 Dariush Bodaghi
Dariush Bodaghi Yuxing Wang2†
Yuxing Wang2† Geng Liu
Geng Liu Xudong Zheng
Xudong Zheng- 1Department of Mechanical Engineering, University of Maine, Orono, ME, United States
- 2Department of Computer Engineering, Rochester Institute of Technology, Rochester, NY, United States
- 3Department of Engineering, King’s College, Wilkes-Barre, PA, United States
- 4Department of Mechanical Engineering, Rochester Institute of Technology, Rochester, NY, United States
This study presents a novel method that combines a computational fluid-structure interaction model with an interpretable deep-learning model to explore the fundamental mechanisms of seal whisker sensing. By establishing connections between crucial signal patterns, flow characteristics, and attributes of upstream obstacles, the method has the potential to enhance our understanding of the intricate sensing mechanisms. The effectiveness of the method is demonstrated through its accurate prediction of the location and orientation of a circular plate placed in front of seal whisker arrays. The model also generates temporal and spatial importance values of the signals, enabling the identification of significant temporal-spatial signal patterns crucial for the network’s predictions. These signal patterns are further correlated with flow structures, allowing for the identification of important flow features relevant for accurate prediction. The study provides insights into seal whiskers’ perception of complex underwater environments, inspiring advancements in underwater sensing technologies.
1 Introduction
Given the limitations of the current underwater navigation systems, there has been a growing interest in advancing underwater sensing capabilities. The increasing scientific, military, and commercial demands, along with the exploration of the uncharted underwater world, have been the driving force behind these developments. Underwater robots employ various navigation systems, including inertial, geophysical, and acoustic systems, to estimate their position, velocity, and acceleration (Stutters et al., 2008; Chutia et al., 2017). However, these systems rely heavily on seafloor maps to prevent collisions and have limited capabilitiesin surveilling, navigating, and maneuvering safely in unpredictable deep waters, while avoiding obstacles and hazardous environments (Kinsey et al., 2006; Leonard and Bahr, 2016).
To address these challenges, different types of sensors are utilized on underwater robots (Chutia et al., 2017; Elshalakani et al., 2020). Camera-based sensors, relying on vision, are commonly used for monitoring the nearby environment and object detection. However, their effectiveness is much lower in dark deep waters, necessitating the use of artificial light sources. This further reduces the sensor’s range and reveals its location, posing a significant disadvantage, particularly in military applications (Yang et al., 2006; Jiang et al. 2019; Jiang et al. 2020). In environments with poor water clarity, the images captured by these sensors suffer from low quality and cloudiness due to light dissipation (scattering) over long distances (Kröger, 2008; Lee et al., 2012). To overcome the limitations of vision-based sensors, sonar sensors are commonly employed to estimate object locations by transmitting acoustic waves (Akyildiz et al., 2004). However, similar to vision sensors, sonar sensors have drawbacks such as high-power consumption and the risk of revealing the sensor’s location (Akyildiz et al., 2004; Elshalakani et al., 2020). Furthermore, the use of sonar sensors can have detrimental effects on various underwater species by altering their natural habitats (Schrope, 2002).
In recent years, there has been significant research on exploring the sensing abilities of phocid seals to address challenges in underwater sensing. These seals possess remarkable whisker arrays that enable them to detect and distinguish hydrodynamic disturbances caused by potential prey. Studies have shown that seals can detect disturbances with speeds as low as 245 μm/s and frequencies ranging from 10 to 100 Hz (Dehnhardt et al., 1998). Behavior studies have demonstrated the seals’ capability to discern the shape and size of upstream objects, track the travel direction of vortex rings, detect miniature submarines, and even follow other seals, by only using their whisker arrays (Dehnhardt et al., 2001; Müller and Kuc, 2007; Wieskotten et al., 2010; 2011). Notably, seals can track objects at considerable distances and detect weak vortices from past movements (Müller and Kuc, 2007). These findings highlight the extraordinary sensitivity, accuracy, and intelligence of seal whisker arrays in detecting hydrodynamic cues. The remarkable sensing abilities of seal whiskers have sparked great interest and inspiration for applications in underwater navigation, object tracking, and object detection. Several biomimetic sensors has already been designed, fabricated and tested in the laboratories (Beam et al., 2013; Kottapalli et al., 2015; Eberhardt et al., 2016; Sayegh et al., 2022; Wang et al., 2022; Zheng et al., 2022; Liu et al., 2023).
Previous studies on the fundamental mechanism of seal whisker sensing have mostly focused on the geometric effects, revealing that the whisker’s unique undulated shape can suppress self-induced vibrations by disrupting the Kármán vortex street in the wake (Hanke et al., 2010; Hans et al., 2013; Beem, 2015; Kottapalli et al., 2015; Morrison et al., 2016). Our recent study (Liu et al., 2019b) further elucidated that the suppression of the vibration is primarily due to the 180° phase shift of the axes of the elliptical cross-sections of seal whisker, which generates stable three-dimensional hair-pin vortices in the wake. Additionally, Beem and Triantafyllou, (2015) found that seal whisker geometry is able to generate large-amplitude “slaloming” motions in vortical flows by extracting energy from passing vortices, thereby enhancing wake capturing sensitivity.
While these findings confirm the high sensitivity of seal whisker sensing, the fundamental mechanism of wake identification and tracking remains largely unknown. Seal whiskers are arranged in a stereotyped grid pattern known as the vibrissa array, and the resulting bend moments at the root of the whiskers collectively form sensory inputs. Recent studies have suggested that seals may correlate temporal-spatial patterns of whisker array signals with surrounding flow patterns for intelligent perception. Various flow features, such as highest velocities, velocity gradients, and wake spatial extension, have been proposed as potential cues for object perception (Wieskotten et al., 2011). Artificial whisker array experiments have also indicated that cross-correlation of bending signals can be used to detect vortices (Glick et al., 2021). Our previous computational studies have shown that whisker array signals can reflect the strength, timing, and trajectories of upstream vortex-induced jets (Liu et al., 2021). Machine-learning models have been used to infer the position of upstream objects based on whisker tip displacements (Elshalakani et al., 2020). However, many questions remain unanswered, including the correlation between mechanical whisker signals and external flow disturbances, the specific flow features seals can sense, and how they interpret flow features to perceive the environment. Further research is needed to unravel these mysteries and advance our understanding of seal whisker sensing.
In this study, we aim to develop a novel method that combines a computational fluid-structure interaction (FSI) model with an interpretable deep-learning model to investigate the fundamental mechanisms underlying seal whisker sensing. The novelty of the method lies in its ability to establish connections between crucial signal patterns, flow characteristics, and attributes of upstream obstacles, thereby enhancing our understanding of the intricate sensing mechanisms. The effectiveness of the method is demonstrated through its ability to accurately predict the location and orientation of a circular plate positioned in front of seal whisker arrays. To achieve this, we placed a circular plate within a free stream, located upstream of the seal’s head. Two whisker arrays were integrated onto the seal’s head, replicating the realistic location, structure, geometry, and orientation observed in reported data. A one-way FSI model was employed to compute the plate-induced wake and subsequent dynamics and signals of the whiskers, represented by the root bending moment. By systematically varying the plate’s location and orientation, we generated diverse wake characteristics and whisker signals.
We further developed and trained an interpretable neural network model to learn and accurately predict the location and orientation of the upstream plate based on the whisker signals. Importantly, the model produces temporal and spatial importance values, allowing for the identification of crucial temporal-spatial signal patterns contributing to the network’s predictions. We then correlated these patterns with flow structures, enabling the identification of important flow features for prediction. The outcomes of this study provide novel insights into seal whiskers’ ability of perceiving and interpreting complex underwater environments, which have the potential to advance the development of underwater sensing technologies.
The rest of the paper is organized as follows: Section 2 introduces the computational model including the governing equations and numerical method, whisker array model, and parametric simulations setup. Section 3 introduces the interpretable deep learning network, including the network architecture, input preparation and training configuration. Section 4 presents the results of parametric simulations and network prediction and interpretation as well as the connection between signals and flow features. Section 4 presents the summary of the results and general conclusion.
2 Computational model
2.1 Governing equations and numerical method
The flow field is modeled using unsteady incompressible Navier-Stokes equations:
where
The current study employs an in-house immersed boundary method based incompressible flow solver, as described by Mittal et al. (2008). The solver discretizes the spatial terms using a cell-centered collocated arrangement of the primitive variables
Each whisker is modeled as elastic structure and dynamics of whisker is governed by the Navier equation:
where
Upon solving the dynamic equation at each time step, the hydrodynamic loadings along each whisker are estimated based on the local flow speeds using a simplified drag-lift model. To achieve this, each individual whisker is discretized into a series of whisker segments, each one spanning one segment. The hydrodynamic force acting on each segment is computed using
2.2 Whisker arrays model
The seal whisker array model, depicted in Figure 1A, is adopted from our previous study (Liu et al., 2021), which closely replicates a real harbor seal’s whisker array. The model comprises of two arrays, each containing 44 individual whiskers, placed on the muzzle of a realistic harbor seal head model. The length, curvature, tapering, location, and orientation of each whisker are taken from the reported data of realistic harbor seal whisker arrays (Murphy, 2013; Graff, 2016) and are illustrated in Figures 1B, C. The undulated geometry of each whisker is constructed using a simplified elliptical cylinder model, superimposed with two sinusoidal undulations with a 180° phase shift of the axes of the elliptical cross-sections (Figure 1D) (Rinehart et al., 2017). The material properties of the whiskers are also determined using experimental data (Hans et al., 2014). For more detailed information, please refer to Liu et al. (2021).
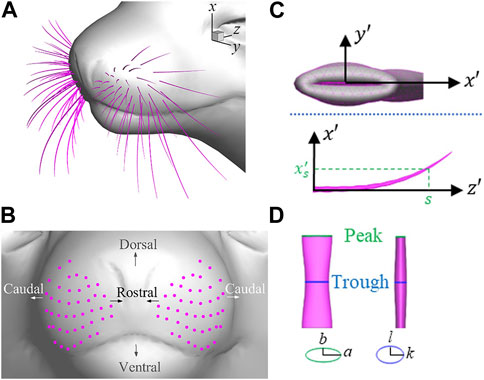
FIGURE 1. (A) Whisker array model of a harbor seal, (B) location of the base of each whisker, (C) Shape of an individual whisker, and (D) Side and front view of one wavelength segment of the seal whisker. The green and blue ellipses are the cross-section at the peak and the trough position, respectively.
2.3 Simulation setup and parametric space
Figure 2 illustrates the simulation setup. A circular plate with a diameter of 40 mm is placed 30 cm in front of the nasal tip of a seal head. This diameter is determined based on previous studies (Wieskotten et al., 2011; Liu et al., 2021) for generating typical-size vortices of prey. The seal head and the plate are immersed into a
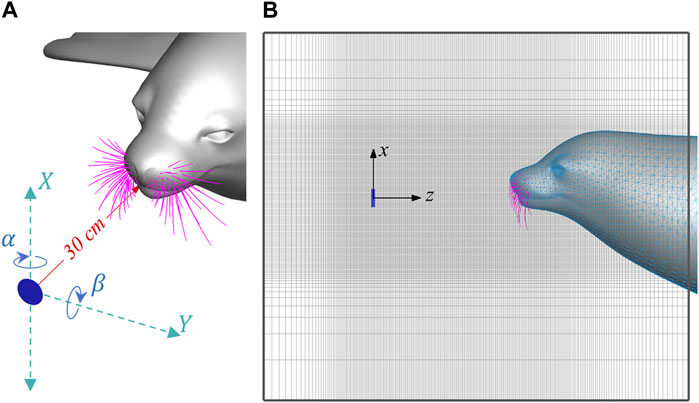
FIGURE 2. (A) Schematics of the upstream plate and X, Y, α and β parameters, and (B) Side view of the computational domain.
The whiskers are discretized using ten-node tetrahedral cells, resulting in a range of 9,700–34,100 cells depending on the length of the whiskers. The whiskers are fixed at the base, preventing any movement or rotation. The solid simulations are performed on the same XSEDE Expanse cluster, with one processor allocated for each whisker. Thus, for each parametric case that includes 88 whiskers, the average computational cost for the solid simulation is 184 CPU hours.
A group of parametric simulations were conducted by systematically varying the location (X, Y) and orientation (α, β) of the circular plate, as defined in Figure 2A. The ranges of these parameters were determined to ensure the wakes would intersect at least one of the whiskers. The selected parameters are: X from −120 mm to 120 mm with an interval of 60 mm, Y from 0 to 200 mm with an interval of 50 mm, with an interval of 50 mm, α from −45° to 45° with an interval of 22.5°, and β from −45° to 45° with an interval of 22.5°. This results in a total of 625 simulation cases.
3 Interpretable deep learning network
3.1 Network architecture
Vision Transformer (ViT) (Vaswani et al., 2017; Dosovitskiy et al., 2020) has demonstrated effective structural interpretation of spatial correlations between pixels in images by dividing raw images into multiple image patches and leveraging comprehensive imaging information. Flexibility of ViT to handle various input sequences allows for the utilization of video information in addition to image information. In this study, we propose a network architecture called Video Transformer (VT) as a straightforward extension of ViT, designed specifically for video processing. VT initially splits the video into spatio-temporal tokens and applies a self-attention model using spatial and temporal blocks to extract features based on spatial and temporal changes of moments. These features are then optimized by passing through transformer blocks to determine the target position and orientation. In this section, we first introduce ViT, followed by a discussion on tubelet embedding for video processing (Arnab et al., 2021), and finally, we present the self-attention model for video processing (Arnab et al., 2021).
3.1.1 ViT
Initially, ViT processes a 2D image,
where MLP consists of two layers with a GELU non-linearity (Hendrycks and Gimpel, 2016). Finally, an MLP is employed to classify the output of L transformer blocks, which can be expressed as:
3.1.2 Tubelet embedding
Given the flexibility of ViT to accommodate any sequence of input tokens
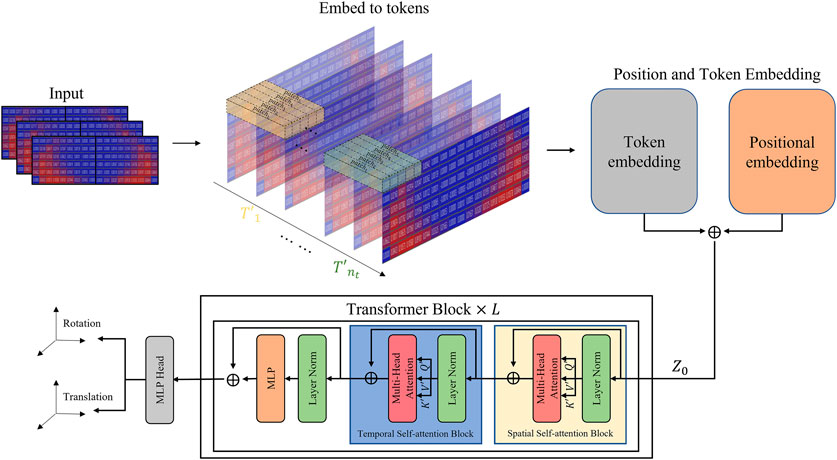
FIGURE 3. Video transformer network structure.
In this context, we define a tubelet with dimensions
3.1.3 Factorized self-attention model
The factorized self-attention model (Ho et al., 2019; Weissenborn et al., 2019; Bertasius et al., 2021) described in the transformer block of Figure 3 processes the spatio-temporal tokens
Specifically, the spatial self-attention block calculates the spatial relationship by reshaping the tokens
Unlike previous approaches, this model does not utilize a classification token since it can handle token reshaping across spatial and temporal dimensions without introducing ambiguities.
3.2 Network input preparation
The primary input signal for the network is derived from the time history of the bending moment at the base of each whisker. The total bending moment is decomposed in two components:
Tactile vibrissal systems in general possess two distinct types of mechanoreceptors in their whiskers, namely Merkel cells (SAI mechanoreceptors) and Pacinian corpuscles (PC mechanoreceptors), specialized in detecting static and dynamic changes (Zimmerman et al., 2014). Merkel cells are adept at sensing static stimuli, such as sustained pressure, while Pacinian corpuscles excel at perceiving dynamic changes, including vibrations and rapid movements. To emulate the underlying nerve system, we further decompose the bending moments into two components: time-averaged (DC) components, denoted as
The subsequent step involves a data transformation process aimed at consolidating the inputs from individual whiskers to generate a signal map, as depicted in Figure 4. Initially, the base locations of each whisker were extracted from the model and then topologically mapped onto a
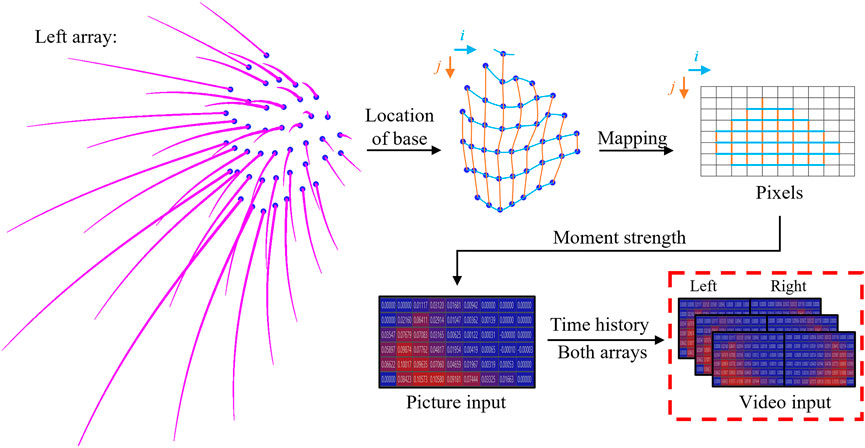
FIGURE 4. Data transformation process to generate the network signal map input from each individual whisker’s data.
3.3 Training configuration
The network training process involves regression training for 1,000 epochs using the Adam optimizer. A batch size of 25 and an initial learning rate of 0.001 are used, along with a multi-step learning rate scheduler. The input samples are divided into train and validation groups using the 5-fold method, with 500 samples for training and 125 samples for validation. Each video sample has a size of
The implementation of the method utilizes PyTorch 1.12.1 and Skorch 0.11.0 (Tietz et al., 2017) frameworks. The training process is performed using an NVIDIA GeForce RTX 2080 Ti GPU with 11GB memory.
4 Results and discussion
This section begins with a comprehensive examination of the influence of the plate’s location and orientation on both the flow field and whisker signals. Subsequently, the outcomes of the interpretable deep learning network are presented, including accuracy measurements and importance value maps of the signals. Lastly, the correlation between the importance value maps, whisker signals, and flow field is analyzed, while also investigating the underlying mechanisms. To facilitate the discussion, five exemplary cases are selected for a detailed investigation, which are summarized in Table 1.
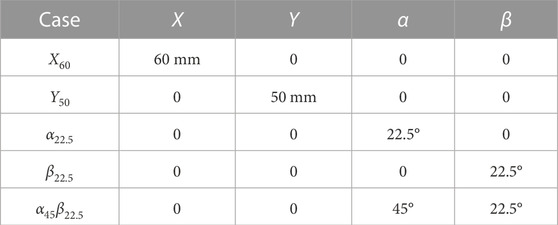
TABLE 1. Definition of the exemplary cases.
4.1 Characteristics of the flow field
Figure 5 shows the wake structure induced by the plate in the exemplary cases, determined by the
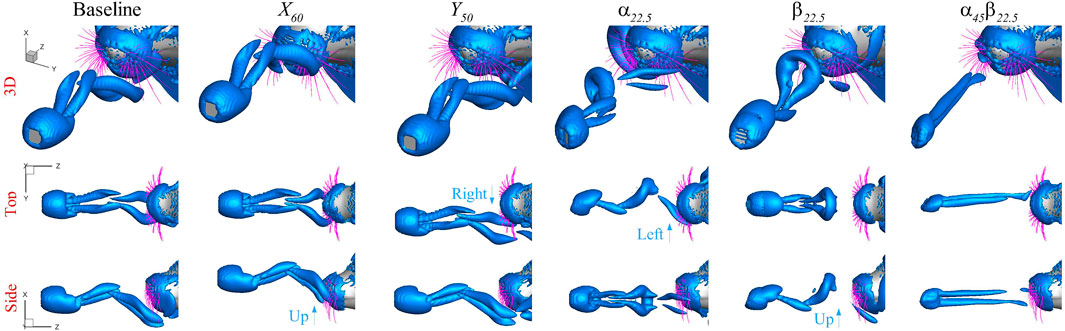
FIGURE 5. Plate induced wake structure determined by the
The vortex shedding frequency also changes with the orientation of the plate. To illustrate the relationship, Figure 6A presents the vortex shedding frequency in the parametric space of α and β. When vortex shedding is absent, a zero value is assigned. The plot reveals that the vortex shedding frequency increases symmetrically along both the α and β axes. When one orientation angle is at its maximum while the other is non-zero, no vortex shedding occurs. To account for the changes in both the vortex shedding frequency and the frontal area of the plate, the Strouhal number was calculated for each case using the formula
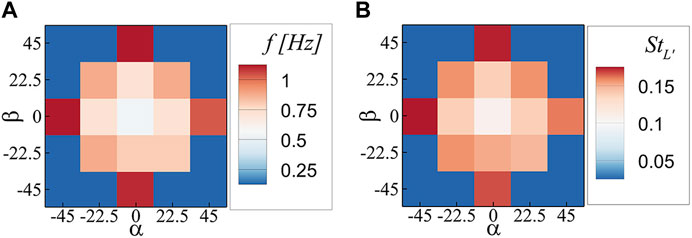
FIGURE 6. (A) Vortex shedding frequency, and (B) Strouhal number; in the parametric space of α and β.
4.2 Whisker signals
We partition the whisker arrays into four regions: LT (left top), LB (left bottom), RT (right top), and RB (right bottom) whiskers, and calculated the average signals,
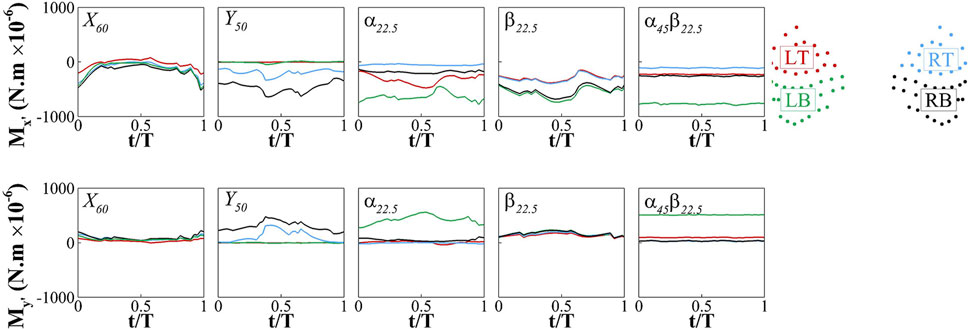
FIGURE 7. Phase-averaged time history of
Overall, the signals at the root of the whisker arrays exhibit temporal and spatial patterns that align with the dynamics of the vortices. There is a clear correlation between the arrival of the vortices and the temporal variation in the signals. The regions where the vortices directly impact the whiskers tend to exhibit stronger signal amplitudes. The distinct vortex dynamics in each case result in different temporal and spatial patterns of the signals, providing an explanation for the discernible differences among the cases.
4.3 Network results
The neural network was trained successfully to predict the location and orientation of the upstream plate. Figure 8A illustrates the best mean squared error (MSE) losses of the training and validation groups over 1,000 epochs, indicating the convergence of the network. The final MSE loss for the training and validation corresponding to the best fold is 0.0255 and 0.1064, respectively. The decision to limit the epochs to 1,000 was based on the observation that the drop in MSE loss for both groups during the last 100 epochs is less than 0.1%. The accuracy of the trained network corresponding to the best fold is depicted in Figure 8B through a comparison of the ground truth and predicted locations and orientations of the upstream plate in the validation cases. The accuracy metric is quantified using the following formula:
where
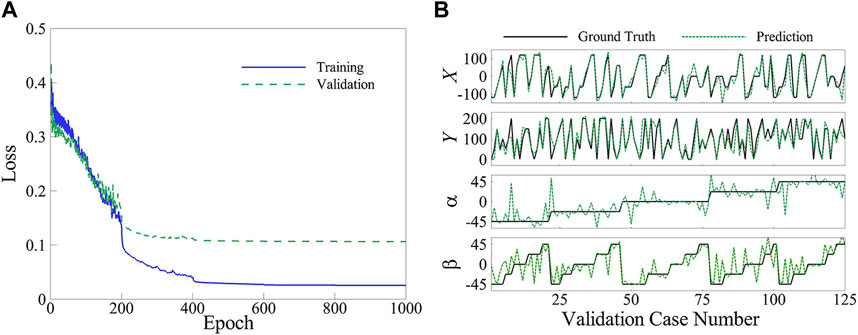
FIGURE 8. (A) Mean squared error loss of the training and validation groups, and (B) Ground truth and predicted values for the validation group.
An essential capability of the interpretable network is its ability to generate temporal and spatial importance of the signals within the trained network, allowing for the identification of significant temporal-spatial signal patterns that contribute to the network’s predictions. In our study, as outlined in Section 3.3, we utilized a temporal dimension of 80 and a time interval of t = 8, resulting in 10 distinct time patches for the prediction. In the
In Figure 9, we present the temporal importance (TI) analysis of the signals in each representative case. Except for the
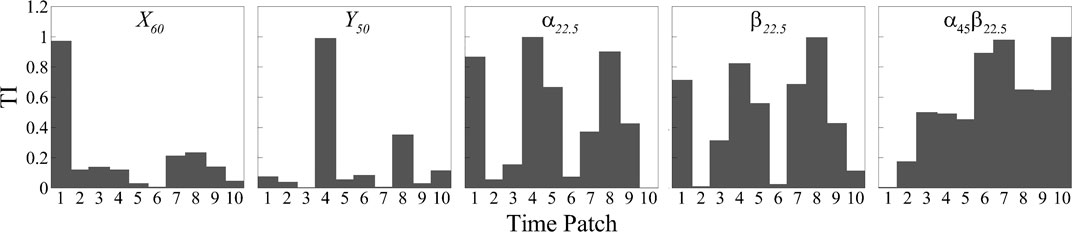
FIGURE 9. Temporal importance map of
The variation in temporal importance patterns highlights the importance of considering the temporal dynamics and their specific correlations with the underlying flow structures when interpreting the prediction process. To illustrate this relationship, we present the vortex structures determined by the
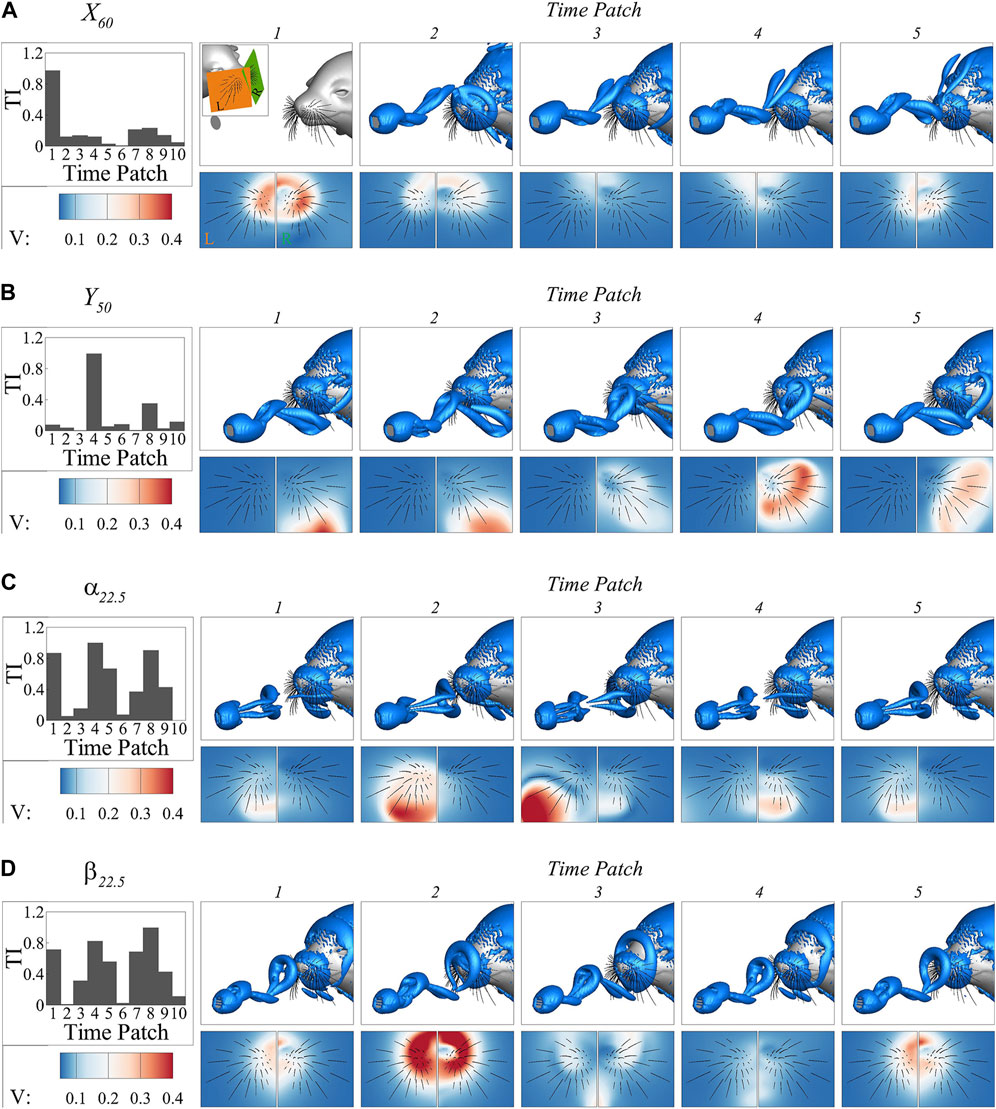
FIGURE 10. The vortex structures determined by the
In the
In the
In the case of
In the case of
The current interpretable deep learning network also provides the importance of each whisker through spatial importance (SI) maps at individual time patch. Figure 11 illustrates the locations of the whiskers with the highest importance values during each of the first five time patches in each case. These whisker locations generally exhibit a strong correlation with areas of strong flow disturbance, as depicted in Figure 10. Furthermore, the spatial distribution of these important whiskers provides insights into the location and size of the vortex rings in the wake. For instance, in the
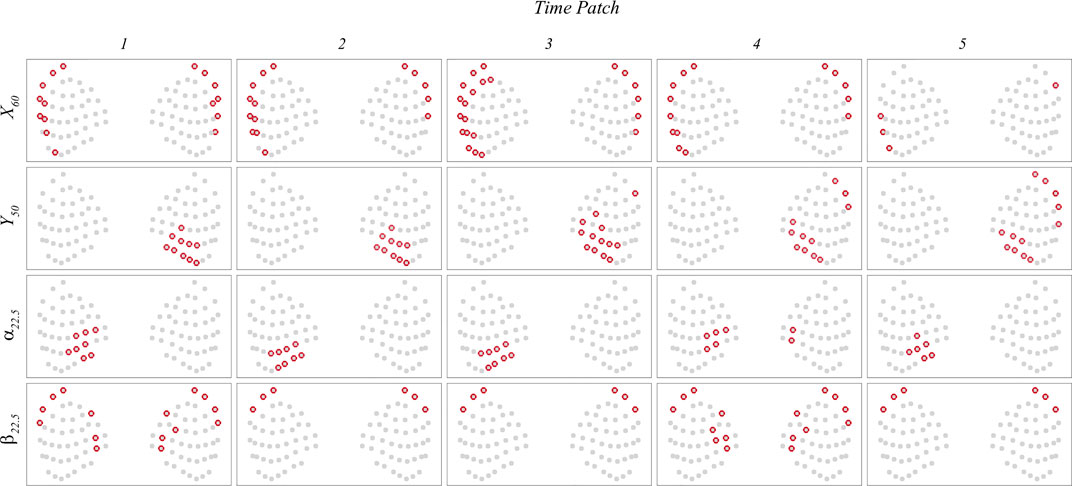
FIGURE 11. The locations of the whiskers with the highest importance values during each of the first five time patches in
To gain a deeper understanding of the signals used for prediction, Figures 12, 13 present the maps of
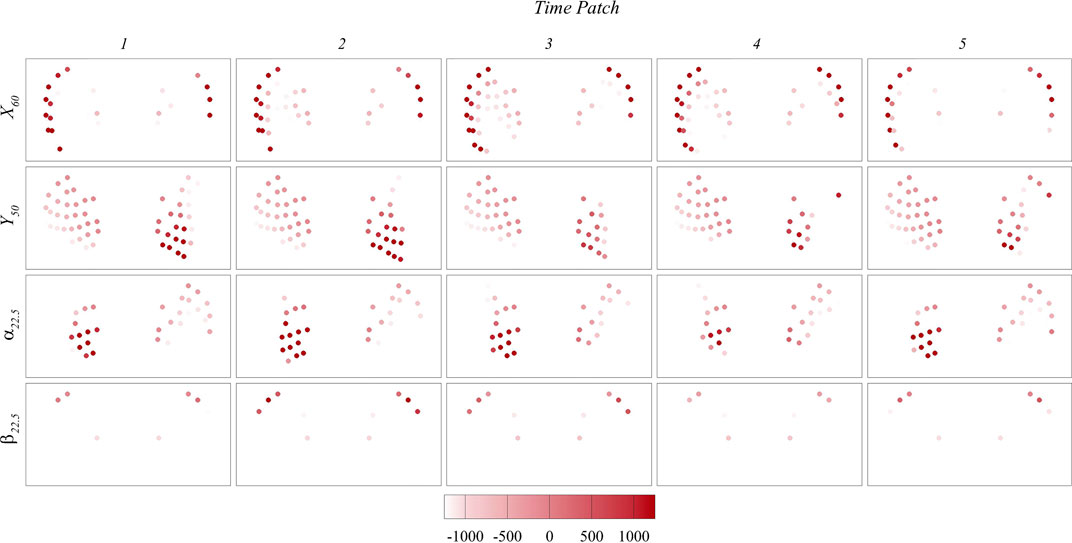
FIGURE 12. Contour maps of
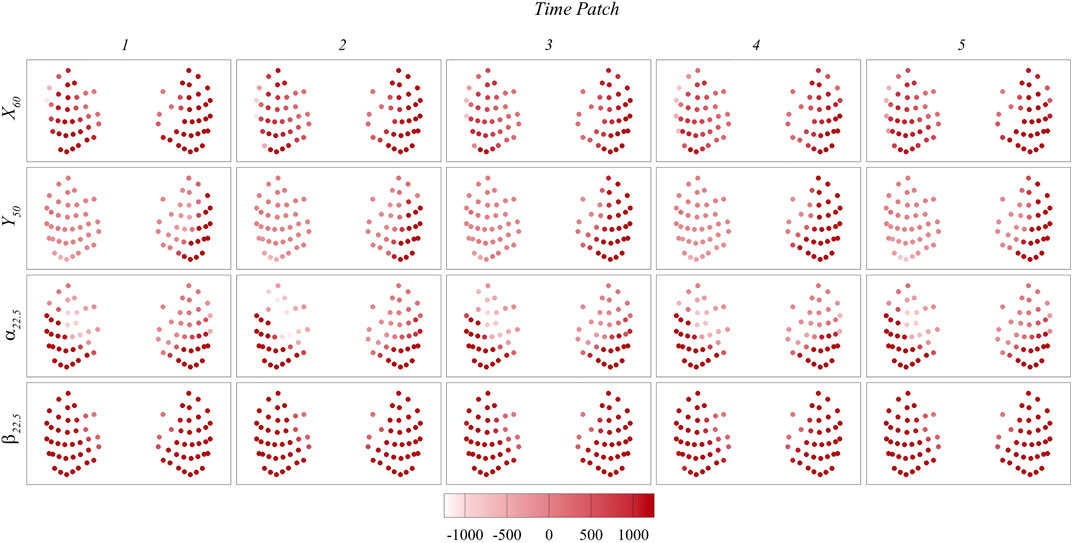
FIGURE 13. Contour maps of
5 Summary and conclusion
In this study, we developed a novel method that combines a computational fluid-structure interaction (FSI) model with interpretable deep-learning models to investigate the fundamental mechanisms of seal whisker sensing. We demonstrate that the method is able to accurately predict the location and orientation of a circular plate upstream of a seal’s head by using the bending moment signals at the root of whisker arrays. The algorithm also reveals crucial temporal-spatial signal patterns utilized in the prediction process, enabling establishing correlations between important signal patterns, flow features, and characteristics of upstream obstacles for enhanced understanding.
The FSI model couples an incompressible flow solver and structural dynamics solver in a one-way coupling manner. We generated diverse wake characteristics and corresponding whisker dynamics by placing a circular plate upstream of a seal’s head and systematically varying the location and orientation of the plate. Two types of wake patterns were observed. For most of the cases, the wake is characterized by two vortex rings shed from two edges of the plate with a phase difference. The translation of the upstream plate solely affects the position of the wake, without altering its structures. However, when the orientation of the plate is changed, it induces an inclination in the wake’s trajectory, a shift in the orientation of the vortices, and an increase of the vortex shedding frequency. Furthermore, the side farther from the inflow corresponds to the location of the stronger vortex, while the side closer to the inflow is associated with the weaker vortex. In contrast, for the cases where one of the orientation angles is the largest while the other is non-zero, no vortex shedding occurs in the wake. Instead, the wake is characterized by two streamwise vortex legs attached to the side of the plate. The whisker signals are represented by the bending moments at the root of each whisker. The signals exhibit temporal and spatial patterns that align with the dynamics of the vortices. There is a clear correlation between the arrival of the vortices and the temporal variation in the signals. The regions where the vortices directly impact the whiskers tend to exhibit stronger signal amplitudes.
We train the network model to learn the location and orientation of the upstream plate based on the whisker signals. The results show that the model is able to predict the location of the plate with the accuracy of 90%–92.0% and the orientation of the plate with the accuracy between 86% and 89%. The analysis on the temporal importance of the signals reveals that the prediction process primarily relies on the time instances when the wake vortices actively interact with the whiskers. When the vortices are at varying distances from the whiskers, such as when the upstream plate is translated, interactions with the vortices in close proximity to the whiskers have a more significant impact, while interactions with the distant vortex ring have minimal influence. In the context of predicting the orientation of the upstream plate, interactions with all vortices can be important. However, interactions involving the vortices that carry more orientation information tend to hold greater significance, even if these vortices do not cause the strongest disturbance and signals. It suggests that the vortices that provide crucial orientation cues play a crucial role in the prediction process, even if their influence on the overall flow disturbance and signal strength is not the most pronounced. Spatially, the whiskers in stronger flow disturbance areas are more important in the prediction. The spatial arrangement of the important whiskers not only captures the vortex ring structure in the wake, but also provides valuable information about the location and size of the rings. In the context of the signals used for prediction, our results suggest that the bending moment generated by the lift force plays a crucial role in the prediction process. The direction of the moment may also be important as important whiskers are all associated with the bending moment in one direction.
Taken together, we demonstrate that the developed interpretable neural network is able to accurately predict the location and orientation of the upstream object of seal whisker arrays. More importantly, it allows for the identification of significant temporal-spatial signal patterns that contribute to the network’s predictions, which can then be further correlated with flow structures, enabling the identification of the crucial flow features that are sensed for accurate prediction. These insights are crucial for the development of intelligent flow sensors capable of accurately perceiving and interpreting complex underwater environments.
We would like to acknowledge that our study utilized a simplified drag-lift model in our one-way coupling approach to estimate the hydrodynamic loadings along each whisker. This simplified model does not take into account the feedback effect of the whiskers on the surrounding flow. In reality, the presence of the whiskers would influence the surrounding flow, resulting in different drag and lift values that could potentially affect the outcomes of our research. However, we believe that the impact of the whiskers on the surrounding flow would be small due to the small diameter of the whiskers (with a mean diameter of 0.66 mm). This diameter is approximately 1-2 orders of magnitude smaller than the scale of the vortices in the wakes, which range from 20 mm to 120 mm. Considering this scale difference, we believe that the one-way coupling approach still provides valuable insights while significantly reducing the computational cost associated with conducting full FSI simulations. It is important to note that future investigations could explore the implications of the two-way coupling between the whiskers and the surrounding flow for a more comprehensive understanding of the hydrodynamic interactions.
In all the simulation cases, we observed that the frequency of the bending moments consistently matched the shedding frequency of the wake vortices. This behavior is a direct consequence of the one-way coupling approach, where the feedback effect of whisker dynamics on the flow was disregarded. It is worth noting, however, that in our previous research (Liu et al., 2019b), we discovered that the distinctive undulated geometry of seal whiskers can effectively mitigate vortex-induced vibration when exposed to steady flows. This unique characteristic enables the whiskers to better synchronize with the wake frequency. Nonetheless, it is important to acknowledge that further investigations are required to explore the interactions between vortex-induced vibration and wake-induced vibration. Understanding these complex dynamics will provide valuable insights into the combined effects and enhance our comprehension of the overall system behavior.
Finally, seal whisker arrays display distinct spatial grid patterns combined with variations in whisker length and orientation. These patterns are believed to play crucial functional roles in enhancing the seals’ sensing capabilities. For instance, it is believed that longer whiskers located on the caudal side of the array can provide a larger detection area, extending the reach and sensitivity to water movements. Moreover, the variation in whisker length and orientation may allow for high sensitivity in wide frequency range and flow direction. Further studies are needed to fully investigate these effects, particularly by comparing the sensing capabilities between different whisker arrangements. The current findings are limited to a single configuration, and more research is necessary to gain a comprehensive understanding.
Data availability statement
The raw data supporting the conclusion of this article will be made available by the authors, without undue reservation.
Author contributions
XZ and QX conceived the original study. XZ, QX, DL, and GL designed the study. DB conducted numerical simulation and data analysis. YW implemented and trained the neural network. DB, YW, GL, DL, QX, and XZ collectively wrote the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by NSF under grant number 214421. The computation was supported by XSEDE award TG-CTS180004 and MCH220042. This study also used the HPC systems of the UMS Advanced Computing Group.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Akyildiz, I. F., Pompili, D., and Melodia, T. (2004). Challenges for efficient communication in underwater acoustic sensor networks. ACM Sigbed Rev. 1 (2), 3–8. doi:10.1145/1121776.1121779
Arnab, A., Dehghani, M., Heigold, G., Sun, C., Lučić, M., and Schmid, C. (2021). “Vivit: A video vision transformer,” in Proceedings of the IEEE/CVF International Conference on Computer Vision, 6836–6846.
Baevski, A., and Auli, M. (2018). Adaptive input representations for neural language modeling. arXiv preprint arXiv:1809.10853.
Beam, H. R., Hildner, M., and Triantafyllou, M. (2013). Calibration and validation of a harbor seal whisker-inspired flow sensor. Smart Mat. Struct. 22, 014012. doi:10.1088/0964-1726/22/1/014012
Beem, H. R. (2015). Passive wake detection using seal whisker-inspired sensing. Woods Hole, MA: Massachusetts Institute of Technology.
Beem, H. R., and Triantafyllou, M. S. (2015). Wake-induced ‘slaloming’ response explains exquisite sensitivity of seal whisker-like sensors. J. Fluid Mech. 783, 306–322. doi:10.1017/jfm.2015.513
Bertasius, G., Wang, H., and Torresani, L. (2021). July Is space-time attention all you need for video understanding? In ICML 2. doi:10.48550/arXiv.2102.05095
Bodaghi, D., Jiang, W., Xue, Q., and Zheng, X. (2021a). Effect of supraglottal acoustics on fluid–structure interaction during human voice production. J. Biomechanical Eng. 143 (4), 041010. doi:10.1115/1.4049497
Bodaghi, D., Xue, Q., Zheng, X., and Thomson, S. (2021b). Effect of subglottic stenosis on vocal fold vibration and voice production using fluid–structure–acoustics interaction simulation. Appl. Sci. 11 (3), 1221. doi:10.3390/app11031221
Chen, J. M., and Fang, Y. C. (1996). Strouhal numbers of inclined flat plates. J. wind Eng. industrial aerodynamics 61 (2-3), 99–112. doi:10.1016/0167-6105(96)00044-x
Chutia, S., Kakoty, N. M., and Deka, D. (2017). A review of underwater robotics, navigation, sensing techniques and applications. Proc. Adv. Robotics, 1–6.
Dehnhardt, G., Mauck, B., and Bleckmann, H. (1998). Seal whiskers detect water movements. Nature 394 (6690), 235–236. doi:10.1038/28303
Dehnhardt, G., Mauck, B., Hanke, W., and Bleckmann, H. (2001). Hydrodynamic trail-following in harbor seals (Phoca vitulina). Science 293 (5527), 102–104. doi:10.1126/science.1060514
Devlin, J., Chang, M. W., Lee, K., and Toutanova, K. (2018). Bert: Pre-training of deep bidirectional transformers for language understanding. arXiv preprint arXiv:1810.04805.
Dosovitskiy, A., Beyer, L., Kolesnikov, A., Weissenborn, D., Zhai, X., Unterthiner, T., et al. (2020). An image is worth 16x16 words: Transformers for image recognition at scale. arXiv preprint arXiv:2010.11929.
Eberhardt, W. C., Wakefield, B. F., Murphy, C. T., Casey, C., Shakhsheer, Y., Calhoun, B. H., et al. (2016). Development of an artificial sensor for hydrodynamic detection inspired by a seal’s whisker array. Bioinspir. Biomim. 11, 056011. doi:10.1088/1748-3190/11/5/056011
Elshalakani, M., Muthuramalingam, M., and Bruecker, C. (2020). A deep-learning model for underwater position sensing of a wake’s source using artificial seal whiskers. Sensors 20 (12), 3522. doi:10.3390/s20123522
Geng, B., Xue, Q., Zheng, X., Liu, G., Ren, Y., and Dong, H. (2017). The effect of wing flexibility on sound generation of flapping wings. Bioinspiration biomimetics 13 (1), 016010. doi:10.1088/1748-3190/aa8447
Glick, R., Muthuramalingam, M., and Brücker, C. (2021). Fluid-structure interaction of flexible whisker-type beams and its implications for flow sensing by pair-wise correlation. Fluids 6 (3), 102. doi:10.3390/fluids6030102
Graff, M. M. (2016). “Mechanisms of whisker based flow sensing,”. Doctoral dissertation (Evanston, IL: Northwestern University).
Hanke, W., Witte, M., Miersch, L., Brede, M., Oeffner, J., Michael, M., et al. (2010). Harbor seal vibrissa morphology suppresses vortex-induced vibrations. J. Exp. Biol. 213 (15), 2665–2672. doi:10.1242/jeb.043216
Hans, H., Miao, J. M., and Triantafyllou, M. S. (2014). Mechanical characteristics of harbor seal (Phoca vitulina) vibrissae under different circumstances and their implications on its sensing methodology. Bioinspiration biomimetics 9 (3), 036013. doi:10.1088/1748-3182/9/3/036013
Hans, H., Miao, J., Weymouth, G., and Triantafyllou, M. (2013). “Whisker-like geometries and their force reduction properties,” in 2013 MTS/IEEE OCEANS-Bergen, Bergen, Norway, 10-14 June 2013 (IEEE), 1–7. doi:10.1109/OCEANS-Bergen.2013.6608113
Hendrycks, D., and Gimpel, K. (2016). Gaussian error linear units (gelus). arXiv preprint arXiv:1606.08415.
Ho, J., Kalchbrenner, N., Weissenborn, D., and Salimans, T. (2019). Axial attention in multidimensional transformers. arXiv preprint arXiv:1912.12180.
Jiang, Y., Ma, Z., and Zhang, D. (2019). Flow field perception based on the fish lateral line system. Bioinspiration biomimetics 14 (4), 041001. doi:10.1088/1748-3190/ab1a8d
Jiang, Y., Zhao, P., Ma, Z., Shen, D., Liu, G., and Zhang, D. (2020). Enhanced flow sensing with interfacial microstructures. Biosurface Biotribology 6 (1), 12–19. doi:10.1049/bsbt.2019.0043
Kinsey, J. C., Eustice, R. M., and Whitcomb, L. L. (2006). “September. A survey of underwater vehicle navigation: Recent advances and new challenges,” in IFAC conference of manoeuvering and control of marine craft 88. 1–12.
Kottapalli, A. G. P., Asadnia, M., Miao, J. M., and Triantafyllou, M. S. (2015). “January. Harbor seal whisker inspired flow sensors to reduce vortex-induced vibrations,” in 2015 28th IEEE International Conference on Micro Electro Mechanical Systems (MEMS) (IEEE), 889–892.
Kröger, R. (2008). “The physics of light in air and water,” in Sensory Evolution on the Threshold-Adaptations in Secondarily Aquatic Vertebrates (University of California Press), 113–119.
Lee, D., Kim, G., Kim, D., Myung, H., and Choi, H. T. (2012). Vision-based object detection and tracking for autonomous navigation of underwater robots. Ocean. Eng. 48, 59–68. doi:10.1016/j.oceaneng.2012.04.006
Leonard, J. J., and Bahr, A. (2016). Autonomous underwater vehicle navigation. Springer handbook of ocean engineering, 341–358.
Liu, G., Jiang, Y., Wu, P., Ma, Z., Chen, H., and Zhang, D. (2023). Artificial whisker sensor with undulated morphology and self-spread piezoresistors for diverse flow analyses. Soft Robot. 10 (1), 97–105. doi:10.1089/soro.2021.0166
Liu, G., Geng, B., Zheng, X., Xue, Q., Dong, H., and Lauder, G. V. (2019a). An image-guided computational approach to inversely determine in vivo material properties and model flow-structure interactions of fish fins. J. Comput. Phys. 392, 578–593. doi:10.1016/j.jcp.2019.04.062
Liu, G., Geng, B., Zheng, X., Xue, Q., Wang, J., and Dong, H. (2018). “An integrated high-fidelity approach for modeling flow-structure interaction in biological propulsion and its strong validation,” in 2018 AIAA Aerospace Sciences Meeting, 1543.
Liu, G., Jiang, W., Zheng, X., and Xue, Q. (2021). Flow-signal correlation in seal whisker array sensing. Bioinspiration Biomimetics 17 (1), 016004. doi:10.1088/1748-3190/ac363c
Liu, G., Xue, Q., and Zheng, X. (2019b). Phase-difference on seal whisker surface induces hairpin vortices in the wake to suppress force oscillation. Bioinspiration Biomimetics 14 (6), 066001. doi:10.1088/1748-3190/ab34fe
Mittal, R., Dong, H., Bozkurttas, M., Najjar, F. M., Vargas, A., and Von Loebbecke, A. (2008). A versatile sharp interface immersed boundary method for incompressible flows with complex boundaries. J. Comput. Phys. 227 (10), 4825–4852. doi:10.1016/j.jcp.2008.01.028
Morrison, H. E., Brede, M., Dehnhardt, G., and Leder, A. (2016). Simulating the flow and trail following capabilities of harbour seal vibrissae with the Lattice Boltzmann Method. J. Comput. Sci. 17, 394–402. doi:10.1016/j.jocs.2016.04.004
Müller, R., and Kuc, R. (2007). Biosonar-inspired technology: Goals, challenges and insights. Bioinspiration biomimetics 2 (4), S146–S161. doi:10.1088/1748-3182/2/4/s04
Murphy, C. T. (2013). Structure and function of pinniped vibrissae. Doctoral dissertation. St. Petersburg, FL: University of South Florida.
Rinehart, A., Shyam, V., and Zhang, W. (2017). Characterization of seal whisker morphology: Implications for whisker-inspired flow control applications. Bioinspiration Biomimetics 12 (6), 066005. doi:10.1088/1748-3190/aa8885
Sayegh, M., Daraghma, H., Mekid, S., and Bashmal, S. (2022). Review of recent bio-inspired design and manufacturing of whisker tactile sensors. Sensors 22 (7), 2705. doi:10.3390/s22072705
Schrope, M. (2002). Whale deaths caused by US Navy's sonar. Nature 415 (6868), 106. doi:10.1038/415106a
Stutters, L., Liu, H., Tiltman, C., and Brown, D. J. (2008). Navigation technologies for autonomous underwater vehicles. IEEE Trans. Syst. Man, Cybern. Part C Appl. Rev. 38 (4), 581–589. doi:10.1109/tsmcc.2008.919147
Tietz, M., Fan, T. J., Nouri, D., and Bossan, B. (2017). skorch: A scikit-learn compatible neural network library that wraps PyTorch.
Van Kan, J. J. I. M. (1986). A second-order accurate pressure-correction scheme for viscous incompressible flow. SIAM J. Sci. Stat. Comput. 7 (3), 870–891. doi:10.1137/0907059
Vaswani, A., Shazeer, N., Parmar, N., Uszkoreit, J., Jones, L., Gomez, A. N., et al. (2017). Attention is all you need. Adv. neural Inf. Process. Syst. 30. doi:10.5555/3295222.3295349
Wang, Q., Li, B., Xiao, T., Zhu, J., Li, C., Wong, D. F., et al. (2019). Learning deep transformer models for machine translation. arXiv preprint arXiv:1906.01787.
Wang, S., Xu, P., Wang, X., Zheng, J., Liu, X., Liu, J., et al. (2022). Scaling autoregressive video models, 97 (15), 107219. arXiv preprint arXiv:1906.02634.
Wieskotten, S., Dehnhardt, G., Mauck, B., Miersch, L., and Hanke, W. (2010). Hydrodynamic determination of the moving direction of an artificial fin by a harbour seal (Phoca vitulina). J. Exp. Biol. 213 (13), 2194–2200. doi:10.1242/jeb.041699
Wieskotten, S., Mauck, B., Miersch, L., Dehnhardt, G., and Hanke, W. (2011). Hydrodynamic discrimination of wakes caused by objects of different size or shape in a harbour seal (Phoca vitulina). J. Exp. Biol. 214 (11), 1922–1930. doi:10.1242/jeb.053926
Yang, Y., Chen, J., Engel, J., Pandya, S., Chen, N., Tucker, C., et al. (2006). Distant touch hydrodynamic imaging with an artificial lateral line. Proc. Natl. Acad. Sci. 103 (50), 18891–18895. doi:10.1073/pnas.0609274103
Zheng, X., Kamat, A. M., Cao, M., and Kottapalli, A. G. P. (2021). Creating underwater vision through wavy whiskers: A review of the flow-sensing mechanisms and biomimetic potential of seal whiskers. J. R. Soc. Interface 18 (183), 20210629. doi:10.1098/rsif.2021.0629
Zheng, X., Kamat, A. M., Krushynska, A. O., Cao, M., and Kottapalli, A. G. P. (2022). 3D printed graphene piezoresistive microelectromechanical system sensors to explain the ultrasensitive wake tracking of wavy seal whiskers. Adv.Funct. Mat. 32, 2207274. doi:10.1002/adfm.202207274
Keywords: bioinspired flow sensing, wake identification, seal whisker, interpretable machine learning, fluid-structure interaction
Citation: Bodaghi D, Wang Y, Liu G, Liu D, Xue Q and Zheng X (2023) Deciphering the connection between upstream obstacles, wake structures, and root signals in seal whisker array sensing using interpretable neural networks. Front. Robot. AI 10:1231715. doi: 10.3389/frobt.2023.1231715
Received: 30 May 2023; Accepted: 17 July 2023;
Published: 03 August 2023.
Edited by:
Xingwen Zheng, The University of Tokyo, JapanReviewed by:
Amar M. Kamat, University of Groningen, NetherlandsZhiqiang Ma, City University of Hong Kong, Hong Kong SAR, China
Copyright © 2023 Bodaghi, Wang, Liu, Liu, Xue and Zheng. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Qian Xue, cWlhbi5YdWVAcml0LmVkdQ==; Xudong Zheng, eHh6ZW1lQHJpdC5lZHU=
†These authors have contributed equally to this work
‡Present address: Geng Liu, Department of Physics and Engineering, University of Scranton, Scranton, PA, United States