
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Sustain. Food Syst. , 27 March 2020
Sec. Crop Biology and Sustainability
Volume 4 - 2020 | https://doi.org/10.3389/fsufs.2020.00022
This article is part of the Research Topic Entomopathogens for Sustainable Food Production View all 6 articles
 Robert G. Hollingsworth1†‡
Robert G. Hollingsworth1†‡ Luis F. Aristizábal2*†
Luis F. Aristizábal2*† Suzanne Shriner2†Gabriel M. Mascarin3†Rafael de Andrade Moral4†
Suzanne Shriner2†Gabriel M. Mascarin3†Rafael de Andrade Moral4† Steven P. Arthurs5†
Steven P. Arthurs5†Since its recent establishment in Hawaii, the coffee berry borer (CBB), Hypothenemus hampei (Coleoptera: Curculionidae), threatens yields, quality, and price of coffee production. A limited number of insecticides (primarily Beauveria bassiana) are used to control CBB with minimal disruption in this agroecosystem. We evaluated two insecticide spray strategies across eight coffee farms in the Kona and Ka‘u districts of Hawaii Island. Coffee growers sprayed insecticides approximately monthly (calendar basis) or else in response to CBB field monitoring data (threshold based). Overall, farms adopting spray thresholds performed more insecticide applications early in the season (May to July), but significantly fewer overall, when compared with calendar-based strategies (i.e., 4–5 vs. 7–11 seasonal sprays, respectively). Generalized linear models assessing the variability in CBB infestation rates, berry penetration, and infection by B. bassiana indicated that threshold-based sprays provided equivalent CBB control compared with calendar ones. When corrected for yield, there were economic savings for threshold- vs. calendar-based spray programs (i.e., cost 5.4 vs. 11.8% of gross yield). Total defects in processed coffee after harvest were statistically similar between the two spray regimes, i.e., 8.5 ± 1.0% and 10.4 ± 1.7%, respectively. We hypothesize that B. bassiana applied early in the season is more effective, since the fungus targets initial CBB infestations when the prolonged location of founder females in the outer berry endosperm favors its infection. Our study suggests that spray timing for CBB based on field monitoring data can reduce costs; however, additional measures, such as field and post-harvest sanitation, are necessary to achieve sustainable CBB control in the Islands.
Worldwide, at least 80 countries in tropical and subtropical regions produce coffee, Coffea arabica L. (Gentianales: Rubiaceae), on over 10 million ha (Vega et al., 2015). According to the International Coffee Organization (ICO) (2019), the global world coffee production was 168.8 million bags in 2018/19, with Brazil, Vietnam, Colombia, Indonesia, Honduras, Ethiopia, and India being the largest coffee producer countries [Food and Agriculture Organization of the United Nations Statistics Division (FAOSTAT), 2020]. In Hawaii, coffee is grown on ≈ 3,200 ha, which produced 16.5 million kg of coffee cherries with a value of US $62.2 million in the 2016–17 season (USDA-NASS, 2017). Although representing a fraction of the 10 billion kg global harvest [International Coffee Organization (ICO), 2019], Hawaiian specialty coffee retains high market prices due to the quality, unique geographic location, fertile volcanic soils, and suitable year-round microclimate (Teuber, 2010).
The arrival of the coffee berry borer (CBB), Hypothenemus hampei Ferrari (Coleoptera: Curculionidae), an invasive insect pest, into the Hawaiian Islands in 2010 (Burbano et al., 2011) threatens the sustainability of coffee production in the Islands. The CBB develops within the coffee bean, resulting in significant losses in production and revenue (Woodill et al., 2014). Strategies for controlling CBB in other countries emphasize an integrated pest management (IPM) approach. Tactics reported include monitoring, deployment of chemical insecticides and biological control agents, as well as cultural control practices focused on crop and post-harvest sanitation of infested berries (Damon, 2000; Jaramillo et al., 2006; Vega et al., 2009, 2015; Aristizábal et al., 2012, 2016; Infante, 2018). In addition to the CBB, several other invasive pests including the coffee leaf miner, Leucoptera coffeella Guerion-Meneville (Lepidoptera: Lyonetidae), and diseases such as coffee leaf rust, Hemileia vastratix Bert et Br., represent a serious threat for the coffee industry (Talhinhas et al., 2017; Pantoja-Gomez et al., 2019).
Among the biological control agents for managing CBB, the entomopathogenic fungus Beauveria bassiana (Basl. Criv.) Vuill. (Hypocreales: Cordycipitaceae) has received great interest, both as a natural disease agent and as a microbial bioinsecticide. The fungus infects adult stages of CBB, especially during periods of high humidity (Bustillo et al., 1999). In field trials in Colombia, application of B. bassiana conidia to fallen berries resulted in about 30–40% infection of CBB and reduced subsequent tree infestations up to 50% (Bustillo et al., 1999; Vera et al., 2011). Another field study reported that a mixture of B. bassiana strains caused 60–66% CBB mortality (Benavides et al., 2012). Some efforts have been made to optimize the efficacy of B. bassiana under field conditions in Mexico (De La Rosa et al., 2000), Ecuador (Sponagel, 1994), and India (Haraprasad et al., 2001).
In Hawaii, natural infection of endemic B. bassiana strains has been noted (Wraight et al., 2018). Insecticides based on the commercial GHA strain of B. bassiana have recently been registered and imported for use in the Islands (Greco et al., 2018). Currently, the use of B. bassiana is recommended to coffee growers in Hawaii within IPM programs for the CBB (Kawabata et al., 2017). In recent years, many growers have applied monthly (calendar) applications of B. bassiana to their entire plantation (Aristizábal, 2018a). However, adoption of this practice has been hampered by the cost of purchasing and applying the product and variability in efficacy due to crop factors and environmental conditions. In theory, field monitoring to locate “hot spots” of activity, including the use of traps to determine peak CBB flight times, can help growers target B. bassiana applications during periods when the CBB is exposed and most vulnerable to infection (Aristizábal et al., 2017a). However, the practicality and effectiveness of field monitoring to support spray decisions for the CBB have not been determined in this region.
In the present study, we surveyed commercial coffee farms in two regions that applied B. bassiana for CBB control. We used a modeling approach to evaluate application strategies (i.e., spraying frequency) as used in an IPM program for CBB. We assessed the economic benefits of field monitoring by comparing CBB control on coffee farms using calendar applications of B. bassiana with those farms that adopted field monitoring and spray thresholds. Since these were commercials farms, it was not possible to include farms where no sprays were used.
Studies were conducted in 2016 and 2017 in seven coffee farms in Kona and one in the Ka'u district of Hawaii Island. Farms were 1–8 ha in area and varied in elevation (300–650 masl) and planting density (1,750–2,400 tree/ha). The cultivars “Typica” and “Caturra” were the most commonly grown and evaluated in this study.
Berry infestation rates by the CBB were monitored at 2–4 week intervals, starting early in the season (May–June) and continuing through harvest (December–January). Assessments at each location used the “30-tree sampling plan” developed in Colombia (Bustillo et al., 1998) and adopted in Hawaii (Aristizábal et al., 2017b, 2018; Kawabata et al., 2017). On each farm, a representative coffee plot was chosen (0.6–1.5 ha), and 30 trees within were randomly selected. One branch containing 40–100 developing green berries in the mid-portion of each tree was chosen, and all infested and non-infested berries counted. Infested berries were identified by the characteristic entrance hole in the berry made by CBB.
To determine the level of B. bassiana mycosis as well as the susceptibility of CBB to future infections, 2–3 infested berries were collected from each sampled tree (i.e., 60–90 berries per farm/sampling date). Infested berries were dissected to determine the position of the colonizing CBB female inside, which determines susceptibility to fungal infection (Figure 1). The presence of B. bassiana on colonizing CBB females (based on visible sporulation appearing as a white mycelial growth) was recorded. CBB positions were evaluated after infested berries had been opened and subsequently classified as AB position (live CBB female without penetration of endosperm) or CD position (live CBB female inside endosperm typically with offspring), or else dead or absent.

Figure 1. Stages of coffee berry borer (CBB) penetration in green berries. (A) “AB position” where female CBB is vulnerable to insecticides (berries 60–140 days old). (B) “CD position” where CBBs are protected within the endosperm (berries >140 days old). (C) CBB in “AB position” killed by Beauveria bassiana. (D) Infested berry with missing CBB.
Growers applied B. bassiana GHA strain (BotaniGard® ES or Mycontrol® ESO, Lam International Cooperation, Butte, MT, USA) throughout the season (Figure 2). Insecticides were applied at label rate (2.36 L/ha) and applied to “run-off” using a backpack sprayer or tractor mounted equipment. Growers adopted either of two spray strategies. In “calendar applications,” sprays were applied every month starting when berries were 60 days old and susceptible to CBB infestation and continued until harvest time. In the second case, growers adopted a “pest threshold–based” approach whereby sprays were carried out when CBB infestation rates reached thresholds ≥ 3% and simultaneously when ≥ 25% of CBBs were in the AB position. These thresholds were based on previous field observations in Latin America, when it was determined that CBB control efficacy can exceed 50% (Baker, 1989; Bustillo et al., 1998; Trujillo et al., 2006). It has been noted that CBB infestation rates > 5% prior to harvesting season can result in economic damage to processed coffee (Bustillo et al., 1998).

Figure 2. Seasonal coffee phenology. (A) Green developing berries where CBB infestation starts. (B) CBB female infected by B. bassiana with conspicuous white mycelium at the tip outgrowing the CBB cadaver. (C) Early-season B. bassiana application with pressurized sprayer. (D) Evaluation of CBB infected with B. bassiana. (E) Red coffee “cherries” at harvest.
To improve spray coverage, a surfactant (Widespread® Max, Loveland Products Inc. Greeley, CO, USA, or Nu Film® P, Miller Chemical & Fertilizer, LLC, Hanover, PA, USA) was used in all cases at 0.3–0.6 L/ha. Some grower applications included a combination (tank mixture) with an insect repellent derived from kaolin-clay (Surround® WP, Tessenderlo Kerly Inc. Phoenix, AZ, USA) applied at 11.3 kg/ha or a pyrethrin-based insecticide (PyronylTM Crop Spray, Central Garden & Pet Company, Schaumburg, IL, USA) applied at 0.88–1.7 L/ha. A limitation of our study was an absence of survey sites where no insecticides were applied, since the widespread threat of CBB across the region made incorporating sufficiently large control blocks impractical.
Finally, the costs of CBB management between the two spray programs were estimated across farms. Growers were surveyed to determine the cost of insecticides and associated labor costs for application.
Assessments were compared between farms adopting the calendar and spray threshold methods. In addition to seasonal CBB infestation data, coffee bean defects were assessed at harvest (October and November). For this procedure, cherry samples (2.5 kg per plot) were randomly collected on two separate occasions. Samples stored in plastic bags were sent to United States Department of Agriculture, Agricultural Research Service (USDA-ARS) Pacific Basin Agricultural Research Center (PBARC) in Hilo, Hawaii, for processing. The dried coffee product was assessed for quality based on Hawaii standards. The parchment coffee was removed to obtain “green beans” (processed coffee) and sieved through different screen sizes. Beans were rated for defects including CBB damage and classified as “extra fancy” (screen size 19), “fancy” (screen size 18), “number one” (screen size 16), or “prime” (other size) (Bittenbender and Smith, 2008).
The proportion of CBB-infested berries was analyzed with binomial generalized linear mixed models, including different quadratic trends over time for each spray system, and random intercepts, slopes, and quadratic terms over time. The proportions of B. bassiana infection and CBB mortality were analyzed with linear trends. The significance of effects was assessed using likelihood-ratio (LR) tests for nested models. The AB/CD position data were fitted with a multivariate linear model to the isometric log-ratio transformation of the category proportions, including different quadratic predictors per spray system over time, and the effect of the farm as a nuisance factor. The significance of the effects was assessed using Pillai's trace test.
The quality of processed coffee was assessed based on the proportional weight of green beans (processed dried coffee) with physical damage (defects) from the samples. These continuous proportion data were analyzed with beta mixed models, including the effect of the spray system as fixed and the effect of the farm as random, and the significance of the effects was assessed using LR tests for nested models. Total and relative costs of CBB management (based on the number of seasonal applications) were compared for each spray program using one-way ANOVA. A normal distribution was assumed for the continuous data, and a Poisson model was used for the count variable (number of spray applications).
Goodness-of-fit for all models was assessed using half-normal plots with a simulated envelope, using package “hnp” (Moral et al., 2017). All analyses were carried out in R software (R Core Team, 2019). The generalized linear mixed models were fitted with package “lme4” (Bates et al., 2015); the beta mixed models were fitted using package “glmmTMB” (Brooks et al., 2017). Graphics were produced with package “ggplot2” (Wickham, 2016).
On average, fewer applications per season were required in threshold-based spray programs (4.4 ± 0.2, range 4–5) compared with calendar-based ones (9.0 ± 1.2, range 7–11; F = 6.23, d.f. = 1, 6, P = 0.013). In farms adopting threshold-based sprays, 75% of B. bassiana applications occurred in the early season (May–July), when colonizing CBB females were observed as taking longer to penetrate immature berries that were unsuitable for oviposition (Figure 1). Supplementary Data for individual farms (Table S1).
The average proportion of infested berries across farms varied from 0.6 to 12.5% over the season. Across farms, CBB infestation rates were highest at early season (5.5 ± 1.1%), declining to 3.5 ± 0.3% by the end of the harvest season (7–9 months later). However, higher seasonal infestation rates were observed in the calendar (high) spraying regime (5.4% ± 0.4) compared to farms that used threshold-based (low) applications (2.8% ± 0.3%) (i.e., LR = 14.6, d.f. = 1, P = 0.00013). This difference represented a 48.2% reduction in the proportion of infested berries among farms adopting threshold-based sprays (Figure 3). The binomial mixed models revealed no interaction between spray system and sample week, fitted with a quadratic (LR = 3.67, d.f. = 1, P = 0.055) or linear trend (LR = 1.04, d.f. = 1, P = 0.307). On the other hand, there was a significant quadratic trend with week (LR = 12.21, d.f. = 1, P = 0.0005). This result supports the idea that the relative CBB infestation pressure is cyclical during the season.
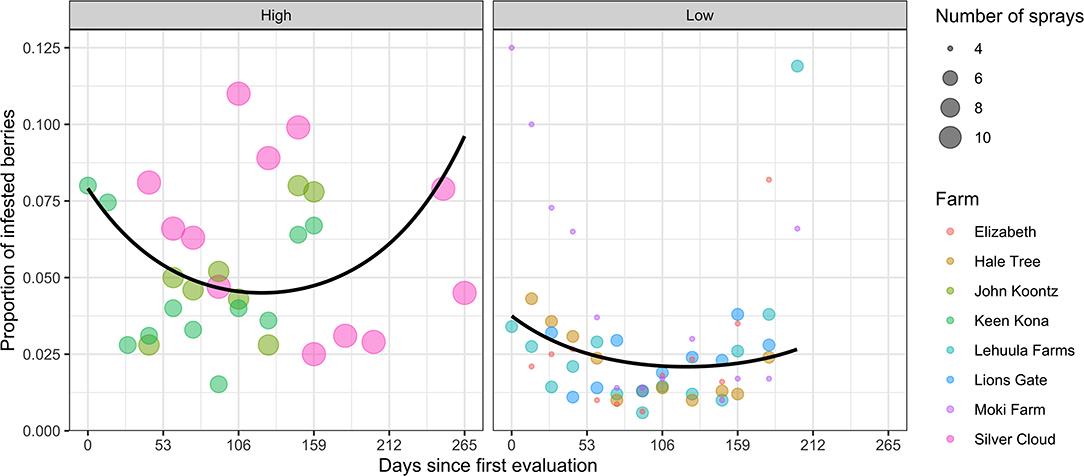
Figure 3. Proportion of CBB-infested berries from eight coffee farms in the Kona and Ka'u districts of Hawaii adopting calendar (high) and threshold (low) spray programs. Data points represent one observation, and curves are fitted means from generalized binomial mixed models with linear and quadratic random effects over time.
The average proportion of CBB with symptomatic B. bassiana sporulation across ranged from 0 to 60% in individual farms. Farms adopting threshold sprays recorded higher infection levels (18.3 ± 1.8%) compared with those using calendar sprays (10.8 ± 1.6%). Moreover, in threshold sprays, CBB infection rates were highest in the early season but declined to similar levels at the end of the season (Figure 4). The fitted models revealed an interaction (spray system × sample week) effect for infected CBB, based on different intercepts and slopes for each spray regime (LR = 4.09, d.f. = 1, P = 0.043), confirming that fungal infection rates are different between these system regimes.
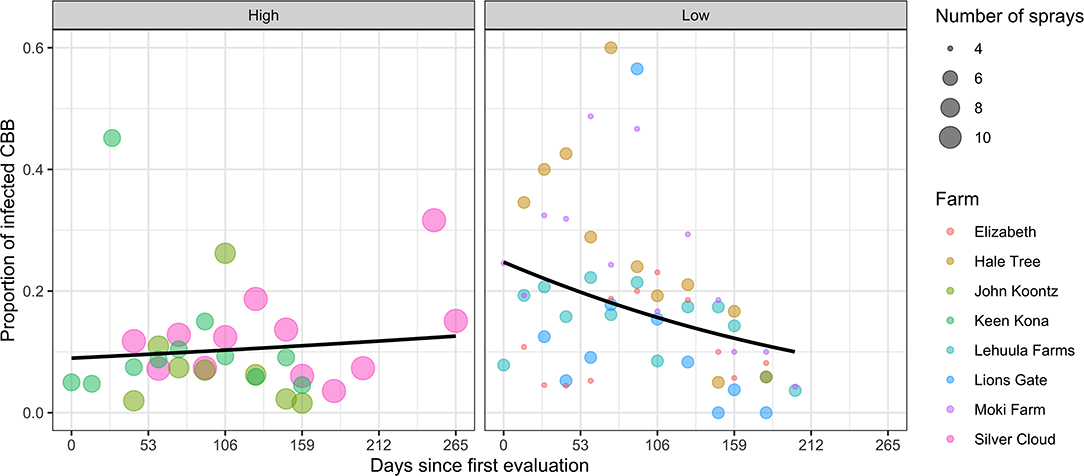
Figure 4. Proportion of CBB-infected with B. bassiana from coffee farms in the Kona and Ka'u districts of Hawaii adopting calendar (high) and threshold (low) spray programs. Data points represent one observation, and curves are fitted means obtained using generalized binomial mixed models with linear and quadratic random effects over time.
The position of CBB inside berries also changed during the season. Most notably, the proportions in the AB and CD position declined and increased with time, respectively (Figure 5). The proportion of CBB in these different categories varied among coffee farms (~F = 3.3, d.f. = 7, 231, P < 0.0001). In threshold-based farms, an approximate threefold increase in the number of CBBs in the CD position was observed at the end of the season. This suggests more late-season reproduction and may explain the corresponding late-season increase in the proportion of AB position (new infestations). The multivariate linear model revealed an interaction effect of spray system × time (F = 6.57, d.f. = 1, 75, P < 0.001). Overall CBB mortality also fluctuated over time. In calendar sprays, an upward trend was observed, compared with a downward trend in threshold sprays, which revealed an interaction (spray system × sample week) effect (LR = 4.53, d.f. = 1, P = 0.033) (Figure 6).
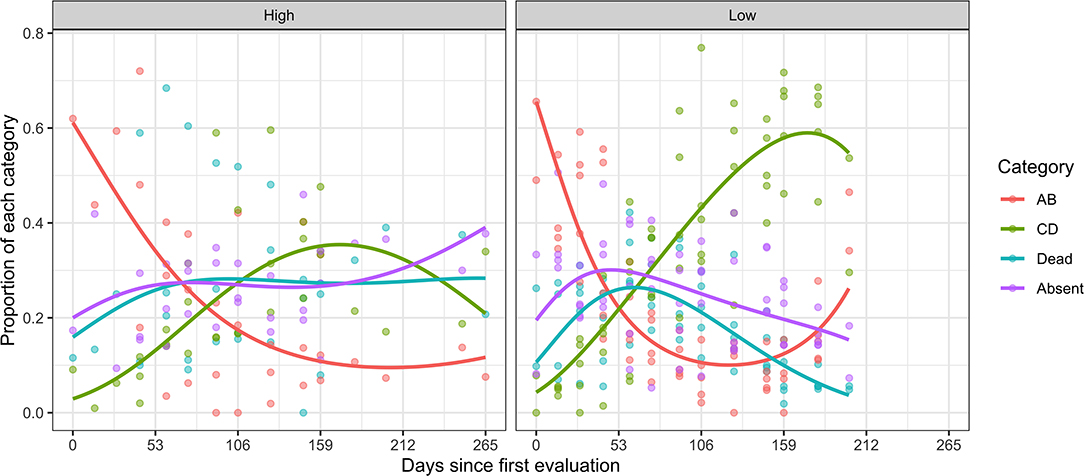
Figure 5. Distribution of the CBB found inside the damaged berries from coffee farms in the Kona and Ka'u districts of Hawaii adopting calendar (high) and threshold (low) spray programs. Data points represent the proportions of CBB in each category, per sample date. The curves were obtained using multivariate linear models based on category proportions, including linear, and quadratic effects of time.
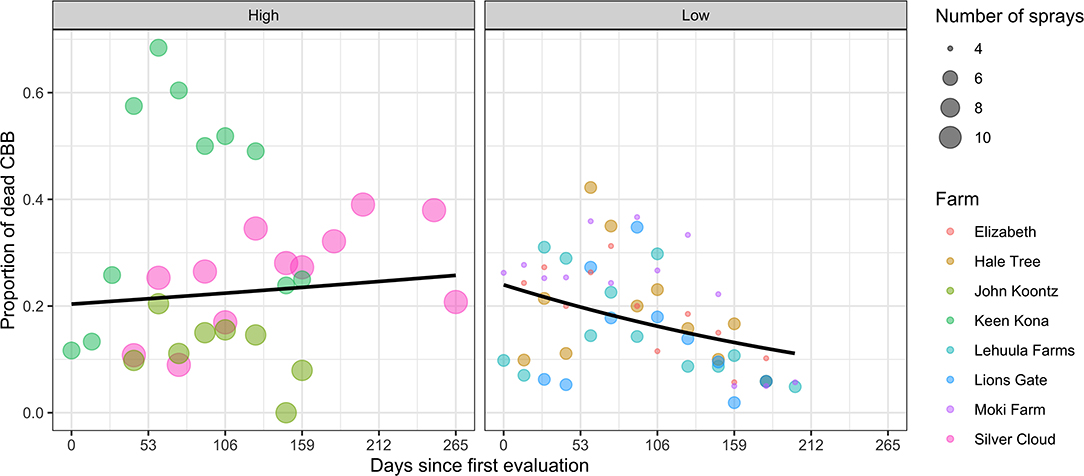
Figure 6. Overall CBB mortality from coffee farms in the Kona and Ka'u districts of Hawaii adopting calendar (high) and threshold (low) spray programs. Data points represent the proportion of CBB mortality for each sample date. Curves are the fitted means obtained using generalized binomial mixed models with linear and quadratic random effects over time.
Total defects in green processed coffee were similar among farms using threshold and calendar applications of B. bassiana (i.e., 8.5 ± 0.96% vs. 10.4 ± 1.7%, respectively). Overall, these differences in processed coffee were statistically similar between both spray systems (LR = 0.694, d.f. = 1, P = 0.405).
Seasonal costs for the CBB insecticide programs (products and labor) were less for threshold-based approaches (F = 7.12, d.f. = 1, 6, P = 0.04), even though the average cost (per spray application) was not different from the calendar-based approaches (F = 0.60, d.f. = 1, 6, P = 0.47) (Table 1). This difference in cost was therefore due to the lower number of applications per season. The yield (kg cherry/ha) was statistically similar between both programs (F = 0.74, d.f. = 1, 6, P = 0.42). Consequently, costs for CBB insecticide programs (based on yield) were proportionally lower in farms using threshold applications (Table 1).
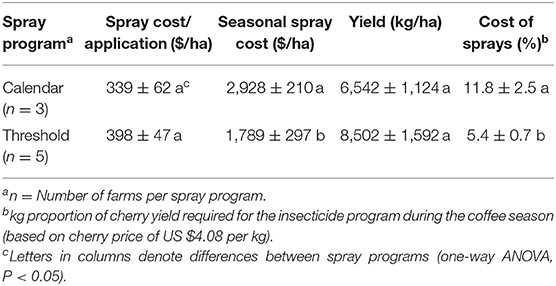
Table 1. Costs for spray program, production of coffee cherry, and relative cost needed to pay for insecticide applications, based on surveys in eight coffee farms in Kona and Ka'u districts, Hawaii, USA (2016/17 season).
Currently, management strategies for the CBB in Hawaii include pruning, sanitation of infested berries, and pesticide applications (Aristizábal et al., 2016, 2017b; Kawabata et al., 2017; Aristizábal, 2018b). Studies investigating additional control tactics, such as the release of CBB-predatory beetles (Follett et al., 2016) and the introduction of specific parasitoids (Follett, 2018), are ongoing. Among the few available insecticides in Hawaii, B. bassiana is compatible with environmental and worker safety concerns (Kawabata et al., 2017; Greco et al., 2018). Since many growers use calendar sprays (LFA observations), we assessed the effectiveness and cost of sprays based on field monitoring data.
Our field studies suggest that using thresholds can reduce the number of seasonal sprays. We observed that 4–5 applications of B. bassiana (alone or mixed with other products) mostly applied early in the season (April–July) provided equivalent CBB control when compared with calendar applications (7–11 sprays) at lower cost. Final defects in processed coffee were similar between farms employing each spray strategy. We hypothesize that this finding can be explained by the relationship between the effectiveness of sprays and seasonal phenology of the CBB.
The early season represents a critical time window for B. bassiana applications since founder females that migrate to new coffee berries are exposed to spray residues. During the early season, we observed higher proportions (20–70%) in AB position, where they are relatively exposed and more vulnerable to infection. Berry infestation rates were also higher during this time. Baker (1999) reported that CBB oviposition is initiated when infested berries have reached >150 days old and >20% of dry weight. Correspondingly, after July, most surviving founder CBBs had penetrated the berries. At this stage, CBBs were less vulnerable because beetles are protected inside the berry endosperm. This would explain why the proportion of CBB in CD position increased across coffee farms and why applications of B. bassiana after July were less effective.
Our field data suggest that pesticides alone may not be a sustainable strategy, since defects on processed coffee generally exceeded 5%, in our results. We noted that these estimates included damage by mechanical process (broken beans), deformations (shell beans), partial or full black beans, partial or full sour beans, fungus damage, and foreign matter. The increased berry infestation rates observed in most farms during the harvest season may be caused by subsequent CBB generations, including those arising from individuals surviving early-season insecticide treatments. Hamilton et al. (2019) reported between 2.1 and 4.9 generations of CBB per year in coffee farms spanning an elevational gradient (200–780 masl) from the Kona and Ka'u districts in Hawaii. The reproduction of the CBB in the mid-season may explain the increases in AB position at harvest, caused by a new CBB generation. This issue was exacerbated in the threshold-based sprays, possibly since few insecticides were applied, which may have increased mid- to late-season reproduction of the CBB. Late-season reestablishment of CBB provides a risk of increased harvest damage and CBB pressure on the following season (Johnson et al., 2019). This new generation of CBB needs to be targeted with insecticides and other strategies to prevent a carry-over of CBB the following season.
Cultural control provides an option to improve late-season CBB management. Several studies (Bustillo et al., 1998; Aristizábal et al., 2011) document that frequent harvesting (2–3 weeks intervals) and/or “strip picking” at the end of the season to remove remaining infested berries can be effective at reducing the carry-over of CBB to the next growing season (Aristizábal et al., 2017b). However, the high cost and shortage of available field labor are challenges faced by coffee growers in Hawaii employing crop sanitation (Aristizábal, 2018a). In our surveys, cultural control was not consistently employed across farms. Consequently, proliferation of CBB from infested berries left on trees or fallen on the ground after harvest (November–January) remains an issue for sustainable CBB management in the region.
Published recommendations for an IPM program designed for the CBB in Hawaii include field monitoring, use of B. bassiana and other insecticides, field sanitation, and post-harvest control (Kawabata et al., 2017; Aristizábal, 2018b). Various IPM strategies for the CBB have been recommended in other regions of the world, including Colombia (Bustillo et al., 1998; Benavides et al., 2002; Aristizábal et al., 2012), Mexico (Barrera et al., 1990), Brazil (Pereira et al., 2012; Olivera et al., 2018), and Africa (Kucel et al., 2009). While an IPM approach is needed, the relatively high cost of field labor in Hawaii remains a limitation for cultural pest control. The cost of B. bassiana may also increase after the federal or state subsidy ends in 2021 (State of Hawaii Plant Industry Division, 2016). Strategies to reduce sampling effort such as sequential sampling plans have been proposed (Aristizábal et al., 2018; Johnson et al., 2018; Pulakkatu-thodi et al., 2018).
In addition to CBB seasonal phenology noted above, the effectiveness of commercial formulations of B. bassiana depends on additional factors. For example, sporulation and secondary transmission of the fungus may depend on environmental variables (Mascarin and Jaronski, 2016). Greco et al. (2018) reported that B. bassiana infections were favored by higher-elevation locations in the Kona district, Hawaii. In Colombia, infection rates between 20 and 75% reflect variability in spray coverage, environmental conditions, and field label dosages (Bustillo et al., 1998, 1999). CBB integrated management should thus reflect the environmental factors in different production regions.
Our study provides evidence for the cost–benefit value of adopting spray thresholds for CBB management in Hawaii. Based on the cherry damage rates we observed, it also highlights the need for field sanitation and post-harvest control measures to mitigate the threat of CBB. Future efforts should investigate the value of such approaches, as well as taking into consideration the regional differences for B. bassiana sprays.
Overall, we observed that B. bassiana applications alone or in a tank mixed with other insecticides reduced CBB populations over the coffee season. Moreover, applications during the early coffee season (April–July) were more effective than applications during the second semester (August–November). This difference was likely due to the location of CBB females (AB position), which favored fungal contamination. While both spray programs (calendar and threshold) provided similar control, the threshold spray strategies was more cost-effective. However, neither spray strategy prevented economic damage to processed coffee. Therefore, additional control practices, such as cultural and sanitation methods, need to be incorporated in a sustainable IPM program for CBB.
The raw data supporting the conclusions of this article are available from the authors as Supplementary Material.
SS obtained the Technical Assistance to Specialty Crops (TASC) Grant and recruited coffee growers for this study. RH, LA, and SS coordinated fieldwork. LA generated research ideas, collected field data and farm information, took photographs, and wrote the first draft. RH and LA worked on the harvest, processing, and analysis of processed coffee quality. GM and RM performed statistical data analyses and generated figures. SA revised the manuscript for technical and scientific accuracy. LA, GM, RM, and SA wrote the final manuscript.
This funding was provided by the Synergistic Hawaii Agriculture Council (SHAC) TASC Grant for CBB (T16GXCBB01, Control of CBB in Hawaii). Additional funding for publication was obtained through a cooperative agreement with the USDA PBARC and SHAC.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
This paper is dedicated to RH (1957–2019, in memoriam) for pioneering work on the management of CBB in Hawaii. We thank the coffee growers from the Kona and Ka'u districts for their participation and support. We also thank John Ross, Melissa Johnson, Nicholas Manoukis, Tracie Masumoto, and Marisa Wall at the USDA-ARS Daniel K. Inouye US PBARC in Hilo, Hawaii, and the Cooperative Extension at the University of Hawaii at Manoa for technical support.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fsufs.2020.00022/full#supplementary-material
Table S1. CBB field infestations and control costs for individual farms.
Aristizábal, L. F. (2018a). Challenges faced by coffee growers establishing an integrated pest management for coffee berry borer in Hawaii. Agric. Res. Technol. 14:555919. doi: 10.19080/ARTOAJ.2018.14.555919
Aristizábal, L. F. (2018b). Controlling the Coffee Berry Borer Through Integrated Pest Management: A Practical Manual for Coffee Growers and Field Workers in Hawaii. Kailua-Kona, HI: Steuben Press.
Aristizábal, L. F., Bustillo, A. E., and Arthurs, S. P. (2016). Integrated pest management of coffee berry borer: strategies from latin America that could be useful for coffee farmers in Hawaii. Insects 7:6. doi: 10.3390/insects7010006
Aristizábal, L. F., Jimenez, M., Bustillo, A. E., and Arthurs, S. P. (2011). Monitoring cultural practices for coffee berry borer Hypothenemus hampei (Coleoptera: Curculionidae: Scolytinae) management in a small coffee farm in Colombia. Fla. Entomol. 94, 685–687. doi: 10.1653/024.094.0334
Aristizábal, L. F., Johnson, M., Shriner, S., Hollingsworth, R., Manoukis, N., Myers, R., et al. (2017b). Integrated pest management of coffee berry borer in Hawaii and Puerto Rico: current status and prospects. Insects 8:123. doi: 10.3390/insects8040123
Aristizábal, L. F., Lara, O., and Arthurs, S. P. (2012). Implementing an integrated pest management program for coffee berry borer in a specialty coffee plantation in Colombia. J. Integr. Pest Manage. 3, G1–G5. doi: 10.1603/IPM11006
Aristizábal, L. F., Shriner, S., Hollingsworth, R., and Arthurs, S. P. (2017a). Flight activity and field infestation relationships for coffee berry borer Hypothenemus hampei (Ferrari) in commercial coffee plantations in Kona and Kau districts, Hawaii. J. Econ. Entomol. 110, 2421–2427. doi: 10.1093/jee/tox215
Aristizábal, L. F., Shriner, S., Hollingsworth, R., Mascarin, G. M., Chaves, B., Matsumoto, T., et al. (2018). Field sampling strategies for coffee berry borer (Coleoptera: Curculionidae: Scolytinae) infesting berries in coffee farms in Hawaii. Int. J. Trop. Insect Sci. 38, 418–426. doi: 10.1017/S174275841800022X
Baker, P. S. (1989). A sampling plan for a control project against the coffee berry borer (Hypothenemus hampei) in Mexico. Trop. Pest Manage. 35, 169–172. doi: 10.1080/09670878909371350
Baker, P. S. (1999). The Coffee Berry Borer in Colombia. Final Report of the DFID – Cenicafé CABI Bioscience IPM for coffee project. Chinchiná: Centro Nacional de Investigaciones de Café. 154.
Barrera, J. F., Moore, D., Abraham, T. J., Murphy, S., and Prior, C. (1990). Biological control of the coffee berry borer, Hypothenemus hampei in Mexico and possibilities for further action. Brighton Crop Protect. Conf. Pest. Dis. 4, 391–396.
Bates, D., Maechler, M., Bolker, B., and Walker, S. (2015). Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67, 1–48. doi: 10.18637/jss.v067.i01
Benavides, P., Bustillo, A. E., Montoya, E. C., Cardenas, R., and Mejía, C. M. (2002). Participacion del control cultural, químico y biologico en el manejo de la broca del cafe. Rev. Colomb. Entomol. 28, 161–166.
Benavides, P., Góngora, C., and Bustillo, A. (2012). “IPM program to control coffee berry borer Hypothenemus hampei, with emphasis on highly pathogenic mixed strains of Beauveria bassiana, to overcome insecticide resistance in Colombia,” in Insecticides-Advances in Integrated Pest Management, 1st Edn, ed F. Perveen (Rijeka: InTech Europe), 512–540.
Bittenbender, H. C., and Smith, V. E. (2008). Growing coffee in Hawaii. College of Tropical Agriculture and Human Resources. University of Hawaii at Manoa. Revised Ed., 40.
Brooks, M. E., Kristensen, K., van Benthem, K. J., Magnusson, A., Berg, C. W., Nielsen, A., et al. (2017). glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 9, 378–400. doi: 10.32614/RJ-2017-066
Burbano, E., Wright, M., Bright, D. E., and Vega, F. E. (2011). New record for the coffee berry borer, Hypothenemus hampei, in Hawaii. J. Insect Sci. 11:117. doi: 10.1673/031.011.11701
Bustillo, A. E., Bernal, M. G., Benavides, P., and Chaves, B. (1999). Dynamics of Beauveria bassiana and Metarhizium anisopliae infecting Hypothenemus hampei (Coleoptera: Scolytidae) populations emerging from fallen coffee berries. Fla Entomol. 82, 491–498. doi: 10.2307/3496468
Bustillo, A. E., Cardenas, M. R., Villalba, D., Orozco, J., Benavides, M. P., and Posada, F. J. (1998). Manejo Integrado de la Broca del Café Hypothenemus Hampei (Ferrari) En Colombia. Cenicafé. Chinchiná: Centro Nacional de Investigaciones de Café. 134.
Damon, A. A. (2000). Review of the biology and control of the coffee berry borer, Hypothenemus hampei (Coleoptera: Scolytidae). Bull. Entomol. Res. 90, 453–465. doi: 10.1017/S0007485300000584
De La Rosa, W., Altorre, R., Barrera, J. F., and Torrello, C. (2000). Effect of Beauveria bassiana and Metarhizium anisopliae (Deuteromycetes) upon the coffee berry borer (Coleoptera: Scolytidae) under field conditions. J. Econ. Entomol. 93, 1409–1414. doi: 10.1603/0022-0493-93.5.1409
Follett, P. A. (2018). “Biological control of coffee berry borer: next steps, in Coffee Berry Borer Conference, Kailua-Kona, Hawaii, April 25, 2018.
Follett, P. A., Kawabata, A., Nelson, R., Asmus, G., Burt, J., Goschke, K., et al. (2016). Predation by at bark beetles (Coleoptera: Silvanidae and Laemophloeidae) on coffee berry borer (Coleoptera: Curculionidae) in Hawaii coffee. Biol. Control 101,152–158. doi: 10.1016/j.biocontrol.2016.07.002
Food Agriculture Organization of the United Nations Statistics Division (FAOSTAT) (2020). Avaliable online at: http://www.fao.org/faostat/en/#data (accessed February 1, 2020).
Greco, B. E., Wright, M. G., Burgueño, J., and Jaronski, S. T. (2018). Efficacy of Beauveria bassiana applications on coffee berry borer across an elevation gradient in Hawaii. Biocontrol Sci. Technol. 28, 995–1013 doi: 10.1080/09583157.2018.1493088
Hamilton, L. J., Hollingsworth, R. G., Sabado-Halpern, M., Manoukis, N. C., Follett, P. A., and Johnson, M. A. (2019). Coffee berry borer (Hypothenemus hampei) (Coleoptera: Curculionidade) development across an elevational gradient on Hawaii Island: applying laboratory degree-day predictions to natural field populations. PLoS ONE 14:e0218321. doi: 10.1371/journal.pone.0218321
Haraprasad, N., Niranjana, S. R., Prakash, H. S., Shetty, H. S., and Wahab, S. (2001). Beauveria bassiana a potential mycopesticide for the efficient control of coffee berry borer, Hypothenemus hampei (Ferrari) in India. Biocontrol Sci. Technol. 11, 25–26. doi: 10.1080/09583150120035675
Infante, F. (2018). Pest management strategies against the coffee berry borer (Coleoptera: Curculionidae: Scolytinae. J. Agr. Food Chem). 66, 5275–5280. doi: 10.1021/acs.jafc.7b04875
International Coffee Organization (ICO) (2019). Trade Statistics. Avaliable online at: http://www.ico.org/trade_statistics.asp (accessed May 26, 2019).
Jaramillo, J., Borgemeister, C., and Baker, P. (2006). Coffee berry borer Hypothenemus hampei (Coleoptera: Curculionidae): searching for sustainable control strategies. Bull. Entomol. Res. 96, 223–233. doi: 10.1079/BER2006434
Johnson, M. A., Fortna, S., Hollingsworth, R. G., and Manoukis, N. C. (2019). Postharvest population reservoirs of coffee berry borer (Coleoptera: Curculionidae) on Hawai'i Island. J. Econ. Entomol. 112, 2833–2841. doi: 10.1093/jee/toz219
Johnson, M. A., Hollingsworth, R., Fortna, S., Aristizábal, L. F., and Manoukis, N. C. (2018). The Hawaii protocol for scientific monitoring of coffee berry borer: a model for coffee agroecosystems worldwide. J. Vis. Exp. 133:57204. doi: 10.3791/57204
Kawabata, A. M., Nakamoto, S. T., and Curtiss, R. T. (2017). Recommendations For Coffee Berry Borer Integrated Pest Management In Hawai'i 2016. University of Hawaii: Mänoa publication. Available online at: http://www.ctahr.hawaii.edu/oc/freepubs/pdf/IP-33.pdf
Kucel, P., Kangire, A., and Egonyu, J. P. (2009). Status and Current Research Strategies for Management of the Coffee Berry Borer (Hypothenemus Hampei Ferr) in Africa. Mukono: Coffee Research Centre; National Crop Resources Research Institute (NaCRRI). 9.
Mascarin, G. M., and Jaronski, S. T. (2016). The production and uses of Beauveria bassiana as a microbial insecticide. World J. Microb. Biot. 32:177. doi: 10.1007/s11274-016-2131-3
Moral, R. A., Hinde, J., and Demétrio, C. G. B. (2017). Half-normal plots and overdispersed models in R: the hnp package. J. Stat. Softw. 81, 1–23. doi: 10.18637/jss.v081.i10
Olivera, C. M., Santos, M. J., Amabile, R. F., Frizzas, M. R., and Bartholo, G. F. (2018). Coffee berry borer in conilon coffee in the Brazilian cerrado: an ancient pest in a new environment. Bull. Entomol. Res. 108, 102–107. doi: 10.1017/S0007485317000530
Pantoja-Gomez, L. M., Corrêa, A. S., de Oliveira, L. O., and Guedes, R. N. C. (2019). Common origin of brazilian and colombian populations of the neotropical coffee leaf miner, Leucoptera coffeella (Lepidoptera: Lyonetiidae). J. Econ. Entomol. 112, 924–931. doi: 10.1093/jee/toy416
Pereira, A. E., Vilela, E., Tinoco, R., de Lima, J. O. G., Fantine, A. K., Morais, E. G. F., et al. (2012). Correlation between numbers captured and infestation levels of the coffee berry-borer, Hypothenemus hampei: a preliminary basis for an action threshold using baited traps. J. Pest Manage. 58, 183–190. doi: 10.1080/09670874.2012.676219
Pulakkatu-thodi, I., Gutierrez-Coarite, R., and Wright, M. G. (2018). Dispersion and optimization of sequential sampling plans for coffee berry borer (Coleoptera: Curculionidae) infestations in Hawaii. Environ. Entomol. 47, 1306–1313. doi: 10.1093/ee/nvy098
R Core Team (2019). A Language and Environment for Statistical Computing. Vienna, Austria: R Foundation for Statistical Computing 2012. Avaliable online at: https://www.R-project.org (accessed December 20, 2019).
Sponagel, K. W. (1994). La broca del Café Hypothenemus Hampei En Plantaciones de Café Robusta en la Amazonia Ecuatoriana. Giessen Wissenschaftlicher, Fachverlag, 192.
State of Hawaii Plant Industry Division (2016). CBB Pesticide Subsidy Program. Available online at: http://hdoa.hawaii.gov/pi/main/cbbsubsidy/ (accessed November 8, 2019).
Talhinhas, P., Batista, D., Diniz, I., Vieira, A., Silva, D. N., Loureiro, A., et al. (2017). The coffee leaf rust pathogen Hemileia vastatrix: one and a half centuries around the tropics. Molec. Plant Pathol. 18, 1039–1051. doi: 10.1111/mpp.12512
Teuber, R. (2010). Geographical indications of origin as a tool of product differentiation: the case of coffee. J. Int. Food Agribus Market. 22, 277–298. doi: 10.1080/08974431003641612
Trujillo, E. H. I., Aristizábal, A. L. F., Bustillo, P. A. E., and Jiménez, Q. M. (2006). Evaluación de métodos para cuantificar poblaciones de broca del café, Hypothenemus hampei (Ferrari) (Coleoptera: Curculionidae: Scolytinae) en fincas de caficultores experimentadores. Rev. Colomb. Entomol. 32, 39–44. Available online at: www.scielo.org.co
USDA-NASS (2017). Hawaii Coffee Marketing Preliminary Season Estimates February 1st, 2017. Available online at: https://www.nass.usda.gov (accessed May 26, 2019).
Vega, F. E., Infante, F., Castillo, A., and Jaramillo, J. (2009). The coffee berry borer, Hypothenemus hampei ((Ferrari) (Coleoptera: Curculionidae): a short review, with recent finding and future research directions. Terr. Arthr. Rev. 2, 129–147. doi: 10.1163/187498209X12525675906031
Vega, F. E., Infante, F., and Johnson, A. J. (2015). “The genus Hypothenemus, with emphasis on H. hampei, the coffee berry borer,”in Bark Beetles, Biology and Ecology of Native and Invasive Species, 1st Edn, eds F. E. Vega and R. W. Hofstetter (London: Elsevier), 427–494. doi: 10.1016/B978-0-12-417156-5.00011-3
Vera, J. T., Montoya, E. C., Benavides, P., and Góngora, C. E. (2011). Evaluation of Beauveria bassiana (Ascomycota: Hypocreales) as a control of the coffee berry borer Hypothenemus hampei (Coleoptera: Curculionidae: Scolytinae) emerging from fallen infested coffee berries on the ground. Biocontrol Sci. Technol. 21, 1–14. doi: 10.1080/09583157.2010.517605
Woodill, A. J., Hemachandra, D., Nakamoto, S. T., and Leung, P. (2014). The economics of coffee production in Hawai‘i. Econ. Issues 25, 1–9. Available online at: www.ctahr.hawaii.edu
Wraight, S. P., Galaini-Wraight, S., Howes, R. L., Castrillo, L. A., Carruthers, R. I., Smith, R. H., et al. (2018). Prevalence of naturally-occurring strains of Beauveria bassiana in populations of coffee berry borer Hypothenemus hampei on Hawai'i Island, with observations on coffee plant-H. hampei-B. bassiana interactions. J. Invertebr. Pathol. 156, 54–72. doi: 10.1016/j.jip.2018.07.008
Keywords: biological control, Hypothenemus hampei, mycoinsecticide, Coffea arabica, coffee farms
Citation: Hollingsworth RG, Aristizábal LF, Shriner S, Mascarin GM, Moral RA and Arthurs SP (2020) Incorporating Beauveria bassiana Into an Integrated Pest Management Plan for Coffee Berry Borer in Hawaii. Front. Sustain. Food Syst. 4:22. doi: 10.3389/fsufs.2020.00022
Received: 04 November 2019; Accepted: 17 February 2020;
Published: 27 March 2020.
Edited by:
Jimmy R. Botella, University of Queensland, AustraliaReviewed by:
Agnieszka Barbara Najda, University of Life Sciences of Lublin, PolandCopyright © 2020 Hollingsworth, Aristizábal, Shriner, Mascarin, Moral and Arthurs. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Luis F. Aristizábal, bGFyaXN0aXphYmFsNzIxQGdtYWlsLmNvbQ==
†These authors have contributed equally to this work
‡Deceased February 2019
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.