
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Sustain. Food Syst. , 12 February 2021
Sec. Crop Biology and Sustainability
Volume 4 - 2020 | https://doi.org/10.3389/fsufs.2020.617065
This article is part of the Research Topic Plant Growth-Promoting Microorganisms for Sustainable Agricultural Production View all 50 articles
 Viviane Cristina Modesto1*
Viviane Cristina Modesto1* Marcelo Andreotti1Allan Hisashi Nakao2Deyvison de Asevedo Soares1Leonardo de Lima Froio1Lourdes Dickmann1Isabô Melina Pascoaloto1Isabela Maria Dias Moyses Fernandes1
Marcelo Andreotti1Allan Hisashi Nakao2Deyvison de Asevedo Soares1Leonardo de Lima Froio1Lourdes Dickmann1Isabô Melina Pascoaloto1Isabela Maria Dias Moyses Fernandes1The use of plant growth-promoting bacteria combined with Integrated Agricultural Production Systems (IAPS) has resulted in productivity increases in the Brazilian Cerrado region. Thus, the objective was to evaluate the effect of inoculating or not inoculating Azospirillum brasilense on corn and palisade grass seeds (Urochloa brizantha cv. Marandu) on growth, yield components, and dry matter yield of the aerial part and grains in an Oxisol cropped in ICLS and under a no-tillage system for 12 years in an irrigated area. The experimental design was a randomized block design with four replications. The treatments were composed of corn (monoculture) with inoculated seed, corn (monoculture) without inoculated seed, corn + palisade grass (intercropping) without inoculation, corn + palisade grass (intercropping) with inoculation in both seeds, corn + palisade grass (intercropping) with inoculation of corn seeds, and corn + palisade grass (intercropping) with inoculation of grass seeds. At the end of each production cycle, the yield and components of corn, corn straw biomass, and dry matter of palisade grass were evaluated. Considering the optimal conditions of soil provided by liming and fertilization at sowing and nitrogen applied in topdressing, the application of A. brasilense in corn seeds did not show its potential. Intercropped systems inoculated or not with A. brasilense on corn seeds increase grain yield, dry matter production of grass, and nutrient accumulation in the straw, providing greater sustainability to the Cerrado no-tillage system.
The intensification of land use for agricultural practices in the Cerrado region is increasing. However, inadequate soil management in this region has led to land degradation, with a consequent reduction in crop yields (Costa et al., 2015). In addition to degradation, the use of species not fitted to local conditions, low plant establishment before grazing, and soil fertility losses due to excessive nutrient extraction by the animal, among other factors, contribute to the degradation of the pastures (Hungria et al., 2016).
Incrementing competitiveness in the agricultural sector through the use of intensive crops as a means of supplying the food demand of developing countries has resulted in the need to replace the extensive traditional production model with systems that enable the maximization of land use as a no-tillage system (NTS) and integrated crop-livestock system (ICLS) (Pariz et al., 2020), as issues related to mitigation of CO2 emissions, water consumption, and overuse of fertilizers have become a worldwide concern (Hu et al., 2016).
However, the accumulation and maintenance of the straw in tropical regions such as the Cerrado are complicated practices due to the high temperatures and dry winter (Cavalli et al., 2018), which favor the rapid decomposition of these residues. However, mulching practices are especially important in these regions. The use of species such as palisade grass and guinea grass intercropping with grains cultures did not cause reductions in kernels' attributes and yield. Instead of this, these grass species have deep root systems that can maintain soil moisture, cycle nutrients, produce biomass, and forage for animal grazing in the off-season (Costa et al., 2020; Mateus et al., 2020). In addition, this system can reduce water losses due to evapotranspiration, as the soil is more protected against the incidence of sun and wind.
With the growing concern for maintaining natural resources and reducing possible losses of fertilizers, especially nitrogen, which are the most demanded by plants, there is a need to search for alternatives that aim to increase their efficiency or meet their need via biological N2 fixation (Yan et al., 2014), aiming at agricultural and environmental sustainability.
In this sense, studies using plant growth-promoting bacteria (PGPB), such as those of the genus Azospirillum, aimed at reducing or even suppressing mineral fertilizers without impairing production, are necessary. Inoculation with certain strains of Azospirillum sp. fitted to edaphoclimatic conditions under field conditions provides greater root development, resulting in greater soil exploitation for water and nutrient absorption. Furthermore, inoculation with these PGPB provides a direct contribution to increase crop yields and enhances the effectiveness of resource production with extensive environmental benefits (Santos et al., 2019).
The genus Azospirillum is the most utilized for corn crop worldwide and comprises 21 species (DSMZ–Deutsche Sammiung von Mikroorganismen und Zellkulturen GmbH, 2019). The strains of A. brasilense Ab-V5 and AB-V6 were released for use in commercial inoculants in maize and wheat in Brazil in 2009 since those strains resulted in increases in grain yield and greater absorption of nutrients (Hungria et al., 2010; Santos et al., 2019). In addition to its ability, PGPB can promote other strategies such as osmotic adjustment, maintenance of roots viability, membrane stability, and accumulation of protein and other metabolites such as proline, which helps in the maintenance of plant metabolism under drought stress (Ngumbi and Kloepper, 2016; Fukami et al., 2018).
However, it is noteworthy that this management does not always replace the use of nitrogen fertilizers and, in some cases, does not allow the reduction of doses to obtain the highest grain yield (Silva et al., 2015). In this context, it was hypothesized that the corn-palisade grass intercropping and the use of PGPB applied in seeds increase grain yield in corn and grass dry matter productivity under irrigated conditions in the low-land Cerrado without reducing the yield of the main crop.
The objective of this study was to evaluate the effect of inoculating or not with Azospirillum brasilense on corn and grass seeds under monoculture or intercropping with palisade grass (Urochloa brizantha cv Marandu) on growth, production components, and yield of dry matter of the aerial part and grains.
The study was composed of two experiments, repeated by two growing seasons (2014/2015 and 2015/2016), carried out in the same area irrigated by a sprinkler (center-pivot) in Central-West Brazil (20°20'S and “51°24'26” W, 370 m above sea level). The climate in this region is Aw, characterized as humid tropical with a rainy season in summer and dry winter, according to Koppen (Unicamp—Centro de Pesquisa Meteorológicas e Climáticas Aplicadas à Agricultura, 2016). The long-term (1956–2013) average annual maximum and minimum temperatures are 31.3 and 18.4°C, respectively. The precipitation rate, maximum and minimum temperatures, and photoperiod of the area of this study were measured (Table 1).
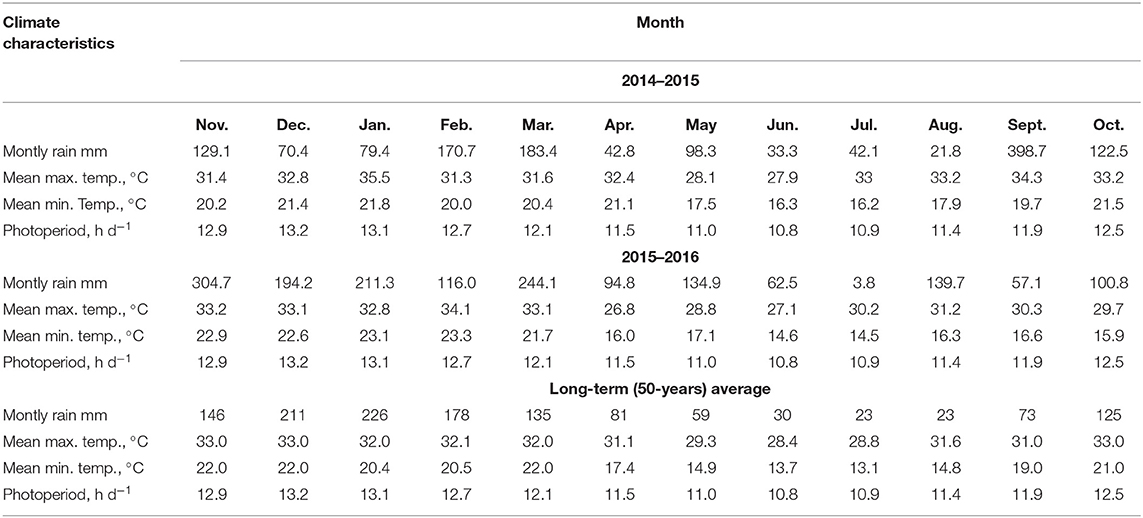
Table 1. Rainfall, maximum and minimum temperatures, and photoperiod during the study period.
The soil in the experimental area was classified as Oxisol (FAO—Food and Agriculture Organization of the United Nations, 2006), clayey, and had a history of 12 years under the NTS, establishment phase, with annual and semi-perennial crops, where soybean was the previous crop.
Before initiating the study in each year, chemical and physical attributes were determined. For physical attributes, the methods described by EMBRAPA–Empresa Brasileira de Pesquisa Agropecuária (1997) were followed, and for chemical attributes, the methodology of van Raij et al. (2001) was followed. The values in the 0.00- to 0.10-m and 0.10- to 0.20-m layers are shown in Table 2.
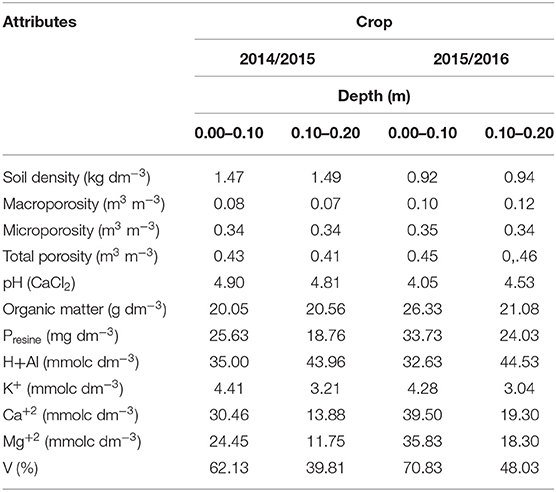
Table 2. Soil physical and chemical attributes at 0.00–0.10 and 0.10–0.20 m depths in 2014/2015 and 2015/2016 crops.
The area was irrigated by a sprinkler (center-pivot), considering the optimal water range for the crops under study. The following equation was used to establish available water capacity (AWC):
where FC is the field capacity (%); PWM, permanent wilting point (%); SD, the soil density (kg dm−3); and ERZD, the effective root zone depth (m).
Those data were obtained from the soil water retention curve, where FC = 20.25%; PWM = 14.58%; SD = 1.48 and 0.93 kg dm−3 (2014/2015 and 2015/2016 crops, respectively); and ERZD = 0.20 m. Therefore, the evaluated soil AWC was 16.78 and 11.54 mm for the 2014/2015 and 2015/2016 crops, respectively.
The water supply was transmitted with a flow of 3.3 mm h−1. Irrigation was applied every time the maximum crop evapotranspiration (ETm) reached 7.43 and 5.11 mm for the respective crop years (<44.3% AWC). The ETm was estimated through the following equation:
where Kc is the crop coefficient and ETo is the reference evapotranspiration.
ETo was calculated through the following equation:
where Kp is the Class A tank coefficient and ECA is the Class A tank evaporation (mm day−1). Water evaporation measurement (mm) was obtained daily from a Class A tank. The Kp was calculated as proposed by Doorenbos and Pruitt (1977) based on the surrounding area, wind speed, and relative air humidity.
The experimental design was a randomized block with four replications, and six treatments consisted of different combinations of intercropping and monoculture corn and palisade grass, as follows: corn (monoculture) with inoculated seed, corn (monoculture) without inoculated seed, corn + palisade grass (intercropping) without inoculation, corn + palisade grass (intercropping) with inoculation in both seeds, corn + palisade grass (intercropping) with inoculation of corn seeds, and corn + palisade grass (intercropping) with inoculation of grass seeds. Each plot consisted of 3.4 m in width and 20 m in length, totaling 68 m2. The usable area in each plot consisted of four central rows, which did not include 1 m at the ends of each plot and two external rows near the edge. In the second year, the plots were allocated at the same location as the first year.
Prior to the implementation of the experiment (October 2014), diazotrophic microorganisms were counted to determine the bacterial population in the number of cells per milliliter, performed by estimating the “Most Likely Number” (MPN) using the MacCrady table in NFB (A. brasilense) semi-solid medium according to the methodology described by Döbereiner et al. (1995), obtaining a value of 9.0 × 108 g−1 cells.
On October 10 of 2014 and 2015, desiccation of the plants in the experimental area was carried out, aiming the eradication of weed and mulching for continuity of the NTS, using the herbicide glyphosate [isopropylamine salt of N–(phosphonomethyl) glycine] (1.44 g acid-equivalent ha−1) at a spray volume of 200 L ha−1. On November 1 of 2015 and 2016, plants were cut with mechanical management using a plant residue crusher.
Simple hybrid DKB 390 YG corn [Zea mays (L.)] was used on the first growing season (2014/15). On the second growing season (2015/16), there was no availability for the same hybrid. Triple hybrid DKB 350 YG was the option for this year. The grass used for this study, in both years, was palisade grass [Urochoa brizantha (A. Rich.) Stapf Marandu].
To ameliorate soil acidity and according to the soil analysis, 30 days prior to sowing, dolomite limestone (40% CaO and 12% MgO) with an 85% effective calcium carbonate equivalence (85% PRNT) Dolomitic limestone was applied over the soil surface without incorporation, aiming to increase base saturation to 70% (0.00–0.20 m) as described by Cantarella and Raij (1997).
At sowing of the respective growing season, 2014/15 and 2015/16, the seeds of corn and palisade grass were inoculated or not with the diazotrophic bacterium A. brasilense (strains Ab-V5 and Ab-V6). The bacteria come from EMBRAPA Soybeam program, selected on Brazil initially for corn and wheat crops (Hungria et al., 2010) at a dose recommended from this mentioned author of 100 ml/25 kg seeds. Inoculation occurred before sowing and in the shade.
The first growing season was sown on November 11, and the second one was sown on November 17. For both years, the corn crop was mechanically sown using a no-till drill at a depth of ~0.05 m in 0.45 m spacing with 3.0 seeds m−1, aiming to reach a final stand close to 60,000 plants ha−1.
In both cropping years, palisade grass was sown on the same day as corn, with another type of seeder-fertilizer, with an alternating double-disc mechanism compatible with NTS, in which the seeds were placed in the fertilizer compartment in the seeder and deposited at a depth of 0.06 m, at a spacing of 0.17 m, using ~7 kg ha−1 of viable pure seeds (CV = 70%) for palisade grass. Thus, the grass seeds were below the corn seeds, following the recommendations of Kluthcouski et al. (2000), with the objective of delaying the emergence of grass in relation to the grain-producing crop and reducing the likely competition among the species in the initial period of the crop development.
Due to the corn crop sowing, in both crops, respectively, the sowing mineral fertilization was performed in the furrow of the grain crop, according to the results obtained in the sampling for initial soil characterization, where 300 kg ha−1 of formulated 08-28-16 (24 kg ha−1 N, 84 kg ha−1 P2O5, and 48 kg ha−1 K2O, respectively) was applied.
At the V6 phenological stage (six fully developed corn leaves), following the recommendations of Cantarella (2007), the topdressing fertilization was carried out by manually applying 120 kg ha−1 N–ammonium sulfate to the corn lines.
During the corn flowering period, in both crop years, after the emission of female inflorescence, that is, when more than 50% of the plants were clogged, and with the presence of stigmas style (hair), the middle-thirds of 15 leaves were collected per plot for free proline quantification (01/30/2015 and 01/17/2016); 0.5 g of fresh leaf limb tissue was used, which was immediately frozen in liquid nitrogen to cease leaf metabolism. After collection, the material was properly stored in a freezer at temperatures below −20°C and analyzed using the methodology described by Bates et al. (1973).
Corn crop cycles for grain production were 140 and 120 days, respectively, after emergence (DAE), in the first and second year, respectively.
One day before the harvest of the 2014/2015 and 2015/2016 sowing seasons, the following morphological components were determined: basal stem diameter (BSD), plant height (PH), and height of ear insertion (HEI). Then, the number of plants and the number of ears contained in the useful area of each experimental unit (two 20-m-long central lines, excluding 1 m from each side row in each plot) were counted to determine the final plant stand (FPS).
Before harvesting (03/07/2015 and 04/01/2016, respectively), to determine the corn yield and yield components, all ears contained in the plot useful area were manually harvested. The following yield components were evaluated: principal main length (EL), principal ear diameter (ED), number of grains/ear (NGE), and 100-seed weight (W100) (130 g kg−1 wet basis) were determined for 10 plants per plot that were randomly chosen from the usable area. Subsequently, the ears were mechanically threshed, and the grains were weighted, so the plot yield was calculated, extrapolated to kg ha−1, and corrected for 130 g kg−1.
Following corn harvesting in the respective harvests, the straw yield resulting from corn and grass dry matter cultivation was evaluated per plot. Therefore, the residues of corn plants were collected in a 2-m line/plot. The grass was collected with a metal square (1.0 × 1.0 m) at a cutting height of 0.30 m, and in each sample, the cut material was dried under forced air circulation at 65°C for 72 h to quantify the total dry matter production of grass and corn straw, both extrapolated to Mg ha−1. Subsequently, the material was ground (Willey-type) and sent to the laboratory where contents of N, P, K, Ca, Mg, and S were determined (Malavolta et al., 1997). Nutrient accumulations per hectare were calculated based on dry matter production and nutritional contents.
Data were normally distributed (W > 0.90) according to the Shapiro-Wilk test, with the results indicating that all data were distributed normally (W > 0.9). Means of the treatment were compared using the LSD test. The effects were considered statistically significant at p ≤ 0.05. Statistical analyses were performed using the “R” software (2020).
The agronomics characteristics PH, HEI, and SBD were significant for both growing seasons (p < 0.05) (Table 3). In the first year, treatments inoculated with A. brasilense in both seeds (corn and palisade grass) were also significant. They obtained higher growth, whereas SBD had higher results for intercropped inoculated only in corn.
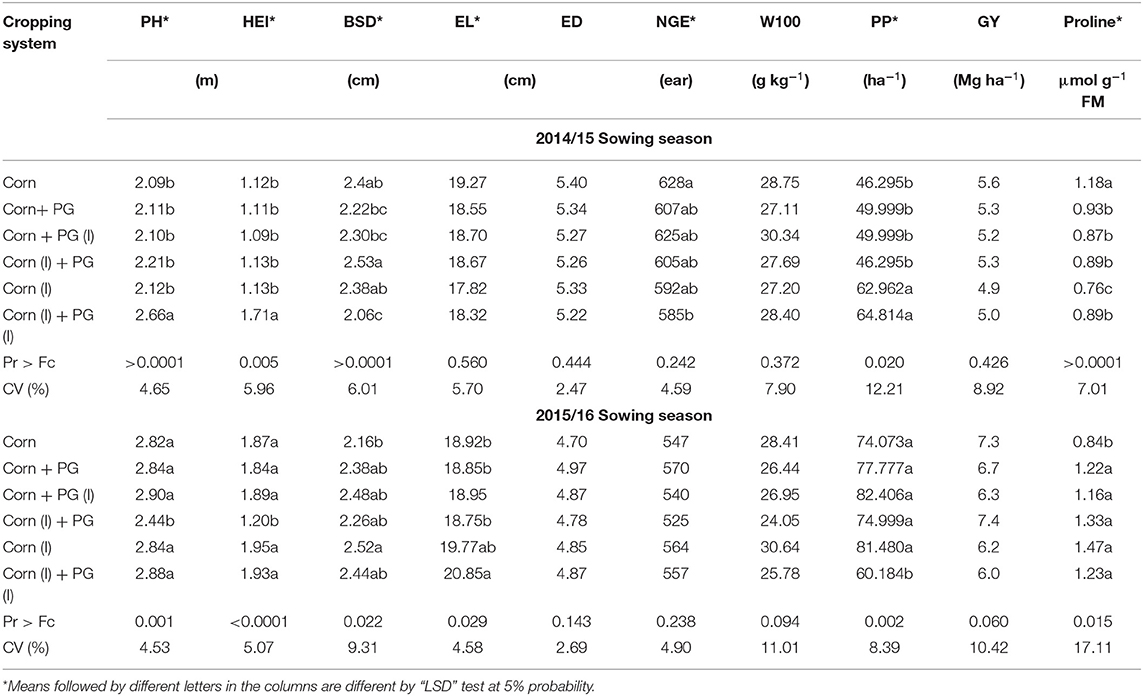
Table 3. Means of plant height (PH), height of ear insertion (HEI), basal steam diameter (BSD), ear diameter (DE), ear length (EL), number of grains per ear (NGE), 100-seed weight (W100), plant population (PP) grain yield (GY) and proline content in intercropped or single corn, A. brasilense (irrigated area) inoculated or not in the seeds in 2014/15 and 2015/16 crops.
At the second growing season, PH and HEI showed better values for all treatments except for the intercropped corn inoculated on corn seeds. Otherwise, for SBD, the monoculture corn inoculated showed a higher diameter. The production attributes EL, ED, NGE, and W100 were not significant for cropping systems or inoculation (p > 0.05), showing that these features are no responses to those systems.
The PP attribute was significant for both growing seasons. In the first season (p = 0.02), PP was higher for corn inoculates in monoculture or intercropped inoculate in both seeds. In the second season (p = 0.002), the only treatment that presents lower PP was the intercropped inoculated in both seeds. On the other hand, PP reached the established amount of 60,000 plants ha−1.
Grain yield was not significant for both seasons (p > 0.05). However, it presents higher GY in the second year. This is related to the fact that in this year, the PP was higher than the first year, resulting in higher productivity.
Proline was significant (p < 0.001) at the first growing season with high levels for monoculture corn without inoculation, whereas in the second growing season, proline was also significant (p = 0.015) with lower levels only for monoculture corn without inoculation.
Corn straw yield was significant for both growing seasons (p < 0.05) (Table 4), although showing different values between years, the system that provided the highest straw production was corn intercropped with palisade grass and inoculated in corn seeds (Table 4).
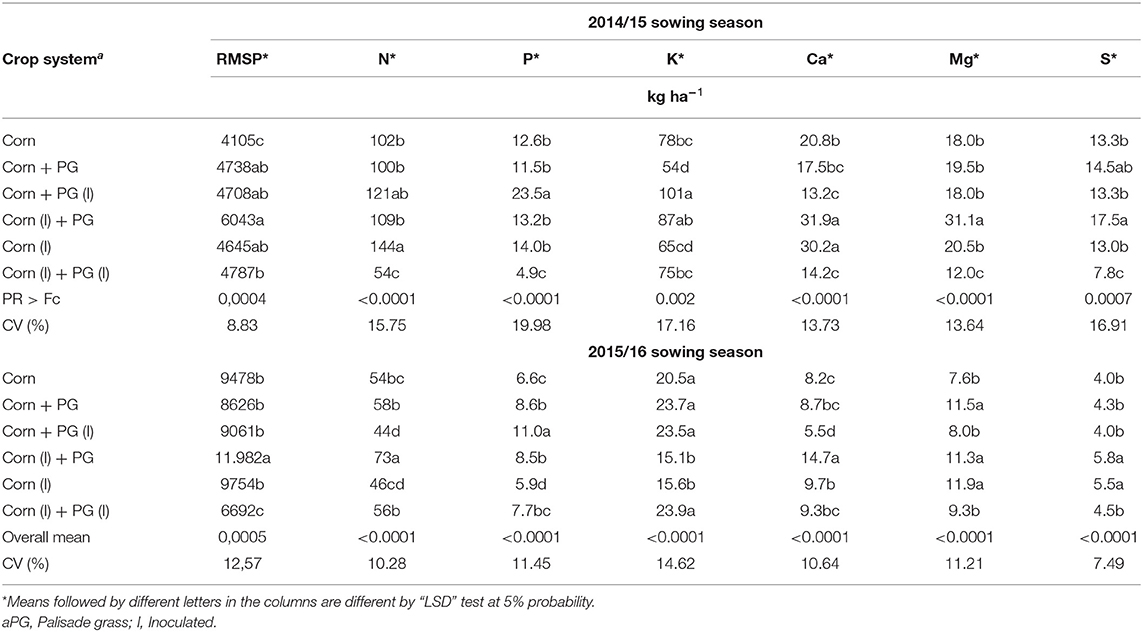
Table 4. Remaining corn straw production (RMSP) and nutrient accumulation in corn straw in 2014/15 and 2015/16 sowing seasons.
There was a significant effect of intercropping corn and palisade grass for both growing seasons (p = 0.004 and p = 0.0005), respectively. Intercropping with A. brasilense in corn seeds inoculated promotes higher corn straw yield.
The macronutrient accumulation in corn straw was significant for the crop systems evaluated (N, P, K, Ca, Mg, and S) for both years (Table 4). It was observed that in the first season, due to the lower MSY and yield production, the nutrient export by plants was not much, showing on average 50% more nutrients accumulated for this year. The macronutrients were not influenced by local inoculation (corn or grass seed).
In the first season, the corn monoculture presented a higher N accumulation. P and K present higher values for intercropped systems with palisade grass inoculated. Ca content was higher for corn inoculated, independent of monoculture or intercropped system. Mg and S found better values for intercropped inoculated in corn seeds.
In the second season, corn intercropped was higher for P, K, Mg, and S, demonstrating higher values for both intercropping and monoculture, independent of the local inoculation.
Palisade grass dry matter yield showed significant results at the first growing season (p < 0.05) (Table 5). However, the effect of the inoculation with PGPB was independent of crop systems. For the next season, because of the better growth of corn, palisade grass shows lower values and not significant results.
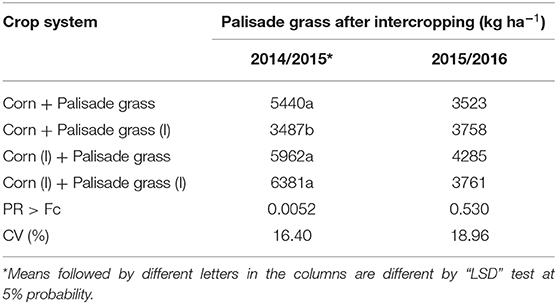
Table 5. Dry matter yield of Palisade grass in an irrigated area after intercropped with corn in 2014/15 and 2015/16 crops.
On the first growing season, PH and HEI (Table 3) were higher when intercropping was inoculated in both seeds. Some reports found that inoculating with this genus of PGBP guaranteed greater plant height and root mass, especially in dry conditions influencing the absorption of nutrients and water from the soil (Fukami et al., 2018; Leite et al., 2018) positively. These results are lower than the results found in the second season. The reduction in PH and consequent HEI is due to the already mentioned fact that the high temperatures lasted throughout the vegetative phase (V8), a period of intense growth of the plant in height (Silva et al., 2015), which will reflect in the lower yield of corn straw.
Plant height is a morphological variable that is directly related to the population density of the crop. Smaller plants tolerate higher densities, while higher plants tolerate lower densities (Matos et al., 2017). It is also known that well-nourished nitrogen plants have a greater ability to assimilate CO2 and synthesize carbohydrates during photosynthesis, resulting in higher growth (Vogel et al., 2013).
As observed at the end of the second season, chemical characteristics of soil improved intercropped systems such as grasses associated with A. brasilense that present a root structure capable of absorbing a larger amount of nutrients and water (Bashan and de-Bashan, 2010) and also improve physical characteristics by way of the capacity of the root system promoting aeration as seen in Table 2.
The more favorable climate conditions in the second year resulted in better conditions of plant development, which justifies the larger plant growth. It is noteworthy that the greater plant height provides a greater productive potential of fresh and dry matter and, consequently, higher straw production for the next crop. However, above-average plant heights may result in greater ease of lodging by wind, rain, and machine traffic, causing productivity losses, a fact not found in this study.
Still, regarding the morphological characteristics, the crop and inoculation systems did not influence the BSD in both years (Table 3). Even at higher densities such as those of the second season, the crop did not show any problems such as lodging and breaking of the plants, a fact common in larger populations, as plants etiolate in search of light.
In stress conditions like the first year, intercropped systems inoculated in corn seeds can promote a higher BSD that works as a reservation of nutrients. In systems with good edaphoclimatic conditions, corn monoculture shows higher PP, in dense crops, the development of plants saved nutrient reservation for its development.
These results corroborate those found by Silva et al. (2015), in that when evaluating intercropping and monoculture corn cultivation systems, at different spacing, and in the Cerrado area, they did not find differences for BSD between the systems. In addition to providing increased breakage and lodging resistance, BSD stores essential soluble solids for photoassimilate translocation and grain formation.
Opposite results for EL and ED were reported by Mumbach et al. (2017) using doses of mineral fertilizer (sowing and mulching) and inoculation with A. brasilense, and for the EL in a study by Moreira et al. (2019) when evaluating the economic performance of corn as a function of inoculant application forms and nitrogen fertilizer doses. This lack of responses to cropping systems may be associated with plant genotype and edaphoclimatic conditions.
The absence of significant results for NGE and W100 (Table 3) can be explained by the uniformity of data from EL and ED. Those results for W100 corroborated with the study of Silva et al. (2015), who also did not observe any significance for this production component when evaluating inoculation with A. brasilense and nitrogen doses in the Cerrado. However, in the second year, results found for exclusive and inoculated corn were better. This is an important component for grain yield and may be altered by any type of stress occurring in the grain filling phase, in which the external factors are the most limiting to plant development (Freitas et al., 2013). In addition, Pariz et al. (2009) reported that the intercropping system of corn with other forages might compromise the translocation of photoassimilates to grains, producing lighter grains.
In that year, PP was better when inoculates in monoculture or intercropped with inoculations in both seeds (2015/16). The average plant ha−1 was lower than the PP established for this study, which was 60,000 plants ha−1. This result is due to the influence of the high temperature on that year when the experiment was conducted (Table 1). The higher temperature, the faster the metabolic process, which leads to early leaf senescence. The ideal temperature for crop development from emergence to flowering is between 24 and 30°C (Cruz et al., 2010).
It can be observed from the evaluation of simultaneous sowing of corn and palisade grass, regardless of the cropping system, that there was no negative influence (Table 3) of competition between species. Corroborating the results of Araújo et al. (2018) and Garcia et al. (2013), both evaluating corn yield in association with palisade grasses, also found that intercropping systems did not influence the final plant stand. Corn PP has direct effects on crop yield and also forage production (Crusciol et al., 2010). This component is related to SBD, and it is expected that the greater supply of water and nutrients will provide a larger plant size and, consequently, a higher PP.
In addition, the larger number of plants ha−1 (75,000 plants ha−1) promotes greater plant growth in height (Calonego et al., 2011; Freitas et al., 2013), with a consequent increase in HEI, corroborating the results of the second year, when PP was over 60,000 plants ha−1. Another factor contributing to the larger PP is that a larger plant population can provide greater height growth, reducing evapotranspiration losses (Queiroz et al., 2016).
On the first growing season, GY has lower values because of the lower PP, a consequence of high temperatures that year, which promotes low germination. Otherwise, production showed satisfactory results, with an average of over 4.9 Mg ha−1 and 7.0 Mg ha−1 in the second season, and more than 116 bags ha−1. In corn, the strains of A. brasilense Ab-V5 and Ab-V6 resulted in an increase in grain yield that reached 27% compared with the non-inoculated control (Hungria et al., 2010). This was not observed in this study because of the different edaphoclimatic conditions.
Although Silva et al. (2015) reported that simultaneous sowing between corn and palisade grass could influence the final grain yield through competition between crops, especially after corn flowering, it was found in this study that the inoculation of corn seeds becomes viable in NTSs with correct input from plant material derived from previous crops because even with the coexistence with forage, higher results for the intercropped systems without any inoculation were achieved.
The most important contribution of the use of A. brasilense is the possibility of increasing dry matter accumulation rate, resulting in an incremented biomass and plant height, accelerating germination and benefiting the root system, and as a consequence, there may be an increase in grain yield due to greater nutrient translocation to grain and reduced competition with grass (Hungria et al., 2010).
Proline is known as an important plant osmoregulator, and its transport is assumed to influence tolerance against stress conditions. In both years, the results of free proline quantification in corn leaves had different levels (Table 2).
Proline metabolism is regulated by the balance between its anabolism and catabolism. Its deposition in parts of the plant may not occur due to an increase in biosynthesis but rather due to its transport and utilization rate (Per et al., 2017). Under normal conditions, proline biosynthesis occurs in the cytosol, while under stress conditions, it may occur in chloroplasts (Rejeb et al., 2014). In agreement with the results observed between the evaluated years, in the first year, the greater evapotranspiration caused the stomata to close so that there was no excessive water loss.
No losses in the utilization rate and transport of the proline contents occurred in the second year as the climatic conditions were more favorable to the development of the crop, thus without stress conditions. Also, the corn cropped in a single system without inoculation showed lower levels of proline; therefore, less deposition and higher utilization rates were found.
Studies show that phytohormones such as auxins, gibberellins, cytokines, ethylene, ABA, and salicylic acid are related to plant growth, development, metabolism, and response to different stress conditions, but these results are still scarce (Iqbal et al., 2009). However, the benefits of phytohormones produced by the action of inoculation with A. brasilense throughout this study can be observed.
The remaining corn straw in NTS on the first growing season was lower compared to the second season. Otherwise, the system that promotes the high straw production was corn intercropped with palisade grass and inoculated in corn seeds (Table 4). The lower remaining corn straw in NTS in the first growing season compared with the second one was expected. The higher temperatures in January 2015 (Table 1) in the flowering phase promotes the aborting of flowers, making the germination infeasible. It is known that moisture shortage affects stomatal functions, reducing the leaf CO2/O2 ratio and inhibiting photosynthesis with concomitant reduction of biomass production (Lopes et al., 2011).
Otherwise, the crop system with corn intercropped with palisade grass inoculated in corn seeds promoted the higher remaining straw, once again indicating the benefits of plant growth-promoting bacteria (PGPB) in that system. This symbiosis between PGPB and plants provided yield productivity gains of grain and forage crops, resulting in a higher accumulation of dry matter after harvest, providing enough straw to maintain NTS.
Another factor that should be taken into account in the implementation of the crop in ICLS is the spatial arrangement and sowing type, which must be defined according to the objective of the system (Borghi et al., 2012) or some water restriction. High densities are recommended only when there is no water restriction (Freitas et al., 2013). The spacing used in this study was 0.45 m, with simultaneous sowing of corn and grass, providing high densities and sufficient dry matter production to maintain the NTS recommended by Kluthcouski et al. (2000), which is >6.0 Mg ha−1.
It was observed that, in the first year, due to lower MRS and GY, the nutrient exportation by plants was lower, showing, on average, 50% more nutrients accumulated for this year. The largest accumulation of N and K stands out, confirming that this nutrient is the most absorbed and accumulated in plant tissues, corroborating the results found by Pariz et al. (2011), Costa et al. (2014), Mendonça et al. (2015), under the same edaphoclimatic conditions.
The soil in the experimental area has been cultivated in SPD for over 12 years when the immobilization of N approaches mineralization (Anghinoni, 2007). Therefore, the accumulation of O.M. and mulch started on the surface, which, combined with corn topdressing fertilization, provided a higher accumulation of this nutrient, especially when inoculated with A. brasilense.
The higher K extraction in intercropping systems is due to the greater competition with palisade grass, which extracts more of this nutrient from the soil. The intercropped system corn + palisade grass is unique in providing K because of the greater amount of exchangeable K in soil (Mateus et al., 2020). However, because it has no structural function in plants, K has a short half-life. Therefore, it is important for the supply of this nutrient in subsurface soil layers for succession crops (Cavalli et al., 2018).
Cropping systems and inoculation with A. brasilense did not influence Ca, Mg, and S. Therefore, the adoption of practices, such as intercropping and use of PGPBs, provides a higher quality of NTS, providing a greater return of nutrients to the soil through greater input of residual biomass. In addition, when the grain-producing crop is intercropped with summer forage grasses, such as Uroclhoas, it is possible to reduce the cost of pasture formation, as a food supply to ruminants in the off-season, where the availability of forage is limited (Costa et al., 2014).
On the first growing season, the effect of inoculation with the recommended doses and strains for corn crop was independent of the inoculated species (corn or grass) (Table 5). Because palisade grass has a vigorous and deep root system, the growth-promoting effects of A. brasilense inoculation on root growth and biomass growth may not have manifested, especially because the crop is irrigated, determining that the plants do not need to deepen the root system in the search for water and nutrients. Similar results were observed by Barducci et al. (2009), in that upon evaluating the inoculation of species of the genera Urochloa and Megathyrsus, they reported that the greater root development of these species makes them more tolerant to water and nutrient deficits and may interfere with the hormonal effects of Azospirillum.
Otherwise, it is important to consider that strains of Azospirillum may differ in their properties to confer tolerance in drought and gains in yield (García et al., 2017). The variation between years is attributed to the overgrowth of corn plants in that year, providing greater shading of the grass, leading to its low metabolic and growth activity. These results are reinforced by Table 1, where corn straw dry matter production was higher in the second year.
When corn straw is added to grass straw production, means of 10.1 and 13.1 kg ha−1 are observed for the first and second year, respectively. In Cerrado regions, where high temperatures and dry winter predominate, these values can supply the balance needed to maintain soil cover and nutrient cycling in this climate, results close to those found by Sá et al. (2015), which ranged from 11.7 to 13.3 kg ha−1 under the same soil and climate conditions.
In that year, when corn was intercropped with palisade grass, regardless of whether or not it is inoculated with A. brasilense, Modesto et al. (2019) observed a greater profitability index as well as operating profit; the positive results found in intercropped systems in the first year and in the second year were 25.6 and 59.77%, respectively, with the average difference of 34.21% due to the variables in the climatic conditions between the years that resulted in higher corn production in the second harvest. These results show that intercropping corn with palisade grass could amortize the production costs of the system and provide grass input to subsequent silage.
Considering the optimal conditions of soil provided by liming and fertilization at sowing and nitrogen applied in topdressing, the application of A. brasilense in corn seeds did not show its potential. Otherwise, in the current study, intercropped systems increase grain yield, dry matter production, and nutrient accumulation in the straw, producing more biomass and providing greater sustainability to NTS in the low-land Cerrado. These results of intercropped systems have great importance, specifically in these tropical regions.
The original contributions presented in the study are included in the article/supplementary files, further inquiries can be directed to the corresponding author.
VM conceived and designed the analysis, collected the data, contributed the data, performed the analysis and wrote the paper. MA conceived and designed the analysis, contributed the data, and wrote the paper. AN and DS collected the data, contributed the data, and performed the analysis. LF, LD, IP, and IF collected the data and performed the analysis. All authors contributed to the article and approved the submitted version.
This work was supported by Scholarship: FAPESP (process number 2014/02697-1).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The data presented in this article were initially reported in the thesis of the corresponding author and is available here: https://repositorio.unesp.br/bitstream/handle/11449/152235/modesto_vc_dr_ilha.pdf?sequence=3&isAllowed=y. We thank FAPESP – Research Support Foundation of the State of São Paulo for granting the first author a doctorate scholarship (Process no. 2014/02697-1).
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fsufs.2020.617065/full#supplementary-material
Anghinoni, I. (2007). “Fertilidade do solo e seu manejo em sistema plantio direto,” in Fertilidade do Solo, eds R. F. Novais, V. V. H. Alvarez, N. F. Barros, R. L. F. Fontes, R. B. Cantarutti, and J. C. L. Neves (Viçosa: Sociedade Brasileira de Ciência do Solo), 873–928.
Araújo, L. S., Branquinho, J. A. S., Silveira, P. M., Silva, L. G. B., Valente, M. S., Siqueira, M. V. R., et al. (2018). Corn yield (Zea mays) intercropped with urochloa brizantha at different row spacing in southeast of Goiás State. Agrarian 42, 307–318. doi: 10.30612/agrarian.v11i42.4335
Barducci, R. S., Costa, C., Crusciol, C. A. C., Borghi, É., Putarov, T. C., and Sarti, L. M. N. (2009). Production of brachiaria brizantha and panicum maximum with corn and nitrogen fertilization. Arch. Zootec. 222, 211–222. doi: 10.4321/S0004-05922009000200006
Bashan, Y., and de-Bashan, L. E. (2010). Chapter two—How the plant growth-promoting bacterium Azospirillum promotes plant growth–a critical assessment. Adv. Agron. 108, 77–136. doi: 10.1016/S0065-2113(10)08002-8
Bates, L. S., Waldren, R. P., and Teare, I. (1973). Rapid determination of free proline for water-stress studies. Plant Soil 39, 205–207. doi: 10.1007/BF00018060
Borghi, E., Crusciol, C. A. C., Nascente, A. S., Mateus, G. P., Martins, P. O., and Costa, C. (2012). Effects of row spacing and intercrop on corn grain yield and forage production of palisade grass. Crop Past. Sci. 63, 1106–1113. doi: 10.1071/CP12344
Calonego, J. C., Poleto, L. C., Domingues, F. N., and Tiritan, C. S. (2011). Yield and growth of corn in different plant arrangements. Agrarian 12, 84–90.
Cantarella, H. (2007). “Nitrogênio,” in Fertilidade do solo, eds R. F. Novais, V. V. H. Alvarez, N. F. Barros, R. L. F. Fontes, R. B. Cantarutti, and J. C. L. Neves (Viçosa: Sociedade Brasileira de Ciência do Solo), 375–470.
Cantarella, H., Raij, B., and van Camargo, C. E. O. (1997). “Cereais,” in Boletim Técnico 100: Recomendação de Adubação e Calagem para o Estado de São Paulo, 2nd Edn, eds B van Raij, H. Cantarella, J. A. Quaggio, and A. M. C. Furlani (Campinas: Instituto Agronômico–IAC), 43–71.
Cavalli, E., Lange, A., Cavalli, C., and Behling, M. (2018). Decomposition and release of nutrients from crop residues on soybean-corn cropping systems. Rev. Bras Ciênc Agr 2:5527. doi: 10.5039/agraria.v13i2a5527
Costa, N. R., Andreotti, M., Buzetti, S., Lopes, K. S. M., Santos, F. G., and Pariz, C. M. (2014). Macronutrient accumulation and decomposition of brachiaria species as a function of nitrogen fertilization during and after intercropping with corn). Rev. Bras. Cienc. Solo. 38, 1223–1233. doi: 10.1590/S0100-06832014000400019
Costa, N. R., Andreotti, M., Crusciol, C. A. C., Pariz, C. M., Bossolani, J. W., Castilhos, A. M., et al. (2020). Can palisade and guinea grass sowing time in intercropping systems affect soybean yield and soil chemical properties? Front. Sustain. Food Syst. 4:81. doi: 10.3389/fsufs.2020.00081
Costa, N. R., Andreotti, M., Lopes, K. S. M., Yokobatake, K. L., Ferreira, J. P., Pariz, C. M., et al. (2015). Soil properties and carbon accumulation in an integrated crop-livestock system under no-tillage. Rev. Bras. Ciênc. Solo 3, 852–863. doi: 10.1590/01000683rbcs20140269
Crusciol, C. A., Soratto, R. P., Borghi, E., and Mateus, G. P. (2010). Benefits of integrating crops and tropical pastures as systems of production. Better Crops 2, 14–16.
Cruz, J. C., Filho, I. A. P., Alvarenga, R. C., Neto, M. M. G., Viana, J. H. M., Oliveira, M. F., et al. (2010). Embrapa Milho e Sorgo Sistemas de Produção, 2 ISSN 1679-012X Versão Eletrônica−6 ªedição. Available online at: https://ainfo.cnptia.embrapa.br/digital/bitstream/item/27037/1/Plantio.pdf (accessed September, 2019).
Döbereiner, J., Baldani, V. L. D., and Baldani, J. I. (1995). Como isolar e identificar bactérias diazotróficas de plantas não-leguminosas. Brasilia: Embrapa SPI.
Doorenbos, J., and Pruitt, W. O. (1977). Guidelines for Predicting Crop Water Requirements. Irrigation and Drainage Paper, 24. Rome: FAO, pp. 179.
DSMZ–Deutsche Sammiung von Mikroorganismen und Zellkulturen GmbH (2019). Prokaryotic Nomenclature up to date. Available online at: https://www.dsmz.de/bacterial-diversity/prokaryotic-nomenclature-up-to-date/prokaryotic-nomeclature-up-to-date.html (accessed November 24, 2020).
EMBRAPA–Empresa Brasileira de Pesquisa Agropecuária (1997). Centro Nacional de Pesquisa de Solos. Rio de Janeiro: Manual de métodos de análise de solo. pp. 212.
Freitas, R. J., Nascente, A. S., and De, S. S. (2013). Population of corn plants intercropped with urochloa ruziziensis. Pesq. Agropec. Trop. 1, 79–87. doi: 10.1590/S1983-40632013000100011
Fukami, J., Ceresini, P., and Hungria, M. (2018). Azospirillum: benefits that go far beyond biological nitrogen fixation. AMB Expr. 8:73. doi: 10.1186/s13568-018-0608-1
Garcia, C. M. P., Andreotti, M., Filho, M. C. M. T., Buzetti, S., Celestrino, T. S., and Lopes, K. S. M. (2013). Agronomic performance of corn and forages species in Crop-Livestock Integration system in the Cerrado. Ciênc. Rural 4, 589–595. doi: 10.1590/S0103-84782013000400005
García, J. E., Maroniche, G., Creus, C., Suárez-Rodríguez, R., Ramirez-Trujillo, J. A., and Groppa, M. D. (2017). In vitro PGPR properties and osmotic tolerance of different Azospirillum native strains and their effects on growthof maize under drought stress. Microbiol. Res. 202, 21–29. doi: 10.1016/j.micres.2017.04.007
Hu, F., Gan, Y., Cui, H., Zhao, C., Fenga, F., Yin, W., et al. (2016). Intercropping corn and wheat with conservation agriculture principles improves water harvesting and reduces carbon emissions in dry areas. Eur. J. Agron. 74, 9–17. doi: 10.1016/j.eja.2015.11.019
Hungria, M., Campo, R. J., Souza, E. M., and Pedrosa, F. O. (2010). Inoculation with selected strains of Azospirillum brasilense and A. lipoferum improves yields of corn and wheat in Brazil. Plant Soil 331, 413–425. doi: 10.1007/s11104-009-0262-0
Hungria, M., Nogueira, M. A., and Araujo, R. S. (2016). Inoculation of Brachiaria spp. with the plant growth-promoting bacterium Azospirillum brasilense: an environment-friendly component in the reclamation of degraded pastures in the tropics. Agric Ecosyst. Environ. 221, 125–131. doi: 10.1016/j.agee.2016.01.024
Iqbal, J., Hua, R., Lin, S., Hatano, R., Feng, M., Lu, L., et al. (2009). CO2 emission in a subtropical red paddy soil (Ultisol) as affected by straw and N-fertilizer applications a case study in Southern China. Agric. Ecosyst. Environ. 131, 292–302. doi: 10.1016/j.agee.2009.02.001
Kluthcouski, J., Cobucci, T., Aidar, H., Yokoyama, L. P., Oliveira, I. P., Costa, J. L. S., et al. (2000). Sistema Santa Fé–Tecnolgia Embrapa: Integração Lavoura-Pecuária pelo consórcio de culturas anuais com forrageiras, em áreas de lavoura, nos sistemas direto e convencional. Circular Técnica, 38. Santo Antônio de Goiás: Embrapa Arroz e Feijão (Circular Técnica, 38), p.28.
Leite, R. D. C., Santos, J. G., Silva, E. L., Alves, C. R., Hungria, M., Leite, R. D. C., et al. (2018). Productivity increase, reduction of nitrogen fertiliser use, and drought-stress mitigation by inoculation of Marandu grass (Urochloa brizantha) with Azospirillum brasilense. Crop Past. Sci. 70, 61–67. doi: 10.1071/CP18105
Lopes, M. S., Araus, J. L., van Heerden, P. D. R., and Foyer, C. H. (2011). Enhancing drought tolerance in C4 crops. J. Exp. Bot. 62, 3135–3153. doi: 10.1093/jxb/err105
Malavolta, E., Vitti, G. C., and Oliveira, A. S. (1997). Avaliação do estado nutricional das plantas: princípios e aplicações, 2nd Edn. Piracicaba: Associação Brasileira para Pesquisa da Potassa e do Fosfato.
Mateus, G. P., Crusciol, C. A. C., Pariz, C. M., Costa, N. R., Borghi, E., Costa, C., et al. (2020). Corn intercropped with tropical perennial grasses as affected by sidedress nitrogen application rates. Nut. Cycl. Agroecosyst. 116, 223–244. doi: 10.1007/s10705-019-10040-1
Matos, F., Oliveira, F. F., Pietroski, M., Muller, P. F., Takeshita, V., and Caione, G. (2017). Use of Azospirillum brasilense to increase the efficiency of nitrogen fertilization on corn. Rev. Bras. Mil. Sor. 1, 131–141. doi: 10.18512/1980-6477/rbms.v16n1p131-141
Mendonça, V. Z., Mello, L. M. M., Andreotti, M., Pariz, C. M., Yano, E. H., and Pereira, F. C. B. L. (2015). Nutrient release from forage straw intercropped with corn and followed by soybean. Rev. Bras. Ciênc. Solo 39, 183–193. doi: 10.1590/01000683rbcs20150666
Modesto, V. C., Andreotti, M., Sabbag, O. J., Soares, D. A, Pechoto, E. A. P, Pascoaloto, I. M, and Nakao, A. M. (2019). Economic analysis of the corn intercropped with Marandu grass as a function of Azospirillum brasilense application. J. Agric. Sci. 11, 387–395. doi: 10.5539/jas.v11n3p387
Moreira, R. C., Valadão, F. C. A., and Valadão, D. D. (2019). Agronomic performance of corn as a function of inoculation with Azospirillum brasilense and nitrogen fertilization. Rev. Ciênc. Agr. 62, 1–10. doi: 10.22491/rca.2019.2865
Mumbach, G. L., Kotowski, F. L. A., Schneider, M. S., Mallmann, É. B., Bonfada, V. O., Portela, E. B., et al. (2017). Response of inoculation with Azospirillum brasilense in wheat and corn of second crop. Rev. Sci. Agr. 2, 97–103. doi: 10.5380/rsa.v18i2.51475
Ngumbi, E., and Kloepper, J. (2016). Bacterial-mediated drought tolerance: current and future prospects. Appl. Soil Ecol. 105, 109–125. doi: 10.1016/j.apsoil.2016.04.009
Pariz, C. M., Andreotti, M., Azenha, M. V., Bergamaschine, A. F., Mello, L. M. M., and Lima, R. C. (2011). Corn grain yield and dry mass of Brachiaria intercrops in the crop-livestock integration system. Ciênc. Rural 5, 875–882. doi: 10.1590/S0103-84782011000500023
Pariz, C. M., Andreotti, M., Tarsitano, M. A. A., Bergamaschine, A. F., Buzetti, S., and Chioderolli, C. A. (2009). Technical and economic performance of corn intercropped with panicum and brachiaria forage in crop-livestock integration system. Pesq. Agropec. Trop. 39, 360–370. doi: 10.5216/pat.v39i4.5651
Pariz, C. M., Costa, N. R., Costa, C., Crusciol, C. A. C., de Castilhos, A. M., Meirelles, P. R. D. L., et al. (2020). An innovative corn to silage- grass-legume intercropping system with oversown black oat and soybean to silage in succession for the improvement of nutrient cycling. Front. Sustain. Food Syst. 4:544996. doi: 10.3389/fsufs.2020.544996
Per, T. S., Khan, N. A., Rddy, O. S., Masood, A., Hasanuzzaman, M., Khan, M. I. R., et al. (2017). Aproaches in modulationg proline metabolismo in plants for salt and drought stress tolerance: phytohormones, mineral, nutrients, and transgenics. Plant Physiol. Biochem. 115, 126–140. doi: 10.1016/j.plaphy.2017.03.018
Queiroz, R. F. D., Chioderoli, C. A., Furlani, C. E. A., Holanda, H. V. D., and Zerbato, C. (2016). Corn intercropped with urochloa ruziziensis under no-tillage system. Pesq. Agropec. Trop. 3, 238–244. doi: 10.1590/1983-40632016v4640464
R Core Team (2020). R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online at: https://www.R-project.org/ (accessed June 26, 2020).
Raij, B., Van Andrade, J. C., Cantarella, H., and Quaggio, J. Á. (2001). Análise quí mica para avaliação da fertilidade de solos tropicais. Campinas: Instituto Agronômico, pp. 284.
Rejeb, B. K., Abdelly, C., and Savouré, A. (2014). How reactive oxygen species and proline face stress together. Plant Physiol. Biochem. 80, 278–284. doi: 10.1016/j.plaphy,.2014.04.007
Sá, J. C. M., Séguy, L., Tivet, F., Lal, R., Bouzinac, S., Borszowskei, P. R., et al. (2015). Carbon depletion by plowing and its restoration by no-till cropping systems in Oxisols of subtropical and tropical agro-ecoregions in Brazil. Land Degrad. Dev. 26, 531–543. doi: 10.1002/ldr.2218
Santos, M. S., Nogueira, M. A., and Hungria, M. (2019). Microbial inoculants: reviewing the past, discussing the present and previewing an outstanding future for the use of beneficial bacteria in agriculture. AMB Expr. 9:205. doi: 10.1186/s13568-019-0932-0
Silva, A. G., Duarte, A. P., Piedade, R. D. C., Costa, H. P., Meireles, K. G. C., and Borges, L. P. (2015). Seed inoculation in off-season corn with Azospirillum and nitrogen topdressing aplication. Rev. Bras. Mil. Sor. 3, 358–370. doi: 10.18512/1980-6477/rbms.v14n3p358-370
Silva, F. A., Freitas, F. C. L., Rocha, P. R. R., Cunha, J. L. X. L., Dombroski, J. L. D., Holanda, M. E. C., et al. (2015). Growing corn for silage in no-tillage and conventional systems under differents periods of hydric defict. Semina 1:340. doi: 10.5433/1679-0359.2015v36n1p327
Vogel, G., Martinkoski, L., and Ruzicki, M. (2013). Effects of Azospirillum brasilense in use of forage grasses: amounts and results. Agropec. Cient Sem. 1, 01–06. doi: 10.30969/acsa.v10i1.471
Vogel, G. F., Martinkoskil, L., Martins, P. J., and Bichel, A. (2013). Agronomic performance of Azospirillum brasilense in rice culture: a review. Rev. Agron. Medio Amb. 3, 567–578.
Keywords: intercropping, no-tillage system, plant growth-promoting bacteria, Urochloa brizantha, Zea mays
Citation: Modesto VC, Andreotti M, Nakao AH, Soares DdA, Froio LdL, Dickmann L, Pascoaloto IM and Fernandes IMDM (2021) Yield and Production Components of Corn Under Straw of Marandu Palisade Grass Inoculated With Azospirillum brasilense in the Low-Land Cerrado. Front. Sustain. Food Syst. 4:617065. doi: 10.3389/fsufs.2020.617065
Received: 13 October 2020; Accepted: 11 December 2020;
Published: 12 February 2021.
Edited by:
Everlon Cid Rigobelo, Universidade Estadual Paulista, BrazilReviewed by:
Muhammad Ali Raza, Sichuan Agricultural University, ChinaCopyright © 2021 Modesto, Andreotti, Nakao, Soares, Froio, Dickmann, Pascoaloto and Fernandes. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Viviane Cristina Modesto, dml2aWFuZS5tb2Rlc3RvQHVuZXNwLmJy
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.