
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Sustain. Food Syst. , 18 December 2020
Sec. Crop Biology and Sustainability
Volume 4 - 2020 | https://doi.org/10.3389/fsufs.2020.606827
This article is part of the Research Topic Plant Growth-Promoting Microorganisms for Sustainable Agricultural Production View all 50 articles
 Neilton Antonio Fiusa Araújo1
Neilton Antonio Fiusa Araújo1 Rafaela Magalhães Brandão2Beatriz Meireles Barguil3Maria das Graças Cardoso2Moacir Pasqual1Renata Alves Lara Silva Rezende1Maysa Mathias Alves Pereira1Victor Hugo Teixeira Buttrós1
Rafaela Magalhães Brandão2Beatriz Meireles Barguil3Maria das Graças Cardoso2Moacir Pasqual1Renata Alves Lara Silva Rezende1Maysa Mathias Alves Pereira1Victor Hugo Teixeira Buttrós1 Joyce Dória1*
Joyce Dória1*Rose essential oil is rich in compounds widely used by the pharmaceutical and cosmetic industry, due to the biological activities it presents. However, obtaining oil is costly, as the yield per plant is low, which requires several techniques that aim to increase its production. The application of growth-promoting bacteria has been studied for this purpose. Thus, the objective of this work was to select efficient bacteria for production and evaluate their influence on the phytotechnical characteristics and composition of the essential oils of roses. Seven species of bacteria were evaluated for the potential to promote growth in vitro, being tested for nitrogen fixation, phosphate solubilization, protease production and auxin production. From bacteria tested, four were selected and inoculated on rose plants of cultivar Black Prince to evaluate the influence on phytotechnical variables of flower and stem and the oil production. The evaluation of the production of roses was performed through the characteristics of the flowers (size, weight, and diameter of the stem) and floral bud. The essential oils from the inoculated flowers were extracted and evaluated in terms of content, yield, and chemical composition. The application of B. acidiceler, B. subtilis and B. pumilus resulted in flowers with a diameter up to 29% larger. The floral stem was increased by up to 24.5% when B. acidiceler and B. pumilus were used. Meanwhile, the stem diameter was around 41% greater in the presence of B. acidiceler, B. subtilis and in the control. Bacillus pumilus also increased the weight of fresh petals (104%) and essential oil yield (26%), changing the chemical composition of the extracted essential oil. Thus, it is concluded that B. acidiceler, B. pumilus, and B. subtilis improved the phytotechnical characteristics of roses. Among bacteria, B. pumilus increased the essential oil content as well as positively changed the chemical composition of the extracted essential oil.
Rose (Rosa hybrida L. - Rosaceae) is one of the three main flowering plants in the world. Currently, there are approximately 35,000 cultivars bred throughout the world (Bendahmane et al., 2013; Kirov et al., 2014) and around 200 species cataloged (Fougère-Danezan et al., 2015). Roses are cultivated around the world for different purposes, from ornamentation to the manufacture of processed products such as perfumes, medicines, and food using bioactive molecules found in their flowers (Gil et al., 2019). Among these molecules, the essential oils (EO) are one of the most researched in roses. EO are secondary metabolites extracted from plants. They are commonly used in the food, cosmetic and pharmaceutical industries, due to their several biological activities, such as antimicrobial, analgesic, antiseptic, carminative, and diuretic; in many cases, EO also show pleasant diverse aromas (Calo et al., 2015; Basak and Guha, 2018; Sarkic and Stappen, 2018).
Brazil has a prominent place in the production of EO, alongside India, China and Indonesia, mainly due to the production of EO extracted from citrus fruits as by-products of the juice industry (Bizzo et al., 2009). EO of roses is rich in terpenic and sesquiterpenic compounds, which are widely explored by the pharmaceutical and cosmetic industries, mainly for their antioxidant capacity (Patrascu and Radoiu, 2016). Other compounds can also be found in other species of roses. In Rosa roxburghii, the EO presents significant amounts of linoleic acid, a powerful poly-unsaturated fatty acid possessing manifold health effects (Yang et al., 2020). The same oil also contains 2′-methylenebis (6-tert-butyl-4-methylphenol) considered a popular antioxidant and anti-aging agent. Phenols and flavonoids are compounds found in Rosa damascena, which presents antioxidant and free radical activity (Nikolova et al., 2019).
Worldwide, rose oil production is based on several species, such as Rosa damascena, Rosa gallica, Rosa centifolia, and Rosa alba L., the first two species being the most used in oil extraction (Kovatcheva et al., 2011). Among the several hybrid cultivars, the rose cv. Black Prince (also known as rose “Barcarolle”) (Avdic et al., 2016) is very widespread throughout Brazil due to its rusticity and beauty. It has medium productivity, high architecture and elongated buttons (Barbosa, 2008). However, there are no reports of oil production of this cultivar, which makes this fact one of the main attractions of the present study.
In Brazil, the production of roses occupies the first position among the cut flowers, corresponding to 30% of the national market (Junqueira and Peetz, 2017). However, one of the problems faced for the acquisition of EO is the low yield per plant, being necessary the use of extraction techniques that increase the yield per flower, or the use of different managements during the cultivation that stimulate the plant production (Pirbalouti et al., 2013).
A new strategy that is being investigated is the use of efficient microorganisms aimed at increasing these molecules of interest. These microorganisms, also known as plant growth promoters (PGP), are used for the most diverse purposes and are capable of providing several benefits (Vejan et al., 2016). In agriculture, the use of PGP has influenced the growth and development of plants, stimulating increased production, aiding in the control of pests and diseases, and activating enzymes capable of producing compounds that improve the quality of products obtained by plants inoculated with them (Saharan and Nehra, 2011; Numan et al., 2018). PGPs can also influence the production of secondary compounds by plants, since many of them have the ability to produce hormones such as auxins, cytokinins and gibberellins or even improve the absorption of nutrients such as nitrogen and phosphorus, which directly affects production and quality of EO from plants (Glick, 2012; Younis et al., 2015).
Bacteria are the most commonly used efficient microorganisms. Studies indicate that bacteria are able to assist plants in their development as well as stimulate the production of compounds that assist in plant defense, such as the production of secondary metabolites responsible for the plant defense system to external factors, which can influence the chemical composition of its aromatic compounds, such as EO (Xie et al., 2019).
In view of the discussed, the objective of this study was to select growth-promoting bacteria for rose cultivation and evaluate their influence on the production of EO.
Seven species of bacteria (Bacillus acidiceler, Bacillus amyloliquefaciens, Bacillus macauenses, Bacillus subtilis, Bacillus pumilus, Pantoea ananatis, Staphylococcus equorum) were isolated from strawberry plants and also from the Crop Collection of Agricultural Microbiology (CCMA) of the Federal University of Lavras, as shown in Table 1.
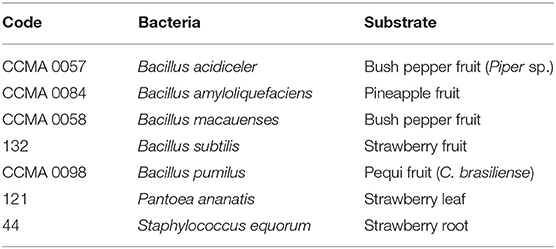
Table 1. Identification and origin of bacteria.
Protease production was determined by the method of Sgroy et al. (2009). Petri dishes with Skim Milk Agar (SMA) culture medium were inoculated with 10 μL of bacterial isolates previously cultivated in Luria-Bertami medium (LB) and incubated at 28°C. Halo formation around the colonies was observed after 24 h after inoculation.
Cultures of bacteria activated in Petri dishes with medium 523 (10 g sucrose, 8 g casein hydrolysate, 4 g yeast extract, 2 g K2HPO4, 0.3 g MgSO4.7H2O, 15 g agar, 1000 mL distillated water) for 24 h were used. Bacteria colonies were transferred to a micro tube containing 1 mL of nutrient broth added of 0.1 g L−1 of L-tryptophan; the material was maintained in agitation for 3 days in the dark. After this period, the micro tubes were centrifuged at 1500 rpm for 3 min and a 0.5 mL aliquot was removed, which was added of 0.5 mL of Salkowiski's reagent. The reaction was incubated for 15 min in the dark, and then a pink or reddish color was observed which signifies the presence of auxins (Kado and Heskett, 1990).
For this test, the bacteria were activated 24 h in advance. 10 μL of bacterial isolates grown in 523 liquid medium were inoculated in dishes containing the NBRIP medium (National Botanical Research Institute's Phosphate growth medium—India) (Nautiyal, 1999) which consists of glucose (10 g), Ca3(PO4)2 (5 g), MgCl2.6H2O (5 g), MgSO4.7H2O (0.25 g), KCl (0.2 g), (NH4)2SO4 (0.1 g), and agar (15 g). The material was incubated at 28°C with 12 h photoperiod. Halo formation was evaluated around the colonies after 24 h after inoculation.
This test was performed using 10 mL flasks containing 5 mL of semi-solid NFB medium, consisting of malic acid (5 g), K2HPO4 (0.5 g), MgSO4.7H2O (0.2 g), NaCl (0.1 g), CaCl2.2H2O (0.02 g), vitamin solution (1 ml), micronutrient solution (2 ml), FeEDTA (4 ml), bromothymol blue (2 ml), KOH (4.5 g), agar (2 g) and pH 6.8 (Döbereiner et al., 1995). Bacteria with 24 h of growth, grown in medium 523 were inoculated in the medium. An inoculation loop was used to collect bacteria and puncture the medium. Then, they were incubated at 28°C, making it possible to observe the halo formation in the upper side of the culture medium, indicating the nitrogen fixation by bacteria.
According to the growth-promoting tests, 4 (four) species of bacteria were selected and inoculated in roses of cultivar Black Prince to evaluate the bacterial effect on the productive characteristics of plants. To prepare the suspension, the bacteria were cultured for 48 h in nutrient broth and the concentration of each suspension was adjusted to 106 cells mL−1. For inoculation, the plants were pruned at 30 cm from the grafting site, removed from the pots and their roots washed until complete exposure. The inoculation in the plants was done by immersing the bare roots in the suspension for 1 h. After that time, the plants were planted in pots with a capacity of 10 L, containing commercial substrate Topstrato®.
The experiment was carried out in a greenhouse, in a randomized block design with 5 treatments (each selected bacteria plus a control that corresponded to plants inoculated with distilled water only) and 5 replicates. The experimental unit consisted of 4 plants per block.
The evaluations started at the first cutting of stems with flowers at the point of harvest (Figure 1). At that point, the floral buds is closed and it changes from a pointed shape to a cup shape. From the first to 5th day, the following phytotechnical characteristics were evaluated: diameter of the closed bud, diameter of the flower at the point of harvest, stem size, stem diameter, stem weight and total weight of petals per flower.

Figure 1. Flower bud opening stages. (A) Bud at the harvest point. (B) Completely open flower.
Rose EO was extracted from flowers at the point of harvest. The plants were acquired from a local producer (Roselanche Floriculture—Garden and Floriculture Brandão, Barbacena, Minas Gerais State, Brazil). The fresh petals were taken to an oven with air circulation for drying at 30°C. After drying, they were weighed and the EO was obtained using ~100 g of dry rose petals with distilled water in a round bottom flask. The plant material was subjected to the hydrodistillation process using the modified Clevenger apparatus for a period of 4 h (Brasil, 2010). Three extractions were performed. EO was separated from the hydrolate by a pentane partition process. The organic part was dried using anhydrous sodium sulfate, filtered and rotary evaporated to eliminate the solvent. EO without solvent residues was protected from light and refrigerated.
The chemical constituents of EO were characterized by gas chromatography coupled to a mass spectrometer (GC-MS, Shimadzu Corporation, model QP2010 Plus, Kyoto, Japan) according to Adams (2007). The following conditions were performed: fused silica capillary column with phase connected to DB5 (30 mx 0.25 mm id, film thickness 0.25 μm); He 5.0 (White Martins, Rio de Janeiro, Brazil) as carrier gas and flow rate of 1 mL min−1. The injector temperature was 220°C and the detector temperature was 240°C. The injection volume of the sample was 0.5 μL diluted in hexane (1%) (Sigma-Aldrich®, St. Louis, MO, USA) with a division ratio of 1:100. Programmed temperature of 60°C increasing to 240°C at a rate of 3°C per min and from 240°C to 300°C at 10°C per min. The final temperature was maintained for 7 min. The operational parameters of the GC-MS followed the conditions: ionization potential (70 eV), ion source temperature (200°C), scan speed of 1000 Da sec−1, scan interval of 0.50 fragments sec−1. Mass analyzes were performed in full scan mode, ranging from 45 to 500 Da. The data regarding chemical constituents were acquired using LabSolutions LC/GC Workstation 2.72. The Van Den Dool and Kratz (1963) equation was used to calculate the retention index (IR = 100n + 100 [(tR (i)–tR (n))/(tR (n + 1) n), where tR (i), tR (n) and tR (n + 1) correspond to the retention times of the tested compounds and standards, respectively. The standards used were the homologous series of n-alkanes (nC8-nC18). The identification of compounds was based on comparison with the retention indices with those of the literature (Adams, 2007) and the mass spectra of the EO constituents with up to 95% similarity were compared with those of the mass spectra libraries FFNSC 1.2, NIST 107, and NIST 21. The quantification of volatile compounds was determined by gas chromatography with a flame ionization detector (DIC) (Shimadzu GC - 2010, Kyoto, Japan). The experimental parameters were the same used by GC-MS, with the exception of the detector temperature, which was 300°C. The percentages of compounds were calculated using the area normalization method.
Due the low amount of extracted oil, the amount was estimated using the difference in weight of the bottle before and after the extraction of the EO. For this purpose, the amber bottles were weighed before extraction and at the end of the extraction process, after the complete evaporation of hexane at room temperature. The weight difference determined the amount of oil extracted. With these values, the yield for each treatment was determined and the content of EO was calculated according to the following formula proposed by Santos et al. (2004): Oil content (%) = [Weight of oil (g) x Dry petal biomass (g)]/100. The yield of EO was expressed in weight of oil per unit weight of plant material in Moisture Free Base (% w/w MFB).
The data were tested for normality and subsequently submitted to ANOVA. In case of significance, the means were compared by the Scott-Knott test (P ≤ 0.05), using the statistical software SISVAR version 5.2 (Ferreira, 2014).
The results of the growth promotion tests showed that the strains of Bacillus acidiceler, B. subtilis, B. pumilus and Staphylococcus equorum were efficient in the phosphate solubilization, protease production and nitrogen fixation tests (Table 2). The B. amyloliquefacies strain was efficient for phosphate solubilization and protease production and the B. macauenses and Pantoea ananatis strains were efficient only for phosphate solubilization and protease production, respectively. None of the strains of bacteria used showed the ability to produce auxins by the test evaluated.
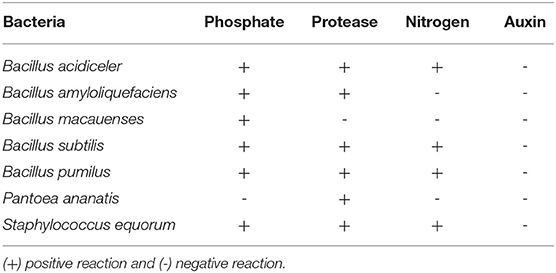
Table 2. Production test of biochemical compounds by bacteria, evaluation of phosphate solubilization, protease production, biological nitrogen fixation, and auxin production.
Phosphate solubilization is a mechanism common to many microorganisms that are associated with plant roots. This solubilization capacity may be related to the decrease in the pH of the medium due to the production of organic acids (such as gluconic acid, oxalic acid, malonic acid, acetic acid, formic acid, citric acid, and succinic acid), which can dissolve the phosphate mineral, as a result of anion exchange, or chelate ions associated with phosphate (Chagas Junior et al., 2010; Tenga et al., 2019). Acidification of the medium is correlated with phosphorus solubilization by acid phosphatase activity, indicating that some microorganisms have developed enzymes capable of releasing inorganic phosphate from organophosphate complexes, in addition to the activity of this enzyme proving to be a good indicator for the mineralization of organic phosphorus (Kumar, 2016; Tenga et al., 2019). Likewise, the capacity for nitrogen fixation is common to many groups of microorganisms and their association due to its beneficial potential has been studied for application in the most varied agricultural crops (Mondani et al., 2019; Schmidt et al., 2019).
Regarding the production of roses, the application of bacteria did not influence the size of the bud before the flower opened (Table 3). However, plants treated with B. acidiceler, B. subtilis, and B. pumilus had higher diameter of open flower. An increase in the weight of the petals of plants treated with B. pumilus was also observed, showing a positive effect of this bacterium in relation to the others evaluated.
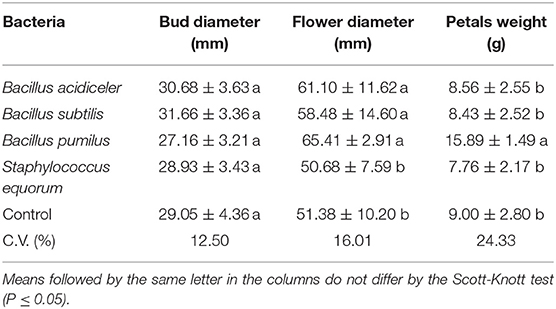
Table 3. Effect of growth-promoting bacteria on the bud diameter, flower diameter and weight of rose petals of the cultivar Black Prince.
Some growth-promoting bacteria can interfere with the production of hormones by plants, which can affect the structure of the roots, increasing their surface area, number and type (Ali et al., 2010; Saharan and Nehra, 2011). In addition to these factors, inoculation with endophytic bacteria can promote increased photosynthesis inducing greater production of carbohydrates by plants (Olanrewaju et al., 2017). In roses, the accumulation of carbohydrates in the flower stems can favor the durability of the flowers in the post-harvest. Thus, the application of endophytic microorganisms that aid in the accumulation of carbohydrates in the flower stems can improve the final quality of the roses, leading to greater durability of the cut stems (Moubayidin et al., 2009; Wang et al., 2014, 2019; Castanheira et al., 2017).
Plants treated with B. acidiceler and B. pumilus had a larger stem size (Table 4). The diameter of the stems was smaller in plants treated with S. equorum and B. pumilus, but only plants treated with S. equorum had lower weight of flower stems.
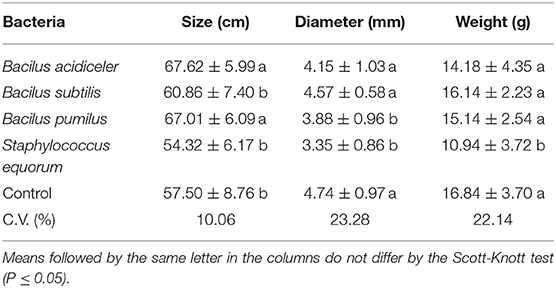
Table 4. Effect of growth-promoting bacteria on size, diameter and weight of stem of roses cv. Black Prince.
The yield of oil extracted from roses treated with B. pumilus was superior to other bacteria. However, a negative effect was observed on the yield of plants treated with B. acidiceler, in which the oil content was also lower in plants treated with this bacterium, with no difference in oil content between the plants treated with the other bacteria and the control (Table 5). However, it was not possible to obtain enough EO to perform the analysis on roses treated with the S. equorum bacterium through the methodology used for extraction.
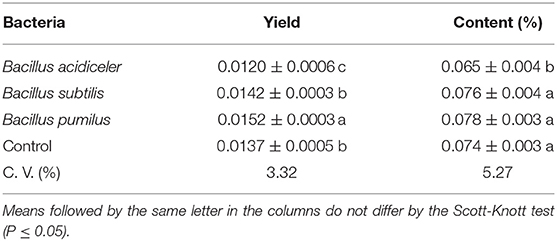
Table 5. Effect of growth-promoting bacteria on yield and content of essential oil of roses cv. Black Prince.
The drying of the material can lead to decreases in the content of EOs and changes in its composition. Although freeze-drying is indicated due to less loss for EO amount, some species may have higher content of EOs when subjected to a convective drying process (Pirbalouti et al., 2013; Roslon et al., 2016).
In addition to phosphate solubilization and nitrogen fixation, growth-promoting microorganisms may be able to produce phytohormones, siderophores and antibiotics, and they are also capable of inducing systemic resistance in the plant (Sgroy et al., 2009). In addition, plants can also interact with these organisms by increasing their growth, since the increase in the amount of EOs is often related to the growth of plant organs due to greater absorption of nutrients; the quality of the oils can also be influenced by the activation of metabolic routes due to the interaction of the plant with microorganisms (Maji et al., 2013; Singh et al., 2019).
B. subtilis strains have the potential to stimulate plant biomass production even under adverse field conditions, such as in saline soils, and to increase chlorophyll production and soluble protein content (Wang et al., 2018). In addition, the application of B. subtilis strains positively influenced the growth and production of crops such as cotton, brachypodium grass (Brachypodium distachyon), arabidopsis (Arabidopsis thaliana) and wheat (Triticum aestivum) (Ali, 2015; Gagné-Bourque et al., 2015; Wang et al., 2018; Andrees et al., 2019).
For the chemical composition of EO of roses, there was a significant difference according to each bacterium used in the treatment of plants (Table 6, Figure 2). The smallest number of components was found in the oils of plants treated with B. acidiceler. However, plants treated with B. pumilus and B. subtilis had higher number of compounds, in which the chemical constituents of carophyllene oxide, α-selinene, limonene, α-farnecene appeared only in plants treated with B. pumilus and teaspiran only in flowers treated with B. subtilis.
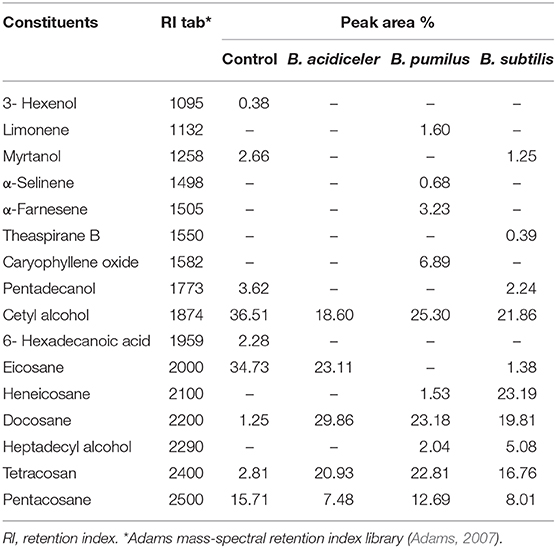
Table 6. Effect of growth-promoting bacteria on chemical composition of essential oil of roses cv. Black Prince extracted by hydro-distillation.
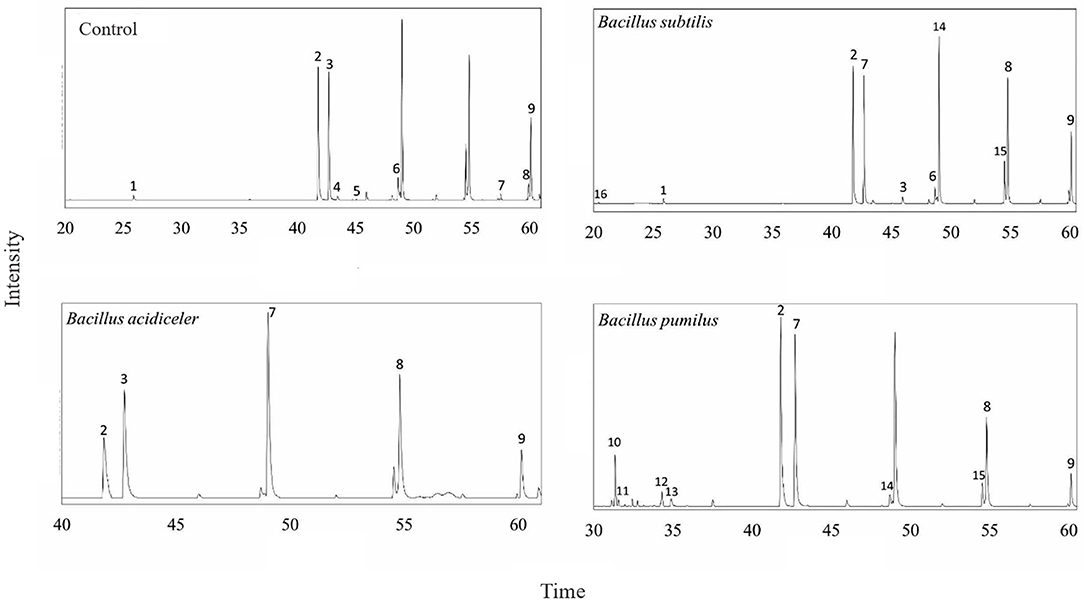
Figure 2. Chromatography of essential oils of rose cv. Black Prince relating the retention time and intensity of each component, according to each growth-promoting bacteria, being: (1) Myrtanol, (2) Cetyl alcohol, (3) Eicosane, (4) Hexadecanoic acid, (5) Hexenol, (6) Pentadecanol, (7) Docosane, (8) Tetracosane, (9) Pentacosane, (10) Caryophyllene, (11) α-Selinene, (12) Limonene, (13) α-Farsene, (14) Heneicosane, (15) Heptadecyl alcohol, (16) Theaspirane.
Limonene is related to the strong smell of orange, which constitutes the final citrus aspect of the aroma of the species under study. This compound has an insecticidal action and can confer resistance to insects in some plants, being widely used in flavor and fragrance industries (Thomas and Bessière, 1989; Sowndhararajan et al., 2015). A-selinene, on the other hand, is a sesquiterpene that is not part of the smell composition of the oils where it is present, but it is part of the chemical composition of the EO of several plants such as guamirim (Calyptranthes concinna), citronella (Cymbopongon nardus), and tiririca (Cyperus rotundus) (Limberger et al., 2002; Lawal and Oyedeji, 2009; Silva et al., 2011).
A-farnesene is a sesquiterpene related to the defense of plants to aphids, in addition to conferring the flavor of some vegetables, being found naturally coating fruits such as apples and pears. In EOs, this compound is also related to the smell of floral-green or woody, being one of the major constituents of ginger oil (Harbone, 1997; Andrade et al., 2012).
Similar to α-farnesene, carophyllene oxide is also a major constituent of EOs from some plants that may contain toxic compounds. This compound has the potential to inhibit the growth of Gram-positive bacteria, and is also toxic to ants and fungi associated with them. In addition, the carophylene oxide found in EOs has already been associated with analgesic and anti-inflammatory action and can induce apoptosis in lymphoma and neuroblastoma cells (Norouzi-Arasi et al., 2006; Judzentienea et al., 2010; Sain et al., 2014). The presence of this compound may indicate an improvement in the medicinal properties of rose oil, as well as influencing their defense against leaf-cutting ants.
The alcohols and esters are the main constituents found in the EO of several varieties and species of roses, configuring more than 80% of the key components of its aromas in some of those plants, where the alcohols are the components used in the evaluation of the quality of the EO of roses (Xiao et al., 2018).
The compounds myrtanol, cetyl alcohol, eicosane, hexadecanoic acid, hexenol, docosane, tetracosan, pentacosane, heneicosane, and heptadecyl alcohol are part of the volatile compounds of EO of rose species intended for the extraction of EOs. However, its presence and quantity in the oil can vary depending on the management, environmental conditions, methodology and time of extraction (Özel et al., 2004; Joichi et al., 2005; Xiao et al., 2018).
The promotion of plant growth has also been proven in strains of B. pumilus, showing its efficiency in the growth, development and production of plants, as well as in resistance to some environmental stresses, such as salinization, drought, and tolerance to heavy metals. Those strains also reduce reactive oxygen species, increasing the content of secondary metabolites and increasing the activity of antioxidant enzymes (Khan et al., 2016; Sirajuddin et al., 2016; Xie et al., 2019). The potential of these microorganisms to stimulate the production and accumulation of secondary metabolites may explain the increase in the EO content in roses and the different compounds present in the chemical composition of the oil extracted from roses inoculated with B. pumilus.
B. acidiceler, B. pumilus and B. subtilis improved the phytotechnical characteristics of roses. Among bacteria, B. pumilus increased the essential oil content as well as positively changed the chemical composition of the extracted essential oil from roses cv. Black Prince.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
NA and JD: research design, experiments administration, supervision, and writing. RB and MMAP: experiments conduction (bacterial analysis) and data discussion. BB, MC, and VB: experiments conduction (rose analysis) and data discussion. MP: visualization and data discussion. RR: writing and critical revision. All authors read and approved the final manuscript.
This work was supported by the National Council for Scientific and Technological Development (CNPq - Brazil).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors would like to thank the National Council for Scientific and Technological Development (CNPQ/Brazil) for scholarships and financial support and also to the Electronic Microscopy Laboratory, the Organic Chemistry Laboratory/Essential Oils, and the Chemical Analysis and Prospecting Center of the Federal University of Lavras for providing the equipment for the analyzes.
Adams, R. P. (2007). Identification of Essential Oils Components by Gas Chromatography/Mass Spectroscopy. Carol Stream, IL: Allured.
Ali, B. (2015). Bacterial auxin signaling: comparative study of growth induction in Arabidopsis thaliana and Triticum aestivum. Turk. J. Bot. 39, 1–9. doi: 10.3906/bot-1401-31
Ali, B., Sabri, A. N., and Hasnain, S. (2010). Rhizobacterial potential to alter auxin content and growth of Vigna radiata (L.). World J. Microbiol. Biotechnol. 26, 1379–1384. doi: 10.1007/s11274-010-0310-1
Andrade, M. A., Cardoso, M. G., Batista, L. R., Mallet, A. C. T., and Machado, S. M. F. (2012). Essential oils of Cinnamomum zeylanicum, Cymbopogon nardus and Zingiber officinale: composition, antioxidant and antibacterial activities. Rev. Ciênc. Agron. 43, 399–408. doi: 10.1590/S1806-66902012000200025
Andrees, H., Haider, M. S., Anjum, T., and Akram, W. (2019). Inducing systemic resistance in cotton plants against charcoal root rot pathogen using indigenous rhizospheric bacterial strains and chemical elicitors. Crop. Prot. 115, 75–83. doi: 10.1016/j.cropro.2018.09.011
Avdic, J., Becic, B., Sarajlic, N., and Arar, K. (2016). Roses (Rosa spp.) in public green spaces of Sarajevo. Food Sci. 61, 209–212.
Basak, S., and Guha, P. (2018). A review on antifungal activity and mode of action of essential oils and their delivery as nano-sized oil droplets in food system. J. Food Sci. Technol. 55, 4701–4710. doi: 10.1007/s13197-018-3394-5
Bendahmane, M., Dubois, A., Raymond, O., and Le Bris, M. (2013). Genetics and genomics of fower initiation and development in roses. J. Exp. Bot. 64, 847–857. doi: 10.1093/jxb/ers387
Bizzo, H. R., Hovell, A. M. C., and Rezende, C. M. (2009). Brazilian essential oils: general view, developments and perspectives. Química Nova 32, 588–594. doi: 10.1590/S0100-40422009000300005
Brasil. (2010). Farmacopeia Brasileira, Vol. 2. Brasília: Agência Nacional de Vigilância Sanitária (ANVISA), 546.
Calo, J. R., Crandall, P. G., O'Bryan, C. A., and Ricke, S. C. (2015). Essential oils as antimicrobials in food systems—A review. Food Control 54, 111–119. doi: 10.1016/j.foodcont.2014.12.040
Castanheira, N. L., Dourado, A. C., Pais, I., Semedo, J., Scotti-Campos, P., Borges, N., et al. (2017). Colonization and beneficial effects on annual ryegrass by mixed inoculation with plant growth promoting bacteria. Microbiol. Res. 198, 47–55. doi: 10.1016/j.micres.2017.01.009
Chagas Junior, A. F., Oliveira, L. A., Oliveira, A. N., and Willerding, A. L. (2010). Capacidade de solubilização de fosfatos e eficiência simbiótica de rizóbios isolados de solos da Amazônia. Acta Scient. Agron. 32, 359–366. doi: 10.4025/actasciagron.v32i2.3185
Döbereiner, J., Baldani, V. L. D., and Baldani, J. I. (1995). Como Isolar e Identificar Bactérias Diazotróficas de Plantas Não-Leguminosas. Brasília: EMBRAPA-SPI.
Ferreira, D. F. (2014). Sisvar: a guide for bootstrap procedures in multiple comparisons. Cienc Agrotec. 38, 109–112. doi: 10.1590/S1413-70542014000200001
Fougère-Danezan, M., Joly, S., Bruneau, A., Gao, X. F., and Zhang, L. B. (2015). Phylogeny and biogeography of wild roses with specific attention to polyploids. Ann. Bot. 115, 275–291. doi: 10.1093/aob/mcu245
Gagné-Bourque, F., Mayer, B. F., Charron, J. B., Vali, H., and Bertrand, A. (2015). Accelerated growth rate and increased drought stress resilience of the model grass Brachypodium distachyon colonized by Bacillus subtilis B26. PLoS ONE 10:e130456. doi: 10.1371/journal.pone.0130456
Gil, C. S., Lim, S. T., Lim, Y. J., Jung, K. H., Na, J. K., and Eom, S. H. (2019). Volatile content variation in the petals of cut roses during vase life. Sci. Hortic. 261, 1–6. doi: 10.1016/j.scienta.2019.108960
Glick, B. R. (2012). Plant growth-promoting bacteria: mechanisms and applications. Scientifica 2012:15. doi: 10.6064/2012/963401
Joichi, A., Yomogida, K., Awano, K., and Ueda, Y. (2005). Volatile components of tea-scented modern roses and ancient Chinese roses. Flavour Fragr. J. 20, 152–157. doi: 10.1002/ffj.1388
Judzentienea, A., Budienea, J., Butkienea, R., Kupcinskieneb, E., Laffont-Schwobc, I., and Masottic, V. (2010). Caryophyllene oxide-rich essential oils of Lithuanian Artemisia campestris ssp. campestris and their toxicity. Nat. Prod. Commun. 5:1981–1984. doi: 10.1177/1934578X1000501232
Junqueira, A. H., and Peetz, M. S. (2017). Brazilian consumption of flowers and ornamental plants: habits, practices and trends. Ornam. Hortic. 23, 178–184. doi: 10.14295/oh.v23i2.1070
Kado, C. I., and Heskett, M. G. (1990). Selective media for isolation of Agrobacterium, Corynebacterium, Erwinia, Pseudomonas and Xanthomonas. Phytopathology 60, 969–976.
Khan, A. X. Q., Zhao, S., Javed, M. T., Khan, K. S., Bano, A., Shen, R. F., et al. (2016). Bacillus pumilus enhances tolerance in rice (Oryza sativa, L.) to combined stresses of NaCl and high boron due to limited uptake of Na+. Environ. Exp. Bot. 124, 120–129. doi: 10.1016/j.envexpbot.2015.12.011
Kirov, I., Van Laere, K., Riek, J., Keyser, E., Van Roy, N., and Khrustaleva, L. (2014). Anchoring linkage groups of the Rosa genetic map to physical chromosomes with tyramide-FISH and EST-SNP markers. PLoS ONE 9:e95793. doi: 10.1371/journal.pone.0095793
Kovatcheva, N., Zheljazkov, V. D., and Astatkie, T. (2011). Productivity, oil content, composition, and bioactivity of oil-bearing rose accessions. HortSci. 46, 710–714. doi: 10.21273/HORTSCI.46.5.710
Kumar, A. (2016). Phosphate solubilizing bacteria in agriculture biotechnology: diversity, mechanism and their role in plant growth and crop yield. Int. J. Adv. Res. 4, 116–124. doi: 10.21474/IJAR01/111
Lawal, O. A., and Oyedeji, A. O. (2009). Chemical composition of the essential oils of Cyperus rotundus L. from South Africa. Molecules 14, 2909–2917. doi: 10.3390/molecules14082909
Limberger, R. P., Simões-Pires, C. A., Sobral, M., Menut, C., Bessiere, J. M., and Henriques, A. T. (2002). Essential oils from Calyptranthes concinna, C. lucida and C. rubella (Myrtaceae). Braz. J. Pharm. Sci. 38:11. doi: 10.1590/S1516-93322002000300011
Maji, D., Barnawal, D., Gupta, A., King, S., Singh, A. K., and Kalra, A. (2013). A natural plant growth promoter calliterpenone from a plant Callicarpa macrophylla Vahl improves the plant growth promoting effects of plant growth promoting rhizobacteria (PGPRs). World J. Microbiol. Biotechnol. 29, 833–839. doi: 10.1007/s11274-012-1238-4
Mondani, F., Khani, K., Honarmand, J. S., and Saeidi, M. (2019). Evaluating effects of plant growth-promoting rhizobacteria on the radiation use efficiency and yield of soybean (glycine max) under water deficit stress condition. Agric. Water Manag. 213, 707–713. doi: 10.1016/j.agwat.2018.11.004
Moubayidin, L., Dimambro, R., and Sabatini, S. (2009). Cytokinin-auxin crosstalk. Trends in Plant Sci. 14, 557–562. doi: 10.1016/j.tplants.2009.06.010
Nautiyal, C. S. (1999). An efficient microbiological growth medium for screening phosphate solubilizing microorganisms. FEMS Microbiol. Lett. 170, 265–270. doi: 10.1111/j.1574-6968.1999.tb13383.x
Nikolova, G., Karamalakova, Y., and Gadjeva, V. (2019). Reducing oxidative toxicity of L-dopa in combination with two different antioxidants: an essential oil isolated from Rosa damascena Mill., and vitamin C. Toxicol. Rep. 6, 267–271. doi: 10.1016/j.toxrep.2019.03.006
Norouzi-Arasi, H., Yavari, I., Chalabian, F., Kiarostami, V., Ghaffarzadeh, F., and Nasirian, A. (2006). Chemical constituents and antimicrobial activities of the essential oil of Acroptilon repens (L.) DC. Flavour Fragr. J. 21, 247–249. doi: 10.1002/ffj.1568
Numan, M., Bashir, S., Khan, Y., Mumtaz, R., Shinwari, Z. K., Khan, A. L., et al. (2018). Plant growth promoting bacteria as an alternative strategy for salt tolerance in plants: a review. Microbiol. Res. 209, 21–32. doi: 10.1016/j.micres.2018.02.003
Olanrewaju, O. S., Glick, B. R., and Babalola, O. O. (2017). Mechanisms of action of plant growth promoting bacteria. World J. Microbiol. Biotechnol. 33, 1–16. doi: 10.1007/s11274-017-2364-9
Özel, M. Z., and Anthony, A., and Clifford, A. A. (2004). Superheated water extraction of fragrance compounds from Rosa canina. Flavour Fragr. J. 19, 354–359. doi: 10.1002/ffj.1317
Patrascu, M., and Radoiu, M. (2016). Rose essential oil extraction from fresh petals using synergetic microwave & ultrasound energy: chemical composition and antioxidant activity assessment. J. Chem. Chem. Eng. 10, 136–142. doi: 10.17265/1934-7375/2016.03.004
Pirbalouti, A. G., Oraie, M., Pouriamehr, M., and Babadi, E. S. (2013). Effects of drying methods on qualitative and quantitative of the essential oil of Bakhtiari savory (Satureja bachtiarica Bunge.). Ind. Crops Prod. 46, 324–327. doi: 10.1016/j.indcrop.2013.02.014
Roslon, W., Wajs-Bonikowska, A., Geszprych, A., and Osinska, E. (2016). Characteristics of essential oil from young shoots of garden angelica (Angelica archangelica L.). J. Essent. Oil Bear. Plants 19, 1462–1470. doi: 10.1080/0972060X.2016.1238322
Saharan, B. S., and Nehra, V. (2011). Plant growth promoting rhizobacteria: a critical review. Life Sci. Medicine Res. 2011, 1–30.
Sain, S., Naoghare, P. K., Devi, S. S., Daiwile, A., Krishnamurthi, K., Arrigo, P., et al. (2014). Beta caryophyllene and caryophyllene oxide, isolated from Aegle marmelos, as the potent anti-inflammatory agents against lymphoma and neuroblastoma cells. Antiinflamm. Antiallergy Agents Med. Chem. 13, 45–55. doi: 10.2174/18715230113129990016
Santos, A. S., Alves, S. M., Figueiredo, F. J. C., and Rocha Neto, O. G. (2004). Descrição de Sistema e Métodos de Extração de Óleos Essenciais e Determinação de Umidade de Biomassa em Laboratório. Comunicado Técnico-Embrapa Belém: Embrapa Amzônia Oriental.
Sarkic, A., and Stappen, I. (2018). Essential oils and their single compounds in cosmetics - a critical review. Cosmetics 5, 1–21. doi: 10.3390/cosmetics5010011
Schmidt, T. M., Thomé, A. H. E., Sperotto, R. A., and Granada, C. E. (2019). Effect of rhizobia inoculation on the development of soil-borne pathogens infecting common bean plants. Europ. J. Plant Pathol. 153, 687–694. doi: 10.1007/s10658-018-1600-y
Sgroy, V., Cassán, F., Masciarelli, O., Del Papa, M. F., Lagares, A., and Luna, V. (2009). Isolation and characterization of endophytic plant growth-promoting (PGPB) or stress homeostasis-regulating (PSHB) bacteria associated to the halophyte Prosopis strombulifera. Appl. Microbiol. Biotechnol. 85, 371–381. doi: 10.1007/s00253-009-2116-3
Silva, C. F., Moura, F. C., Mendes, M. F., and Pessoa, F. L. P. (2011). Extraction of citronella (Cymbopogon nardus) essential oil using supercritical CO2: experimental data and mathematical modeling. Braz. J. Chem. Eng. 28, 343–350. doi: 10.1590/S0104-66322011000200019
Singh, S., Tripathia, A., Majia, D., Awasthia, A., Vajpayeeb, P., and Kalraa, A. (2019). Evaluating the potential of combined inoculation of Trichoderma harzianum and Brevibacterium halotolerans for increased growth and oil yield in Mentha arvensis under greenhouse and field conditions. Ind. Crops Prod. 131, 173–181. doi: 10.1016/j.indcrop.2019.01.039
Sirajuddin, K. A., Ali, L., Chaudhary, H. J., Munis, M. F. H., and Masood, S. (2016). Bacillus pumilus alleviates boron toxicity in tomato (Lycopersicum esculentum L.) due to enhanced antioxidant enzymatic activity. Scient. Hortic. 200, 178–185. doi: 10.1016/j.scienta.2016.01.024
Sowndhararajan, K., Cho, H., Yu, B., and Kim, S. (2015). Effect of olfactory stimulation of isomeric aroma compounds, (+)-limonene and terpinolene on human electroencephalographic activity. Eur. J. Integr. Med. 7, 561–566. doi: 10.1016/j.eujim.2015.08.006
Tenga, Z., Shaoa, W., Zhanga, K., Huoa, Y., and Lia, M. (2019). Characterization of phosphate solubilizing bacteria isolated from heavy metal contaminated soils and their potential for lead immobilization. J. Environ. Manage. 231, 189–197. doi: 10.1016/j.jenvman.2018.10.012
Van Den Dool, H., and Kratz, P. D. (1963). A generalization of the retention index system including linear temperature programmed gas-liquid partition chromatography. J. Chromatogr. 11, 463–471.
Vejan, P., Abdullah, R., Khadiran, T., Ismail, S., and Boyce, A. N. (2016). Role of plant growth promoting rhizobacteria in agricultural sustainability - a review. Molecules 21, 573–590. doi: 10.3390/molecules21050573
Wang, Q., Ye, J., Wu, Y., Luo, S., Chen, B., Ma, L., et al. (2019). Promotion of the root development and Zn uptake of Sedum alfredii was achieved by an endophytic bacterium Sasm05. Ecotox. Environ. Safe. 172, 97–104. doi: 10.1016/j.ecoenv.2019.01.009
Wang, W., Wu, Z., He, Y., Huang, Y., Li, X., and Ye, B. (2018). Plant growth promotion and alleviation of salinity stress in Capsicum annuum L. by Bacillus isolated from saline soil in Xinjiang. Ecotox. Environ. Safe. 164, 520–529. doi: 10.1016/j.ecoenv.2018.08.070
Wang, Y. Y., Yang, X. E., Zhang, X. C., Dong, L. X., Zhang, J., Wei, Y. Y., et al. (2014). Improved plant growth and Zn accumulation in grains of rice (Oryza sativa L.) by inoculation of endophytic microbes isolated from a Zn hyperaccumulator, Sedum alfredii Hance. J. Agric. Food Chem. 62, 1783–1791. doi: 10.1021/jf404152u
Xiao, Z., Luo, J., Niu, Y., and Wu, M. (2018). Characterization of key aroma compounds from different rose essential oils using gas chromatography-mass spectrometry, gas chromatography-olfactometry and partial least squares regression. Nat. Prod. Res. 32, 1567–1572. doi: 10.1080/14786419.2017.1389933
Xie, Z., Chu, Y., Zhang, W., Lang, D., and Zhang, X. (2019). Bacillus pumilus alleviates drought stress and increases metabolite accumulation in Glycyrrhiza uralensis Fisch. Environ. Exp. Bot. 158, 99–106. doi: 10.1016/j.envexpbot.2018.11.021
Yang, Q. Q., Zhang, D., Farha, A. K., Yang, X., Li, H. B., Kong, K. W., et al. (2020). Phytochemicals, essential oils, and bioactivities of an underutilized wildfruit Cili (Rosa roxburghii). Ind. Crop. Prod. 143, 1–8. doi: 10.1016/j.indcrop.2019.111928
Keywords: Bacillus sp, biological activity, content, efficient microorganisms, Rosa hybrida L.
Citation: Araújo NAF, Brandão RM, Barguil BM, Cardoso MdG, Pasqual M, Rezende RALS, Pereira MMA, Buttrós VHT and Dória J (2020) Plant Growth-Promoting Bacteria Improve Growth and Modify Essential Oil in Rose (Rosa hybrida L.) cv. Black Prince. Front. Sustain. Food Syst. 4:606827. doi: 10.3389/fsufs.2020.606827
Received: 15 September 2020; Accepted: 04 November 2020;
Published: 18 December 2020.
Edited by:
Everlon Cid Rigobelo, São Paulo State University, BrazilReviewed by:
Akbar Karami, Shiraz University, IranCopyright © 2020 Araújo, Brandão, Barguil, Cardoso, Pasqual, Rezende, Pereira, Buttrós and Dória. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Joyce Dória, am95Y2UuZG9yaWFAdWZsYS5icg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.