
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Sustain. Food Syst. , 20 February 2020
Sec. Sustainable Food Processing
Volume 4 - 2020 | https://doi.org/10.3389/fsufs.2020.00014
 Marika Pellegrini1†‡
Marika Pellegrini1†‡ Chiara Rossi1‡
Chiara Rossi1‡ Sara Palmieri1
Sara Palmieri1 Francesca Maggio1
Francesca Maggio1 Clemencia Chaves-López1
Clemencia Chaves-López1 Claudio Lo Sterzo1
Claudio Lo Sterzo1 Antonello Paparella1
Antonello Paparella1 Dario De Medici2
Dario De Medici2 Antonella Ricci1*
Antonella Ricci1* Annalisa Serio1*
Annalisa Serio1*Chemical disinfectants represent one of the commonly used practice in minimally processed vegetables food-chain. However, the scarce safety and sustainability of these agents force food industry to move toward more sustainable “green washing solutions.” Among the latter, while the application of plant derivates for the control of several pathogens is already well-known, the potential anti-Salmonella activity of Coriandrum sativum seeds derivates is still unexplored and was therefore investigated in this study. In detail, Minimal Inhibitory Concentration (MIC) and Minimal Bactericidal Concentration (MBC) of different coriander seed derivates (i.e., essential oil, hydrosol, and ethanolic extract) were determined by broth dilution against six Salmonella enterica strains isolated from fresh and minimally processed fruits and vegetables. Only the essential oil (EO) was effective in vitro with strain-dependent results. In addition, when mixed in co-culture, the strains were more sensitive to the essential oil treatment. Chemical investigations allowed to define (s)-(+)-linalool as major compound in the essential oil, and to underline interesting phenolic content with correlated antioxidant capacity. A cocktail of three strains of different serovars was selected and employed for a preliminary in situ trial on stick carrots. The obtained results allowed to establish that the application of coriander seed EO at concentrations of 5 μL mL−1 was able to reduce and contain the growth of the Salmonella cocktail up to 24 h at 10°C. Good sensory evaluation results were obtained by applying this EO concentration as washing treatment, especially in terms of color parameter. Further studies should be undertaken to emphasize the upstream activity, improving the formulation or exploiting a combined effect with other sanitizers or treatments (e.g., physical treatments). The present study contributes to the knowledge on coriander derivates activity against Salmonella spp. and on the potential application as sustainable washing treatment in removing this pathogen from fresh cut carrots.
Minimally processed vegetables (MPV) are defined as fresh, raw vegetables processed to supply ready-to-eat or ready-to-use foods (Nguyen-the and Carlin, 1994). These products can harbor a variety of spoilage microorganisms and a particular concern is related to the potential presence of cold-tolerant pathogens such as Listeria monocytogenes, Escherichia coli, Bacillus cereus, Aeromonas hydrophila, Yersinia enterocolitica, and Salmonella spp. Among them, Salmonella spp. represent a significant public health concern, being responsible of several foodborne outbreaks (Cai et al., 2016; Rossi et al., 2019). This food-borne pathogen, usually linked to poultry meat and products, is also found in a wide variety of fruits and vegetables (Pui et al., 2011) such as carrots (Daucus carota) (Gutiérrez-Alcántara et al., 2016; Liu et al., 2019) and fresh unpasteurized carrot juice (Torres-Vitela et al., 2013).
The major risks of Salmonella spp. contamination in the food industry are related to their occurrence in plants. In addition, strains belonging to this genus form biofilms on many food contact surfaces of processing equipment (e.g., cutting boards, knives) with consequent cross-contamination of vegetables (Lo Fo Wong et al., 2002; Doulgeraki et al., 2016). Effective intervention methods able to reduce Salmonella infections are based on washing and sanitization procedures, principally utilizing chemical disinfectants (e.g., chlorinated water, Silveira et al., 2017; Ramos et al., 2020). However, these chemical agents present several disadvantages in terms of safety and sustainability. Food industry should move away from these chemical disinfectants, therefore the implementation of washing steps with effective “green washing solutions” is of great importance (Gil et al., 2009).
Among these sustainable and innovative alternatives, essential oils (EOs), their distillation co-products (hydrosols) and extracts obtained from various plant materials have been investigated for their use in the food sector, since many of them have shown to possess antimicrobial, antioxidant and food preservative properties (Paparella et al., 2016; D'Amato et al., 2018), including interesting potentials in reducing Salmonella spp. on fruit and vegetables (Raybaudi-Massilia et al., 2006; Gündüz et al., 2009; da Silva et al., 2016; Rossi et al., 2019).
In one of our previous in vitro study (Pellegrini et al., 2018) thyme, savory, oregano and coriander seeds EOs obtained from Abruzzo territory cultivations, expressed an interesting anti-Salmonella activity. The majority of these EOs, however, can alter the taste of food or exceed acceptable flavor thresholds (Gutierrez et al., 2009; Dolati et al., 2016) due to strong balsamic notes of thymol (Calín-Sánchez et al., 2013, 2015). The high concentration of linalool (75%) in coriander seeds EO, on the other hand, may suggest mild aromatic characteristics more suitable for food application.
Research reports on composition, antioxidant and antimicrobial activity of Coriandrum sativum seeds extracts are limited, especially concerning food processing applications. Thus, the aims of the present work were to (i) enrich the knowledge regarding in vitro anti-Salmonella activity of coriander seeds derivates and to (ii) investigate their potential application as washing treatment on stick carrots MPV.
C. sativum seeds, retrieved from a local farmer in Abruzzo, were subjected to different extraction procedures to obtain coriander seed EO (CEO), hydrosol (CH), and extract (CX).
The CEO was steam-distilled from the whole seeds (1 kg) without any further processing, by means of a E0105 12 lt PLUS Essential Oils Extractor (Albrigi Luigi Srl, Italy) for 2 h. After distillation, CEO was transferred to an amber glass vial with anhydrous sodium sulfate (Sigma Aldrich, USA), conditioned with argon and sealed. The CH was recovered from the same extraction process (i.e., remaining aqueous fraction after EO removal) and transferred to sterile tubes. Both collected CEO and CH were stored under refrigeration at 4°C. The distillation was conducted in triplicate.
The CX was obtained through rapid solid-liquid dynamic extraction (RSLDE) performed by means of 500 mL Naviglio extractor® (Atlas Filtri, Padua, Italy). Ten hundred g of the matrix were subjected to the RSLDE process with 250 mL of absolute ethanol (using a reduction chamber unit) as follows: 30 cycles (with a maximum pressure of 8 bar), each cycle composed by 12 hits in the dynamic phase (2 min duration) and a duration of the static phase of 2 min. At the end of the extraction process, the ethanol extract was filtered, collected in a pear-shaped evaporation flask and the solvent was removed by means of the Laborota 4,000 rotary evaporator (Heidolph–Schwabach, Germany). The obtained CX was transferred to an amber glass vial and stored under refrigeration at 4°C. The extraction was performed three times.
Six Salmonella enterica of different serovars were used in the study: S. Derby, S. Thompson, S. Napoli and S. Typhimurium monophasic variant, S. Kasenyi and S. Veneziana were isolated from fresh and minimally processed fruits and vegetables as previously described (Losio et al., 2015; Rossi et al., 2019). Salmonella strains were obtained from stock cultures maintained at −80°C in cryovials in 20% v/v glycerol/tryptic soy broth (TSB, Oxoid Thermofisher, Rodano. Italy). All strains were cultivated routinely on TSB at 37°C and stored at 4°C on tryptic soy agar slants.
C. sativum derivatives were studied for their MIC values according to the microdilution method, as described by CLSI guidelines (Clinical and Laboratory Standards Institute, 2011). The CEO was dissolved in sterile PBS (Phosphate Buffer Saline) 50 mM pH 7.0 to reach the initial concentration of 40 μL mL−1, as already described (Mazzarrino et al., 2015). The hydrosol was used as it stands, while the evaporated RSLDE extract was diluted with distilled water (40 mg mL−1). The inocula were prepared from overnight broth cultures (early stationary growth phase), and suspensions were adjusted to the required microbial load (5 × 105 CFU mL−1). A positive (100 μL of TSB plus 100 μL inoculum) and a negative control (200 μL of sterile TSB) were considered for each strain. The MIC value was considered as the lowest EO concentration that prevented growth after 48 h of incubation at 37°C. The MBC, the lowest concentration of the essential oil at which incubated microorganisms are completely killed, were confirmed by re-inoculating on agar plates with 10 μL of each culture medium from the microplates.
MBCs and further investigations were carried out only on CEO, as for the other derivates not satisfying MIC values were recorded. These assays were performed for all six strains in single culture and for some strains in co-culture.
According to MIC/MBC results, the 2 most sensitive serovars were selected and coupled with two other serovars. The cocktails were constituted as follows: cocktail 1 (C1): S. Thompson, S. Derby and S. Napoli; cocktail 2 (C2): S. Thompson, S. Derby and S. Typhimurium. All the analyses were performed in triplicate.
A turbidimetric analysis was conducted to evaluate inactivation/growth kinetics of the strains in presence of different CEO concentrations. Inocula were prepared as described in section Determination of Minimal Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC) and incubated at 37°C. The plates were automatically scanned and recorded every 15 min for 2 days by the OmniLog incubator/reader (Biolog Inc., Hayward, USA). The scanning technology used by Omnilog records the color change in the well as digital Omnilog units. Data were recorded using the OmniLog PM software provided by the same company and then exported into Microsoft Excel Professional for further data processing. The values were expressed as mean of replicated growth curves. The experimental data were fitted according to the equation of Baranyi and Roberts (1994), by means of DMFit software (https://browser.combase.cc/DMFit.aspx) to estimate growth parameters such as lag phase length and maximum growth value, as previously described (Mazzarrino et al., 2015).
The CEO was investigated for its chemical composition through GC-MS analysis, following the procedure previously described by Pellegrini et al. (2018).
The antioxidant capacity (TEAC/ABTS, DPPH, FRAP) and total phenolic content (TPC) were also investigated.
The TEAC/ABTS assay was determined as described by Masaldan and Iyer (2011). The TEAC/ABTS results of the samples were estimated in terms of mg Trolox equivalent (TE)/g EO as the mean of three replicates.
The FRAP was determined by using the potassium ferricyanide-ferric chloride method described by Oyaizu (1986). The FRAP of the samples was estimated in terms of mg Trolox equivalent (TE)/g EO as the mean of three replicates.
The DPPH radical-scavenging activity of the EOs methanolic solutions was measured according to the method described by Brand-Williams et al. (1995). Results were expressed in terms of mg Trolox equivalent (TE)/g EO as the mean of three replicates.
The TPC was determined by the Folin–Ciocâlteu method described by El-Lateef Gharib and Teixeira da Silva (2013). The TPC results were expressed in terms of mg Gallic acid equivalents (GAE)/g EO as the mean of three replicates.
Fresh carrots (Daucus carota L.) were purchased at a local market and those with defects were discarded. Selected carrots were washed to reduce the native spoiling flora as reported by Martínez-Hernández et al. (2017); then, the carrot samples were manually peeled by a hand potato peeler and cut into 4 × 0.3 cm sticks.
Three Salmonella serovars were used in the study as a cocktail. In particular, the inoculum of S. Thompson, S. Derby and S. Napoli was standardized in PBS at about 5 × 105 CFU mL−1. The carrot samples were aseptically submerged into Salmonella suspension for 30 min at room temperature (Ruiz-Cruz et al., 2007) and stand to dry 30 min in a class II biological safety cabinet. CEO was diluted in sterile PBS as described in section Determination of Minimal Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC) to reach the final concentration of 5 μL mL−1. The carrot samples were aseptically transferred into an open sterile glass jar, containing the CEO solution. Also control samples were included, dipping the carrot samples in the same way in PBS alone. All samples were dipped into the appropriate solution for 2 min, then the excess solution was drained off and samples were air-dried for 30 min in a biological safety cabinet (Gonzalez et al., 2004). Carrots sticks were aseptically placed in polyethylene bags to prevent water loss and stored under aerobic conditions for 48 h at 10°C (abuse temperature).
The samples (25 g) were aseptically transferred in stomacher bags containing 225 mL of sterile saline solution, homogenized in stomacher (Lab Blender 400, Seward Medical, UK) for 5 min, then serial decimal dilutions were prepared in sterile saline solution. Salmonella counts were determined using Xilose Lysine Deoxicholate agar–XLD (Oxoid–Rodano, Italy), after incubation at 37°C for 24 h. The analyses were carried out in duplicate at the following intervals: 0, 1, 24, 48 h, considering time 1 h as time after the application of the treatment. To assess the absence of Salmonella spp. in non-inoculated stick carrots, control samples were included and followed for the same time of storage.
Sensory evaluation was performed as proposed by Cui et al. (2017). 30 people (15 males and 15 females) aged 26–60 years were recruited from the staff of the University of Teramo. Control and treated carrot sticks were served at room temperature separately. Unsalted crackers and mineral water (room temperature) were provided to clean the palate between samples. The hedonic scale consisted of 9 levels (1: dislike extremely/ extremely bland and 9: like extremely/extremely flavorful), in which the panelists evaluated the different attributes (i.e., color, aroma, taste, and overall acceptability).
Microbiological counts were converted to Log CFU g−1 and all results were expressed as mean ± standard deviation. Data were subjected to analysis of variance (ANOVA) and pair-comparison with the same group was achieved applying Tukey's post-hoc test procedure at p < 0.05. Correlations between TPC and AOC results were calculated by means of Pearson Correlation. Statistical tests were carried out using XLSTAT 2017 (Addinsoft, Paris, France).
The C.sativum seeds extracts were tested for their antimicrobial activity against six S. enterica strains of different serovars, previously isolated from vegetables, and belonging to the most recently isolated Salmonella serotypes in Italy in this kind of products (Rossi et al., 2019).
The results obtained from MIC/MBC assay were reported in Table 1. As showed, among the tested extracts, the CEO was the only one able to exert an antibacterial activity against S. enterica serovars, with a strain-dependent sensitivity. Indeed, S. Typhimurium and S. Kasenyi showed high resistance to treatment with CEO, whose MIC and MBC values (μL mL−1) were higher than the maximum concentration employed in the analysis (20 μL mL−1). Otherwise, the most sensitive strains were S. Derby and S. Thompson, for which the effect was both bacteriostatic and bactericidal at a concentration of 5 μL mL−1. The essential oil effectiveness was also observed for S. Napoli and S. Veneziana, however the inhibitory concentrations of CEO for the two strains were 20 μL mL−1.
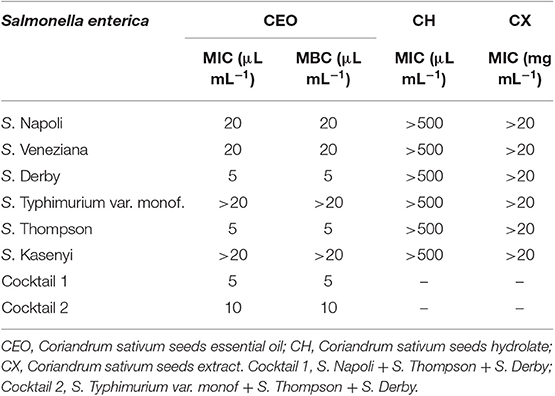
Table 1. Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC) of Coriandrum sativum extracts against single strains and cocktails of Salmonella enterica.
In order to employ a cocktail consisting of 3 Salmonella strains for an in situ study, the evaluation of the MIC and MBC of co-cultured microorganisms was performed. Specifically, following the MIC/MBC results, the two most CEO sensitive serovars (S. Derby and S. Thompson) were selected and both S. Napoli and S. Typhimurium were tested as additional strains.
Table 1 shows also the results of the MIC and MBC values of coriander EO on Salmonella strains co-cultures. The comparison clearly shows that Salmonella serovars inoculated in ternary cultures, are more sensitive to the treatment with CEO. S. Napoli in fact, was individually inhibited at the 20 μL mL−1 CEO concentration, nevertheless, when co-cultured in presence of S. Thompson and Derby (Cocktail 1) a concentration of 5 μL mL−1 was able to exert bactericidal effect. A similar behavior was observed for S. Typhimurium; while in single culture, a CEO concentration >20 μL mL−1 was requested for the inhibition, only 10 μL mL−1 concentration were sufficient to inhibit C2 co-culture. According to these results, C1 co-culture (S. Napoli + S. Thompson + S. Derby) was selected for in situ testing.
In Figures 1A,B the inactivation/growth kinetics of the two assayed cocktails (C1 and C2, respectively) were presented.
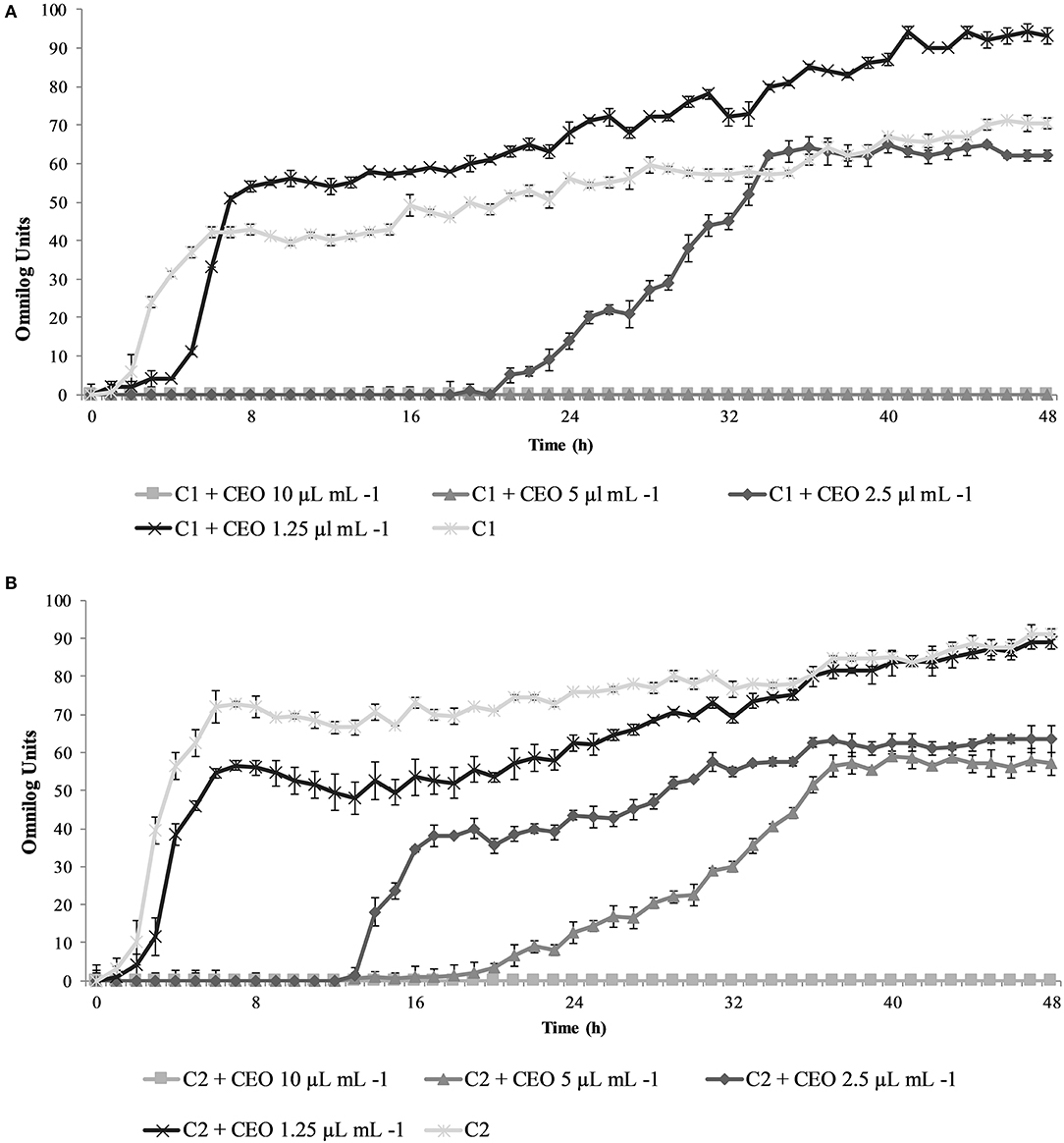
Figure 1. Growth/inhibition dynamics of Salmonella strains in co-cultures, in presence of different concentrations of Coriandrum sativum seeds essential oil (μL mL−1) during 48 h of incubation at 37°C. (A) S. Thompson, S. Derby and S. Napoli co-colture (C1); (B) S. Thompson, S. Derby and S. Typhimurium co-culture (C2).
Regarding C1, as displayed in Figure 1A, from the wells where different concentrations of CEO were present, different kinetics with respect to the control (C1) were recorded. In particular, at 10 and 5 μL mL−1 no growth was observed along the 48 h of analysis (lethal concentrations); at 2.5 μL mL−1 concentrations, a Lag phase extension was detected (22 h with respect to 1.1 hour observed for control). In presence of 1.25 μL mL−1, after a slight initial Lag phase elongation of about 4 h, a subsequent increase of microbial growth was detected, being the maximum growth value of 87.4 Omnilog Units in spite of 64.8 Omnilog Units observed for C1, according to the data modeling.
In reference to C2 (Figure 1B), only for 10 μL mL−1 a lethal effect was observed. The Lag phase extension was greater at concentration of 5 μL mL−1 (21.8 h), followed by 2.5 μL mL−1 (9.6 h); both concentrations also determined lower maximum growth values (59 and 61.5 Omnilog Units respectively) than the control (86 Omnilog Units), confirming the antimicrobial action of CEO at those concentrations. On the contrary, 1.25 μL mL−1 CEO concentration seemed to exert very slight effects on the C2 cocktail growth; its recorded kinetic in fact, was almost similar to the control one, with a sharp increase in the first hours of incubation and a similar final growth value.
Table 2 reports the chromatographic characterization by GC-MS of CEO, which allowed to identify the 95% of the volatile mixture. From the obtained results, it was underlined that CEO was constituted essentially by monoterpenes. In particular, (S)-(+)-linalool accounted for the 78% of the total volatile mixture; the remaining 17% of identified compounds was composed only by five other monoterpenes, with a major contribution of trans-β-ocimene, cis-geraniol, and camphor.
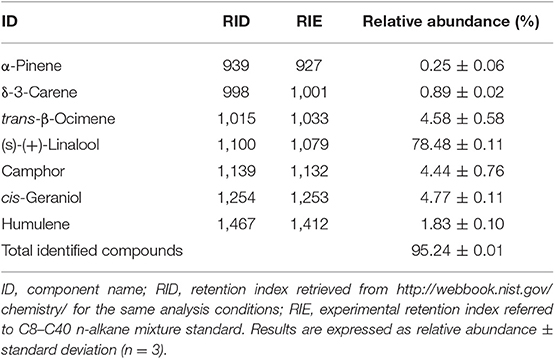
Table 2. Gas Chromatography-Mass Spectrometry (GC-MS) characterization of Coriandrum sativum seeds essential oil.
The antioxidant activity assays demonstrated interesting results (Table 3), in terms of radical scavenging activity (TEAC/ABTS and DPPH) and ferric reducing antioxidant power (FRAP). These results were also strongly positively correlated (r > 0.9; p < 0.05) with total phenolic content.
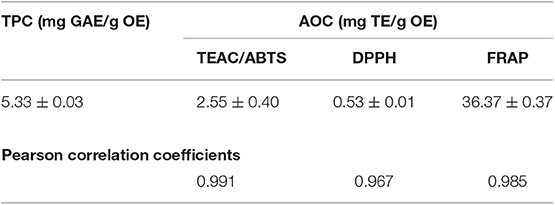
Table 3. Coriandrum sativum seeds essential oil: Total Phenolic Content (TPC) estimation, by Folin-Ciocâlteu method, and Antioxidant Activity Capacity (AOC) assays by Trolox Equivalent Antioxidant Capacity with 2,20-azinobis-(3-ethylbenzothiazoline-6-sulfonic acid (TEAC/ABTS), 2,2-diphenyl-1-picrylhydrazyl (DPPH) and Ferric Reducing Antioxidant Power (FRAP).
The results of the preliminary study about CEO anti-Salmonella activity on stick carrots are displayed in Figure 2. First of all, a slight “matrix effect” was recorded: in fact, at time 1, the not treated inoculated samples (C), showed a difference in microbial load of about 0.8 Log with respect to the inoculum. This effect could be related to the specific complexity of food matrix, that interfere with bacterial cells. Furthermore, in untreated control samples (C), the counts of Salmonella were almost stable during the storage time.

Figure 2. Evolution of Salmonella counts during refrigerated storage of stick carrots (48 h, 10°C) samples treated with 5 μL mL−1 Coriandrum sativum seeds essential oil and in control ones. For the same assay results followed by the same case-letter are not significantly different according to Tukey' HSD post hoc test (p > 0.05).
From Figure 2 it is also possible to infer how immediately after performing the treatment with the essential oil (1 h), the application of CEO on carrots determined a significant reduction (p < 0.05) in S. enterica of about 1 Log CFU g−1. After 24 h, the difference between treated samples and control samples still persist, while after 2 days this reduction was lost, with counts comparable to those of the control.
The analysis of uninoculated carrots revealed the absence of Salmonella spp.
Sensory evaluation results were reported in Table 4. As can be observed, any negative alteration was observed among treated and untreated samples in terms of aroma, taste and overall acceptability. For these parameters no significant differences (p > 0.05) were recorded between the investigated samples. Color was positively affected by CEO washing treatment, obtaining higher scores (p < 0.05) with respect to the control.
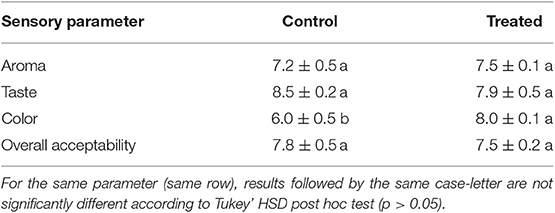
Table 4. Sensory evaluation results obtained for stick carrots treated with Coriandrum sativum seeds essential oil washing treatment and for control stick carrots.
The results of the present study showed that, among the C. sativum seeds derivatives, the only effective, at the tested concentrations, was the essential oil.
This better anti-Salmonella activity could be due to the chemical compounds contained in the essential oil. As reported in literature, (s)-(+)-Linalool, the major chemical component of CEO, has a remarkable antimicrobial activity (Aelenei et al., 2019), but the antimicrobial effect of the essential oil was probably due not only to its contribution. In fact, it is well-known that different biological activities of plant derivatives can be a function of synergistic interactions among major and minor compounds (Moon et al., 2006; Lima Oliveira et al., 2018). In example, camphor is present in other herbs and essential oils (basil, marjorane, rosemary, sage), and it is known for its antimicrobial activity, related to the ability to disturb the membrane phospholipid bilayer and to interact with enzyme and proteins (Bouazama et al., 2017). Its presence in low percentages in the studied coriander essential oils could boost (s)-(+)-linalool activity, still remaining within the safety daily doses established by EFSA (2008).
The chemical composition results obtained for the CEO were in accordance with literature data (Lo Cantore et al., 2004; Mandal and Mandal, 2015). A similar chemical composition was also obtained in our previous study (Pellegrini et al., 2018) for an essential oil recovered from coriander seeds purchased from the same local producer, but belonging to a different seeds batch. A different situation emerged when antioxidant activity and total phenolic content results were compared with those obtained in this previous study (Pellegrini et al., 2018). In fact, the essential oil recovered from the new batch of seeds gave better results. In fact, although the chemical composition is very similar, small variations can have an impact on the biological activity of the essential oils, and particularly the minor components with synergistic effects with major ones (Lima Oliveira et al., 2018). Further comparison of our results with literature data was not possible due to the different origin of vegetal matrix, AOC assays procedures and expression of results. However, the values obtained were comparable with those usually reported for essential oils (Ruiz-Navajas et al., 2013; Wang et al., 2017; Yashin et al., 2017).
The tested in vitro anti-Salmonella activity was quite interesting, even considering only the serovars singularly. The antimicrobial activity of coriander seed essential oils with similar (s)-(+)-linalool content has been previously investigated by different authors (Burdock and Carabin, 2009). In the study of Ildiz et al. (2018), the growth inhibition of several bacterial species, including S. Typhi, through the use of CEO at different concentrations (1,250–5,000 μg mL−1), was described. Moreover, Delaquis et al. (2002) showed antimicrobial activity of CEO able to inhibit L. monocytogenes and S. aureus, although without efficacy against P. fragi and S. Typhimurium. Although the Gram negative-bacteria seem to be more tolerant to essential oils treatment than Gram-positive ones, due to the lipopolysaccharides that protect them from hydrophobic compounds (Hyldgaard et al., 2012), the CEO primary mechanism of action is membrane damage, similarly in both Gram-positive and Gram-negative bacteria (Silva et al., 2011).
The higher sensitivity of Salmonella cocktails to the CEO treatment with respect to the single cultures might be due to hormesis phenomenon, nevertheless also the antagonism or competition established between the different cultures should be considered. Probably, to recover the stress caused by the oil, the microorganisms compete for nutrients or energy, hindering each other. Thanissery and Smith (2014) also observed this Salmonella behavior when 3 different serovars (Heidelberg, Montevideo, and Enteritidis) were treated with rosemary and clove essential oils in single and mixed culture.
The prolonged Lag period of Salmonella cocktails and reduced growth rate in presence of CEO are compatible with cell response to stressing events and represent a measure of the stress suffered by the cells. The cell in this phase in fact recovers a physiological state that allows the start over of multiplication. The speed of recovery of this condition corresponds to the extension of the Lag phase, depending on the time necessary for the different metabolic processes required (Serio et al., 2010). In our case the stressful event was the exposure to sub-lethal concentrations of CEO.
In addition to successful results obtained from in vitro studies, coriander essential oil underlined good potential as novel sustainable washing agent against S. enterica on carrot samples. This study, in fact, demonstrated that the application of CEO (5 μL mL−1) for a short time (2 min) effectively decreased Salmonella populations in artificially inoculated stick carrots up to 1 day of storage time. However, at the end of the storage period the populations slightly increased on the treated sample, with counts comparable with those of the control. These results, in addition to highlighting the capability of Salmonella spp. to survive and growth at low temperature, also evidence the ability of Salmonella to adapt to the stress conditions caused by the CEO. In this context, Kalily et al. (2017) demonstrated the adaptation of S. Senftenberg to linalool, which also conferred better protection to other antimicrobial treatments. In contrast with our findings, Ndoti-Nembe et al. (2015) in a study conducted on refrigerated mini-carrots, observed no significant reduction of the S. Typhimurium load at day 1 of treatment with savory EO (0.35% v/v) while a reduction of about 2 Log CFU g−1 was shown at day 9 of this treatment.
Although CEO was not effective during the entire examined storage time, the obtained results suggest that this extract could be used in combination with other natural substances with synergetic effects or in carrier systems (e.g., microemulsions, liposome).
Moreover, the stick carrots treatment with CEO washing solution did not cause any adverse effects on the organoleptic properties of the stick carrots. This factor should not be underestimated since the application of plant derivates, especially essential oils, usually interfere with the product aroma and taste (Lv et al., 2011). Although applied at low concentration, in fact, the intense aroma of essential oils has negative impact on food organoleptic properties, often exceeding the consumers' thresholds of acceptability (Hyldgaard et al., 2012).
Furthermore, carrot color was positively affected by this treatment. This last parameter is an important quality attribute. In particular, cut products are subjected to increased exposed surface area, that can lead to an increase in tissues respiration and subsequent protein, lipids and carbohydrates degradation (Xylia et al., 2018). The color preserving activity should be ascribed to phenolic compounds and to their related antioxidant capacity. These antioxidants, in fact, are well-known compounds utilized to prevent, retard or delay oxidative reaction and increase color stability (Mandal and Mandal, 2015).
To our knowledge, this is the first report on anti-Salmonella activity of C. sativum seeds essential oil on carrots. Thus, our findings contribute to enrich the knowledge on the antimicrobial activity of this essential oil against Salmonella spp. and on its potentiality as washing treatment in the control of S. enterica on fresh cut carrots. The in vitro study underlined a positive antimicrobial activity against the selected strains of different serovars, both tested singularly and in cocktails. The in situ study allowed to confirm the effectiveness of the essential oil in real matrix, where good sensory evaluation results were obtained upon the utilization of this washing treatment. The chemical investigations allowed to obtain interesting results in terms of total phenolic content and related antioxidant activity that probably contributed to the color stability observed in carrot products.
Future studies should be addressed to optimize the upstream activity, enhancing the essential oil permeability in cells by ameliorating vehicle systems (e.g., microemulsions, liposome) or by coupling this treatment with another one (e.g., physical treatments) to extend its efficacy during time.
The data that support the findings of this study are available from the corresponding authors (AR and AS) upon reasonable request.
MP: C. sativum seeds derivates extraction, chemical characterization, and draft preparation. CR: microbiological analyses and draft preparation. SP: support in chemical characterization. FM: support in microbiological analyses. CC-L and CL: data analyses. AP: experimental idea. DD: strains provision. AR and AS: data discussion and manuscript revision.
CR was supported by the European Social Fund (ESF) and the Italian Ministry of Education, University and Research (MIUR), through the National Operational Program for Research and Innovation, PON R&I 2014–2020, Action 1.2 ‘AIM: Attraction and International Mobility’ (AIM1894039-2).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Aelenei, P., Rimbu, C., Guguianu, E., Dimitriu, G., Aprotosoaie, A., Brebu, M., et al. (2019). Coriander essential oil and linalool – interactions with antibiotics against gram-positive and gram-negative bacteria. Lett. Appl. Microbiol. 68, 156–164. doi: 10.1111/lam.13100
Baranyi, J., and Roberts, T. A. (1994). A dynamic approach to predicting bacterial growth in food. Int. J. Food Microbiol. 23, 277–294. doi: 10.1016/0168-1605(94)90157-0
Bouazama, S., Harhar, H., Costa, J., Desjobert, J. M., Talbaoui, A., and Tabyaoui, M. (2017). Chemical composition and antibacterial activity of the essential oils of Lavandula pedunculata and Lavandula dentata. J. Mat. Environ. Sci. 8, 2154–2160.
Brand-Williams, W., Cuvelier, M. E., and Berset, C. (1995). Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 28, 25–30. doi: 10.1016/S0023-6438(95)80008-5
Burdock, G. A., and Carabin, I. G. (2009). Safety assessment of coriander (Coriandrum sativum L.) essential oil as a food ingredient. Food Chem. Toxicol. 47, 22–34. doi: 10.1016/j.fct.2008.11.006
Cai, Y., Tao, J., Jiao, Y., Fei, X., Zhou, L., Wang, Y., et al. (2016). Phenotypic characteristics and genotypic correlation between Salmonella isolates from a slaughterhouse and retail markets in Yangzhou, China. Int. J. Food Microbiol. 222, 56–64. doi: 10.1016/j.ijfoodmicro.2016.01.020
Calín-Sánchez, Á., Figiel, A., Lech, K., Szumny, A., and Carbonell-Barrachina, Á. A. (2013). Effects of drying methods on the composition of thyme (Thymus vulgaris L.) essential oil. Dry. Technol. 31, 224–235. doi: 10.1080/07373937.2012.725686
Calín-Sánchez, Á., Figiel, A., Lech, K., Szumny, A., Martínez-Tomé, J., and Carbonell-Barrachina, Á. A. (2015). Dying methods affect the aroma of Origanum majorana L. analyzed by GC–MS and descriptive sensory analysis. Ind. Crops Prod. 74, 218–227. doi: 10.1016/j.indcrop.2015.04.067
Clinical and Laboratory Standards Institute (2011). Performance Standards for Antimicrobial Susceptibility Testing; Twenty-First Informational Supplement. Wayne, PA: CLSI.
Cui, H. Y., Yuan, L., Li, W., and Lin, L. (2017). Edible films incorporated with chitosan and Artemisia annua oil nanoliposomes for inactivation of E. coli O157:H7 on cherry tomatoes. Int. J. Food Sci. Technol. 52, 687–698. doi: 10.1111/ijfs.13322
da Silva, J. P., de Souza, E. F., Della Modesta, R. C., Gomes, I. A., Freitas-Silva, O., and de Melo Franco, B. D. (2016). Antibacterial activity of nisin, oregano essential oil, EDTA, and their combination against Salmonella Enteritidis for application in mayonnaise. Vigil. Sanit. Debate 4, 83–91. doi: 10.3395/2317-269x.00308
D'Amato, S., Serio, A., Chaves-López, C., and Paparella, A. (2018). Hydrosols: biological activity and potential as antimicrobials for food applications. Food Control 86, 126–137. doi: 10.1016/j.foodcont.2017.10.030
Delaquis, P. J., Stanich, K., Girard, B., and Mazza, G. (2002). Antimicrobial activity of individual and mixed fraction of dill, celandra, coriander and eucalyptus essential oil. Int. J. food Microbiol. 74, 101–109. doi: 10.1016/S0168-1605(01)00734-6
Dolati, M., Rezaei, K., Vanak, Z. P., and Movahed, S. (2016). Study of the effects of essential oils of cumin, savory and cardamom as natural antioxidants on the flavor and oxidative stability of soybean oil during the storage. J. Essent. Oil-Bearing Plants 19, 176–184. doi: 10.1080/0972060X.2014.935030
Doulgeraki, A. I., Papaioannou, M., and Nychas, G.-J. E. (2016). Targeted gene expression study of Salmonella enterica during biofilm formation on rocket leaves. Food Sci. Technol. 65, 254–260. doi: 10.1016/j.lwt.2015.08.017
EFSA (2008). Camphor in flavourings and other food ingredients with flavouring properties. Opinion of the scientific panel on food additives, flavourings, processing aid and materials in contact with food on a request from the commission. EFSA J. 729, 1–15. doi: 10.2903/j.efsa.2008.729
El-Lateef Gharib, F. A., and Teixeira da Silva, J. A. (2013). Composition, total phenolic content and antioxidant activity of the essential oil of four Lamiaceae herbs. Med. Aromat. Plant Sci. Biotechnol. 7, 19–27.
Gil, M. I., Selma, M. V., López-Gálvez, F., and Allende, A. (2009). Fresh-cut product sanitation and wash water disinfection: problems and solutions. Int. J. Food Microbiol. 134, 37–45. doi: 10.1016/j.ijfoodmicro.2009.05.021
Gonzalez, R. J., Luo, Y., Ruiz-Cruz, S., and McEvoy, J. L. (2004). Efficacy of sanitizers to inactivate Escherichia coli O157:H7 on fresh-cut carrot shreds under simulated process water conditions. J. Food Prot. 67, 2375–80. doi: 10.4315/0362-028X-67.11.2375
Gündüz, G. T., Gönül, S. A., and Karapinar, M. (2009). Efficacy of myrtle oil against Salmonella typhimurium on fresh produce. Int. J. Food Microbiol. 130, 147–150. doi: 10.1016/j.ijfoodmicro.2009.01.010
Gutierrez, J., Bourke, P., Lonchamp, J., and Barry-Ryan, C. (2009). Impact of plant essential oils on microbiological, organoleptic and quality markers of minimally processed vegetables. Innov. Food Sci. Emerg. Technol. 10, 195–202. doi: 10.1016/j.ifset.2008.10.005
Gutiérrez-Alcántara, E. J., Gómez-Aldapa, C. A., Román-Gutiérrez, A. D., Rangel-Vargas, E., González-Olivares, L. G., and Castro-Rosas, J. (2016). Antimicrobial activity of roselle Hibiscus sabdariffa calyx extracts on culture media and carrots against multidrug-resistant Salmonella strains isolated from raw carrots. J. Food Saf. 36, 450−458. doi: 10.1111/jfs.12259
Hyldgaard, M., Mygind, T., Meyer, R. L., and Debabov, D. (2012). Essential oils in food preservation: mode of action, synergies, and interactions with food matrix components. Front. Microbiol. 3:12. doi: 10.3389/fmicb.2012.00012
Ildiz, N., Kiliç, A. B., and Konca, Y. (2018). Phytochemical composition of Coriandrum sativum L. (coriander) seeds and antibacterial effects on laying hens. Extraction 12:115.
Kalily, E., Hollander, A., Korin, B., Cymerman, I., and Yaron, S. (2017). Adaptation of Salmonella enterica serovar Senftenberg to linalool and its association with antibiotic resistance and environmental persistence. Appl. Environ. Microbiol. 83, e03398–16. doi: 10.1128/AEM.03398-16
Lima Oliveira, P. D., de Oliveira, K. A. R., dos Santos Vieira, W. A., Câmara, M. P. S., and de Souza, E. L. (2018). Control of anthracnose caused by Colletotrichum species in guava, mango and papaya using synergistic combinations of chitosan and Cymbopogon citratus (D.C. ex Nees) Stapf. essential oil. Int. J. Food Microbiol. 266, 87–94. doi: 10.1016/j.ijfoodmicro.2017.11.018
Liu, Q., Jin, X., Feng, X., Yang, H., and Fu, C. (2019). Inactivation kinetics of Escherichia coli O157: H7 and Salmonella typhimurium on organic carrot (Daucus carota L.) treated with low concentration electrolyzed water combined with short-time heat treatment. Food Control 106:106702. doi: 10.1016/j.foodcont.2019.06.028
Lo Cantore, P., Iacobellis, N. S., De Marco, A., Capasso, F., and Senatore, F. (2004). Antibacterial activity of Coriandrum sativum L. and Foeniculum vulgare Miller var. vulgare (Miller) essential oils. J. Agric. Food Chem. 52, 7862–7866. doi: 10.1021/jf0493122
Lo Fo Wong, D. M. A., Hald, T., van der Wolf, P. J., and Swanenburg, M. (2002). Epidemiology and control measures for Salmonella in pigs and pork. Livest. Prod. Sci. 76, 215–222. doi: 10.1016/S0301-6226(02)00121-5
Losio, M. N., Pavoni, E., Bilei, S., Bertasi, B., Bove, D., Capuano, F., et al. (2015). Microbiological survey of raw and ready-to-eat leafy green vegetables marketed in Italy. Int. J. Food Microbiol. 210, 88–91. doi: 10.1016/j.ijfoodmicro.2015.05.026
Lv, F., Liang, H., Yuan, Q., and Li, C. (2011). In vitro antimicrobial effects and mechanism of action of selected plant essential oil combinations against four food-related microorganisms. Food Res. Int. 44, 3057–3064. doi: 10.1016/j.foodres.2011.07.030
Mandal, S., and Mandal, M. (2015). Coriander (Coriandrum sativum L.) essential oil: chemistry and biological activity. Asian Pac. J. Trop. Biomed. 5, 421–428. doi: 10.1016/j.apjtb.2015.04.001
Martínez-Hernández, G. B., Amodio, M. L., and Colelli, G. (2017). Carvacrol-loaded chitosan nanoparticles maintain quality of fresh-cut carrots. Innov. Food Sci. Emerg. Technol. 41, 56–63. doi: 10.1016/j.ifset.2017.02.005
Masaldan, S., and Iyer, V. V. (2011). Antioxidant and antiproliferative activities of methanolic extract of Aloe vera leaves in human cancer cell lines. J. Pharm. Res. 4, 2791–2796.
Mazzarrino, G., Paparella, A., Chaves-López, C., Faberi, A., Sergi, M., Sigismondi, C., et al. (2015). Salmonella enterica and Listeria monocytogenes inactivation dynamics after treatment with selected essential oils. Food Control 50, 794–803. doi: 10.1016/j.foodcont.2014.10.029
Moon, T., Wilkinson, J. M., and Cavanagh, H. M. A. (2006). Antiparasitic activity of two Lavandula essential oils against Giardia duodenalis, Trichomonas vaginalis and Hexamita inflata. Parasitol. Res. 99:722. doi: 10.1007/s00436-006-0234-8
Ndoti-Nembe, A., Vu, K. D., Han, J., Doucet, N., and Lacroix, M. (2015). Antimicrobial effects of nisin, essential oil, and γ-Iirradiation treatments against high load of Salmonella Typhimurium on mini-carrots. J. Food Sci. 80, M1544–M1548. doi: 10.1111/1750-3841.12918
Nguyen-the, C., and Carlin, F. (1994). The microbiology of minimally processed fresh fruits and vegetables. Crit. Rev. Food Sci. Nutr. 34, 371–401. doi: 10.1080/10408399409527668
Oyaizu, M. (1986). Studies on products of browning reaction: antioxidative activity of products of browning reaction prepared from glucosamine. Japan. J. Nutr. 44, 307–315. doi: 10.5264/eiyogakuzashi.44.307
Paparella, A., Mazzarrino, G., Chaves-López, C., Rossi, C., Sacchetti, G., Guerrieri, O., et al. (2016). Chitosan boosts the antimicrobial activity of Origanum vulgare essential oil in modified atmosphere packaged pork. Food Microbiol. 59, 23–31. doi: 10.1016/j.fm.2016.05.007
Pellegrini, M., Ricci, A., Serio, A., Chaves-López, C., Mazzarrino, G., D'Amato, S., et al. (2018). Characterization of EOs obtained from Abruzzo autochthonous plants: antioxidant and antimicrobial activities assessment for food application. Foods 7:19. doi: 10.3390/foods7020019
Pui, C. F., Wong, W. C., Chai, L. C., Nillian, E., Ghazali, F. M., Cheah, Y. K., et al. (2011). Simultaneous detection of Salmonella spp., Salmonella Typhi and Salmonella Typhimurium in sliced fruits using multiplex PCR. Food Control 22, 337–342. doi: 10.1016/j.foodcont.2010.05.021
Ramos, B., Brandão, T. R., Teixeira, P., and Silva, C. L. (2020). Biopreservation approaches to reduce Listeria monocytogenes in fresh vegetables. Food Microbiol. 85:103282. doi: 10.1016/j.fm.2019.103282
Raybaudi-Massilia, R. M., Mosqueda-Melgar, J., and MartÍn-Belloso, O. (2006). Antimicrobial activity of essential oils on Salmonella Enteritidis, Escherichia coli, and Listeria innocua in fruit juices. J. Food Prot. 69, 1579–1586. doi: 10.4315/0362-028X-69.7.1579
Rossi, C., Chaves-López, C., Smole MoŽina, S., Di Mattia, C., Scuota, S., Luzzi, I., et al. (2019). Salmonella enterica adhesion: effect of Cinnamomum zeylanicum essential oil on lettuce. LWT Food Sci. Technol. 111, 16–22. doi: 10.1016/j.lwt.2019.05.026
Ruiz-Cruz, S., Acedo-Félix, E., Díaz-Cinco, M., Islas-Osuna, M. A., and González-Aguilar, G. A. (2007). Efficacy of sanitizers in reducing Escherichia coli O157:H7, Salmonella spp. and Listeria monocytogenes populations on fresh-cut carrots. Food Control 18, 1383–1390. doi: 10.1016/j.foodcont.2006.09.008
Ruiz-Navajas, Y., Viuda-Martos, M., Sendra, E., Perez-Alvarez, J. A., and Fernández-López, J. (2013). In vitro antioxidant and antifungal properties of essential oils obtained from aromatic herbs endemic to the southeast of Spain. J. Food Prot. 76, 1218–1225. doi: 10.4315/0362-028X.JFP-12-554
Serio, A., Chiarini, M., Tettamanti, E., and Paparella, A. (2010). Electronic paramagnetic resonance investigation of the activity of Origanum vulgare L. essential oil on the Listeria monocytogenes membrane. Lett. Appl. Microbiol. 51, 149–157. doi: 10.1111/j.1472-765X.2010.02877.x
Silva, F., Ferreira, S., Queiroz, J. A., and Domingues, F. C. (2011). Coriander (Coriandrum sativum L.) essential oil: its antibacterial activity and mode of action evaluated by flow cytometry. J Med. Microbiol. 60, 1479–1486. doi: 10.1099/jmm.0.034157-0
Silveira, J. B., Hessel, C. T., and Tondo, E. C. (2017). Inactivation of Salmonella Enteritidis on lettuces used by minimally processed vegetable industries. J. Infect. Dev. Countr. 11, 34–41. doi: 10.3855/jidc.8224
Thanissery, R., and Smith, D. P. (2014). Marinade with thyme and orange oils reduces Salmonella Enteritidis and Campylobacter coli on inoculated broiler breast fillets and whole wings. Poult. Sci. 93, 1258–1262. doi: 10.3382/ps.2013-03697
Torres-Vitela, M. D. R., Gómez Aldapa, C. A., Cerna-Cortes, J. F., Villarruel-López, A., Rangel-Vargas, E., and Castro-Rosas, J. (2013). Presence of indicator bacteria, diarrheagenic E. coli pathotypes and Salmonella in fresh carrot juice from Mexican restaurants. Lett. Appl. Microbiol. 56, 180–185. doi: 10.1111/lam.12030
Wang, H. F., Hway, K. H., Yang, C. H., and Huang, K. F. (2017). Anti-oxidant activity and major chemical component analyses of twenty-six commercially available essential oils. J. Food Drug Anal. 25, 881–889. doi: 10.1016/j.jfda.2017.05.007
Xylia, P., Chrysargyris, A., Botsaris, G., and Tzortzakis, N. (2018). Mint and pomegranate extracts/oils as antibacterial agents against Escherichia coli O157:H7 and Listeria monocytogenes on shredded carrots. J. Food Saf. 38:e12423. doi: 10.1111/jfs.12423
Keywords: Coriandrum sativum, essential oil, anti-Salmonella activity, Salmonella enterica, Minimal processed vegetables, carrots, in vitro study, in situ study
Citation: Pellegrini M, Rossi C, Palmieri S, Maggio F, Chaves-López C, Lo Sterzo C, Paparella A, De Medici D, Ricci A and Serio A (2020) Salmonella enterica Control in Stick Carrots Through Incorporation of Coriander Seeds Essential Oil in Sustainable Washing Treatments. Front. Sustain. Food Syst. 4:14. doi: 10.3389/fsufs.2020.00014
Received: 24 October 2019; Accepted: 31 January 2020;
Published: 20 February 2020.
Edited by:
Cristobal N. Aguilar, Universidad Autónoma de Coahuila, MexicoReviewed by:
Guadalupe Virginia Nevárez-Moorillón, Autonomous University of Chihuahua, MexicoCopyright © 2020 Pellegrini, Rossi, Palmieri, Maggio, Chaves-López, Lo Sterzo, Paparella, De Medici, Ricci and Serio. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Antonella Ricci, YXJpY2NpQHVuaXRlLml0; Annalisa Serio, YXNlcmlvQHVuaXRlLml0
†Present Address: Marika Pellegrini, Department of Life, Health and Environmental Sciences, University of L'Aquila, L'Aquila, Italy
‡These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.