 Shigeru Miyagawa1,2,3*
Shigeru Miyagawa1,2,3* Rob DeSalle4
Rob DeSalle4 Vitor Augusto Nóbrega5
Vitor Augusto Nóbrega5 Remo Nitschke6,7
Remo Nitschke6,7 Mercedes Okumura8
Mercedes Okumura8 Ian Tattersall9
Ian Tattersall9- 1Department of Linguistics and Philosophy, Massachusetts Institute of Technology, Cambridge, MA, United States
- 2Biosciences Institute, University of São Paulo, São Paulo, Brazil
- 3Research Center for Super-Smart Society, Seikei University, Tokyo, Japan
- 4American Museum of Natural History, Institute for Comparative Genomics, New York, NY, United States
- 5Department of Linguistics, University of São Paulo, São Paulo, Brazil
- 6Institute for the Interdisciplinary Study of Language Evolution, University of Zurich, Zurich, Switzerland
- 7Department of Linguistics, University of Arizona, Tucson, AZ, United States
- 8Laboratory of Human Evolutionary Studies, Department of Genetics and Evolutionary Biology, University of São Paulo, São Paulo, Brazil
- 9American Museum of Natural History, Division of Anthropology, New York, NY, United States
Recent genome-level studies on the divergence of early Homo sapiens, based on single nucleotide polymorphisms, suggest that the initial population division within H. sapiens from the original stem occurred approximately 135 thousand years ago. Given that this and all subsequent divisions led to populations with full linguistic capacity, it is reasonable to assume that the potential for language must have been present at the latest by around 135 thousand years ago, before the first division occurred. Had linguistic capacity developed later, we would expect to find some modern human populations without language, or with some fundamentally different mode of communication. Neither is the case. While current evidence does not tell us exactly when language itself appeared, the genomic studies do allow a fairly accurate estimate of the time by which linguistic capacity must have been present in the modern human lineage. Based on the lower boundary of 135 thousand years ago for language, we propose that language may have triggered the widespread appearance of modern human behavior approximately 100 thousand years ago.
1 Introduction
More than any other trait, language defines us as human. Yet there is no clear agreement on when this crucial feature emerged in our evolution. Some who have studied the archaeological record suggest that language emerged in our lineage around 100 thousand years ago (kya) (Tattersall, 2012, 2017, 2018; Wadley, 2021), while others have claimed that some form of language preceded the emergence of modern humans (Albessard-Ball and Balzeau, 2018; Botha, 2020). Indeed, it has been argued [e.g., by Progovac (2016) and Dediu and Levinson (2018)] that language is not uniquely the property of the lineage that produced H. sapiens. Here we accept the reasoning of that behaviors compatible with language and the consistent exercise of symbolic thinking are detectable only in the archaeological record of H. sapiens (Tattersall, 2012; Berwick et al., 2013; Berwick and Chomsky, 2016), and approach the issue of the antiquity of language in our species by showing that, although it is not yet possible to identify the time when a linguistic capacity emerged, genomic evidence allows us to establish with reasonable certainty the latest point at which it must have been present in early H. sapiens populations.
Over the past 15 years, numerous studies have addressed the question of exactly when the first division occurred in the original stem population of early H. sapiens. While those studies do not tell us exactly when language emerged, they allow us to make a reasonable estimate of the lower boundary of the possible time range for this key occurrence. H. sapiens emerged as an anatomically distinctive entity by about 230kya (Vidal et al., 2022). Sometime after that speciation event, the first division occurred, with all descendant populations of that division having full-fledged language. From this universal presence of language, we can deduce that some form of linguistic capacity must have been present before the first population divergence. If the linguistic capacity had emerged in humans after the initial divergence, one would expect to find modern human populations that either do not have language, or that have some communication capacity that differs meaningfully from that of all other human populations. Neither is the case. The 7,000 or so languages in the world today share striking similarities in the ways in which they are constructed phonologically, syntactically, and semantically (Eberhard et al., 2023).
Genomic studies of early H. sapiens population broadly agree that the first division from the original stem is represented today by the Khoisan peoples of Southern Africa (Schlebusch et al., 2012). This conclusion was reached early on Vigilant et al. (1989), Knight et al. (2003), Tishkoff et al. (2007), and Veeramah et al. (2012), and it has more recently been bolstered by studies using newer genomic techniques (Fan et al., 2019; Lorente-Galdos et al., 2019; Schlebusch et al., 2017; Schlebusch et al., 2020; Pakendorf and Stoneking, 2021). The term “Khoisan” refers to a bio-genetic affiliation that is linked both to a proposed ancestor-group and to some modern peoples, living in present-day South Africa, who include modern speakers of the Khoe-Khwadi, Tuu, and Ju-ǂHoan languages that have some genetic affiliation to the first divergence of the human population (Güldemann and Sands, 2009; du Plessis, 2014). It follows that, if we can identify when the first division occurred, we can with reasonable certainty consider that date to define the lower boundary of when human language was present in the ancestral modern human population. Based on the results of studies focusing on whole genome single nucleotide polymorphisms (SNPs), we estimate that this first division occurred at approximately 135kya. 1
Huybregts (2017) was the first to attempt to pinpoint the timing of the first division in this way. Although he suggested a date of ~125kya, close to our estimate of ~135kya, his estimate was necessarily based on a fairly narrow set of studies showing a remarkably variable range. The studies he examined ranged from the clearly implausible 300kya (Scally and Durbin, 2012), to 180kya (Rito et al., 2013) and as little as 100kya (Schlebusch et al., 2012). Pakendorf and Stoneking (2021) later listed several studies proposing that the first division was older than 160kya (Fan et al., 2019; Lorente-Galdos et al., 2019; Schlebusch et al., 2020), along with four others, from 140 to 110kya, that overlapped with the range suggested by Huybregts (Gronau et al., 2011; Veeramah et al., 2012; Mallick et al., 2016; Song et al., 2017). Several newer studies now allow us to approach the age of the first division with greater precision.
2 Divergence time meta-analysis
The literature was searched with combinations of the terms “Khoisan,” “divergence time,” “DNA” and “molecular clock.” We considered only published literature (excluding biorXiv and thesis documents). We also excluded any publication that we deemed a review of previously published work. In this way we obtained 15 publications that contained time estimates for the divergence time of the Khoisan lineage from other human lineages. Publication dates ranged from 2007 to 2023 (Table 1). For each publication we recorded the upper and lower time estimates of divergence, the sample size (both number of individuals and number of populations used, when possible), whether the marker used was maternal (mtDNA), paternal (Y chromosome) or autosomal. In addition, we briefly recorded the informatics methodology that was used to make the estimates. We use a median value of divergence time (Table 2) to summarize the 15 studies for age of the African lineages.
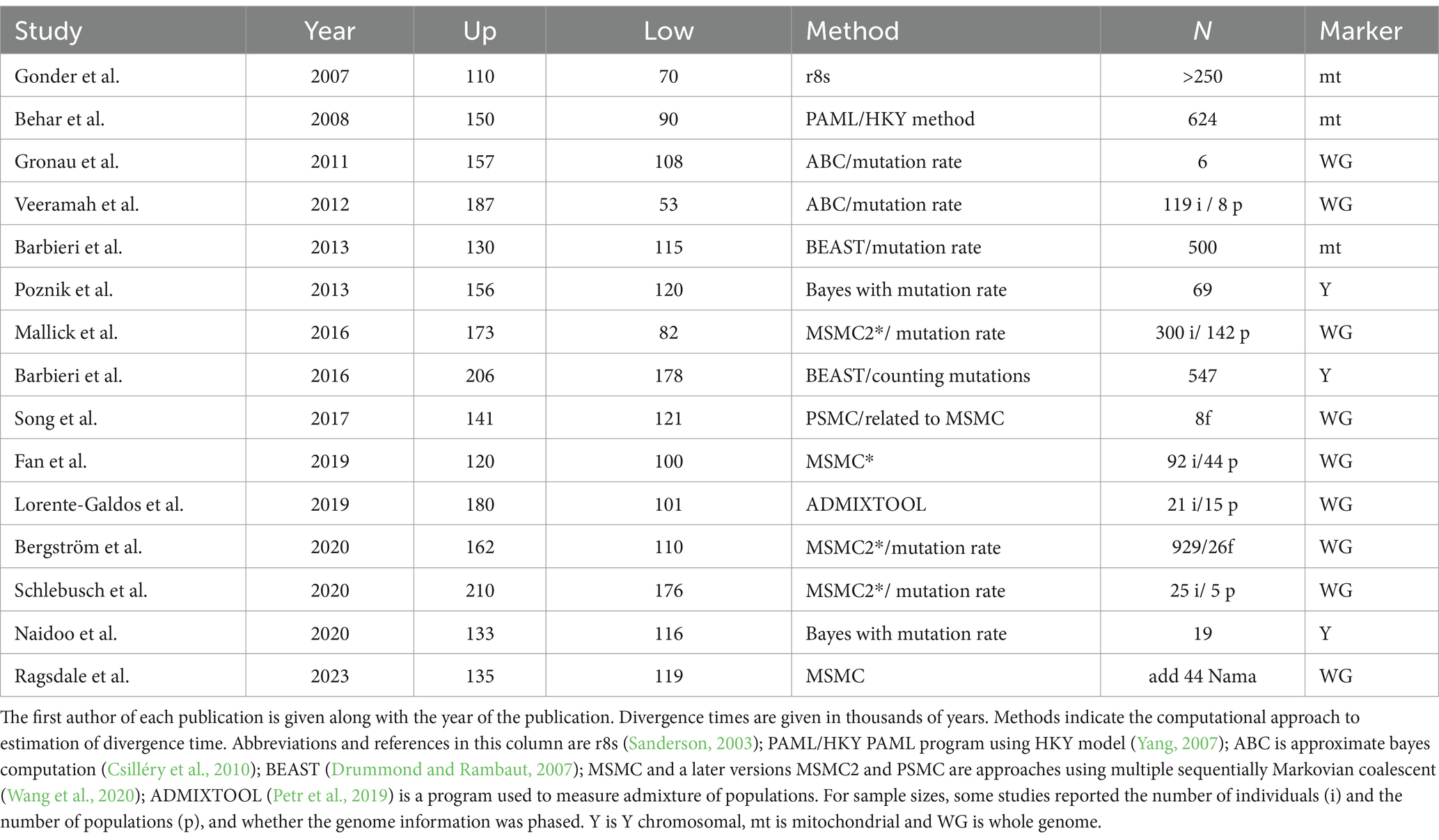
Table 1. Summary of estimates of divergence times for Khoisan lineage.
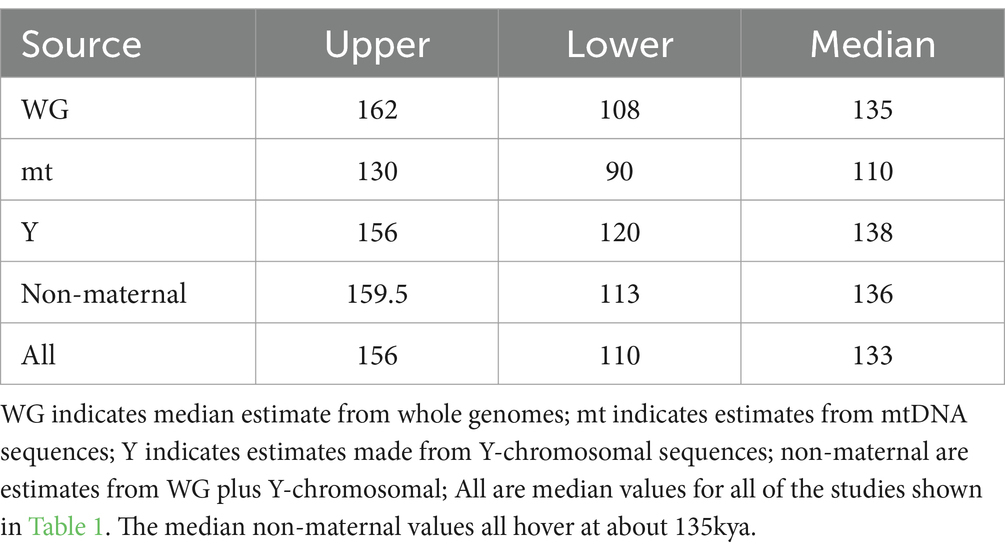
Table 2. Median estimates for Khoisan divergence.
2.1 Results
To attempt to bring more clarity to the question, we looked at a significantly larger body of work than was available to Huybregts. Estimates of the divergence times of early human populations using molecular markers have been made since the 1980s, when molecular data first began to be available for human evolution research (Soodyall and Jenkins, 1992; Barbujani, 1997; Scozzari et al., 1999; Seielstad et al., 1999; Harris and Hey, 1999; Zhivotovsky et al., 2003). Most of those estimates were made using single genes, or fragments of single genes, or microsatellites. Here we focus on estimates made using whole genome SNPs data, which came onto the scene post-2005. All works are listed in Table 1.
2.1.1 Markers used
These studies vary with respect to the markers used. On the one hand, there are studies using the uniparentally inherited Y chromosome (Y) and mitochondrial DNA (mt) markers; and on the other there are those that use whole genome data. Irrespective of study type, the methods used to determine divergence times are almost exclusively Bayesian, the most precise of them using the recently developed Multiple Sequentially Markovian Coalescent (MSMC) methodology (otherwise known as a “stairway” approach, since it results in a diagram that resembles a stairway). Table 1 summarizes 15 studies (three mt, three Y, and nine whole genome). With the caveats that the estimates of divergence times in those studies were made both with different markers and using different statistical methods for time estimation, we can use them to make an upper and lower boundary for the Khoisan divergence event. Upper estimates of the initial Khoisan divergence from other populations range from 110kya to 210kya, while lower estimates range from 53kya to 178kya. And while these distributions overlap considerably, we can make a fairly replicable estimate of divergence from the data in Table 1, using median values to arrive at ranges and an overall estimate for the divergence time.
2.1.2 Median estimates
The median estimates of divergence for the Khoisan lineage are shown in Table 2. The estimate from whole genome comparisons alone has a median value of 136kya (±23kya). The discrepancy previously noted between male and female lineage divergences (Wilder et al., 2004; Lippold et al., 2014) is also observed here (mt median divergence = 110kya ± 30kya, versus Y chromosomal median divergence = 138kya ±18kya).
While some imprecision in the molecular clock data cannot be eliminated with current techniques, the agreement of those median estimates based on non-maternal markers is clear. It is reasonable from these median estimates to conclude that the original divergence of the Khoisan lineage took place at about 135kya (± 20kya), with the divergence times of other African language lineages lying subsequent to this time.
3 Language as a trigger for modern human behavior
These genomic studies of early H. sapiens suggest that linguistic potential must have been present in the H. sapiens population at the latest by 135kya.
At present, we cannot go back further to pinpoint the date by which language itself emerged. What we can do is to look forward and see how, subsequent to 135kya, language may have had a direct hand in shaping modern human behaviors. To do this we focus on the widespread appearance of suggestive symbolically mediated behaviors, such as the Blombos cave ochres with regular-pattern incisions (Henshilwood et al., 2004; Henshilwood et al., 2009) and the geometric patterns engraved on ostrich eggshells found at two nearby locations, the Diepkloof (Texier et al., 2010; Texier et al., 2013) and Klipdrift Shelters (Henshilwood et al., 2014). While complex behaviors such as burial of the dead and occasional bodily decoration do appear to have occurred sporadically among Neanderthals and other extinct hominins (Frayer et al., 2006; García-Diez et al., 2013; Rodriguez-Vidal et al., 2014; Peresani et al., 2014; Joordens et al., 2015; Radovčić et al., 2015; Hoffmann et al., 2018; Majkić et al., 2018a, 2018b), it appears to have been only within H. sapiens, and subsequent to about around 100kya, that such behaviors eventually became systematized across the population (Tattersall, 2012, 2017, 2018; Wadley, 2021). The indicators concerned include such features as the use of pigments (Henshilwood et al., 2011), the employment of pierced marine shells for ornaments and body decorations (Henshilwood et al., 2004; Vanhaeren et al., 2006; d’Errico et al., 2009a), the engraving of non-figurative motifs, complex technologies (Powell et al., 2009; Grove, 2016) and ultimately the earliest representational objects (Henshilwood et al., 2018).
While we find sporadic occurrences of analogous behaviors somewhat earlier in human evolution, and in different contexts (Rodriguez-Vidal et al., 2014; Peresani et al., 2014; Joordens et al., 2015; Radovčić et al., 2015; Hoffmann et al., 2018; Li et al., 2019), it is only after around 100kya that we see such behaviors become routine and normalized in H. sapiens (Tattersall, 2012, 2017, 2018; Wadley, 2021). This suggests that linguistic capacity was fully in place before the widespread and normalized appearance of modern human behavior.
What might the role of language have been in the formation and spreading of modern human behavior? Huybregts (2017) argues that, while the linguistic capacity was present in H. sapiens prior to 125kya, it only existed as a system strictly internal to our mind, as suggested by Chomsky (2010, 2016), and did not become externalized as a communication system until sometime later, which puts it in the timeline of those other modern human behaviors. The ~77kya Blombos engraved ochres have similarly been considered as proxies for language, and not necessarily to have been preceded by it (Henshilwood and Marean, 2003, 2006; d’Errico et al., 2009b; Henshilwood and Dubreuil, 2009; Henshilwood and d'Errico, 2011). Some have suggested that language is a prerequisite for symbolically-mediated behavior (Henshilwood et al., 2004; Henshilwood and Dubreuil, 2009).
We wish to note the specific role that language may have played in organizing, and hence systematizing, modern behavior. Our proposal is similar to earlier suggestions by Henshilwood and others, but is based on a concrete and verifiable date of approximately 135 kya as the lower boundary for the presence of language. As the most complex communication tool yet devised in nature, it had a direct and enormous impact on all facets of human life. Language, with its complex system of mental representations and rules for combining them, is able to create new ways to connect existing symbols and predict new ways of behavior. This is, perhaps, what we see in the time gap between the lower boundary of 135kya for language, and the beginnings of the emergence of rich and normative symbolic behavior starting around 100kya. A way to interpret this gap is that language was central in organizing and systematizing modern human behavior.
As an alternative to the view that language was the trigger for modern human behavior (Tattersall, 2012, 2018), a number of archaeologists favor the view of an incremental, cumulative assembly of such modern human behavior as symbolic material culture, technological diversification, the use of diverse raw materials, and use of extensive social networks (Scerri and Will, 2023). One way to combine this view with ours is to consider language as the trigger that both accelerated and consolidated all these gradual processes that were taking place over the Middle Stone Age. One clear point of agreement with these scholars is that all believe that modern human behaviors emerged within Middle to Late Pleistocene in Africa.
4 The picture that emerges
Based on the recent genetic studies of early H. sapiens, we have pinpointed approximately 135kya as the moment at which some linguistic capacity must have been present in the human population. Looking forward from this event, modern human behaviors such as body decoration and the production of ochre pieces with symbolic engravings appeared as normative and persistent behaviors around 100kya. We believe that the time lag implied between the lower boundary of when language was present (135kya) and the emergence of normative modern human behaviors across the population suggests that language itself was the trigger that transformed nonlinguistic early H. sapiens (who nonetheless already possessed “language-ready” brains acquired at the origin of the anatomically distinctive species) into the symbolically-mediated beings familiar today. This development of the most sophisticated communication device in evolution allowed our ancestors to accelerate and consolidate symbolically-mediated behaviors until they became the norm for the entire species.
Data availability statement
Publicly available datasets were analyzed in this study. This data can be found at: the data utilized in this manuscript is available in the set of articles explored. In the manuscript, you can find all relevant references to the publications where the data is presented.
Author contributions
SM: Conceptualization, Formal analysis, Funding acquisition, Investigation, Project administration, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. RD: Data curation, Formal analysis, Investigation, Methodology, Resources, Validation, Visualization, Writing – original draft. VN: Formal analysis, Investigation, Methodology, Resources, Validation, Visualization, Writing – review & editing. RN: Investigation, Methodology, Writing – review & editing. MO: Formal analysis, Investigation, Methodology, Writing – review & editing. IT: Formal analysis, Investigation, Methodology, Writing – original draft.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. SM and VN are funded by the São Paulo Research Foundation (FAPESP), grant numbers 2018/18900-1 and 2023/03196-5, respectively.
Acknowledgments
We would like to thank the reviewer for their valuable comments and remarks on this article, which have helped us enhance its scope and address key issues.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Footnotes
1. ^It is important to note that in the current debate on human language evolution, at least two major perspectives are considered across different disciplines engaged with this topic. The first is the gradualist view, which sees the modern complexity of human language as the result of an incremental evolutionary process (Pinker and Bloom, 1990; Jackendoff, 1999, 2002; Tallerman, 2014, 2017; Progovac, 2015, Martins and Boeckx, 2019, a.o.). The second is the saltational view, which posits that most components of present-day linguistic capacity were exapted and assembled in a sudden, catastrophic event, giving rise to a cognitive faculty whose complexity did not emerge from gradual tinkering over eons (Bickerton, 1990, 1998; Berwick, 1998; Chomsky, 2002, 2005, 2016; Di Sciullo, 2013; Nóbrega and Miyagawa, 2015; Watanabe et al., 2017; Berwick and Chomsky, 2016, 2019, 2011, a.o.). For a comprehensive overview of these two perspectives, see Progovac (2019). The empirical implications of this article, however, do not allow for a definitive determination of which approach is on the right track. Rather, it provides insight into the earliest possible boundary by which language was available, without necessarily committing to the specific evolutionary changes that led to its emergence. In any case, any proposal suggesting that linguistic capacity emerged later than 135kya (e.g., Klein and Edgar, 2002; Chomsky, 2005) appears, in our view, highly implausible.
References
Albessard-Ball, L., and Balzeau, A. (2018). Of tongues and men: a review of morphological evidence for the evolution of language. J. Lang. Evol. 3, 79–89. doi: 10.1093/jole/lzy001
Barbujani, G. (1997). DNA variation and language affinities. Am. J. Hum. Genet. 61, 1011–1014. doi: 10.1086/301620
Berwick, R. C. (1998). “Language evolution and the minimalist program: the origins of syntax” in Approaches to the evolution of language: social and cognitive bases. eds. J. R. Hurford, M. Studdert-Kennedy, and C. Knight (Cambridge: Cambridge University Press), 320–340.
Berwick, R., and Chomsky, N. (2011). “The biolinguistic program. The current state of its development” in The Biolinguistic Enterprise: New perspectives on the evolution and nature of the human language faculty, ed. By A. M. Di Sciullo and C. Boeckx (Oxford: Oxford University Press), 19–41.
Berwick, R. C., and Chomsky, N. (2016). Why only us: language and evolution. Cambridge, MA: MIT Press.
Berwick, R. C., and Chomsky, N. (2019). All or nothing: no half-merge and the evolution of syntax. PLoS Biol. 17:e3000539. doi: 10.1371/journal.pbio.3000539
Berwick, R. C., Hauser, M. D., and Tattersall, I. (2013). Neanderthal language? Just-so stories take center stage. Front. Psychol. 4:671. doi: 10.3389/fpsyg.2013.00671
Bickerton, D. (1998). “Catastrophic evolution: the case for a single step from protolanguage to full human language” in Approaches to the evolution of language: Social and cognitive bases. eds. J. R. Hurford, M. Studdert-Kennedy, and C. Knight (Cambridge: Cambridge University Press), 341–358.
Botha, R. (2020). Neanderthal language: Demystifying the linguistic powers of our extinct cousins. Cambridge: Cambridge University Press.
Chomsky, N. (2005). Three factors in language design. Linguist. Inq. 36, 1–22. doi: 10.1162/0024389052993655
Chomsky, N. (2010). “Some simple evo devo theses: how true might they be for language?” in The evolution of human language: Biolinguistic perspectives. eds. R. K. Larson, V. Déprez, and H. Yamakido (Cambridge: Cambridge University Press), 45–62.
Csilléry, K., Blum, M. G. B., Gaggiotti, O. E., and François, O. (2010). Approximate Bayesian computation (ABC) in practice. Trends Ecol. Evol. 25, 410–418. doi: 10.1016/j.tree.2010.04.001
d’Errico, F., Vanhaeren, M., Barton, N., Bouzouggar, A., Mienis, H. K., Richter, D., et al. (2009a). Additional evidence on the use of personal ornaments in the middle Paleolithic of North Africa. Proc. Natl. Acad. Sci. U. S. A. 106, 16051–16056. doi: 10.1073/pnas.0903532106
d’Errico, F., Vanhaeren, M., Henshilwood, C. S., Lawson, G., Maureille, B., Gambier, D., et al. (2009b). “From the origin of language to the diversification of languages: what can archaeology and paleoanthropology say?” in Becoming eloquent: advances in the emergence of language, human cognition, and modern cultures. eds. F. d’Errico and J.-M. Hombert (Amsterdam: John Benjamins), 13–68.
Dediu, D., and Levinson, S. C. (2018). Neanderthal language revisited: not only us. Curr. Opin. Behav. Sci. 21, 49–55. doi: 10.1016/j.cobeha.2018.01.001
Di Sciullo, A. M. (2013). Exocentric compounds, language and proto-language. Lang. Inf. Soc. 20, 1–25. doi: 10.29211/soli.2013.20..001
Drummond, A. J., and Rambaut, A. (2007). BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol. Biol. 7, 214–218. doi: 10.1186/1471-2148-7-214
du Plessis, M. (2014). The damaging effects of romantic mythopoeia on Khoesan linguistics. Crit. Arts 28, 569–592. doi: 10.1080/02560046.2014.929217
Eberhard, D. M., Simons, G. F., and Fennig, C. D. (2023). Ethnologue: Languages of the world. 26th Edn. Dallas, TX: SIL International.
Fan, S., Kelly, D. E., Beltrame, M. H., Hansen, M. E. B., Mallick, S., Ranciaro, A., et al. (2019). African evolutionary history inferred from whole genome sequence data of 44 indigenous African populations. Genome Biol. 20, 82–14. doi: 10.1186/s13059-019-1679-2
Frayer, D. W., Orschiedt, J., Cook, J., Russell, M. D., and Radovčić, J. (2006). Krapina 3: cut Marks and ritual behavior? Period. Biol. 108, 519–524.
García-Diez, M., Fraile, B. O., and Maestu, I. B. (2013). Neanderthal graphic behavior: the pecked pebble from Axlor Rockshelter (Northern Spain). J. Anthropol. Res. 69, 397–410. doi: 10.3998/jar.0521004.0069.307
Gronau, I., Hubisz, M. J., Gulko, B., Danko, C. G., and Siepel, A. (2011). Bayesian inference of ancient human demography from individual genome sequences. Nat. Genet. 43, 1031–1034. doi: 10.1038/ng.937
Grove, M. (2016). Population density, mobility, and cultural transmission. J. Archaeol. Sci. 74, 75–84. doi: 10.1016/j.jas.2016.09.002
Güldemann, T., and Sands, B. (2009). “What click languages can and Can’t tell us about language origins” in The cradle of language. eds. R. Botha and C. Knight (Oxford: Oxford University Press), 204–218.
Harris, E. E., and Hey, J. (1999). X chromosome evidence for ancient human histories. Proc. Natl. Acad. Sci. U. S. A. 96, 3320–3324. doi: 10.1073/pnas.96.6.3320
Henshilwood, C. S., d’Errico, F., van Niekerk, K., Coquinot, Y., Jacobs, Z., Lauritzen, S. E., et al. (2011). A 100,000-year-old ochre-processing workshop at Blombos cave, South Africa. Science 334, 219–222. doi: 10.1126/science.1211535
Henshilwood, C. S., d’Errico, F., van Niekerk, K., Dayet, L., Queffelec, A., and Pollarolo, L. (2018). An abstract drawing from the 73,000-year-old levels at Blombos cave, South Africa. Nature 562, 115–118. doi: 10.1038/s41586-018-0514-3
Henshilwood, C. S., d’Errico, F., Vanhaeren, M., van Niekerk, K., and Jacobs, Z. (2004). Middle stone age Shell beads from South Africa. Science 304:404. doi: 10.1126/science.1095905
Henshilwood, C. S., d’Errico, F., and Watts, I. (2009). Engraved ochres from the middle stone age levels at Blombos cave, South Africa. J. Hum. Evol. 57, 27–47. doi: 10.1016/j.jhevol.2009.01.005
Henshilwood, C. S., and d'Errico, F. (2011). “Middle stone age engravings and their significance to the debate on the emergence of symbolic material culture” in Homo Symbolicus: the Dawn of language, imagination and spirituality. eds. C. S. Henshilwood and F. d’Errico (Amsterdam: John Benjamins Publishing Company), 75–96.
Henshilwood, C. S., and Dubreuil, B. (2009). “Reading the artefacts: gleaning language skills from the middle stone age in southern Africa” in The cradle of language. eds. R. Botha and C. Knight (Oxford: Oxford University Press), 41–61.
Henshilwood, C. S., and Marean, C. W. (2003). The origin of modern human behaviour: a review and critique of models and test implications. Curr. Anthropol. 44, 627–651. doi: 10.1086/377665
Henshilwood, C. S., and Marean, C. W. (2006). “Remodelling the origins of modern human behaviour” in The prehistory of Africa: Tracing the lineage of modern man. ed. H. Soodyall (Johannesburg: Jonathan Ball Publishers), 31–46.
Henshilwood, C. S., van Niekerk, K., Wurz, S., Delagnes, A., Armitage, S. J., Rifkin, R. F., et al. (2014). Klipdrift shelter, southern cape, South Africa: preliminary report on the Howiesons Poort layers. J. Archaeol. Sci. 45, 284–303. doi: 10.1016/j.jas.2014.01.033
Hoffmann, D. L., Angelucci, D. E., Villaverde, V., Zapata, J., and Zilhão, J. (2018). Symbolic use of marine shells and mineral pigments by Iberian Neandertals 115,000 years ago. Sci. Adv. 4:eaar5255. doi: 10.1126/sciadv.aar5255
Huybregts, R. (2017). Phonemic clicks and the mapping asymmetry: how language emerged and speech developed. Neurosci. Biobehav. Rev. 81, 279–294. doi: 10.1016/j.neubiorev.2017.01.041
Jackendoff, R. (1999). Possible stages in the evolution of the language capacity. Trends Cogn. Sci. 3, 272–279. doi: 10.1016/s1364-6613(99)01333-9
Jackendoff, R. (2002). Foundations of language: Brain, meaning, grammar, evolution. Oxford, UK: Oxford University Press.
Joordens, J. C. A., d’Errico, F., Wesselingh, F. P., Munro, S., de Vos, J., Wallinga, J., et al. (2015). Homo erectus at Trinil on Java used shells for tool production and engraving. Nature 518, 228–231. doi: 10.1038/nature13962
Knight, A., Underhill, P. A., Mortensen, H. M., Zhivotovsky, L. A., Lin, A. A., Henn, B. M., et al. (2003). African Y chromosome and mtDNA divergence provides insight into the history of click languages. Curr. Biol. 13, 464–473. doi: 10.1016/S0960-9822(03)00130-1
Li, Z., Doyon, L., Li, H., Wang, Q., Zhang, Z., Zhao, Q., et al. (2019). Engraved bones from the archaic hominin site of Lingjing, Henan Province. Antiquity 93, 886–900. doi: 10.15184/aqy.2019.81
Lippold, S., Xu, H., Ko, A., Li, M., Renaud, G., Butthof, A., et al. (2014). Human paternal and maternal demographic histories: insights from high-resolution Y chromosome and mtDNA sequences. Investig. Genet. 5, 1–17. doi: 10.1186/2041-2223-5-13
Lorente-Galdos, B., Lao, O., Serra-Vidal, G., Santpere, G., Kuderna, L. K. F., Arauna, L. R., et al. (2019). Whole-genome sequence analysis of a Pan-African set of samples reveals archaic gene flow from an extinct basal population of modern humans into sub-Saharan populations. Genome Biol. 20, 77–15. doi: 10.1186/s13059-019-1684-5
Majkić, A., d’Errico, F., Milošević, S., Mihailović, D., and Dimitrijević, V. (2018a). Sequential Incisions on a Cave Bear Bone from the Middle Paleolithic of Pešturina Cave, Serbia. Journal of Archaeological Method and Theory 25: 69–116. doi: 10.1007/s10816-017-9331-5
Majkić, A., d’Errico, F., and Stepanchuk, V. (2018b). Assessing the Significance of Palaeolithic Engraved Contexts: A Case Study from the Mousterian Site of Kiik-Koba, Crimea. PLoS ONE 13: e0195049. doi: 10.1371/journal.pone.0195049
Mallick, S., Li, H., Lipson, M., Mathieson, I., Gymrek, M., Racimo, F., et al. (2016). The Simons genome diversity project: 300 genomes from 142 diverse populations. Nature 538, 201–206. doi: 10.1038/nature18964
Martins, P. T., and Boeckx, C. (2019). Language evolution and complexity considerations: the no half-merge fallacy. PLoS Biol. 17:e3000389. doi: 10.1371/journal.pbio.3000389
Nóbrega, V. A., and Miyagawa, S. (2015). The precedence of syntax in the rapid emergence of human language in evolution as defined by the integration hypothesis. Front. Psychol. 6:271. doi: 10.3389/fpsyg.2015.00271
Pakendorf, B., and Stoneking, M. (2021). The genomic prehistory of peoples speaking Khoisan languages. Hum. Mol. Genet. 30, R49–R55. doi: 10.1093/hmg/ddaa221
Peresani, M., Dallatorre, S., Astuti, P., Dal Colle, M., Ziggiotti, S., and Peretto, C. (2014). Symbolic or utilitarian? Juggling interpretations of Neanderthal behavior: new inferences from the study of engraved stone surfaces. J. Anthropol. Sci. 92, 233–255. doi: 10.4436/JASS.92007
Petr, M., Vernot, B., and Kelso, J. (2019). Admixr—R package for reproducible analyses using ADMIXTOOLS. Bioinformatics 35, 3194–3195. doi: 10.1093/bioinformatics/btz030
Pinker, S., and Bloom, P. (1990). Natural language and natural selection. Behav. Brain Sci. 13, 707–727. doi: 10.1017/S0140525X00081061
Powell, A., Shennan, S., and Thomas, M. G. (2009). Late Pleistocene demography and the appearance of modern human behavior. Science 324, 1298–1301. doi: 10.1126/science.1170165
Progovac, L. (2016). A gradualist scenario for language evolution: precise linguistic reconstruction of early human (and Neandertal) grammars. Front. Psychol. 7:1714. doi: 10.3389/fpsyg.2016.01714
Progovac, L. (2019). A critical introduction to language evolution: current controversies and future prospects. Cham, Switzerland: Springer.
Radovčić, D., Sten, A. O., Radovčić, J., and Frayer, D. W. (2015). Evidence for Neandertal jewelry: modified white-tailed eagle claws at Krapina. PLoS One 10:e0119802. doi: 10.1371/journal.pone.0119802
Rito, T., Richards, M. B., Fernandes, V., Alshamali, F., Cerny, V., Pereira, L., et al. (2013). The first modern human dispersals across Africa. PLoS One 8:e80031. doi: 10.1371/journal.pone.0080031
Rodriguez-Vidal, J., d’Errico, F., Pacheco, F. G., Blasco, R., Rosell, J., Jennings, R. P., et al. (2014). A rock engraving made by Neanderthals in Gibraltar. Proc. Natl. Acad. Sci. U. S. A. 111, 13301–13306. doi: 10.1073/pnas.1411529111
Sanderson, M. J. (2003). r8s: inferring absolute rates of molecular evolution and divergence times in the absence of a molecular clock. Bioinformatics 19, 301–302. doi: 10.1093/bioinformatics/19.2.301
Scally, A., and Durbin, R. (2012). Revising the human mutation rate: implications for understanding human evolution. Nat. Rev. Genet. 13, 745–753. doi: 10.1038/nrg3295
Scerri, E. M. L., and Will, M. (2023). The revolution that still Isn’t: the origins of behavioral complexity in Homo sapiens. J. Hum. Evol. 179:103358. doi: 10.1016/j.jhevol.2023.103358
Schlebusch, C. M., Malmström, H., Günther, T., Sjödin, P., Coutinho, A., Edlund, H., et al. (2017). Southern African ancient genomes estimate modern human divergence to 350,000 to 260,000 years ago. Science 358, 652–655. doi: 10.1126/science.aao6266
Schlebusch, C. M., Sjödin, P., Breton, G., Günther, T., Naidoo, T., Hollfelder, N., et al. (2020). Khoe-san genomes reveal unique variation and confirm the deepest population divergence in Homo sapiens. Mol. Biol. Evol. 37, 2944–2954. doi: 10.1093/molbev/msaa140
Schlebusch, C. M., Skoglund, P., Sjödin, P., Gattepaille, L. M., Hernandez, D., Jay, F., et al. (2012). Genomic variation in seven Khoe-san groups reveals adaptation and complex African history. Science 338, 374–379. doi: 10.1126/science.1227721
Scozzari, R., Cruciani, F., Santolamazza, P., Malaspina, P., Torroni, A., Sellitto, D., et al. (1999). Combined use of Biallelic and microsatellite Y-chromosome polymorphisms to infer affinities among African populations. Am. J. Hum. Genet. 65, 829–846. doi: 10.1086/302538
Seielstad, M., Bekele, E., Ibrahim, M., Touré, A., and Traoré, M. (1999). A view of modern human origins from Y chromosome microsatellite variation. Genome Res. 9, 558–567. doi: 10.1101/gr.9.6.558
Song, S., Sliwerska, E., Emery, S., and Kidd, J. M. (2017). Modeling human population separation history using physically phased genomes. Genetics 205, 385–395. doi: 10.1534/genetics.116.192963
Soodyall, H., and Jenkins, T. (1992). Mitochondrial DNA polymorphisms in Khoisan populations from southern Africa. Ann. Hum. Genet. 56, 315–324. doi: 10.1111/j.1469-1809.1992.tb01159.x
Tallerman, M. (2014). No syntax saltation in language evolution. Lang. Sci. 46, 207–219. doi: 10.1016/j.langsci.2014.08.002
Tallerman, M. (2017). Can the integration hypothesis account for language evolution? J. Neurolinguistics 43, 254–262. doi: 10.1016/j.jneuroling.2016.06.006
Tattersall, I. (2012). Masters of the planet: The search for our human origins. New York: Palgrave Macmillan.
Tattersall, I. (2017). “Why was human evolution so rapid?” in Human paleontology and prehistory. eds. A. Marom and E. Hovers (Cham: Springer).
Tattersall, I. (2018). An evolutionary framework for the Acquisition of Symbolic Cognition by Homo sapiens. Comp. Cogn. Behav. Rev. 3, 99–114. doi: 10.3819/ccbr.2008.30006
Texier, J.-P., Porraz, G., Parkington, J., Rigaud, J. P., Poggenpoel, C., Miller, C., et al. (2010). A howiesons poort tradition of engraving ostrich eggshell containers dated to 60,000 years ago at diepkloof rock shelter, South Africa. Proc. Natl. Acad. Sci. U. S. A. 107, 6180–6185. doi: 10.1073/pnas.0913047107
Texier, J.-P., Porraz, G., Parkington, J., Rigaud, J. P., Poggenpoel, C., and Tribolo, C. (2013). The context, form and significance of the MSA engraved ostrich eggshell collection from Diepkloof rock shelter, Western cape, South Africa. J. Archaeol. Sci. 40, 3412–3431. doi: 10.1016/j.jas.2013.02.021
Tishkoff, S. A., Reed, F. A., Renciaro, A., Voight, B. F., Babbitt, C. C., et al. (2007). Convergent adaptation of human lactase persistence in Africa and Europe. Nat. Genet. 39, 31–40. doi: 10.1038/ng1946
Vanhaeren, M., d’Errico, F., Stringer, C., James, S. L., Todd, J. A., and Mienis, H. K. (2006). Middle Paleolithic Shell beads in Israel and Algeria. Science 312, 1785–1788. doi: 10.1126/science.1128139
Veeramah, K. R., Wegmann, D., Woerner, A., Mendez, F. L., Watkins, J. C., Destro-Bisol, G., et al. (2012). An early divergence of KhoeSan ancestors from those of other modern humans is supported by an ABC-based analysis of autosomal resequencing data. Mol. Biol. Evol. 29, 617–630. doi: 10.1093/molbev/msr212
Vidal, C. M., Lane, C. S., Asrat, A., Barfod, D. N., Mark, D. F., Tomlinson, E. L., et al. (2022). Age of the oldest known Homo sapiens from eastern Africa. Nature 601, 579–583. doi: 10.1038/s41586-021-04275-8
Vigilant, L., Pennington, R., Harpending, H., Kocher, T. D., and Wilson, A. C. (1989). Mitochondrial DNA sequences in single hairs from a southern African population. Proc. Natl. Acad. Sci. U. S. A. 86, 9350–9354. doi: 10.1073/pnas.86.23.9350
Wadley, L. (2021). What stimulated rapid, cumulative innovation after 100,000 years ago? J. Archaeol. Method Theory 28, 120–141. doi: 10.1007/s10816-020-09499-y
Wang, K., Mathieson, I., O’Connell, J., and Schiffels, S. (2020). Tracking human population structure through time from whole genome sequences. PLoS Genet. 16:e1008552. doi: 10.1371/journal.pgen.1008552
Watanabe, S., Hofman, M., and Shimizu, T. (2017). “Integration hypothesis: a parallel model of language development in evolution” in Evolution of the brain, cognition, and emotion in vertebrates. eds. S. Watanabe, M. Hofman, and T. Shimizu (Tokyo: Springer Japan), 225–247.
Wilder, J. A., Mobasher, Z., and Hammer, M. F. (2004). Genetic evidence for unequal effective population sizes of human females and males. Mol. Biol. Evol. 21, 2047–2057. doi: 10.1093/molbev/msh214
Yang, Z. (2007). PAML 4: phylogenetic analysis by maximum likelihood. Mol. Biol. Evol. 24, 1586–1591. doi: 10.1093/molbev/msm088
Keywords: human population genomics, early human population divergences, language evolution, modern human behavior, modern human cognition
Citation: Miyagawa S, DeSalle R, Nóbrega VA, Nitschke R, Okumura M and Tattersall I (2025) Linguistic capacity was present in the Homo sapiens population 135 thousand years ago. Front. Psychol. 16:1503900. doi: 10.3389/fpsyg.2025.1503900
Edited by:
Barnaby James Wyld Dixson, The University of Queensland, AustraliaReviewed by:
Ljiljana Progovac, Wayne State University, United StatesCopyright © 2025 Miyagawa, DeSalle, Nóbrega, Nitschke, Okumura and Tattersall. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Shigeru Miyagawa, bWl5YWdhd2FAbWl0LmVkdQ==