 Ingrid Rivera-Iñiguez1,2†‡
Ingrid Rivera-Iñiguez1,2†‡ Claudia Hunot-Alexander2†
Claudia Hunot-Alexander2† Maricruz Sepúlveda-Villegas1
Maricruz Sepúlveda-Villegas1 Liliana Campos-Medina1,3
Liliana Campos-Medina1,3 Sonia Roman1*
Sonia Roman1*- 1Department of Genomic Medicine in Hepatology, Civil Hospital of Guadalajara, “Fray Antonio Alcalde”, Health Sciences Center, University of Guadalajara, Guadalajara, Jalisco, Mexico
- 2Department of Human Reproduction Clinics, Infant Growth and Development, Institute of Human Nutrition, Health Sciences Center, Guadalajara, Jalisco, Mexico
- 3Doctoral Program in Molecular Biology in Medicine, Health Sciences Center, University of Guadalajara, Guadalajara, Jalisco, Mexico
Introduction: Appetitive traits are influenced by the interplay between genetic and environmental factors. This study aimed to explore the relationship between gene polymorphisms involved in the regulation of energy balance and food reward and appetitive traits in young Mexican subjects.
Methods: This cross-sectional study involved 118 university freshman undergraduates who completed the Adult Eating Behaviour Questionnaire for Spanish speakers (AEBQ-Esp) to assess their appetitive traits. A real-time PCR system was employed to determine gene polymorphisms involved in energy balance (LEP rs7799039, MC4R rs17782313, FTO rs9939609, GHRL rs696217), and reward system (DRD2/ANKK1 Taq1A rs1800497 and COMT rs4680).
Results: The mean age of participants was 20.14 ± 3.95 years, 71.2% were women and their mean BMI was 23.52 ± 4.05 kg/m2. COMT Met allele carriers presented a significantly higher “Emotional overeating” mean score than Val allele carriers (2.63 ± 0.70 vs. 2.23 ± 0.70, p = 0.028). The MC4R CC + CT genotype correlated positively with “Emotional overeating” (Phi = 0.308, p = 0.01). The COMT MetMet+MetVal genotype correlated with higher “Emotional overeating” (r = 0.257, p = 0.028; Phi = 0.249, p = 0.033). The protective genotype FTO TT correlated positively with “Emotional undereating” (Phi = 0.298, p = 0.012). Carriers of the risk genotype MC4R CC + CT presented a higher risk of “Emotional overeating” than TT carriers (OR = 2.4, 95% CI 1.3–4.8, p = 0.034). Carriers of the risk genotype COMT MetMet+MetVal (OR = 3.4, 95% CI 1.1–10.3, p = 0.033), were associated with a higher risk of “Emotional overeating” than ValVal carriers. The protective FTO genotype TT was associated with “Emotional undereating” (OR = 1.8, 95% CI 1.1–9.1, p = 0.014).
Discussion: The study found a relationship between the protective genotypes of FTO TT and “Emotional undereating” and risk genotypes of COMT Met/Met+Met/Val and MC4R CC + CT with “Emotional overeating.” These genetic factors may increase weight gain by enhancing hedonic food consumption and reducing satiety control. Future studies should focus on replication studies in ethnically diverse young adults and life stages to explore the relationship between polymorphisms and appetitive traits and weight. This will help tailor personalized nutrigenetic strategies to counteract disordered eating patterns leading to obesity and associated co-morbidities.
1 Introduction
Western societies and countries with emerging market economies such as Mexico are immersed in an environment conducive to the development of obesity (1). Given this rise, the “Behavioral Susceptibility Theory” (BST) proposes that some individuals are more sensitive to external food cues or “Food Responsiveness” and less responsive to internal cues for satiety or “Satiety Responsiveness,” therefore, increasing the risk of obesity (2, 3). The BST suggests that human body weight and appetitive traits are influenced by the interaction between genetic factors with the surrounding environment (4–6). “Food approach” and/or “food avoidance” appetitive traits, can be measured using psychometric questionnaires such as the Adult Eating Behavior Questionnaire (AEBQ) and its validated Spanish version (AEBQ-Esp) (7, 8). These traits are linked to higher body weight changes spanning from infancy to adulthood (5, 7).
Twin studies and Genome-Wide Association Studies (GWAS) have explained how genetic susceptibility to obesity is influenced by appetite; having a heightened sensitivity to internal hunger and fullness signals (“Satiety responsiveness”) and enjoying food (“Enjoyment of food”), as well as wanting to eat in response to the sight, smell, or taste of food (“Food responsiveness”), with heritability estimates of 63 and 75%, respectively (9). In the Gemini cohort, appetitive traits such as “Slowness in eating” (84%) and “Satiety responsiveness” (72%) were found to be highly genetically determined, while a moderate effect was associated with “Food responsiveness” (59%) and “Enjoyment of food” (53%) (10). Several extensive adult cohorts spanning the UK, the US, Canada, France, and Finland consistently reveal associations between genetic susceptibility and appetitive traits related to “Food responsiveness” such as “External eating,” “Uncontrolled eating” and “Disinhibition” (11–14). Notably, a longitudinal study of 2,464 British adults from the Whitehall II cohort demonstrated that “Disinhibition” mediated 34% of the association between a polygenic risk score (PRS) for obesity (comprising 97 genetic variants) in 20-year BMI trajectories during midlife (15).
Responsiveness to food cues, refers to the desire to eat when encountering food-related stimuli like the appearance, aroma, or taste of food (3). This aspect is linked to hedonic processes associated with pleasure and reward (16) regulated in the brain reward system. Gene polymorphisms related to the reward system such as the Dopamine Receptor 2 (DRD2/ANKK1) gene and Catechol-O-methyltransferase (COMT) seem to influence appetitive traits, by promoting hedonic food consumption and reducing satiety control (17). The A1 allele from the DRD2/ANKK1 TaqIA1 (rs1800497) polymorphism contributes to decreased DRD2 receptor density and increases L-DOPA activity, impairing the reward system. The A1 allele is highly prevalent in Mexicans and contributes to detrimental dietary quality and metabolic disturbances (18). The COMT gene regulates emotional, cognitive, and appetitive processes (19, 20). The rs4680 (Val158Met) polymorphism in the COMT gene has been related to appetite conditioning (19).
Internal satiety signals such as hunger and fullness are regulated by hormones in the energy balance system (21). Three genes encode important components related to energy intake and energy expenditure balance such as Leptin (LEP), ghrelin (GHRL), melanocortin-4-receptor (MC4R), and the fat mass and obesity-associated gene (FTO) (22, 23). Leptin, the hormone that regulates fullness is codified by the Leptin (LEP) gene and the G allele from the −2548G > A, rs7799039 polymorphism of this gene has been linked with lower levels of postprandial fullness (24). The rs696217 polymorphism (Leu72Met) of the GHRL gene affects hunger and satiety. Individuals with the Met72Met genotype exhibit greater consumption of sugars and bread compared to carriers of the Leu72Leu genotype (25). There is evidence that individuals showing genetic risk factors exhibit varying scores related to “food approach” and “food avoidant” appetitive traits, as observed among 180 young Mexican subjects, where circulating IgG autoAbs reacting to ghrelin and leptin were evaluated (26). Additionally, MC4R regulates energy homeostasis and appetite signals (27, 28). The C allele from the −188 kb T > C, rs17782313 MC4R polymorphism is associated with higher BMI (29) and higher appetite (30). In a study with 221 obese Chilean children, the rs17782313 variant was associated with “Satiety responsiveness” and “Enjoyment of food” scores (31). This variant may be linked to childhood obesity, impacting factors such as “Enjoyment of food” and “Satiety responsiveness” and possibly “Eating in the absence of hunger” according to a case–control study involving 139 normal-weight and 238 obese children aged 6–12 years (32). The T > A, rs9939609 FTO gene polymorphism expressed in the hypothalamus, regulating food intake and energy expenditure is known to be related to higher BMI. Studies in children examining the relationship between appetitive traits and FTO reveal that those with the AA genotype are more prone to overweight or obesity. This inclination is attributed to lower “Satiety responsiveness,” heightened “Food responsiveness,” and increased “Enjoyment of food” in children and adolescents (33, 34).
Studies indicate that young adults, especially university students, undergoing a transitional life stage face challenges such as balancing work and study responsibilities, as well as managing stress (35). These stressors can adversely affect their eating behaviors, putting them at a heightened risk of developing disordered eating patterns (36, 37) and the development of obesity (38). However, little is known of the relationship that exists between different polymorphisms that participate in the regulation of food consumption and appetitive traits in young people, which could serve to tailor intervention in this age group and move toward a more personalized medicine and nutrition approach. Therefore, this study aimed to explore the relationship between polymorphisms of genes involved in energy balance (LEP rs7799039, MC4R rs17782313, FTO rs9939609, GHRL rs696217), and reward system (DRD2/ANKK1 rs1800497 and COMT rs4680) and appetitive traits subscales of the AEBQ-Esp in young Mexican subjects.
2 Materials and methods
2.1 Participants and procedure
This study was carried out at the Department of Genomic Medicine in Hepatology from January 2018–2019. Due to the 2019 COVID pandemic, research activities were postponed and resumed from July to December 2022. The study involved primarily freshman undergraduates selected from the Health Sciences Center (Centro Universitario de Ciencias de la Salud, CUCS) at the Universidad de Guadalajara, a public non-fee-paying university. Students who did not have Spanish as their first language were not included in the research. Researchers initiated contact with university administrators, who, in turn, approached lecturers responsible for undergraduate courses, seeking their willingness to involve their classes in the study. The researchers secured informed consent from the students before commencing data collection. Ethical approval (number CI-08218) was obtained by the Institutional Review Board.
2.2 Measurement of appetitive traits
Appetitive traits were assessed using the AEBQ-Esp, a 30-item self-report questionnaire known for its validity and reliability in measuring seven distinct appetitive traits (8). The three “food approach” traits consisted of “Food responsiveness” (four items, e.g., I am always thinking about food), “Emotional overeating” (five items, e.g., I eat more when I’m angry), and “Enjoyment of food” (three items, e.g., I love food). Four “Food avoidance” traits included: “Satiety responsiveness” (four items, e.g., I get full up easily), “Emotional undereating” (five items, e.g., I eat less when I’m upset), “Food fussiness” (five items, e.g., I enjoy tasting new foods) and “Slowness in eating” (four items, e.g., I eat slowly). All participants responded to a 5-point Likert scale, ranging from 1 (“strongly disagree”) to 5 (“strongly agree”). Mean scores were calculated for each subscale and classified as dichotomous variables, as “high-score” for “food approach” traits if their score on each subscale was greater than three, likewise, they were classed as “low-score” for “food avoidance” traits if their score for each subscale was less than three.
2.3 DNA extraction and genotyping
A peripheral blood sample was obtained to perform the genomic DNA extraction from leukocytes with a modified Salting Out technique (39); a quantified concentration of 20 ng/μl with Nanodrop 2000c was stored at −70°C. A real-time PCR system was performed with predesigned TaqMan Single Nucleotide Polymorphism (SNP) Genotyping Assays (Applied Biosystems, Life Technologies, Camarillo, CA, USA) for the determination of LEP rs7799039 (C___1328079_10), MC4R rs17782313 (C__32667060_10), FTO rs9939609 (C__30090620_10), GHRL rs696217 (C___3151003_20), DRD2/ANKK1 Taq1A rs1800497 (C___7486676_10) and COMT rs4680 polymorphisms (C__25746809_50) using a 96-well plate and then read on a Step One Plus thermocycler (Applied Biosystems). Thermal cycling conditions were the following: activation stage at 95°C for 10 min, denaturation stage of 40 cycles at 95°C for 15 s. And the annealing/extension stages at 60°C for 1 min.
2.4 Sociodemographic data
Participants reported their age, university, work status (full-time job; part-time job; self-employed; not working), marital status (not married; married; lives with partner), living status (at home; away from home), both parents’ work (full-time job; part-time job; self-employed; has different jobs; unemployed; does not have a father or mother) and both parents’ education (primary school; GCSE levels; A levels; bachelors; postgraduate). The latter information was used as a proxy for socioeconomic status (data not shown) (40).
2.5 Anthropometry
Height was measured without shoes to the nearest 0.1 cm (Seca stadiometer); weight was measured in kilograms to the nearest 100 g (Tanita weighing scale); and body mass index (BMI) was the ratio of weight (kg), by height (m2). BMI cut-off points were as follows: healthy weight (18.5–24.9 kg/m2) overweight (25–29.9 kg/m2), and obesity (>30 kg/m2).
2.6 Statistical analyses
The Kolmogorov–Smirnov test was applied to know the distribution of the analyzed data. Continuous variables were presented as mean ± standard deviations (SD), and categorical variables as percentages where appropriate. Cronbach’s alpha was performed to test internal reliability among the AEBQ-Esp subscales, considering an acceptable value of greater than 0.7. Comparisons were conducted using the T-test for normally distributed data and the Mann–Whitney U test for non-normally distributed data. Correlation analyses were conducted using point biserial correlations for normally distributed data, Kendall’s correlation for non-normally distributed data, and the Phi-Coefficient was also calculated. The association of genotypes with the subscales was estimated with an odds ratio (OR) test and a 95% confidence interval (CI). A p-value of <0.05 was considered significant. Statistical analysis was conducted with SPSS software (version 25.0; SPSS Inc., Chicago, IL, USA), significance level of p < 0.05. The Hardy–Weinberg equilibrium (HWE) was obtained with the Arlequin software for Windows (version 3.1; Berne, Switzerland).
3 Results
3.1 Population’s descriptive characteristics
Demographic, clinical, and genetic characteristics are presented in Table 1. In this study, 118 young adults (20.14 ± 3.95 years) were evaluated (71.2% women). The mean BMI was 23.52 ± 4.05 kg/m2.
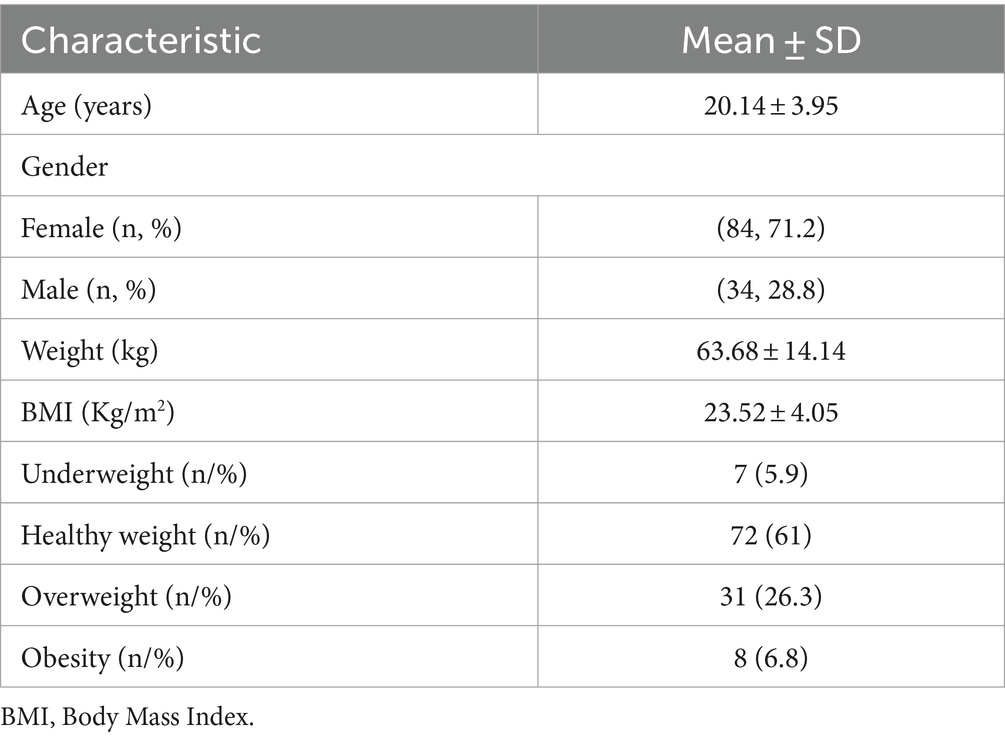
Table 1. Population demographics and clinical characteristics (n = 118).
Most subjects were healthy weight (61%), followed by 33.1% of excess weight, and 5.9% were underweight.
3.2 Descriptive data for appetitive traits and AEBQ-Esp internal validation
The means and standard deviations of the seven appetitive traits of the AEBQ-Esp subscales and their internal reliability are shown in Table 2. Cronbach’s alpha value >0.7 demonstrated good internal reliability in five of the seven traits, only the “Food fussiness” and “Slowness in eating” subscales were < 0.5.
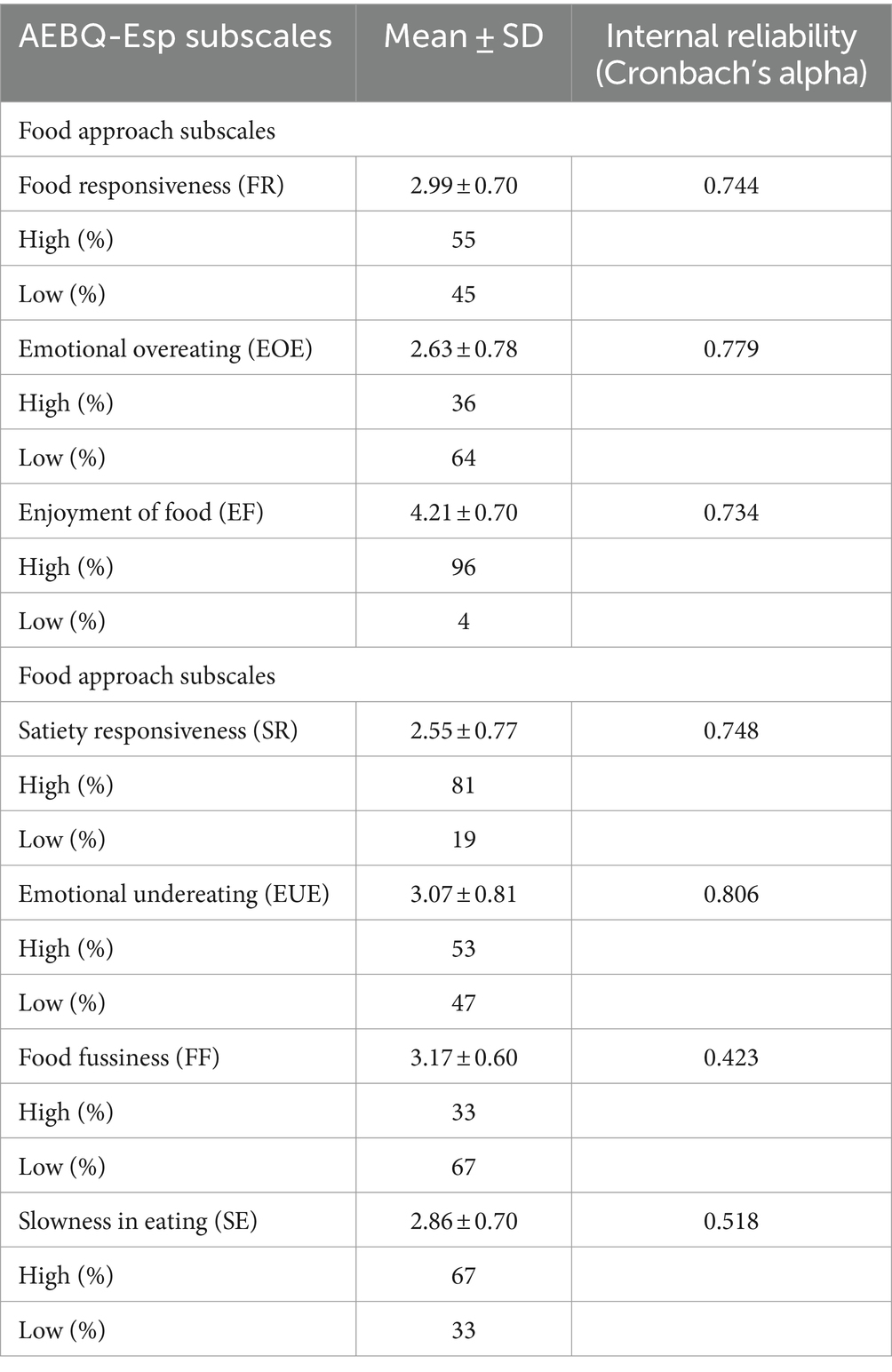
Table 2. AEBQ-Esp subscales and internal reliability.
3.3 Allele and genotype frequencies of the analyzed gene polymorphism
The allele and genotype distribution of the analyzed gene polymorphisms are shown in Table 3. All analyzed polymorphisms were in Hardy–Weinberg equilibrium (HWE) (p > 0.05). A different number of samples were analyzed for each polymorphism. For the MC4R, the T allele was more prevalent in the population (88%), and the absence of the genotype CC was observed. The frequency of the G allele was higher in the genes LEP, COMT, and DRD2; also in these groups, the heterozygote (AG/GA) was the most frequent genotype. In the FTO gene, the frequency of the T allele was 70%, and 48% of homozygotes. In the case of the GHRL gene, the G allele had a frequency of 85%, while the TT genotype was present in only one subject. All analyzed polymorphisms were in Hardy–Weinberg equilibrium (HWE) (p > 0.05).
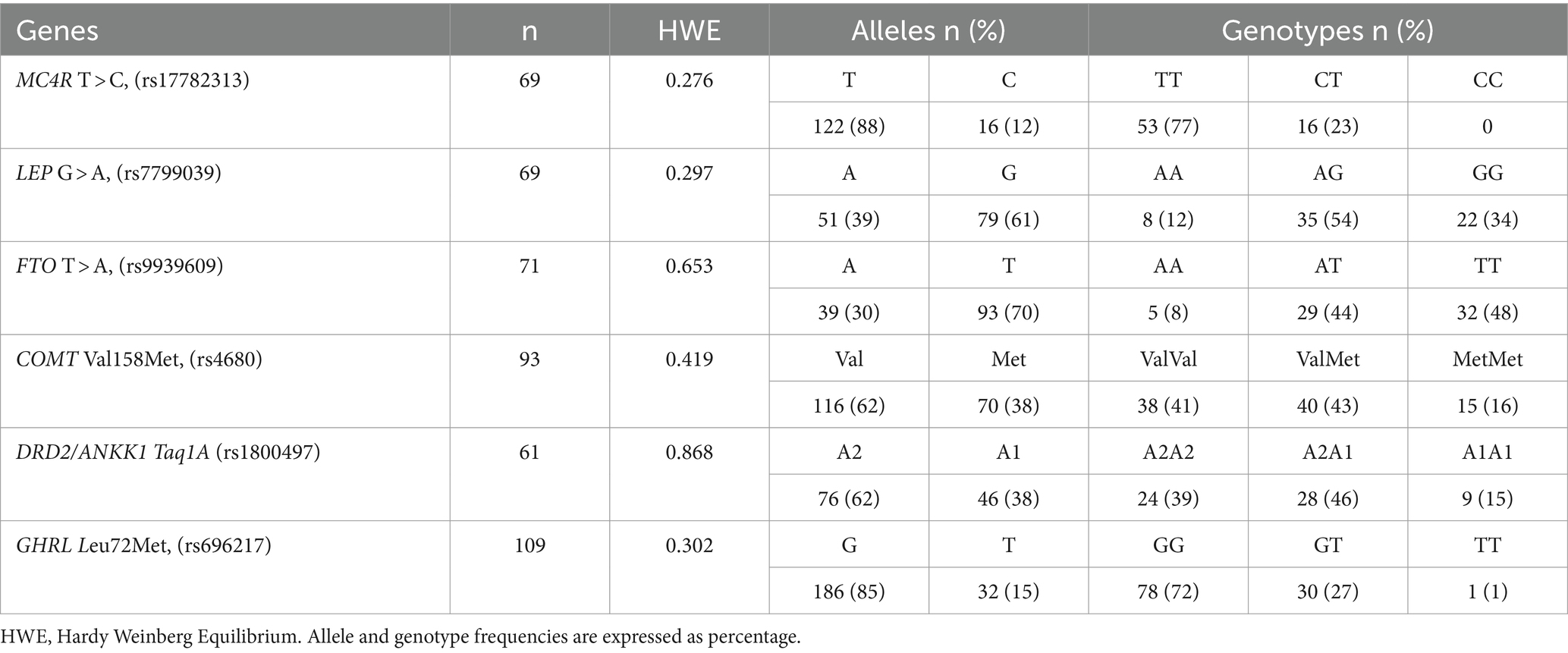
Table 3. Alleles and genotypes frequency of the gene polymorphisms.
3.4 Appetitive traits according to gene polymorphisms
The means and SD of the AEBQ-Esp subscales according to the dominant model of the studied gene polymorphisms are presented in Table 4. There was a tendency toward carriers of the MC4R C allele to have lower “Emotional overeating” (2.42 ± 0.65, p = 0.08). Similarly, LEP G allele carriers (2.87 ± 0.65, p = 0.06) tended to present lower “Slowness in eating.” FTO carriers tended to have higher “Emotional undereating” (3.11 ± 0.79, p = 0.061). Met allele carriers from the COMT gene tended to present higher “Food fussiness” (3.40 ± 0.37, p = 0.06) and presented significantly higher “Emotional overeating” (2.63 ± 0.70, p = 0.028) than Val allele carriers. No other significant differences were found.
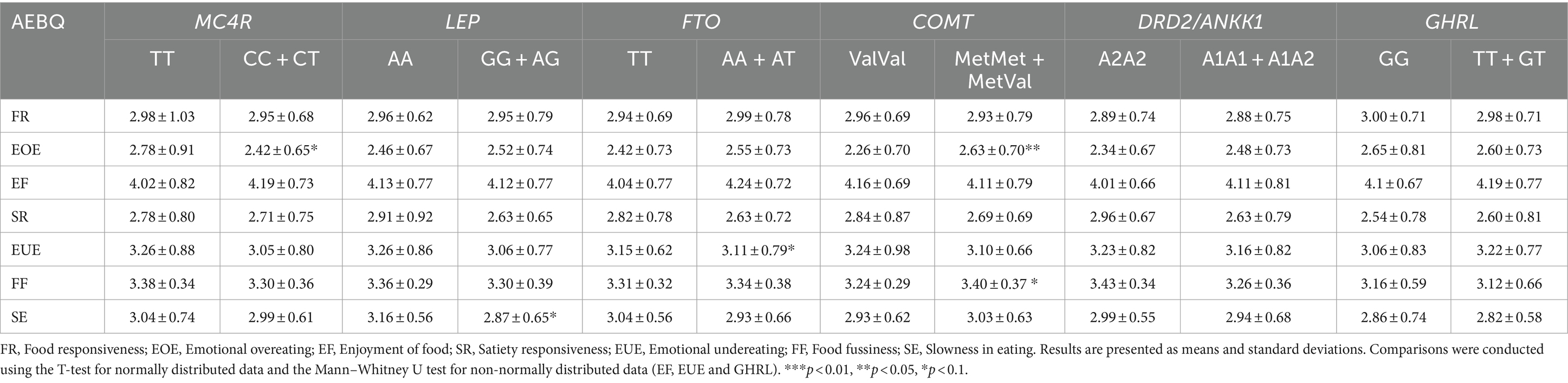
Table 4. Appetitive traits according to gene polymorphisms.
3.5 Correlations between gene polymorphisms and appetitive traits
The correlation analysis between genetic models and mean appetitive trait values is listed in Table 5, and correlations between gene polymorphisms and appetitive traits as dichotomous variables can be found in Table 6. In terms of “food approach” subscales, there was a tendency toward a positive correlation between the MC4R CC + CT genotype and “Emotional overeating” mean scores (r = 0.211, p = 0.082, Table 5), as well as with the categorical variable (Phi = 0.308, p = 0.010, Table 6). Similarly, the COMT MetMet+MetVal genotype correlated with higher “Emotional overeating” mean scores (r = 0.257, p = 0.028; Phi = 0.249, p = 0.033). Carriers of the higher risk genotype MetMet+MetVal were positively correlated with all “food approach” traits (Phi = 0.378, p = 0.002, Table 6). There was a tendency toward a correlation of the high-risk genotype LEP GG + AG with higher “Enjoyment of food” mean scores (r = 0.209, p = 0.082, Table 6).
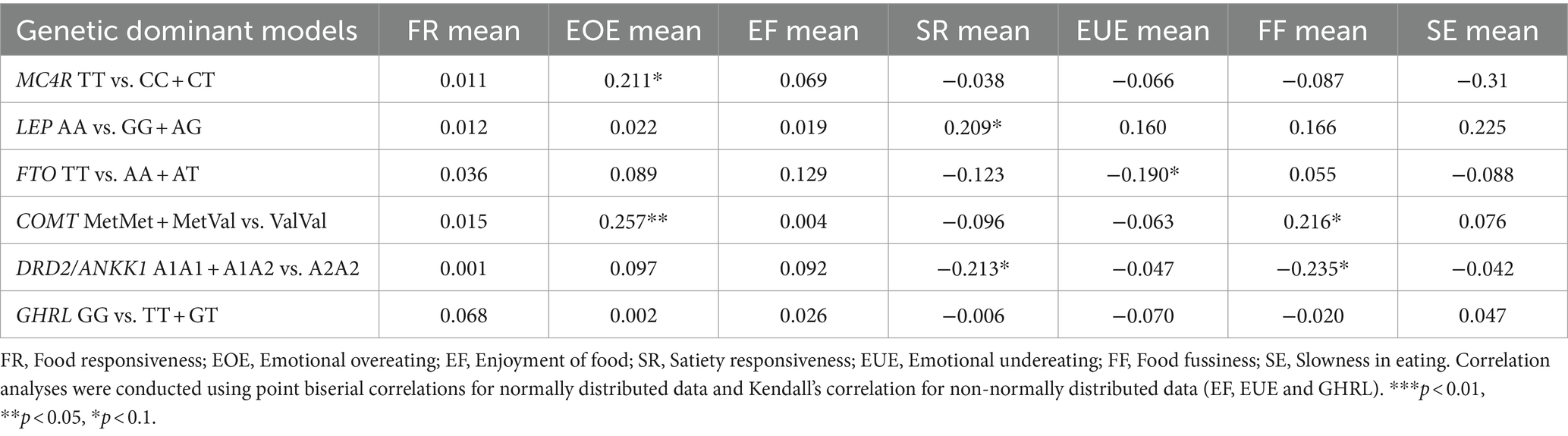
Table 5. Correlations between gene polymorphisms and appetitive traits mean values.
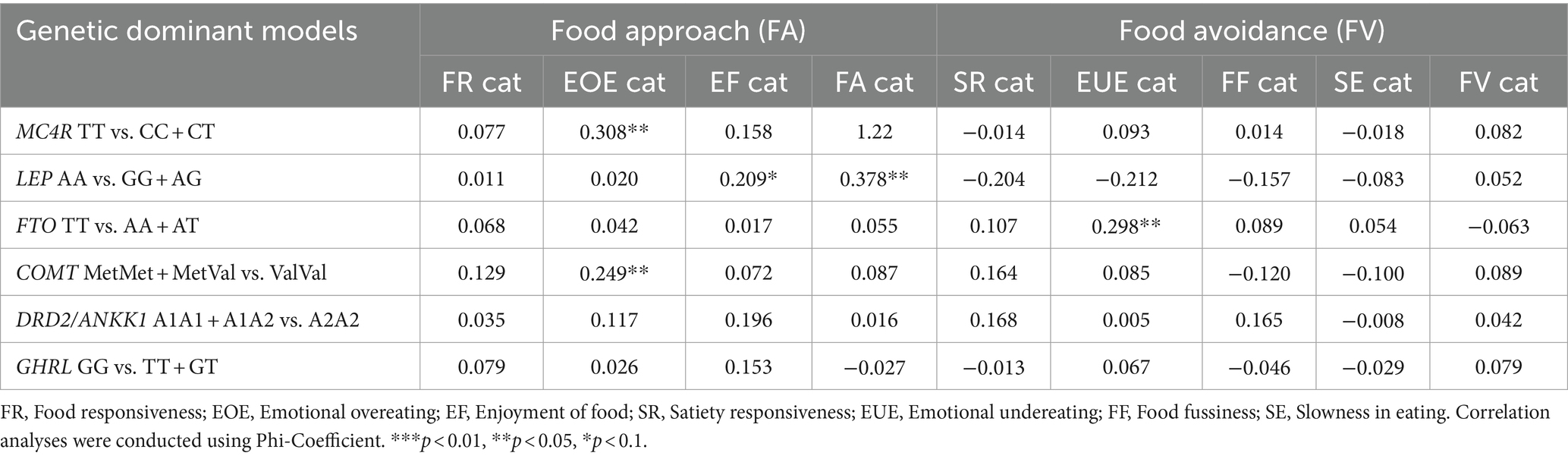
Table 6. Correlations between gene polymorphisms and appetitive traits as dichotomous variable.
As part of the relationships between gene polymorphism and “food avoidant” traits, we found that the protective genotype LEP AA was positively correlated with “Satiety responsiveness” (r = 0.209, p = 0.085). In contrast, there was a tendency toward a negative correlation between the risk genotype DRD2 A1A1 + A1A2 with lower “Satiety responsiveness” (r = −0.213, p = 0.08). The high-risk genotype FTO AA+AT was negatively correlated to “Emotional undereating”; however, it was not significant (r = −0.190, p = 0.061). There was a significant positive correlation between the protective FTO genotype with “Emotional undereating” as a categorical variable (Phi = 0.298, p = 0.012). In this study there were only correlation tendencies with “Food fussiness” and gene polymorphisms; COMT high-risk genotype MetMet+MetVal and “Food fussiness” (r = 0.216, p = 0.066), and the protective genotype DRD2 A2A2 (r = −0.235, p = 0.60).
3.6 Association between gene polymorphisms and appetitive traits
Carriers of the risk genotype MC4R CC + CT presented a higher risk of “Emotional overeating” than TT carriers (OR = 2.4, 95% CI 1.3–4.8, p = 0.034). Similarly, carriers of the risk genotype COMT MetMet+MetVal, were associated with a higher risk of “Emotional overeating” than Val carriers (OR = 3.4, 95% CI 1.1–10.3, p = 0.033). The protective genotype TT from FTO was associated with higher “Emotional undereating” (OR = 1.8, 95% CI 1.1–9.1, p = 0.014) (Data not shown).
4 Discussion
Mexico presents one of the highest rates of obesity worldwide, and there is limited data in the Mexican population related to appetitive traits and genetic risk factors that contribute to obesity. This appears to be the first study that explores the relationship between gene polymorphisms related to energy balance, hunger, satiety, and reward with scores of “food approach” and “food avoidant” appetitive trait subscales in young healthy weight (BMI: 23.52 ± 4.05) Mexican adults. To determine the appetitive traits in this population, the AEBQ-Esp was used, which has previously been validated in the Mexican population (8). In this study, the results reveal that the AEBQ-Esp with the exclusion of the “Food fussiness” and “Slowness in eating” subscales, were internally reliable (0.734–0.906); therefore, we recommend care be taken with regards to results interpreted using these two subscales. These results differ from the original validation of the AEBQ-Esp, where alpha Cronbach values were 0.70 and 0.91, respectively, for these subscales (8). Lower internal reliability values have also been found when mothers of children (41) and toddlers (42) complete the AEBQ-Esp questionnaire, in samples of Mexican subjects. This could be due to the size of the population obtained in this sample.
In the present study, we found important relationships between the analyzed gene polymorphisms with scores of “food approach” (“Emotional overeating” and “Enjoyment of Food”) and “food avoidant” (“Satiety responsiveness,” “Food fussiness” and “Emotional undereating”) appetitive traits. In terms of “Emotional overeating,” MC4R has been related to obesity by disrupting energy homeostasis and appetite signals (27, 28). Specifically, the rs17782313 polymorphism of MC4R has been previously associated with higher BMI and higher appetite (30). Moreover, when C carriers are exposed to unhealthy eating patterns, the risk of developing depressive feelings increases (43). Our analysis revealed a tendency toward a correlation between the MC4R (rs17782313) CC + CT genotype and “Emotional overeating” mean value (r = 0.211, p = 0.082) and as a categorical variable (Phi = 0.308, p = 0.010). Moreover, carriers of the risk genotype MC4R CC + CT presented a higher risk of “Emotional overeating” than TT carriers (OR = 2.4, 95% CI 1.3–4.8, p = 0.034). This is similar to what was found in an Iranian cross-sectional study in adults overweight and obese, where obese individuals carriers of the C risk allele of MC4R (rs17782313) exhibited a higher mean score of “Emotional eating” assessed with the self-report Emotional Eating Questionnaire (EEQ). In that study, MC4R rs17782313 was also associated with appetite, energy intake, nutrients, stress, ghrelin, and cortisol levels (44). In contrast, in a study with adults with obesity from Chile, “Uncontrolled eating” and “Emotional eating” measured using the Three-Factor Eating Questionnaire-R18, showed that carriers of the risk allele rs17782313 presented higher scores of “Emotional undereating” compared to non-carriers (45).
The COMT gene plays a key role in the reward system, where it regulates emotional, cognitive, and appetite processes (19, 20). Moreover, it is important to highlight that in the Mexican population, a higher prevalence of the Met allele (63%) has been reported (46), which contrasts with the prevalence in other populations. The Met allele has been reported to be associated with a personality trait such as “worrier” and with impulsivity and negative emotions (47, 48). In line with this, in the present study, we found that the high-risk genotype COMT (rs4680) MetMet+MetVal was positively correlated with the mean scores (r = 0.257, p = 0.028), and categorical variables of “Emotional overeating” (Phi = 0.249, p = 0.033). In addition, we only found that carriers of the higher risk genotype MetMet+MetVal were positively correlated with “food approach” traits (Phi = 0.378, p = 0.002). In contrast, in a cross-sectional study of 3-4-year-old children conducted in Brazil, results showed that the COMT Val158Met polymorphism contributed to a higher intake of palatable food. That study revealed that carriers of the Val allele presented a higher intake of fats when compared to MetMet homozygotes (49).
In terms of “food avoidant” traits, we found that the protective genotype LEP AA was positively correlated with “Satiety responsiveness” (r = 0.209, p = 0.085) and a tendency toward a negative correlation between the risk genotype DRD2 A1A1 + A1A2 and “Satiety responsiveness” (r = −0.213, p = 0.08). Valladares found that carriers of the class II allele from the non-translatable hypervariable region at the 3′ end of the gene LEP (TTTC)n, had higher scores of “Slowness in eating” vs. non-carriers (31). It has been reported previously, that rs7799039 is associated with higher consumption of energy intake (50). These studies so far, highlight that there is a need for more studies to determine whether polymorphisms of LEP could be influencing emotional overeating.
Moreover, in this study, we found a tendency toward a negative correlation between the high-risk genotype FTO AA+AT and “Emotional undereating” (r = −0.190, p = 0.061). There was a significant positive correlation between the protective genotype from the FTO with “Emotional undereating” as a categorical variable (Phi = 0.298, p = 0.012). The protective FTO genotype TT was associated with higher “Emotional undereating” (OR = 1.8, 95% CI 1.1–9.1, p = 0.014). In contrast, Valladares and colleagues, in their study conducted among Chilean children with obesity, did not find any relationships between “Emotional undereating” and the genetic polymorphisms studied in that population (31).
On the other hand, “Food fussiness” is a “food avoidance” trait, that refers to the reluctance to eat both new and familiar foods, pickiness of the flavor and texture of foods, and it has been mostly explored among children (51, 52). Interestingly, although not significant, in this study, we found a tendency toward the presence of the COMT high-risk genotype MetMet+MetVal and higher “Food fussiness” (r = 0.216, p = 0.066), and the protective genotype DRD2 A2A2 with lower “Food fussiness” (r = −0.235, p = 0.60). However, care should be taken not to put too much weight on these results, as the “Food fussiness” “subscale was not reliable.” The genes that contribute to “Food fussiness” have not been identified yet. However, it has been previously reported that “Food fussiness” is highly heritable (53). Potentially a larger sample size study that explores the role of the COMT and DRD2 genes and “Food fussiness” would be necessary. According to data from the Gemini twin study, genetic correlations have been observed between “Food fussiness” and preferences for vegetables and fruits (54). It would be worth analyzing if the COMT and DRD2 genes influence the intake of fruits and vegetables in a larger population.
Finally, in the context of the global obesity epidemic including in Mexico, these results show the importance of moving toward a more personalized medicine and nutrition approach, toward assessing risk, and finding novel ways to prevent and treat diseases (55). Furthermore, the presence of the respective risk alleles among the studied genes underscores the need for preventive strategies against further weight gain among the population (56). In the present study, 61% of participants presented healthy weight and the prevalence of overweight and obesity was 33.1%. Therefore, a future perspective could be including a bigger sample size to perform analysis according to participant’s BMI. Also, appetitive traits can be targeted to create behavioral strategies for weight reduction when personalized profiles are generated for individuals (57). Also, appetitive traits reflect the relationship that individuals have with food and response to food cues in the environment, which initiate in childhood and continue until adolescence as a risk factor for the development of eating disorders (58) is a further complication in this age group.
Our results need to be considered in view of several limitations. We recognize that in a cross-sectional study, it is complex to infer causal relationships because it only involves a one-time measurement, and we did not consider previous individual environmental factors that might influence appetitive traits including the 2019 COVID pandemic. Similarly, the pandemic affected our ability to conduct research activities, but we resumed activities and finalized the data analysis in 2022. Young adults may tend to under-report certain traits perceived as less desirable, such as higher “Food responsiveness,” while potentially over-reporting traits considered more favorable, like higher “Satiety responsiveness.” Conversely, it’s worth noting that social desirability tends to increase with age and be more prevalent in females (59). In addition, in this study, we had higher participation of female than male students. These young adults were recruited from the Health Sciences Center at the Universidad de Guadalajara, in which more than 60% of the students are females, specifically students enrolled in Nutrition are within a healthy BMI. We recognize that the characteristics and size of this sample may limit generalizing the results and detecting further associations. Including more male participants may allow us to identify associations related to gender. However, it has been suggested that certain appetitive traits influence young females to be more susceptible to develop overweight or obesity (60). We also did not evaluate physical activity levels which seems to have a modifying effect on certain SNPs such as the rs993609 FTO polymorphism (61, 62) and it is possible that individuals who engage in higher physical activity levels could have lesser levels of stress or negative emotions that influence emotional overeating. Despite the limitations of this study, it is important to recognize that young university adults experience a drastic transition in their lifestyles, which together with their genetic background may influence certain appetitive traits that could increase the risk of overweight and obesity. Multiple environmental factors and genes are involved in eating behavior, so there is a need to continue to expand the number of genes related to energy balance, appetite, satiety, and food reward and conduct longitudinal studies in different populations.
5 Conclusion
In this study the risk genotypes of COMT Met+Met and MC4R TT correlated with higher “Emotional overeating” and the protective FTO genotype TT was also associated with higher “Emotional undereating.” These results may facilitate understanding the genetic influence of weight gain by promoting hedonic food consumption and reductions in satiety regulation and could signal the need for more tailored strategies, including personalized medicine, personalized nutrition, or behavior-based treatment strategies that can help improve eating behaviors in young subjects. Similarly, identifying individuals at a higher risk of presenting appetite traits that facilitate cravings and food overconsumption could help prevent the development of obesity at earlier stages, even starting as early as childhood. These results may also help decision-makers to create public health recommendations and policy changes related to environmental factors that could be triggers of these high-risk appetitive traits. These findings require replication in different culturally diverse young adults, as well as in different stages of the life course. Future studies should also examine the prospective relationship between polymorphisms and appetitive traits and explore how these associations may relate to changes in weight and how engagement in healthy lifestyle behaviors could help modulate these associations.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Ethics statement
The studies involving humans were approved by Comités de Investigación, Ética en Investigación y Bioseguridad del Centro Universitario de Ciencias de la Salud, Universidad de Guadalajara. The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
Author contributions
IR-I: Conceptualization, Project administration, Resources, Visualization, Writing – review & editing, Formal analysis, Funding acquisition, Investigation, Writing – original draft. CH-A: Conceptualization, Formal analysis, Funding acquisition, Investigation, Project administration, Visualization, Writing – original draft, Writing – review & editing, Resources. MS-V: Formal analysis, Investigation, Visualization, Writing – review & editing. LC-M: Formal analysis, Investigation, Visualization, Writing – review & editing. SR: Conceptualization, Formal analysis, Funding acquisition, Investigation, Project administration, Resources, Supervision, Visualization, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. CH-A was funded by the Programa para el Desarrollo Profesional Docente [PRODEP (UDG-EXB-684)] – Secretaría de Educación Pública (SEP) – Mexico.
Acknowledgments
We would like to thank administrators and Faculty at the Centro Universitario de Ciencias de la Salud (CUCS), Universidad de Guadalajara, for allowing us to collect the data presented in this study. We would also like to thank students from the Instituto de Nutrición Humana, CUCS, Universidad de Guadalajara, and from the Departamento de Medicina Genómica en Hepatologia, Hospital Civil de Guadalajara, “Fray Antonio Alcalde,” who helped with the data collection.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The handling editor OR-L declared a past co-authorship with the author SR.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. García-Chávez, CG, Monterrubio-Flores, E, Ramírez-Silva, I, Aburto, TC, Pedraza, LS, and Rivera-Dommarco, J. Food contribution to total daily energy intake in the Mexican population older than five years. Salud Publica Mex. (2020) 62:166–80. doi: 10.21149/10636
2. Carnell, S, and Wardle, J. Appetite and adiposity in children: evidence for a behavioral susceptibility theory of obesity. Am J Clin Nutr. (2008) 88:22–9. doi: 10.1093/ajcn/88.1.22
3. Llewellyn, CH, and Fildes, A. Behavioural Susceptibility Theory: Professor Jane Wardle and the Role of Appetite in Genetic Risk of Obesity. Curr Obes Rep. (2017) 6:38–45. doi: 10.1007/s13679-017-0247-x
4. Llewellyn, C, and Wardle, J. Behavioral susceptibility to obesity: gene-environment interplay in the development of weight. Physiol Behav. (2015) 152:494–501. doi: 10.1016/j.physbeh.2015.07.006
5. Kininmonth, A, Smith, A, Carnell, S, Steinsbekk, S, Fildes, A, and Llewellyn, C. The association between childhood adiposity and appetite assessed using the child eating behavior questionnaire and baby eating behavior questionnaire: a systematic review and meta-analysis. Obes Rev. (2021) 22:e13169. doi: 10.1111/obr.13169
6. Llewellyn, CH, Kininmonth, AR, Herle, M, Nas, Z, Smith, AD, Carnell, S, et al. Behavioural susceptibility theory: the role of appetite in genetic susceptibility to obesity in early life. Philos Trans R Soc Lond Ser B Biol Sci. (2023) 378:20220223. doi: 10.1098/rstb.2022.0223
7. Hunot, C, Fildes, A, Croker, H, Llewellyn, CH, Wardle, J, and Beeken, RJ. Appetitive traits and relationships with BMI in adults: development of the adult eating behaviour questionnaire. Appetite. (2016) 105:356–63. doi: 10.1016/j.appet.2016.05.024
8. Hunot-Alexander, C, Arellano-Gómez, LP, Smith, AD, Kaufer-Horwitz, M, Vásquez-Garibay, EM, Romero-Velarde, E, et al. Examining the validity and consistency of the Adult Eating Behaviour Questionnaire-Español (AEBQ-Esp) and its relationship to BMI in a Mexican population. Eat Weight Disord. (2022) 27:651–63. doi: 10.1007/s40519-021-01201-9
9. Carnell, S, Haworth, CMA, Plomin, R, and Wardle, J. Genetic influence on appetite in children. Int J Obes. (2008) 32:1468–73. doi: 10.1038/ijo.2008.127
10. Llewellyn, CH, van Jaarsveld, CHM, Johnson, L, Carnell, S, and Wardle, J. Nature and nurture in infant appetite: analysis of the Gemini twin birth cohort. Am J Clin Nutr. (2010) 91:1172–9. doi: 10.3945/ajcn.2009.28868
11. Jacob, R, Drapeau, V, Tremblay, A, Provencher, V, Bouchard, C, and Pérusse, L. The role of eating behavior traits in mediating genetic susceptibility to obesity. Am J Clin Nutr. (2018) 108:445–52. doi: 10.1093/ajcn/nqy130
12. Konttinen, H, Llewellyn, C, Wardle, J, Silventoinen, K, Joensuu, A, Männistö, S, et al. Appetitive traits as behavioural pathways in genetic susceptibility to obesity: a population-based cross-sectional study. Sci Rep. (2015) 5:14726. doi: 10.1038/srep14726
13. Cornelis, MC, Rimm, EB, Curhan, GC, Kraft, P, Hunter, DJ, Hu, FB, et al. Obesity susceptibility loci and uncontrolled eating, emotional eating and cognitive restraint behaviors in men and women. Obesity. (2014) 22:E135–41. doi: 10.1002/oby.20592
14. de Lauzon-Guillain, B, Clifton, EA, Day, FR, Clément, K, Brage, S, Forouhi, NG, et al. Mediation and modification of genetic susceptibility to obesity by eating behaviors. Am J Clin Nutr. (2017) 106:996–1004. doi: 10.3945/ajcn.117.157396
15. Brunner, EJ, Maruyama, K, Shipley, M, Cable, N, Iso, H, Hiyoshi, A, et al. Appetite disinhibition rather than hunger explains genetic effects on adult BMI trajectory. Int J Obes. (2021) 45:758–65. doi: 10.1038/s41366-020-00735-9
16. Blundell, JE, and Finlayson, G. Is susceptibility to weight gain characterized by homeostatic or hedonic risk factors for overconsumption? Physiol Behav. (2004) 82:21–5. doi: 10.1016/j.physbeh.2004.04.021
17. Salgado-Pineda, P, Delaveau, P, Blin, O, and Nieoullon, A. Dopaminergic contribution to the regulation of emotional perception. Clin Neuropharmacol. (2005) 28:228–37. doi: 10.1097/01.wnf.0000185824.57690.f0
18. Rivera-Iñiguez, I, Panduro, A, Ramos-Lopez, O, Villaseñor-Bayardo, SJ, and Roman, S. DRD2/ANKK1 TaqI A1 polymorphism associates with overconsumption of unhealthy foods and biochemical abnormalities in a Mexican population. Eat Weight Disord. (2019) 24:835–44. doi: 10.1007/s40519-018-0596-9
19. Klucken, T, Kruse, O, Wehrum-Osinsky, S, Hennig, J, Schweckendiek, J, and Stark, R. Impact of COMT Val158Met-polymorphism on appetitive conditioning and amygdala/prefrontal effective connectivity. Hum Brain Mapp. (2015) 36:1093–101. doi: 10.1002/hbm.22688
20. Vai, B, Riberto, M, Poletti, S, Bollettini, I, Lorenzi, C, Colombo, C, et al. Catechol-O-methyltransferase Val(108/158)Met polymorphism affects fronto-limbic connectivity during emotional processing in bipolar disorder. Eur Psychiatry. (2017) 41:53–9. doi: 10.1016/j.eurpsy.2016.10.002
21. Harrold, JA, Dovey, TM, Blundell, JE, and Halford, JCG. CNS regulation of appetite. Neuropharmacology. (2012) 63:3–17. doi: 10.1016/j.neuropharm.2012.01.007
22. Krashes, MJ, Lowell, BB, and Garfield, AS. Melanocortin-4 receptor-regulated energy homeostasis. Nat Neurosci. (2016) 19:206–19. doi: 10.1038/nn.4202
23. Speakman, JR . FTO effect on energy demand versus food intake. Nature. (2010) 464:E1; discussion E2. doi: 10.1038/nature08807
24. Dougkas, A, Yaqoob, P, Givens, DI, Reynolds, CK, and Minihane, AM. The impact of obesity-related SNP on appetite and energy intake. Br J Nutr. (2013) 110:1151–6. doi: 10.1017/S0007114513000147
25. Sanchez-Murguia, T, Torres-Castillo, N, Magaña-de la Vega, L, Rodríguez-Reyes, SC, Campos-Pérez, W, and Martínez-López, E. Role of Leu72Met of GHRL and Gln223Arg of LEPR variants on food intake, subjective appetite, and hunger-satiety hormones. Nutrients. (2022) 14:2100. doi: 10.3390/nu14102100
26. Espinoza-García, AS, Hunot-Alexander, C, Martínez-Moreno, AG, Vázquez-Solorzano, R, Porchas-Quijada, M, and Reyes-Castillo, Z. IgG antibodies reacting with ghrelin and leptin are correlated with body composition and appetitive traits in young subjects. Appetite. (2022) 168:105685. doi: 10.1016/j.appet.2021.105685
27. Baldini, G, and Phelan, KD. The melanocortin pathway and control of appetite-progress and therapeutic implications. J Endocrinol. (2019) 241:R1–R33. doi: 10.1530/JOE-18-0596
28. Yeo, GSH, Chao, DHM, Siegert, AM, Koerperich, ZM, Ericson, MD, Simonds, SE, et al. The melanocortin pathway and energy homeostasis: from discovery to obesity therapy. Mol Metab. (2021) 48:101206. doi: 10.1016/j.molmet.2021.101206
29. Lombard, Z, Crowther, NJ, van der Merwe, L, Pitamber, P, Norris, SA, and Ramsay, M. Appetite regulation genes are associated with body mass index in black South African adolescents: a genetic association study. BMJ Open. (2012) 2:e000873. doi: 10.1136/bmjopen-2012-000873
30. Hammad, MM, Abu-Farha, M, Hebbar, P, Cherian, P, al Khairi, I, Melhem, M, et al. MC4R variant rs17782313 associates with increased levels of DNAJC27, ghrelin, and visfatin and correlates with obesity and hypertension in a kuwaiti cohort. Front Endocrinol. (2020) 11:437. doi: 10.3389/fendo.2020.00437
31. Valladares, M, Obregón, AM, Weisstaub, G, Burrows, R, Patiño, A, Ho-Urriola, J, et al. Association between feeding behavior, and genetic polymorphism of leptin and its receptor in obese Chilean children. Nutr Hosp. (2014) 31:1044–51. doi: 10.3305/nh.2015.31.3.8049
32. Ho-Urriola, J, Guzmán-Guzmán, IP, Smalley, SV, González, A, Weisstaub, G, Domínguez-Vásquez, P, et al. Melanocortin-4 receptor polymorphism rs17782313: association with obesity and eating in the absence of hunger in Chilean children. Nutrition. (2014) 30:145–9. doi: 10.1016/j.nut.2013.05.030
33. Emond, JA, Tovar, A, Li, Z, Lansigan, RK, and Gilbert-Diamond, D. FTO genotype and weight status among preadolescents: assessing the mediating effects of obesogenic appetitive traits. Appetite. (2017) 117:321–9. doi: 10.1016/j.appet.2017.07.009
34. Wardle, J, Llewellyn, C, Sanderson, S, and Plomin, R. The FTO gene and measured food intake in children. Int J Obes. (2009) 33:42–5. doi: 10.1038/ijo.2008.174
35. Hill, D, Conner, M, Clancy, F, Moss, R, Wilding, S, Bristow, M, et al. Stress and eating behaviours in healthy adults: a systematic review and meta-analysis. Health Psychol Rev. (2022) 16:280–304. doi: 10.1080/17437199.2021.1923406
36. Quick, VM, and Byrd-Bredbenner, C. Disturbed eating behaviours and associated psychographic characteristics of college students. J Hum Nutr Diet. (2013) 26:53–63. doi: 10.1111/jhn.12060
37. Stice, E, Desjardins, CD, Rohde, P, and Shaw, H. Sequencing of symptom emergence in anorexia nervosa, bulimia nervosa, binge eating disorder, and purging disorder and relations of prodromal symptoms to future onset of these disorders. J Abnorm Psychol. (2021) 130:377–87. doi: 10.1037/abn0000666
38. Carnell, S, Benson, L, Pryor, K, and Driggin, E. Appetitive traits from infancy to adolescence: using behavioral and neural measures to investigate obesity risk. Physiol Behav. (2013) 121:79–88. doi: 10.1016/j.physbeh.2013.02.015
39. Miller, SA, Dykes, DD, and Polesky, HF. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res. (1988) 16:1215. doi: 10.1093/nar/16.3.1215
41. Hunot-Alexander, C, Curiel-Curiel, CP, Romero-Velarde, E, Vásquez-Garibay, EM, Mariscal-Rizo, A, Casillas-Toral, E, et al. Intergenerational transmission of appetite: associations between mother-child dyads in a Mexican population. PLoS One. (2022) 17:e0264493. doi: 10.1371/journal.pone.0264493
42. Hunot-Alexander, C, González-Toribio, J, Vásquez-Garibay, EM, Larrosa-Haro, A, Casillas-Toral, E, and Curiel-Curiel, CP. Validity and reliability of the baby and child eating behavior questionnaire, toddler version (BEBQ-Mex and CEBQ-T-Mex) in a low sociodemographic sample recruited in a Mexican hospital. Behav Sci. (2021) 11:168. doi: 10.3390/bs11120168
43. Hajmir, MM, Mirzababaei, A, Clark, CCT, Ghaffarian-Ensaf, R, and Mirzaei, K. The interaction between MC4R gene variant (rs17782313) and dominant dietary patterns on depression in obese and overweight women: a cross sectional study. BMC Endocr Disord. (2023) 23:83. doi: 10.1186/s12902-023-01335-0
44. Rahati, S, Qorbani, M, Naghavi, A, and Pishva, H. Association and interaction of the MC4R rs17782313 polymorphism with plasma ghrelin, GLP-1, cortisol, food intake and eating behaviors in overweight/obese Iranian adults. BMC Endocr Disord. (2022) 22:234. doi: 10.1186/s12902-022-01129-w
45. Vega, JA, Salazar, G, Hodgson, MI, Cataldo, LR, Valladares, M, Obregón, AM, et al. Melanocortin-4 receptor gene variation is associated with eating behavior in Chilean adults. Ann Nutr Metab. (2016) 68:35–41. doi: 10.1159/000439092
46. González-Castro, TB, Tovilla-Zárate, C, Juárez-Rojop, I, Pool García, S, Genis, A, Nicolini, H, et al. Distribution of the Val108/158Met polymorphism of the COMT gene in healthy Mexican population. Gene. (2013) 526:454–8. doi: 10.1016/j.gene.2013.05.068
47. Bieliński, M, Jaracz, M, Lesiewska, N, Tomaszewska, M, Sikora, M, Junik, R, et al. Association between COMT Val158Met and DAT1 polymorphisms and depressive symptoms in the obese population. Neuropsychiatr Dis Treat. (2017) 13:2221–9. doi: 10.2147/NDT.S138565
48. Stein, DJ, Newman, TK, Savitz, J, and Ramesar, R. Warriors versus worriers: the role of COMT gene variants. CNS Spectr. (2006) 11:745–8. doi: 10.1017/S1092852900014863
49. Galvão, ACS, Krüger, RC, Campagnolo, PDB, Mattevi, VS, Vitolo, MR, and Almeida, S. Association of MAOA and COMT gene polymorphisms with palatable food intake in children. J Nutr Biochem. (2012) 23:272–7. doi: 10.1016/j.jnutbio.2010.12.004
50. Crovesy, L, and Rosado, EL. Interaction between genes involved in energy intake regulation and diet in obesity. Nutrition. (2019) 67-68:110547. doi: 10.1016/j.nut.2019.06.027
51. Russell, CG, and Russell, A. Biological and psychosocial processes in the development of children’s appetitive traits: insights from developmental theory and research. Nutrients. (2018) 10:692. doi: 10.3390/nu10060692
52. Kininmonth, AR, Herle, M, Tommerup, K, Haycraft, E, Farrow, C, Croker, H, et al. Parental feeding practices as a response to child appetitive traits in toddlerhood and early childhood: a discordant twin analysis of the Gemini cohort. Int J Behav Nutr Phys Act. (2023) 20:39. doi: 10.1186/s12966-023-01440-2
53. Smith, AD, Herle, M, Fildes, A, Cooke, L, Steinsbekk, S, and Llewellyn, CH. Food fussiness and food neophobia share a common etiology in early childhood. J Child Psychol Psychiatry. (2017) 58:189–96. doi: 10.1111/jcpp.12647
54. Fildes, A, van Jaarsveld, CHM, Cooke, L, Wardle, J, and Llewellyn, CH. Common genetic architecture underlying young children’s food fussiness and liking for vegetables and fruit. Am J Clin Nutr. (2016) 103:1099–104. doi: 10.3945/ajcn.115.122945
55. Roman, S, Ramos-Lopez, O, and Panduro, A. Genomic medicine in hepatology: towards personalized medicine in obesity and chronic liver disease. Ann Hepatol. (2023) 28:100875. doi: 10.1016/j.aohep.2022.100875
56. Panduro, A, Roman, S, Milán, RG, Torres-Reyes, LA, and Gonzalez-Aldaco, K. Personalized nutrition to treat and prevent obesity and diabetes In: Z Cheng , editor. Nutritional signaling pathway activities in obesity and diabetes. Burlington House, Piccadilly, London, W1J 0BA: The Royal Society of Chemistry. (2020).
57. Hunot-Alexander, C, Croker, H, Fildes, A, Johnson, F, and Beeken, RJ. Brief ‘Appetitive Trait Tailored Intervention’: development in a sample of adults with overweight and obesity. Behav Chang. (2022) 39:106–22. doi: 10.1017/bec.2021.22
58. Herle, M, Stavola, BD, Hübel, C, Abdulkadir, M, Ferreira, DS, Loos, RJF, et al. A longitudinal study of eating behaviours in childhood and later eating disorder behaviours and diagnoses. Br J Psychiatry. (2020) 216:113–9. doi: 10.1192/bjp.2019.174
59. Soubelet, A, and Salthouse, TA. Influence of social desirability on age differences in self-reports of mood and personality. J Pers. (2011) 79:741–62. doi: 10.1111/j.1467-6494.2011.00700.x
60. Campos-Nonato, I, Galván-Valencia, Ó, Hernández-Barrera, L, Oviedo-Solís, C, and Barquera, S. Prevalencia de obesidad y factores de riesgo asociados en adultos mexicanos: resultados de la Ensanut 2022. Salud Publica Mex. (2023) 65:s238–47. doi: 10.21149/14809
61. Andersen, MK, Ängquist, L, Bork-Jensen, J, Jonsson, AE, Stinson, SE, Sandholt, CH, et al. Physical activity and insulin sensitivity independently attenuate the effect of FTO rs9939609 on obesity. Diabetes Care. (2023) 46:985–92. doi: 10.2337/dc22-2078
Keywords: polymorphisms, AEBQ, appetitive traits, young adults, personalized nutrition, tailoring, eating patterns
Citation: Rivera-Iñiguez I, Hunot-Alexander C, Sepúlveda-Villegas M, Campos-Medina L and Roman S (2024) Relationship between energy balance and reward system gene polymorphisms and appetitive traits in young Mexican subjects. Front. Nutr. 11:1373578. doi: 10.3389/fnut.2024.1373578
Edited by:
Omar Ramos-Lopez, Universidad Autónoma de Baja California, Tijuana, MexicoReviewed by:
Daniela Abigail Navarro, Ariel University, IsraelNanette Steinle, United States Department of Veterans Affairs, United States
Copyright © 2024 Rivera-Iñiguez, Hunot-Alexander, Sepúlveda-Villegas, Campos-Medina and Roman. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sonia Roman, c29uaWFtYXJpYXJvbWFuQGhvdG1haWwuY29t
†These authors have contributed equally to this work and share first authorship
‡PRESENT ADDRESS
Ingrid Rivera-Iñiguez, Department of Pediatrics, University of California San Diego School of Medicine, San Diego, CA, United States