 Yinxia Huang†
Yinxia Huang† Chuanzhi Zhu
Chuanzhi Zhu Liping Pan
Liping Pan Zongde Zhang
Zongde Zhang
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Cell. Infect. Microbiol. , 25 July 2023
Sec. Molecular Bacterial Pathogenesis
Volume 13 - 2023 | https://doi.org/10.3389/fcimb.2023.1218583
This article is part of the Research Topic Factors associated with drug resistance and virulence of Mycobacterium tuberculosis View all 8 articles
Tuberculosis (TB) is a widespread infectious disease caused by Mycobacterium tuberculosis (M. tb), which has been a significant burden for a long time. Post-translational modifications (PTMs) are essential for protein function in both eukaryotic and prokaryotic cells. This review focuses on the contribution of protein acetylation to the function of M. tb and its infected macrophages. The acetylation of M. tb proteins plays a critical role in virulence, drug resistance, regulation of metabolism, and host anti-TB immune response. Similarly, the PTMs of host proteins induced by M. tb are crucial for the development, treatment, and prevention of diseases. Host protein acetylation induced by M. tb is significant in regulating host immunity against TB, which substantially affects the disease’s development. The review summarizes the functions and mechanisms of M. tb acetyltransferase in virulence and drug resistance. It also discusses the role and mechanism of M. tb in regulating host protein acetylation and immune response regulation. Furthermore, the current scenario of isoniazid usage in M. tb therapy treatment is examined. Overall, this review provides valuable information that can serve as a preliminary basis for studying pathogenic research, developing new drugs, exploring in-depth drug resistance mechanisms, and providing precise treatment for TB.
In 2021, an estimated 10.6 million people (95% UI: 9.9–11 million) worldwide contracted tuberculosis (TB), marking an increase of 4.5% from 10.1 million (95% UI: 9.5–10.7 million) in 2020. During the same period, the TB incidence rate (new cases per 100,000 population per year) increased by 3.6% (WHO, 2022). TB is one of the most fatal infectious diseases, and its connection with HIV/AIDS is especially tragic (Riou and Althaus, 2020). HIV suppresses the immune system, making individuals more susceptible to Mycobacterium tuberculosis (M. tb) infections, hastening the progression to active TB, and increasing latent TB reactivation by 20-fold (Pawlowski et al., 2012; Wang Y., et al, 2022). It is concerning that HIV-infected individuals are more likely to develop drugresistant TB in Oceania and Eastern Europe. Additionally, HIVXDR-TB has become increasingly common among elderly people (Zhou et al., 2023).
Protein post-translational modifications (PTMs) are reversible mechanisms of cellular adaptation to changing environmental conditions. PTMs such as phosphorylation, acetylation, ubiquitination, and pupylation play a crucial role in mycobacterial virulence, pathogenesis, and metabolism. Approximately one third of the annotated M. tb proteome is modified post-translationally, and many of these proteins are essential for mycobacterial survival. Understanding the signaling pathways and PTMs may assist clinical strategies and drug development for M. tb (Budzik et al., 2020; Arora et al., 2021). Among PTMs, protein acetylation plays a crucial role in mycobacterial virulence, pathogenesis, and metabolism. In eukaryotes, protein acetylation is involved in almost all biological processes, including transcriptional regulation, protein translation, central metabolism, protein stability, signal transduction, and pathogen virulence (Carabetta and Cristea, 2017; Nakayasu et al., 2017; Christensen et al., 2019; Shvedunova and Akhtar, 2022). Recently, protein acylation has received increased attention due to its involvement in several mitochondrial, nuclear, and cytosolic processes (Glozak et al., 2005; Finkel et al., 2009; Norris et al., 2009; Kim and Yang, 2011). Protein acetylation is a dynamic equilibrium process in which the acetyl group of acetyl-coA is transferred to the N-a-amino group of protein or N-lysine protein group under the action of acetyltransferase or deacetylated transferase. Initially, it was believed to be an epigenetic modification of chromatin-related proteins, such as histones (Bernal et al., 2014). However, it is now suggested that acetylation modification plays important roles in biological processes. With the progress of mass spectrometry technology, the role of protein acetylation modification in the occurrence and development of diseases has become an important direction and focus of current research. Acetylation modification is a conserved post-translational modification discovered on histones in 1964 and is closely related to biological processes such as gene transcription regulation and protein function (Allfrey et al., 1964). In recent years, more and more studies have found that protein acetylation plays an indispensable role in the occurrence, development, and outcome of TB. Understanding the role and mechanism of new protein acetylation modification in the regulation of host anti-TB immunity is a current research focus on the epigenetic mechanism of TB. This may provide new targets for TB prevention, diagnosis, and host-directed therapy (HDT) for TB (Kilinç et al., 2021).
In this review, we will systematically discuss the new progress in the research of M. tb acetylation modification and related acetyltransferases. This will provide a theoretical basis and research ideas for exploring the development of novel anti-TB drugs targeting M. tb acetyltransferase, new mechanisms of drug resistance, and precise treatment. Additionally, we will explore how the metabolism of isoniazid (INH), a commonly used drug in TB therapy, depends on the N-acetyl transferase 2 (NAT2) enzyme.
In brief, the review will expatiate the following three parts:
1. acetylation modification of proteins & the role of acetyltransferase in M. tb;
2. acetylation modification of proteins in TB patients;
3. N-acetyltransferase acetylation (NAT) polymorphisms & TB treatment.
The whole flow diagram of the article is below (Figure 1).
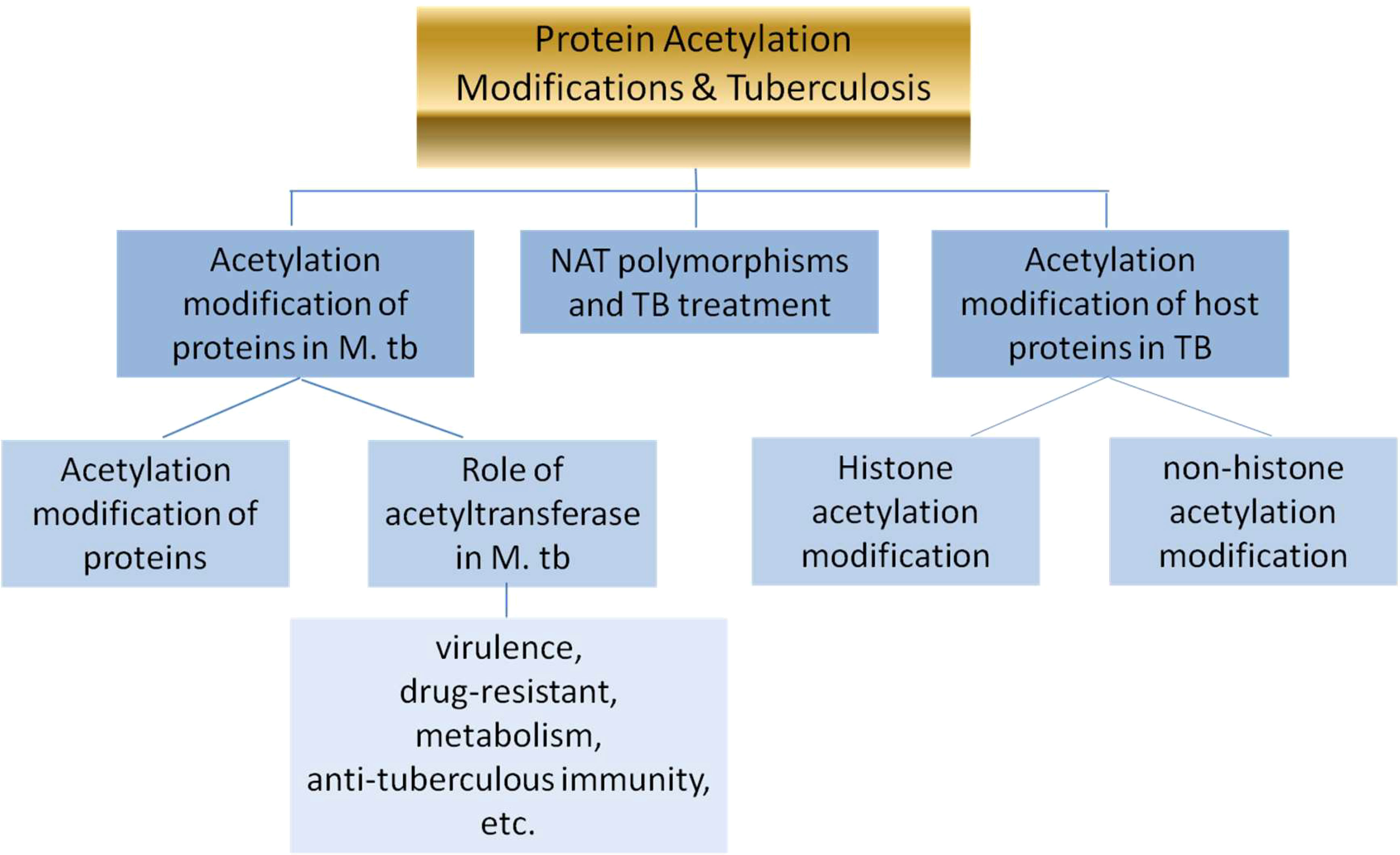
Figure 1 The whole flow diagram of the article.
Acetylation modification of proteins is a critical mechanism of cellular adaptation to changing environmental conditions, and is also implicated in the virulence, pathogenesis and metabolism of M. tb. The recent study has identified 1128 acetylation sites on 658 M. tb proteins, and Gene Ontology (GO) analysis of the acetylome revealed that acetylated proteins are involved in the regulation of diverse cellular processes including metabolism and protein synthesis (Xie et al., 2015). Singh KK. et al. showed that acetylation of response regulator protein MtrA inhibited phosphorylation modifications thereby promoting division of M. tb (Singh et al., 2020). The acetyltransferase of M. tb is involved in the modification of various small molecular substrates, including antibiotics, amino acids, and other molecules, thereby regulating transcription, translation, protein folding, and metabolic pathways. The bioinformatics analysis revealed the existence of 47 potential acetyltransferases in M. tb (Table 1), among which three genes, namely Rv2747, Rv3341, and Rv1653, encode essential acetyltransferases (Xie et al., 2019). Additionally, M. tb acetyltransferase can interact with host immune signaling proteins and modulate the host innate immune response against TB (Burckhardt and Escalante-Semerena, 2020. Shvedunova and Akhtar, 2022). The identification of acetylated proteins and acetyltransferases in M. tb provides a theoretical basis and research ideas for the development of novel anti-TB drugs targeting M. tb acetyltransferase, new mechanisms of drug resistance, and precise treatment.
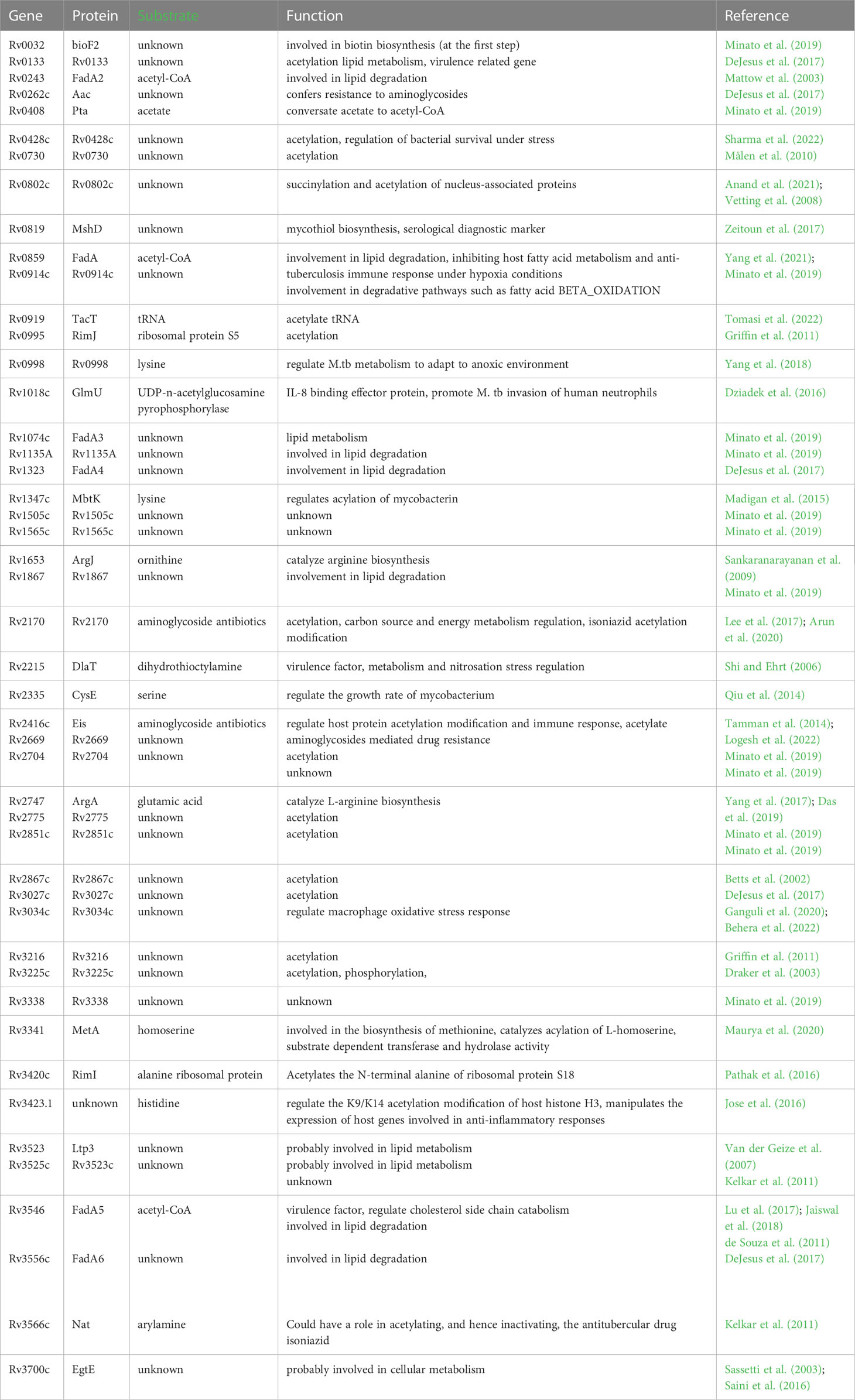
Table 1 47 potential acetyltransferases of M. tb.
The success of M. tb as a pathogen is partly attributed to its ability to sense and respond to dynamic host microenvironments. Protein acetylation modification plays a key role in bacterial virulence and pathogenicity (Ren et al., 2017). Various M. tb acetyltransferases have been identified and confirmed to act as virulence factors. The acetyltransferase Pat, encoded by Rv0998, has been shown to have acetylase activity that is directly regulated by cAMP binding in vitro (Nambi et al., 2010; Xu et al., 2011). Studies have demonstrated that the acetylation of a conserved lysine 193 (K193) within the C-terminal DNA-binding domain of the cyclic AMP (cAMP) receptor protein (CRP) reduces its DNA-binding ability and inhibits transcriptional activity. The reversible acetylation status of CRP K193 has been shown to significantly affect mycobacterial growth phenotype, alter the stress response, and regulate the expression of biologically relevant genes (Di et al., 2023). Shi SP. et al. (Shi and Ehrt, 2006) generated a Rv2215/dlaT knockout strain and tested its ability to grow, resist nitrosative stress, and cause disease in mice, which demonstrated that Rv2215/dlaT is required for optimal growth of M. tb. DlaT encodes dihydrolipoamide acyltransferase, which together with the pyruvate dehydrogenase E1 component (AceE) and dihydrolipoamide dehydrogenase (Lpd) constitutes pyruvate dehydrogenase (PDH) in M. tb. PDH catalyzes the oxidation of pyruvate by NAD to acetyl-coenzyme A (acetyl-CoA) and CO2. Acetyl-CoA then feeds into the tricarboxylic acid (TCA) cycle.
Although the existence of M. tb acetyltransferases as virulence factors has been discovered, the specific targets of these acetyltransferases and the exact molecular mechanisms affecting M. tb virulence remain to be studied and clarified.
The continuing spread of drug-resistant TB is one of the most urgent and difficult challenges facing global TB control. Studies have found that the activity of most of the existing ten kinds of anti-TB drugs, such as aminoglycosides, chloramphenicol, streptomycin, fluoroquinolones and other drugs is regulated by acetylation modification (Schwarz et al., 2004; Reygaert, 2018). For instance, in addition to mutations in katG, nhA, ahpC, kasA, and ndh genes, isoniazid (INH) resistance is associated with acetyltransferase Rv2170, which catalyzes the transfer of acetyl group from acetyl CoA to INH to form acetylated INH (Silva et al., 2003). The acetylated INH is then decomposed into isonicotinic acid and acetylhydrazine, overcoming INH toxicity and producing resistance (Arun et al., 2020). Furthermore, acetylation modification can also affect the metabolic rate of INH in vivo, thereby affecting its therapeutic effect in different individuals (Jing et al., 2020).
The enhanced intracellular survival (Eis) protein encoded by M. tb is an acetyl transferase that targets aminoglycoside antibiotics. Zaunbrecher et al. and Houghton et al. have found that EIS-mediated acetylation modification can inactivate kanamycin, capreomycin and other drugs (Zaunbrecher et al., 2009; Houghton et al., 2013). Reeves et al. found that transcription regulator WhiB7 promoted kanamycin acetylation by enhancing the transcription of Eis genes, and Eis itself was also regulated by acetylation modification (Reeves et al., 2013). Moreover, small molecule inhibitors targeting Eis have also been developed rapidly in recent years. Garzan et al. found that Eis inhibitors can be effectively applied in kanamycin adjuvant combination therapy, which provides a new solution for drug resistance (Garzan et al., 2016; Garzan et al., 2017; Punetha et al., 2020). However, the effect of Eis acetylation on its own activity and its mechanism in regulating aminoglycoside drug resistance remains unclear (Birhanu et al., 2017). Additionally, it has been reported that Rv0262c encoded aminoglycoside 2’-n-acetyltransferase can also acetylate all known aminoglycoside antibiotics, including ribomycin, neomycin B, gentamicin and tobramycin containing 2’ amino, etc. (Hegde et al., 2001). Correspondingly, Rv3225c-encoded acetyltransferase has a low level of aminoglycoside modification activity on aminoglycoside antibiotics, which can lead to resistance of M. tb to aminoglycoside antibiotics through acetylation modification (Kim et al., 2006). Meanwhile, N-acetyl cysteine can artificially increase respiration and additional ROS accumulation, which enhances moxifloxacin lethality in M. tb-infected cultured macrophages and mice. Addition of ROS stimulators to fluoroquinolone treatment of TB constitutes a new direction for suppressing the transition of MDR-TB to XDR-TB (Singh et al., 2022).
In summary, the regulation of drug acetylation modification by M. tb through acetyltransferase is an important cause of drug resistance, according to the studies mentioned above. These studies suggest that small molecule inhibitors targeting M. tb acetyltransferase activity can be developed directly as new anti-TB drugs and can also promote the anti-TB effect of existing drugs by enhancing their sensitivity or preventing drug tolerance.
In the metabolic pathway, approximately 90% of metabolic enzymes in the metabolic pathway, including tricarboxylic acid cycle, gluconeogenesis, glycolysis, glycogen metabolism, fatty acid metabolism, and urea cycle, undergo acetylation modification (Zhao et al., 2010). Rv2170 has been found to possess lysine acetyltransferase activity, which can affect the glyoxylic acid metabolism or tricarboxylic acid cycle by reducing the lysine residues of Isocitrate lyase or Isocitrate dehydrogenase through acetylation modification (Lee et al., 2017). Moreover, the deacetylation of DosR at K182 promotes the hypoxia response in M. tb and enhances the transcription of DosR-targeted genes. Rv0998 has been identified as an acetyltransferase that mediates the acetylation of DosR at K182. Deletion of Rv0998 also promoted the adaptation of M. tb to hypoxia and the transcription of DosR-targeted genes. Mice infected with an M. tb strain containing acetylation-defective DosRK182R had much lower bacterial counts and less severe histopathological impairments compared with those infected with the wild-type strain (Yang et al., 2018). Additionally, Rv0998 has been shown to regulate carbon flux, change oxidation reaction, and reduce tricarboxylic acid cycle reaction, which may contribute to M. tb survival in mice (Bi et al., 2018; Rittershaus et al., 2018). The acetylase activity of Rv0998 is regulated by metabolism-related products, including cAMP, acetyl-CoA, and the deacetylase Rv1151c (Bi et al., 2018). These findings suggest that targeting M. tb acetyltransferase in its own metabolic pathway could be a potential pathway for anti-TB therapy.
Furthermore, TB is linked to human metabolism, and individuals with diabetes and other metabolic disorders have a higher risk of M. tb infection (Bernal et al., 2014). Therefore, investigating the effect of M. tb acetyltransferase on host metabolism could be a promising new strategy for developing anti-TB therapy targeting metabolism-related enzymes.
TB arises from the interplay between bacterial virulence and host immunity. The virulence factors of M. tb enable it to evade the host immune system and survive within the host (Zhu et al., 2019). For example, Kim et al. (Kim et al., 2012) found that Eis protein inhibits JNK-dependent autophagy, phagosome maturation and Reactive Oxygen Species (ROS) production through acetylation of DUSP16/MKP-7 at K55 site. Duan Liang et al. (Duan et al., 2016) found that Eis protein inhibits macrophage autophagy by increasing histone H3 acetylation, up-regulating IL-10 expression, and then activating AKT/mTOR/P70S6K pathway. Rv3423.1, a novel histone acetyltransferase from M. tb, has been shown to mediate acetylation at the H3K9/K14 positions by co-localizing with the host chromatin in the nucleus. By binding to the host chromatin, Rv3423.1 may manipulate the expression of host genes involved in anti-inflammatory responses, allowing M. tb to evade clearance and survive in the intracellular environment (Jose et al., 2016). Another protein secreted by mycobacteria under hypoxia, FadA (Fatty-acid degradation A), acts as an acetyltransferase that converts host acetyl-CoA to acetoacetyl-CoA. This reduces the acetyl-CoA level and suppresses H3K9Ac-mediated expression of the host proinflammatory cytokine Il-6, thereby promoting granuloma progression (Yang et al., 2021). Eis also acetylates M. tb HU (MtHU), which leads to reduced DNA interactions and altered DNA compaction capacity of NAP (Ghosh et al., 2016). Overexpression of Eis can result in excessive acetylation of HU and genomic decompression. Given the importance of HU for M. tb survival, it is possible that its acetylation by Eis is also linked to drug resistance and survival.
Thus, understanding the role of acetyltransferases in host immunity against TB may offer a new therapeutic approach to TB infection.
Protein acetylation and deacetylation is catalyzed by protein acetyltransferases and deacetylases, respectively, of which several families exist. There are two types of protein acetylation: the acetylation of proteins at the ε-amino group and the acetylation of the a-amino group of the N-terminal amino acid (Bernal et al., 2014; Khadela et al., 2022). While the acetylation of the α-amino group of the N-terminal amino acid of proteins is very rare in bacteria, it is frequent in eukaryotes (30 - 80% of proteins) and archaea (14 - 29% of proteins) (Polevoda and Sherman, 2003; Soppa, 2010). On the other hand, the acetylation of proteins at the ε-amino group of internal lysine residues is a widely distributed PTMs, frequent in all domains of life. In eukaryotes, the physiological relevance of N-ε-lysine protein acetylation is well demonstrated. It has been demonstrated that the 8-amino group of multiple lysine side chains in histones can be acetylated to manipulate gene expression by regulating chromatin tightness or influencing transcription factor binding in promoters and distal enhancers, as well as histone DNA interactions (Roth et al., 2001; Yuan et al., 2009; Bannister and Kouzarides, 2011; Barnes et al., 2019). For the first time, Choudhary et al. identified the existence of acetylation modification at 3600 lysine sites on 1750 proteins, suggesting that lysine acetylation has a wide range of regulatory effects (Choudhary et al., 2009). Non-histone acetylation plays a key role in physiological and pathological processes, including the regulation of enzyme activity, protein degradation, protein interaction, subcellular localization, chromatin regulation and metabolism (Drazic et al., 2016; Narita et al., 2019). Abnormal protein acetylation or deacetylation is closely related to many diseases, such as leukemia, cancer, diabetes, infectious diseases, cardiovascular and nervous system related diseases and so on (Timmermann et al., 2001; Morales-Tarré et al., 2021).
During M. tb infection, the host anti-TB immune response was regulated by M. tb-induced gene expression, which is one of the strategies for its intracellular survival and progression of TB. Research reports that M. tb-infected macrophages inhibit histone H3 acetylation (H3Ac) in the interleukin-12B (IL-12B) promoter region, leading to down-regulation of IL-12B expression and inhibition of Th1 type immune response. This promotes M. tb survival in the host (Chandran et al., 2015). Wang et al. found that M. tb infection inhibit HLA-DR gene expression by regulating the recruitment of HDAC complex in the HLA-DR promoter to enable its intracellular survival (Wang et al., 2005). Chen et al. found that the expression of H3K14Ac in peripheral blood lymphocytes of TB patients was reduced, especially the specific enrichment in the promoter region of TNF-α and IL-12B was decreased, which was related to the survival rate of TB patients (Chen et al., 2017). In addition, the up-regulated expression of HDAC1 inhibits the expression of H3K14Ac and plays a role in the outcome of active pulmonary TB and its clinical treatment. Moores et al. found that M. tb regulates the expression of matrix metalloproteinases (MMP-1 and MMP-3) via HDAC and histone acetyltransferase (HAT) activity and the manipulation of histone acetylation modification, which is a key factor in TB immune pathogenesis (Moores et al., 2017). These studies suggest that acetylation of histones, or acetylation of specific lysine sites, is associated with intracellular survival of M. tb and the development of TB.
Recently, the studies of histone acetylome-wide associations (HAWAS) showed that there were at least 2000 differences in acetylation sites associated with differential gene expression in the whole genome of peripheral granulocytes and monocytes of TB patients and healthy people. Histone acetylation quantitative trait locus (haQTL) analysis revealed candidate causal immunophenotypic changes in different populations of granulocyte and monocyte haQTL. M. tb infection regulates the differential enrichment of the inward rectifier potassium channel subfamily promoter J member 15 (CNJ15) of H3K27Ac, which enhances cell apoptosis and promotes M. tb clearance in vitro (Del Rosario et al., 2022). On the other hand, trained immunity, proposed by Netea et al. and Joosten et al., has become an important new evaluation index system for host immune protection induced by TB vaccines (Netea et al., 2016; Joosten et al., 2018). Post-immunization mediated trained immunity (mainly affecting H3K27Ac) of BCG or MTBVAC, an active M. tb candidate vaccine, can enhance the production of cytokines by monocytes and thus provide immune protection (Tarancón et al., 2020; Sheng and Cristea, 2021). Li et al. found that BCG infection can up-regulate the expression level of p300 in mature THP-1 cell lines and regulate the acetylation level of histone H3 and AP-2α. It was further demonstrated that trichostatin A (TSA), a broad-spectrum histone deacetylase inhibitor, enhances the enrichment of the toll-like receptor2 (TLR2) promoter by regulating the acetylation of AP-2α. Furthermore, promoter transcriptional activity was increased to up-regulate TLR2 gene expression (Li et al., 2013). Pennini et al. found that activation of TLR2 inhibits IFN-induced acetylation of histones H3 and H4 (Pennini et al., 2006). Therefore, targeted regulation of acetylation of specific lysine sites in histones may be an important way to enhance the host’s effective resistance to M. tb infection and/or promote immune clearance.
In addition to histone acetylation, non-histone acetylation also plays a crucial role in regulating cellular processes. Like histone proteins, non-histone proteins are also modified by histone acetyltransferases and HDACs. Various studies have reported that non-histone acetylation plays an essential role in the occurrence, development, and outcome of infectious diseases caused by viruses, bacteria, and other pathogens such as DNA virus, influenza virus, rabies virus, and Salmonella typhi infection (Wu et al., 2010; Green et al., 2018; Song et al., 2020). Furthermore, non-histone acetylation modification has been found to regulate autophagy, apoptosis, and inflammasome activation in innate immune responses (Wan et al., 2017; Son et al., 2020; Pagán et al., 2022) Autophagy-related proteins such as ATG5, ATG7, ATC8, and ATG12 can be acetylated by p300, leading to inhibition of autophagy (Lee and Finkel, 2009; Battaglioni et al., 2022; Wang D. et al., 2022). On the other hand, NAD+-dependent histone deacetylase Sirt1 can deacetylate ATG5, ATG7, and LC3, promoting autophagy occurrence (Lee et al., 2008).
In the process of M. tb infection, studies have shown that Sirt1 activation induced by M. tb infection can activate autophagy by directly mediating MAP1LC3B/LC3B deacetylation, which may limit the growth of intracellular M. tb (Iqbal et al., 2021). These findings suggest that acetylated autophagy-related proteins play a key role in regulating autophagy activation and inhibition, and the role of autophagy in host anti-M. tb infection has been established (Pellegrini et al., 2021). Non-histone deacetylation mediated by Sirt family proteins is suggested to be significant in understanding M. tb-mediated inflammatory response and discovering new drug targets (Cheng et al., 2017; Bhaskar et al., 2020; Yang et al., 2021; Yang et al., 2022). In another recent experiment, Brandenburg J. et al. showed that Wnt family member 6 (WNT6) promotes foam cell formation during TB by regulating key lipid metabolism genes including Acetyl Coenzyme A Carboxylase (ACC2). These findings open new perspectives for host-directed adjunctive treatment of pulmonary TB (Brandenburg et al., 2021).
Based on the above studies, non-histone acetylation modification is shown to be effective in host anti-TB immunity. Additionally, non-histone modifications involved in cell signal transduction, protein interaction, protein aggregation, protein degradation, and subcellular localization may also play a critical role in regulating host anti-TB immunity during M. tb infection.
Arylamine N-acetyltransferase comprises N-acetyltransferase 1 (NAT1) and N-acetyltransferase 2 (NAT2) in humans (Hein et al., 2022). NAT2 is mainly expressed in the liver and the GI tract (Husain et al., 2007), and is responsible for the N-acetylation polymorphism observed in human populations (Weber and Hein, 1985; McDonagh et al., 2014; Agundez and Garcia-Martin, 2018; Mitchell, 2020). There are several single nucleotide polymorphisms (SNPs) in the coding exon of the NAT2 gene, which are inherited as NAT2 haplotypes and genotypes and confer rapid, intermediate, and slow acetylator phenotypes that modify drug metabolism (Hein, 2009; Hein and Millner, 2021).
Although isoniazid (INH) remains one of the major first-line drugs, the extensive use of INH to treat active and latent TB infections is compromised by INH-induced hepatotoxicity and liver failure (Hall et al., 2009; Sterling et al., 2020). The NAT2 genotype dependent pharmacokinetic parameters measured in human subjects have been confirmed by measurement of INH N-acetylation both in vitro and in situ in cryopreserved human hepatocytes and the TB patient (Doll et al., 2017; Hein and Millner, 2021). A study conducted in Indonesia revealed that patients with TB and the slow-acetylator phenotype caused by NAT2 variants are highly susceptible to drug-induced liver injury caused by anti-TB drugs, confirming the association between slow-acetylator NAT2 variants and susceptibility to drug-induced liver injury in an Indonesian population (Yuliwulandari et al., 2016). Furthermore, recent studies concluded that INH N-acetylation in human subjects differs significantly with respect to rapid, intermediate, and slow acetylator NAT2 genotypes in terms of plasma half-life, bioavailability (area under the curve), plasma metabolic ratio of INH to N-acetyl-INH, and clearance. The meta-analysis studies report that slow acetylators were significantly more likely to experience hepatotoxicity from INH treatment for TB than rapid acetylators (Khan et al., 2019; Richardson et al., 2019).
Personalized therapy, also known as host-directed therapy (HDT), is being developed in many recent studies for conditions such as TB caused by M. tb. Epigenetic processes, including acetylation modification, play a crucial role in the development of personalized HDT (Marimani et al., 2018). One of the most focused themes among these studies is NAT2 polymorphisms. The paradigm for NAT2 phenotype-dependent dosing strategies is presented as a value of pharmacogenomics-guided isoniazid therapy for the prevention and treatment of TB. Béranger Agathe et al. have demonstrated that NAT2 genotype is the most impactful factor of INH metabolism, compared with low-birth-weight (LBW) and preterm infant born (Béranger et al., 2022). Phenotype-dependent dosing strategies aim to reduce the risk of adverse reactions, increase therapeutic efficacy, reduce costs, and improve patient care and disease prevention. Several studies have proposed pharmacogenomics-guided INH therapy for TB (Matsumoto et al., 2014; Jung et al., 2015; Choi et al., 2017; Motta et al., 2018; Suvichapanich et al., 2018; Jing et al., 2020). A recent study conducted in the USA and Brazil found that the clearance rates of INH were lowest in predicted slow acetylators (median 19.3 L/hr), moderate in intermediate acetylators (median 41.0 L/hr), and highest in fast acetylators (median 46.7 L/hr) (Verma et al., 2021). Moreover, there are significant differences in the distribution of NAT2 gene polymorphisms among different nationalities and races, the anti-TB treatment regimens adopted by patients are different, and the tolerance and exclusion standards of the subjects are different (Suarez-Kurtz et al., 2016; Zahra et al., 2020; Zhang et al., 2021). Depending on the NAT2 genotype of the patients, several studies have evaluated isoniazid doses of 2.5 mg/kg (0.5 times standard dose), 5 mg/kg (standard dose), and 7.5 mg/kg (1.5 times standard dose) for slow, intermediate, and fast metabolizers, respectively. As a result, a better treatment success rate was achieved, and the occurrence of liver function injury was reduced (Azuma et al., 2013; Huerta-García et al., 2020).
All these results suggest that understanding the diversity of drug-related genetic markers is critical for individualized drug-gene therapy programs in ethnic minorities in China and populations highly mixed with these ethnic groups. The above studies could make personalized TB treatment dosing available in reality. Pharmacogenomic-guided dosing can help achieve consistent drug levels and improve clinical outcomes.
As one main type of epigenetics in TB, acetylation plays a crucial role in aiding M. tb survival in the host, rendering the host vulnerable to the pathogen, and activating the host’s immune system against the invading pathogen. Hence, the study of acetylation processes is crucial for comprehending the progression of M. tb, identifying ideal candidates for therapeutic targets, minimizing drug toxicity, and monitoring the efficacy of administered therapy in developing personalized medication regimens. There are still some limitations of current research on protein acetylation within TB. We still lack dynamic change analysis of the acetylated protein expression. Future research may focus on the dynamic changes of acetylated protein expression in M. tb and host at different time points, which will provide more effective HDT targets for drug treatment of TB.
YH and CZ contributed to conception, design and drafting the manuscript. LP and ZZ contributed to conception and critically revised the manuscript. All authors contributed to the article and approved the submitted version.
This work was supported by the National Natural Science Foundation of China (82172279) and the Beijing Hospitals Authority Innovation Studio of Young Staff Funding (202136).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Agundez, J. A., Garcia-Martin, E. (2018). “Chapter 1.3, arylamine n-acetyltransferases in health and disease: from pharmacogenetics to drug discovery and diagnostics,” in Human arylamine n-acetyltransferases type 2: phenotype correlation with genotypeA clinical perspective. Eds. Laurieri, N., Sim, E. (Singapore: World Scientific Publishing), ISBN: ISBN: 978-981-3232-00-6.
Allfrey, V. G., Faulkner, R., Mirsky, A. E. (1964). Acetylation and methylation of histones and their possible role in the regulation of RNA synthesis. Proc. Natl. Acad. Sci. U.S.A. 51 (5), 786–794. doi: 10.1073/pnas.51.5.786
Anand, C., Santoshi, M., Singh, P. R., Nagaraja, V. (2021). Mycobacterium tuberculosis Rv0802c is an acyltransferase that succinylates and acetylates nucleoid-associated protein HU. Microbiol. (Reading) 167 (7). doi: 10.1099/mic.0.001058
Arora, G., Bothra, A., Prosser, G., Arora, K., Sajid, A. (2021). Role of post-translational modifications in the acquisition of drug resistance in mycobacterium tuberculosis. FEBS J. 288 (11), 3375–3393. doi: 10.1111/febs.15582
Arun, K. B., Madhavan, A., Abraham, B., Balaji, M., Sivakumar, K. C., Nisha, P., et al. (2020). Acetylation of isoniazid is a novel mechanism of isoniazid resistance in mycobacterium tuberculosis. Antimicrob. Agents Chemother. 65 (1). doi: 10.1128/AAC.00456-20
Azuma, J., Masako, O., Kubota, R., Yokota, S., Nagai, T., Tsuyuguchi, K., et al. (2013). NAT2 genotype guided regimen reduces isoniazid-induced liver injury and early treatment failure in the 6-month four-drug standard treatment of tuberculosis: a randomized controlled trial for pharmacogenetics-based therapy. Eur. J. Clin. Pharmacol. 69 (5), 1091–1101. doi: 10.1007/s00228-012-1429-9
Bannister, A. J., Kouzarides, T. (2011). Regulation of chromatin by histone modifications. Cell Res. 21 (3), 381–395. doi: 10.1038/cr.2011.22
Barnes, C. E., English David, M., Cowley Shaun, M. (2019). Acetylation & Co: an expanding repertoire of histone acylations regulates chromatin and transcription. Essays Biochem. 63 (1), 97–107. doi: 10.1042/EBC20180061
Battaglioni, S., Benjamin, D., Wälchli, M., Maier, T., Hall, M. N. (2022). mTOR substrate phosphorylation in growth control. Cell 185 (11), 1814–1836. doi: 10.1016/j.cell.2022.04.013
Behera, A., Jain, P., Ganguli, G., Biswas, M., Padhi, A., Pattanaik, K. P., et al. (2022). Mycobacterium tuberculosis acetyltransferase suppresses oxidative stress by inducing peroxisome formation in macrophages. Int. J. Mol. Sci. 23 (5). doi: 10.3390/ijms23052584
Béranger, A., Bekker, A., Solans, Belén P., Cotton, M. F., Mirochnick, M., Violari, A., et al. (2022). Influence of NAT2 genotype and maturation on isoniazid exposure in low-Birth-Weight and preterm infants with or without human immunodeficiency virus (HIV) exposure. Clin. Infect. Dis. 75 (6), 1037–1045. doi: 10.1093/cid/ciac001
Bernal, V., Castaño-Cerezo, S., Gallego-Jara, J., Écija-Conesa, A., de Diego, T., Iborra, JoséL., et al. (2014). Regulation of bacterial physiology by lysine acetylation of proteins. N Biotechnol. 31 (6), 586–595. doi: 10.1016/j.nbt.2014.03.002
Betts, J. C., Lukey, P. T., Robb, L. C., McAdam, R. A., Duncan, K. (2002). Evaluation of a nutrient starvation model of mycobacterium tuberculosis persistence by gene and protein expression profiling. Mol. Microbiol. 43 (3), 717–731. doi: 10.1046/j.1365-2958.2002.02779.x
Bhaskar, A., Kumar, S., Khan, M. Z., Singh, A., Dwivedi, V. P., Nandicoori, V. K. (2020). Host sirtuin 2 as an immunotherapeutic target against tuberculosis. Elife 9. doi: 10.7554/eLife.55415
Bi, J., Gou, Z., Fengzhu, Z., Chen, Y., Gan, J., Liu, J., et al. (2018). Acetylation of lysine 182 inhibits the ability of mycobacterium tuberculosis DosR to bind DNA and regulate gene expression during hypoxia. Emerg. Microbes Infect. 7 (1), 108. doi: 10.1038/s41426-018-0112-3
Birhanu, A. G., Yimer, S. A., Holm-Hansen, C., Norheim, G., Aseffa, A., Abebe, M., et al. (2017). N- and O-acetylation in mycobacterium tuberculosis lineage 7 and lineage 4 strains: proteins involved in bioenergetics, virulence, and antimicrobial resistance are acetylated. J. Proteome Res. 16 (11), 4045–4059. doi: 10.1021/acs.jproteome.7b00429
Brandenburg, J., Marwitz, S., Tazoll, S. C., Waldow, F., Kalsdorf, B., Vierbuchen, T., et al. (2021). WNT6/ACC2-induced storage of triacylglycerols in macrophages is exploited by mycobacterium tuberculosis. J. Clin. Invest. 131 (16). doi: 10.1172/JCI141833
Budzik, J. M., Swaney, D. L., Jimenez-Morales, D., Johnson, J. R., Garelis, N. E., Repasy, T., et al. (2020). Dynamic post-translational modification profiling of mycobacterium tuberculosis-infected primary macrophages. Elife 9 (undefined). doi: 10.7554/eLife.51461
Burckhardt, R. M., Escalante-Semerena, J. C. (2020). NSmall-molecule acetylation by GCN5-related -acetyltransferases in bacteria. Microbiol. Mol. Biol. Rev. 84 (2). doi: 10.1128/MMBR.00090-19
Carabetta, V. J., Cristea, I. M. (2017). Regulation, function, and detection of protein acetylation in bacteria. J. Bacteriol. 199 (16). doi: 10.1128/JB.00107-17
Chandran, A., Antony, C., Jose, L., Mundayoor, S., Natarajan, K., Kumar, R.A. (2015). Mycobacterium tuberculosis infection induces HDAC1-mediated suppression of IL-12B gene expression in macrophages. Front. Cell Infect. Microbiol. 5 (undefined). doi: 10.3389/fcimb.2015.00090
Chen, Y.-C., Chao, T.-Y., Leung, S.-Y., Chen, C.-J., Wu, C.-C., Fang, W.-F., et al. (2017). Histone H3K14 hypoacetylation and H3K27 hypermethylation along with HDAC1 up-regulation and KDM6B down-regulation are associated with active pulmonary tuberculosis disease. Am. J. Transl. Res. 9 (4), 1943–1955.
Cheng, C. Y., Gutierrez, N. M., Marzuki, M. B., Lu, X., Foreman, T. W., Paleja, B., et al (2017). Host sirtuin 1 regulates mycobacterial immunopathogenesis and represents a therapeutic target against tuberculosis. Sci. Immunol. 2 (9). doi: 10.1126/sciimmunol.aaj1789
Choi, R., Jeong, B. Ho, Koh, W. J., Lee, S. Y. (2017). Recommendations for optimizing tuberculosis treatment: therapeutic drug monitoring, pharmacogenetics, and nutritional status considerations. Ann. Lab. Med. 37 (2), 97–107. doi: 10.3343/alm.2017.37.2.97
Choudhary, C., Kumar, C., Gnad, F., Nielsen, M. L., Rehman, M., Walther, T. C., et al. (2009). Lysine acetylation targets protein complexes and co-regulates major cellular functions. Science 325 (5942), 834–840. doi: 10.1126/science.1175371
Christensen, D. G., Baumgartner, J. T., Xie, X., Jew, K. M., Basisty, N., Schilling, B., et al. (2019). Mechanisms, detection, and relevance of protein acetylation in prokaryotes. mBio 10 (2). doi: 10.1128/mBio.02708-18
Das, U., Singh, E., Dharavath, S., Tiruttani, S. U. K., Pal, R. K., Vijayan, R., et al. (2019). Structural insights into the substrate binding mechanism of novel ArgA from mycobacterium tuberculosis. Int. J. Biol. Macromol. 125 (undefined), 970–978. doi: 10.1016/j.ijbiomac.2018.12.163
DeJesus, M. A., Gerrick, E. R., Xu, W., Park, S. W., Long, J. E., Boutte, C. C., et al. (2017). Comprehensive essentiality analysis of the mycobacterium tuberculosis genome via saturating transposon mutagenesis. mBio 8 (1). doi: 10.1128/mBio.02133-16
Del Rosario, R. C.H., Poschmann, J., Lim, C., Cheng, C. Y., Kumar, P., Riou, C., et al. (2022). Histone acetylome-wide associations in immune cells from individuals with active mycobacterium tuberculosis infection. Nat. Microbiol. 7 (2), 312–326. doi: 10.1038/s41564-021-01049-w
de Souza, Gustavo A., Leversen, N. A., Målen, H., Wiker, H. G. (2011). Bacterial proteins with cleaved or uncleaved signal peptides of the general secretory pathway. J. Proteomics 75 (2), 502–510. doi: 10.1016/j.jprot.2011.08.016
Di, Y., Xu, S., Chi, M., Hu, Y., Zhang, X., Wang, H., et al. (2023). Acetylation of cyclic AMP receptor protein by acetyl phosphate modulates mycobacterial virulence. Microbiol. Spectr. 11 (1), e0400222. doi: 10.1128/spectrum.04002-22
Doll, M. A., Salazar-González, RaúlA, Bodduluri, S., Hein, D. W. (2017). NArylamine -acetyltransferase 2 genotype-dependent -acetylation of isoniazid in cryopreserved human hepatocytes. Acta Pharm. Sin. B. 7 (4), 517–522. doi: 10.1016/j.apsb.2017.05.003
Draker, K.-A., Boehr, D. D., Elowe, N. H., Noga, T. J., Wright, G. D. (2003). Functional annotation of putative aminoglycoside antibiotic modifying proteins in mycobacterium tuberculosis H37Rv. J. Antibiot (Tokyo) 56 (2), 135–142. doi: 10.7164/antibiotics.56.135
Drazic, A., Myklebust, L. M., Ree, R., Arnesen, T. (2016). The world of protein acetylation. Biochim. Biophys. Acta 1864 (10), 1372–1401. doi: 10.1016/j.bbapap.2016.06.007
Duan, L., Yi, M., Chen, J., Li, S., Chen, W. (2016). Mycobacterium tuberculosis EIS gene inhibits macrophage autophagy through up-regulation of IL-10 by increasing the acetylation of histone H3. Biochem. Biophys. Res. Commun. 473 (4), 1229–1234. doi: 10.1016/j.bbrc.2016.04.045
Dziadek, B., Brzostek, A., Grzybowski, M., Fol, M., Krupa, A., Kryczka, J., et al. (2016). Mycobacterium tuberculosis AtsG (Rv0296c), GlmU (Rv1018c) and SahH (Rv3248c) proteins function as the human IL-8-Binding effectors and contribute to pathogen entry into human neutrophils. PloS One 11 (2), e0148030. doi: 10.1371/journal.pone.0148030
Finkel, T., Deng, C.-X., Mostoslavsky, R. (2009). Recent progress in the biology and physiology of sirtuins. Nature 460 (7255), 587–591. doi: 10.1038/nature08197
Ganguli, G., Pattanaik, K. P., Jagadeb, M., Sonawane, A. (2020). Mycobacterium tuberculosis Rv3034c regulates mTORC1 and PPAR-γ dependant pexophagy mechanism to control redox levels in macrophages. Cell Microbiol. 22 (9), e13214. doi: 10.1111/cmi.13214
Garzan, A., Willby, M. J., Green, K. D., Tsodikov, O. V., Posey, J. E., Garneau-Tsodikova, S. (2016). M. tuberculosis discovery and optimization of two eis inhibitor families as kanamycin adjuvants against drug-resistant. ACS Med. Chem. Lett. 7 (12), 1219–1221. doi: 10.1021/acsmedchemlett.6b00261
Garzan, A., Willby, M. J., Ngo, H. X., Gajadeera Chathurada, S., Green Keith, D., Holbrook Selina, Y. L., et al. (2017). Combating enhanced intracellular survival (Eis)-mediated kanamycin resistance of mycobacterium tuberculosis by novel Pyrrolo[1,5-a]pyrazine-Based eis inhibitors. ACS Infect. Dis. 3 (4), 302–309. doi: 10.1021/acsinfecdis.6b00193
Ghosh, S., Padmanabhan, B., Anand, C., Nagaraja, V. (2016). Lysine acetylation of the mycobacterium tuberculosis HU protein modulates its DNA binding and genome organization. Mol. Microbiol. 100 (4), 577–588. doi: 10.1111/mmi.13339
Glozak, M. A., Sengupta, N., Zhang, X., Seto, E. (2005). Acetylation and deacetylation of non-histone proteins. Gene 363 (undefined), 15–23. doi: 10.1016/j.gene.2005.09.010
Green, K. D., Biswas, T., Pang, A. H., Willby, M. J., Reed, M. S., Stuchlik, O., et al. (2018). Acetylation by eis and deacetylation by Rv1151c of mycobacterium tuberculosis HupB: biochemical and structural insight. Biochemistry 57 (5), 781–790. doi: 10.1021/acs.biochem.7b01089
Griffin, J. E., Gawronski, J. D., Dejesus, M. A., Ioerger, T. R., Akerley, B. J., Sassetti, C. M. (2011). High-resolution phenotypic profiling defines genes essential for mycobacterial growth and cholesterol catabolism. PloS Pathog. 7 (9), e1002251. doi: 10.1371/journal.ppat.1002251
Hall, R. G., Leff, R. D., Gumbo, T. (2009). Treatment of active pulmonary tuberculosis in adults: current standards and recent advances. insights from the society of infectious diseases pharmacists. Pharmacotherapy 29 (12), 1468–1481. doi: 10.1592/phco.29.12.1468
Hegde, S. S., Javid-Majd, F., Blanchard, J. S. (2001). Overexpression and mechanistic analysis of chromosomally encoded aminoglycoside 2’-n-acetyltransferase (AAC(2’)-ic) from mycobacterium tuberculosis. J. Biol. Chem. 276 (49), 45876–45881. doi: 10.1074/jbc.M108810200
Hein, D. W. (2009). N-acetyltransferase SNPs: emerging concepts serve as a paradigm for understanding complexities of personalized medicine. Expert Opin. Drug Metab. Toxicol. 5 (4), 353–366. doi: 10.1517/17425250902877698
Hein, D. W., Doll, M. A., Habil, M. R. (2022). NHuman -acetyltransferase 1 and 2 differ in affinity towards acetyl-coenzyme a cofactor and -Hydroxy-Arylamine carcinogens. Front. Pharmacol. 13 (undefined). doi: 10.3389/fphar.2022.821133
Hein, D. W., Millner, L. M. (2021). Arylamine n-acetyltransferase acetylation polymorphisms: paradigm for pharmacogenomic-guided therapy- a focused review. Expert Opin. Drug Metab. Toxicol. 17 (1), 9–21. doi: 10.1080/17425255.2021.1840551
Houghton, J. L., Green, K. D., Pricer, R. E., Mayhoub, A. S., Garneau-Tsodikova, S. (2013). Unexpected n-acetylation of capreomycin by mycobacterial eis enzymes. J. Antimicrob. Chemother. 68 (4), 800–805. doi: 10.1093/jac/dks497
Huerta-García, A. P., Medellín-Garibay, S. E., Ortiz-Álvarez, A., Magaña-Aquino, M., Rodríguez-Pinal, C. J., Portales-Pérez, D. P., et al. (2020). Population pharmacokinetics of isoniazid and dose recommendations in Mexican patients with tuberculosis. Int. J. Clin. Pharm. 42 (4), 1217–1226. doi: 10.1007/s11096-020-01086-1
Husain, A., Zhang, X., Doll, M. A., States, J.C., Barker, D. F., Hein, D. W. (2007). Identification of n-acetyltransferase 2 (NAT2) transcription start sites and quantitation of NAT2-specific mRNA in human tissues. Drug Metab. Dispos. 35 (5), 721–727. doi: 10.1124/dmd.106.014621
Iqbal, I. K., Bajeli, S., Sahu, S., Bhat, S. A., Kumar, A. (2021). Hydrogen sulfide-induced GAPDH sulfhydration disrupts the CCAR2-SIRT1 interaction to initiate autophagy. Autophagy 17 (11), 3511–3529. doi: 10.1080/15548627.2021.1876342
Jaiswal, A. K., Husaini, S. H. A., Amarjeet., K., Subbarao, N. (2018). Designing novel inhibitors against mycobacterium tuberculosis FadA5 (acetyl-CoA acetyltransferase) by virtual screening of known anti-tuberculosis (bioactive) compounds. Bioinformation 14 (6), 327–336. doi: 10.6026/97320630014327
Jing, W., Zong, Z., Tang, B., Wang, J., Zhang, T., Wen, Shu’an, et al. (2020). Population pharmacokinetic analysis of isoniazid among pulmonary tuberculosis patients from China. Antimicrob. Agents Chemother. 64 (3). doi: 10.1128/AAC.01736-19
Joosten, S. A., van Meijgaarden, K. E., Arend, S. M., Prins, C., Oftung, F., Korsvold, G. E., et al. (2018). Mycobacterial growth inhibition is associated with trained innate immunity. J. Clin. Invest. 128 (5), 1837–1851. doi: 10.1172/JCI97508
Jose, L., Ramachandran, R., Bhagavat, R., Gomez, R. L., Chandran, A., Raghunandanan, S., et al. (2016). Hypothetical protein Rv3423.1 of mycobacterium tuberculosis is a histone acetyltransferase. FEBS J. 283 (2), 265–281. doi: 10.1111/febs.13566
Jung, J. Ah, Kim, T.-E., Lee, H., Jeong, B.-H., Park, H. Y., Jeon, K., et al. (2015). A proposal for an individualized pharmacogenetic-guided isoniazid dosage regimen for patients with tuberculosis. Drug Des. Devel Ther. 9 (undefined), 5433–5438. doi: 10.2147/DDDT.S87131
Kelkar, D. S., Kumar, D., Kumar, P., Balakrishnan, L., Muthusamy, B., Yadav, A. K., et al. (2011). Proteogenomic analysis of mycobacterium tuberculosis by high resolution mass spectrometry. Mol. Cell Proteomics 10 (12), M111.011627. doi: 10.1074/mcp.M111.011445
Khadela, A., Chavda, V. P., Postwala, H., Shah, Y., Mistry, P., Apostolopoulos, V. (2022). Epigenetics in tuberculosis: immunomodulation of host immune response. Vaccines (Basel) 10 (10). doi: 10.3390/vaccines10101740
Khan, S., Mandal, R. K., Elasbali, A. M., Dar, S. A., Jawed, A., Wahid, M., et al. (2019). NAT2Pharmacogenetic association between gene polymorphisms and isoniazid induced hepatotoxicity: trial sequence meta-analysis as evidence. Biosci. Rep. 39 (1). doi: 10.1042/BSR20180845
Kilinç, Gül, Saris, A., Ottenhoff, T. H.M., Haks, MariëlleC (2021). Host-directed therapy to combat mycobacterial infections. Immunol. Rev. 301 (1), 62–83. doi: 10.1111/imr.12951
Kim, K. H., An, D. Ri, Song, J., Yoon, Ji Y, Kim, H. S., Yoon, H. J., et al. (2012). Mycobacterium tuberculosis eis protein initiates suppression of host immune responses by acetylation of DUSP16/MKP-7. Proc. Natl. Acad. Sci. U S A. 109 (20), 7729–7734. doi: 10.1073/pnas.1120251109
Kim, S. C., Sprung, R., Chen, Y., Xu, Y., Ball, H., Pei, J., et al. (2006). Substrate and functional diversity of lysine acetylation revealed by a proteomics survey. Mol. Cell. 23 (4), 607–618. doi: 10.1016/j.molcel.2006.06.026
Kim, G.-W., Yang, X.-J. (2011). Comprehensive lysine acetylomes emerging from bacteria to humans. Trends Biochem. Sci. 36 (4), 211–220. doi: 10.1016/j.tibs.2010.10.001
Lee, In H, Cao, L., Mostoslavsky, R., Lombard, D. B., Liu, J., Bruns, N. E., et al. (2008). A role for the NAD-dependent deacetylase Sirt1 in the regulation of autophagy. Proc. Natl. Acad. Sci. U S A. 105 (9), 3374–3379. doi: 10.1073/pnas.0712145105
Lee, In H, Finkel, T. (2009). Regulation of autophagy by the p300 acetyltransferase. J. Biol. Chem. 284 (10), 6322–6328. doi: 10.1074/jbc.M807135200
Lee, W., VanderVen, B. C., Walker, S., Russell, D. G. (2017). Novel protein acetyltransferase, Rv2170, modulates carbon and energy metabolism in mycobacterium tuberculosis. Sci. Rep. 7 (1), 72. doi: 10.1038/s41598-017-00067-1
Li, M., Li, Xi, Wang, E., Luo, E. (2013). Upregulation of toll-like receptor 2 gene expression by acetylation of AP-2 alpha in THP-1 cells, a human monocytic cell line. Int. J. Biochem. Cell Biol. 45 (8), 1594–1599. doi: 10.1016/j.biocel.2013.05.003
Logesh, R., Lavanya, V., Jamal, S., Ahmed, N. (2022). Designing of a chimeric vaccine using EIS (Rv2416c) protein against mycobacterium tuberculosis H37Rv: an immunoinformatics approach. Appl. Biochem. Biotechnol. 194 (1), 187–214. doi: 10.1007/s12010-021-03760-0
Lu, R., Schaefer, C. M., Nesbitt, N. M., Kuper, J., Kisker, C., Sampson Nicole, S. (2017). Catabolism of the cholesterol side chain in mycobacterium tuberculosis is controlled by a redox-sensitive thiol switch. ACS Infect. Dis. 3 (9), 666–675. doi: 10.1021/acsinfecdis.7b00072
Madigan, C. A., Martinot, A. J., Wei, J.-R., Madduri, A., Cheng, T.-Y., Young, D. C., et al. (2015). Lipidomic analysis links mycobactin synthase K to iron uptake and virulence in m. tuberculosis. PloS Pathog. 11 (3), e1004792. doi: 10.1371/journal.ppat.1004792
Målen, H., Pathak, S., Søfteland, T., de Souza Gustavo, A., Wiker, H. G. (2010). Definition of novel cell envelope associated proteins in triton X-114 extracts of mycobacterium tuberculosis H37Rv. BMC Microbiol. 10 (undefined), 132. doi: 10.1186/1471-2180-10-132
Marimani, M., Ahmad, A., Duse, A. (2018). The role of epigenetics, bacterial and host factors in progression of mycobacterium tuberculosis infection. Tuberculosis 113, undefined 113, 200–214. doi: 10.1016/j.tube.2018.10.009
Matsumoto, T., Ohno, M., Azuma, J. (2014). Future of pharmacogenetics-based therapy for tuberculosis. Pharmacogenomics 15 (5), 601–607. doi: 10.2217/pgs.14.38
Mattow, J., Schaible, U. E., Schmidt, F., Hagens, K., Siejak, F., Brestrich, G., et al. (2003). Comparative proteome analysis of culture supernatant proteins from virulent mycobacterium tuberculosis H37Rv and attenuated m. bovis BCG Copenhagen. Electrophoresis 24 (null), 3405–3420. doi: 10.1002/elps.200305601
Maurya, B., Pochet, L., Wouters, J., Colaço, M., Misquith, S. (2020). MetA (Rv3341) from mycobacterium tuberculosis H37Rv strain exhibits substrate dependent dual role of transferase and hydrolase activity. Biochimie 179 (undefined), 113–126. doi: 10.1016/j.biochi.2020.09.013
McDonagh, E. M., Boukouvala, S., Aklillu, E., Hein David, W., Altman Russ, B., Klein Teri, E. (2014). PharmGKB summary: very important pharmacogene information for n-acetyltransferase 2. Pharmacogenet. Genomics 24 (8), 409–425. doi: 10.1097/FPC.0000000000000062
Minato, Y., Gohl, D. M., Thiede, J. M., Chacón, J. M., Harcombe, W. R., Maruyama, F., et al. (2019). Genomewide assessment of mycobacterium tuberculosis conditionally essential metabolic pathways. mSystems 4 (4). doi: 10.1128/mSystems.00070-19
Mitchell, S. C. (2020). N-acetyltransferase: the practical consequences of polymorphic activity in man. Xenobiotica 50 (1), 77–91. doi: 10.1080/00498254.2019.1618511
Moores, R. C., Brilha, S., Schutgens, F., Elkington, P. T., Friedland, J. S. (2017). Epigenetic regulation of matrix metalloproteinase-1 and -3 expression in mycobacterium tuberculosis infection. Front. Immunol. 8. doi: 10.3389/fimmu.2017.00602
Morales-Tarré, O., Alonso-Bastida, R., Arcos-Encarnación, B., Pérez-Martínez, L., Encarnación-Guevara, S. (2021). Protein lysine acetylation and its role in different human pathologies: a proteomic approach. Expert Rev. Proteomics. 18 (11), 949–975. doi: 10.1080/14789450.2021.2007766
Motta, I., Calcagno, A., Bonora, S. (2018). Pharmacokinetics and pharmacogenetics of anti-tubercular drugs: a tool for treatment optimization? Expert Opin. Drug Metab. Toxicol. 14 (1), 59–82. doi: 10.1080/17425255.2018.1416093
Nakayasu, E. S., Burnet, M. C., Walukiewicz, H. E., Wilkins, C. S., Shukla, A. K., Brooks, S., et al. (2017). Ancient regulatory role of lysine acetylation in central metabolism. mBio 8 (6). doi: 10.1128/mBio.01894-17
Nambi, S., Basu, N., Visweswariah, S. S. (2010). cAMP-regulated protein lysine acetylases in mycobacteria. J. Biol. Chem. 285 (32), 24313–24323. doi: 10.1074/jbc.M110.118398
Narita, T., Weinert, B. T., Choudhary, C. (2019). Functions and mechanisms of non-histone protein acetylation. Nat. Rev. Mol. Cell Biol. 20 (3), 156–174. doi: 10.1038/s41580-018-0081-3
Netea, M. G., Joosten, L. A.B., Latz, E., Mills, K. H.G., Natoli, G., Stunnenberg, H. G., et al. (2016). Trained immunity: a program of innate immune memory in health and disease. Science 352 (6284), aaf1098. doi: 10.1126/science.aaf1098
Norris, K. L., Lee, J.-Y., Yao, T.-P. (2009). Acetylation goes global: the emergence of acetylation biology. Sci. Signal 2 (97), pe76. doi: 10.1126/scisignal.297pe76
Pagán, A. J., Lee, L. J., Edwards-Hicks, J., Moens, C. B., Tobin, D. M., Busch-Nentwich, E. M., et al. (2022). mTOR-regulated mitochondrial metabolism limits mycobacterium-induced cytotoxicity. Cell 185 (20), 3720–3738.e13. doi: 10.1016/j.cell.2022.08.018
Pathak, D., Bhat, A. H., Sapehia, V., Rai, J., Rao, A. (2016). Biochemical evidence for relaxed substrate specificity of nα-acetyltransferase (Rv3420c/rimI) of mycobacterium tuberculosis. Sci. Rep. 6 (undefined), 28892. doi: 10.1038/srep28892
Pawlowski, A., Jansson, M., Sköld, M., Rottenberg, M. E., Källenius, G. (2012). Tuberculosis and HIV co-infection. PloS Pathog. 8 (2), e1002464. doi: 10.1371/journal.ppat.1002464
Pellegrini, J. M., Tateosian, N. L., Morelli, M. P., García, V. E. (2021). Shedding light on autophagy during human tuberculosis. A Long Way to Go. Front. Cell Infect. Microbiol. 11. doi: 10.3389/fcimb.2021.820095
Pennini, M. E., Pai, R. K., Schultz, D. C., Boom, W.H., Harding, C. V. (2006). Mycobacterium tuberculosis 19-kDa lipoprotein inhibits IFN-gamma-induced chromatin remodeling of MHC2TA by TLR2 and MAPK signaling. J. Immunol. 176 (7), 4323–4330. doi: 10.4049/jimmunol.176.7.4323
Polevoda, B., Sherman, F. (2003). N-terminal acetyltransferases and sequence requirements for n-terminal acetylation of eukaryotic proteins. J. Mol. Biol. 325 (4), 595–622. doi: 10.1016/s0022-2836(02)01269-x
Punetha, A., Ngo, H. X., Holbrook, S. Y.L., Green, K. D., Willby, M. J., Bonnett, S. A., et al. (2020). Structure-guided optimization of inhibitors of acetyltransferase eis from mycobacterium tuberculosis. ACS Chem. Biol. 15 (6), 1581–1594. doi: 10.1021/acschembio.0c00184
Qiu, J., Ma, Y., Owusu, L., Jiang, T., Xin, Yi (2014). Functional analysis of serine acetyltransferase from mycobacterium smegmatis. J. Basic Microbiol. 54 (7), 670–677. doi: 10.1002/jobm.201300858
Reeves, A. Z., Campbell, P. J., Sultana, R., Malik, S., Murray, M., Plikaytis, B. B., et al. (2013). Aminoglycoside cross-resistance in mycobacterium tuberculosis due to mutations in the 5’ untranslated region of whiB7. Antimicrob. Agents Chemother. 57 (4), 1857–1865. doi: 10.1128/AAC.02191-12
Ren, J., Sang, Yu, Lu, J., Yao, Y.-F. (2017). Protein acetylation and its role in bacterial virulence. Trends Microbiol. 25 (9), 768–779. doi: 10.1016/j.tim.2017.04.001
Reygaert, W. C. (2018). An overview of the antimicrobial resistance mechanisms of bacteria. AIMS Microbiol. 4 (3), 482–501. doi: 10.3934/microbiol.2018.3.482
Richardson, M., Kirkham, J., Dwan, K., Sloan, D. J., Davies, G., Jorgensen, A. L. (2019). NAT2 variants and toxicity related to anti-tuberculosis agents: a systematic review and meta-analysis. Int. J. Tuberc Lung Dis. 23 (3), 293–305. doi: 10.5588/ijtld.18.0324
Riou, J., Althaus, C. L. (2020). Pattern of early human-to-human transmission of wuhan 2019 novel coronavirus, (2019-nCoV), December 2019 to January 2020. Euro Surveill 25 (4). doi: 10.2807/1560-7917.ES.2020.25.4.2000058
Rittershaus, E. S.C., Baek, S.-H., Krieger, I. V., Nelson, S. J., Cheng, Y.-S., Nambi, S., et al. (2018). A lysine acetyltransferase contributes to the metabolic adaptation to hypoxia in mycobacterium tuberculosis. Cell Chem. Biol. 25 (12), 1495–1505.e3. doi: 10.1016/j.chembiol.2018.09.009
Roth, S. Y., Denu, J. M., Allis, C. D. (2001). Histone acetyltransferases. Annu. Rev. Biochem. 70 (undefined), 81–120. doi: 10.1146/annurev.biochem.70.1.81
Saini, V., Cumming, B. M., Guidry, L., Lamprecht, D. A., Adamson, J. H., Reddy, V. P., et al. (2016). Ergothioneine maintains redox and bioenergetic homeostasis essential for drug susceptibility and virulence of mycobacterium tuberculosis. Cell Rep. 14 (3), 572–585. doi: 10.1016/j.celrep.2015.12.056
Sankaranarayanan, R., Garen, C. R., Cherney, M. M., Yuan, M., Lee, C., James, M. N. G. (2009). Preliminary X-ray crystallographic analysis of ornithine acetyltransferase (Rv1653) from mycobacterium tuberculosis. Acta Crystallogr. Sect F Struct. Biol. Cryst Commun. 65 (null), 173–176. doi: 10.1107/S1744309109000360
Sassetti, C. M., Boyd, D. H., Rubin, E. J. (2003). Genes required for mycobacterial growth defined by high density mutagenesis. Mol. Microbiol. 48 (1), 77–84. doi: 10.1046/j.1365-2958.2003.03425.x
Schwarz, S., Kehrenberg, C., Doublet, Benoît, Cloeckaert, A. (2004). Molecular basis of bacterial resistance to chloramphenicol and florfenicol. FEMS Microbiol. Rev. 28 (5), 519–542. doi: 10.1016/j.femsre.2004.04.001
Sharma, A., Kumar, A., Rashid, M., Amnekar, R. V., Gupta, S., Kaur, J. (2022). A phagosomally expressed gene, rv0428c, of mycobacterium tuberculosis demonstrates acetyl transferase activity and plays a protective role under stress conditions. Protein J. 41 (2), 260–273. doi: 10.1007/s10930-022-10044-x
Sheng, X., Cristea, I. M. (2021). The antiviral sirtuin 3 bridges protein acetylation to mitochondrial integrity and metabolism during human cytomegalovirus infection. PloS Pathog. 17 (4), e1009506. doi: 10.1371/journal.ppat.1009506
Shi, S., Ehrt, S. (2006). Dihydrolipoamide acyltransferase is critical for mycobacterium tuberculosis pathogenesis. Infect. Immun. 74 (1), 56–63. doi: 10.1128/IAI.74.1.56-63.2006
Shvedunova, M., Akhtar, A. (2022). Modulation of cellular processes by histone and non-histone protein acetylation. Nat. Rev. Mol. Cell Biol. 23 (5), 329–349. doi: 10.1038/s41580-021-00441-y
Silva, Márcia S N, Senna, S. G., Ribeiro, M. O., Valim, Andréia R. M., Telles, M. A., Kritski, A., et al. (2003). Mutations in katG, inhA, and ahpC genes of Brazilian isoniazid-resistant isolates of mycobacterium tuberculosis. J. Clin. Microbiol. 41 (9), 4471–4474. doi: 10.1128/JCM.41.9.4471-4474.2003
Singh, K. K., Athira, P. J., Bhardwaj, N., Singh, D. P., Watson, U., Saini, D. K. (2020). M. tuberculosisAcetylation of response regulator protein MtrA in regulates its repressor activity. Front. Microbiol. 11 (undefined). doi: 10.3389/fmicb.2020.516315
Singh, A., Zhao, X., Drlica, K. (2022). Fluoroquinolone heteroresistance, antimicrobial tolerance, and lethality enhancement. Front. Cell Infect. Microbiol. 12 (undefined). doi: 10.3389/fcimb.2022.938032
Son, S. M., Park, So J, Stamatakou, E., Vicinanza, M., Menzies, F. M., Rubinsztein, D. C. (2020). Leucine regulates autophagy via acetylation of the mTORC1 component raptor. Nat. Commun. 11 (1), 3148. doi: 10.1038/s41467-020-16886-2
Song, Z.-M., Lin, H., Yi, X.-M., Guo, W., Hu, M.-M., Shu, H.-B. (2020). KAT5 acetylates cGAS to promote innate immune response to DNA virus. Proc. Natl. Acad. Sci. U S A. 117 (35), 21568–21575. doi: 10.1073/pnas.1922330117
Soppa, Jörg (2010). Protein acetylation in archaea, bacteria, and eukaryotes. Archaea 2010 (undefined). doi: 10.1155/2010/820681
Sterling, T. R., Njie, G., Zenner, D., Cohn, D. L., Reves, R., Ahmed, A., et al. (2020). Guidelines for the treatment of latent tuberculosis infection: recommendations from the national tuberculosis controllers association and CDC 2020. MMWR Recomm Rep. 69 (1), 1–11. doi: 10.15585/mmwr.rr6901a1
Suarez-Kurtz, G., Fuchshuber-Moraes, M., Struchiner, C. J., Parra, E. J. (2016). Single nucleotide polymorphism coverage and inference of n-acetyltransferase-2 acetylator phenotypes in wordwide population groups. Pharmacogenet. Genomics 26 (8), 363–369. doi: 10.1097/FPC.0000000000000225
Suvichapanich, S., Fukunaga, K., Zahroh, H., Mushiroda, T., Mahasirimongkol, S., Toyo-Oka, L., et al. (2018). NAT2 ultra-slow acetylator and risk of anti-tuberculosis drug-induced liver injury: a genotype-based meta-analysis. Pharmacogenet. Genomics 28 (7), 167–176. doi: 10.1097/FPC.0000000000000339
Tamman, H., Ainelo, A., Ainsaar, K., Hõrak, R. (2014). A moderate toxin, GraT, modulates growth rate and stress tolerance of pseudomonas putida. J. Bacteriol 196 (1), 157–169. doi: 10.1128/JB.00851-13
Tarancón, R., Domínguez-Andrés, J., Uranga, S., Ferreira, AnaísaV, Groh, L. A., Domenech, M., et al. (2020). New live attenuated tuberculosis vaccine MTBVAC induces trained immunity and confers protection against experimental lethal pneumonia. PloS Pathog. 16 (4), e1008404. doi: 10.1371/journal.ppat.1008404
Timmermann, S., Lehrmann, H., Polesskaya, A., Harel-Bellan, A. (2001). Histone acetylation and disease. Cell Mol. Life Sci. 58 (null), 728–736. doi: 10.1007/pl00000896
Tomasi, F. G., Hall, A. M. J., Schweber, J. T. P., Dulberger, C. L., Kerry., M., Liu, Q., et al. (2022). A tRNA-acetylating toxin and detoxifying enzyme in mycobacterium tuberculosis. Microbiol. Spectr. 10 (3), e0058022. doi: 10.1128/spectrum.00580-22
Van der Geize, R., Yam, K., Heuser, T., Wilbrink, M. H., Hara, H., Anderton, M. C., et al. (2007). A gene cluster encoding cholesterol catabolism in a soil actinomycete provides insight into mycobacterium tuberculosis survival in macrophages. Proc. Natl. Acad. Sci. U.S.A. 104 (6), 1947–1952. doi: 10.1073/pnas.0605728104
Verma, R., Patil, S., Zhang, N., Moreira, F. M.F., Vitorio, M. T., da Santos, A. S., et al. (2021). NAT2A rapid pharmacogenomic assay to detect polymorphisms and guide isoniazid dosing for tuberculosis treatment. Am. J. Respir. Crit. Care Med. 204 (11), 1317–1326. doi: 10.1164/rccm.202103-0564OC
Vetting, M. W., Errey, J. C., Blanchard, J. S. (2008). Rv0802c from mycobacterium tuberculosis: the first structure of a succinyltransferase with the GNAT fold. Acta Crystallogr. Sect F Struct. Biol. Cryst Commun. 64 (null), 978–985. doi: 10.1107/S1744309108031679
Wan, W., You, Z., Xu, Y., Zhou, Li, Guan, Z., Peng, C., et al. (2017). mTORC1 phosphorylates acetyltransferase p300 to regulate autophagy and lipogenesis. Mol. Cell. 68 (2), 323–335.e6. doi: 10.1016/j.molcel.2017.09.020
Wang, Y., Curry, H. M., Zwilling, B. S., Lafuse, W. P. (2005). Mycobacteria inhibition of IFN-gamma induced HLA-DR gene expression by up-regulating histone deacetylation at the promoter region in human THP-1 monocytic cells. J. Immunol. 174 (9), 5687–5694. doi: 10.4049/jimmunol.174.9.5687
Wang, Y., Jing, W., Liu, J., Liu, M. (2022). Global trends, regional differences and age distribution for the incidence of HIV and tuberculosis co-infection from 1990 to 2019: results from the global burden of disease study 2019. Infect. Dis. (Lond). 54 (11), 773–783. doi: 10.1080/23744235.2022.2092647
Wang, D., Xu, C., Yang, W., Chen, J., Ou, Y., Guan, Y., et al. (2022). E3 ligase RNF167 and deubiquitinase STAMBPL1 modulate mTOR and cancer progression. Mol. Cell. 82 (4), 770–784.e9. doi: 10.1016/j.molcel.2022.01.002
WHO (2022) 2022, GLobal tuberculosis report. Available at: https://www.who.int/teams/global-tuberculosis-programme/tb-reports.
Wu, S., Ye, Z., Liu, X., Zhao, Y., Xia, Y., Steiner, A., et al. (2010). Salmonella typhimurium infection increases p53 acetylation in intestinal epithelial cells. Am. J. Physiol. Gastrointest. Liver Physiol. 298 (5), G784–G794. doi: 10.1152/ajpgi.00526.2009
Xie, L., Wang, X., Zeng, J., Zhou, M., Duan, X., Li, Q., et al. (2015). Proteome-wide lysine acetylation profiling of the human pathogen mycobacterium tuberculosis. Int. J. Biochem. Cell Biol. 59 (undefined), 193–202. doi: 10.1016/j.biocel.2014.11.010
Xie, L., Yang, W., Fan, X., Xie, J. (2019). Comprehensive analysis of protein acetyltransferases of human pathogen mycobacterium tuberculosis. Biosci. Rep. 39 (12). doi: 10.1042/BSR20191661
Xu, H., Hegde, S. S., Blanchard, J. S. (2011). Reversible acetylation and inactivation of mycobacterium tuberculosis acetyl-CoA synthetase is dependent on cAMP. Biochemistry 50 (26), 5883–5892. doi: 10.1021/bi200156t
Yang, H., Chen, J., Chen, Y., Jiang, Y., Ge, B., Hong, L. (2021). Sirt1 activation negatively regulates overt apoptosis in mtb-infected macrophage through bax. Int. Immunopharmacol. 91 (undefined), 107283. doi: 10.1016/j.intimp.2020.107283
Yang, H., Sha, W., Liu, Z., Tang, T., Liu, H., Qin, L., et al. (2018). Lysine acetylation of DosR regulates the hypoxia response of mycobacterium tuberculosis. Emerg. Microbes Infect. 7 (1), 34. doi: 10.1038/s41426-018-0032-2
Yang, H., Wang, F., Guo, X., Liu, F., Liu, Z., Wu, X., et al. (2021). Interception of host fatty acid metabolism by mycobacteria under hypoxia to suppress anti-TB immunity. Cell Discovery 7 (1), 90. doi: 10.1038/s41421-021-00301-1
Yang, L., Wang, L., Tang, Q., Liu, Y., Meng, C., Sun, S., et al. (2022). Hsa_circ_0093884 bound to RNA-binding protein RPS3 ameliorates hepatocyte inflammation in anti-tuberculosis drug-induced liver injury by competitively activating SIRT1. Int. Immunopharmacol. 110 (undefined), 109018. doi: 10.1016/j.intimp.2022.109018
Yang, X., Wu, L., Ran, Y., Xu, Ao, Zhang, B., Yang, X., et al. (2017). Crystal structure of l-glutamate n-acetyltransferase ArgA from mycobacterium tuberculosis. Biochim. Biophys. Acta Proteins Proteom 1865 (12), 1800–1807. doi: 10.1016/j.bbapap.2017.09.009
Yuan, J., Pu, M., Zhang, Z., Lou, Z. (2009). Histone H3-K56 acetylation is important for genomic stability in mammals. Cell Cycle 8 (11), 1747–1753. doi: 10.4161/cc.8.11.8620
Yuliwulandari, R., Susilowati, R. W., Wicaksono, B. D., Viyati, K., Prayuni, K., Razari, I., et al. (2016). NAT2 variants are associated with drug-induced liver injury caused by anti-tuberculosis drugs in Indonesian patients with tuberculosis. J. Hum. Genet. 61 (6), 533–537. doi: 10.1038/jhg.2016.10
Zahra, M. A., Kandeel, M., Aldossary, S. A., Al-Taher, A. (2020). Study on genotyping polymorphism and sequencing of n-acetyltransferase 2 (NAT2) among Al-ahsa population. BioMed. Res. Int. 2020 (undefined), 8765347. doi: 10.1155/2020/8765347
Zaunbrecher, M.A., Sikes, R.D., Metchock, B., Shinnick, T. M., Posey, J. E. (2009). Overexpression of the chromosomally encoded aminoglycoside acetyltransferase eis confers kanamycin resistance in mycobacterium tuberculosis. Proc. Natl. Acad. Sci. U S A. 106 (47), 20004–20009. doi: 10.1073/pnas.0907925106
Zeitoun, H., Bahey-El-Din, M., Kassem, M. A., Aboushleib, H. M. (2017). Mycothiol acetyltransferase (Rv0819) of mycobacterium tuberculosis is a potential biomarker for direct diagnosis of tuberculosis using patient serum specimens. Lett. Appl. Microbiol. 65 (6), 504–511. doi: 10.1111/lam.12801
Zhang, T., Li, Q., Dong, B., Liang, X., Jia, M., Bai, J., et al. (2021). CYPs, VKORC1, NAT2, DPYDGenetic polymorphism of drug metabolic gene and of five ethnic minorities in heilongjiang province, northeast China. Pharmgenomics Pers. Med. 14 (undefined), 1537–1547. doi: 10.2147/PGPM.S339854
Zhao, S., Xu, W., Jiang, W., Yu, W., Lin, Y., Zhang, T., et al. (2010). Regulation of cellular metabolism by protein lysine acetylation. Science 327 (5968), 1000–1004. doi: 10.1126/science.1179689
Zhu, C., Cai, Yi, Zhu, J., Zhang, L., Xing, A., Pan, L., et al. (2019). Histone deacetylase inhibitors impair the host immune response against mycobacterium tuberculosis infection. Tuberculosis 118 (undefined), 101861. doi: 10.1016/j.tube.2019.101861
Zhou, C., Li, Y., Huang, Y., Liang, D., Cui, Z., Huang, L., et al. (2023). Evaluation of the diagnostic efficacy of EC-test for latent tuberculosis infection in ambulatory people living with HIV. AIDS. doi: 10.1097/QAD.0000000000003573
Keywords: Mycobacterium tuberculosis, TB, post-translational modification, acetylation, N-acetyltransferase acetylation
Citation: Huang Y, Zhu C, Pan L and Zhang Z (2023) The role of Mycobacterium tuberculosis acetyltransferase and protein acetylation modifications in tuberculosis. Front. Cell. Infect. Microbiol. 13:1218583. doi: 10.3389/fcimb.2023.1218583
Received: 07 May 2023; Accepted: 29 June 2023;
Published: 25 July 2023.
Edited by:
Quanxin Long, Chongqing Medical University, ChinaCopyright © 2023 Huang, Zhu, Pan and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Liping Pan, cGFubGlwaW5nMjAwNkAxNjMuY29t; Zongde Zhang, enpkNDE3QDE2My5jb20=
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.