
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Trop. Dis. , 06 July 2022
Sec. Vector Biology
Volume 3 - 2022 | https://doi.org/10.3389/fitd.2022.853405
This article is part of the Research Topic Mosquitoes and Their Role in Disease Transmission: Recent Advances and Innovations View all 9 articles
 Mahamoudou Balboné1
Mahamoudou Balboné1 Dieudonné Diloma Soma2Moussa Namountougou2,3Samuel Fogné Drabo1Hamadou Konaté2,3Oumarou Toe3Koama Bayili2Georges Benson Meda2
Dieudonné Diloma Soma2Moussa Namountougou2,3Samuel Fogné Drabo1Hamadou Konaté2,3Oumarou Toe3Koama Bayili2Georges Benson Meda2 Roch Kounbobr Dabiré2
Roch Kounbobr Dabiré2 Olivier Gnankine1*
Olivier Gnankine1*Introduction: Malaria and dengue are two main vector-borne infectious diseases transmitted by Anopheles gambiae and Aedes aegypti, respectively, in tropical and subtropical regions. The concern for environmental safety and the increasing development of resistance to the chemical in main arthropod vectors raises interest in the search for botanicals such as essential oils (EOs) that can be used in vector control.
Methods: Larvicidal bioassays were performed according to the WHO standard methods using Ae. aegypti and An. gambiae larvae collected from Bobo-Dioulasso and in the Kou Valley (VK), respectively, two areas located in Houet Province of western Burkina Faso. Kisumu and Bora bora, the susceptible strains of An. gambiae and Ae. aegypti, respectively, were used as controls. OEs extracted from leaves of five aromatic plants, Cymbopogon citratus, Cymbopogon nardus, Eucalyptus camaldulensis, Lippia multiflora, and Ocimum americanum, naturally growing in Burkina Faso were tested. The pyriproxyfen was used as a positive control.
Results: As a result, the lethal concentrations (LC50) for Ae. aegypti from Bobo-Dioulasso and An. gambiae from VK ranged from 41.9 to 103.8 ppm and 39.5 to 138.1 ppm, respectively. As for LC90 values, they ranged from 74.6 to 311.3 ppm for Ae. aegypti from Bobo-Dioulasso and from 90.2 to 328.9 ppm for An. gambiae from VK. Among the EOs tested, L. multiflora showed the highest activity against all the strains of An. gambiae and Ae. aegypti larvae. No difference in terms of LC50 values was found between L. multiflora and pyriproxyfen used as a positive control on An. gambiae larvae. It is not the case for Aedes aegypti populations, where pyriproxyfen remains the most toxic. Overall, An. gambiae populations were the most susceptible to EOs tested as compared to Ae. aegypti populations.
Conclusion: Our study furthers our knowledge of the larvicidal activity of EOs in the western part of Burkina and opens new avenues in their putative use in vector control strategies.
Vector-borne diseases are human diseases caused by parasites, viruses, or bacteria transmitted by vectors. Each year, more than 700,000 deaths occur worldwide due to vector-borne diseases such as dengue, yellow fever, Japanese encephalitis, leishmaniasis, Chagas disease, onchocerciasis, malaria, schistosomiasis, and human African trypanosomiasis (1). Among these diseases, malaria remains the most serious vector-borne disease, affecting between 300 and 500 million people, and 1.4 to 2.6 million deaths annually throughout the world were recorded. More than 40% of the world’s population lives in malarious areas (2).
According to the epidemiological bulletin, in the first half of 2018, Burkina Faso recorded 3,501,245 cases of malaria, including 1,002 deaths, for a case-fatality rate of 0.7% (3). The same is true for dengue, where the number of reported cases worldwide peaked in 2019, and all WHO regions were affected (4). In Burkina Faso, an increase in the weekly incidence of dengue cases was noted from week 31 of the year 2017 (5).
Current mosquito control strategies depend primarily on chemical insecticides. The discovery, development, and use of chemicals have reduced the interest in plant products. However, widespread use of these insecticides in public health and agriculture for the control of vector and pest species has favored many concerns such as the increasing physiological resistance in major vector species, environmental pollution, and toxic hazards to human and other non-target organisms due to their broad spectrum of activity (6–9). As a result, there has been an increased interest in developing potential alternative or additional control methods/tools that are effective against the target vector species, which are environmentally safe, biodegradable, and low cost and can be used by individuals and communities in specific situations (10, 11). Therefore, in recent years, various workers have been concentrating their efforts on the search for natural products derived from plants as an alternative to conventional insecticides used in controlling vectors for which resistance was detected (12). Among many natural products, essential oils (EOs) and their constituents have received considerable attention in the search for new pesticides and have been found to possess an insecticidal potential (13).
EOs show several interesting properties. First, they easily penetrate insect cuticles, which increases their bioavailability (14). This property could be of interest if it results in a shortened stay of insects on treated surfaces. Second, EO compounds such as acyclic or monocyclic monoterpenes are small and volatile molecules that might have spatial repellency properties. For example, insect sensilla are specialized for detecting odorants and have been shown to respond to volatile monoterpenes (14). Finally, active compounds in EOs may have a specific mode of action, which makes them good alternatives to the use of pyrethroids.
The EO of Lippia multiflora is used as a spice, a meal condiment, a drink flavoring (15), and a mouthwash. In addition to insecticidal activities, Abena et al. (16) found analgesic, antipyretic, and anti-inflammatory effects of L. multiflora oil. Cymbopogon citratus EO has antimicrobial activity against Helicobacter pylori, a bacterium responsible for gastroduodenal diseases (17). Cymbopogon nardus is a medicinal plant because of its analgesic effect and is widely used in gastronomy (18). Also, several studies noticed the insecticidal properties of EO of C. nardus, C. citratus, Ocimum americanum, and Eucalyptus camaldulensis (18–21).
In Burkina Faso, some studies on the insecticidal effects of EOs with mosquitoes in general and mosquito larvae in particular have involved strains restricted to Ouagadougou (21–23). Little is known about the EO susceptibility in Anopheles and Aedes populations in the western part of this country. Moreover, previous studies did not take into account the positive control in the various tests.
Our study aimed at evaluating the larvicidal activities of EOs of five local aromatic plants from Burkina Faso, including C. citratus, C. nardus (DC.) Stapf, E. camaldulensis Dehn, L. multiflora, and O. americanum (Wild.) A.J. Paton in terms of lethal concentrations (LC50 and LC90) upon field populations of Aedes aegypti from Bobo-Dioulasso and Anopheles gambiae from the Vallée du Kou (VK).
An. gambiae larvae were collected from district number 7 of the Vallée du Kou (Bama) (11°24′N and 4°24′O), a rice-growing area located 30 km north of Bobo-Dioulasso. Ae. aegypti larvae were collected from tires in various urban settings in Bobo-Dioulasso, the economic capital city of Burkina Faso. Bobo-Dioulasso and Bama are two localities in the province of Houet in western Burkina Faso. The larvae were collected from June to October 2021 and transported to the insectary of the Institute of Research in Health Sciences (IRSS) located in Bobo-Dioulasso where 3 and 4 larval instars were sorted according to Filho (24). Larvae from laboratory reference strains were used as controls. The insectary conditions were 27°C ± 2°C temperature, 80% ± 10% relative humidity, and 12-h light and 12-h dark photoperiod.
The five plant species tested, C. citratus, C. nardus (Linn.), E. camaldulensis, L. multiflora, and O. americanum, were used in this current study. They were chosen firstly due to endogenous data provided from informants questioned in areas investigated as well as the data from the literature. These plants were collected from the garden of the Research Institute of Applied Sciences and Technologies (IRSAT). The identification of the plants and extraction of EOs were performed by IRSAT. All the EOs were extracted from the leaves of the test plants by hydrodistillation (HD) using a clevenger-type apparatus and stored in a dark glass bottle at 4°C prior to use. The oils thus obtained were separated from water in the condenser and stored in airtight containers under refrigeration (4°C) till their later use for larval bioassays. The major compounds of these EOs were obtained by gas chromatography–mass spectrometry (GC-MS).
The larval bioassay tests were carried out following the standard WHO larval bioassay test method (25). They were carried out in the laboratory of the Institute of Research in Health Sciences located in Bobo-Dioulasso. Clear plastic cups with capacities of 200 ml each were used for the larvicidal bioassays. An appropriate amount of each EO was dissolved in acetone to prepare 1 ml of stock solution. Fresh stock solutions of each of the above stock solutions were prepared by adding distilled water to produce the required concentrations (50, 100, 150, and 200 ppm). Four replicates were carried out for each test concentration and species of mosquito larvae. Twenty active third and fourth larval instars of An. gambiae and Ae. aegypti in 10 ml of distilled water were transferred into each clear plastic cup that contained 139 ml of distilled water. One milliliter of the mixing solution of EO and acetone was added to each cup that contained 149 ml of distilled water to give a final solution of 150 ml with the desired concentrations. Two replicates of the control were carried out simultaneously with 149 ml of distilled water and 1 ml of acetone. Pyriproxyfen, the reference chemical larvicidal, was used at different concentrations as a positive control.
Analyses were performed by using the statistical software XLSTAT version 2,015.1.01. The LC50, LC90, and the 95% CIs were calculated by probit analysis using the same statistical software in order to compare the larvicidal potency of the plants and susceptibility of the test mosquito larvae. LC50 and LC90 values were judged as significantly different between the EOs (p < 0.05) if the CIs did not overlap (14, 26). In all the tests, no control mortality was detected after the 24-h exposure; hence, no correction was required based on Abbot’s formula.
The major compounds of the 5 EOs are summarized in Table 1. The EO of C. citratus was found to be rich in geranial (48.1%), neral (35.8%), and myrcene (11%). The EO of C. nardus was predominantly composed of citronellal (41.7%), geraniol (20.8%), and β-elemene (11%). The EO of E. camaldulensis was rich in 1,8-cineole (59.5%) and α-pinene (9.17%). The OE of L. multiflora was characterized by p-cymene (25.27%), β-caryophyllene (12.7%), and thymol (11.88%). O. americanum EO was characterized by a high percentage of 1,8-cineole (31.22%) followed by camphor (12.73%).
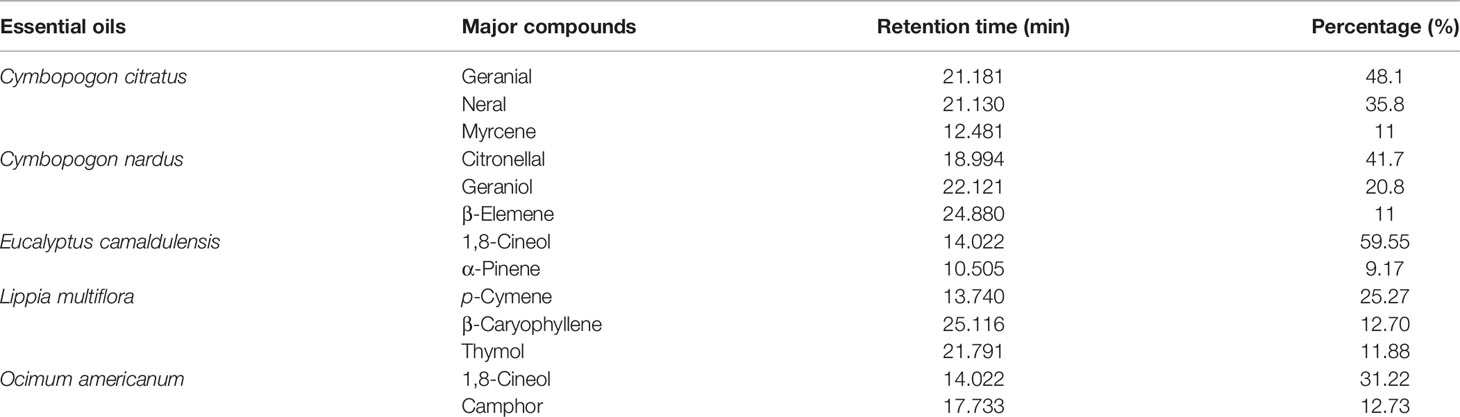
Table 1 Major compounds of the 5 essential oils tested on larvae of Anopheles gambiae and Aedes aegypti.
EOs from 5 plant species demonstrated larvicidal activities against susceptible strains and field-collected 3 and 4 larval instars of An. gambiae. Overall, five EOs were the most toxic on 3 and 4 larval instars of An. gambiae susceptible strain. On the susceptible strain larvae of An. gambiae (Kisumu), the LC50 and LC90 values were from 3.9 to 75.8 ppm and from 29.5 to 193.5 ppm, respectively (Table 2). On field-collected larvae of An. gambiae (VK), the LC50 and LC90 values were between 39.5 to 138.1 ppm and from 90.2 to 328.9 ppm, respectively (Table 3).

Table 2 Median and 90% lethal concentrations and their Confidence Limits (CLs) of plant essential oils against 3 and 4 larval instars of Anopheles gambiae susceptible strain from Kisumu.
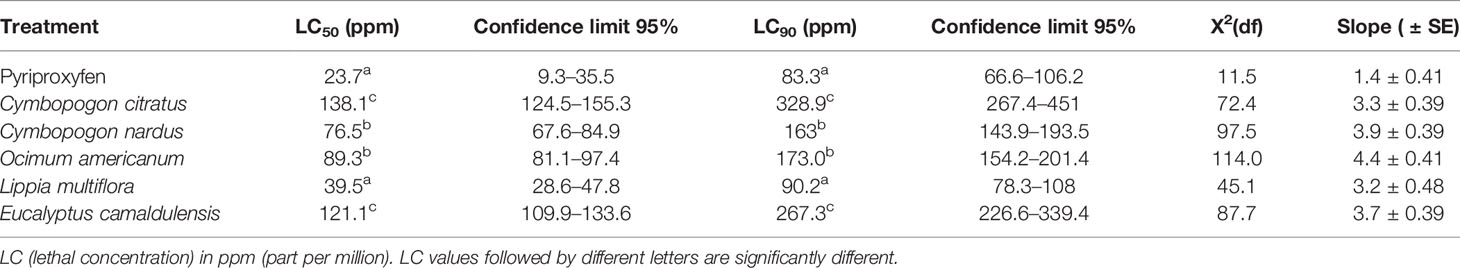
Table 3 Median and 90% lethal concentrations and their Confidence Limits (CLs) of plant essential oils against 3 and 4 larval instars of Anopheles gambiae from VK, field strain.
Regarding LC50 and LC90 values, L. multiflora exhibited the highest larvicidal activity against the 2 strains of An. gambiae (LC50 = 3.9 ppm, LC90 = 29.5 ppm for Kisumu larvae and LC50 = 39.5 ppm and LC90 = 90.2 ppm for VK larvae).
As for O. americanum, it exhibited the weakest larvicidal activity (LC50 = 75.8 ppm; LC90 = 193.5 ppm) for Kisumu larvae, while C. citratus showed the weakest larvicidal activity (LC50 = 138.1 ppm; LC90 = 328.9 ppm) on VK larvae.
Furthermore, oils from E. camaldulensis, C. nardus, and C. citratus still showed strong larvicidal activities after L. multiflora with LC50 values <50 ppm and LC90 values <91 ppm on Kisumu larvae. On VK larvae, EOs of C. nardus, O. americanum, and E. camaldulensis showed strong larvicidal activities after L. multiflora with LC50 values <140 ppm and LC90 values <270 ppm.
Only the EO of L. multiflora gives LC50 and LC90 values close to that of the positive control pyriproxyfen against Kisumu and VK larvae.
Based on the overlap of the CIs of the LC50 and LC90 values, no significant difference was observed between L. multiflora and pyriproxyfen on the two An. gambiae strains tested.
All EOs from five (5) plant species exhibited toxicity against susceptible strain larvae and 3 and 4 larval instars of Ae. aegypti field-collected strain. Tables 4 and 5 show the LC50 and LC90 values of the EOs of five (5) plants tested against susceptible strains and 3 and 4 larval instars of Ae. aegypti field-collected strain.
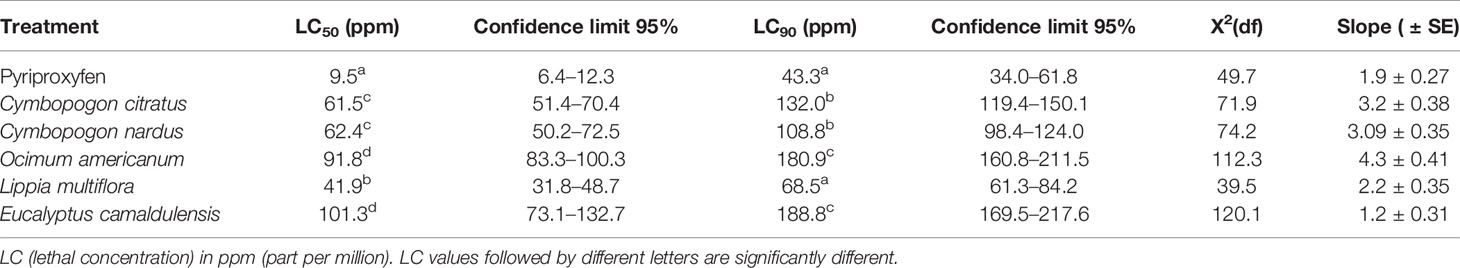
Table 4 Median and 90% lethal concentrations and their Confidence Limits (CLs) of plant essential oils against 3 and 4 larval instars of Aedes aegypti susceptible strain from the Bora bora.
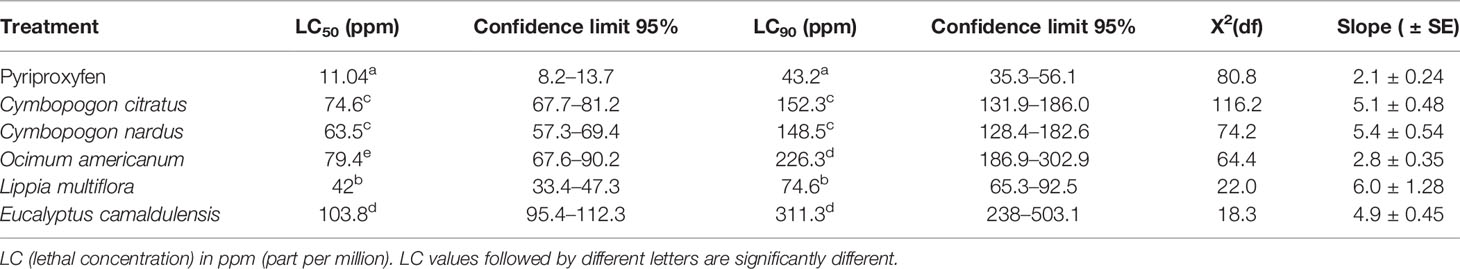
Table 5 Median and 90% lethal concentrations and their Confidence Limits (CLs) of plant essential oils against 3 and 4 larval instars of Aedes aegypti from Bobo-Dioulasso, field strain.
As in the case of An. gambiae populations, among the EOs tested, L. multiflora was the most toxic EO on the two (2) strains of Ae. aegypti tested. Indeed, LC50 and LC90 values were 42 and 74.6 ppm, respectively, for L. multiflora tested on the susceptible strain of Ae. aegypti (Bora bora) and 3 and 4 larval instars of Ae. aegypti field-collected strain from Bobo-Dioulasso and lower for those from other EOs tested where the LC50 and LC90 values ranged from 61.5 to 103.8 ppm and 108.8 to 311.3 ppm, respectively.
For the Bora bora strain, LC50 and LC90 values were 61.5, 62.4, 91.8, 41.9, and 101.3 ppm and 132, 108.8, 180.9, 68.5, and 188.8 ppm for EOs from C. citratus, C. nardus, E. camaldulensis, L. multiflora, and O. americanum, respectively (Table 4). These values were close to those obtained with the field strain collected from Bobo-Dioulasso (LC50 and LC90 values were 74.6, 63.5, 79.4, 41.9, and 103.8 ppm and 152.3, 148.5, 226.3, 74.6, and 1,053.6 ppm with EOs from C. citratus, C. nardus, E. camaldulensis, L. multiflora, and O. americanum, respectively) (Table 5). EO of E. camaldulensis remains the least toxic to both strains of Ae. aegypti regarding the LC50 and LC90 values above 100 and 188 ppm, respectively.
Based on the overlap of the CIs of the LC50 and LC90 values, there were no significant differences between L. multiflora and pyriproxyfen at LC90 with Bora bora. Also, there were no significant differences between C. citratus and C. nardus regarding LC50 and LC90 values with the two Ae. aegypti strains tested. Also, there were no significant differences between E. camaldulensis and O. americanum regarding LC90 with the two Ae. aegypti strains tested and LC50 with Bora bora. It is also the same with C. citratus and C. nardus, in regard to LC50 and LC90 values found in two Ae. aegypti strains tested.
Up to now, long-lasting insecticidal nets (LLIN) and indoor residual spraying (IRS) remain the two main tools targeting mainly adults An. gambiae and to lesser extent Ae. aegypti vectors. Thus, there is an urgent need to develop innovative tools and techniques including EOs used against eggs, larvae instar, and adults in order to increase the success of vector control (11).
Our study aimed at evaluating the bioefficacy of five EOs on Ae. aegypti and An. gambiae larvae according to the WHO standard methods. Most studies have already performed the larvicidal tests but did not include the chemical control pyriproxyfen, an insect growth regulator (IGR). IGR acts by mimicking one of these hormones or directly perturbating cuticle formation/deposition or lipid biosynthesis.
In this current study, all EOs tested exhibited variable insecticide activity against the larvae of An. gambiae and Ae. aegypti. Regarding LC50 and LC90 values, the EOs from C. citratus, C. nardus, E. camaldulensis, L. multiflora, and O. americanum exhibited higher larvicidal activities against third and fourth instar laboratory-reared larvae of An. gambiae and Ae. aegypti after 24 h of exposure, the most toxic of all being that of L. multiflora. LC50 of EO from E. camaldulensis was the least toxic against Ae. aegypti, whereas C. citratus remains the least toxic against An. gambiae. These data are not in agreement with those found by Manh et al. (27), who reported that on Ae. aegypti larvae, C. citratus EO (LC50 = 120.6 ppm) was less effective than that of E. camaldulensis (LC50 = 33.7 ppm).
Our results obtained with C. nardus on An. gambiae in this study were below those obtained by Ahouansou et al. (19), who obtained a value of 97.3 ppm for LC50 on An. gambiae. Our results were similar to those reported by Solon et al. (28) on Ae. aegypti populations. In their study, O. americanum and C. citratus exhibited values of LC50 equaling 67 and 69 ppm, respectively. However, the results with O. americanum were below those reported by Wangrawa et al. (21) on An. gambiae whose LC50 was 209.84 ppm.
The LC50 and LC90 values obtained with the L. multiflora in this current study were lowest than those reported by Bassole et al. (23). Recently, the same trend was observed by Yameogo et al. (22), who showed also the toxic effect of L. multiflora against Ae. aegypti populations from Tabtenga, Ouagadougou.
Interestingly, the high toxicity of the L. multiflora EO could result in the presence of three major components: thymol, p-cymene, and β-caryophyllene. According to Folashade (29), thymol, germacrene D, p-cymene, thymyl acetate, and sabinene were found in EOs of L. multiflora in Ghana and seem to be involved in Lippia toxicity. Conversely, the lower larvicidal activities of C. citratus, C. nardus, E. camaldulensis, and O. americanum oils could be explained by the minor effect of piperitone, geraniol, and 1.8-cineole against Ae. aegypti and An. gambiae larvae. This could explain the differences observed in biological activity among the EOs.
Regarding the LC50 and LC90 values, the bioefficacy of L. multiflora on An. gambiae strains is closer to that of the positive control used in this study, which is pyriproxyfen, the reference larvicide used against larvae. This would open new alternatives for mosquito control.
Globally, in An. gambiae strain populations, the laboratory-reared larvae were found to be the most susceptible to EOs than those from the field population (VK), known for their resistance to pyrethroid (30–32).
As observed in An. gambiae populations, Ae. aegypti larvae were the most susceptible to L. multiflora after the chemical pyriproxyfen. The LC50 values found in this study were slightly lower than those observed in Ae. aegypti populations from Ouagadougou (22), explaining the occurrence of the phenotypic resistance to the main chemical.
As shown by Namountougou et al. (33), tests performed with deltamethrin on Ae. aegypti populations collected from Bobo-Dioulasso showed mortality rates reaching 89.62% and 82.72% for 2013 and 2014, respectively. Mortality rates of Ae. aegypti collected from Ouagadougou were 50.7% and 20.7% for 2013 and 2014, respectively. All these values were lower than 90%, suggesting a resistance. It is very urgent to search for an alternative to chemicals. Our data will help to develop innovative strategies based on EOs.
Our results showed that the use of EOs as larvicides is a promising strategy. Therefore, they could be the object of particular attention in the search for new natural, selective, and biodegradable larvicidal products that can be used in public health vector control programs against An. gambiae and Ae. aegypti in particular.
The original contributions presented in the study are included in the article/supplementary materials, further inquiries can be directed to the corresponding author/s.
MB and OG designed the study. DS critically supervised the study. MB, HK, GM, and OT carried out the laboratory experiments. MB, SD, and OG analyzed and interpreted the data and drafted the manuscript. OG, RD, and MN revised the manuscript. All authors contributed to the article and approved the submitted version.
Funding for this study was provided partly by the TWAS 18-163 RG/BIO/AF/AC_G-FR3240303649 and the Centre d’excellence Africain (CEA) en Innovations biotechnologiques pour l’elimination des maladies a transmission vectorielle. I also thanks LAMIVECT (Laboratoire Mixte International sur les Maladies à transmissions Vectorielle) for supporting publication fees.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We are indebted to “Institut de recherche en Sciences Appliquées et technologiques” (IRSAT) for providing us with essential oils.
3. Ministere de la Santé du Burkina Faso, Direction des statistiques sectorielles. Rapport Final. (2018).
4. WHO. Noncommunicable Diseases (2021). Available at: https://www.who.int/news-room/fact-sheets/detail/n.
5. Ministère de la Santé. “Ministère de la Santé du Burkina Faso, Direction des statistiques sectorielles“Flambée de cas de dengue au Burkina Faso,”. In: Sitrep Dengue, vol. 26 p.11. (2017).
6. Minjas JN. “Laboratory Observation on the Toxicity of Swartzia Madaguscariensis (Leguminosae) Extract to Mosquito Larvae. Trans R Soc Trop Med Hyg (1986) 80:460–1. doi: 10.1016/0035-9203(86)90345-7
7. WHO. Vector Resistance for Pesticides. World Health Organization Technical Report Serial 818 Vol. 17. . Geneva: WHO (1992) p. 62 pp. 499–504.
8. Hemingway J. Parasitology: Enhanced New Ways to Control Malaria. Science (2004) 303:1984–5. doi: 10.1126/science.1096548
9. Gnankine O, Bassolé IHN, Chandre F, Glitho IA, Akogbeto M, Dabiré RK, et al. Insecticide Resistance in Bemisia Tabaci Gennadius (Homoptera: Aleyrodidae) and Anopheles Gambiae Giles (Diptera: Culicidae) Could Compromise the Sustainability of Malaria Vector Control Strategies in West Africa. Acta Tropica (2013) 128(1):7–17. doi: 10.1016/j.actatropica.2013.06.004
10. Redwane A, Lazrek HB, Bouallam S, Markouk M, Amarouch H, Jana M. Larvicidal Activity of Extracts From Querus Lusitania Var. Infectoria Galls (Oliv). J Ethnophamacol (2002) 79:261–3. doi: 10.1016/S0378-8741(01)00390-7
11. Gnankineí O, Bassoleí ILHN. Essential Oils as an Alternative to Pyrethroids ‘ Resistance Against Anopheles Species Complex Giles. molecules (2017), 22(10):1321. doi: 10.3390/molecules22101321
12. Zoubiri S, Baaliouamer A. Potentiality of Plants as Source of Insecticide Principles. J Saudi Chem Soc (2014) 18(6):925–38. doi: 10.1016/j.jscs.2011.11.015
13. Tripathi AK, Upadhyay S, Bhuiyan M, Bhattacharya PR. A Review on Prospects of Essential Oils as Biopesticide in Insect-Pest Management. Pharmacol Phytother (2009) 1(5):52–63.
14. Deletre E, Martin T, Campagne P, Bourguet D, Cadin A, Menut C, et al. Repellent, Irritant and Toxic Effects of 20 Plant Extracts on Adults of the Malaria Vector Anopheles Gambiae Mosquito. PloS one (2013) 8(12):1–10. doi: 10.1371/journal.pone.0082103
15. Abegaz B, Asfaw N, Lwande W. Constituents of the Essential Oils From Wild and Cultivated Lippia Adoensis Hochst. Ex Walp. J Essent Oil Res (1993) 5(5):487–91. doi: 10.1080/10412905.1993.9698268
16. Daferera BD, Sokmena A, Sokmen M, Polissiou M. Antimicrobial and Antioxidant Activities of the Essential Oil and Various Extracts of Salvia Tomentosa Miller (Lamiaceae). Food Chem (2005) 90(3):333–40. doi: 10.1016/j.foodchem.2003.09.013
17. Ohno T, Kita M, Yamaoka Y, Imamura S, Yamamoto T, Mitsufuji S, et al. Antimicrobial Activity of Essential Oils and Other Plant Extracts. Helicobacter (2003) 8:207–15. doi: 10.1046/j.1523-5378.2003.00146.x
18. Abena AA, Diatewa GM, Gakosso M, Gbeassor TH, Hondi-Assah, Ouamba JM. Comparative Chemical and Analgesic Properties of Essential Oils of Cymbopogon Nardus (L) Rendle of Benin and Congo. Afr J Tradit Complement Altern Med (2007) 4(3):267–72. doi: 10.4314/ajtcam.v4i3.31218
19. Ahouansou AC, Fagla SRM, Tokoudagba JM, Toukourou H, Badou YK, Gbaguidi FA. Chemical Composition and Larvicidal Activity of the Essential Oil of Cymbopogon Nardus (L.) Rendle on Anopheles Gambiae. Int J Biol Chem Sci (2019) 13(3):1861. doi: 10.4314/ijbcs.v13i3.53
20. Drabo SF, Olivier G, Bassolé IHN, Nébié RC, Laurence M. Susceptibility of MED-Q1 and MED-Q3 Biotypes of Bemisia Tabaci (Hemiptera: Aleyrodidae) Populations to Essential and Seed Oils. J Econ Entomol (2017) 110(3):1031–8. doi: 10.1093/jee/tox100
21. Wangrawa DW, Badolo A, Guenne S, Guelbeíogo WM, Kiendreíbeogo M, Sagnon N, et al. Larvicidal and Oviposition- Deterrence Activities of Four Local Plant Extracts From Burkina Faso Against Anopheles Gambiae s. L. (Diptera: Culicidae). Int J Mosquito Res (2016) 3(6):11–9.
22. Yaméogo F, Wangrawa DW, Sombié A, Sanon A, Badolo A. Insecticidal Activity of Essential Oils From Six Aromatic Plants Against Aedes Aegypti, Dengue Vector From Two Localities of Ouagadougou, Burkina Faso. Arthropod Plant Interact (2021) 15(4):627–34. doi: 10.1007/s11829-021-09842-4
23. Bassoleí IHN, Guelbeíogo WM, Neíbieí R, Costantini C, Sagnon NF, Kabore ZI, et al. Ovicidal and Larvicidal Activity Against Aedes Aegypti and Anopheles Gambiae Complex Mosquitoes of Essential Oils Extracted From Three Spontaneous Plants of Burkina Faso. Parasitologia (2003) 45:23–6. doi: 10.1007/s11829-021-09842-4
24. Pereira Filho AA, Pessoa GCD, Yamaguchi LF, Stanton MA, Serravite AM, Pereira RHM, et al. Larvicidal Activity of Essential Oils From Piper Species Against Strains of Aedes Aegypti (Diptera: Culicidae) Resistant to Pyrethroids. Front Plant Sci (2021) 12:685864. doi: 10.3389/fpls.2021.685864
25. WHO. World Malaria Report 2015 - Summary. (2015). Available at: http://www.who.int/malaria/publications/world-malaria-report-2015/report/en/.
26. Petersen JL, Floore TG, Brogdon WG. Diagnostic Dose of Synergized D-Phenothyrin for Insecticide Susceptibility Testing by Bottle Bioassay. J Am Mosq Control Assoc (2004) 20:183–8.
27. Manh HD, Hue DT, Hieu NTT, Tuyen DTT, Tuyet OT. The Mosquito Larvicidal Activity of Essential Oils From Cymbopogon and Eucalyptus Species in Vietnam. Insects (2020) 11(2):1–7. doi: 10.3390/insects11020128
28. Cavalcanti ES, Morais SM, Lima MA, Santana EW. Larvicidal Activity of Essential Oils from Brazilian Plants Against Aedes aegypti L. Mem Inst Oswaldo Cruz (2004) 99(5):541–4. doi: 10.1590/S0074-02762004000500015
29. Folashade KO, Omoregie EH. Essential Oil of Lippia Multiflora Moldenke : A Review. J Appl Pharm Sci (2012) 02(1):15–23.
30. Namountougou M, Soma DD, Kientega M, Balboneí M, Kaboreí DPA, Drabo SF, et al. Insecticide Resistance Mechanisms in Anopheles Gambiae Complex Populations From Burkina Faso, West Africa. Acta Tropica (2019) 197:1–93201–12. doi: 10.1016/j.actatropica.2019.105054
31. Chandre F, Darrier LM, Akogbeto M, Faye O, Mouchet J, Guillet P. Status of Pyrethroid Resistance in Anopheles Gambiae Sensu Lato. Bulletin of the World Health Organization (1999) 77(3):230–4.
32. Poda SB, Soma DD, Hien A, Namountougou M, Gnankineí O, Diabateí A, et al. Targeted Application of an Organophosphate ‐ Based Paint Applied on Windows and Doors Against Anopheles Coluzzii Resistant to Pyrethroids Under Real Life Conditions in Vallíe Du Kou, Burkina Faso (West Africa). Malaria J (2018) 17(136):1–9. doi: 10.1186/s12936-018-2273-x
33. Namountougou M, Soma DD, Balboneí M, Kaboreí AD, Kientega M, Hien A, et al. Monitoring Insecticide Susceptibility in Aedes Aegypti Populations From the Two Biggest Cities, Ouagadougou and Bobo-Dioulasso, in Burkina Faso: Implication of Metabolic Resistance. Trop Med Infect Dis (2020) 5:84. doi: 10.3390/tropicalmed5020084
Keywords: resistance, essential oils, larvicidal activities, lethal concentration, Bobo-Dioulasso
Citation: Balboné M, Soma DD, Namountougou M, Drabo SF, Konaté H, Toe O, Bayili K, Meda GB, Dabiré RK and Gnankine O (2022) Essential Oils From Five Local Plants: An Alternative Larvicide for Anopheles gambiae s.l. (Diptera: Culicidae) and Aedes aegypti (Diptera: Culicidae) Control in Western Burkina Faso. Front. Trop. Dis 3:853405. doi: 10.3389/fitd.2022.853405
Received: 12 January 2022; Accepted: 30 March 2022;
Published: 06 July 2022.
Edited by:
Cleber Galvão, Oswaldo Cruz Foundation (Fiocruz), BrazilReviewed by:
Carlos Manuel Dutok-Sánchez, Universidade Federal do Amapá, BrazilCopyright © 2022 Balboné, Soma, Namountougou, Drabo, Konaté, Toe, Bayili, Meda, Dabiré and Gnankine. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Olivier Gnankine, b2xpZ25hbmtpbmVAZ21haWwuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.