 Jansller Luiz Genova1*
Jansller Luiz Genova1* Paulo Evaristo Rupolo2Liliana Bury de Azevedo2Daniela Henz2Silvana Teixeira Carvalho2Marcos Kipper3Giovana de Arruda Castelo Gonçalves4Hellen Lazarino Oliveira Vilela1Tiago Junior Pasquetti4Newton Tavares Escocard de Oliveira2Andrei Roberto Manelli Dietrich2Paulo Levi de Oliveira Carvalho2
Paulo Evaristo Rupolo2Liliana Bury de Azevedo2Daniela Henz2Silvana Teixeira Carvalho2Marcos Kipper3Giovana de Arruda Castelo Gonçalves4Hellen Lazarino Oliveira Vilela1Tiago Junior Pasquetti4Newton Tavares Escocard de Oliveira2Andrei Roberto Manelli Dietrich2Paulo Levi de Oliveira Carvalho2- 1Animal Science Department, Universidade Federal de Viçosa, Viçosa, MG, Brazil
- 2Animal Science Department, Universidade Estadual do Oeste do Paraná, Marechal Cândido Rondon, PR, Brazil
- 3Elanco Animal Health Incorporated Company, São Paulo, SP, Brazil
- 4Animal Science Department, Universidade Estadual de Mato Grosso do Sul, Aquidauana, MS, Brazil
This study aimed to assess the effects of β-mannanase supplementation in metabolizable energy (ME)-reduced diets containing xylanase-phytase on performance, fecal score, blood biochemical and immunological profile, apparent total tract digestibility (ATTD), digesta passage rate, fecal microbiome, carcass traits and meat quality in finisher pigs (n = 40 entire male hybrid, 26.0 ± 0.9 kg) randomly assigned to 1 of 4 dietary treatments: a control diet containing isolated phytase and xylanase valued at 40 kcal of ME/kg (CD0), CD0 + β-mannanase (0.3 g/kg valued at 30 kcal of ME/kg) (CD70), CD0 + β-mannanase (0.3 g/kg valued at 45 kcal of ME/kg) (CD85), and CD0 + β-mannanase (0.3 g/kg valued at 60 kcal of ME/kg) (CD100), with 10 pen replicates. Pigs fed CD0 diet showed (P = 0.002) greater ADFI. However, pigs fed CD0 diet showed (P = 0.009) lower G:F than those provided CD70 or CD85 diets. A greater (P < 0.001) superoxide dismutase concentration was observed in pigs fed CD70 diet. Pigs fed CD85 diet showed (P = 0.002) greater digestible protein than pigs fed CD0 or CD100 diets. Pigs fed CD70 diet showed an increase of 11.3% in digestible protein than those fed CD0 diet. In addition, greater (P < 0.001) digestible energy was observed in pigs fed CD85 diet. Pigs fed CD0 or CD100 diets showed greater (P < 0.05) Firmicutes:Bacteroidota ratio than those fed CD85 diet. The Muribaculaceae was more abundant (P = 0.030) in pigs fed CD70 diet than in those fed CD0 diet. The Prevotella was more abundant (P = 0.045) in pigs fed CD85 diet than in those fed CD100 diet. In conclusion, β-mannanase supplementation in diets containing xylanase-phytase allows reducing 85 kcal of ME/kg because it improves gain to feed ratio, energy and protein usage, and backfat thickness without metabolic and intestinal ecosystem disorders in finisher pigs.
1. Introduction
Plant-based ingredients widely used in the diets of pigs possess significant amounts of antinutritional factors (1, 2). These antinutritional substances, such as β-mannans (1, 3–5), phytate molecules (3, 6), and xylans (4, 6), are not digested by endogenous enzymes, and compromise the use of nutrients and energy metabolism in non-ruminant animals (5).
Based on this, dietary supplementation of β-mannanase has been attributed to the hydrolysis of β-mannans reducing the immune response capacity induced by feeding (2), and energy expenditure for immune system activation (5). This nutritional strategy also allows the use of phytase enzyme, known to improve the availability of phosphorus and calcium in diets containing phytate molecules (3), and providing additional energy and improving energy efficiency (7). In addition, the antinutritional effects of non-starch polysaccharides (NSP) provided by xylans highlight the importance of using the xylanase enzyme (1). Xylanase breakdowns the plant cell wall releasing nutrients within the cell and reduce digesta viscosity (6).
Diets supplemented with a blend of these enzymes may be of economic-environmental-nutritional interest. Indeed, β-mannanase has been previously reported to reduce feed to gain ratio and increase nutrient ATTD (5). Greater phosphorus and lower neutral detergent fiber digestibility were reported when combined xylanase-phytase were supplemented in the diet of grower pigs (6). Greater blood glucose concentration and lower backfat thickness were observed in finisher pigs fed β-mannanase-xylanase (8); however, no effect on ATTD in grower pigs provided diets containing phytase and β-mannanase were observed (3).
To date, no studies have been conducted to assess the effects of the dietary association of these enzymes on the fecal microbiome, total digesta passage rate, and fecal consistency score in finisher pigs. Here, a study was conducted based on the hypothesis that β-mannanase supplementation in ME-reduced diets improves ATTD, intestinal digesta viscosity, and intestinal microbiome, supporting growth performance and health compared to the diet without β-mannanase supplementation.
Therefore, this study assessed the effects of β-mannanase associated with xylanase-phytase on growth performance, fecal score, biochemical and immunological blood profile, ATTD, total digesta passage rate, fecal microbiome, carcass traits, and meat quality in finisher pigs fed ME-reduced diets.
2. Materials and methods
2.1. Animals, experimental design, housing, and dietary treatments
A total of 40 entire male hybrid pigs (26.0 ± 0.9 kg BW) from a commercial line (Landrace × Large White) were used. Pigs were allotted to 1 of 4 dietary treatments in a randomized complete block design with 10 pen replicates and 1 animal per pen as the experimental unit. Blocks were based on the initial BW of pigs.
At the beginning of the experiment, animals were weighed and identified with numbered ear tags. Pigs were housed in a masonry facility with 2 rows (with a central aisle) of concrete floor pens (6.3 m2). All pens were equipped with a semiautomatic front feeder and a nipple waterer.
Room temperature and relative humidity were recorded by a data logger (Hygro-Thermometer, model RT811) located in the middle of the experimental facility. Temperature and ventilation were controlled via side curtains and trees on both sides of the facility. Room temperature and relative humidity averaged 20.4 ± 6.6°C and 63.6 ± 19.3%, respectively.
The experimental period lasted 52 days and was divided into 2 phases: finisher I (d 0 to 22) and finisher II (d 22 to 52). Diets (Table 1) were formulated to meet the nutritional requirements of pigs in each phase (9) and offered as mash, and ad libitum. All diets were corn- and soybean meal-based with industrial amino acids, and were isonutritional with variations only in soybean oil and inert content.
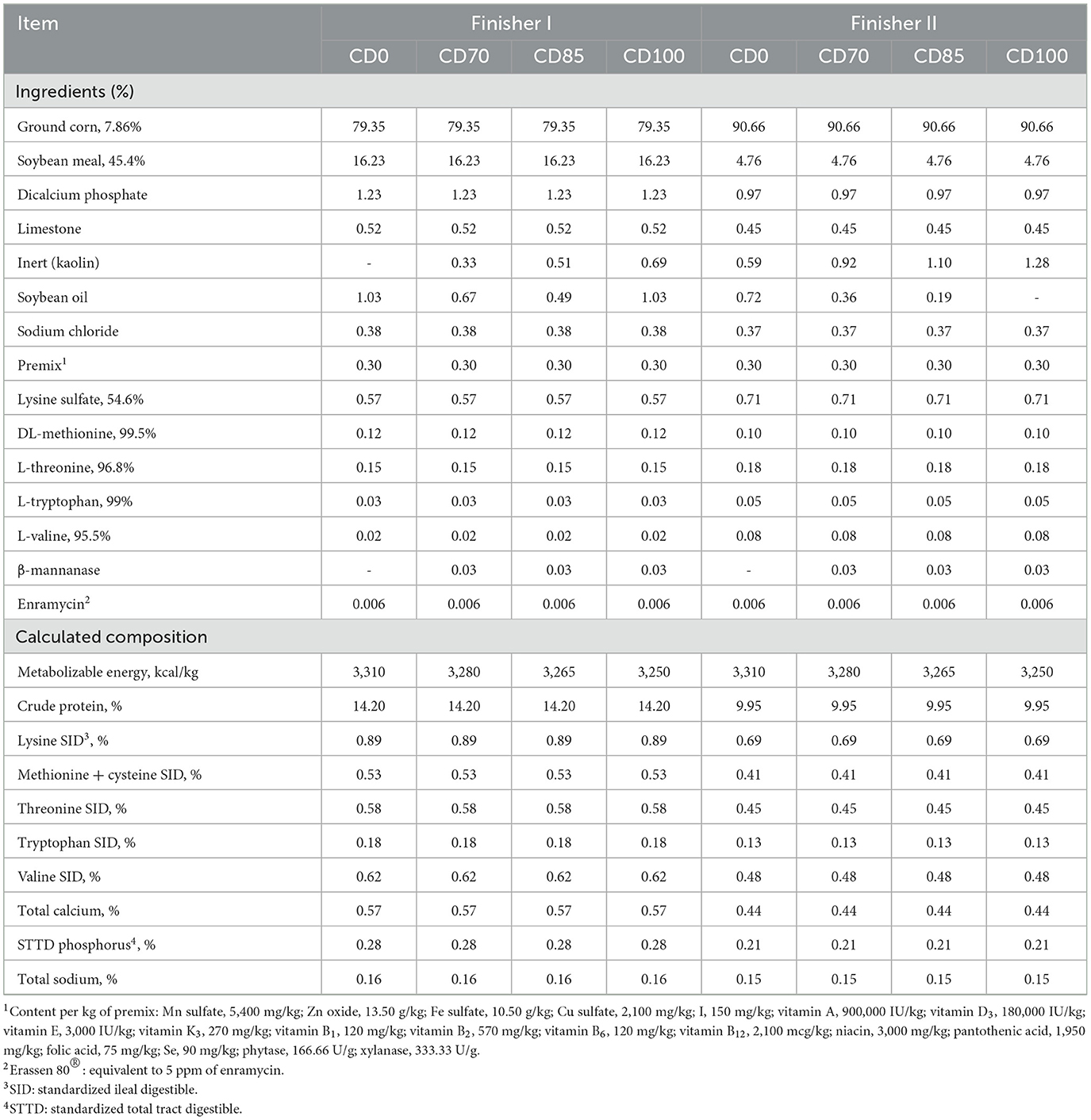
Table 1. Composition of diets provided to finisher pigs (as-fed basis).
Dietary treatments were: (1) a control diet containing isolated phytase and xylanase valued at 40 kcal of ME/kg (CD0), (2) CD0 + β-mannanase (0.3 g/kg valued at 30 kcal of ME/kg) (CD70), (3) CD0 + β-mannanase (0.3 g/kg valued at 45 kcal of ME/kg) (CD85), and (4) CD0 + β-mannanase (0.3 g/kg valued at 60 kcal of ME/kg) (CD100).
2.2. Traits of the tested enzymes
Xylanase (Sunhy Biology Co., Ltd, Wuhan, HB, China; registration no. PR-08978 03462) was a product obtained from Trichoderma longibrachiatum. A U of xylanase is the amount of enzyme that releases 1 micromol of reducing sugar from a xylan solution (5 mg/mL) at 37°C and pH 5.5. Phytase (Sunhy Biology Co., Ltd, Wuhan, HB, China; registration no. PR 000267-4.000005) was a product from Aspergillus niger with the activity of 1,000 U/g of dry solid at 37°C and pH 5.5. β-mannanase (Elanco Animal Health, Inc., São Paulo, SP, Brazil; registration no. SP-59122 30011, HemicellTM HT) was obtained from Paenibacillus lentus. A U of β-mannanase is the amount of enzyme that releases 0.72 mcg of reducing sugars (equivalent to D-mannose) per min from goma locust (mannans concentration of 88%) at 40°C and pH 7.5.
2.3. Growth performance and fecal consistency score
Animals had free access to diets and water throughout the experiment. Offered diets and leftovers were recorded daily using a digital scale (model UL-50, DIGI-TRON, Curitiba, PR, Brazil) to determine the average daily feed intake (ADFI, g/day). Pigs were weighed at the beginning and end of each experimental phase using a 2 bars digital scale (model ULB-3000, IWM bivolt, Curitiba, PR, Brazil). Initial BW (IBW, kg), final BW (FBW, kg), average daily gain (ADG, g/day), and gain to feed ratio (G:F, g:g) were determined.
Fecal consistency score was assessed via partial feces collection at the end of finisher phases. Before feces collection, all pens (08:00) were cleaned and animals were monitored for a 12-h period. During this period, fecal samples were collected right after defecation, except for the lower part that was in contact with the floor. Feces were packed in plastic bags and kept in a thermal box (4°C) until the end of the collection period. Then, the samples were homogenized and 2 subsamples of 110 g each were weighed in a scale (model M4102, Bel engineering, Monza, Italy) and dried in a forced-air oven (Tecnalbrand, SF-325 NM model; Piracicaba, SP, Brazil) at 55°C for 72 h for dry matter determination (10). Values were tabulated and classified according to fecal consistency, following the adapted methodology (11).
2.4. Blood sampling and blood profile analysis
Animals fasted for 8 h at the end of the finisher II phase. Blood samples (≅10 mL) were withdrawn from the anterior cranial vena cava using 1.2 × 40 mm needles and 20 mL syringes. Samples were transferred to 1 of 3 tubes containing potassium fluoride, EDTA, or no anticoagulant. All tubes were previously identified, placed into a thermal box (4°C), and sent to the blood laboratory for further analysis. Plasma or serum was isolated from blood by centrifugation (Centrilab analog centrifuge, model 80-2B) at 3,000 g for 10 min. Then, ≅3 mL of plasma or serum were transferred to previously identified polyethylene tubes (Eppendorf-type) and stored at −20°C until analysis of urea (enzymatic-colorimetric method), glucose (enzymatic-colorimetric method), total cholesterol (enzymatic-colorimetric method), total protein (enzymatic-biuret method), and albumin (bromocresol green colorimetric) of 10 animals per treatment.
All analyses were performed in the blood laboratory of Unioeste via spectrophotometry with the aid of an analyzer (Bel SPECTRO S05) using commercial kits (Gold Analisa Diagnóstica—Belo Horizonte, MG, Brazil).
Globulin was calculated as the difference between total protein and plasma albumin. Blood samples from 8 animals per treatment were stored at −80°C and sent to a private laboratory (Curitiba, PR, Brazil) where serum concentrations of superoxide dismutase (SOD), glutathione S-transferase, and immunoglobulins M were determined via the immunoturbidimetry method.
2.5. Apparent total tract digestibility and total digesta passage rate
The insoluble acid ash marker (IAA, celite®) was added to the diets (10 g/kg diet) at the end of the finisher II phase to assess ATTD using partial feces collection (indirect method) (12). The diets containing the marker were homogenized in a vertical mixer for 10 min. These diets were fed to pigs for 3 days before feces collection. On the fourth day, partial feces collection was performed following the adapted methodology (13). The beginning and end of the diet supply and the feed intake per pen were recorded. Feces were collected for 12 h on the last day of the supply of the diets containing the marker. During collection, feces were packed in polyethylene plastic bags (previously identified) and kept in thermal boxes containing ice (4°C). After this period, the feces were stored at −18°C for further analysis.
Afterward, the samples were thawed and homogenized. Two subsamples (110 g each) were weighed in a scale (bel engineering, model M4102, Monza, Italy) and dried in a forced-air oven (Tecnalbrand, SF-325 NM model; Piracicaba, SP, Brazil) at 55°C for 72 h, according to the methodologies (10). Then, the samples were ground in a micro-powder grinding mill (R-TE-350; Tecnal Scientific Equipment, Piracicaba, SP, Brazil) and stored in plastic containers previously identified for laboratory analyses.
Insoluble acid ash marker was analyzed via hydrochloric acid (4N) digestion, following the adapted procedures (13). The chemical composition of diets and feces samples was determined according to the methodologies described (10). The gross energy in diets and feces samples was determined in a bomb calorimeter (IKA®, model C200, USA).
Based on the results of laboratory analyses, the recovery percentage of IAA and the ATTD coefficients of dry matter (ADCDM), organic matter (ADCOM), crude protein (ADCCP), and gross energy (ADCGE) were calculated. Digestible nutrients and energy were determined as a percentage of digestible dry matter (DDM), digestible organic matter (DOM), digestible protein (DP), and kcal/kg of digestible energy (DE), according to the established equations (12).
The total digesta passage rate was assessed via fecal marker excretion at the end of the finisher phases, according to the adapted methodology (14). Before supplying the diets containing the marker, a quantified portion of the diet was weighed with 1.5% of the marker (iron oxide) and homogenized to ensure the intake in a single meal. One h before the evaluation, all diet was removed from the feeder of pens and placed in identified containers to be returned to the respective feeder afterward. Diets containing the marker were supplied following the same sequence used to withdraw diets without marker. The supplying time and the time when animals consumed all the marked diet (h 0) were recorded per pen. Pens were monitored to identify the defection of marked feces. The defecation time was recorded accordingly. The total digesta passage rate was calculated based on the time (in min) between the marked diet consumption and the excretion of marked feces.
2.6. Fecal microbiome
At the end of finisher II phase, rectum feces samples from 6 pigs per treatment were collected and immediately placed in sterile Eppendorf-type tubes using swabs. Right after collection, the samples were stored at −80°C until analysis.
A commercial kit (ZR Fecal DNA MiniPrep® from Zymo Research) was used to extract DNA from samples following the manufacturer's instructions. The integrity of the extracted DNA was assessed via 1% agarose gel electrophoresis. The extracted DNA was quantified via spectrophotometry at 260 nm.
A segment of approximately 460 bases of the hypervariable region V3-V4 of the ribosomal gene 16S rRNA was amplified using the universal primers described by the methodology. The PCR conditions were as follows: 95°C for 3 min, 25 cycles at 95°C for 30 s, 55°C for 30 s, and 72°C for 30 s, followed by a step at 72°C for 5 min. A metagenomics library was built from the amplified using a commercial kit (Nextera DNA Library Preparation Kit, Illumina®). The amplified were pooled and sequenced in Illumina's MiSeqTM sequencer® (15).
Readings were analyzed in the quantitative insights into microbial ecology (QIIME2) platform (16). The following procedures were performed: removal of low-quality sequences, filtration, chimera's removal, and taxonomic classification. Sequences were classified into bacterial genera via amplicon sequence variants (ASVs) identification, in this case, the homology between sequences when compared against a database. The 2019 edition (SILVA 138) of the SILVA ribosomal sequence database (17) was used to compare the sequences.
To generate the classification of bacterial communities via ASVs identification, 25,610 readings per sample were used. Thus, data were normalized and samples with different number of readings were not compared. The samples of identifiers 29,160 and 29,167 were removed due to the low number of readings (< 15,000). They were retrieved after the quality filtering steps.
2.7. Slaughter procedures, carcass traits, and meat quality
On day 52 of the experimental period, all animals (n = 10/treatment) were fasted for 12 h and then transported for 6 h (a total of 18 h of fasting) to a commercial abattoir (Medianeira, PR, Brazil) with federal certification. Pigs were slaughtered using carbon dioxide stunning, followed by exsanguination.
All analyses were performed and calculated according to the methodologies described (18). The quantitative carcass traits such as backfat thickness, muscle percentage in the carcass, lean meat percentage, and lean meat amount were measured in the slaughterhouse using a swine carcass typing pistol (model UltraFom 300, Carometec). The carcass weight was determined using a scale placed in the slaughter line. Then, hot carcass yield, meat yield, and amount of chilled meat were calculated.
Carcass length was measured after a cold shock in the cold chamber. Measurements were taken from the cranial edge of the atlas to the cranial edge of the aitch bone. A sample (≅30 cm) of the l. thoracis muscle was collected between the last thoracic vertebra and the first lumbar vertebra (caudal to cranial direction). Samples were immediately packed in the identified polyethylene plastic bags, placed in thermal boxes (4°C), and transported to the Animal Products Technology Laboratory (APTL) belonging to Unioeste.
Then, pH value in the l. thoracis muscle was measured using a portable pHmeter (model AK103, Asko produtos eletrônicos Ltda, São Leopoldo, RS, Brazil) in the area of the last rib 4 and 24 h post mortem. For the measurements taken 4 h post mortem, the carcasses were submitted to 180 min of cooler shock, as follows: first stage: from −18°C to −15°C; second stage: from −15°C to −12°C; and third stage: from −10°C to −8°C.
At the APTL, samples were refrigerated (≅2°C) for 24 h and then the backfat thickness and loin depth were measured using a digital pachymeter (MTX, stainless hardened). To determine the loin eye area (LEA) of the l. thoracis muscle, samples were scanned using a scanner printer (Officejet 4500 Desktop - G510a, HP, São Paulo, SP, Brazil). A black box was used to block the lighting and improve the image quality. Then, readings were performed using a Software (imageJ 1.53e - Java).
Meat color was assessed after muscle oxygenation via air exposure for 15 min. Color analyses were performed using a Minolta CR400 colorimeter device (Konica Minolta Holdings, inc. Tokyo, Japan) and the results were expressed using the CIELAB color system. Color parameters were measured as L* (luminosity), a* (red-green component), and b* (yellow-blue component), which represent the saturation (chroma or purity) and the tint (color or hue). With these results, the saturation of the l. thoracis muscle was calculated.
Marbling was determined using photographic standards and a 7-point scale (1 = traces of marbling and, 7 = excessive marbling). The subjective color analysis was performed using a 6-point scale (1 = light color and, 6 = trend to red).
Afterward, samples were boned and the l. thoracis muscle was cross-sectioned into four 2.5-cm subsamples. The subsamples were used to determine drip loss (DL), thaw loss (TL), cooking loss (CL), shear force (SF), and chemical analyses. Subsample 1 was used to assess DL. The remaining subsamples were packed in the identified polyethylene bags and stored at−18°C until analyses. The losses were expressed as the percentage of lost water in relation to the original sample weight. Cooking loss was performed sequentially in a grill (Britannia brand, multi grill 2). Shear force analysis was performed using 6 cores (1.5 cm) removed from subsample 2 (TL and CL sequentially) using a stainless-steel cylinder sampler. Subsequently, the cores were submitted to a TA.HD.plus texture meter (model Texture Analyser, Stable Micro Systems) equipped with a standard shear blade calibrated for force (15 g), deformation (20 mm), and speed (2.0 mm/s).
Subsample 3 was thawed in a refrigerator at a controlled temperature (4°C). Fat and connective tissue were withdrawn using a knife. Then, the subsamples were ground in a microprocessor and packed in the originally identified bags to determine moisture, ash, and crude protein. The ether extract was performed according to the AOCS methodology (Am 5-04, 2017) using an Ankom extractor (model XT15, NY, USA). Subsample 4 was kept frozen as a backup. The in vivo loin depth and backfat thickness were assessed in the lumbar area P2 in finisher II pigs using an Aloka ultrasound (Echo Camera model - SSD-500 vet, Tokyo, Japan).
2.8. Statistical procedures
A Student standardized residuals analysis was performed before one-way analysis of covariance (ANCOVA) and variance (ANOVA), in which values >3 standard deviations were considered outliers. The normality of experimental errors and the homogeneity of variance of errors among treatments were evaluated using Shapiro-Wilk and Levene tests, respectively. For antioxidant enzyme data, outliers were identified via ROUT test (Q = 1%) and the normality was assessed via D'Agostino-Pearson test. Data on growth performance were analyzed using the following model:
The effects of the factors in the model were described as: Yijk = average observation of the dependent variable in each plot, measured in the i-th class of treatment, in the j-th block, and the k-th replication; μ = overall mean effect; Ti = fixed effect of treatment classes, i = (1, 2, 3, and 4); bj = random effect of block, j = (1 and 2); β = regression coefficient of Y over X; Xijk = average observation of the covariate (initial BW) in each plot, measured in the i-th class of treatment, in the j-th block, and the k-th replication; = overall mean for covariate X; εijk = random error of the plot associated with level i, block j, and replication k. For other variables, the statistical model used was the one mentioned above, no covariate effect.
Treatment effect on dependent variables was verified via ANCOVA or ANOVA. Treatment significance was set at P < 0.10 when the power of the test was < 80%. Multiple comparisons among treatment means were performed according to the post hoc test of Tukey and t-Student at 5% and 10% of probability, respectively. All statistical analyses were performed using the procedures of the SAS University Edition (SAS Inst. Inc., Cary, NC, USA). All normally distributed data were reported as means and their pooled SEM.
For the fecal microbiome, the statistical comparison among the groups in the analyses of alpha diversity and the relative abundances of taxa among all experimental groups was performed via Wilcoxon non-parametric test at P < 0.05. Statistical analyses for beta diversity were performed through permutational multivariate analysis of variance (PERMANOVA) in the QIIME2 pipeline. A total of 10,000 permutations was used. Alpha diversity analyses were calculated using phyloseq (19) and microbiome (20) libraries.
3. Results
3.1. Growth performance and fecal consistency score
Pigs fed CD0 diet showed (P = 0.002) greater ADFI than pigs fed other dietary treatments (Table 2). However, pigs fed CD0 diet showed (P = 0.009) lower G:F than those provided CD70 or CD85 diets. Although no difference among dietary treatments was observed in the finisher II phase, pigs fed ME-reduced diets containing the enzymes combination-maintained growth performance. No dietary treatment effect on the fecal consistency score was observed in finisher pigs.
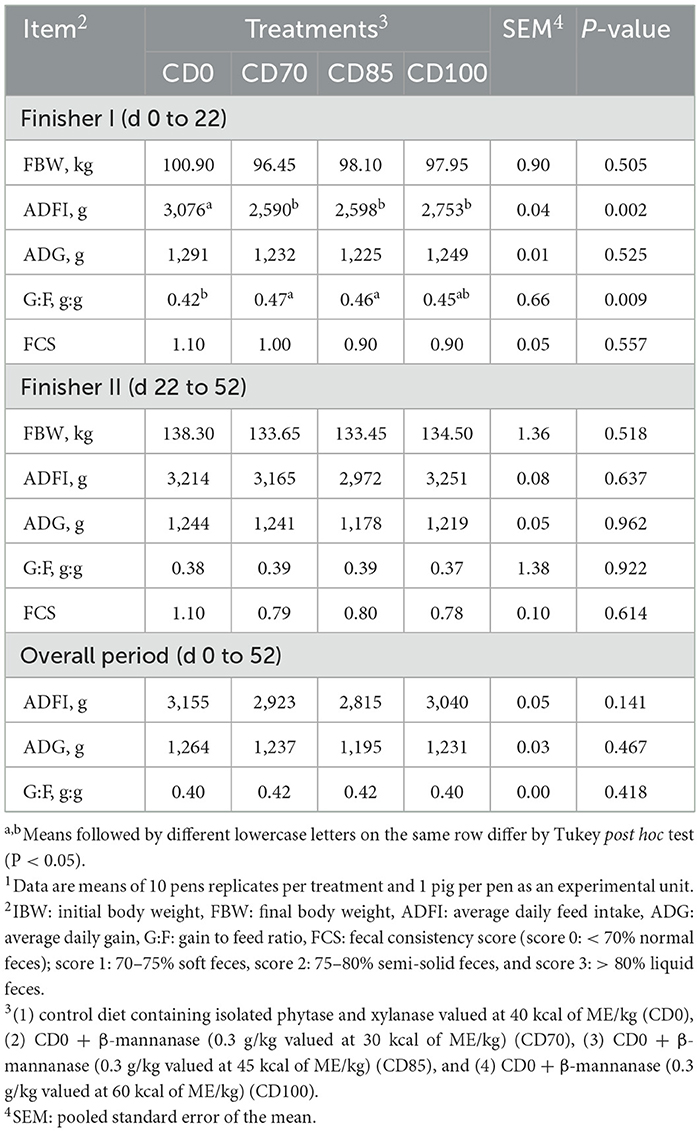
Table 2. Effect of β-mannanase supplementation in diets containing xylanase-phytase and reduced metabolizable energy on performance and fecal consistency score in finisher pigs1.
3.2. Blood biochemical and immune profile
A greater (P < 0.001) SOD concentration was observed in pigs fed CD70 diet compared to other dietary treatments (Table 3). No dietary treatment effect on the biochemical blood profile was observed in finisher pigs.
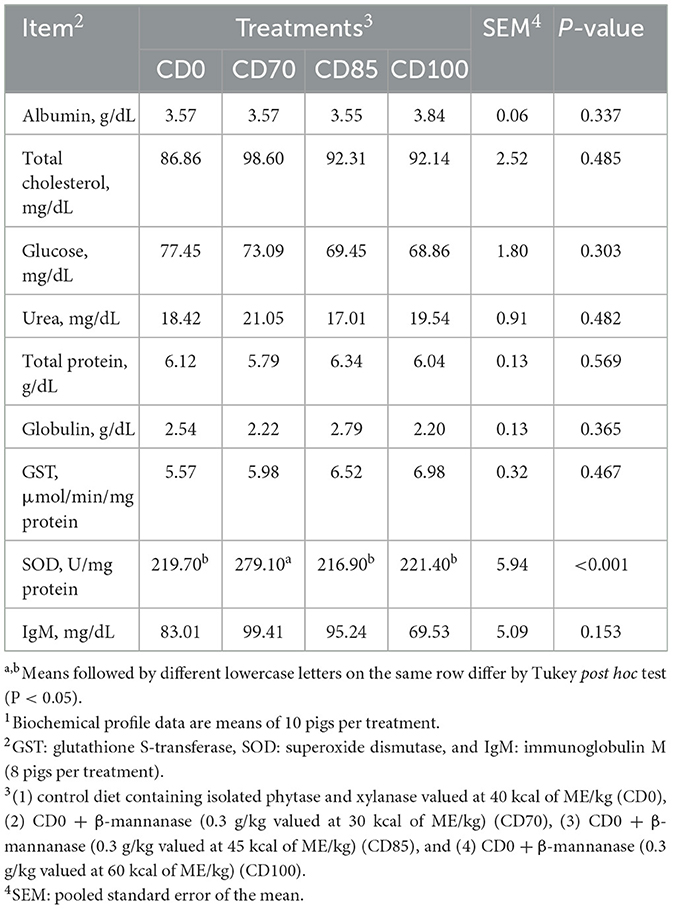
Table 3. Effect of β-mannanase supplementation in diets containing xylanase-phytase and reduced metabolizable energy on blood biochemical and immunological profile in finisher pigs on day 521.
3.3. Apparent total tract digestibility and total digesta passage rate
Pigs fed CD85 diet showed (P = 0.002) greater DP than pigs fed CD0 or CD100 diets. Pigs fed CD70 diet showed an increase of 11.3% in DP than those fed CD0 diet (Table 4). In addition, greater (P < 0.001) DE was observed in pigs fed CD85 diet compared to other dietary treatments. No effect of dietary treatments on the passage rate of total digesta was observed in finisher pigs.
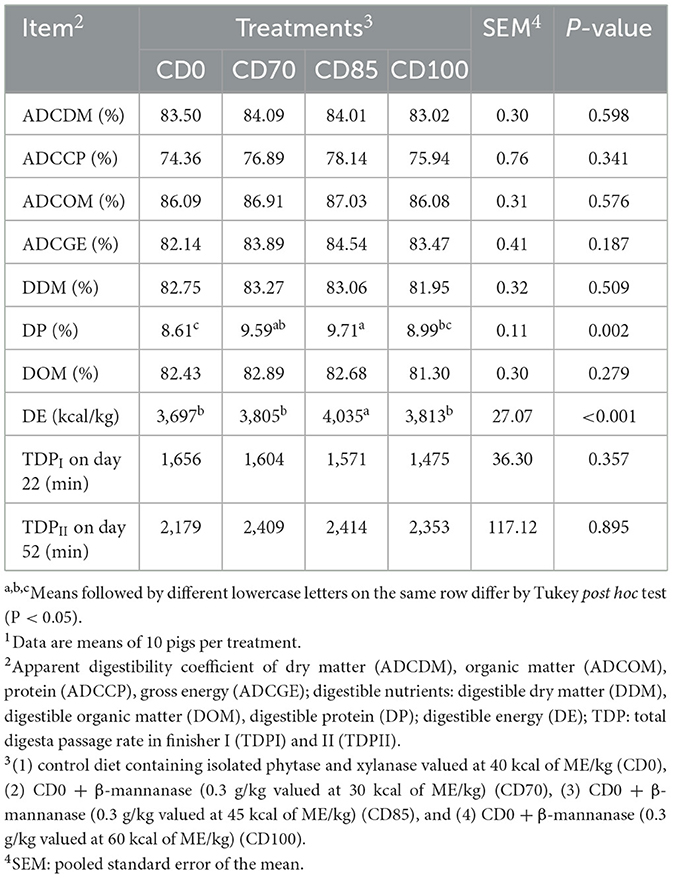
Table 4. Effect of β-mannanase supplementation in diets containing xylanase-phytase and reduced metabolizable energy on apparent total tract digestibility (dry matter basis) on day 52, and total digesta passage rate in finisher pigs on days 22 and 521.
3.4. Fecal microbiome
No difference among treatments was observed via the alpha diversity test (Shannon, Evenness Pielou, Simpson Index, Fisher, total number of observed OTUs, and Chao 1) in finisher pigs (Figure 1). Beta diversity was estimated via Bray-Curtis, Jaccard, UniFrac, and Weighted Unifrac parameters (Figure 2); however, no differences among dietary treatments were observed in finisher pigs.
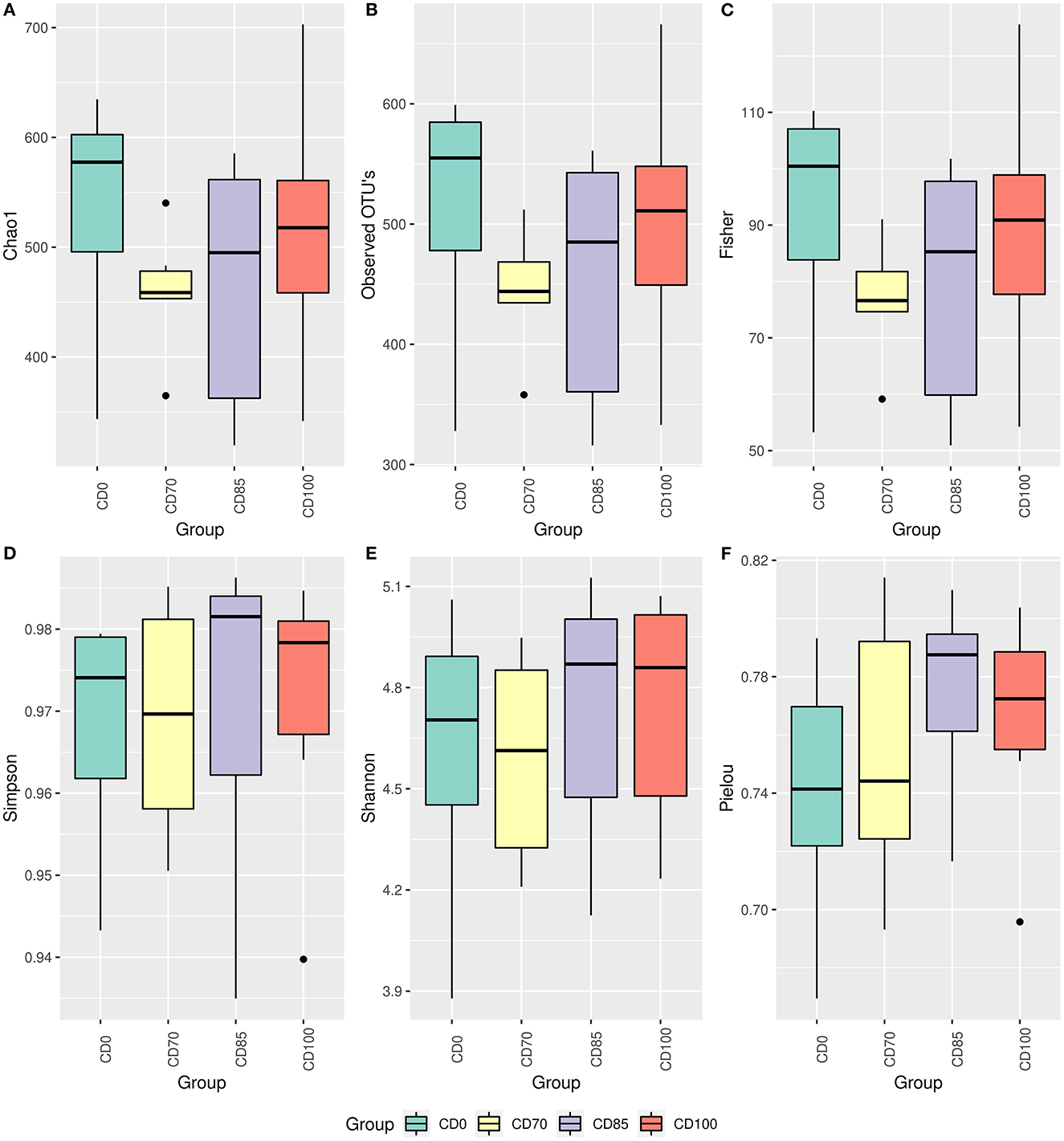
Figure 1. Alpha diversity estimated by parameters Chao1 (A), observed OTUs (B), Fisher (C), Simpson (D), Shannon (E), and Pielou (F) in finisher pigs fed 1 of 4 dietary treatments: a control diet containing isolated phytase and xylanase valued at 40 kcal of ME/kg (CD0), CD0 + β-mannanase (0.3 g/kg valued at 30 kcal of ME/kg) (CD70), CD0 + β-mannanase (0.3 g/kg valued at 45 kcal of ME/kg) (CD85), and CD0 + β-mannanase (0.3 g/kg valued at 60 kcal of ME/kg) (CD100). Data are averages of 6 pigs per dietary treatment.

Figure 2. Beta diversity estimated by parameters Bray-Curtis (A), Jaccard (B), Unifrac (C), and Weighted Unifrac (D) in finisher pigs fed 1 of 4 dietary treatments: a control diet containing isolated phytase and xylanase valued at 40 kcal of ME/kg (CD0), CD0 + β-mannanase (0.3 g/kg valued at 30 kcal of ME/kg) (CD70), CD0 + β-mannanase (0.3 g/kg valued at 45 kcal of ME/kg) (CD85), and CD0 + β-mannanase (0.3 g/kg valued at 60 kcal of ME/kg) (CD100). Data are averages of 6 pigs per dietary treatment.
The most abundant phyla we observed were Firmicutes, Bacteroidota (previously described as Bacteroidetes), Proteobacteria, and Spirochaetota (previously described as Spirochaetes) (Figure 3A). The classes Clostridia, Bacteroidia, Negativicutes, Bacilli, Gammaproteobacteria, and Spirochaetia showed the largest populations (Figure 3B). The most abundant orders were Bacteroidales, Oscillospirales, Veillonellales, Lachnospirales, Clostridiales, Acidaminococcales, Christensenellales, Enterobacterales, Lactobacillales, Treponematales, Selenomonadales, and Peptostreptococcales (Figure 3C).
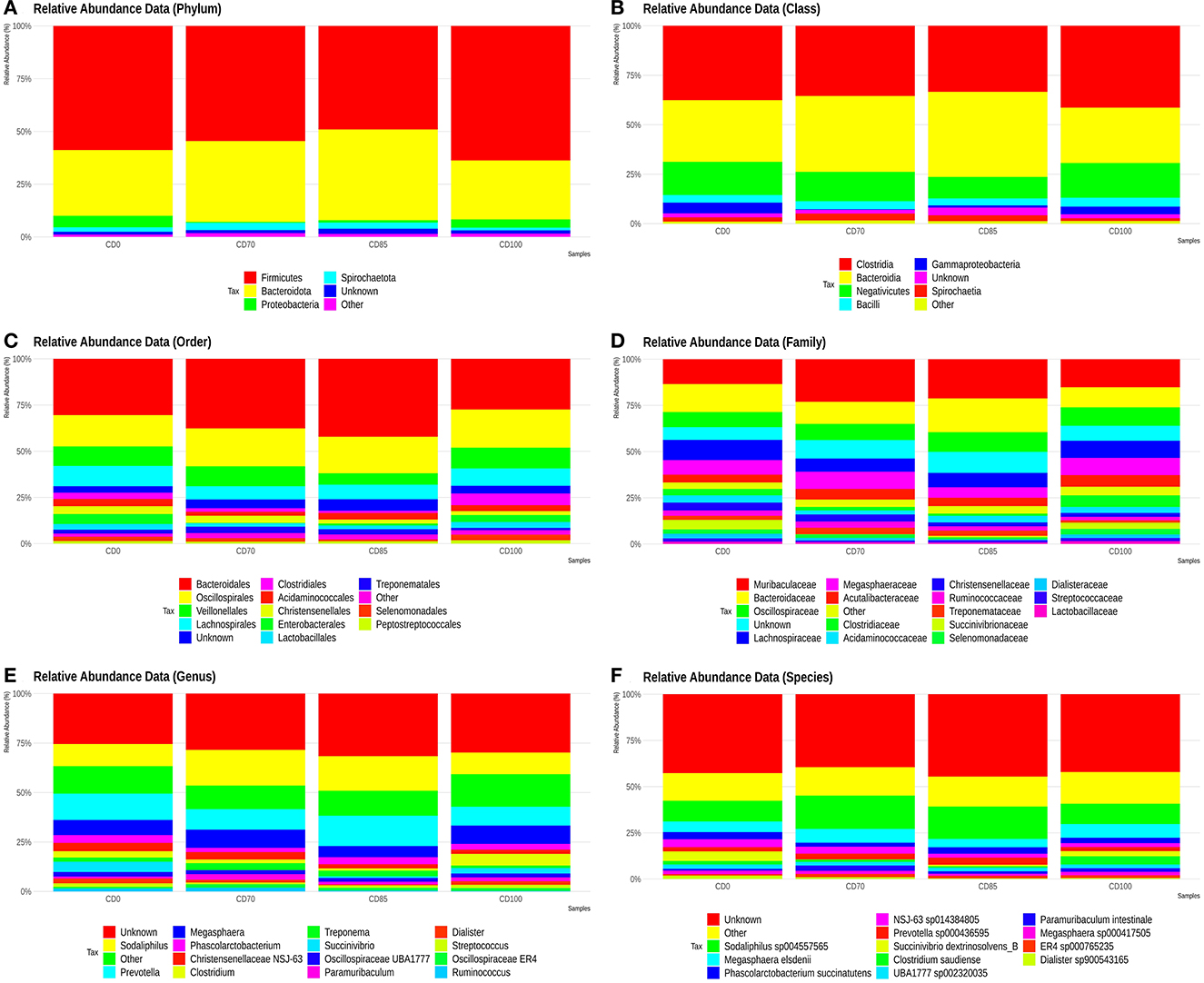
Figure 3. Relative abundance of phyla (A), classes (B), orders (C), families (D), genera (E), and species (F) presents in finisher pigs fed 1 of 4 dietary treatments: a control diet containing isolated phytase and xylanase valued at 40 kcal of ME/kg (CD0), CD0 + β-mannanase (0.3 g/kg valued at 30 kcal of ME/kg) (CD70), CD0 + β-mannanase (0.3 g/kg valued at 45 kcal of ME/kg) (CD85), and CD0 + β-mannanase (0.3 g/kg valued at 60 kcal of ME/kg) (CD100). Data are averages of 6 pigs per dietary treatment.
The most abundant families were Muribaculaceae, Bacteroidaceae, Oscillospiraceae, Lachnospiraceae, Megasphaeraceae, Acutalibacteraceae, Clostridiaceae, Acidaminococcaceae, Christensenellaceae, Ruminococcaceae, Treponemataceae, Succinivibriononaceae, Selenomonadaceae, Dialisteraceae, Streptococcaceaeaceae, and Lactobacillaceae (Figure 3D).
The most abundant genera were Sodaliphilus, Prevotella, Megasphaera, Phascolarctobacterium, Christensenellaceae NSJ-63, Clostridium, Treponema, Succinivibrio, Oscillospiraceae UBA1777, Paramuribaculum, Dialister, Streptococcus, Oscillospiraceae ER4, and Ruminococcus (Figure 3E).
The species Sodaliphilus sp004557565, Megasphaera elsdenii, Phascolarctobacterium succinatutens, NSJ-63 sp014384805, Prevotella sp000436595, Succinivibrio dextrinosolvens_B, Clostridium saudiense, UBA1777 sp002320035, Paramuribaculum intestinale, Megasphaera sp000417505, ER4 sp000765235, and Dialister sp900543165 showed the largest abundances (Figure 3F).
In addition, pigs fed CD0 diet showed (P = 0.049) greater Firmicutes:Bacteroidota ratio (FBR) than those provided with CD85 diet (Figure 4). However, pigs fed CD100 diet showed (P = 0.011) greater FBR than those fed CD85 diet. We analyzed only the taxon that showed different (P < 0.05) average relative abundance among dietary treatments. Therefore, the Muribaculaceae family was more abundant (P = 0.030) in pigs fed CD70 diet than in those fed CD0 diet (Figure 5). In addition, the Prevotella genus was more abundant (P = 0.045) in pigs fed CD85 diet than in those fed CD100 diet (Figure 6).
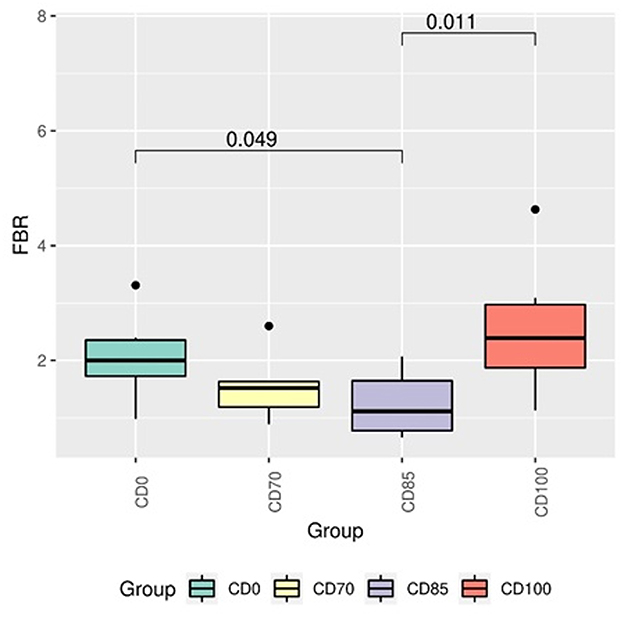
Figure 4. Firmicutes:Bacteroidetes ratio in finisher pigs fed 1 of 4 dietary treatments: a control diet containing isolated phytase and xylanase valued at 40 kcal of ME/kg (CD0), CD0 + β-mannanase (0.3 g/kg valued at 30 kcal of ME/kg) (CD70), CD0 + β-mannanase (0.3 g/kg valued at 45 kcal of ME/kg) (CD85), and CD0 + β-mannanase (0.3 g/kg valued at 60 kcal of ME/kg) (CD100). Data are averages of 6 pigs per dietary treatment. Means differed by Wilcoxon test (P < 0.05).
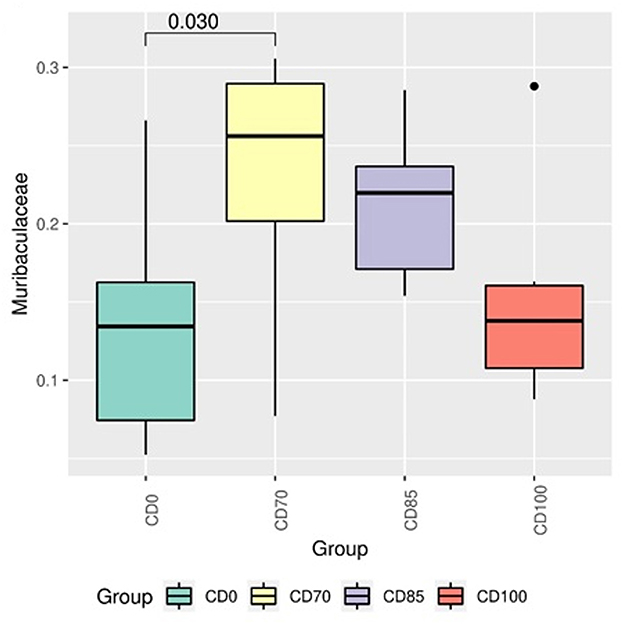
Figure 5. Differential abundance analysis of taxon of the Muribaculaceae family in finisher pigs fed 1 of 4 dietary treatments: a control diet containing isolated phytase and xylanase valued at 40 kcal of ME/kg (CD0), CD0 + β-mannanase (0.3 g/kg valued at 30 kcal of ME/kg) (CD70), CD0 + β-mannanase (0.3 g/kg valued at 45 kcal of ME/kg) (CD85), and CD0 + β-mannanase (0.3 g/kg valued at 60 kcal of ME/kg) (CD100). Data are averages of 6 pigs per dietary treatment. Means differed by Wilcoxon test (P < 0.05).
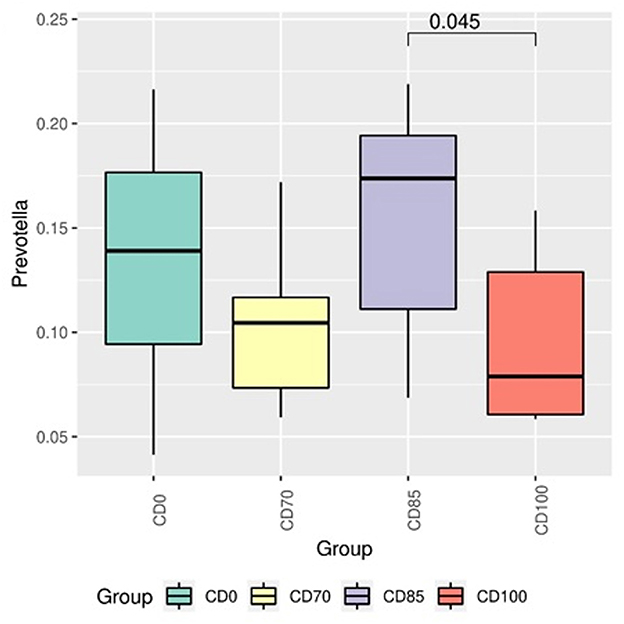
Figure 6. Differential abundance analysis of taxon of the Prevotella genus in finisher pigs fed 1 of 4 dietary treatments: a control diet containing isolated phytase and xylanase valued at 40 kcal of ME/kg (CD0), CD0 + β-mannanase (0.3 g/kg valued at 30 kcal of ME/kg) (CD70), CD0 + β-mannanase (0.3 g/kg valued at 45 kcal of ME/kg) (CD85), and CD0 + β-mannanase (0.3 g/kg valued at 60 kcal of ME/kg) (CD100). Data are averages of 6 pigs per dietary treatment. Means differed by Wilcoxon test (P < 0.05).
3.5. Carcass traits and meat quality
Pigs fed CD0 diet showed (P = 0.094) greater backfat thickness measured with ultrasound than pigs fed CD70 or CD85 diets (Table 5). In addition, animals fed CD0 or CD85 diets showed (P = 0.060) greater pH24h in the l. thoracis muscle than those fed CD100 diet.
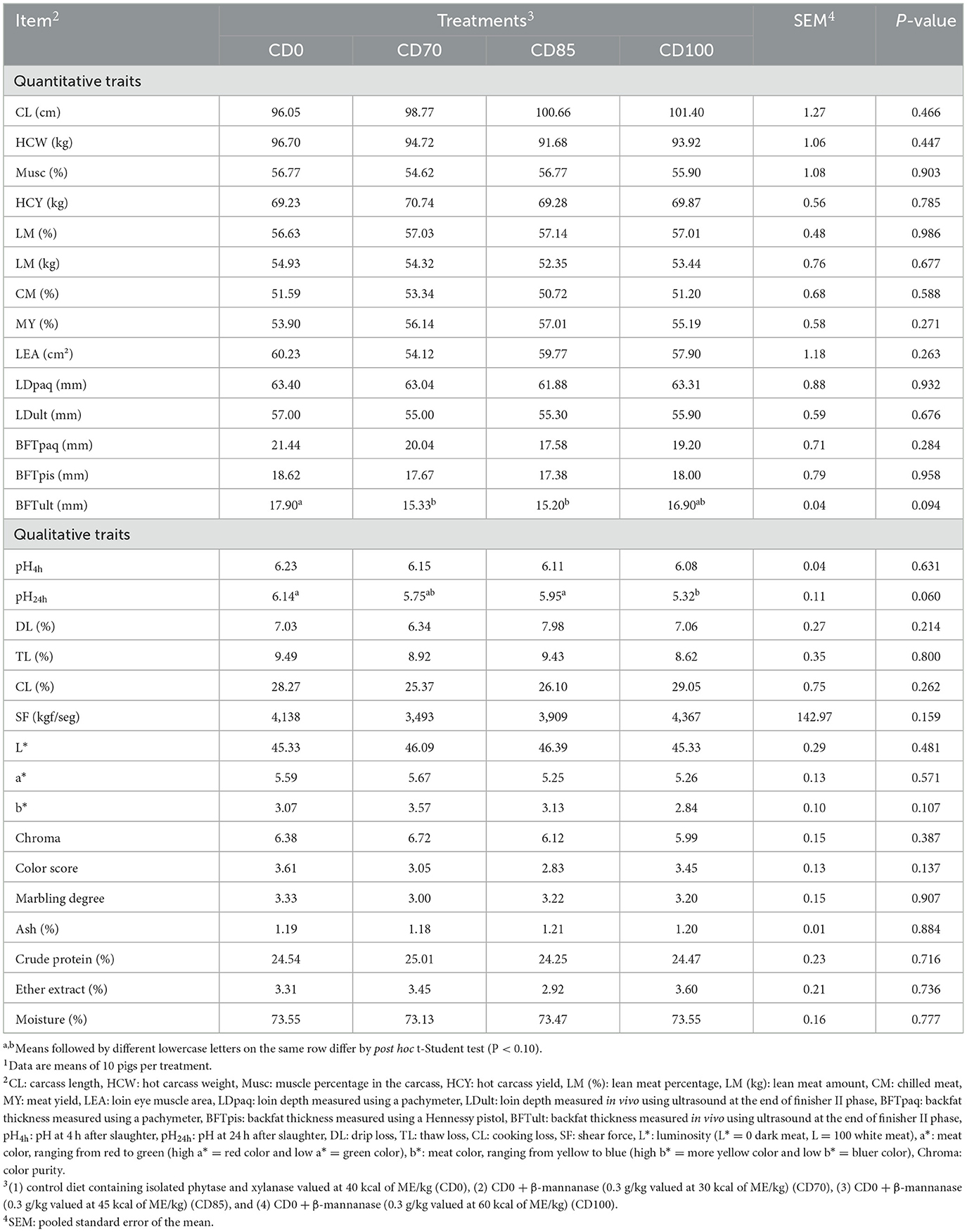
Table 5. Effect of β-mannanase supplementation in diets containing xylanase-phytase and reduced metabolizable energy on carcass traits and meat quality in finisher pigs on day 521.
4. Discussion
In the present study, animals were healthy throughout the experiment. However, pigs fed diets supplemented with β-mannanase supported growth performance due to the combined effect of these enzymes in the hydrolysis of antinutritional factors, and as energy sparing and extra energy supply (2, 4, 7). The energy-saving effect of diets supplemented with β-mannanase is attributed to an unnecessary immune deactivation caused by the β-mannans in plant products (2).
A lower SOD concentration is due to the different enzymatic antioxidant system in response to oxidative stress. When pigs are fed diets with reduced ME, the metabolism is changed to use body reserves such as energy and lipids (21). As a result, the process of nutrient oxidation produces energy for animal metabolism; however, energy production is also a source of free radicals (22). In the present study, the greater SOD activity in pigs was performed to eliminate reactive free radicals, as previously reported (23). However, this improved antioxidant capacity did not favor greater energy and nutrient usage in pigs fed CD70 diet.
A previous study (8) reported a higher glucose concentration in pigs fed diets supplemented with β-mannanase-xylanase. The authors mentioned above explained this result based on successful enzyme hydrolysis of NSP, unlike our study, where no differences among the dietary treatments in the biochemical blood profile were observed. The mechanisms of action of these enzymes are supported by the greater usage of hydrolyzed nutrients that favors absorption by the enterocytes in the small intestine (1). However, in the present study, a reduction of 100 kcal of ME/kg diet did not promote greater ATTD in pigs, even with β-mannanase supplementation. This result did not impair the nutrient ATTD coefficients and did not affect the occurrence of intestinal disorders such as diarrhea or increasing digesta viscosity.
On other hand, the results suggested that feeding the CD85 diet to finisher pigs promoted greater DP and DE compared to other dietary treatments, explained by a successful degradation of NSP (8) that improves nutrient usage and energy efficiency due to the increased effectiveness of host enzymes. This mechanism is performed by β-mannanase-xylanase enzymes via breaking down cell walls containing NSP (24) and reducing digesta viscosity (6). However, we did not observe changes on fecal consistency score or the total digesta passage rate in pigs.
Furthermore, β-mannanase supplementation in diets has been previously reported to stimulate the activity of endogenous enzymes (5) and hence favor a greater ATTD in pigs. The effects of phytase on ATTD of nutrients other than calcium and phosphorus have not been well established yet (6). However, the association of dietary xylanase-phytase has been reported to break down cell walls and release more phytic acid to be broken down by phytase (25).
Usually, the consumption of diets with greater energy content promotes increased backfat thickness, as well as the additional energy effect that can be provided by phytase in pig diets (7), which agrees with the results we observed. Pigs fed CD85 diet showed lower backfat thickness measured in vivo with ultrasound due to greater energy digestibility even with reduced ME dietary content (8). Although the animals fed CD70 diet had lower DE, the backfat thickness was positively influenced in the animals of this dietary treatment.
Contrary to our observation, enzyme supplementation increased energy digestibility and no effect on backfat thickness in pigs due to dietary energy content was observed in a previous study (24). However, our finding was similar to the one reported by (8), who also observed lower backfat thickness in pigs fed diets supplemented with β-mannanase-xylanase.
Phosphorus and phytate grouped with arabinoxylans have been previously reported (24) to increase redness and reduce water retention in the meat of pigs when exposed to xylanase-phytase action. A similar effect was not observed in the present study; however, we observed dietary treatment effects on pH24, which is related to meat quality regarding water retention capacity, color, softness, juiciness, and flavor. Overall, our results agree with those reported by (26), who summarized the higher quality traits of meat from finisher pigs and estimated values of 5.54 for pH24h, L* of 46.6, CL of 25.8%, and chroma of 6.2.
In the present study, differences in pH24h among dietary treatments are attributed to muscle glycogen concentration (although not determined in the present study), which largely depends on the diet provided to animals. The lower pH24h value in the meat of pigs fed CD100 diet is related to a greater rate of lactic acid-producing pyruvate, as evidenced by (27). Based on these pH24h values in meat, pigs fed CD0 and CD100 diets showed meats classified as DFD and PSE, respectively (18).
Bacterial diversity in the gastrointestinal tract was assessed in the present study because this is crucial in modulating intestinal functionality and is essential for metabolism, ATTD, and nutrient usage. In general, the balance of the commensal microbiota plays a role in the health of the host. This role is attributed to the diversity of genera and species that possesses protective function, reduces pathogens, inhabit intestinal surfaces, and produces antimicrobial substances (28). The above-mentioned roles attributed to intestinal microbiota promoted animal health during the experimental period.
No effects of dietary treatments on alpha and beta diversity were observed in finisher pigs. This lack of effect could be attributed to the dynamism of microbial communities and different profiles in the gastrointestinal tract segments. In our study, the most prevalent bacterial phyla in pigs were Firmicutes, Proteobacteria, and Bacteroidota, which agrees with previous studies (29, 30). Firmicutes and Bacteroidota are the phyla of greatest representation and paramount importance for gastrointestinal homeostasis (31). An increased incidence of Firmicutes may also be negatively correlated with the presence of potentiality pathogenic bacteria in the intestine of pigs (32). According to (33), a greater presence of organisms of the phylum Firmicutes may create a hostile intestinal environment for pathogenic bacteria colonization.
In a previous study (34), FBR was reported as widely accepted as an evaluative parameter beneficial for intestinal health; therefore, changes in this proportion can trigger several pathologies (35–37). In a study conducted by (38), a greater FBR in pigs was related to improved energy efficiency and growth performance. In addition, similar results were reported in a study conducted on poultry (39); however, these findings differ from our results.
In the present study, the Ruminococcaceae, Lactobacillaceae, and Lachnospiraceae families showed relative abundance in pigs. These families compose the central microbiota of the distal intestine portion and are found in similar proportions in the colon and feces (40). The CD70 diet has positively modulated the growth of Ruminococcaceae family in finisher pigs. This family produces xylanases, cellulases, α-glucosidases, α and β-galactosidases providing greater energy usage (41). In addition, bacteria belonging to the Ruminococcaceae family degrade complex carbohydrates. A reduction in this family has been associated with the use of calorie-rich diets and/or enhanced with carbohydrates (42). This effect could support the reduced Ruminococcaceae occurrence in pigs fed CD0 diet.
Furthermore, no treatment effect on the abundance of Prevotellaceae and Rikenellaceae families was observed in pigs. This finding is associated, in several studies, with a low G:F. Similarly, the Christensenellaceae family was not affected by treatments, which was related to improvement G:F in pigs (43), although a lower G:F was observed in finisher I pigs fed CD0 diet.
However, the Lachnospiraceae family was abundant in the pig gastrointestinal microbiota in both dietary treatments in the present study. This family is known to produce butyric acid (29), which plays a role in maintaining intestinal epithelium structure (41). In addition, the fecal microbiome in finisher pigs showed a relative abundance of families associated with short-chain fatty acids synthesis as final products of sugar fermentation, for example, the Oscillospiraceae (44) and Christensenellaceae families (45).
Prevotella is part of the phylum Bacteroidetes, which participates in immune system modulation, metabolic syndromes, and brain-intestine axis regulation (46). This genus was more abundant in pigs fed CD85 diet than in those fed CD100 diet. This result suggests that these changes are related to the different energy content among diets. In fact, Prevotella has been reported to play a role in carbohydrate metabolism, such as the degradation of polysaccharides and oligosaccharides usage (47). When analyzed together, part of the modulations observed in pigs occurred in families and genera that play a crucial role in gastrointestinal tract homeostasis.
5. Conclusion
Based on the assessing criteria in this study, β-mannanase supplementation in diets containing xylanase-phytase allows reducing 85 kcal of ME/kg because it improves the gain to feed ratio, energy and protein usage, and backfat thickness without metabolic and intestinal ecosystem disorders in finisher pigs. Furthermore, reducing dietary ME alters the fecal microbiome in finisher pigs regardless of the combined enzymes.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Ethics statement
All experimental procedures performed were approved by the Ethics Committee on the use of production animals at the Universidade Estadual do Oeste do Paraná (Authorization number 17/2022).
Author contributions
PC, MK, and TP: conceptualization, data curation, and project management. JG, PR, LA, DH, and SC: methodology. JG and NO: software. JG, PR, and NO: statistical analysis, formal analysis, and writing—original draft preparation. PC, SC, and MK: validation. PR, PC, LA, DH, and SC: investigation. JG, PR, MK, GG, HV, and TP: writing—review and editing. PC, SC, AD, and MK: supervision. All authors contributed to the article and approved the submitted version.
Funding
The authors declare that this study received funding from Elanco Animal Health Incorporated Company. The funder had the following involvement in the study: conceptualization, data curation, project management, validation, writing-review and editing, and supervision.
Acknowledgments
We thank the efforts and support in the research from the Elanco Animal Health Incorporated Company, Universidade Estadual do Oeste do Paraná, and the Copagril Agroindustrial Cooperative.
Conflict of interest
MK was employed by Elanco Animal Health Incorporated Company.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Liu S, Ma C, Liu L, Ning D, Liu Y, Dong B. β-Xylosidase and β-mannosidase in combination improved growth performance and altered microbial profiles in weanling pigs fed a corn-soybean meal-based diet. Asian-Australas J Anim Sci. (2019) 32:1734. doi: 10.5713/ajas.18.0873
2. Vangroenweghe F, Poulsen K, Thas O. Supplementation of a β-mannanase enzyme reduces post-weaning diarrhea and antibiotic use in piglets on an alternative diet with additional soybean meal. Porc Health Manag. (2021) 7:1–12. doi: 10.1186/s40813-021-00191-5
3. Mok CH, Lee JH, Kim BG. Effects of exogenous phytase and β-mannanase on ileal and total tract digestibility of energy and nutrient in palm kernel expeller-containing diets fed to growing pigs. Anim Feed Sci Technol. (2013) 186:209–13. doi: 10.1016/j.anifeedsci.2013.10.008
4. Tiwari UP, Chen H, Kim SW, Jha R. Supplemental effect of xylanase and mannanase on nutrient digestibility and gut health of nursery pigs studied using both in vivo and in vitro models. Anim Feed Sci Technol. (2018) 245:77–90. doi: 10.1016/j.anifeedsci.2018.07.002
5. Kipper M, Andretta I, Quadros VRD, Schroeder B, Pires PGDS, Franceschina CS, França I. Performance responses of broilers and pigs fed diets with β-mannanase. Rev Bras Zootec. (2020) 49:1–11. doi: 10.37496/rbz4920180177
6. Yang YY, Fan YF, Cao YH, Guo PP, Dong B, Ma YX. Effects of exogenous phytase and xylanase, individually or in combination, and pelleting on nutrient digestibility, available energy content of wheat and performance of growing pigs fed wheat-based diets. Asian-Australas J Anim Sci. (2017) 30:57–63. doi: 10.5713/ajas.15.0876
7. Silva CA, Callegari MA, Dias CP, Bridi AM, Pierozan CR, Foppa L, et al. Increasing doses of phytase from Citrobacter braakii in diets with reduced inorganic phosphorus and calcium improve growth performance and lean meat of growing and finishing pigs. PLoS ONE. (2019) 14:e0217490. doi: 10.1371/journal.pone.0217490
8. Cho JH, Kim IH. Effects of beta mannanase and xylanase supplementation in low energy density diets on performances, nutrient digestibility, blood profiles and meat quality in finishing pigs. Asian J Anim Vet Adv. (2013) 8:622–30. doi: 10.3923/ajava.2013.622.630
9. Rostagno HS, Albino LFT, Hannas MI, Donzele JL, Sakomura NK, Perazzo FG, et al. Tabelas Brasileiras para aves e suínos: composição de alimentos e exigências nutricionais. Viçosa, MG: UFV (2017).
10. Silva DJ, Queiroz AC. Análises de alimentos (métodos químicos e biológicos). Viçosa, MG: UFV (2002).
11. Hart GK, Dobb GJ. Effect of a fecal bulking agent on diarrhea during enteral feeding in the critically ill. JPEN J Parenter Enteral Nutr. (1988) 12:465–8. doi: 10.1177/0148607188012005465
12. Sakomura NK, Rostagno HS. Métodos de pesquisa em nutrição de monogástricos. Jaboticabal, SP: FUNEP (2016).
13. Kavanagh S, Lynch PB, O'Mara F, Caffrey PJ, A. comparison of total collection and marker technique for the measurement of apparent digestibility of diets for growing pigs. Anim Feed Sci Technol. (2001) 89:49–58. doi: 10.1016/S0377-8401(00)00237-6
14. Owusu-Asiedu AJFJ, Patience JF, Laarveld B, Van Kessel AG, Simmins PH, Zijlstra RT. Effects of guar gum and cellulose on digesta passage rate, ileal microbial populations, energy and protein digestibility, and performance of grower pigs. J Anim Sci. (2006) 84:843–52. doi: 10.2527/2006.844843x
15. Degnan PH, Ochman H. Illumina-based analysis of microbial community diversity. ISME J. (2012) 6:183–94. doi: 10.1038/ismej.2011.74
16. Caporaso JG, Lauber CL, Walters WA, Berg-Lyons D, Lozupone CA, Turnbaugh PJ, et al. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc Natl Acad Sci. (2011) 108:4516–22. doi: 10.1073/pnas.1000080107
17. Yilmaz P, Parfrey LW, Yarza P, Gerken J, Pruesse E, Quast C, et al. The SILVA and “all-species living tree project (LTP)” taxonomic frameworks. Nucleic Acids Res. (2013) 42:D643–48. doi: 10.1093/nar/gkt1209
18. Bridi AM, Silva CA. Métodos de avaliação da carcaça e da carne suína. Londrina, PR: Midigraft (2009).
19. McMurdie PJ, Holmes S. phyloseq: an R package for reproducible interactive analysis and graphics of microbiome census data. PLoS One. (2013) 8:e61217. doi: 10.1371/journal.pone.0061217
20. Lahti L, Shetty, S,. Introduction to the Microbiome R Package. (2018). Available online at: https://microbiome.github.io/tutorials/ (Accessed March 15, 2022).
21. Belhadj Slimen I, Najar T, Ghram A, Abdrrabba M. Heat stress effects on livestock: molecular, cellular and metabolic aspects, a review. J Anim Physiol Anim Nutr. (2016) 100:401–12. doi: 10.1111/jpn.12379
22. Celi P, Gabai G. Oxidant/antioxidant balance in animal nutrition and health: the role of protein oxidation. Front Vet Sci. (2015) 2:48. doi: 10.3389/fvets.2015.00048
23. Li Z, Tang L, Liu N, Zhang F, Liu X, Jiang Q, et al. Comparative effects of compound enzyme and antibiotics on growth performance, nutrient digestibility, blood biochemical index, and intestinal health in weaned pigs. Front Microbiol. (2021) 12:768767. doi: 10.3389/fmicb.2021.768767
24. Cho JH, Park JH, Lee DH, Lee JM, Song TH, Kim IH. Effects of xylanase supplementation on growth performance, digestibility, fecal gas emission, and meat quality in growing–finishing pigs. Can J Anim Sci. (2016) 97:95–100. doi: 10.1139/CJAS-2015-0198
25. Kim JC, Sands JS, Mullan BP, Pluske JR. Performance and total-tract digestibility responses to exogenous xylanase and phytase in diets for growing pigs. Anim Feed Sci Technol. (2008) 142:163–72. doi: 10.1016/j.anifeedsci.2007.07.004
26. Sardi L, Gastaldo A, Borciani M, Bertolini A, Musi V, Garavaldi A, et al. Pre-slaughter sources of fresh meat quality variation: The case of heavy pigs intended for protected designation of origin products. Animals. (2020) 10:2386. doi: 10.3390/ani10122386
27. Yin Y, Liu Y, Duan G, Han M, Gong S, Yang Z, et al. The effect of dietary leucine supplementation on antioxidant capacity and meat quality of finishing pigs under heat stress. Antioxidants. (2022) 11:1373. doi: 10.3390/antiox11071373
28. Duda-Chodak A, Tarko T, Satora P, Sroka P. Interaction of dietary compounds, especially polyphenols, with the intestinal microbiota: a review. Eur J Nutr. (2015) 54:325–41. doi: 10.1007/s00394-015-0852-y
29. Gresse R, Durand FC, Dunière L, Blanquet-Diot S, Forano E. Microbiota composition and functional profiling throughout the gastrointestinal tract of commercial weaning piglets. Microorganisms. (2019) 7:343. doi: 10.3390/microorganisms7090343
30. Kim BR, Shin J, Guevarra RB, Lee JH, Kim DW, Seol KH, et al. Deciphering diversity indices for a better understanding of microbial communities. J Microbiol Biotechnol. (2017) 27:2089–93. doi: 10.4014/jmb.1709.09027
31. Rychlik I. Composition and function of chicken gut microbiota. Animals. (2020) 10:103. doi: 10.3390/ani10010103
32. Mulder IE, Schmidt B, Stokes CR, Lewis M, Bailey M, Aminov RI, et al. Environmentally-acquired bacteria influence microbial diversity and natural innate immune responses at gut surfaces. BMC Biol. (2009) 7:1–20. doi: 10.1186/1741-7007-7-79
33. Molist F, Manzanilla EG, Pérez JF, Nyachoti CM. Coarse, but not finelyground, dietary fibre increases intestinal firmicutes:bacteroidetes ratio and reduces diarrhoea induced by experimental infection in piglets. Br J Nutr. (2012) 108:9–15. doi: 10.1017/S0007114511005216
34. Wang Z, Tang Y, Long L, Zhang H. Effects of dietary L-theanine on growth performance, antioxidation, meat quality and intestinal microflora in white feather broilers with acute oxidative stress. Front Vet Sci. (2022) 696: 889485. doi: 10.3389/fvets.2022.889485
35. Magne F, Gotteland M, Gauthier L, Zazueta A, Pesoa S, Navarrete P, Balamurugan R. The firmicutes/bacteroidetes ratio: a relevant marker of gut dysbiosis in obese patients? Nutrients. (2020) 12:1474. doi: 10.3390/nu12051474
36. Stojanov S, Berlec A, Štrukelj B. The influence of probiotics on the firmicutes/bacteroidetes ratio in the treatment of obesity and inflammatory bowel disease. Microorganisms. (2020) 8:1–16. doi: 10.3390/microorganisms8111715
37. Turnbaugh PJ, Ley RE, Mahowald MA, Magrini V, Mardis ER, Gordon JI. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature. (2006) 444:1027–31. doi: 10.1038/nature05414
38. Zhao W, Wang Y, Liu S, Huang J, Zhai Z, He C, et al. The dynamic distribution of porcine microbiota across different ages and gastrointestinal tract segments. PLoS ONE. (2015) 10:1–13. doi: 10.1371/journal.pone.0117441
39. Xu Y, Yang H, Zhang L, Su Y, Shi D, Xiao H, et al. High-through put sequencing technology to reveal the composition and function of cecal microbiota in Dagu chicken. BMC Microbiol. (2016) 16:1–9. doi: 10.1186/s12866-016-0877-2
40. Gierse LC, Meene A, Schultz D, Schwaiger T, Karte C, Schröder C, et al. A multi-omics protocol for swine feces to elucidate longitudinal dynamics in microbiome structure and function. Microorganisms. (2020) 8:1–20. doi: 10.3390/microorganisms8121887
41. Biddle A, Stewart L, Blanchard J, Leschine S. Untangling the genetic basis of fibrolytic specialization by Lachnospiraceae and Ruminococcaceae in diverse gut communities. Diversity. (2013) 5:627–40. doi: 10.3390/d5030627
42. Lagkouvardos I, Lesker TR, Hitch TCA, Gálvez EJC, Smit N, Neuhaus K, et al. Sequence and cultivation study of Muribaculaceae reveals novel species, host preference, and functional potential of this yet undescribed family. Microbiome. (2019) 7:28. doi: 10.1186/s40168-019-0637-2
43. Quan J, Cai G, Ye J, Yang M, Ding R, Wang X, et al. A global comparison of the microbiome compositions of three gut locations in commercial pigs with extreme feed conversion ratios. Sci Rep. (2018) 8:4536. doi: 10.1038/s41598-018-22692-0
44. Beaumont M, Cauquil L, Bertide A, Ahn I, Barilly C, Gil L, et al. Gut microbiota-derived metabolite signature in suckling and weaned piglets. J Proteome Res. (2021) 20:982–94. doi: 10.1021/acs.jproteome.0c00745
45. Morotomi M, Nagai F, Watanabe Y. Description of Christensenella minutagen. Nov., sp nov, isolated from human faeces, which forms a distinct branch in the order Clostridiales, and proposal of Christensenellaceae fam nov. Int J Syst Evol Microbiol. (2012) 62:144–9. doi: 10.1099/ijs.0.026989-0
46. Gibiino G, Lopetuso LR, Scaldaferri F, Rizzatti G, Binda C, Gasbarrini A. Exploring bacteroidetes: metabolic key points and immunological tricks of our gut commensals. Dig Liver Dis. (2018) 50:635–9. doi: 10.1016/j.dld.2018.03.016
Keywords: blood profile, carcass-meat traits, digestibility, exogenous enzymes, fecal microbiome, pig performance
Citation: Genova JL, Rupolo PE, Azevedo LBd, Henz D, Carvalho ST, Kipper M, Gonçalves GdAC, Vilela HLO, Pasquetti TJ, Oliveira NTEd, Dietrich ARM and Carvalho PLdO (2023) β-mannanase supplementation in diets reduced in 85 kcal metabolizable energy/kg containing xylanase-phytase improves gain to feed ratio, nutrient usage, and backfat thickness in finisher pigs. Front. Vet. Sci. 10:1144692. doi: 10.3389/fvets.2023.1144692
Received: 14 January 2023; Accepted: 27 February 2023;
Published: 16 March 2023.
Edited by:
Huansheng Yang, Hunan Normal University, ChinaReviewed by:
Damiano Cavallini, University of Bologna, ItalyHong Hu, Anhui Science and Technology University, China
Copyright © 2023 Genova, Rupolo, Azevedo, Henz, Carvalho, Kipper, Gonçalves, Vilela, Pasquetti, Oliveira, Dietrich and Carvalho. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jansller Luiz Genova, amFuc2xsZXIuZ2Vub3ZhQHVmdi5icg==