
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Sustain. Food Syst., 23 April 2020
Sec. Agroecology and Ecosystem Services
Volume 4 - 2020 | https://doi.org/10.3389/fsufs.2020.00040
This article is part of the Research TopicMoving from a Curative to Preventative Pest Management ParadigmView all 14 articles
 Sara M. Kross1,2,3*
Sara M. Kross1,2,3* Breanna L. Martinico2
Breanna L. Martinico2 Ryan P. Bourbour2
Ryan P. Bourbour2 Jason M. Townsend2,4Chris McColl3
Jason M. Townsend2,4Chris McColl3 T. Rodd Kelsey3
T. Rodd Kelsey3Agriculture-dominated landscapes harbor significantly diminished biodiversity. Woody vegetation along field margins can provide farmers with ecosystem services and benefit biodiversity. However, when crops are damaged by the biodiversity harbored in such vegetation, farmers are reluctant to incorporate field margin habitat onto their land and may even actively remove such habitats. We investigated how damage by both insect pests (sunflower moth, Homoeosoma electellum) and avian pests to sunflower (Helianthus annuus) seed crops varied as a function of field-margin and landscape-scale habitat, as well as by bird abundance and diversity. Surveys for insect damage, avian abundance, and bird damage were carried out over 2 years in 30 different fields. The mean percentage of moth-damaged sunflowers sampled was nearly four times higher in fields that had bare or weedy margins (23.5%; $877/ha) compared to fields with woody vegetation (5.9%; $220/ha) and was positively associated with landscape-scale habitat complexity. Birds damaged significantly fewer sunflower seeds (2.7%) than insects, and bird damage was not affected by field margin habitat type, landscape-scale habitat variables, or avian abundance, but was significantly higher along field edges compared to ≥ 50 m from the field edge. Avian species richness nearly doubled in fields with woody margin habitat compared to fields with bare/weedy margins in both the breeding season and in fall. These results indicate that the benefits of planting or retaining woody vegetation along sunflower field margins could outweigh the ecosystem disservices related to bird damage, while simultaneously increasing the biodiversity value of intensively farmed agricultural landscapes.
In the face of significant losses of both diversity and abundance of avian species (Rosenberg et al., 2019), farming agroecosystems represent a critical frontline for improving vast tracts of land for the conservation of biodiversity beyond the reserve system (Kremen and Merenlender, 2018; Grass et al., 2019). Establishing and protecting agroecosystems that harness functional diversity to provide ecosystem services at the farm and landscape level may simultaneously decrease chemical inputs and increase biodiversity (Daily et al., 2000; Bommarco et al., 2013; Weier et al., 2018; Kleijn et al., 2019). For example, establishing or maintaining strips of woody vegetation along field margins can increase the diversity, abundance, and corresponding ecosystem services of pollinators (Garibaldi et al., 2011; Sardiñas et al., 2016; M'Gonigle et al., 2015), arthropod predators (Eilers and Klein, 2009; Gareau et al., 2013), and birds (Heath et al., 2017; Gonthier et al., 2019). Likewise, higher amounts of natural habitat within agricultural landscapes (landscape-level complexity) can also increase biodiversity and associated ecosystem services (Chaplin-Kramer et al., 2011; Klein et al., 2012; Karp et al., 2018; Gonthier et al., 2019).
Increasing structural complexity within the working agricultural landscape can enhance the conservation value of larger tracts of protected land situated on the periphery of agricultural lands (Heath et al., 2017; Kremen and Merenlender, 2018). By integrating habitat complexity into agricultural landscapes, dispersal corridors, access to food sources, and greater genetic connectivity may occur across a wide diversity of taxa (Isbell et al., 2017). Such changes to the agricultural landscape could therefore have dual benefits for both landscape-scale biodiversity (DeClerck et al., 2010) and on-farm sustainability in the form of ecosystem services including biological pest control (Mitchell et al., 2013). However, natural habitat in agroecosystems can also harbor pests, which is true for avian pests of many seed and fruit crops worldwide (De Grazio, 1978; Gebhardt et al., 2011; Kross et al., 2012; Schäckermann et al., 2014). Farmers that perceive birds as detrimental to their crops will take action to deter birds (Kross et al., 2018), often by removing field margin habitat (Gennet et al., 2013) or utilizing commercially available bird deterrents such as gas guns, reflective tape, or netting (Baldwin et al., 2013), all of which can be costly for both farmers and non-target wildlife. Bird depredation of crops therefore not only has direct economic implications for growers, but can lead farmers to oppose conservation programs within agricultural communities and on their own properties (Kross et al., 2018).
Farmers are the primary decision makers for land management choices within agricultural regions, and their decisions, including those related to implementation of integrated pest management (IPM), are mostly based on direct economic returns (Kleijn et al., 2019). Ecosystem services, when enhanced by integrating habitat complexity into the working agricultural landscape, can serve as a part of IPM and contribute to a more sustainable and holistic preventative pest management paradigm (Stenberg, 2017). However, the effects of natural vegetation on biological control are complex and can vary with crop type, seasonality, farm management, and the demographic effects of interactions between natural enemies and pests (Karp et al., 2018; Settele and Settle, 2018). There is a strong need to provide clear, balanced empirical information to better inform habitat-modification based IPM strategies on the working, farm-scale level. This goal can be confused at times when, for example, studies into the detrimental behaviors of birds rarely focus on potentially beneficial impacts, and similarly, studies into beneficial pest-control services from birds rarely focus on damage that the same species may cause to crops (Pejchar et al., 2018; but see a few recent exceptions: Peisley et al., 2016; Gonthier et al., 2019). Therefore, disentangling the relationships between landscape- and field-level habitat complexity and crop damage from insect and avian pests has critical implications for both habitat management and preventative pest management in agroecosystems.
In California, one of the world's most productive and intensive farming regions, <4% of potential field margins have been planted with woody vegetation such as hedgerows (Brodt et al., 2009); field margins therefore have significant potential for increasing the biodiversity conservation value of California's working agricultural landscape. However, farmer surveys in California showed that major obstacles to hedgerow implementation included uncertainty around the potential ecosystem service benefits of hedgerows, along with concern that these hedgerows could harbor plant, insect, and vertebrate pests (Brodt et al., 2009). Research to provide empirical documentation of the costs and benefits of planting (and in some cases, retaining) such habitats is therefore critical to inform land management decisions.
Avian species and their abundance and diversity relationships to landscape structure can, in particular, create a still-unresolved conundrum in the analysis of costs and benefits to farmers (Pejchar et al., 2018). Local- and landscape-scale habitat influences both pests and their potential predators at varying scales and effect sizes (Karp et al., 2018). At the same time, some species of birds can provide ecosystem services, in the form of insect pest control, during the breeding season, and then can become major pests of the same crops when they switch to a more omnivorous diet in the fall (Figure 1).
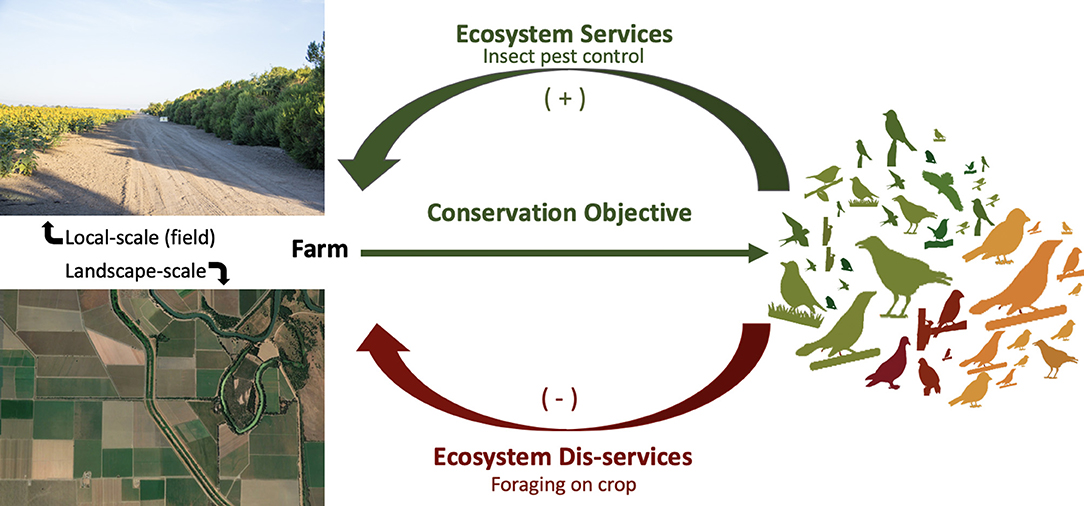
Figure 1. A conceptual framework for the potential effects of landscape- and local-scale habitat complexity on avian diversity (represented by bird silhouettes), and potential benefits (ecosystem services in the form of insect pest control) or costs (ecosystem disservices in the form of bird damage to crops), which can also vary with seasonality and diet, that affect yields for farmers.
In the present study, we analyzed the potentially conflicting roles of avian species within the unique sunflower (Helianthus annuus) seed growing region of California's Central Valley. We investigated the effects of both field-margin and landscape-scale habitat complexity on the occurrence of (1) potential benefits to farmers in the form of (A) avian insectivory leading to reduced occurrence of major invertebrate pests of sunflower seeds; and also (2) of potential costs to farmers in the form of (B) insect damage to sunflower seeds, and (C) bird damage to sunflower seeds.
California's Central Valley is one of the most productive agricultural landscapes in the world, producing over 25% of the fresh produce consumed in the United States (USDA 2015), and valued at over $45 billion (USD) per year. Over 95% of the Central Valley's riparian and wetland ecosystems have been replaced by highly intensive agriculture and urban development (Katibah, 1984; Frayer et al., 1989), with remnant native habitat existing only in fragmented and isolated patches. Nevertheless, some native biodiversity in this region persists despite the highly human-modified landscape (Heath et al., 2017).
Each year, sunflower is grown for hybrid seed production on an average of 20,234 ha (50,000 acres) across California's Sacramento Valley, producing over 31,750 tons, valued at ~$70 million/year (Long et al., 2019). California's Central Valley produces over 95% of the United States' hybrid sunflower seeds, and over 25% of global sunflower seeds (Long et al., 2019). Sunflowers grown for seed are valued 5–10 times higher than the commercial oil crops for which they are used (Long et al., 2019), and growers therefore have a low threshold for damage. All sunflower fields in our study were grown for the same seed company and therefore were grown using the same standard sunflower production field-management practices (Long et al., 2019). This study was conducted within conventional fields (i.e., non-organic fields), but no growers reported utilizing insecticides on their fields over the duration of this study.
The sunflower moth (Homoeosoma electellum) is a major pest of sunflowers in North America and is the predominant insect pest of the crop in California (Long et al., 2019). Female sunflower moths lay eggs among the florets of sunflowers in early bloom, and eggs take 2–5 days to hatch. After hatching, larvae remain on the face of flowers for 8 days before boring into the developing seeds where they can cause losses of 30–60% of a crop (Long et al., 2019). Birds are the predominant vertebrate pest of sunflower crops around the world (De Grazio, 1978; Schäckermann et al., 2014; Ernst et al., 2019; Long et al., 2019). In North America, and in our study area, Icterid birds and the non-native European starling (Sturnus vulgaris) cause the most damage to sunflowers because of their omnivorous diets and behavior of foraging in large flocks. In Israel, bird damage to sunflowers is often concentrated to the edges nearest to habitat (specifically, large trees) that can act as shelter for birds, and can also be affected by landscape-scale habitat (Schäckermann et al., 2014), however flocking birds often cause damage within the interior of fields as well (e.g., European starlings in New Zealand vineyards; Kross et al., 2012).
The presence or absence of woody habitat along field margins has been demonstrated to affect avian diversity and abundance in our study area (Heath et al., 2017), and at the time of our study the effects of habitat on sunflower moth damage was not described. To quantify the influence of habitat complexity on the bird community and sunflower damage, we included fields with woody margin habitat (n = 6 in 2014 and n = 12 in 2015) and fields with bare or weedy field margins in (n = 7 2014 and n = 5 in 2015), for a total of 30 fields (Figure 2). To quantify local (field) habitat complexity, at 5 evenly spaced locations along each 200-m long field margin transect (used for bird counts, see 2.6 below), we collected data on the maximum height of vegetation within 10 m of the of the transect, we estimated the number of canopy layers (out of a possible 7 predetermined canopy layers), and used satellite imagery to measure the width of field margin vegetation perpendicular to the focal field (see Heath et al., 2017 for details).
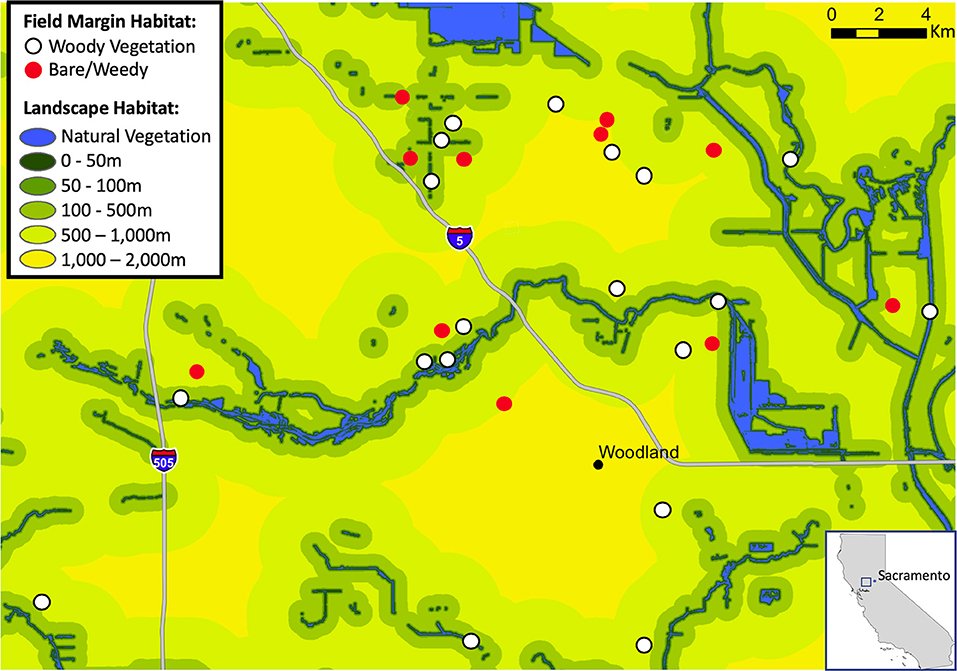
Figure 2. Map showing sunflower field locations at varying distances from natural habitat (blue) across an intensive agriculture landscape. Sunflower fields had either bare/weedy field margin habitat (red points), or had woody vegetation field margin habitat (white points).
Because the variables describing field margin habitat (height, width, and number of vegetation layers) were highly correlated, we used a Principle Components Analysis (PCA) to reduce these into two orthogonal axes that explained over 95.5% of the variance among them. The two axes, PC1 and PC2, were included as predictor variables in our candidate models for sunflower damage and for bird abundance and richness. PC1 explained 86.2% of the variability among habitat variables and was negatively associated with all three variables, whereas PC2 was positively associated with habitat width and height, and negatively associated with habitat layers. Therefore, if PC1 is a positive predictor of damage, we would expect less damage at sites with habitat that is taller, wider and has more layers (because of the inverse relationship). If PC2 is a positive predictor of damage, we would expect less damage at sites with more habitat layers and more damage at sites with taller/wider habitat.
To quantify and incorporate landscape habitat complexity into our study design, we selected fields at varying distances from natural habitat, which in our study area consists mainly of remnant and restored riparian areas (Figure 2). We used pre-existing habitat data for our study area (CA DWR, 2008; Geographic Information Center, 2009), and added polygons for any trees within 800 m of each transect that were not included in the existing dataset (e.g., trees lining driveways, trees around homesteads). To calculate the distance to riparian areas, we first created a distance raster that encompassed the entire study area by using the Euclidean distance algorithm in ArcGIS 10.1 (ESRI, 2010). We used the riparian vegetation GIS dataset (habitats classified as native riparian, blue oak woodland, valley foothill riparian, fresh emergent wetland, saline emergent wetland, and valley foothill riparian) as the “source” input for the algorithm and set the output grid cell size to 10 m. Each field's transect center point was then buffered by 50 m, and we calculated the distance from each grid cell within the buffer to the nearest riparian vegetation polygon. The mean distance for all cells within each buffer was calculated as the distance value for each field. We also calculated the mean proportion area consisting of natural habitat at concentric buffer distances of 100, 200, 400, and 800 m, which have been shown to be relevant scales for riparian bird species in the Central Valley (Seavy et al., 2009).
To account for collinearity among landscape-scale habitat complexity variables, we ran separate models using the predictor variables for landscape-scale habitat complexity (either distance to the nearest natural habitat or percent natural cover at varying distances) for both the insect damage (Table S1) and the bird damage (Table S2) models.
In 2015, we created exclosures to prevent vertebrates (birds and bats) from accessing sunflowers (see Maas et al., 2019 for a review of exclosure methods). Exclosures consisted of nylon bird netting (No-Knot Bird Netting ¾ polypropylene mesh, Bird B Gone Inc®, Irvine, CA) draped over an area 4 sunflowers in width and ~20 flowers in length (for a total of ~80 plants) and secured to cover the tops of the flowers to a height of ~2–4 feet above the ground. Birds were never observed inside our exclosures. Exclosures were installed in late spring, prior to the onset of bloom (which is when sunflower moth typically lay eggs on the flowers), and were checked and maintained over the entire growing season until final damage estimates were made in August-September. We set up four exclosures in each field, with the closest end of each exclosure located 5, 10, 50, and 100 m from the edge of the field. Due to last minute changes in the harvest schedule at some fields, we were able to collect damage data from the exclosures at nine different fields.
We sampled from 10 sunflowers at distances from 0 to 200 m from the field edge. In 2014, we collected observations of both insect and bird damage from each site at 0, 10, 20, 30, 40, 50, 75, 100, 150, and 200 m from the field edge. In 2015, we collected observations from each site at 5, 10, 50, and 100 m from the field edge because we found in 2014 that bird damage dropped to close to 0 at distances beyond 50 m (see Figure 3C), and that insect damage was largely unchanged by distance from the field edge. Estimates for insect and bird damage in 2015 were taken from sunflowers within exclosures and from sunflowers that were ~10 m from the exclosures (parallel to the field margin), but only data from non-enclosed sunflowers was used in our comparative analysis of insect damage.
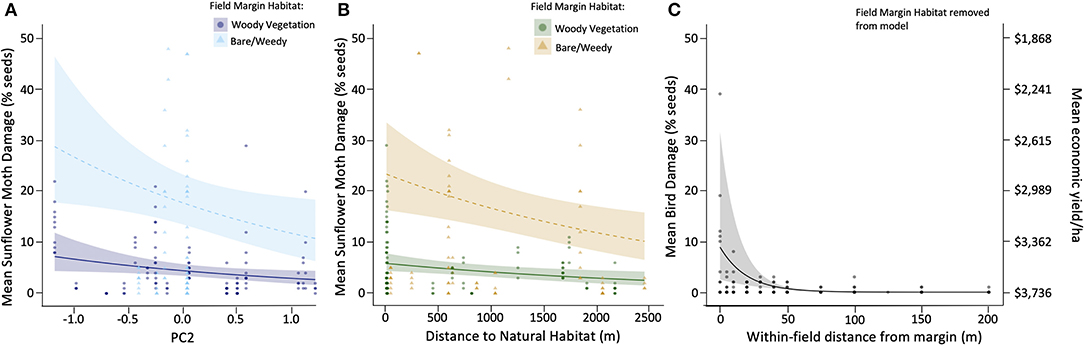
Figure 3. Model estimates (and 95% confidence intervals) of percent of sunflower seeds damaged (primary axis) as a function of the presence (darker colored lines) or absence (lighter colored lines) of woody vegetation along field edges and, (A) increasing field margin height and width (PC2), (B) the distance to the nearest natural habitats; and (C) percent seeds damaged by birds as a function of the distance of sampling points within each field from the nearest field margin. Raw data points are shown, with darker points indicating multiple overlapping points. The secondary axis for economic yield (right side of figure) applies to all 3 panels and shows the mean value (from 2015 to 2018) of sunflower seed crop in the region ($3,736 USD) equivalent to 0% damage, and decreasing as percent damage increases on the primary axis.
We quantified both bird and insect damage by visually inspecting 10 sunflowers within each sampling area. Sunflowers were chosen by reaching out to select a plant stalk, so the seed-bearing area of each plant was not seen until after the plant was selected. Observers moved a few steps along and between rows to select each new flower. Bird damage was characterized by missing seeds. We were careful to avoid classifying wind-damaged seeds as bird damage. These seeds were generally removed from larger continuous areas of the sunflower head, whereas seeds removed by birds were in patchy sections or removed singularly. Insect damage was characterized by an area of visible frass (insect excrement and webbing) on the surface of multiple sunflower seeds. Seeds under the frass were often shrunken or visibly damaged. All areas that were under frass were classified as insect-damaged.
To estimate the percent of seeds on each sunflower that were damaged, we used a pre-cut circular piece of galvanized steel chicken-wire that was marked to allow for easy measurement of the flowers. Sunflower heads were classified into different size classes based on the diameter (to the nearest 1.3 cm, or 0.5 inches) of the seed-bearing area on each plant. We then estimated the number of hexagons on the wire (to the nearest ¼ hexagon) that was damaged by birds or damaged by insects on each sunflower head. Using the flower circumference and the known area within each hexagon of our grid, we were then able to calculate the percent of each sunflower head that was damaged by birds, and the total that was damaged by insects. For each sampling location, we aggregated the data from the 10 flowers for a single mean for percent damage. To estimate yield, damage from insects and damage from birds were summed for a total percent damage to each sunflower, since both types of damage result in a direct loss of yield for growers.
We used published data on mean sunflower yields and economic value for the Sacramento Valley from 2015 to 2018 (Long et al., 2018) to calculate the reduction in gross earnings for farmers as a result of insect and bird damage in response to significant predictor variables. Mean sunflower yields were 1,260 lbs/acre (1,412 kg/ha; range 1,076–1,748 kg/ha) after seed companies cleaned and removed non-viable seeds and non-seed material from field harvests (Long et al., 2018). Seeds were valued at a mean value of $1.2/lb [$0.54/kg; range of $0.41–0.68/kg (Long et al., 2018)]. We calculated the economic effect size of insect or bird damage by multiplying the scaled effect sizes from our model estimates with these mean values, assuming that yields were valued at $3,736/ha if they sustained no bird or insect damage.
We conducted four bird surveys at each site, two in summer (June 9–July 2) and two in fall (August 5–September 16). All bird surveys were conducted by trained observers and timed to coincide with sunflower bloom in the summer (when sunflower moths typically lay eggs on the flowers), and immediately prior to the seed harvest in the fall. All counts were conducted between dawn and 10 a.m. and were not conducted in very cold (<3 C) or very hot weather (>24 C), in high winds, or in heavy precipitation. Counts were also re-scheduled if there were any farm workers or machinery in our focal field. The order in which fields were visited for counts was randomized within geographical groupings of fields. We conducted two counts per visit at each field: one to quantify the birds utilizing the field margin habitat, and another to quantify the birds utilizing the field interior. To count birds utilizing field margin habitat, observers walked a 200 m transect over 10 min, counting all birds detected by sight or sound within 20 m of the field margin, but not within the field itself. To count birds utilizing the field interior, observers returned to the mid-point of the transect, allowed 5 min for birds to settle, and then conducted a 10-min point count focused only on birds that were observed within the field (a half circle with an ~200 m radius from our center point). We truncated the detection distance at field margins because margin habitat varied across sites. We assumed that intra-species detectability was the same within all sunflower fields, since sunflowers were at similar levels of maturation and height at the time of our surveys. Sunflowers are a tall (2–3 m) and densely planted crop, so most birds detected within the crop were of individuals flying into or out of fields, or singing/calling within a field.
For all statistical models, we included as predictor variables in our maximal models the continuous variables for the distance from the nearest natural habitat (or proportion natural cover within concentric distance bands), PC1, PC2, as well as the categorical variable for whether the field had a weedy or bare edge (simple edge habitat) or had woody field margin habitat (complex edge habitat). For insect and bird damage to sunflowers, we also included the distance into the field the sample was collected (as both a linear and quadratic predictor, to account for potentially non-linear effects of distance on insect and bird damage). We simplified the maximal models by removing interactions, then main effects, until no further reduction in residual deviance (measured using Akaike's Information Criterion) was obtained. For all regression analyses, we considered candidate models with ΔAIC ≤ 2 and chose the most parsimonious model. All data was analyzed in R (R Core Team, 2015).
We used a Wilcoxon rank-sum test to compare the total insect damage observed inside exclosures and in adjacent non-exclosure locations.
Only the data from the non-exclosure sampling locations were used for investigating the effects of habitat variables on sunflower damage. Sunflower moth damage and bird damage were analyzed in separate models. We assessed our data for appropriate model distributions using quantile comparison plots (“qqplot” in Fox and Weisberg, 2019), then the fitdistr function (Venables and Ripley, 2002). We also used likelihood ratio tests to compare the final models to alternative likely distributions. For both insect and bird damage to sunflowers, we used generalized linear models with a negative binomial family of errors (“glm.nb” in MASS package; Venables and Ripley, 2002).
We ran eight separate generalized linear regressions with a poisson family of errors for avian species richness and abundance along the field edge and within the field interior for data collected in summer and in fall (Table S3).
There was no significant difference between sunflower damage from insects inside exclosures (vertebrates excluded; mean = 3.40 ± 0.61% damage) compared to areas outside of exclosures (vertebrates present; mean = 3.08 ± 0.47% damage; W = 67,828, p = 0.21; Figure S1).
Sunflower moth damage was almost four-times higher at sites with bare or weedy field margin habitat (23.46 ± 1.41%) compared to sites with woody vegetation (5.89 ± 1.16%; z = 7.12, p < 0.001). There was a slight decrease in sunflower moth damage as habitat height and width (PC2) increased (z = −2.75, p = 0.005; Figure 3A). Model selection revealed that the variable for mean distance to natural habitat was the most parsimonious landscape-scale habitat variable in our insect damage models (Table 1) and had a significant reduction in damage as distance from natural habitat increased (z = −2.25, p = 0.02; Figure 3B).
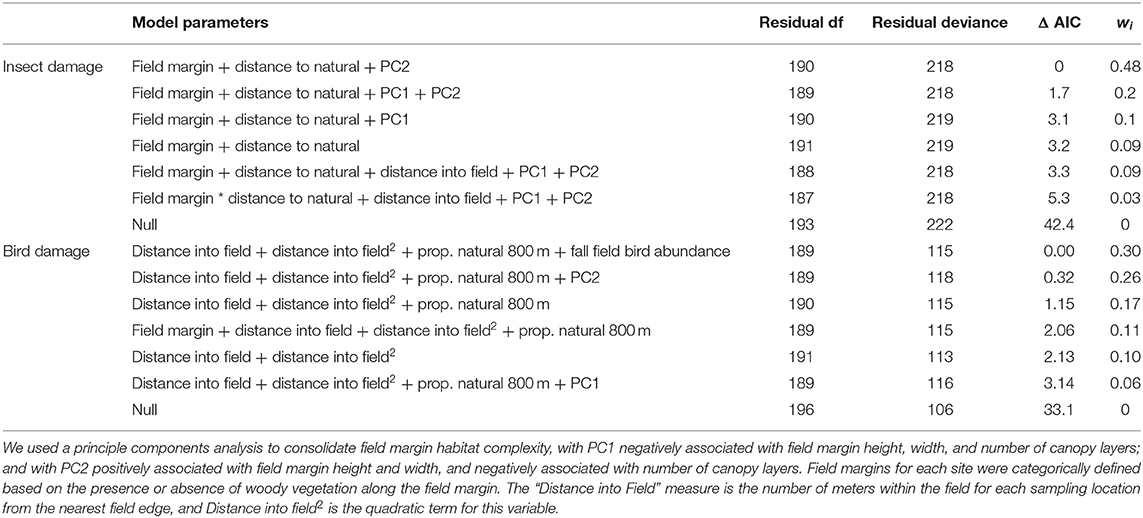
Table 1. Model selection for candidate models explaining sunflower moth damage to sunflower seeds using the distance to nearest natural habitat as a measure of landscape-scale habitat complexity, and for candidate models explaining bird damage to sunflower seeds using the proportion natural habitat within 800m of each site.
The most parsimonious model for bird damage included the proportion of natural habitat within 800 m of the field as the landscape-scale habitat variable (Table 1) and had a marginally significant increase in bird damage (z = 1.96, p = 0.05). Bird damage was highest at the edge of fields, regardless of the presence of field margin habitat, and dropped quickly to near 0% within 50 m of the field edge (Figure 3C). This effect was driven primarily by distance from field edge, with the linear (z = −4.38, p < 0.001) and quadratic values (z = 2.93, p = 0.003) for distance from field edge retained in the final model.
Our damage models estimate that the presence of woody field margin habitat results in significant changes to yield and therefore economic value of sunflower crops. Multiplying these results by the mean value of sunflower seeds in the region ($3,736/ha) allows for a coarse estimate of the economic implications of damage to sunflowers. For example, at sites adjacent to natural vegetation, farmers would expect to lose $877/ha in yields due to sunflower moth damage at sites with bare/weedy vegetation along the field margin, compared to $220/ha in lost yields due to sunflower moth damage at sites with woody vegetation, but this difference would lessen with increasing distance from natural areas as overall insect damage also declined (Figure 3B, secondary axis). To put this into perspective, the mean cost of applying insecticides to treat for sunflower moth is $292/ha (the equivalent of losses of ~7.8% damage), so our results suggest that fields with more complex, woody margins would be likely to remain under an economic threshold that would trigger growers to apply insecticides, whereas sites with bare/weedy margins mostly incur damage above that threshold (Figures 3A,B, secondary axis). Bird damage at the field edge would result in $100/ha in lost yields but that would decline to negligible damage within 50 m of the field edge (Figure 3C, secondary axis).
We observed 70 different avian species during our summer counts, and 74 species during our fall counts. These included California ‘Bird Species of Special Concern′ (Shuford and Gardali, 2008) like northern harrier (Circus hudsonius), yellow warbler (Setophaga petechia), and California “Threatened” species like Swainson's hawk (Buteo swainsoni), and tri-colored blackbird (Agelaius tricolor, 13 individuals observed at one site). During our summer counts, 64 different bird species utilized sunflower field edges and 49 species utilized field interiors. During our fall counts, we observed 69 species utilizing sunflower field edges and 46 species utilizing field interiors. The most abundant birds observed in fields interiors during the fall were Icterid species including Brewer's (Euphagus cyanocephalus) and red-winged blackbirds (Agelaius phoeniceus), European starling (Sturnus vulgaris), American goldfinch (Spinus tristis), house finch (Haemorhous mexicanus), and mourning dove (Zenaida macroura), all of which are species that are likely to damage sunflower seeds- although mourning doves most likely feed on seeds that have been knocked to the ground.
For our summer counts, avian species richness (Figure 4A) and abundance (Figure 4B) along field edges were positively associated with increasing field margin habitat complexity (Table 2, Table S4). Increasing field margin habitat was associated with lower avian species richness within fields in summer (PC1, Figure 4C, Table 2). For summer field interiors, avian species richness was negatively associated with increasing field margin habitat complexity (PC1, Figure 4C, Table 2). Avian abundance within field interiors in the summer was negatively associated with increasing height and width of field margin habitat (PC2, Figure 4D, Table 2). Model results for landscape scale habitat complexity, measured as the distance to nearest natural habitat, indicate that fields located further from natural habitat had lower avian species richness and abundance along field edges in the summer (but not field interiors, Table 2).
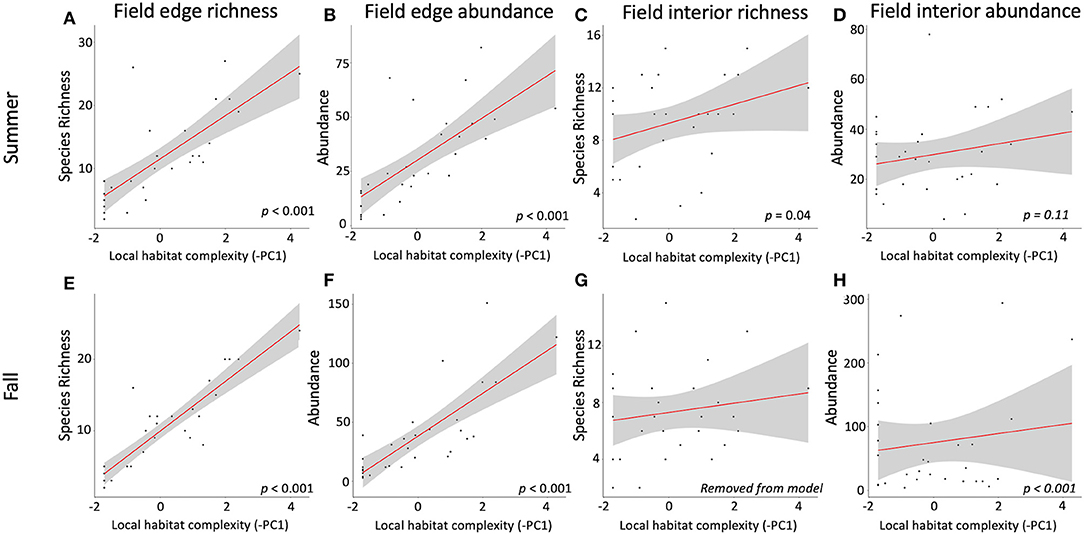
Figure 4. Avian species richness and abundance along sunflower field edges and within sunflower field interiors in Summer (top row) and Fall (bottom row) as a function of increasing field margin habitat height, width, and number of canopy layers (-PC1). Statistical significance of PC1 variable as a predictor in a linear regression for each response variable is shown bottom right in each panel. Panels show (A) summer field edge richness, (B) summer field edge abundance, (C) summer field interior richness, (D) summer field interior abundance, (E) fall field edge richness, (F) fall field edge abundance, (G) fall field interior richness, and (H) fall field interior abundance.
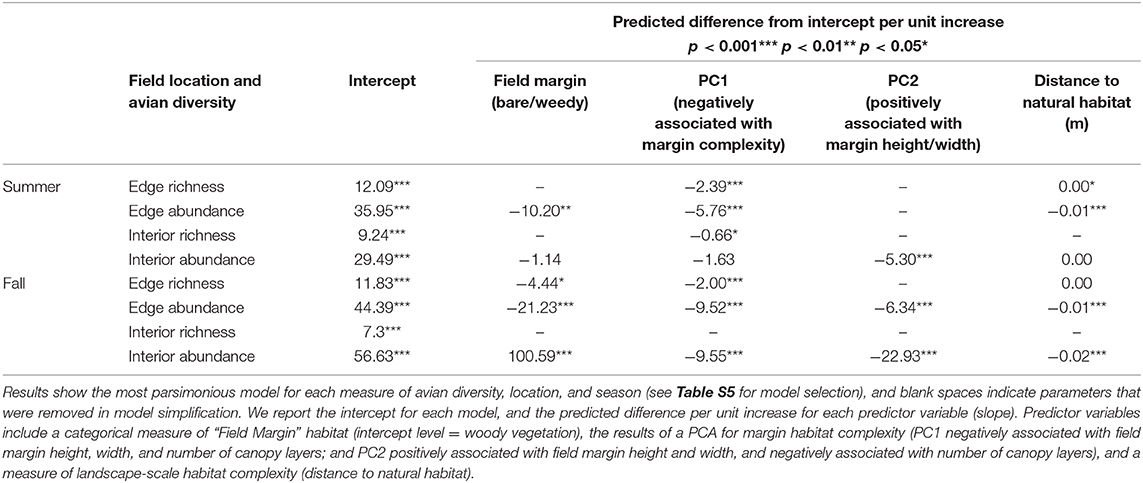
Table 2. Model estimates for each of eight models for avian species richness and abundance measured either along the edge of sunflower fields, or within the field interiors, in either summer or fall.
In the fall, avian species richness (Figure 4E) and abundance (Figure 4F) along field edges, and avian abundance within field interiors (Figure 4H) were positively associated with increasing field margin (local) habitat complexity (PC1; Table 2, Table S4). Avian species richness within field interiors in the fall was not significantly correlated with PC1 (Figure 4G). However, avian abundance along field edges and within field interiors was negatively associated with PC2, which is associated positively with margin habitat height and width (Table 2). Fall avian abundance was significantly lower along bare/weedy field edges (estimated mean reduction of 21.23 birds/transect) compared to along edges with woody vegetation (estimated mean of 44.39 birds/transect), but the opposite was true within field interiors, where there was an estimated increase of 100.59 more birds per transect at sites with bare/weedy edges compared to fields with woody vegetation along their edges (intercept = 56.63, Table 2). Only the abundance of birds within field interiors was significantly driven by landscape-scale habitat complexity, with fewer birds in fields further from natural habitat patches (at a rate of ~2 birds per 100 m, Table 2). Avian species richness in field interiors was not correlated with any of our predictor variables for local or landscape scale habitat complexity.
Our results suggest that sunflower growers would benefit from planting or maintaining woody vegetation alongside their fields since insect damage was significantly higher at sites without field margin vegetation, while bird damage was not driven by field margin habitat. In fact, bird damage at our 30 fields was similar across sites with and without field margin habitat. Furthermore, within sunflower fields across all distances from the field margin, sunflower moth damage was significantly higher than bird damage, and was the main source of yield loss for sunflower growers in our area. The pest control service benefits that farmers receive from field margin vegetation therefore outweigh the potential ecological disservices associated with bird damage to sunflowers. Our results also indicate a clear benefit for biodiversity, with significantly higher species richness and avian abundance along field margins that had woody habitat. Combined, these results support the assertion that diversified farming systems can provide both farmers and broader society with multiple additive ecosystem services (Kremen and Miles, 2012).
Our exclosures did not reveal an effect of bird foraging on sunflower moth damage. This could be the result of small sample size (n = 36 exclosures in 9 fields), or these results could indicate that foliage-gleaning birds and bats are not a major predator of sunflower moth. We hypothesize that the patterns of sunflower moth damage we observed were driven by either increased predation pressure from invertebrates, or from aerially-hunting bats and birds (which would not have been affected by the exclosures). Because of their nocturnal nature, adult sunflower moths are likely to be targeted more by nocturnal arthropod predators and/or bats (both of which would not be affected by the presence of exclosures) than by the predominantly diurnal avian predators. Studies in California have shown that the presence of habitat along field margins is associated with increased diversity and abundance of beneficial insects including natural enemies (Eilers and Klein, 2009; Gareau et al., 2013; Morandin et al., 2014), and with increased bat activity (Kelly et al., 2016), suggesting that our results could be driven by either or both of these groups of predators. Alternative hypotheses to explain our findings could be that woody vegetation along field margins present physical barriers to sunflower moths, or that increasing sunflower resources further from natural habitat dilute concentrations (and therefore damage) of sunflower moths, especially if natural habitats are the source of sunflower moths (e.g., see Tscharntke et al., 2016; O'Rourke and Petersen, 2017). Further research is clearly needed in this system.
Our results demonstrate that both insect and bird damage increased with landscape-scale habitat complexity. California's Central Valley is largely dominated by agriculture, with few corridors of remnant natural habitat along riparian areas (Figure 2). In this landscape, such corridors of natural habitat may be a source of migrating sunflower moths, and may also provide roosting habitat for large flocks of icterid birds in the autumn. Natural areas may therefore be a greater source of pests than they are a source of natural enemies (e.g., Hypothesis 2 in Tscharntke et al., 2016). However, our results show that regardless of how complex the landscape a farm is embedded within is, retaining or planting woody vegetation along field margins leads to a decrease in insect damage, and has no effect on total bird damage compared to sites with low levels of local habitat complexity. Importantly, this is also the scale at which farmers make decisions about planting, and therefore has the largest implications for rapid changes on private lands. Native hedgerows, the primary method for farmers to plant new woody vegetation along field margins, are also an important sources of pollination services for sunflowers (Sardiñas et al., 2016) and support pollinator metacommunity dynamics (Ponisio et al., 2019).
Habitat loss, largely a result of agriculture, is a primary driver of alarming trends of decreasing avian abundance in North America (Rosenberg et al., 2019). Both local (e.g., Hinsley and Bellamy, 2000; Batary et al., 2012; Heath et al., 2017; Gonthier et al., 2019) and landscape (e.g., Railsback and Johnson, 2014; Heath et al., 2017; Karp et al., 2018; Gonthier et al., 2019) habitat complexity have been linked to increased diversity of avian communities in farmlands, and may boost the conservation value of intensive agricultural landscapes. We found that avian species richness was positively associated with local-scale and landscape-scale habitat complexity. The presence of woody vegetation also led to higher avian abundance along field edges, but was associated with decreased avian abundance within sunflower fields in the fall, although this did not drive a reduction in bird damage in our study.
Birds on farms can provide multiple, overlapping, and complex benefits and costs for farmers at multiple scales (Pejchar et al., 2018). Individual species can be beneficial to a crop in some seasons and detrimental in others, or may benefit one crop and cause damage to another. Birds may also disrupt other natural trophic cascades that benefit farmers (Grass et al., 2017). Importantly, while our results indicate a net benefit of woody vegetation along field margins for both sunflower yields and avian diversity in California, sunflowers in other regions (Peer et al., 2003; Schäckermann et al., 2014; Ernst et al., 2019) suffer from economically significant bird-damage to the same crop. Therefore, we caution that land managers and scientists should consider local climate, habitat availability, agricultural practices, and avian communities before translating our findings into management changes in other regions.
Increasing natural habitat in intensive agricultural landscapes can provide numerous ecosystem services and support biodiversity. However the risk of also attracting pests is a major cause of farmer reluctance to plant or retain such habitat (Brodt et al., 2009). Finding solutions that lead to landscapes that benefit both wildlife and farming is essential to ensuring food security and a thriving biodiversity in the future (Bommarco et al., 2013; Kremen and Merenlender, 2018). Our study demonstrates that while landscape habitat complexity leads to slight increases in insect and bird damage, but also increases bird diversity and abundance. Regardless of landscape-scale habitat, fields with local habitat complexity have higher yields compared to fields with bare or weedy edges, and fields with woody vegetation along field margins also harbor a greater diversity and abundance of birds. To move into a more preventative pest management paradigm, encouraging farmers in California to plant or retain woody vegetation along field margins will simultaneously increase sunflower seed yields and increase the diversity and abundance of birds.
Data will be made available on the Columbia University Library Digital Repository, https://doi.org/10.7916/d8-k1sp-wd06.
The animal study was reviewed and approved by Institutional Animal Care and Use Committee (IACUC #18033) at the University of California, Davis.
SK, TK, and JT conceived the ideas and designed methodology. SK, BM, and RB field assistants collected the data. CM performed the landscape analysis. SK analyzed the data and led the writing of the manuscript. All authors contributed critically to the drafts and gave final approval for publication.
Fieldwork was funded by the David H. Smith Conservation Research Fellowship to SK, who was hosted by J. Eadie at UC Davis.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank the landowners and growers who provided access and information for this study especially Button and Turkovich, Joe Muller & Sons, Bullseye Farms, Citrona Farms, and Bypass Farms. Pioneer Hi-Bred International allowed for this research to be conducted, and we received significant logistical advice from A. Anderson. We also received invaluable study design advice from R. Long and numerous local pest control advisors. R. Oliver, N. Brehaut, K. Shaw, S. Lei, J. Wall, and E. Barry helped with field work. D. Gonthier and two reviewers provided valuable feedback on this paper. An earlier version of this manuscript has been released as a Pre-Print at BioRxiv (Kross et al., 2019).
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fsufs.2020.00040/full#supplementary-material
Baldwin, R. A., Salmon, T. P., Schmidt, R. H., and Timm, R. M. (2013). Wildlife pests of California agriculture: regional variability and subsequent impacts on management. Crop Protect. 46, 29–37. doi: 10.1016/j.cropro.2012.12.016
Batary, P., Kovacs-Hostyanszki, A., Fischer, C., Tscharntke, T., and Holzschuh, A. (2012). Contrasting effect of isolation of hedges from forests on farmland vs. woodland birds. Commun. Ecol. 13, 155–161. doi: 10.1556/ComEc.13.2012.2.4
Bommarco, R., Kleijn, D., and Potts, S. G. (2013). Ecological intensification: harnessing ecosystem services for food security. Trends Ecol. Evol. 28, 230–238. doi: 10.1016/j.tree.2012.10.012
Brodt, S., Klonsky, K., Jackson, L., Brush, S. B., and Smukler, S. (2009). Factors affecting adoption of hedgerows and other biodiversity-enhancing features on farms in California, USA. Agrofor. Syst. 76, 195–206. doi: 10.1007/s10457-008-9168-8
Chaplin-Kramer, R., Tuxen-Bettman, K., and Kremen, C. (2011). Value of wildland habitat for supplying pollination services to Californian agriculture. Rangelands 33, 33–41. doi: 10.2111/1551-501X-33.3.33
Daily, G. C., Soderqvist, T., Aniyar, S., Arrow, K., Dasgupta, P., Ehrlich, P. R., et al. (2000). Ecology - the value of nature and the nature of value. Science 289, 395–396. doi: 10.1126/science.289.5478.395
De Grazio, J. W. (1978). “World bird damage problems,” in Proceedings of the 8th Vertebrate Pest Conference (Sacramento, CA).
DeClerck, F. A. J., Chazdon, R., Holl, K. D., Milder, J. C., Finegan, B., Martinez-Salinas, A., et al. (2010). Biodiversity conservation in human-modified landscapes of Mesoamerica: past, present and future. Biol. Conserv. 143, 2301–2313. doi: 10.1016/j.biocon.2010.03.026
Eilers, E. J., and Klein, A. M. (2009). Landscape context and management effects on an important insect pest and its natural enemies in almond. Biol. Control 51, 388–394. doi: 10.1016/j.biocontrol.2009.07.010
Ernst, K., Elser, J., Linz, G., Kandel, H., Holderieath, J., DeGroot, S., et al. (2019). The economic impacts of blackbird (Icteridae) damage to sunflower in the USA. Pest Manag. Sci. 75, 2910–2915. doi: 10.1002/ps.5486
Frayer, W. E., Peters, D. D., and Pywell, H. R. (1989). Wetlands of the California Central Valley: Status and Trends. Portland, OR: U. S. Fish and Wildlife Service.
Gareau, T. L. P., Letourneau, D. K., and Shennan, C. (2013). Relative Densities of natural enemy and pest insects within California hedgerows. Environ. Entomol. 42, 688–702. doi: 10.1603/EN12317
Garibaldi, L. A., Steffan-Dewenter, I., Kremen, C., Morales, J. M., Bommarco, R., Cunningham, S. A., et al. (2011). Stability of pollination services decreases with isolation from natural areas despite honey bee visits: habitat isolation and pollination stability. Ecol. Lett. 14, 1062–1072. doi: 10.1111/j.1461-0248.2011.01669.x
Gebhardt, K., Anderson, A. M., Kirkpatrick, K. N., and Shwiff, S. A. (2011). A review and synthesis of bird and rodent damage estimates to select California crops. Crop Protect. 30, 1109–1116. doi: 10.1016/j.cropro.2011.05.015
Gennet, S., Howard, J., Langholz, J., Andrews, K., Reynolds, M. D., and Morrison, S. A. (2013). Farm practices for food safety: an emerging threat to floodplain and riparian ecosystems. Front. Ecol. Environ. 11, 236–242. doi: 10.1890/120243
Geographic Information Center (2009). Vegetation – Central Valley Riparian and Sacramento Valley [ds1000].
Gonthier, D. J., Sciligo, A. R., Karp, D. S., Lu, A., Garcia, K., Juarez, G., et al. (2019). Bird services and disservices to strawberry farming in Californian agricultural landscapes. J. Appl. Ecol. 56, 1948–1959. doi: 10.1111/1365-2664.13422
Grass, I., Lehmann, K., Thies, C., and Tscharntke, T. (2017). Insectivorous birds disrupt biological control of cereal aphids. Ecology 98, 1583–1590. doi: 10.1002/ecy.1814
Grass, I., Loos, J., Baensch, S., Batáry, P., Librán-Embid, F., Ficiciyan, A., et al. (2019). Land-sharing/-sparing connectivity landscapes for ecosystem services and biodiversity conservation. People Nat. 1, 262–272. doi: 10.1002/pan3.21
Heath, S. K., Soykan, C. U., Velas, K. L., Kelsey, R., and Kross, S. M. (2017). A bustle in the hedgerow: woody field margins boost on farm avian diversity and abundance in an intensive agricultural landscape. Biol. Conserv. 212, 153–161. doi: 10.1016/j.biocon.2017.05.031
Hinsley, S. A., and Bellamy, P. E. (2000). The influence of hedge structure, management and landscape context on the value of hedgerows to birds: a review. J. Environ. Manage. 60, 33–49. doi: 10.1006/jema.2000.0360
Isbell, F., Adler, P. R., Eisenhauer, N., Fornara, D., Kimmel, K., Kremen, C., et al. (2017). Benefits of increasing plant diversity in sustainable agroecosystems. J. Ecol. 105, 871–879. doi: 10.1111/1365-2745.12789
Karp, D. S., Chaplin-Kramer, R., Meehan, T. D., Martin, E. A., DeClerck, F., Grab, H., et al. (2018). Crop pests and predators exhibit inconsistent responses to surrounding landscape composition. Proc. Natl. Acad. Sci. U.S.A. 115, E7863–E7870. doi: 10.1073/pnas.1800042115
Katibah, E. E. (1984). “A brief history of riparian forests in the Central Valley of California,” in California Riparian Systems, eds R. E. Warner and K. M. Hendrix (Berkeley, CA: University of California Press), 23–30.
Kelly, R. M., Kitzes, J., Wilson, H., and Merenlender, A. (2016). Habit diversity promotes bat activity in a vineyard landscape. Agric. Ecosyst. Environ. 223, 175–181. doi: 10.1016/j.agee.2016.03.010
Kleijn, D., Bommarco, R., Fijen, T. P. M., Garibaldi, L. A., Potts, S. G., van der Putten, W. H., et al. (2019). Ecological Intensification: bridging the gap between science and practice. Trends Ecol. Evol. 34, 154–166. doi: 10.1016/j.tree.2018.11.002
Klein, A.-M., Brittain, C., Hendrix, S. D., Thorp, R., Williams, N., and Kremen, C. (2012). Wild pollination services to California almond rely on semi-natural habitat: WIld pollination services to California almond. J. Appl. Ecol. 49, 723–732. doi: 10.1111/j.1365-2664.2012.02144.x
Kremen, C., and Merenlender, A. M. (2018). Landscapes that work for biodiversity and people. Science 362:eaau6020. doi: 10.1126/science.aau6020
Kremen, C., and Miles, A. (2012). Ecosystem services in biologically diversified versus: coventional farming systems: benefits, externalities, and trade-offs. Ecol. Soc. 17:40. doi: 10.5751/ES-05035-170440
Kross, S. M., Ingram, K. P., Long, R. F., and Niles, M. T. (2018). Farmer perceptions and behaviors related to wildlife and on-farm conservation actions. Conserv. Lett. 11:e12364. doi: 10.1111/conl.12364
Kross, S. M., Martinico, B. L., Bourbour, R. P., Townsend, J. M., McColl, C., and Kelsey, T. R. (2019). Net effects of field and landscape scale habitat on insect and bird damage to sunflowers. bioRxiv doi: 10.1101/804328
Kross, S. M., Tylianakis, J. M., and Nelson, X. J. (2012). Effects of introducing threatened falcons into vineyards on abundance of passeriformes and bird damage to grapes. Conserv. Biol. 26, 142–149. doi: 10.1111/j.1523-1739.2011.01756.x
Long, R., Gulya, T., Light, S., Khaled, B., Konrad, M., and Roland, M. D. (2019). Sunflower Hybrid Seed Production in California. University of California, Agriculture and Natural Resources.
Long, R. F., Light, S. E., Galla, M., and Murdock, J. (2018). Sample Costs to Produce Sunflowers for Hybrid Seed in the Sacramento Valley. UC Agriculture and Natural Resources Cooperative Extension, Agriculture Issues Center, UC Davis Department of Agriculture and Resource Economics.
Maas, B., Heath, S., Grass, I., Cassano, C., Classen, A., Faria, D., et al. (2019). Experimental field exclosure of birds and bats in agricultural systems — methodological insights, potential improvements, and cost-benefit trade-offs. Basic Appl. Ecol. 35, 1–12. doi: 10.1016/j.baae.2018.12.002
M'Gonigle, L. K., Ponisio, L. C., Cutler, K., and Kremen, C. (2015). Habitat restoration promotes pollinator persistence and colonization in intensively managed agriculture. Ecol. Appl. 25, 1557–1565. doi: 10.1890/14-1863.1
Mitchell, M. G. E., Bennett, E. M., and Gonzalez, A. (2013). Linking Landscape connectivity and ecosystem service provision: current knowledge and research gaps. Ecosystems 16, 894–908. doi: 10.1007/s10021-013-9647-2
Morandin, L. A., Long, R. F., and Kremen, C. (2014). Hedgerows enhance beneficial insects on adjacent tomato fields in an intensive agricultural landscape. Agric. Ecosyst. Environ. 189, 164–170. doi: 10.1016/j.agee.2014.03.030
O'Rourke, M. E., and Petersen, M. J. (2017). Extending the ‘resource concentration hypothesis’ to the landscape-scale by considering dispersal mortality and fitness costs. Agric. Ecosyst. Environ. 249, 1–3. doi: 10.1016/j.agee.2017.07.022
Peer, B. D., Homan, H. J., Linz, G. M., and Bleier, W. J. (2003). Impact of blackbird damage to sunflower: bioenergetic and economic models. Ecol. Appl. 13, 248–256. doi: 10.1890/1051-0761(2003)013[0248:IOBDTS]2.0.CO;2
Peisley, R. K., Saunders, M. E., and Luck, G. W. (2016). Cost-benefit trade-offs of bird activity in apple orchards. PeerJ 4:e2179. doi: 10.7717/peerj.2179
Pejchar, L., Clough, Y., Ekroos, J., Nicholas, K. A., Olsson, O., Ram, D., et al. (2018). Net effects of birds in agroecosystems. Bioscience. doi: 10.1002/eap.2115
Ponisio, L. C., Valpine, P., M'Gonigle, L. K., and Kremen, C. (2019). Proximity of restored hedgerows interacts with local floral diversity and species' traits to shape long-term pollinator metacommunity dynamics. Ecol. Lett. 22, 1048–1060. doi: 10.1111/ele.13257
R Core Team (2015). R: A Language and Environment for Statistical Computing. Vienne: R Foundation for Statistical Computing.
Railsback, S. F., and Johnson, M. D. (2014). Effects of land use on bird populations and pest control services on coffee farms. Proc. Natl. Acad. Sci. U.S.A. 111, 6109–6114. doi: 10.1073/pnas.1320957111
Rosenberg, K. V., Dokter, A. M., Blancher, P. J., Sauer, J. R., Smith, A. C., Smith, P. A., et al. (2019). Decline of the North American avifauna. Science 366, 120–124. doi: 10.1126/science.aaw1313
Sardiñas, H. S., Tom, K., Ponisio, L. C., Rominger, A., and Kremen, C. (2016). Sunflower (Helianthus annuus) pollination in California's Central Valley is limited by native bee nest site location. Ecol. Appl. 26, 438–447. doi: 10.1890/15-0033
Schäckermann, J., Weiss, N., Wehrden, H., von, and Klein, A.-M. (2014). High trees increase sunflower seed predation by birds in an agricultural landscape of Israel. Front. Ecol. Evol. 2:35. doi: 10.3389/fevo.2014.00035
Seavy, N. E., Viers, J. H., and Wood, J. K. (2009). Riparian bird response to vegetation structure: a multiscale analysis using LiDAR measurements of canopy height. Ecol. Appl. 19, 1848–1857. doi: 10.1890/08-1124.1
Settele, J., and Settle, W. H. (2018). Conservation biological control: improving the science base. Proc. Natl. Acad. Sci. U.S.A. 115, 8241–8243. doi: 10.1073/pnas.1810334115
Shuford, W. D., and Gardali, T. (eds.). (2008). California Bird Special Concern. Vol. 1. Western Field Ornithologists; California Department of Fish and Game.
Stenberg, J. A. (2017). A conceptual framework for integrated pest management. Trends Plant Sci. 22, 759–769. doi: 10.1016/j.tplants.2017.06.010
Tscharntke, T., Karp, D. S., Chaplin-Kramer, R., Batáry, P., DeClerck, F., Gratton, C., et al. (2016). When natural habitat fails to enhance biological pest control – Five hypotheses. Biol. Conserv. 204, 449–458. doi: 10.1016/j.biocon.2016.10.001
Venables, W. N., and Ripley, B. D. (2002). Modern Applied Statistics With S. New York, NY: Springer.
Keywords: agroecology, crop damage, ecosystem services, farm, hedgerow, integrated pest management, pest control, landscape
Citation: Kross SM, Martinico BL, Bourbour RP, Townsend JM, McColl C and Kelsey TR (2020) Effects of Field and Landscape Scale Habitat on Insect and Bird Damage to Sunflowers. Front. Sustain. Food Syst. 4:40. doi: 10.3389/fsufs.2020.00040
Received: 31 October 2019; Accepted: 20 March 2020;
Published: 23 April 2020.
Edited by:
David Gonthier, University Press of Kentucky, United StatesReviewed by:
Alistair John Campbell, Embrapa Amazônia Oriental, BrazilCopyright © 2020 Kross, Martinico, Bourbour, Townsend, McColl and Kelsey. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sara M. Kross, c21rMjI1OEBjb2x1bWJpYS5lZHU=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.