successional pathways. Biodivers. Conservat. 25, 2543–2561. doi: 10.1007/s10531-015-1020-7
Briske, D. D., Ash, A. J., Derner, J. D., and Huntsinger, L. (2014). Commentary: a critical assessment of the policy endorsement for holistic management. Agric. Syst. 125, 50–53. doi: 10.1016/j.agsy.2013.12.001
Briske, D. D., Derner, J. D., Brown, J. R., Fuhlendorf, S. D., Teague, W. R., Havstad, K. M., et al. (2008). Rotational grazing on rangelands: reconciliation of perception and experimental evidence. Rangel. Ecol. Manage. 61, 3–17. doi: 10.2111/06-159R.1
Brown, J., and Kothmann, M. (2009). Rotational grazing and the profession and practice of rangeland management. Rangelands 31, 3–4. doi: 10.2111/1551-501X-31.5.3
Brunson, M. W., and Huntsinger, L. (2008). Ranching as a conservation strategy: can old ranchers save the new West? Rangel. Ecol. Manage. 61, 137–147. doi: 10.2111/07-063.1
Chimhowu, A., and Woodhouse, P. (2006). Customary vs private property rights? Dynamics and trajectories of vernacular land markets in Sub-Saharan Africa. J. Agr. Change 6, 346–371. doi: 10.1111/j.1471-0366.2006.00125.x
Cossins, N. J., and Upton, M. (1987). The Borana pastoral system of southern Ethiopia. Agric. Syst. 25, 199–218. doi: 10.1016/0308-521X(87)90020-5
Coughenour, M. B., Ellis, J. E., Swift, D. M., Coppock, D. L., Galvin, K., McCabe, J. T., et al. (1985). Energy extraction and use in a nomadic pastoral ecosystem. Science 230, 619–625. doi: 10.1126/science.230.4726.619
Crawford, C. L., Volenec, Z. M., Sisanya, M., Kibet, R., and Rubenstein, D. I. (2019). Behavioral and ecological implications of bunched, rotational cattle grazing in East African Savanna Ecosystem. Rangel. Ecol. Manage. 72, 204–209. doi: 10.1016/j.rama.2018.07.016
Cunnison, I. (1966). Baggara Arabs: Power and the Lineage in a Sudanese Nomad Tribe. Oxford: Clarendon Press.
Dahl, G. (1979a). Suffering Grass: Subsistence and Society of Waso Borana. Department of Social Anthropology, University of Stockholm.
Dahl, G. (1979b). “Ecology and equality: the Boran case,” in Pastoral Production and Society (Cambridge: Cambridge University Press), 261–281.
De Ridder, N. D., and Wagenaar, K. T. (1984). A comparison between the productivity of traditional livestock systems and ranching in eastern Botswana. ILCA Newsletter 3, 5–6.
Derner, J. D., Lauenroth, W. K., Stapp, P., and Augustine, D. J. (2009). Livestock as ecosystem engineers for grassland bird habitat in the western Great Plains of North America. Rangel. Ecol. Manage. 62, 111–118. doi: 10.2111/08-008.1
Donihue, C. M., Porensky, L. M., Foufopoulos, J., Riginos, C., and Pringle, R. M. (2013). Glade cascades: indirect legacy effects of pastoralism enhance the abundance and spatial structuring of arboreal fauna. Ecology 94, 827–837. doi: 10.1890/12-0856.1
Dwyer, M. J., and Istomin, K. V. (2008). Theories of nomadic movement: a new theoretical approach for understanding the movement decisions of Nenets and Komi reindeer herders. Hum. Ecol. 36:521. doi: 10.1007/s10745-008-9169-2
Dyson-Hudson, R., and Dyson-Hudson, N. (1969). Subsistence herding in Uganda. Sci. Am. 220, 76–89. doi: 10.1038/scientificamerican0269-76
Eldridge, D. J., Bowker, M. A., Maestre, F. T., Roger, E., Reynolds, J. F., and Whitford, W. G. (2011). Impacts of shrub encroachment on ecosystem structure and functioning: towards a global synthesis. Ecol. Lett. 14, 709–722. doi: 10.1111/j.1461-0248.2011.01630.x
Elhadi, Y. A., Nyariki, D. M., and Wasonga, O. V. (2015). Role of camel milk in pastoral livelihoods in Kenya: contribution to household diet and income. Pastoralism 5:8. doi: 10.1186/s13570-015-0028-7
Ellis, J. E., and Swift, D. M. (1988). Stability of African pastoral ecosystems: alternate paradigms and implications for development. J. Range Manage. 41, 450–459. doi: 10.2307/3899515
Engler, J. O., and von Wehrden, H. (2018). Global assessment of the non-equilibrium theory of rangelands: revisited and refined. Land Use Policy 70, 479–484. doi: 10.1016/j.landusepol.2017.11.026
Epp, H., and Dyck, I. (2002). Early human-bison population interdependence in the plains ecosystem. Great Plains Res. J. Natural Soc. Sci. 615, 323–337.
Erlandson, J. M., and Braje, T. J. (2013). Archeology and the Anthropocene. Anthropocene 4, 1–7. doi: 10.1016/j.ancene.2014.05.003
Estell, R. E., Havstad, K. M., Cibils, A. F., Fredrickson, E. L., Anderson, D. M., Schrader, T. S., et al. (2012). Increasing shrub use by livestock in a world with less grass. Rangel. Ecol. & Manage. 65, 553–562. doi: 10.2111/REM-D-11-00124.1
Faye, B. (2020). How many large camelids in the world? A synthetic analysis of the world camel demographic changes. Pastoralism 10:25. doi: 10.1186/s13570-020-00176-z
Fernandez-Gimenez, M. E. (2002). Spatial and social boundaries and the paradox of pastoral land tenure: a case study from postsocialist Mongolia. Hum. Ecol. 30, 49–78. doi: 10.1023/A:1014562913014
Foley, S. F., Gronenborn, D., Andreae, M. O., Kadereit, J. W., Esper, J., Scholz, D., et al. (2013). The Palaeoanthropocene–the beginnings of anthropogenic environmental change. Anthropocene 3, 83–88. doi: 10.1016/j.ancene.2013.11.002
Fuhlendorf, S. D., and Engle, D. M. (2004). Application of the fire–grazing interaction to restore a shifting mosaic on tallgrass prairie. J. Appl. Ecol. 41, 604–614. doi: 10.1111/j.0021-8901.2004.00937.x
Fuhlendorf, S. D., Engle, D. M., Elmore, R. D., Limb, R. F., and Bidwell, T. G. (2012). Conservation of pattern and process: developing an alternative paradigm of rangeland management. Rangel. Ecol. Manage. 65, 579–589. doi: 10.2111/REM-D-11-00109.1
Fuhlendorf, S. D., Fynn, R. W., McGranahan, D. A., and Twidwell, D. (2017). “Heterogeneity as the basis for rangeland management,” in Rangeland Systems, ed D. D. Briske (Cham: Springer), 169–196. doi: 10.1007/978-3-319-46709-2_5
Fuhlendorf, S. D., Harrell, W. C., Engle, D. M., Hamilton, R. G., Davis, C. A., and Leslie, D. M. Jr. (2006). Should heterogeneity be the basis for conservation? Grassland bird response to fire and grazing. Ecol. Appl. 16, 1706–1716. doi: 10.1890/1051-0761(2006)016(1706:SHBTBF)2.0.CO
Fuhlendorf, S. D., Townsend, D. E., Elmore, R. D., and Engle, D. M. (2010). Pyric-herbivory to promote rangeland heterogeneity: evidence from small mammal communities. Rangel. Ecol. Manage. 63, 670–678. doi: 10.2111/REM-D-10-00044.1
Fynn, R. W. (2012). Functional resource heterogeneity increases livestock and rangeland productivity. Rangel. Ecol. Manage. 65, 319–329. doi: 10.2111/REM-D-11-00141.1
Fynn, R. W., Augustine, D. J., and Fuhlendorf, S. D. (2019). “Managing browsing and grazing ungulates,” in The Ecology of Browsing and Grazing II, eds I. J Gordon and H. H. T. Prins (Cham: Springer), 321–338. doi: 10.1007/978-3-030-25865-8_14
Fynn, R. W., Augustine, D. J., Peel, M. J., and de Garine-Wichatitsky, M. (2016). Strategic management of livestock to improve biodiversity conservation in African savannahs: a conceptual basis for wildlife-livestock coexistence. J. Appl. Ecol. 53, 388–397. doi: 10.1111/1365-2664.12591
Galaty, J. G. (1982). Being “Maasai”; being “people-of-cattle”: ethnic shifters in East Africa. Am. Ethnol. 9, 1–20. doi: 10.1525/ae.1982.9.1.02a00010
Gebremichael, B., and Girmay, S. (2019). Camel milk production and marketing: pastoral areas of Afar, Ethiopia. Pastoralism 9:16. doi: 10.1186/s13570-019-0147-7
Grant, R. C., Botha, J., Grant, T. C., Peel, M. J., and Smit, I. P. (2019). When less is more: heterogeneity in grass patch height supports herbivores in counter-intuitive ways. Afr. J. Range For. Sci. 36, 1–8. doi: 10.2989/10220119.2018.1543209
Gregorini, P. (2019). Grazing in Future Multi-Scapes: From Thoughtscapes to Landscapes, Creating Health from the Ground Up. Proposal for an international workshop, Lincoln University. Available online at: https://web.cvent.com/event/3bcbdfc4-ff78-4f6d-804d-7bdedfeb8c8d/summary (accessed March 2021).
Gripne, S. L. (2005). Grassbanks: An evaluation of a conservation tool (Graduate Student theses, dissertations, & professional papers). University of Montana, 9585. Available online at: https://scholarworks.umt.edu/etd/9585 (accessed March 2021).
Gutwein, M., and Goldstein, J. H. (2013). Integrating conservation and financial objectives on private rangelands in northern Colorado: rancher and practitioner perceptions. Rangel. Ecol. Manage. 66, 330–338. doi: 10.2111/REM-D-11-00206.1
Gwynne, M. D., and Bell, R. H. V. (1968). Selection of vegetation components by grazing ungulates in the Serengeti National Park. Nature 220, 390–393. doi: 10.1038/220390a0
Hammer, E. (2014). Local landscape organization of mobile pastoralists in southeastern Turkey. J. Anthropol. Archaeol. 35, 269–288. doi: 10.1016/j.jaa.2014.06.001
Havstad, K., Peters, D., Allen-Diaz, B., Bartolome, J., Bestelmeyer, B., Briske, D., et al. (2009). “The western United States rangelands: a major resource,” in Grassland: Quietness and Strength for a New American Agriculture, Vol. 142, eds W. Wedin and S. L. Fales (Madison, Wisconsin, WI: ASA-CSSA-SSSA), 75–93. doi: 10.2134/2009.grassland.c5
Hawkins, H. J. (2017). A global assessment of Holistic Planned Grazing™ compared with season-long, continuous grazing: meta-analysis findings. Afr. J. Range For. Sci. 34, 65–75. doi: 10.2989/10220119.2017.1358213
Heady, H. F. (1961). Continuous vs. specialized grazing systems: a review and application to the California annual type. J. Range Manage. 14, 182–193. doi: 10.2307/3895147
Herren, U. J. (1990). “The commercial sale of camel milk from pastoral herds in the Mogadishu hinterland, Somalia,” in Pastoral Development Network (London: Overseas Development Institute).
Hoben, A. (1988). “The political economy of land tenure in Somalia,” in Land and Society in Contemporary Africa, eds R. E. Downs, and S. P. Reyna (Hanover: University Press of New England), 192–220.
Holdo, R. M., Holt, R. D., Sinclair, A. R., Godley, B. J., and Thirgood, S. (2011). “Migration impacts on communities and ecosystems: empirical evidence and theoretical insights,” in Animal Migration: A Synthesis, eds E. J. Milner-Gulland, J. M. Fryxell, and A. R. Sinclair (Oxford University Press), 131–143. doi: 10.1093/acprof:oso/9780199568994.003.0009
Holechek, J. L., Pieper, R. D., and Herbel, C. H. (2001). Range Management. Principles and Practices. New Jersey, NJ: Prentice-Hall.
Honeychurch, W., and Makarewicz, C. A. (2016). The archaeology of pastoral nomadism. Ann. Rev. Anthropol. 45, 341–359. doi: 10.1146/annurev-anthro-102215-095827
Huho, J. M., Ngaira, J. K., and Ogindo, H. O. (2011). Living with drought: the case of the Maasai pastoralists of northern Kenya. Educa. Res. 2, 779–789.
Huntsinger, L. (2016). Enabling sustainable pastoral landscapes: building social capital to restore natural capital. Options Méditerranéennes 116, 315–325.
Huntsinger, L., Forero, L. C., and Sulak, A. (2010). Transhumance and pastoralist resilience in the western United States. Pastoralism 1, 1–15.
Huntsinger, L., and Hopkinson, P. (1996). Sustaining rangeland landscapes: a social and ecological process. J. Range Manage. 49, 167–173. doi: 10.2307/4002689
Huntsinger, L., Sayre, N. F., and Macaulay, L. (2014). “Ranchers, land tenure, and grass-roots governance: maintaining pastoralist use of rangelands in the US in three different settings,” in The Governance of Rangelands: Collective Action for Sustainable Pastoralism, eds P. M. Herrera, J. Davies, and P. M. Baena (London: Routledge), 62–93.
Illius, A.W., and O'Connor, T.G. (1999). On the relevance of nonequilibrium concepts to arid and semiarid grazing systems. Ecol. Appl. 9, 798–813. doi: 10.1890/1051-0761(1999)009(0798:OTRONC)2.0.CO
Istomin, K. V., and Dwyer, M. J. (2010). Dynamic mutual adaptation: human-animal interaction in reindeer herding pastoralism. Hum. Ecol. 38, 613–623. doi: 10.1007/s10745-010-9341-3
Jacobs, A. H. (1975). “Maasai pastoralism in historical perspective,” in Pastoralism in Tropical Africa, ed. T. Monod (Oxford: Oxford University Press), 406–425. doi: 10.4324/9780429490781-16
Jahnke, H. E. (1982). Livestock Production Systems and Livestock Development in Tropical Africa, Vol. 35. Kiel: Kieler Wissenschaftsverlag Vauk.
Johnson, S. G. (1998). Oaks at the Edge: Land Use Change in the Woodlands of Central Sierra Nevada, California. Berkeley, CA: University of California.
Kagunyu, A. W., and Wanjohi, J. (2014). Camel rearing replacing cattle production among the Borana community in Isiolo County of Northern Kenya, as climate variability bites. Pastoralism 4, 1–5. doi: 10.1186/s13570-014-0013-6
Kay, K. (2016). Breaking the bundle of rights: conservation easements and the legal geographies of individuating nature. Environ. Plan. A Econ. Space 48, 504–522. doi: 10.1177/0308518X15609318
Kerven, C. (2004). “The influence of cold temperatures and snowstorms on rangelands and livestock in northern Asia,” in Rangelands at Equilibrium and Non-equilibrium, ed S. Vetter. (Cape Town: Programme for Land and Agrarian Studies (PLAAS) School of Government, University of the Western Cape), 41–55.
Kerven, C., Robinson, S., and Behnke, R. (2020). Pastoralism at scale on the Kazakh rangelands: from clans to workers to ranchers. Food Syst. 4:590401. doi: 10.3389/fsufs.2020.590401
Khazanov, A. M. (1994). Nomads and the Outside World, 2nd Edn. Cambridge: Cambridge University Press.
Krätli, S. (2008). Cattle breeding, complexity and mobility in a structurally unpredictable environment: the WoDaaBe herders of Niger. Nomadic Peoples 12, 11–41. doi: 10.3167/np.2008.120102
Krätli, S., and Schareika, N. (2010). Living off uncertainty: the intelligent animal production of dryland pastoralists. Eur. J. Dev. Res. 22, 605–622. doi: 10.1057/ejdr.2010.41
Lesorogol, C. K. (2010). “Creating common grazing rights on private parcels: how new rules produce incentives for cooperative land management,” in Cooperation in Economy and Society, ed R. Marshall (Lanham, MD: Altamira Press), 239–258.
Liao, C., Agrawal, A., Clark, P. E., Levin, S. A., and Rubenstein, D. I. (2020). Landscape sustainability science in the drylands: mobility, rangelands and livelihoods. Landscape Ecol. 1–15. doi: 10.1007/s10980-020-01068-8
Liffmann, R. H., Huntsinger, L., and Forero, L. C. (2000). To ranch or not to ranch: home on the urban range? J. Range Manage. 53, 362–370. doi: 10.2307/4003745
Limb, R. F., Fuhlendorf, S. D., Engle, D. M., Weir, J. R., Elmore, R. D., and Bidwell, T. G. (2011). Pyric–herbivory and cattle performance in grassland ecosystems. Rangel. Ecol. Manage. 64, 659–663. doi: 10.2111/REM-D-10-00192.1
Lind, J., Sabates-Wheeler, R., Caravani, M., Kuol, L., and Nightingale, D. (2020). Newly evolving pastoral and post-pastoral rangelands of Eastern Africa. Pastoralism 10:24. doi: 10.1186/s13570-020-00179-w
López-Carr, D., Pricope, N. G., Aukema, J. E., Jankowska, M. M., Funk, C., Husak, G., et al. (2014). A spatial analysis of population dynamics and climate change in Africa: potential vulnerability hot spots emerge where precipitation declines and demographic pressures coincide. Popul. Environ. 35, 323–339. doi: 10.1007/s11111-014-0209-0
Maestas, J.D., Knight, R.L., and Gilgert, W.C. (2003). Biodiversity across a rural land-use gradient. Conservat. Biol. 17, 1425–1434. doi: 10.1046/j.1523-1739.2003.02371.x
Maestre, F. T., Salguero-Gomez, R., and Quero, J. L. (2012). It is getting hotter in here: determining and projecting the impacts of global environmental change on drylands. Phil. Trans. R. Soc. B 367, 3062–3075. doi: 10.1098/rstb.2011.0323
Marshall, F., Reid, R. E., Goldstein, S., Storozum, M., Wreschnig, A., Hu, L., et al. (2018). Ancient herders enriched and restructured African grasslands. Nature 561, 387–390. doi: 10.1038/s41586-018-0456-9
McAllister, R. R., Gordon, I. J., Janssen, M. A., and Abel, N. (2006). Pastoralists' responses to variation of rangeland resources in time and space. Ecol. Appl. 16, 572–583. doi: 10.1890/1051-0761(2006)016(0572:PRTVOR)2.0.CO
McGranahan, D. A., Hovick, T. J., Dwayne Elmore, R., Engle, D. M., Fuhlendorf, S. D., Winter, S. L., et al. (2016). Temporal variability in aboveground plant biomass decreases as spatial variability increases. Ecology 97, 555–560. doi: 10.1890/15-0906
McInturff, A., Xu, W., Wilkinson, C. E., Dejid, N., and Brashares, J.S. (2020). Fence ecology: frameworks for understanding the ecological effects of fences. BioScience 70, 971–985. doi: 10.1093/biosci/biaa103
McNaughton, S. J. (1979). Grazing as an optimization process: grass-ungulate relationships in the Serengeti. Am. Natural. 113, 691–703. doi: 10.1086/283426
McNaughton, S. J. (1983). Serengeti grassland ecology: the role of composite environmental factors and contingency in community organization. Ecol. Monogr. 53, 291–320. doi: 10.2307/1942533
McNaughton, S. J. (1984). Grazing lawns: animals in herds, plant form, and coevolution. Am. Natural. 124, 863–886. doi: 10.1086/284321
Megersa, B., Markemann, A., Angassa, A., Ogutu, J. O., Piepho, H. P., and Zárate, A. V. (2014). Livestock diversification: an adaptive strategy to climate and rangeland ecosystem changes in southern Ethiopia. Hum. Ecol. 42, 509–520. doi: 10.1007/s10745-014-9668-2
Megersa, B., Markemann, A., Angassa, A., and Zárate, A. V. (2013). The role of livestock diversification in ensuring household food security under a changing climate in Borana, Ethiopia. Food Security 6, 15–28. doi: 10.1007/s12571-013-0314-4
Miehe, G., Miehe, S., Kaiser, K., Reudenbach, C., Behrendes, L., Duo, L., et al. (2009). How old is pastoralism in Tibet? An ecological approach to the making of a Tibetan landscape. Palaeogeogr. Palaeoclimatol. Palaeoecol. 276, 130–147. doi: 10.1016/j.palaeo.2009.03.005
Mirzabaev, A., Ahmed, M., Werner, J., Pender, J., and Louhaichi, M. (2016). Rangelands of Central Asia: challenges and opportunities. J. Arid Land 8, 93–108. doi: 10.1007/s40333-015-0057-5
Muchiru, A. N., Western, D., and Reid, R.S. (2009). The impact of abandoned pastoral settlements on plant and nutrient succession in an African savanna ecosystem. J. Arid Environ. 73, 322–331. doi: 10.1016/j.jaridenv.2008.09.018
Muchiru, A. N., Western, D. J., and Reid, R. S. (2008). The role of abandoned pastoral settlements in the dynamics of African large herbivore communities. J. Arid Environ. 72, 940–952. doi: 10.1016/j.jaridenv.2007.11.012
Mwangi, E. (2007). Subdividing the commons: Distributional conflict in the transition from collective to individual property rights in Kenya's Maasailand. World Dev. 35, 815–834. doi: 10.1016/j.worlddev.2006.09.012
Noor, I. M., Guliye, A. Y., Tariq, M., and Bebe, B. O. (2013). Assessment of camel and camel milk marketing practices in an emerging peri-urban production system in Isiolo County, Kenya. Pastoralism 3:28. doi: 10.1186/2041-7136-3-28
Ocaido, M., Muwazi, R. T., and Opuda-Asibo, J. (2009). Financial analysis of livestock production systems around Lake Mburo National Park, in South Western Uganda. Livestock Res. Rural Dev. 21:70. doi: 10.1007/s11250-009-9315-x
O'Connor, T. G., Puttick, J. R., and Hoffman, M. T. (2014). Bush encroachment in southern Africa: changes and causes. Afr. J. Range For. Sci. 31, 67–88. doi: 10.2989/10220119.2014.939996
O'Reagain, P. J., and Turner, J. R. (1992). An evaluation of the empirical basis for grazing management recommendations for rangeland in southern Africa. J. Grassland Soc. Southern Afr. 9, 38–49. doi: 10.1080/02566702.1992.9648297
Österle, M. (2008). From cattle to goats: the transformation of East Pokot pastoralism in Kenya. Nomadic Peoples 12, 81–91. doi: 10.3167/np.2008.120105
Ostrom, E. (1990). Governing the Commons: The Evolution of Institutions for Collective Action. Cambridge, UK: Cambridge University Press.
Otte, M. J., and Chilonda, P. (2002). Cattle and Small Ruminant Production Systems in sub-Saharan Africa: A Systematic Review. Rome: FAO.
Owen-Smith, N., and Novellie, P. (1982). What should a clever ungulate eat? Am. Natural. 119, 151–178. doi: 10.1086/283902
Paine, R. (1994). Herds of the Tundra: A Portrait of Saami Reindeer Pastoralism. Washington, DC: Smithsonian Institute Press.
Penning de Vries, F. W. T., and Djiteye, M. A. (1982). The Productivity of Sahelian Rangeland: a Study of Soils. Wageningen: Vegetation and the Exploitation of This Natural Resource, Centre for Agricultural Publishing and Documentation.
Plieninger, T., Ferranto, S., Huntsinger, L., Kelly, M., and Getz, C. (2012). Appreciation, use, and management of biodiversity and ecosystem services in California's working landscapes. Environ. Manage. 50, 427–440. doi: 10.1007/s00267-012-9900-z
Porensky, L. M., Bucher, S. F., Veblen, K. E., Treydte, A. C., and Young, T. P. (2013a). Megaherbivores and cattle alter edge effects around ecosystem hotspots in an African savanna. J. Arid Environ. 96, 55–63. doi: 10.1016/j.jaridenv.2013.04.003
Porensky, L. M., and Veblen, K. E. (2012). Grasses and browsers reinforce landscape heterogeneity by excluding trees from ecosystem hotspots. Oecologia 168, 749–759. doi: 10.1007/s00442-011-2123-9
Porensky, L. M., Wittman, S. E., Riginos, C., and Young, T. P. (2013b). Herbivory and drought interact to enhance spatial patterning and diversity in a savanna understory. Oecologia 173, 591–602. doi: 10.1007/s00442-013-2637-4
Pricope, N. G., Husak, G., Lopez-Carr, D., Funk, C., and Michaelsen, J. (2013). The climate-population nexus in the East African Horn: emerging degradation trends in rangeland and pastoral livelihood zones. Global Environ. Change 23, 1525–1541. doi: 10.1016/j.gloenvcha.2013.10.002
Reid, R.S., and Ellis, J.E. (1995). Impacts of pastoralists on woodlands in South Turkana, Kenya: livestock-mediated tree recruitment. Ecol. Appl. 5, 978–992. doi: 10.2307/2269349
Rennie, T., Light, D., Rutherford, A., Miller, M., Fisher, I., Pratchett, D., et al. (1977). Beef cattle productivity under traditional and improved management in Botswana. Trop. Anim. Health Product. 9, 1–6. doi: 10.1007/BF02297380
Rissman, A., and Sayre, N. F. (2012). Conservation outcomes and social relations: a comparative study of private ranchland conservation easements. Soc. Natural Resour. 25, 523–538. doi: 10.1080/08941920.2011.580419
Roman, J., and Palumbi, S. R. (2003). Whales before whaling in the North Atlantic. Science 301, 508–510. doi: 10.1126/science.1084524
Rosenberg, A. A., Bolster, W. J., Alexander, K. E., Leavenworth, W. B., Cooper, A. B., and McKenzie, M. G. (2005). The history of ocean resources: modeling cod biomass using historical records. Front. Ecol. Environ. 3, 78–84. doi: 10.1890/1540-9295(2005)003(0078:THOORM)2.0.CO
Sandford, S. (1981). Review of World Bank Livestock Activities in Dry Tropical Africa. Unpublished Consultancy Report. Washington, DC: World Bank.
Sayre, N. F. (2005). Working wilderness: the Malpai Borderlands Group and the Future of the Western Range. Tucson: Rio Nuevo Publishers.
Sayre, N. F. (2017). The Politics of Scale: A History of Rangeland Science. Chicago: University of Chicago Press.
Schareika, N. (2001). Environmental knowledge and pastoral migration among the Wodaabe of south-eastern Niger. Nomadic Peoples 5, 65–88. doi: 10.3167/082279401782310934
Schareika, N., Brown, C., and Moritz, M. (2020). Critical transitions from pastoralism to ranching in central Africa. Curr. Anthropol. 62. doi: 10.1086/713248
Schlütz, F., and Lehmkuhl, F. (2009). Holocene climatic change and the nomadic Anthropocene in Eastern Tibet: palynological and geomorphological results from the Nianbaoyeze Mountains. Q. Sci. Rev. 28, 1449–1471. doi: 10.1016/j.quascirev.2009.01.009
Schorger, A. W. (1955). The Passenger Pigeon: Its Natural History and Extinction. Norman: University of Oklahoma Press; Blackburn Press.
Schwinning, S., and Sala, O. E. (2004). Hierarchy of responses to resource pulses in arid and semi-arid ecosystems. Oecologia 141, 211–220. doi: 10.1007/s00442-004-1520-8
Scoones, I. (1995). Exploiting heterogeneity: habitat use by cattle in dryland Zimbabwe. J. Arid Environ. 29, 221–237. doi: 10.1016/S0140-1963(05)80092-8
Sneath, D. (2003). Land use, the environment and development in post-socialist Mongolia. Oxford Dev. Stud. 31, 441–459. doi: 10.1080/1360081032000146627
Spengler, R. N. (2014). Niche dwelling vs. niche construction: landscape modification in the bronze and iron ages of central Asia. Hum. Ecol. 42, 813–821. doi: 10.1007/s10745-014-9697-x
Stammler, F. (2005). “Reindeer nomads meet the market: culture,” in Property and Globalisation at the End of the Land, Vol. 6 Halle Studies in the Anthropology of Eurasia (Muenster: Lit Publishers).
Stépanoff, C. (2012). Human-animal “joint commitment” in a reindeer herding system. Hau J. Ethnogr. Theor. 2, 287–312. doi: 10.14318/hau2.2.015
Sternberg, T. (2012). Piospheres and pastoralists: vegetation and degradation in steppe grasslands. Hum. Ecol. 40, 811–820. doi: 10.1007/s10745-012-9539-7
Stevens, N., Lehmann, C. E. R., Murphy, B. P., and Durigan, G. (2017). Savanna woody encroachment is widespread across three continents. Global Change Biol. 23, 235–244. doi: 10.1111/gcb.13409
Stoddart, L. A., Smith, A. D., and Box, T. W. (1975). Range Management. New York, NY: McGraw-Hill Co.
Strickon, A. (1965). “The Euro-American ranching complex,” in Man, Culture, and Animals. The Role of Animals in Human Ecological Adjustments, eds A. Leeds and A. P. Vayda (Washington, DC: Publication No. 78 of the American Association for the Advancement of Science), 229–258.
Strydom, P. E. (2008). Do indigenous Southern African cattle breeds have the right genetics for commercial production of quality meat? Meat Sci. 80, 86–93. doi: 10.1016/j.meatsci.2008.04.017
Strydom, P. E., Frylinck, L., Van der Westhuizen, J., and Burrow, H. M. (2008). Growth performance, feed efficiency and carcass and meat quality of tropically adapted breed types from different farming systems in South Africa. Austral. J. Exp. Agric. 48, 599–607. doi: 10.1071/EA06057
Teague, R., Provenza, F., Kreuter, U., Steffens, T., and Barnes, M. (2013). Multi-paddock grazing on rangelands: why the perceptual dichotomy between research results and rancher experience? J. Environ. Manage. 128, 699–717. doi: 10.1016/j.jenvman.2013.05.064
Teague, R., Provenza, F., Norton, B., Steffens, T., Barnes, M., Kothmann, M., et al. (2008). “Benefits of multi-paddock grazing management on rangelands: limitations of experimental grazing research and knowledge gaps,” in Grasslands: Ecology, Management and Restoration, ed G. H. Schroder (New York, NY: Nova Science Publishers), 41–80.
Tomich, T. P., Brodt, S., Ferris, H., Galt, R., Horwath, W. R., Kebreab, E., et al. (2011). Agroecology: a review from a global-change perspective. Ann. Rev. Environ. Resour. 36, 193–222. doi: 10.1146/annurev-environ-012110-121302
Turner, M. D. (1998a). Long-term effects of daily grazing orbits on nutrient availability in Sahelian West Africa: I. Gradients in the chemical composition of rangeland soils and vegetation. J. Biogeogr. 25, 669–682. doi: 10.1046/j.1365-2699.1998.2540669.x
Turner, M. D. (1998b). Long-term effects of daily grazing orbits on nutrient availability in Sahelian West Africa: 2. Effects of a phosphorus gradient on spatial patterns of annual grassland production. J. Biogeogr. 25, 683–694. doi: 10.1046/j.1365-2699.1998.2540683.x
Turner, M. D. (1999). “The role of social networks, indefinite boundaries and political bargaining in maintaining the ecological and economic resilience of the transhumance systems of Sudano-Sahelian West Africa,” in Managing Mobility in African Rangelands: The Legitimization of Transhumance, ed M. Niamir-Fuller (London: Intermediate Technology Publications), 97–123. doi: 10.3362/9781780442761.005
Turton, D. (1994). “Mursi political identity and warfare: the survival of an idea,” in Ethnicity and Conflict in the Horn of Africa, eds K. Fukui and J. Markakis (London: James Currey), 15–32.
Tyler, N., Hanssen-Bauer, I., Førland, E., and Nellemann, C. (2020). The shrinking resource base of pastoralism: Saami reindeer herding in a climate of change. Front. Sustain. Food Syst. 4:585685. doi: 10.3389/fsufs.2020.585685
Van Auken, O. W. (2000). Shrub invasions of North American semiarid grasslands. Ann. Rev. Ecol. System. 31, 197–215. doi: 10.1146/annurev.ecolsys.31.1.197
Van Auken, O. W. (2009). Causes and consequences of woody plant encroachment into western North American grasslands. J. Environ. Manage. 90, 2931–2942. doi: 10.1016/j.jenvman.2009.04.023
Van de Laar, A. (1990). Framework for the analysis of common pool natural resources. ISS Working Paper Ser. General Ser. 77, 1–62.
van der Waal, C., Kool, A., Meijer, S. S., Kohi, E., Heitkönig, I. M., de Boer, W. F., et al. (2011). Large herbivores may alter vegetation structure of semi-arid savannas through soil nutrient mediation. Oecologia 165, 1095–1107. doi: 10.1007/s00442-010-1899-3
Vehrs, H. P. (2016). Changes in landscape vegetation, forage plant composition and herding structure in the pastoralist livelihoods of East Pokot, Kenya. J. Eastern Afr. Stud. 10, 88–110. doi: 10.1080/17531055.2015.1134401
Venter, Z. S., Hawkins, H. J., and Cramer, M. D. (2017). Implications of historical interactions between herbivory and fire for rangeland management in African savannas. Ecosphere 8:e01946. doi: 10.1002/ecs2.1946
Ventresca Miller, A. R., Spengler, R., Haruda, A., Miller, B., Wilkin, S., Robinson, S., et al. (2020). Ecosystem engineering among ancient pastoralists in northern Central Asia. Front. Earth Sci. 8:168. doi: 10.3389/feart.2020.00168
Volpato, G., and King, E. G. (2019). From cattle to camels: trajectories of livelihood adaptation and social-ecological resilience in a Kenyan pastoralist community. Regional Environ. Change 19, 849–865. doi: 10.1007/s10113-018-1438-z
von Wehrden, H., Hanspach, J., Kaczensky, P., Fischer, J., and Wesche, K. (2012). Global assessment of the non-equilibrium concept in rangelands. Ecol. Appl. 22, 393–399. doi: 10.1890/11-0802.1
Wade, R. (1987). The management of common property resources: finding a cooperative solution. World Bank Res. Observer 2, 219–234. doi: 10.1093/wbro/2.2.219
Wako, G., Tadesse, M., and Angassa, A. (2017). Camel management as an adaptive strategy to climate change by pastoralists in southern Ethiopia. Ecol. Proc. 6, 1–12. doi: 10.1186/s13717-017-0093-5
Walker, B. (2019). “Guiding rangelands into an uncertain future,” in Presentation to the Australian Rangelands Conference (Canberra, ACT: Australian Rangelands Society). doi: 10.1071/RJ20051
Watson, E. E., Kochore, H. H., and Dabasso, B. H. (2016). Camels and climate resilience: adaptation in northern Kenya. Hum. Ecol. 44, 701–713. doi: 10.1007/s10745-016-9858-1
Western, D. (1982). The environment and ecology of pastoralists in arid savannas. Dev. Change 13, 183–211. doi: 10.1111/j.1467-7660.1982.tb00117.x
Woodhouse, P. (2003). African enclosures: a default mode of development. World Dev. 31, 1705–1720. doi: 10.1016/S0305-750X(03)00140-2
Yagodin, V. N., and Amirov, S. S. (2014). Ancient hunting constructions on the Ustyurt Plateau. Saiga News 18, 18–20.
Keywords: pastoralism, ranching, herd composition, migration, rangeland tenure
Citation: Behnke RH (2021) Grazing Into the Anthropocene or Back to the Future? Front. Sustain. Food Syst. 5:638806. doi: 10.3389/fsufs.2021.638806
Received: 07 December 2020; Accepted: 22 March 2021;
Published: 30 April 2021.
Edited by:
Iain James Gordon, Australian National University, AustraliaReviewed by:
Dan Rubenstein, Princeton University, United StatesLynn Huntsinger, University of California, Berkeley, United States
Randall Boone, Colorado State University, United States
Copyright © 2021 Behnke. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Roy H. Behnke, cm95aGJlaG5rZUBnbWFpbC5jb20=