 Felipe Carvalho Novaes
Felipe Carvalho Novaes Jean Carlos Natividade
Jean Carlos Natividade
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Psychol. , 09 January 2023
Sec. Evolutionary Psychology
Volume 13 - 2022 | https://doi.org/10.3389/fpsyg.2022.874261
This article is part of the Research Topic A 150 Years’ Celebration of Darwin’s Book on Human Evolution and Sexual Selection: Its Legacy and Future Prospects View all 21 articles
Cultural innovations, such as tools and other technical articles useful for survival, imply that creativity is an outcome of evolution. However, the existence of purely ornamental items obfuscates the functional value of creativity. What is the functional or adaptive value of aesthetic and intellectual ornaments? Recent evidence shows a connection between ornamental creativity, an individual’s attractiveness, and their reproductive success. However, this association is not sufficient for establishing that creativity in humans evolved by sexual selection. In this critical review, we synthesize findings from many disciplines about the mechanisms, ontogeny, phylogeny, and the function of creativity in sexual selection. Existing research indicates that creativity has the characteristics expected of a trait evolved by sexual selection: genetic basis, sexual dimorphism, wider variety in males, influence of sex hormones, dysfunctional expressions, an advantage in mating in humans and other animals, and psychological modules adapted to mating contexts. Future studies should investigate mixed findings in the existing literature, such as creativity not being found particularly attractive in a non-WEIRD society. Moreover, we identified remaining knowledge gaps and recommend that further research should be undertaken in the following areas: sexual and reproductive correlates of creativity in non-WEIRD societies, relationship between androgens, development, and creative expression, as well as the impact of ornamental, technical and everyday creativity on attractiveness. Evolutionary research should analyze whether being an evolved signal of genetic quality is the only way in which creativity becomes sexually selected and therefore passed on from generation to generation. This review has gone a long way toward integrating and enhancing our understanding of ornamental creativity as a possible sexual selected psychological trait.
“Sexual selection made our brains wasteful, if not wasted: it transformed a small, efficient ape-style brain into a huge, energy-hungry handicap spewing out luxury behaviors like conversation, music, and art.” (Miller, 2000, p. 134).
Being creative secures undeniable practical benefits for survival. Crows and chimpanzees use twigs and create tools by modifying these twigs to better perform the desired aim (Reader et al., 2016). Chimpanzees, for instance, use such tools for termite fishing (Sanz et al., 2009). Humans have also created tools (e.g., handaxes) and various implements (e.g., clothes) that make it easier to get food and survive in diverse environments (Puccio, 2017). However, what would the evolutionary benefits be of body decoration, cave paintings, literary classics, philosophical treatises, or guitar solos?
Darwin’s (1871) answer to that question was sexual selection. The ability to make tools must have evolved by viability selection, for it helped in survival in hostile environments, while the aesthetic skills required to create artistic performances and products would have evolved through sexual selection because they contributed to mate attraction. The extravagance of bird song and plumage, together with humans’ conspicuous drive to produce art and other forms of ornaments (e.g., self-grooming: Valentova et al., 2022; daily behaviors: Kapoor et al., 2021; and humor: Kaufman et al., 2007) would have a common evolutionary root (Darwin, 1871). The aesthetic sensibility, artistic capacities (including musicality), creative capacities, and creative motivation necessary to produce and appreciate (two different traits) these aesthetically conspicuous manifestations (e.g., art, music, paint, dance, humor, and metaphors) constitute a mental trait named ornamental creativity (Figure 1).
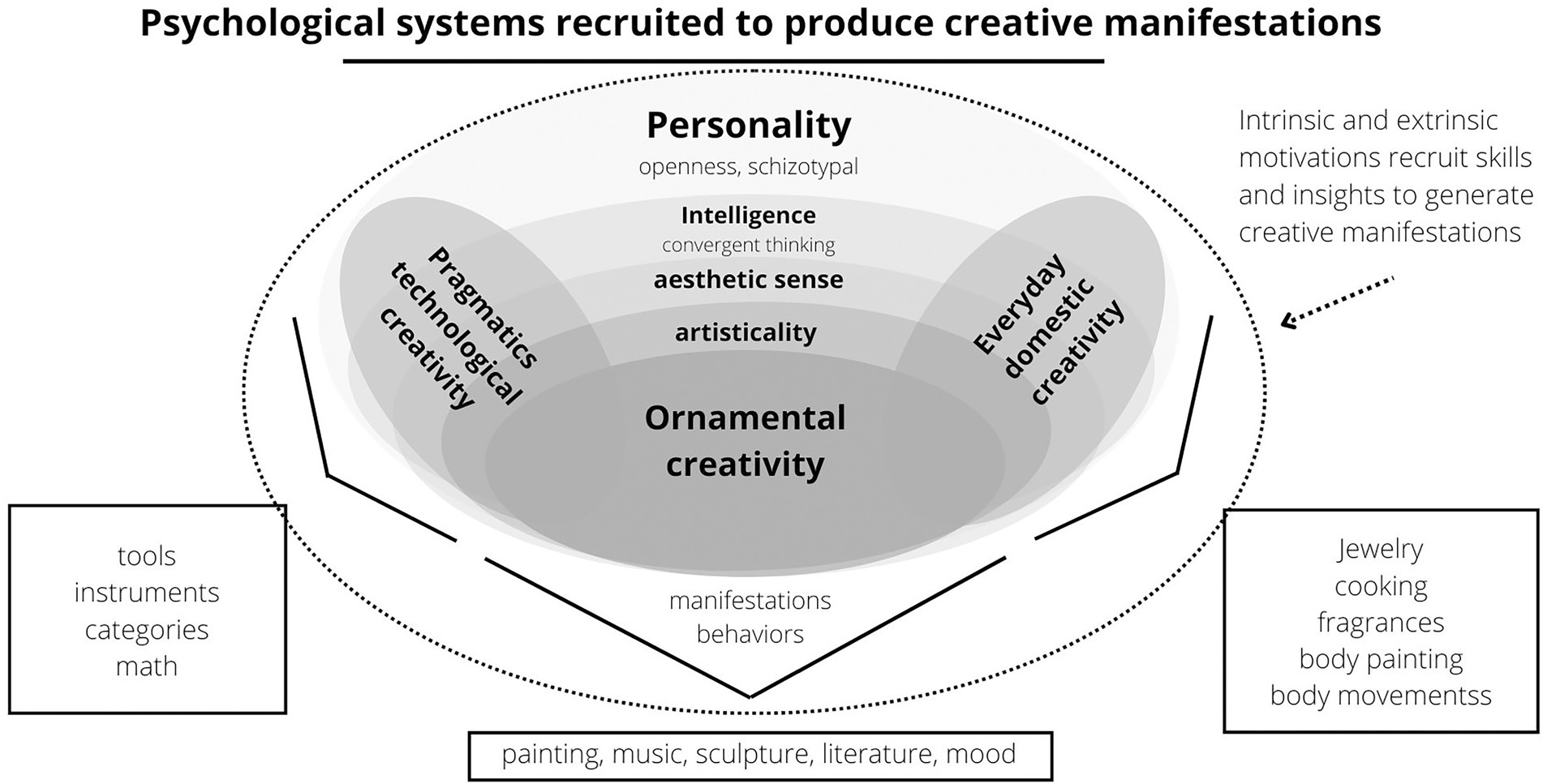
Figure 1. This scheme synthesizes the main variables associated with creativity and the possible relationships between them. A schematic illustration of a possible psychological structure of creativity. People who are more open to new experiences are more curious, flexible, and original, i.e., more creative and intelligent. Creativity involves divergent and convergent thinking, two characteristics associated with intelligence. Each type of creativity would operate according to its psychological mechanisms that are activated in the face of specific contexts and stimuli. These contexts and stimuli can have a reproductive nature, such as situations related to mate attraction or the visualization of a physically attractive partner. According to Feist (2001), ornamental creativity would have evolved by sexual selection because it was more conspicuous (e.g., making artistic pieces), just like the peacock’s plumage (Møller and Petrie, 2002, but also Askew, 2014 and Thavarajah et al., 2016), while technical creativity would have evolved by viability selection because it was more beneficial for survival (i.e., making tools). We also propose that reproductive motivations can mobilize everyday creativity; after all, people use their creativity and aesthetic sensitivity daily to beautify themselves (e.g., using makeup and clothes that enhance the most attractive features of the face and body, respectively; Stephen and Luoto, 2021; Varella et al., 2017; Valentova et al., 2022). Importantly, sexual selection would have shaped the psychological propensities to perform these manifestations and to enjoy them.
Recent studies seem to confirm that more creative people, particularly in the ornamental/aesthetic aspect, are considered more attractive in Western, educated, industrialized, rich, and democratic societies (see Karamihalev, 2013; Lebuda et al., 2021). Nevertheless, more than that is needed to show that creativity evolved by sexual selection.
A complete explanation must consider an ethological analysis, which holistically synthesizes evidence regarding mechanisms, ontogeny, phylogeny, and evolutionary function (Tinbergen, 1963), as well as a nomological network based on theoretical, cross-cultural, hunter-gatherer, phylogenetic, genetic, psychological, medical, and physiological evidence (Schmitt and Pilcher, 2004). The more evidence in that nomological network, the greater the chances that a mental trait is a psychological adaptation (Schmitt and Pilcher, 2004). Psychological adaptations are cognitive modules evolved to solve problems recurring in the environment of evolutionary adaptedness of a species (Schmitt and Pilcher, 2004). The consequences of these modules are flexible and adaptive behaviors, which allow them to be inherited by future generations as tendencies to develop the same modules ontogenetically (Schmitt and Pilcher, 2004). However, modules do not fossilize. How, then, to recognize them?
Human psychological adaptations can be recognized by criteria such as high efficiency, high complexity, high modularity, low phenotypic variance, low genotypic variance, low heritability, universality across cultures and individuals (Miller, 2000). However, the criteria used to identify adaptations evolved by viability selection differ from those used to identify adaptations evolved by sexual selection (Miller, 2000). Some of the ornamental adaptations evolved by sexual selection are fitness indicators. Effective fitness indicators are costly and wasteful, like a peacock’s plumage (Miller, 2000; Møller and Petrie, 2002; Askew, 2014; Thavarajah et al., 2016). Effective fitness indicators are also simple because they do not need to convey much information or to recruit many resources from the organism; they only need to “create a discriminable signal perceivable at a reasonable distance that reliably indicates a single quantity” (Miller, 2000). Fitness indicators must be sensitive to the covariation of different capacities to indicate their quality level, such as a peahen that notices a peacock’s exuberant tail and becomes inclined to mate with it (Miller, 2000). Furthermore, fitness indicators tend to vary more in the population than adaptations shaped by viability selection (Miller, 2000). That greater variability allows individuals to be sexually selected according to the display of the most costly indicator (Miller, 2000).
This review integrates existing evidence to explore whether creativity is or is not a psychological adaptation evolved through sexual selection. There are earlier attempts to review evidence about the evolution of mental abilities (Miller and Todd, 1998; Klasios, 2013), including creativity (e.g., Karamihalev, 2013), but these earlier attempts do not use the nomological approach presented here (Miller, 2000; Schmitt and Pilcher, 2004; Lewis et al., 2017).
Before proceeding, we must make some critical caveats. We do not suppose that creativity has evolved only by sexual selection. The idea is that the selection of creative partners has possibly overemphasized this ability, allowing it to be co-opted for ornamental purposes (Miller, 2000, 2001; Winegard et al., 2018). We are also not assuming that a trait evolved by sexual selection originated from sexual selection. Traits can initially evolve by viability selection and then be co-opted and exapted by sexual selection (e.g., foot fetish or bodily piercings; Luoto, 2019a). We are also not saying that sexual selection is only about sex differences (see Hooper and Miller, 2008; Janicke and Fromonteil, 2021), nor only male-biased sex differences (Miller, 2013; Varella et al., 2014, 2017; Rosenthal and Ryan, 2022). Further, sexual selection is not a proximate motivation (e.g., Bach could have been religiously motivated when composing music but still could have increased mating success because of his musical success; see Miller, 2000; Varella et al., 2013). Also, sexual selection is not only mate attraction but also relationship maintenance (long-term), intrasexual competition, intersexual conflict, and parenting (Kenrick et al., 2010; Petrie, 2021; Shuker and Kvarnemo, 2021). Furthermore, we are not claiming that ornamental creative ability is the only route to differential reproduction; other such domains include, but are not limited to, sports, physical enhancements, resource acquisition, parenting skills, and nepotism (Manning and Taylor, 2001; Stephen and Luoto, 2021; Walter et al., 2021; Varella et al., 2022).
Before approaching the multiple lines of evidence supporting creativity as a sexually selected trait, it is necessary to characterize creativity and its constituent variables (Figure 1).
Creativity is synonymous with originality and effectiveness (Runco and Jaeger, 2012). Creativity includes original aesthetic manifestations and precise imitations (non-original) of aesthetic manifestations. Creative people are often called innovative and inventive (Said-Metwaly et al., 2017). Research on creativity has focused on individual-level cognitive aspects as divergent thinking and intelligence (Runco and Jaeger, 2012; Karwowski et al., 2016b) and personality, mainly plasticity and openness (Karwowski et al., 2016b). From a process-focused perspective, creativity is a process of blind variation and selective retention of original ideas (Jung et al., 2013). Creative products result from these individual characteristics and processes and manifest in multiple domains, such as everyday, scholarly, performance, scientific, and artistic domains (Kapoor et al., 2021).
All of these creative domains are costly because successfully navigating them depends on healthy brains, and healthy brains are costly (i.e., energy-intensive and susceptible to instabilities throughout development; Miller, 2001). However, ornamental or aesthetic manifestations may be more expensive because they require a lot of energy expenditure without the practical benefits in return (from the point of view of survival; Feist, 2001). That would be the case for creativity employed in such domains as art and body beautification.
Creativity is often operationally defined as divergent thinking, although the connection between creative capacity, creative achievement, and divergent thinking is not always clear (Hornberg and Reiter-Palmon, 2017). Divergent thinking is the ability to come up with solutions, answers, or questions in response to an open problem of a visual or verbal nature (Hornberg and Reiter-Palmon, 2017). The level of divergent thinking depends on originality and fluency, where originality is the number of responses distinct from those of other individuals, and fluency is the overall number of responses (Hornberg and Reiter-Palmon, 2017). Divergent thinking alone does not guarantee creativity, and a dose of convergent thinking is also needed, which is defined as the ability to select the most helpful ideas from among those generated by the divergent associative process (Cropley, 2006).
Openness to experience and extroversion are the two personality components most consistently associated with creativity (Karwowski and Lebuda, 2016; Hornberg and Reiter-Palmon, 2017; Vartanian et al., 2018). Individuals higher in openness to experience are flexible, curious, less conventional, and tend to seek sensations and stimulation. The search for sensations and stimulation are occasionally pointed out as characteristics of extroversion as well (Karwowski and Lebuda, 2016). Openness and extroversion make up a second-order factor called plasticity in a model called Big Two, which positively predicts creativity and beliefs about creative capacity (Puryear et al., 2017; Feist, 2019). Openness to experiences is divided in sub-dimensions. The intellect sub-dimension, linked to intellectual curiosity, predicts success in science; the openness sub-dimension, linked to desire of adventure, predicts success in the arts (Feist, 1998).
Schizotypy is a personality feature that seems to be associated with creativity. Schizotypal traits are characterized by positive symptoms, such as magical thinking, unusual perceptual experiences, impulsive nonconformity, and negative symptoms, such as introversion, emotional instability, and cognitive disarray (Holt, 2019). Schizotypy is positively related to originality and divergent thinking (e.g., Wang et al., 2018; Holt, 2019). Exaggerated schizotypy leads to exaggerated and unexpected associations, which leads to exaggerated creativity, observed mainly in artists (Acar and Sen, 2013; Carter et al., 2019; Aguilera and Rodríguez-Ferreiro, 2021). Artists often display “healthy schizotypy,” that is, higher creativity with no psychosis symptoms (Holt, 2019; Rantala et al., 2022). For example, poets tend to be more schizotypal, associating less obviously related ideas (Acar and Sen, 2013).
Autistic traits may also be associated with creativity (Baron-Cohen et al., 2001). Subjects with autism are inhibited in fluency and flexibility, but they have high levels of attention to details and originality; thus, subjects with autism are creative for different reasons, compared to the general population (for a meta-analysis, see Pennisi et al., 2020). Further, individuals with non-clinical autistic phenotype score lower on self-report creativity scales but exhibit greater creative performance in tasks involving drawing (Jankowska et al., 2019) and greater convergent thinking in anagram solution tasks (Abu-Akel et al., 2020).
Intelligence is also involved with creativity. Intelligence (i.e., g factor or cognitive ability) is the capacity to think, plan, solve problems, and adapt to the environment (Cattell, 1963). More specifically, intelligence is a general factor that emerges from performance in specific and interrelated domains (e.g., verbal, spatial, mathematical). It comprises the ability to reason (fluid intelligence) and the ability to acquire knowledge (crystallized intelligence; Cattell, 1963; see also Kovacs and Conway, 2019).
Solving problems requires intelligence. However, it also requires originality and thinking beyond the obvious. Thus, it is difficult to distinguish between creativity and intelligence. Creative people also tend to be intelligent (Kim, 2008; Karwowski et al., 2021). For example, a meta-analysis that included 11,418 people showed that success in mathematics (a field with performance closely linked to IQ) is associated with creativity (Bicer et al., 2021). For instance, fluid intelligence and originality are positively correlated (Silvia, 2008). Higher fluid intelligence is associated with a higher use of metaphors (Silvia and Beaty, 2012). Individuals with better memory show more divergent thinking (Silvia and Nusbaum, 2013). Those with higher divergent thinking have higher verbal, figural (Cho et al., 2010), and visuospatial reasoning (Kell et al., 2013). In other words, intelligence is a necessary condition of creativity (Guilford, 1967).
These relationships can also manifest in specific fields. Scientists need to analyze problems systematically (which demands intelligence) to reach innovative outcomes (creativity; Karwowski et al., 2021). Elaborating disruptive theories requires “thinking outside the box” and pattern recognition (Feist, 1998). On the other hand, artists can work by associating ideas more freely (Boyd, 2010; Karwowski et al., 2021). Thus, intelligence and creativity are essential to the arts and sciences, but technical fields are more cognitively loaded than the arts (Feist, 1998; Park et al., 2007).
Aesthetic sensibility is the ability to assess the quality of sensory stimuli, such as abstract drawings and human faces. Eibl-Eibesfeldt (1989) describes three levels in human aesthetic psychology: (1) the basic level, which we have shared with most vertebrates and includes regularities, symmetry, harmony, and superstimuli; (2) the human-specific level, which is universal and relates to a human “sense of order” that underlies more specific attributes such as balance, rhythm, rhyme, and harmony; (3) the local culture level which is related to traditions and tastes shared within each human population.
The capacity to perform aesthetic appraisals (visual) is associated with intelligence, divergent thinking (figural but non-verbal), and personality (aesthetic openness; Myszkowski et al., 2014). Creative ornamental products are judged more on their aesthetic rather than technical merit, thus, they are expected to rely more on aesthetic sensibility (Feist, 2001). Personality also seems to be associated with aesthetic sensibility. People with greater openness to experience and schizotypy tend to appreciate artistic activities more (Feist and Brady, 2004) and have higher aesthetic motivation (Furnham, 2021).
It is necessary to emphasize that aesthetic and artistic sensibility are not synonymous. Esthetic sensitivity is a more general capability than artistic; the two are independent adaptations of each other (Varella et al., 2011). For example, many animals distinguish different types of human art; however, few animals find it as reinforcing as humans do (e.g., Varella, 2021), which points to the specificity of human artistic appreciation (Watanabe, 2013).
Why do human beings employ their creativity in making original works of art? In line with previous proposals, we believe that part of the answer concerns sexual selection (Darwin, 1871; Low, 1979; Eibl-Eibesfeldt, 1989; Zahavi and Zahavi, 1997; Miller and Todd, 1998; Miller, 2000; Miller, 2001; De Block and Dewitte, 2007; Baer and Kaufman, 2008; Dutton, 2009; Varella et al., 2011, 2017, 2022; Verpooten and Nelissen, 2012; De Ridder and Vanneste, 2013; Karamihalev, 2013; Westphal-Fitch and Fitch, 2018; Luoto, 2019a). However, to test this hypothesis, the evidence must be collected from diverse levels of analysis (mechanisms, development, stimuli, phylogeny, and function; Tinbergen, 1963; Varella et al., 2012; Fitch, 2015). More recently, this approach has been expanded by investigating cultural, social, biological, and environmental inputs that activate psychological modules (Lewis et al., 2017; Luoto, 2019a). To answer whether creativity evolved by sexual selection, one needs to establish whether different sources of evidence converge toward indicating that ornamental creativity serves reproductive ends (Table 1).

Table 1. Predictions based on sexual selection and supporting evidence.
There are polymorphisms in alleles of genes associated with dopaminergic systems, such as genes DRD2, DAT, COMT, DRD4, and TPH1, which are also associated with creative abilities and achievements (e.g., Reuter et al., 2006; Runco et al., 2011; Mayseless et al., 2013; Zabelina et al., 2016; Luoto, 2019b). Such genes also appear to play a role in sexual selection. For example, D4 dopamine receptor gene variation is linked to infidelity and sexual promiscuity (Garcia et al., 2010; Acevedo et al., 2020). Furthermore, genes associated with creativity and preference for creative partners are correlated, which shows that creativity has been subject to sexual selection at some level, perhaps by assortative mating (Verweij et al., 2014).
The selection of these genes linked to creativity has an ancient history. Modern humans have over 200 unique non-protein-encoding genes that regulate the co-expression of many other protein-encoding genes in coordinated networks underlying modern capabilities such as creativity, which are not found in chimpanzees or Neanderthals (Zwir et al., 2022).
These genes provide a part of the biological substrate necessary for creativity; however, they do not necessarily imply the development of creative capacities. Inheriting the propensity to develop the ability differs from having the ability, which depends on adequate stimulation throughout ontogenetic development as well as a host of other neurodevelopmental and biopsychosocial factors.
Dopamine has a role in creativity and sexuality, which suggests a link with sexual selection (Garcia et al., 2010; Acevedo et al., 2020). Dopaminergic activity is also related to psychomotor agitation behaviors such as eye blinking, that is a known clinical marker of accelerated dopaminergic activity observed in schizophrenic patients and non-clinical creative individuals (Akbari Chermahini and Hommel, 2009).
Testosterone is also related to creativity. For instance, men and women usually present a peak of musical talent from puberty (Hassler, 1992). Testosterone seems to enhance musical performance up to a certain level, but performance drops above this level (Hassler, 1992). Hassler (1992) conjectures that the positive effect of the hormone on musical creativity is mediated by its influence on spatial reasoning. In fact, children trained in a musical instrument have better indicators of intelligence and creativity (Benz et al., 2016; but see Mosing et al., 2014 for a discussion of genetic and practice effects). Artists of both sexes have a lower 2D:4D ratio, which suggests these individuals sustained more influence of testosterone during intrauterine development (Sluming and Manning, 2000; Crocchiola, 2014). Furthermore, there is a positive correlation between lower 2D:4D, amount of Nobel prizes, and scientific publications (van der Linden et al., 2020). On the other hand, concentrations of salivary testosterone and preference for sophistication levels of music are negatively correlated (Doi et al., 2018).
Hormonal influence in creative expressions can be detected in the patterns of drawings made by boys and girls. For instance, between 5 and 6 years old, boys draw more objects, use fewer colors and prefer cool colors, while girls draw more people and flowers, using more colors (Abraham, 2016). Girls with congenital adrenal hyperplasia (who consequently produce more androgens than the female average) present drawing patterns more akin to those made by boys (Iijima et al., 2001).
Though women with high testosterone display intense creative activity, the pattern seems more consistent in men. The masculine peak of artistic output happens in married men from 30 to 40 years old, extending beyond 40 for singles (Kanazawa, 2000). Men write 10 times more books and other literary outputs, accounting for more entries in the Guinness World Records (Lange, 2011; Lange and Euler, 2014).
So what, after all, is the role of testosterone in creativity? Androgen levels may be positively linked to increased performance and personality. For example, increasing performance in skills such as spatial reasoning and increasing willingness to take risks, compete, and seek novelty (Feist, 2019; Luoto and Varella, 2021). That would partially explain the male prominence in scientific and artistic fields, even without differences in creativity averages between the sexes (see Goldin and Rouse, 2000; Mosing et al., 2015).
Could brain architecture influence creativity and thus affect mate selection? More creative people exhibit brain laterality leaning to the right; however, the findings are mixed. For example, more schizotypal people use the right hand less (Somers et al., 2009) and show greater left asymmetry in the use of the senses (Lindell, 2014), which suggests greater right lateralization of the brain hemispheres. Whereas the left hemisphere is associated with access to more specific semantic networks, the right hemisphere is connected to more diffuse networks, connecting more general ideas in the semantic network, which may explain the activation of this hemisphere during divergent thinking (Lindell, 2014).
Lateralization of the brain hemispheres appears to be associated with testosterone. For example, boys with higher testosterone levels at puberty show greater right brain lateralization (Beking et al., 2018). Boys with higher levels of intrauterine testosterone showed the opposite lateral (i.e., left) activation at puberty (Beking et al., 2018), which seems more associated with an autistic profile (Castelli et al., 2002).
If creativity evolved through sexual selection, it would logically follow that creative partners will be in higher demand. Studies have shown that creativity is a trait desired in romantic partners by Americans (Buss, 1989; Buss et al., 1990; Li et al., 2011), Brazilians (Souza et al., 2016; Novaes, 2022), Chinese (Chang et al., 2011), Singaporeans (Li et al., 2011), Indians (Kamble et al., 2014), and others (Buss et al., 1990; Watkins, 2017; but see Lebuda et al., 2021). The importance given to creativity varies. Creativity, especially of the ornamental/aesthetic type (Kaufman et al., 2016; see Figure 1), starts to matter in mating as soon as primary preferences (such as physical beauty) are satisfied (Li et al., 2002). Creativity is so essential in attractiveness that it can make someone more attractive than social status (Buss and Barnes, 1986; Novaes, 2022), physical appearance (Watkins, 2017), or intelligence (Prokosch et al., 2009).
The power of creativity can explain the universal preference for creative partners in partner retention. Couples that engage in novel and stimulating activities become closer (Aron et al., 2005). Thinking about romantic relationships stimulates creativity more than thinking about casual sex (Campbell and Fletcher, 2015). However, the consequences change depending on the type of creativity considered. For example, while everyday creativity increases romantic love, artistic creativity decreases it – but they did not elaborate explanations for these results in evolutionary terms (Campbell and Kaufman, 2017).
Fitness indicators are subject to instability in their development, affecting their carriers’ reproductive success (Klasios, 2013). For example, peacocks may have trouble developing their extravagant plumage because of deleterious mutations, environmental stress, or parasites (Møller and Petrie, 2002; Askew, 2014; Thavarajah et al., 2016). Likewise, developmental problems can impair brain development and the display of mental and, by extension, creative ornaments (Miller and Todd, 1998; Shaner et al., 2004; Del Giudice et al., 2010). Some psychological disorders may result from ontogenetically or evolutionarily disturbed mental adaptations (Figure 2; Rantala et al., 2019, 2021, 2022).
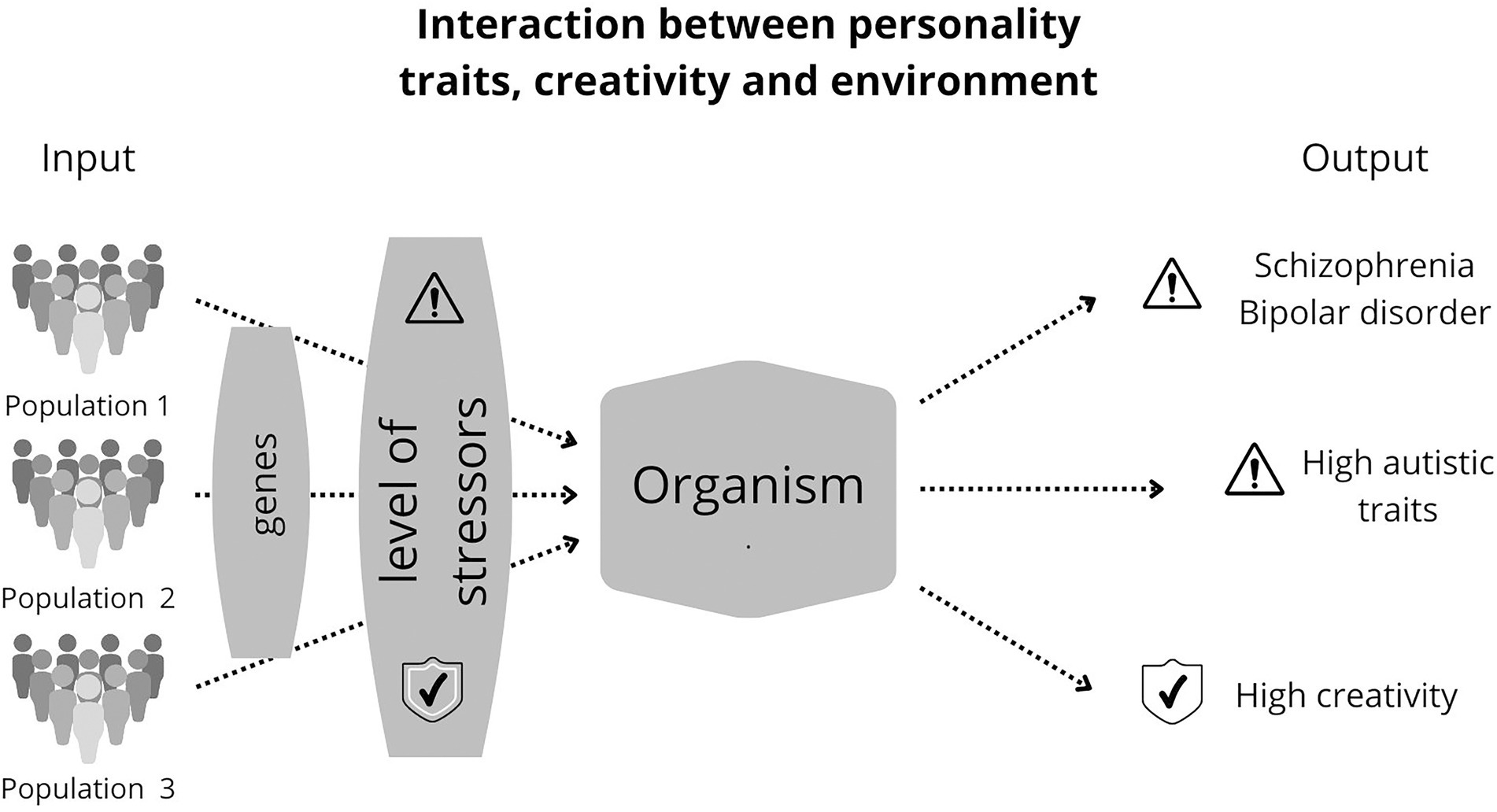
Figure 2. Consequences of stress on people with schizotypal and autistic traits. Creative people are moderately schizotypal. In other words, schizotypal people are often imaginative, associating ideas in unusual ways. People with schizophrenia and bipolar disorder also have these characteristics, sometimes manifesting them in a “dysfunctional way,” e.g., in a paranoid way, but not in an artistic way. That “dysfunctional manifestation” of creativity can occur because of stressors that disrupt normal nervous system development. Thus, individuals with the same genetic propensities for high creativity may manifest it in the form of a disorder or the neurotypical form, depending on how much the stressors have affected ontogeny. This figure shows three examples of populations, each subjected to different levels of environmental stress. The symbol with “!” means high levels of stressors, while the symbol “ok” means tolerable levels of stressors, that is, those that do not significantly impair neural development.
Schizotypal traits are linked to mental ornaments that involve creativity and can be displayed in various ways, such as verbally and visually (Shaner et al., 2004). Dysfunctional levels of schizotypy characterize schizophrenic people, and they suffer from verbal difficulties, such as a disorganized expression of thoughts through language (Shaner et al., 2004). Despite high levels of schizotypy, schizophrenic and bipolar people have difficulty expressing creativity (Acar and Runco, 2012; Acar et al., 2018). The association between schizotypy and creativity follows an “inverted U” shape, growing to a point beyond which one decreases while the other increases (Abraham, 2014). That explains the ambiguous findings on disorders and creativity (Acar and Runco, 2012; Acar and Sen, 2013; Acar et al., 2018).
Mood disorders are also associated with creativity only to a certain extent. A meta-analysis showed that (verbal) creativity and bipolarity are positively associated (Taylor, 2017). Epidemiological analyses reveal a higher prevalence of bipolar disorder among academics and artists (Kyaga et al., 2011). Likewise, people with a high expression of autistic traits are more prevalent in fields such as engineering and mathematics (Morsanyi et al., 2012). However, the association between creativity and success declines as the severity of psychopathological symptoms increases (Pennisi et al., 2020) – see Figure 2.
The shared genetic basis between mental disorders, personality, and creativity can explain these associations. In other words, the genetic risk of developing schizophrenia and bipolar disorder predicts artistic achievements positively (Power et al., 2015). More precisely, higher polygenic risk for bipolar disorder is associated with a higher tendency for divergent thinking (Takeuchi et al., 2021). Also, the polygenic risk of developing schizophrenia is positively associated with risk behavior (Li et al., 2020), which is linked to creativity (Feist, 2019). The same genetic component involved in verbal ability is involved in schizophrenia, which may explain verbal proficiency drawbacks in schizophrenic patients (Jonsdottir et al., 2021). Similarly, autism shares some of the genetic basis of intelligence, which can explain the relation of the autistic phenotype with the improvement in cognitive performance and convergent thinking, which are essential to creativity (Crespi, 2016). In addition, there may be some relationship between autism, creativity, and artisticality (Kellman, 1998; Spikins et al., 2018).
The connection between creativity and personality occurs on many levels. For example, creativity, intelligence, openness to experience, and extraversion share a genetic basis (Kandler et al., 2016), which may be a consequence of assortative mating (Conroy-Beam et al., 2019). This genetic basis is involved in dopaminergic systems (Gocłowska et al., 2019). Such systems are activated during process that constitute creative thinking, such as divergent thinking, exploratory activities, and the search for novelty (Vartanian et al., 2018; Gocłowska et al., 2019), commonly associated with individuals high in extroversion and openness (Feist, 2019).
In relation to sex differences, studies have shown that women are more open to experiences than men, whereas others found no gender distinction (Weisberg et al., 2011; Natividade and Hutz, 2015; Schmitt et al., 2017). Differences may be found in the openness to experience subfactors, with women more open than men and men more intellectual than women (Weisberg et al., 2011). Regarding extroversion, women, on average, are more extroverted than men (Weisberg et al., 2011; Natividade and Hutz, 2015; Schmitt et al., 2017).
Creative behaviors may result from psychological structures evolved to solve adaptive problems. Such psychological structures consist of capacities, perceptions, and motivations shaped to generate adaptive outputs (Lewis et al., 2017). This framework has recently been applied to “artisticality” (Varella et al., 2011, 2017; Luoto, 2019a; Varella, 2021) and “musicality” (Bispham, 2009). Here, we propose a similar psychological framework for creativity. That is, creative behaviors of any kind (everyday, academic, performative, scientific, and artistic; Kapoor et al., 2021) will result from evolved psychological structures (creative capacity, aesthetic sense, and motivation) capable of generating cultural creative outputs.
Psychological adaptations are related to intrinsic motivation to perform certain activities. In fact, children and adults from different cultures exercise their creative capacity in games and other activities in search of fun, pleasure, and affective social interactions (Boyd, 2010; Moraes et al., 2022). This creative play may or may not involve aesthetic sensibility (e.g., drawing and painting; Myszkowski et al., 2014, 2018). Further, this exercise of creative and aesthetic capacities is so spontaneous that it does not even need artistic training (Frois and Eysenck, 1995; Boyd, 2010).
In other words, there is motivation early on in development to put creative and aesthetic capabilities into action. Motivation can be defined as organizing and coordinating aspects of behaviors that arise from a wide variety of internal, environmental, and social sources and is manifested at many levels of behavioral and neural organization (Shizgal, 2001). There is intrinsic and extrinsic motivation. Intrinsic motivation is the will or impulse to engage and sustain engagement in a given activity; extrinsic motivation involves engaging in an activity to have rewards external to the activity itself (Ryan and Deci, 2017). People often engage in creative activities because they are intrinsically motivated by the activity, as in the case of involvement with music (Bispham, 2009) and art (Varella, 2021). It is not by chance that individuals who choose art-related courses are more motivated by intrinsic reasons (e.g., using their talents) than extrinsic ones (e.g., parental and media influence, earnings, status; Varella, 2021).
Furthermore, intrinsic motivation did not vary significantly between arts careers (e.g., music, dance, theater, visual arts, and literary studies), suggesting a specific motivational system for general artistic abilities that underlies the expression of all artistic modalities (Varella, 2021). Offering rewards (extrinsic motivation) for creative performance can even decrease the quality of products generated by children (Amabile and Gitomer, 1984) and possibly in other animals, as anecdotally observed in a chimpanzee (Morris, 1962). Motivation in this area appears very early in development (Bispham, 2009; Varella, 2021). Children already draw, and babies are entertained by the mother’s vocalizations and movements, which may indicate the perception of rhythm, fundamental for appreciation and aesthetic productions in music and dance (Trevarthen, 1999; Bispham, 2009).
Some people are more creative than others, which is spontaneously evidenced since childhood (Feist, 2004). As children age, more cognitive maturity ensues, and it may be accompanied by a greater investment of creativity in specific domains, such as visual art, music, literature, or science (Barbot and Tinio, 2015). If creativity has evolved by sexual selection, it is expected that its increase follows changes beginning at puberty, when there is a boost in the release of androgens (Miller, 2000; Miller, 2001). However, that boost is not linear. During puberty, there is a drop in gray matter and the number of dopamine receptors; this likely explains the decrease in the cognitive aspect of creativity and the increase of the aspects related to personality, such as openness, thrill-seeking and novelty (Barbot and Tinio, 2015).
The relationship between androgens and creative expression could indicate the existence of sexual dimorphism in creativity, but, considering physical and psychological characteristics in general, humans have milder sexual dimorphism than other species (Janicke and Fromonteil, 2021). More specifically, humans tend to be more monomorphic because both sexes are heavily invested in offspring and, therefore, more selective in choosing mates (Stewart-Williams and Thomas, 2013). That likely led both females and males to evolve and develop fitness indicators for mate attraction (Stewart-Williams and Thomas, 2013). That would explain the similarity in creativity between men and women and the greater variability in different creative domains in both sexes (details in Table 1; Varella et al., 2017; Nakano et al., 2021).
Evidence on the variability of creativity in childhood is mixed (Lau and Cheung, 2015; He, 2018). There is evidence of greater male variability in adulthood (He and Wong, 2011; He, 2018), which has been replicated in several countries in African (Karwowski et al., 2016a) and European (Karwowski et al., 2016b) continents, as well as the United States (Taylor and Barbot, 2021), with a few exceptions (He et al., 2013; Ju et al., 2015; Lau and Cheung, 2015). For instance, a study conducted in Poland reported greater male variability in all ages in the performance at a task that involved completing a drawing as creatively as possible (Karwowski et al., 2016b). In a second study, various domains of creativity were examined in a sample of people of diverse ages and schooling. There were no sex differences in the means in performance or potential, but there was sex differences on specific domains of creativity (e.g., Karwowski et al., 2016b), as well as a greater male variability (e.g., Karwowski et al., 2016b).
However, the variability in creativity in each sex may also depend on the creative domain evaluated. He (2018) measured divergent thinking and creative problem-solving in Hong Kong university students. Greater male variability was found in divergent thinking tasks involving images but not verbal tasks (He, 2018). See Table 1.
Average creativity between men and women is highly variable among studies, even among creative domains (Abraham, 2016). In childhood, girls perform better than boys (Cheung and Lau, 2010; Hemdan and Kazem, 2019). In adults, most studies show female superiority (45.2%), while others find no sex differences (31.5%; Nakano et al., 2021). In a study on 3–7-year-olds, girls were better represented at the top of creativity distribution, while in the age range of 19–23-year-olds, the boys had the best representation at the top (He et al., 2015). In a longitudinal study that lasted four years in Hong Kong, boys and girls from 8 to 11 years old displayed improvement in creativity, with female superiority (He, 2018), but boys’ creativity increased from age 15 and surpassed that of girls at 16 (He, 2018).
The disparity between men and women may also be perceived in personality traits associated with creativity. For example, teenage girls are more extroverted and open to experiences (personality traits associated with creativity) than boys of the same age, which corroborates the evidence of higher creativity in girls in this age range (De Bolle et al., 2015). These disparities in personality may stem from male and female timing in sexual maturation. Girls enter puberty earlier, which seems to explain their improvement in socialization and cognitive performance ahead of boys (De Bolle et al., 2015).
If creativity is a result of evolution, it is to be expected that there would be similar characteristics in other species (e.g., Cauchard et al., 2013). Likewise, if this process is partly due to sexual selection, it makes sense to assume that creativity plays a role in the reproduction of humans and other species.
Species that develop in hostile and unstable environments tend to have an intense social life, large brains, extended youth, and an ability to learn; that is to say, these species are marked by higher phenotypic plasticity, which carries advantages in solving adaptive and new problems (Lefebvre, 2013). This is the case with some primates, including Homo sapiens, and birds, like crows (Lefebvre et al., 2004; Sanz et al., 2009; Lefebvre, 2013; Bandini and Harrison, 2020). For instance, crows choose hook-shaped twigs, ideal for “fishing” food from hard-to-reach places (Taylor, 2014). Chimpanzees fashion sprigs to feed on termites from inside trunks (Bandini and Harrison, 2020). An anecdotal sample showed that a male chimpanzee used plant leaves and branches to emit specific sounds that caught females’ attention (Bandini and Harrison, 2020).
Animals more inclined to be innovative tend to be neophilic, i.e., they run more risks and seek novelty and sensations (Kaufman et al., 2011). Orangutans illustrate the case of a neophobic primate known for innovating less in natural environments; on the other hand, chimpanzees are more neophilic (van Schaik et al., 2016; Bandini and Harrison, 2020). Just like humans, these plastic behaviors are associated with dopamine in various brain systems (Kaufman et al., 2011).
There has been shown sex differences in innovation. Creativity can be used by the less dominant sex as an alternative sexual and foraging strategy, as it happens among chimpanzees, where females are the most habitual tool users (Reader and Laland, 2001). However, Lefebvre (2013) states that males innovate more than females not only in primates, but also in birds and in ancestral humans. Using tools to enable more accessible food gathering brings evident benefits for survival and less obvious ones for reproduction. Males with privileged access to food gain advantages in the social hierarchy, ensuring access to allies and females (Reader and Laland, 2001).
Some displays of creativity are more ornamental or aesthetic than pragmatic, impacting mating (Miller, 2001). The bowerbird is a classic example of an ornamental manifestation of cognition in non-human animals. The male satin bowerbird increases its reproductive success by decorating the bower with blue objects (Borgia, 1986). To do this, males need an aesthetic sense to decorate the bower, just as females need an aesthetic sense to evaluate the best-decorated bower. Depending on the definition of art and aesthetics used, the bowers produced and appreciated by bowerbirds can be considered non-human examples of art (Endler, 2012). This aesthetic sense needs a certain cognitive complexity typically present in altricial species (Boogert et al., 2011).
In fact, females in several non-human animal species seem to prefer males who tend to exhibit better cognitive performance. For instance, males that build the fanciest bowers have greater reproductive success, and these males are better at problem-solving (e.g., getting food from a box; Keagy et al., 2009). Also, female budgerigars remain closer to males that manifest more complex problem-solving (Chen et al., 2019). The same behavior is observed in eastern meadow voles (Microtus pennsylvanicus) and North Island robins (Petroica longipes; Spritzer et al., 2005; Shaw et al., 2019). These behaviors suggest that males who exhibit greater cognitive complexity tend to attract females. However, research is unclear as to whether exhibiting cognitive behaviors attracts females or whether some other trait, indirectly correlated with cognition, generates that attraction.
As an altricial species (or a secondary altricial species, see Portmann, 1969), humans also have a large brain and prolonged development that favor cognitive flexibility. The genus Homo seems to have been expressing itself creatively for thousands of years. Homo erectus already used pigments 800,000 years ago likely for decorative purposes in Southern Africa; 450,000 years ago, they already scratched arbitrary lines on mollusk shells, just as chimpanzees seem to do; 40,000 years ago, Homo sapiens made cave paintings and decorated tools from the American continent to Asia (Høgh-Olesen, 2018).
The advent of decorated tools suggests that technical and ornamental displays of creativity have co-evolved. Tools probably started to be produced to solve practical problems, but ornamenting these tools could have social functions (e.g., identification in the group hierarchy), which characterize weapons and uniforms in more recent human history (Menninghaus, 2019). However, these creative and artistic manifestations may be collective activities of the whole group, not restricted to specialized individuals, i.e., professional artists (e.g., Mackie, 2015; Rabazo-Rodríguez et al., 2017).
A specific (and controversial) hypothesis about Paleolithic tools’ pragmatic and ornamental function is that of the sexy handaxe (Kohn and Mithen, 1999). According to this hypothesis, handaxes would have the same ornamental function as the bowerbird’s decorated bower (Kohn and Mithen, 1999). An effective handaxe just for hunting would not need to be overly symmetrical, big, or heavy, making them difficult to use as a weapon (Menninghaus, 2019). Thus, such artifacts could also have social functions, such as signaling identity and potential as a reproductive partner, which could indicate the level of status, dominance, and/or aesthetic skills. It is also possible that the symmetry of artifacts takes advantage of sensory system biases that make specific patterns more attractive than others (such as more symmetrical faces; Gangestad and Thornhill, 2003). In the aggregate, such findings as reviewed above suggest that displaying aesthetic and ornamental capabilities linked to creativity is not restricted to the present but also to the evolutionary past.
The available evidence indicates that creative manifestations—mainly ornamental ones, but also pragmatic ones—may play a unique role in attractiveness. But why? In the following sections, we will examine evidence related to various theories (see also Davis and Arnocky, 2022).
According to the good genes model, traits selected by sexual selection confer indirect advantages to the offspring, such as genetic quality (see Davis and Arnocky, 2022). “Providing” good genes is essential in species with low male parental investment (Trivers, 1972; Zahavi, 1975). For example, peacocks display their striking plumage, and bowerbirds display their decorated bowers that serve as honest signals of genetic and/or phenotypic quality (Zahavi, 1975; Borgia, 1986). The mental fitness hypothesis uses the same logic applied to the mind. According to that hypothesis, higher-than-average creativity and intelligence would lead to improved reproductive success by indirectly indicating genetic quality (Miller and Todd, 1998; Miller, 2000, 2001; Karamihalev, 2013). Genetic quality means fewer harmful mutations (Klasios, 2013). These mutations can disrupt the organism’s development, including the brain (Klasios, 2013). Therefore, displaying creative products ultimately indicates that the producer is healthy and has an efficient brain (Miller and Todd, 1998; Klasios, 2013).
If the mental fitness hypothesis is correct, there will be associations between creativity (and related cognitive variables) and indicators of genetic quality, and these indicators may be associated with health (Zahavi, 1975; Miller, 2000). One of the cognitive performance indicators most associated with health is intelligence (possibly the cognitive part of creativity). Higher intelligence is related to longevity and a lower risk of suffering from certain diseases, such as hypertension, heart problems, and Alzheimer’s disease (Arden et al., 2009a, 2015; Deary et al., 2019). That can be explained by the fact that intelligent people have healthier behaviors and would be subject to less risk (Gale et al., 2010).
Biological factors can also explain the relationship between intelligence and health. Deleterious mutations cause instabilities in the organism’s development, impairing brain development and cognitive performance (Gajos and Beaver, 2017). Paternal age is a known indicator of detrimental mutations (Gajos and Beaver, 2017). Children of older fathers are more at risk of having autism, attention-deficit/hyperactivity disorder, bipolar disorder, schizophrenia, and depression (D’Onofrio et al., 2014; Gajos and Beaver, 2017; Woodley of Menie and Kanazawa, 2017). More specifically, older fathers tend to have a little less intelligent children (even controlling for other variables), with a reduction of.84 to 1.23 points in g factor for each additional decade in paternal age since conception (Arslan et al., 2014; D’Onofrio et al., 2014; Woodley of Menie, 2015; Gajos and Beaver, 2017).
It is expected that characteristics affected by instabilities in development are interrelated. For instance, developmental disturbances lead to higher bilateral asymmetry in the body; the higher the asymmetry, the lower the IQ (Banks et al., 2010). Genetic mutations that hinder normal development also impact the quality of sperm (Jeffery et al., 2016). One study found a positive correlation between sperm quality and intelligence (Arden et al., 2009b); however, another, more recent one, did not (DeLecce et al., 2020).
Geary (2019) suggested that detrimental mutations that affect cognitive performance involve small inefficiencies in cellular processes. More precisely, genetic problems would affect the workings of cellular organelles, such as mitochondria, in charge of cell energy production and operation of the immune system and the brain; difficulties in cellular energy production would affect neurons, which in turn would impact brain functioning (Geary, 2019). Up to now, this notion seems largely speculative, though promising (see Savi et al., 2020; Ujma and Kovacs, 2020). However, other studies provide little supporting evidence for this hypothesis, while some studies have even reported contradictory evidence. For example, Mosing et al. (2015) showed significant correlations between musical aptitude and genetic quality measures only in females, when one would expect to find this result also in men.
Health and cognitive performance may also be associated in other species. For instance, better cognitive performance in bees often indicates an absence of parasites (Gegear et al., 2006). That happens because fighting infectious agents is as costly as investing in complex cognition; thus, there is a trade-off between immune function and cognitive performance (Boogert et al., 2011). Thus, being healthy, and having good learning and problem-solving skills simultaneously, can reflect high levels of genetic quality, or low parasite load. That is compatible with evidence showing that males who are better at problem-solving and learning and have better inhibitory ability have healthier offspring and are, on average, preferred in mating contexts (Spencer et al., 2005; Minter et al., 2017).
Though cognitive qualities imply health, these qualities may be valuable in the mating market for providing more advantages in obtaining resources that may later increase reproductive success (Stephen and Luoto, 2021). Cauchard et al. (2017) suggest this by showing that males of a bird species with better cognitive performance care more about their offspring. However, this hypothesis does not explain why females of promiscuous species (i.e., in which the females do not need the male’s resources) also mate with males having good cognitive qualities (Borgia, 1986; Keagy et al., 2011, 2012).
Fitness indicators vary more than traits evolved for other functions not related to fitness. This variation is due to the number of genes associated with these traits (pleiotropy), the susceptibility to mutations that affect development, and the sexual selection pressure that selects the trait according to the “more is better” logic (Miller, 2000). The heritability of attributes, manifestations, and achievements in a creative domain in men and women suggests that creativity can be a fitness indicator (details in Table 1).
If creativity is a mental adaptation evolved by sexual selection to indicate fitness potential, then it is expected to show high variability in the population due to the large number of genes involved in the expression of this adaptation (pleiotropy; Miller, 2000). Creativity shows high heritability and high variability in men and women, depending on the creative domain considered (see Miller, 2013; Varella et al., 2017; Table 1).
Yet what role does creativity have in attractiveness? Are there sex differences in the role of creativity in attractiveness? Men seem more interested than women in creative activities, and more engaged than women in creative behaviors in the past, according to a scale used to measure interest in creative activities and creative behaviors (Beaussart et al., 2012). Men are also more creative with unexpected flirting behaviors (White et al., 2018). A recent meta-analysis showed a male advantage in creative performance that the authors attributed to cultural factors (Hora et al., 2021). However, previous reviews found no sex/gender differences in creative ability or creative achievement in general but in specific domains of creativity (Baer and Kaufman, 2008).
Why are men more engaged and prominent in art than women, despite their similar creative performance? Men and women can use their creative potential in different ways. For instance, in comedy women use their creativity more in assessing humor, while men use theirs more in producing humor (Greengross et al., 2020). Similarly, one study mentioned the presence of more women (69%) than men in the front rows of music concerts (Sluming and Manning, 2000).
Sociocultural factors can also play a role in these sex differences. Indeed, men are better represented in artistic fields, but it is also a fact that for most of history, women have had fewer chances and fewer incentives to engage and achieve prominence in art (Varella et al., 2017; Rosenthal and Ryan, 2022). A meta-analysis of cross-cultural studies shows that women are “more artistic,” which suggests that sex differences in engagement in these fields may vary culturally (Ellis et al., 2008). Varella et al. (2010) found that women actually appreciate more than men an unknown instrumental piece of music, that women report to appreciate more music in general than men, and that women also report singing more than men. Another study showed a greater number of women in samples of gifted students in art-related courses (Holahan et al., 1995). Given the divergent findings, further studies are needed to verify whether sex differences concerning artistic manifestations exist and whether are more explainable by psychological or cultural factors (Hora et al., 2021).
Anecdotal evidence suggests that creativity is attractive. Creative geniuses in art and science (e.g., Lord Byron, Albert Einstein, Pablo Picasso, Van Gogh, and Charles Chaplin) are known for having had many casual sex partners, marriages, and children (Karamihalev, 2013). We have already shown evidence that creativity is universally attractive, but in this section, we will discuss specific evidence in more detail.
It is difficult to say whether this is a causal relationship or merely an association, but studies have shown that, for example, men with a larger artistic output (e.g., poets and painters) have a larger number of sexual partners (Clegg et al., 2011; Beaussart et al., 2012; Lange and Euler, 2014). Mosing et al. (2015) showed that boys had higher music achievement than girls. Furthermore, there was a negative association between sociosexuality, music aptitude and achievement in both sexes (Mosing et al., 2015). The authors indicated that these results are in accordance with the mutual mate choice model, in which both sexes utilize music to attract partners in a long-term reproductive strategy. But other studies show the opposite in terms of reproductive strategy. For instance, female and male poets and painters had more sexual partners (i.e., short-term reproductive strategy) than controls from other non-artistic professions (Nettle and Clegg, 2006). After reading vignettes describing a man with different levels of creativity and resources, women consider creative men with fewer resources as more attractive than less creative men with abundant resources; an limitation of this study was the low number of participants (41 women; Haselton and Miller, 2006). Male faces become more attractive if presented alongside creative text or music (Marin et al., 2017; Watkins, 2017; Marin and Rathgeber, 2022). The attributed attractiveness is even greater when men produce more complex (compared with less complex) musical patterns (e.g., Charlton, 2014; see Bongard et al., 2019). Furthermore, men, more than women, prefer songs with more complex, technical musical patterns and those less present in popular styles of music (Colley, 2008; Ord, 2020). The male preference for complex and technical music may be explained as a consequence of the evolved aesthetic propensities to impress women (which contradict the mutual mate choice model). An alternative explanation is that observed male preference is a byproduct with no signaling component per se. Artists and other creative people tend to be more open to experiences than other professionals, and more open people are likely to be more erotically inclined and less sexually restricted (Natividade and Hutz, 2016).
But the artistic propensities of both sexes can be used to attract mates (intersexual selection) and to compete for mates (intrasexual competition; Varella et al., 2022). For example, women propensities to visual and circus arts were related to intersexual selection, while literary and musical arts were related to both elevated inter-and intrasexual selection (Varella et al., 2022). In men, circus arts were related intersexual selection and visual arts with intrasexual competition (Varella et al., 2022).
Everyday displays of creativity also seem to impact attractiveness. For instance, women are attracted by men who can employ metaphors, make them laugh at jokes, and have significant verbal prowess (Lange et al., 2014; Gao et al., 2017; Greengross et al., 2020). Summing up, ornamental signs of creativity seem to be mainly linked to male attractiveness, though some suggest they also increase the attractiveness of creative women (Kaufman et al., 2016). However, some studies contradict the association between creativity, openness, and unrestricted sexuality. For instance, though better musical performance increases attractiveness (Madison et al., 2018), individuals with a more prominent musical output take longer to have their first intercourse, and women have fewer sexual partners as their musical output grows (Mosing et al., 2015).
In general, costly signaling develops in promiscuous species, in which the only expected male investment is genetic quality. Humans are diversified in their reproductive strategies and may enter short-term relationships in some situations, a kind of partnership akin to the promiscuity of other species (Gangestad and Simpson, 2000; Schmitt, 2005; Buss and Schmitt, 2019). Hence, if creativity signals good genes, it is expected that creative and original people are more attractive for casual relationships, as it seems to occur among bowerbirds (Borgia, 1986; Miller, 2001; Keagy et al., 2009). Findings regarding a preference for creative partners for short-term relationships are inconclusive at best. Some studies propose that women prefer creative men for long-term partnerships (Madison et al., 2018), for short-term ones (Haselton and Miller, 2006; Charlton, 2014; Mosing et al., 2015), or both (Griskevicius et al., 2006; Prokosch et al., 2009).
For instance, Madison et al. (2018) and Charlton (2014) have shown that presenting men with music increased their attractiveness for short-term relationships. Prokosch et al. (2009) recorded men as they performed four activities demanding verbal intelligence and creativity and then showed the videos for women to evaluate them. Intelligence predicted male attractiveness for long-term relationships, while creativity predicted attractiveness for short-and long-term relationships (Prokosch et al., 2009). Similarly, in two studies, male artists had a greater interest in long-term relationships, but one of the studies showed an association between being more successful in the career and the larger quantity of children, a sign of a larger reproductive effort characteristic of short-term relationships (Clegg et al., 2011; Mosing et al., 2015). The attractiveness of creativity in short-and long-term relationships may contradict the relationship between creativity and good genes; but it may also indicate that, precisely because it signals good genes, creativity becomes attractive in long-term relationships, in which people are more demanding (for other evolutionary theories, see Dissanayake, 2008; Luoto, 2019a; Mehr et al., 2021; Savage et al., 2021).
If creativity is a kind of costly signaling, creative men will be more desirable to women in the fertile window of the menstrual cycle. The higher chance of getting pregnant at this time would heighten sexual appeal (Stern et al., 2021) and the preference for men with a better genetic constitution (see Thomas et al., 2021). Some evidence confirms that women in their fertile window would rather have casual relationships with creative men (e.g., Charlton, 2014; Marin et al., 2017). Haselton and Miller (2006) showed that women in the fertile window preferred short-term relationships with creative men (regardless of the men’s amount of resources). Furthermore, women were more creative during their fertile phase (Galasinska and Szymkow, 2021; Galasinska and Szymkow, 2022).
Some psychopathologies may be dysfunctional expressions of ornamental creativity as if the fitness indicator had failed or overshot its optimum. Indeed, people with schizotypal traits attract partners using metaphors and verbal proficiency (Lange et al., 2016; Gao et al., 2017). On the other hand, people with schizophrenia have cognitive and linguistic problems that make it difficult to express such “verbal ornaments” and, therefore, also impair their attractiveness (Shaner et al., 2004). Schizophrenic symptoms peak at puberty, when levels of circulating androgens increase, influencing secondary sexual traits. The increase in symptom severity is related to dopaminergic antagonists, which in humans and other animals are associated with sexuality and reproduction (Shaner et al., 2004; Del Giudice et al., 2010). In addition to being associated with creativity, schizotypal and autistic traits would theoretically be linked to short-and long-term reproductive strategies, respectively (Del Giudice et al., 2010). That may explain the relationship between creativity, personality, and sexual selection. Individuals with an autistic phenotype invest more in offspring and favor long-term relationships (Del Giudice et al., 2014; Ponzi et al., 2016). Autistic creativity is mainly characterized by convergent thinking, exhibiting greater pragmatic creativity. According to Del Giudice et al. (2010), the prevalence of autism is currently linked to the selection of genes that are associated with greater systematization, greater attention to detail, and the ability to innovate in technical fields, which may have become more helpful from the Holocene, a period characterized by agriculture (Harpending and Cochran, 2002). However, attention to detail is also crucial in archeological cave paintings (Kellman, 1998; Spikins et al., 2018). These psychological skills would be worth greater prestige and access to resources in these cultural contexts, also leading to a higher number of mating opportunities for individuals with these skills, particularly men (Henrich and Gil-White, 2001; Del Giudice et al., 2010). That would suggest that the reproductive benefit of this kind of creativity lies in facilitating the acquisition of resources, not necessarily in signaling good genes.
On the other hand, schizotypal individuals may have occupied the role of shamans in traditional societies (Dein and Littlewood, 2011). That would explain ancestral artistic displays as part of religious rituals rather than pure art, the latter being more common in contemporary art (Høgh-Olesen, 2018). Theoretically, artistic skills and a more original personality would improve access to short-term relationships, which is difficult to infer from ancestral societies, yet studying a hunter-gatherer population, Smith et al. (2017) found that the value of good storytellers is reflected in the fact that they also have increased reproductive success and receive more resources than less-skilled storytellers. It is also conceivable that this ornamental creativity associated with schizotypy does not guarantee many privileges in these societies. A study of the Meru, a semi-nomadic tribe in Kenya, showed that more creative people had more resources, although they had fewer children, fewer grandchildren, and fewer wives/husbands (Lebuda et al., 2021). That is the opposite of what is expected from an adaptive perspective, namely that creativity leads to reproductive success. One possible explanation is that the attraction to creative partners is recent in human history, in societies that value disruption, innovation, and rapid change. Traditional societies value stability, traditions, and rules. Or, yet, creativity was adaptive from a sexual selection point of view in our environment of evolutionary adaptedness (EEA), but is not adaptive anymore because societies and environments (including the Meru) have changed so drastically.
Socioecological stimuli and contexts exert an important influence on creativity. The stimuli and contexts that inspire men and women to perform more creatively may reveal the influence of sexual selection. For instance, men behave more creatively (produce better creative descriptions of abstract paintings) after seeing photos of attractive women. That suggests that a romantic stimulus activates cognitive/neural mechanisms linked to reproduction, leading men to perform better on tasks that might impress women. The increase in male creativity was maintained even when participants performed creative tasks after imagining themselves having a short-term or long-term relationship (Griskevicius et al., 2006). Nevertheless, female creativity increased in the face of greater assurance that the potential partner would invest in a long-term relationship, which may indicate that creativity was selected in the female sex to deal with a reproductive strategy more focused on attracting investment from partners in a committed relationship (Griskevicius et al., 2006).
This study reinforces the idea that creativity evolved by sexual selection in men and women (Baer and Kaufman, 2008; Varella et al., 2011, 2014, 2017; Miller, 2013). The increase in female creativity, given the possibility of short-and long-term relationships, indicates that ornamental creativity may have evolved for signaling both good genes and an ability to acquire resources.
It is common for females to select males based on their ability to contribute resources to offspring (Andersson, 1994; Davis and Arnocky, 2022). Parental care is very important to the human species, and resources are one of the forms of parental investment (Andersson, 1994), which is why men with more access to resources are universally considered more attractive, all other things being equal (Buss and Schmitt, 2019; Walter et al., 2020), regardless of the level of gender equality (Zhang et al., 2019). Indeed, intelligence and creativity are critical to activities that implicate access to resources, e.g., academic and professional activities, increasing attractiveness in modern societies (see Lebuda et al., 2021). Intelligence and creativity may not be attractive in themselves but as markers of resource-related potential. Thus, it is possible that intelligence and creativity are not considered attractive in societies (as may be the case with the Meru) where access to resources does not depend on intelligence or creativity.
A possible counter-argument is that even if creativity is attractive because it increases access to resources, this would not necessarily exclude its possible role as an indicator of good genes (Luoto et al., 2019a). After all, individuals able to obtain resources in a given context must be healthy enough to participate in activities that require physical and cognitive effort. As a consequence, it may be that the more attractive women think they are, the more they prefer men who rank high on signs of health and social status, because women’s attractiveness is a fungible currency on the mating market that can be “exchanged” for traits that women seek (Buss and Shackelford, 2008). Creativity can be a reliable sign of both health and status. This would explain why women prefer creative men for short-term and long-term relationships (e.g., Griskevicius et al., 2006; Prokosch et al., 2009).
According to Winegard et al. (2018), cognitive resources increase attractiveness only when they provide culturally valuable assets. More precisely, inventing new technologies or artistic products confer prestige, which can be “exchanged” for resources and mate partners (Henrich and Gil-White, 2001). This model emphasizes intrasexual competition more than intersexual selection (Winegard et al., 2018). Intrasexual and intersexual competition are two main mechanisms of sexual selection (Darwin, 1871; Puts, 2016). In intrasexual competition, individuals (usually males) compete with others of the same sex for access to the opposite sex, which leads to the evolution of ornaments (e.g., plumage, singing, dancing) and/or weapons (e.g., horns, talons, fangs); whereas in intersexual selection, males mostly display their ornaments and weapons directly to the opposite sex (Berglund et al., 1996). In humans, males and females are involved with intersexual selection and intrasexual competition (see, e.g., Stewart-Williams and Thomas, 2013). It is possible that ornamental creativity evolved to be useful in intrasexual competition as a way of impressing other men, in addition to being useful in intersexual selection (Winegard et al., 2018). Accordingly, Winegard et al. (2018) mention historical, philosophical, and literary treatises written when most women were not able to read. The contents of such treatises do not seem to appeal to female interests (as pointed out by feminist authors; e.g., Irigaray, 1985) since they deal with war, politics, military strategy, and metaphysics. Men who were more prolific in these fields garnered more prestige among other men, securing access to valuable social assets, such as resources and protection for themselves and their families (Henrich and Gil-White, 2001; Winegard et al., 2018). Thus, men with superior cognitive abilities will also tend to rise in social status hierarchies.
This model based on competition for status shows advantages over the “cultural courtship” model championed by Miller (2000). Miller (2013) states that men and women have been shaped by evolution to display mental abilities indicative of genetic quality (see also Stewart-Williams and Thomas, 2013). A possible flaw in this hypothesis is the belief that humans have always chosen their partners freely and individually. Actually, for most of human history, and still in most traditional cultures, marriages are arranged, i.e., the bride’s family, usually fathers and brothers-in-law, interfere in the groom’s choice (Apostolou, 2017). In other words, throughout history, mate selection has been about advertising one’s attractiveness to other men.
Many male physical traits seem to bear on dominance, but not necessarily on attractiveness, such as a male face, beard, and muscles (Puts, 2010). Creativity seems to have its use in intrasexual competition, and lyrics about the male world seem to illustrate that (e.g., Black Sabbath, ACDC, and Metallica); but it is also true that some such cultural expressions are directed toward the opposite sex (e.g., ‘N Sync and Back Street Boys; see Winegard et al., 2018). For example, among professional male guitarists, the time spent playing the instrument positively predicted desire for casual sex with women, and the speed in playing positively predicted a desire to impress other men (DeLecce et al., 2022). Varella et al. (2022) showed that men and women use their artistic propensities in intersexual selection (female-biased) and intrasexual competition (male-biased). Creativity and aesthetic sense also probably participated in the female intrasexual competition since body beautification is universally crucial in female attractiveness (Varella et al., 2017).
The theories of costly signaling and mental fitness (Zahavi, 1975; Miller, 2000, 2001) have become very popular in explaining the existence of abnormal phenotypes in many species, including creative manifestations in humans (Luoto et al., 2019a). However, Darwin (1871) believed something else: for him, sexual selection picked flashy traits for arbitrary aesthetic reasons (Prum, 2012; Davis and Arnocky, 2022). There is evidence that Darwin’s insight was correct. In many species, individuals are selected to mate through a process called sensory exploration (Verpooten and Nelissen, 2012). For instance, female guppies tend to copulate with males with more markedly orange spots around their bodies. That is not so because these males are genetically better, but because their orange spots co-opt the female’s sensory system, shaped by natural selection to find food of the same color (see Verpooten and Nelissen, 2012). This sensory co-optation process may lead to a “runaway selection,” in which each new generation of males develops phenotypes ever more extravagant and unrelated to any underlying aptitude (Prum, 2012).
Showy phenotypes may also arise if they make the offspring attractive (this is known as the “sexy son hypothesis”). In this process, the alleles of the most selective females spread and are inherited by their offspring, as daughters become more selective and sons showier (Prokop et al., 2012). Such dynamics feedback on themselves on the grounds of attractiveness advantage (which is, in this case, more arbitrary than in the selection based on “good genes”). Hence, while sexual selection based on the “sexy son” phenomenon promises more reproductively successful offspring for being more attractive according to arbitrary patterns, selection based on “good genes” provides offspring that are successful because of better health (Prokop et al., 2012).
However, to Miller (2001), the process of sensory exploration (and, one assumes, the benefits based on “sexy son”) would not continue to be arbitrary under all possible scenarios. Miller (2000) has suggested that ornaments would become so intricate that they will come to depend on the expression of ever more genes; at this stage in complexity, pleiotropy would grow, as well as the threshold of genetic quality required to go on sporting an ever costlier phenotype. Furthermore, sexual selection based on sensory exploration should be more common in not very social species, where the first step to mating is finding a partner. However, primates are social species; thus, locating a potential partner is no problem. On the contrary, the hard part is selecting the best option (Miller, 2000; Verpooten and Nelissen, 2012). Thus, if Miller (2000) is correct, genetic quality becomes a part of sexual selection at some point.
What are the implications of this plurality of mechanisms of sexual selection for the evolution of creativity? Creativity may have evolved by viability selection and also by sexual selection (Luoto et al., 2019a), but the details of this transition are uncertain. Creativity may have begun to grow initially due to general cognitive growth, which is useful for problem-solving. Its effects would have then started to affect other traits that are more useful in partner selection, such as the ability to get food (which in Neolithic humans may happen as social status increases: Winegard et al., 2018). The pressure on expanding cognitive performance would have enhanced this ability to the point that it could only be sustained by individuals having at least enough genetic quality to afford its energy costs (Miller, 2000). From a certain point on, practical benefits of cognitive ability do not increase together with the growth in cognitive ability, which is when conspicuous (i.e., ornamental, fruitless) forms of cognitive ability and creativity to exhibit fitness may arise.
Ornamental traits may be an exaptation, i.e., pragmatic creative capacities evolved initially by viability selection and were later co-opted by sexual selection as phylogenetic exaptations related to aesthetic and artistic production and appreciation (Varella et al., 2011; Luoto et al., 2019a). For instance, probably the first handaxes built by human ancestors would have been used to assist in taking down prey and cutting up carcasses; however, they would gradually be co-opted for aesthetic uses, acquiring ornaments and a more symmetrical look (Mithen, 2003). As an extension of this point of view, creative individuals can reap advantages through functional and/or ornamental extended phenotypes that they have created or acquired. For instance, clothes, cars, and houses were invented for practical reasons (e.g., protection, locomotion) and later acquired ornamental functions that signal (and extend) personal attributes, such as interest in certain forms of romantic involvement, intelligence, and creativity (Luoto, 2019a).
The role of culture in creativity may go beyond exaptations or extended phenotypes. Recently, many traits have been studied that would result from gene–culture coevolution (Bender, 2019). Briefly, this evolutionary process is characterized by selecting genes based on cultural pressures. The evolution of the ability to write is an enlightening example. Human beings did not evolve to read and write. Writing is a result of learning specific cultural techniques that co-opt brain areas shaped initially to deal with other adaptive issues (Parkinson and Wheatley, 2015); however, as soon as writing spreads and starts generating social benefits, genes associated with a greater ability to learn to read and write were selected (Overmann, 2016). This process in which learned responses acquire a genetic base is called the Baldwin effect, which has been used to explain complex aspects of human cognition (Baldwin, 2018).
That process may have supported the fast growth in complexity in hominine cognitive abilities (see recent proposals of this idea in the context of music: Podlipniak, 2017, 2021; Mehr et al., 2021; Savage et al., 2021). Neuroplasticity may have allowed learning ornamental techniques that initially aimed at reinforcing intergroup cohesion and establishing some form of individual or group identity (Garofoli, 2015). These distinctive ornaments may have begun to bring reproductive advantages for cultural reasons, which is when Baldwin effect’s ability to produce ever more complex ornaments may have been genetically incorporated. The use of the Baldwin Effect here is admittedly speculative. Further studies are needed to explore the relations between culture, plasticity, genetics, and evolution.
Did creativity evolve by sexual selection? This article aimed to answer this question considering a pluralistic Tinbergian perspective (Tinbergen, 1963; Fitch, 2015) and a nomological network of evidence (Schmitt and Pilcher, 2004; Konner, 2021). That is the most up-to-date and comprehensive review to integrate and organize an interdisciplinary body of evidence to answer this question about creativity. In summary, our findings suggest that sexual selection likely influenced the evolution of creativity (Table 1); however, the sub-process behind this influence is unclear. Creativity has multiple adaptive functions regarding sexual selection (for an example on musicology, see Fitch, 2015). More specifically, creativity may have evolved by sexual selection not only because it indicates good genes, potential to acquire resources, dominance, or because it is a useful ability in the production of artifacts (e.g., bowers, paintings, music) that co-opt sensory biases to attract attention. It is probable that creativity is connected to all of these aspects. Evolutionary psychologists must test hypotheses derived from processes of sexual selection other than good genes and costly signaling (Luoto, 2019a; Davis and Arnocky, 2022).
Evidence related to sex differences supports that both sexes have developed fitness indicators (Miller, 2013; see Table 1). Overall, no evidence was found of average differences in creative ability or achievement between adult men and women (Baer and Kaufman, 2008), but sexual dimorphism shows up when specific creative domains are evaluated (Ellis et al., 2008; Varella et al., 2010; Savage et al., 2015; Greengross et al., 2020; Hora et al., 2021; Nakano et al., 2021). Other evidence suggests greater creativity among women (Varella et al., 2017; Nakano et al., 2021). Women showed higher mean and greater variability, compared with men, in creative tasks involving language; men showed higher mean and greater variability in creative tasks involving figures and drawing (He et al., 2015; Lau and Cheung, 2015; Karwowski et al., 2016a,b; Taylor and Barbot, 2021). Such sexual differences are compatible with those found in academic and professional preferences, in which women predominate in areas that involve language and men, in areas that involve spatial reasoning (e.g., Wright et al., 2015).
Furthermore, greater variability appears positively associated with greater phenotypic plasticity, which has been linked to the action of androgenic and dopaminergic systems (Del Giudice et al., 2018; Janicke et al., 2021; see Table 1). Creativity is related with greater phenotypic plasticity (Feist, 2019), dopaminergic system (Reuter et al., 2006; Runco et al., 2011; Mayseless et al., 2013), and testosterone levels in both sexes (Hassler, 1992; Sluming and Manning, 2000; Crocchiola, 2014). However, paradoxically, testosterone promotes left brain lateralization, but creativity is linked to right lateralization (Beking et al., 2018). Future studies should investigate the possible role of androgens in male and female creativity and the mechanisms that promote this association.
Evidence on the relationship between psychopathology and creativity supports the predictions of the mental fitness theory (Miller and Todd, 1998; Miller, 2001). Disorders such as schizophrenia, bipolarity, and autism seem to be dysfunctional manifestations of schizotypal and autistic phenotypes linked to creativity, possibly due to genetic predisposition, developmental instabilities, and evolutionarily novel lifestyle factors such as low-grade systemic inflammation and chronic stress (Rantala et al., 2021, 2022).
Evidence shows that creativity is sexy: it is considered attractive in potential mating partners, at least in WEIRD societies (Buss, 1989; Li et al., 2011; Kaufman et al., 2016; Souza et al., 2016). Future studies should verify the role that creativity plays in attractiveness in traditional societies (e.g., Lebuda et al., 2021). Furthermore, future studies should verify the theoretical framework (e.g., good genes) that explains the attractiveness and evolutionary function of different types of creativity (e.g., aesthetic and pragmatic).
According to the mental fitness theory, ornamental manifestations of creativity would be more attractive in short-term relationships (e.g., Haselton and Miller, 2006; Charlton, 2014; Mosing et al., 2015) because they indicate good genes. But other studies suggest that ornamental creativity may also evolved to retain partners in a long-term relationship (Varella et al., 2017; Winegard et al., 2018). Alternatively, artistic capacities could have evolved because they are able to produce something that the human mind finds beautiful (Darwin, 1871; Prum, 2012).
Despite making an important contribution to the literature on mating preferences by bringing together multiple studies and interpreting how much they support the hypothesis of the evolution of creativity by sexual selection, there are limitations to this review, which future studies may overcome. The first is that, despite the extent of the review performed here, this article is not a systematic review or a meta-analysis.
A second limitation is the focus on mental fitness theory. This focus resulted from our deliberate choice to consider theories and evidence supporting (or not) the evolution of creativity by sexual selection. It was for this reason that we added a discussion about creativity being attractive because it signals good genes or because it almost always (at least in WEIRD societies) equates to resource acquisition. Future evolutionary research on creativity should compare evidence related to sexual selection and other theories, such as exaptations, co-opted by-product and cultural evolution (Luoto, 2019a). This review has gone a long way toward integrating and enhancing our understanding of ornamental creativity as a possible sexual selected psychological trait.
FN: main argument, writing, and formatation. JN: main argument and revision. All authors contributed to the article and approved the submitted version.
We would like to appreciate for the enriching advice provided by both peers and the editor of the volume. Without these
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abraham, A. (2014). Is there an inverted-U relationship between creativity and psychopathology? Front. Psychol. 5:750. doi: 10.3389/fpsyg.2014.00750
Abraham, A. (2016). Gender and creativity: an overview of psychological and neuroscientific literature. Brain Imaging Behav. 10, 609–618. doi: 10.1007/s11682-015-9410-8
Abu-Akel, A., Webb, M. E., de Montpellier, E., Von Bentivegni, S., Luechinger, L., Ishii, A., et al. (2020). Autistic and positive schizotypal traits respectively predict better convergent and divergent thinking performance. Think. Skills Creat. 36:100656. doi: 10.1016/j.tsc.2020.100656
Acar, S., Chen, X., and Cayirdag, N. (2018). Schizophrenia and creativity: a meta-analytic review. Schizophr. Res. 195, 23–31. doi: 10.1016/j.schres.2017.08.036
Acar, S., and Runco, M. A. (2012). Psychoticism and creativity: a meta-analytic review. Psychol. Aesthet. Creat. Arts 6, 341–350. doi: 10.1037/a0027497
Acar, S., and Sen, S. (2013). A multilevel meta-analysis of the relationship between creativity and schizotypy. Psychol. Aesthet. Creat. Arts 7:214. doi: 10.1037/a0031975
Acevedo, B. P., Poulin, M. J., Collins, N. L., and Brown, L. L. (2020). After the honeymoon: neural and genetic correlates of romantic love in newlywed marriages. Front. Psychol. 11:634. doi: 10.3389/fpsyg.2020.00634
Aguilera, M., and Rodríguez-Ferreiro, J. (2021). Differential effects of schizotypy dimensions on creative personality and creative products. Creat. Res. J. 33, 202–208. doi: 10.1080/10400419.2020.1866895
Akbari Chermahini, S., and Hommel, B. (2009). The (b) link between creativity and dopamine: spontaneous eye blink rates predict divergent, but not convergent thinking. Cognition 115, 458–465. doi: 10.1016/j.cognition.2010.03.007
Amabile, T. M., and Gitomer, J. (1984). Children’s artistic creativity: effects of choice in task materials. Personal. Soc. Psychol. Bull. 10, 209–215. doi: 10.1177/0146167284102006
Apostolou, M. (2017). Individual mate choice in an arranged marriage context: evidence from the standard cross-cultural sample. Evol. Psychol. Sci. 3, 193–200. doi: 10.1007/s40806-017-0085-9
Arden, R., Gottfredson, L. S., and Miller, G. (2009a). Does a fitness factor contribute to the association between intelligence and health outcomes? Evidence from medical abnormality counts among 3654 US veterans. Intelligence 37, 581–591. doi: 10.1016/j.intell.2009.03.008
Arden, R., Gottfredson, L. S., Miller, G., and Pierce, A. (2009b). Intelligence and semen quality are positively correlated. Intelligence 37, 277–282. doi: 10.1016/j.intell.2008.11.001
Arden, R., Luciano, M., Deary, I. J., Reynolds, C. A., Pedersen, N. L., Plassman, B. L., et al. (2015). The association between intelligence and lifespan is mostly genetic. Int. J. Epidemiol. 45, 178–185. doi: 10.1093/ije/dyv112
Aron, A., Fisher, H., Mashek, D. J., Strong, G., Li, H., and Brown, L. L. (2005). Reward, motivation, and emotion systems associated with early-stage intense romantic love. J. Neurophysiol. 94, 327–337. doi: 10.1152/jn.00838.2004
Arslan, R. C., Penke, L., Johnson, W., Iacono, W. G., and McGue, M. (2014). The effect of paternal age on offspring intelligence and personality when controlling for parental trait levels. PLoS One 9:e90097. doi: 10.1371/journal.pone.0090097
Askew, G. N. (2014). The elaborate plumage in peacocks is not such a drag. J. Exp. Biol. 217, 3237–3241. doi: 10.1242/jeb.107474
Baer, J., and Kaufman, J. C. (2008). Gender differences in creativity. J. Creat. Behav. 42, 75–105. doi: 10.1002/j.2162-6057.2008.tb01289.x
Bandini, E., and Harrison, R. A. (2020). Innovation in chimpanzees. Biol. Rev. 95, 1167–1197. doi: 10.1111/brv.12604
Banks, G. C., Batchelor, J. H., and McDaniel, A. (2010). Smarter people are (a bit) more symmetrical: a meta-analysis of the relationship between intelligence and fluctuating asymmetry. Intelligence 38, 393–401. doi: 10.1016/j.intell.2010.04.003
Barbot, B., and Tinio, P. P. L. (2015). Where is the “g” in creativity? A specialization–differentiation hypothesis. Front. Hum. Neurosci. 8:1041. doi: 10.3389/fnhum.2014.01041
Baron-Cohen, S., Wheelwright, S., Skinner, R., Martin, J., and Clubley, E. (2001). The autism-Spectrum quotient (AQ): evidence from Asperger syndrome/high-functioning autism, males and females, scientists and mathematicians. J. Autism Dev. Disord. 31, 5–17. doi: 10.1023/A:1005653411471
Beaussart, M. L., Kaufman, S. B., and Kaufman, J. C. (2012). Creative activity, personality, mental illness, and short-term mating success. J. Creat. Behav. 46, 151–167. doi: 10.1002/jocb.11
Beking, T., Geuze, R. H., Van Faassen, M., Kema, I. P., Kreukels, B. P. C., and Groothuis, T. G. G. (2018). Prenatal and pubertal testosterone affect brain lateralization. Psychoneuroendocrinology 88, 78–91. doi: 10.1016/j.psyneuen.2017.10.027
Bender, A. (2019). The role of culture and evolution for human cognition. Top. Cogn. Sci. 12, 1–18. doi: 10.1111/tops.12449
Benz, S., Sellaro, R., Hommel, B., and Colzato, L. S. (2016). Music makes the world go round: the impact of musical training on non-musical cognitive functions—a review. Front. Psychol. 6:2023. doi: 10.3389/fpsyg.2015.02023
Berglund, A., Bisazza, A., and Pilastro, A. (1996). Armaments and ornaments: an evolutionary explanation of traits of dual utility. Biol. J. Linn. Soc. 58, 385–399. doi: 10.1111/j.1095-8312.1996.tb01442.x
Bicer, A., Chamberlin, S., and Perihan, C. (2021). A meta-analysis of the relationship between mathematics achievement and creativity. J. Creat. Behav. 55, 569–590. doi: 10.1002/jocb.474
Bispham, J. C. (2009). Music’s “design features”: musical motivation, musical pulse, and musical pitch. Music. Sci. 13, 41–61. doi: 10.1177/1029864909013002041
Bongard, S., Schulz, I., Studenroth, K. U., and Frankenberg, E. (2019). Attractiveness ratings for musicians and non-musicians: an evolutionary-psychology perspective. Front. Psychol. 10:2627. doi: 10.3389/fpsyg.2019.02627
Boogert, N. J., Fawcett, T. W., and Lefebvre, L. (2011). Mate choice for cognitive traits: a review of the evidence in nonhuman vertebrates. Behav. Ecol. 22, 447–459. doi: 10.1093/beheco/arq173
Borgia, G. (1986). Sexual selection in bowerbirds. Sci. Am. 254, 92–100. doi: 10.1038/scientificamerican0686-92
Boyd, B. (2010). On the Origin of Stories: Evolution, Cognition, and Fiction, Cambridge, MA: Harvard University Press.
Buss, D. M. (1989). Sex differences in human mate preferences: evolutionary hypotheses tested in 37 cultures. Behav. Brain Sci. 12, 1–14. doi: 10.1017/S0140525X00023992
Buss, D. M., Abbott, M., Angleitner, A., Asherian, A., Biaggio, A., Blanco-Villasenor, A., et al. (1990). International preferences in selecting mates: a study of 37 cultures. J. Cross-Cult. Psychol. 21, 5–47. doi: 10.1177/0022022190211001
Buss, D. M., and Barnes, M. (1986). Preferences in human mate selection. J. Pers. Soc. Psychol. 50, 559–570. doi: 10.1037/0022-3514.50.3.559
Buss, D. M., and Schmitt, D. P. (2019). Mate preferences and their behavioral manifestations. Annu. Rev. Psychol. 70, 23–34. doi: 10.1146/annurev-psych-010418-103408
Buss, D. M., and Shackelford, T. K. (2008). Attractive women want it all: good genes, economic investment, parenting proclivities, and emotional commitment. Evol. Psychol. 6, 134–146. doi: 10.1177/147470490800600116
Campbell, L., and Fletcher, G. J. (2015). Romantic relationships, ideal standards, and mate selection. Curr. Opin. Psychol. 1, 97–100. doi: 10.1016/j.copsyc.2015.01.007
Campbell, K., and Kaufman, J. (2017). Do you pursue your heart or your art? Creativity, personality, and love. J. Fam. Issues 38, 287–311. doi: 10.1177/0192513X15570318
Carter, C., Hass, R. W., Charfadi, M., and Dinzeo, T. J. (2019). Probing linear and nonlinear relations among schizotypy, hypomania, cognitive inhibition, and creativity. Creat. Res. J. 31, 83–92. doi: 10.1080/10400419.2019.1580091
Castelli, F., Frith, C., Happé, F., and Frith, U. (2002). Autism, Asperger syndrome and brain mechanisms for the attribution of mental states to animated shapes. Brain: A J. Neurol. 125, 1839–1849. doi: 10.1093/brain/awf189
Catchpole, C. K. (1987). Bird song, sexual selection and female choice. Trends Ecol. Evol. 2, 94–97. doi: 10.1016/0169-5347(87)90165-0
Cattell, R. B. (1963). Theory of fluid and crystallized intelligence: a critical experiment. J. Educ. Psychol. 54, 1–22. doi: 10.1037/h0046743
Cauchard, L., Angers, B., Boogert, N. J., Lenarth, M., Bize, P., and Doligez, B. (2017). An experimental test of a causal link between problem-solving performance and reproductive success in wild great tits. Front. Ecol. Evol. 5:107. doi: 10.3389/fevo.2017.00107
Cauchard, L., Boogert, N. J., Lefebvre, L., Dubois, F., and Doligez, B. (2013). Problem-solving performance is correlated with reproductive success in a wild bird population. Anim. Behav. 85, 19–26. doi: 10.1016/j.anbehav.2012.10.005
Chang, L., Wang, Y., Shackelford, T. K., and Buss, D. M. (2011). Chinese mate preferences: cultural evolution and continuity across a quarter of a century. Personal. Individ. Differ. 50, 678–683. doi: 10.1016/j.paid.2010.12.016
Charlton, B. D. (2014). Menstrual cycle phase alters women’s sexual preferences for composers of more complex music. Proc. R. Soc. B Biol. Sci. 281:20140403. doi: 10.1098/rspb.2014.0403
Chen, J., Zou, Y., Sun, Y.-H., and Ten Cate, C. (2019). Problem-solving males become more attractive to female budgerigars. Science 363, 166–167. doi: 10.1126/science.aau8181
Cheung, P. C., and Lau, S. (2010). Gender differences in the creativity of Hong Kong school children: comparison by using the new electronic Wallach–Kogan creativity tests. Creat. Res. J. 22, 194–199. doi: 10.1080/10400419.2010.481522
Cho, S. H., Nijenhuis, J. T., Van Vianen, A. E., Kim, H. B., and Lee, K. H. (2010). The relationship between diverse components of intelligence and creativity. J. Creativity Behav. 44, 125–137. doi: 10.1002/j.21626057.2010.tb01329.x
Clegg, H., Miell, D., and Nettle, D. (2011). Status and mating success amongst visual artists. Front. Psychol. 2:310. doi: 10.3389/fpsyg.2011.00310
Coleman, S. W., Patricelli, G. L., Coyle, B., Siani, J., and Borgia, G. (2007). Female preferences drive the evolution of mimetic accuracy in male sexual displays. Biol. Lett. 3, 463–466. doi: 10.1098/rsbl.2007.0234
Colley, A. (2008). Young people’s musical taste: relationship with gender and gender-related traits. J. Appl. Soc. Psychol. 38, 2039–2055. doi: 10.1111/j.1559-1816.2008.00379.x
Conroy-Beam, D., Roney, J. R., Lukaszewski, A. W., Buss, D. M., Asao, K., Sorokowska, A., et al. (2019). Assortative mating and the evolution of desirability covariation. Evol. Hum. Behav. 40, 479–491. doi: 10.1016/j.evolhumbehav.2019.06.003
Coon, H., and Carey, G. (1989). Genetic and environmental determinants of musical ability in twins. Behav. Genet. 19, 183–193. doi: 10.1007/BF01065903
Crespi, B. J. (2016). Autism as a disorder of high intelligence. Front. Neurosci. 10:300. doi: 10.3389/fnins.2016.00300
Crocchiola, D. (2014). Art as an indicator of male fitness: does prenatal testosterone influence artistic ability? Evol. Psychol. 12, 521–533. doi: 10.1177/147470491401200303
Cropley, A. (2006). In praise of convergent thinking. Creat. Res. J. 18, 391–404. doi: 10.1207/s15326934crj1803_13
D’Onofrio, B. M., Rickert, M. E., Frans, E., Kuja-Halkola, R., Almqvist, C., Sjölander, A., et al. (2014). Paternal age at childbearing and offspring psychiatric and academic morbidity. JAMA Psychiat. 71, 432–438. doi: 10.1001/jamapsychiatry.2013.4525
Darwin, C. (1871). The Descent of Man and Selection in Relation to Sex. Princeton, NJ: Princeton University Press.
Davis, A. C., and Arnocky, S. (2022). Darwin versus Wallace: aesthetic evolution and preferential mate choice. Front. Psychol. 2588:862385. doi: 10.3389/fpsyg.2022.862385
De Block, A., and Dewitte, S. (2007). Mating games: cultural evolution and sexual selection. Biol. Philos. 22, 475–491. doi: 10.1007/s10539-006-9041-y
De Bolle, M., De Fruyt, F., McCrae, R. R., Löckenhoff, C. E., Costa, P. T. Jr., Aguilar-Vafaie, M. E., et al. (2015). The emergence of sex differences in personality traits in early adolescence: a cross-sectional, cross-cultural study. J. Pers. Soc. Psychol. 108:171. doi: 10.1037/a0038497
De Ridder, D., and Vanneste, S. (2013). The artful mind: sexual selection and an evolutionary neurobiological approach to esthetic appreciation. Perspect. Biol. Med. 56, 327–340. doi: 10.1353/pbm.2013.0029
Deary, I. J., Harris, S. E., and Hill, W. D. (2019). What genome-wide association studies reveal about the association between intelligence and physical health, illness, and mortality. Curr. Opin. Psychol. 27, 6–12. doi: 10.1016/j.copsyc.2018.07.005
Dein, S., and Littlewood, R. (2011). Religion and psychosis: a common evolutionary trajectory? Transcult. Psychiatry 48, 318–335. doi: 10.1177/1363461511402723
Del Giudice, M., Angeleri, R., Brizio, A., and Elena, M. R. (2010). The evolution of autistic-like and schizotypal traits: a sexual selection hypothesis. Front. Psychol. 1:41. doi: 10.3389/fpsyg.2010.00041
Del Giudice, M., Barrett, E. S., Belsky, J., Hartman, S., Martel, M. M., Sangenstedt, S., et al. (2018). Individual differences in developmental plasticity: a role for early androgens? Psychoneuroendocrinology 90, 165–173. doi: 10.1016/j.psyneuen.2018.02.025
Del Giudice, M., Klimczuk, A. C., Traficonte, D. M., and Maestripieri, D. (2014). Autistic-like and schizotypal traits in a life history perspective: diametrical associations with impulsivity, sensation seeking, and sociosexual behavior. Evol. Hum. Behav. 35, 415–424. doi: 10.1016/j.evolhumbehav.2014.05.007
DeLecce, T., Fink, B., Shackelford, T., and Abed, M. G. (2020). No evidence for a relationship between intelligence and ejaculate quality. Evol. Psychol. 18:1474704920960450. doi: 10.1177/1474704920960450
DeLecce, T., Pazhoohi, F., Szala, A., and Shackelford, T. K. (2022). Extreme metal guitar skill: A case of male–male status seeking, mate attraction, or byproduct? Evol. Behav. Sci.. Advance online publication. doi: 10.1037/ebs0000304
Dissanayake, E. (2008). “The arts after Darwin: does art have an origin and adaptive function?”, In World art Studies: Exploring Concepts and Approaches, eds K. Zijlemans and W. Dammevan Amsterdam: Valiz, 241–263.
Doi, H., Basadonne, I., Venuti, P., and Shinohara, K. (2018). Negative correlation between salivary testosterone concentration and preference for sophisticated music in males. Personal. Individ. Differ. 125, 106–111. doi: 10.1016/j.paid.2017.11.041
Ellis, L., Hershberger, S., Field, E., Wersinger, S., Pellis, S., Geary, D., et al. (2008). Sex Differences: Summarizing more than a Century of Scientific Research. New York: Taylor & Francis.
Endler, J. A. (2012). Bowerbirds, art and aesthetics: are bowerbirds artists and do they have an aesthetic sense? Commun. Integr. Biol. 5, 281–283. doi: 10.4161/cib.19481
Feist, G. J. (1998). A meta-analysis of personality in scientific and artistic creativity. Personal. Soc. Psychol. Rev. 2, 290–309. doi: 10.1207/s15327957pspr0204_5
Feist, G. J. (2001). Natural and sexual selection in the evolution of creativity. Bull. Psychol. Arts 2, 11–16.
Feist, G. J. (2004). “The evolved fluid specificity of human creative talent,” in Creativity: From Potential to Realization. eds. R. J. Sternberg, L. E. Grigorenko, and J. L. Singer (Washington, DC: American Psychological Association), 57–82.
Feist, G. J. (2019). Creativity and the big two model of personality: plasticity and stability. Curr. Opin. Behav. Sci. 27, 31–35. doi: 10.1016/j.cobeha.2018.07.005
Feist, G. J., and Brady, T. R. (2004). Openness to experience, non-conformity, and the preference for abstract art. Empir. Stud. Arts 22, 77–89. doi: 10.2190/Y7CA-TBY6-V7LR-76GK
Fitch, W. T. (2015). Four principles of bio-musicology. Philos. Trans. R. Soc. B: Biol. Sci. 370:20140091. doi: 10.1098/rstb.2014.0091
Frois, J. P., and Eysenck, H. J. (1995). The visual aesthetic sensitivity test applied to Portuguese children and fine arts students. Creat. Res. J. 8, 277–284. doi: 10.1207/s15326934crj0803_6
Fukui, H. (2001). Music and testosterone: a new hypothesis for the origin and function of music. Ann. N. Y. Acad. Sci. 930, 448–451. doi: 10.1111/j.1749-6632.2001.tb05767.x
Furnham, A. (2021). Demographic, personality trait and personality disorder correlates of aesthetic motivation. Imagin. Cogn. Pers. 40, 333–350. doi: 10.1177/0276236620942917
Gajos, J. M., and Beaver, K. M. (2017). The role of paternal age in the prediction of offspring intelligence. J. Genet. Psychol. 178, 319–333. doi: 10.1080/00221325.2017.1377678
Galasinska, K., and Szymkow, A. (2021). The more fertile, the more creative: changes in Women’s creative potential across the ovulatory cycle. Int. J. Environ. Res. Public Health 18:5390. doi: 10.3390/ijerph18105390
Galasinska, K., and Szymkow, A. (2022). Enhanced originality of ideas in women during ovulation: a within-subject study. Front. Psychol. 2643:859108. doi: 10.3389/fpsyg.2022.859108
Gale, C. R., Batty, G. D., Tynelius, P., Deary, I. J., and Rasmussen, F. (2010). Intelligence in early adulthood and subsequent hospitalisation and admission rates for the whole range of mental disorders: longitudinal study of 1,049,663 men. Epidemiology 21:70. doi: 10.1097/EDE.0b013e3181c17da8
Gangestad, S. W., and Simpson, J. A. (2000). The evolution of human mating: trade-offs and strategic pluralism. Behav. Brain Sci. 23, 573–587. doi: 10.1017/S0140525X0000337X
Gangestad, S. W., and Thornhill, R. (2003). Facial masculinity and fluctuating asymmetry. Evol. Hum. Behav. 24, 231–241. doi: 10.1016/S1090-5138(03)00017-5
Gao, Z., Gao, S., Xu, L., Zheng, X., Ma, X., Luo, L., et al. (2017). Women prefer men who use metaphorical language when paying compliments in a romantic context. Sci. Rep. 7:40871.
Garamszegi, L. Z., Zagalska-Neubauer, M., Canal, D., Blázi, G., Laczi, M., Nagy, G., et al. (2018). MHC-mediated sexual selection on birdsong: generic polymorphism, particular alleles and acoustic signals. Mol. Ecol. 27, 2620–2633. doi: 10.1111/mec.14703
Garcia, J. R., MacKillop, J., Aller, E. L., Merriwether, A. M., Wilson, D. S., and Lum, J. K. (2010). Associations between dopamine D4 receptor gene variation with both infidelity and sexual promiscuity. PLoS One 5:e14162. doi: 10.1371/journal.pone.0014162
Garofoli, D. (2015). Do early body ornaments prove cognitive modernity? A critical analysis from situated cognition. Phenomenol. Cogn. Sci. 14, 803–825. doi: 10.1007/s11097-014-9356-0
Geary, D. C. (2019). The spark of life and the unification of intelligence, health, and aging. Curr. Dir. Psychol. Sci. 28:0963721419829719. doi: 10.1177/0963721419829719
Gegear, R. J., Otterstatter, M. C., and Thomson, J. D. (2006). Bumble-bee foragers infected by a gut parasite have an impaired ability to utilize floral information. Proc. R. Soc. B Biol. Sci. 273, 1073–1078. doi: 10.1098/rspb.2005.3423
Gocłowska, M. A., Ritter, S. M., Elliot, A. J., and Baas, M. (2019). Novelty seeking is linked to openness and extraversion, and can lead to greater creative performance. J. Pers. 87, 252–266. doi: 10.1111/jopy.12387
Goldin, C., and Rouse, C. (2000). Orchestrating impartiality: the impact of" blind" auditions on female musicians. Am. Econ. Rev. 90, 715–741. doi: 10.1257/aer.90.4.715
Greengross, G., Silvia, P. J., and Nusbaum, E. C. (2020). Sex differences in humor production ability: a meta-analysis. J. Res. Pers. 84:103886. doi: 10.1016/j.jrp.2019.103886
Grigorenko, E. L., LaBuda, M. C., and Carter, A. S. (1992). Similarity in general cognitive ability, creativity, and cognitive style in a sample of adolescent Russian twins. Acta. Genet. Med. Gemellol. (Roma) 41, 65–72. doi: 10.1017/S000156600000252X
Griskevicius, V., Cialdini, R. B., and Kenrick, D. T. (2006). Peacocks, Picasso, and parental investment: the effects of romantic motives on creativity. J. Pers. Soc. Psychol. 91, 63–76. doi: 10.1037/0022-3514.91.1.63
Harpending, H., and Cochran, G. (2002). In our genes. Proc. Natl. Acad. Sci. 99, 10–12. doi: 10.1073/pnas.012612799
Harrison, M. A., and Hughes, S. M. (2017). Sex drugs and rock and roll: evidence supporting the storied trilogy. Hum. Ethol. Bull. 32, 63–84. doi: 10.22330/heb/323/063-084
Haselton, M. G., and Miller, G. F. (2006). Women’s fertility across the cycle increases the short-term attractiveness of creative intelligence. Hum. Nat. 17, 50–73. doi: 10.1007/s12110-006-1020-0
Hassler, M. (1992). Creative musical behavior and sex hormones: musical talent and spatial ability in the two sexes. Psychoneuroendocrinology 17, 55–70. doi: 10.1016/0306-4530(92)90076-J
He, W. (2018). A four-year longitudinal study of the sex-creativity relationship in childhood, adolescence, and emerging adulthood: findings of mean and variability analyses. Front. Psychol. 9:2331. doi: 10.3389/fpsyg.2018.02331
He, W. J., and Wong, W. C. (2011). Gender differences in creative thinking revisited: findings from analysis of variability. Personal. Individ. Differ. 51, 807–811. doi: 10.1016/j.paid.2011.06.027
He, M. W. J., Wong, W. C., and Hui, A. N. N. (2015). “Gender differences in means and variability on creative thinking: patterns in childhood, adolescence, and emerging adulthood,” in Creativity, Culture and Development. eds. A. G. Tan and C. Perleth (Singapore: Springer), 85–98.
He, W., Wong, W., Li, Y., and Xu, H. (2013). A study of the greater male variability hypothesis in creative thinking in mainland China: male superiority exists. Personal. Individ. Differ. 55, 882–886. doi: 10.1016/j.paid.2013.07.017
Hemdan, A. H., and Kazem, A. M. (2019). Creativity development of high-achieving students. Creat. Res. J. 31, 296–308. doi: 10.1080/10400419.2019.1641684
Henrich, J., and Gil-White, F. J. (2001). The evolution of prestige: freely conferred deference as a mechanism for enhancing the benefits of cultural transmission. Evol. Hum. Behav. 22, 165–196. doi: 10.1016/S1090-5138(00)00071-4
Holahan, C. K., Sears, R. R., and Cronbach, L. J. (1995). The Gifted Group in Later Maturity. Stanford, CA: Stanford University Press.
Holt, N. J. (2019). The expression of schizotypy in the daily lives of artists. Psychol. Aesthet. Creat. Arts 13:359. doi: 10.1037/aca0000176
Hooper, P. L., and Miller, G. F. (2008). Mutual mate choice can drive costly signaling even under perfect monogamy. Adapt. Behav. 16, 53–70. doi: 10.1177/1059712307087283
Hora, S., Badura, K. L., Lemoine, G. J., and Grijalva, E. (2021). A meta-analytic examination of the gender difference in creative performance. J. Appl. Psychol. Advance online publication. doi: 10.1037/apl0000999
Hornberg, J., and Reiter-Palmon, R. (2017). “Creativity and the big five personality traits: is the relationship dependent on the creativity measure?,” in The Cambridge Handbook of Creativity and Personality Research (Cambridge: Cambridge University Press), 275–293.
Iijima, M., Arisaka, O., Minamoto, F., and Arai, Y. (2001). Sex differences in children’s free drawings: a study on girls with congenital adrenal hyperplasia. Horm. Behav. 40, 99–104. doi: 10.1006/hbeh.2001.1670
Janicke, T., and Fromonteil, S. (2021). Sexual selection and sexual size dimorphism in animals. Biol. Lett. 17:20210251. doi: 10.1098/rsbl.2021.0251
Jankowska, D. M., Omelańczuk, I., Czerwonka, M., and Karwowski, M. (2019). Exploring links between creative abilities, creative personality and subclinical autistic traits. Personal. Individ. Differ. 142, 226–231. doi: 10.1016/j.paid.2018.05.008
Jeffery, A. J., Pham, M. N., Shackelford, T. K., and Fink, B. (2016). Does human ejaculate quality relate to phenotypic traits? Am. J. Hum. Biol. 28, 318–329. doi: 10.1002/ajhb.22805
Jonsdottir, G. A., Einarsson, G., Thorleifsson, G., Magnusson, S. H., Gunnarsson, A. F., Frigge, M. L., et al. (2021). Genetic propensities for verbal and spatial ability have opposite effects on body mass index and risk of schizophrenia. Intelligence 88:101565. doi: 10.1016/j.intell.2021.101565
Ju, C., Duan, Y., and You, X. (2015). Retesting the greater male variability hypothesis in mainland China: a cross-regional study. Personal. Individ. Differ. 72, 85–89. doi: 10.1016/j.paid.2014.07.021
Jung, R. E., Mead, B. S., Carrasco, J., and Flores, R. A. (2013). The structure of creative cognition in the human brain. Front. Hum. Neurosci. 7:330. doi: 10.3389/fnhum.2013.00330
Kamble, S., Shackelford, T. K., Pham, M., and Buss, D. M. (2014). Indian mate preferences: continuity, sex differences, and cultural change across a quarter of a century. Personal. Individ. Differ. 70, 150–155. doi: 10.1016/j.paid.2014.06.024
Kanazawa, S. (2000). Scientific discoveries as cultural displays: a further test of Miller’s courtship model. Evol. Hum. Behav. 21, 317–321. doi: 10.1016/S1090-5138(00)00051-9
Kandler, C., Riemann, R., Angleitner, A., Spinath, F. M., Borkenau, P., and Penke, L. (2016). The nature of creativity: the roles of genetic factors, personality traits, cognitive abilities, and environmental sources. J. Pers. Soc. Psychol. 111, 230–249. doi: 10.1037/pspp0000087
Kapoor, H., Reiter-Palmon, R., and Kaufman, J. C. (2021). Norming the muses: establishing the psychometric properties of the Kaufman domains of creativity scale. J. Psychoeduc. Assess. 39, 680–693. doi: 10.1177/07342829211008334
Karamihalev, S. (2013). Why creativity is sexy: a review of the evidence of sexual selection for creative abilities in humans. J. Eur. Psychol. Stud. 4, 78–86. doi: 10.5334/jeps.bb
Karwowski, M., Czerwonka, M., Wiśniewska, E., and Forthmann, B. (2021). How is intelligence test performance associated with creative achievement? A meta-analysis. J. Intell. 9:28. doi: 10.3390/jintelligence9020028
Karwowski, M., Jankowska, D. M., Gajda, A., Marczak, M., Groyecka, A., and Sorokowski, P. (2016a). Greater male variability in creativity outside the WEIRD world. Creat. Res. J. 28, 467–470. doi: 10.1080/10400419.2016.1229978
Karwowski, M., Jankowska, D. M., Gralewski, J., Gajda, A., Wiśniewska, E., and Lebuda, I. (2016b). Greater male variability in creativity: a latent variables approach. Think. Skills Creat. 22, 159–166. doi: 10.1016/j.tsc.2016.10.005
Karwowski, M., and Lebuda, I. (2016). The big five, the huge two, and creative self-beliefs: a meta-analysis. Psychol. Aesthet. Creat. Arts 10:214. doi: 10.1037/aca0000035
Kaufman, A. B., Butt, A. E., Kaufman, J. C., and Colbert-White, E. N. (2011). Towards a neurobiology of creativity in nonhuman animals. J. Comp. Psychol. 125:255. doi: 10.1037/a0023147
Kaufman, S. B., Kozbelt, A., Bromley, M. L., and Miller, G. (2007). “The role of creativity and humor in mate selection,” in Mating Intelligence eds. G. Geher and G. Miller (Hillsdale, NJ: Lawrence Erlbaum Associates Publishers), 253–288.
Kaufman, S. B., Kozbelt, A., Silvia, P., Kaufman, J. C., Ramesh, S., and Feist, G. J. (2016). Who finds bill gates sexy? Creative mate preferences as a function of cognitive ability, personality, and creative achievement. J. Creat. Behav. 50, 294–307. doi: 10.1002/jocb.78
Kawase, H., Okata, Y., and Ito, K. (2013). Role of huge geometric circular structures in the reproduction of a marine pufferfish. Sci. Rep. 3, 1–5. doi: 10.1038/srep02106
Keagy, J., Savard, J.-F., and Borgia, G. (2009). Male satin bowerbird problem-solving ability predicts mating success. Anim. Behav. 78, 809–817. doi: 10.1016/j.anbehav.2009.07.011
Keagy, J., Savard, J. F., and Borgia, G. (2011). Complex relationship between multiple measures of cognitive ability and male mating success in satin bowerbirds, ptilonorhynchus violaceus. Anim. Behav. 81, 1063–1070. doi: 10.1016/j.anbehav.2011.02.018
Keagy, J., Savard, J. F., and Borgia, G. (2012). Cognitive ability and the evolution of multiple behavioral display traits. Behav. Ecol. 23, 448–456. doi: 10.1093/beheco/arr211
Kell, H. J., Lubinski, D., Benbow, C. P., and Steiger, J. H. (2013). Creativity and technical innovation: spatial ability’s unique role. Psychol. Sci. 24, 1831–1836. doi: 10.1177/0956797613478615
Kellman, J. (1998). Ice age art, autism, and vision: how we see/how we draw. Stud. Art Educ. 39, 117–131. doi: 10.2307/1320464
Kennair, L. E. O., Wade, T. J., Tallaksen, M. T., Grøntvedt, T. V., Kessler, A. M., Burch, R. L., et al. (2022). Perceived effectiveness of flirtation tactics: the effects of sex, mating context and individual differences in US and Norwegian samples. Evol. Psychol. 20:14747049221088011. doi: 10.1177/14747049221088011
Kenrick, D. T., Griskevicius, V., Neuberg, S. L., and Schaller, M. (2010). Renovating the pyramid of needs: contemporary extensions built upon ancient foundations. Perspect. Psychol. Sci. 5, 292–314. doi: 10.1177/1745691610369469
Kim, K. H. (2008). Meta-analyses of the relationship of creative achievement to both IQ and divergent thinking test scores. J. Creat. Behav. 42, 106–130. doi: 10.1002/j.2162-6057.2008.tb01290.x
Klasios, J. (2013). Cognitive traits as sexually selected fitness indicators. Rev. Gen. Psychol. 17, 428–442. doi: 10.1037/a0034391
Kohn, M., and Mithen, S. (1999). Handaxes: products of sexual selection? Antiquity 73, 518–526. doi: 10.1017/S0003598X00065078
Konner, M. (2021). Nine levels of explanation. Hum. Nat. 32, 748–793. doi: 10.1007/s12110-021-09414-8
Kovacs, K., and Conway, A. R. A. (2019). What Is IQ? Life Beyond “General Intelligence.”. Curr. Dir. Psychol. Sci. 28, 1–6. doi: 10.1177/0963721419827275
Kyaga, S., Lichtenstein, P., Boman, M., Hultman, C., Långström, N., and Landen, M. (2011). Creativity and mental disorder: family study of 300 000 people with severe mental disorder. Br. J. Psychiatry 199, 373–379. doi: 10.1192/bjp.bp.110.085316
Lange, B. P., and Euler, H. A. (2014). Writers have groupies, too: high quality literature production and mating success. Evol. Behav. Sci. 8, 20–30. doi: 10.1037/h0097246
Lange, B. P., Hennighausen, C., Brill, M., and Schwab, F. (2016). Only cheap talk after all? New experimental psychological findings on the role of verbal proficiency in mate choice. Psychol. Lang. Commun. 20, 1–22. doi: 10.1515/plc-2016-0001
Lange, B. P., Zaretsky, E., Schwarz, S., and Euler, H. A. (2014). Words won’t fail: experimental evidence on the role of verbal proficiency in mate choice. J. Lang. Soc. Psychol. 33, 482–499. doi: 10.1177/0261927X13515886
Lau, S., and Cheung, P. C. (2015). A gender-fair look at variability in creativity: growth in variability over a period versus gender comparison at a time point. Creat. Res. J. 27, 87–95. doi: 10.1080/10400419.2015.992685
Lebuda, I., Sorokowski, P., Groyecka-Bernard, A., Marczak, M., Gajda, A., Jankowska, D. M., et al. (2021). Creativity, mating, and reproductive successes outside the WEIRD world. Creat. Res. J. 33, 255–263. doi: 10.1080/10400419.2020.1870816
Lefebvre, L. (2013). Brains, innovations, tools and cultural transmission in birds, non-human primates, and fossil hominins. Front. Hum. Neurosci. 7:245. doi: 10.3389/fnhum.2013.00245
Lefebvre, L., Reader, S. M., and Sol, D. (2004). Brains, innovations and evolution in birds and primates. Brain Behav. Evol. 63, 233–246. doi: 10.1159/000076784
Lewis, D. M., Al-Shawaf, L., Conroy-Beam, D., Asao, K., and Buss, D. M. (2017). Evolutionary psychology: a how-to guide. Am. Psychol. 72, 353–373. doi: 10.1037/a0040409
Li, N. P., Bailey, J. M., Kenrick, D. T., and Linsenmeier, J. A. W. (2002). The necessities and luxuries of mate preferences: testing the tradeoffs. J. Pers. Soc. Psychol. 82, 947–955. doi: 10.1037/0022-3514.82.6.947
Li, N. P., Valentine, K. A., and Patel, L. (2011). Mate preferences in the US and Singapore: a cross-cultural test of the mate preference priority model. Personal. Individ. Differ. 50, 291–294. doi: 10.1016/j.paid.2010.10.005
Li, H., Zhang, C., Cai, X., Wang, L., Luo, F., Ma, Y., et al. (2020). Genome-wide association study of creativity reveals genetic overlap with psychiatric disorders, risk tolerance, and risky behaviors. Schizophr. Bull. 46, 1317–1326. doi: 10.1093/schbul/sbaa025
Lindell, A. K. (2014). On the interrelation between reduced lateralization, schizotypy, and creativity. Front. Psychol. 5:813. doi: 10.3389/fpsyg.2014.00813
Low, B. S. (1979). “Sexual selection and human ornamentation,” in Evolutionary Biology and Human Social Behavior. eds. N. A. Chagnon and W. Irons (Boston: Duxbury Press), 462–487. Available at: https://hraf.yale.edu/ehc/documents/355
Luoto, S. (2019a). An updated theoretical framework for human sexual selection: from ecology, genetics, and life history to extended phenotypes. Adapt. Hum. Behav. Physiol. 5, 1–55. doi: 10.1007/s40750-018-0103-6
Luoto, S. (2019b). Response to commentaries: life history genetics, fluid intelligence, and extended phenotypes. Adapt. Hum. Behav. Physiol. 5, 112–115. doi: 10.1007/s40750-019-0109-8
Luoto, S., and Varella, M. A. C. (2021). Pandemic leadership: sex differences and their evolutionary–developmental origins. Front. Psychol. 12:633862. doi: 10.3389/fpsyg.2021.633862
Macdougall-Shackleton, S. A. (1997). “Sexual selection and the evolution of song repertoires,” in Current Ornithology. Current Ornithology. eds. V. Nolan, E. D. Ketterson, and C. F. Thompson, vol. 14 (Boston, MA: Springer).
Mackie, M. E. (2015). Estimating age and sex: Paleodemographic identification using rock art hand sprays, an application in Johnson County, Wyoming. J. Archaeol. Sci. Rep. 3, 333–341. doi: 10.1016/j.jasrep.2015.06.023
Madison, G., Holmquist, J., and Vestin, M. (2018). Musical improvisation skill in a prospective partner is associated with mate value and preferences, consistent with sexual selection and parental investment theory: implications for the origin of music. Evol. Hum. Behav. 39, 120–129. doi: 10.1016/j.evolhumbehav.2017.10.005
Manning, J. T., and Taylor, R. P. (2001). Second to fourth digit ratio and male ability in sport: implications for sexual selection in humans. Evol. Hum. Behav. 22, 61–69. doi: 10.1016/S1090-5138(00)00063-5
Marin, M. M., and Rathgeber, I. (2022). Darwin’s sexual selection hypothesis revisited: musicality increases sexual attraction in both sexes. Front. Psychol. doi: 10.3389/fpsyg.2022.971988
Marin, M. M., Schober, R., Gingras, B., and Leder, H. (2017). Misattribution of musical arousal increases sexual attraction towards opposite-sex faces in females. PLoS One 12:e0183531. doi: 10.1371/journal.pone.0183531
May, J. L., and Hamilton, P. A. (1980). Effects of musically evoked affect on women’s interpersonal attraction toward and perceptual judgments of physical attractiveness of men. Motiv. Emot. 4, 217–228. doi: 10.1007/BF00995420
Mayseless, N., Uzefovsky, F., Shalev, I., Ebstein, R. P., and Shamay-Tsoory, S. G. (2013). The association between creativity and 7R polymorphism in the dopamine receptor D4 gene (DRD4). Front. Hum. Neurosci. 7:502. doi: 10.3389/fnhum.2013.00502
Mehr, S. A., Krasnow, M. M., Bryant, G. A., and Hagen, E. H. (2021). Origins of music in credible signaling. Behav. Brain Sci. 44:E60. doi: 10.1017/S0140525X20000345
Menninghaus, W. (2019). Aaesthetics After Darwin: The Multiple Origins and Functions of Art. Boston: Academic Studies Press.
Miller, G. F. (2000). Mental traits as fitness indicators: expanding evolutionary psychology’s adaptationism. Ann. N. Y. Acad. Sci. 907, 62–74. doi: 10.1111/j.1749-6632.2000.tb06616.x
Miller, G. F. (2001). Aesthetic fitness: how sexual selection shaped artistic virtuosity as a fitness indicator and aesthetic preferences as mate choice criteria. Bull. Psychol. Arts 2, 20–25.
Miller, G. F. (2013). Mutual mate choice models as the red pill in evolutionary psychology: Long delayed, much needed, ideologically challenging, and hard to swallow. Psychol. Inq. 24, 207–210. doi: 10.1080/1047840X.2013.817937
Miller, G. F., and Todd, P. M. (1998). Mate choice turns cognitive. Trends Cogn. Sci. 2, 190–198. doi: 10.1016/S1364-6613(98)01169-3
Miller, G., Tybur, J. M., and Jordan, B. D. (2007). Ovulatory cycle effects on tip earnings by lap dancers: economic evidence for human estrus? Evol. Hum. Behav. 28, 375–381. doi: 10.1016/j.evolhumbehav.2007.06.002
Minter, R., Keagy, J., and Tinghitella, R. M. (2017). The relationship between male sexual signals, cognitive performance, and mating success in stickleback fish. Ecol. Evol. 7, 5621–5631. doi: 10.1002/ece3.3091
Mithen, S. (2003). “Handaxes: the first aesthetic artefacts,” in Evolutionary Aesthetics. eds. E. Voland and K. Grammer (Berlin, Heidelberg: Springer), 261–275.
Møller, A. P., and Petrie, M. (2002). Condition dependence, multiple sexual signals, and immunocompetence in peacocks. Behav. Ecol. 13, 248–253. doi: 10.1093/beheco/13.2.248
Moraes, Y. L., Valentova, J. V., and Varella, M. A. C. (2022). The evolution of playfulness, play and play-like phenomena in relation to sexual selection. Front. Psychol. doi: 10.3389/fpsyg.2022.925842
Morris, D. (1962). The Biology of Art: A Study of the Picture-Making Behaviour of the Great Apes and its Relationship to Human Art. London: Methuen
Morsanyi, K., Primi, C., Handley, S. J., Chiesi, F., and Galli, S. (2012). Are systemizing and autistic traits related to talent and interest in mathematics and engineering? Testing some of the central claims of the empathizing–systemizing theory. Br. J. Psychol. 103, 472–496. doi: 10.1111/j.2044-8295.2011.02089.x
Mosing, M. A., Madison, G., Pedersen, N. L., Kuja-Halkola, R., and Ullén, F. (2014). Practice does not make perfect: no causal effect of music practice on music ability. Psychol. Sci. 25, 1795–1803. doi: 10.1177/0956797614541990
Mosing, M. A., Verweij, K. J. H., Madison, G., Pedersen, N. L., Zietsch, B. P., and Ullén, F. (2015). Did sexual selection shape human music? Testing predictions from the sexual selection hypothesis of music evolution using a large genetically informative sample of over 10,000 twins. Evol. Hum. Behav. 36, 359–366. doi: 10.1016/j.evolhumbehav.2015.02.004
Myszkowski, N., Çelik, P., and Storme, M. (2018). A meta-analysis of the relationship between intelligence and visual “taste” measures. Psychol. Aesthet. Creat. Arts 12, 24–33. doi: 10.1037/aca0000099
Myszkowski, N., Storme, M., Zenasni, F., and Lubart, T. (2014). Is visual aesthetic sensitivity independent from intelligence, personality and creativity? Personal. Individ. Differ. 59, 16–20. doi: 10.1016/j.paid.2013.10.021
Nakano, T. D. C., Oliveira, K. D. S., and Zaia, P. (2021). Gender differences in creativity: a systematic literature review. Psicologia: Teoria e Pesquisa 37, 1–10. doi: 10.1590/0102.3772e372116
Natividade, J. C., and Hutz, C. S. (2015). Short form scale of descriptors of the five personality factors: pros and cons. Psico 46, 79–89. doi: 10.15448/1980-8623.2015.1.16901
Natividade, J. C., and Hutz, C. S. (2016). Personal characteristics associated with sexuality can be classified into seven dimensions in Brazil. Personal. Individ. Differ. 97, 88–97. doi: 10.1016/j.paid.2016.03.030
Nettle, D., and Clegg, H. (2006). Schizotypy, creativity and mating success in humans. Proc. R. Soc. B Biol. Sci. 273, 611–615. doi: 10.1098/rspb.2005.3349
Novaes, F. (2022). Sapiossexualidade: A influência da inteligência e da criatividade na criatividade e na seleção de parceiros [Sapiosexuality: The Influence of Intelligence and Creativity on Attractiveness and Sexual Selection]. PhD thesis. Rio de Janeiro (RJ): Pontifical Catholic University of Rio de Janeiro.
Ord, M. (2020). ““Every noise at once”: online music discovery maps and cosmopolitan subjectivities” in The Bloomsbury Handbook of Popular Music and Social Class. ed. I. Peddie (New York: Bloomsbury Publishing Inc), 117–133.
Östlund-Nilsson, S., and Holmlund, M. (2003). The artistic three-spined stickleback (Gasterosteous aculeatus). Behav. Ecol. Sociobiol. 53, 214–220. doi: 10.1007/s00265-002-0574-z
Overmann, K. A. (2016). Beyond writing: the development of literacy in the ancient near east. Camb. Archaeol. J. 26, 285–303. doi: 10.1017/S0959774316000019
Park, G., Lubinski, D., and Benbow, C. P. (2007). Contrasting intellectual patterns predict creativity in the arts and sciences: tracking intellectually precocious youth over 25 years. Psychol. Sci. 18, 948–952. doi: 10.1111/j.1467-9280.2007.02007.x
Parkinson, C., and Wheatley, T. (2015). The repurposed social brain. Trends Cogn. Sci. 19, 133–141. doi: 10.1016/j.tics.2015.01.003
Pennisi, P., Laura, G., Giusy, M., and Michela, C. (2020). Autism, autistic traits and creativity: a systematic review and meta-analysis. Cogn. Process. 22, 1–36. doi: 10.1007/s10339-020-00992-6
Petrie, M. (2021). Evolution by sexual selection. Front. Ecol. Evol. 950:786868. doi: 10.3389/fevo.2021.786868
Piffer, D., and Hur, Y.-M. (2014). Heritability of creative achievement. Creat. Res. J. 26, 151–157. doi: 10.1080/10400419.2014.901068
Podlipniak, P. (2017). The role of the Baldwin effect in the evolution of human musicality. Front. Neurosci. 11:542. doi: 10.3389/fnins.2017.00542
Podlipniak, P. (2021). The role of canalization and plasticity in the evolution of musical creativity. Front. Neurosci. 15:267. doi: 10.3389/fnins.2021.607887
Ponzi, D., Henry, A., Kubicki, K., Nickels, N., Wilson, M. C., and Maestripieri, D. (2016). Autistic-like traits, sociosexuality, and hormonal responses to socially stressful and sexually arousing stimuli in male college students. Adapt. Hum. Behav. Physiol. 2, 150–165. doi: 10.1007/s40750-015-0034-4
Portmann, A. (1969). Biologische Fragmente zu einer Lehre vom Menschen [A Zoologist Looks at Humankind] Schwabe, Basel, Germany trans Schaefer J (1990).
Power, R. A., Steinberg, S., Bjornsdottir, G., Rietveld, C. A., Abdellaoui, A., Nivard, M. M., et al. (2015). Polygenic risk scores for schizophrenia and bipolar disorder predict creativity. Nat. Neurosci. 18, 953–955. doi: 10.1038/nn.4040
Prokop, Z. M., Michalczyk, Ł., Drobniak, S. M., Herdegen, M., and Radwan, J. (2012). Meta-analysis suggests choosy females get sexy sons more than “good genes”. Evol. Int. J. Org. Evol. 66, 2665–2673. doi: 10.1111/j.1558-5646.2012.01654.x
Prokosch, M. D., Coss, R. G., Scheib, J. E., and Blozis, S. A. (2009). Intelligence and mate choice: intelligent men are always appealing. Evol. Hum. Behav. 30, 11–20. doi: 10.1016/j.evolhumbehav.2008.07.004
Prum, R. O. (2012). Aesthetic evolution by mate choice: Darwin’s really dangerous idea. Philos. Trans. R. Soc. London B: Biol. Sci. 367, 2253–2265. doi: 10.1098/rstb.2011.0285
Puccio, G. J. (2017). From the dawn of humanity to the 21st century: creativity as an enduring survival skill. J. Creat. Behav. 51, 330–334. doi: 10.1002/jocb.203
Puryear, J. S., Kettler, T., and Rinn, A. N. (2017). Relationships of personality to differential conceptions of creativity: a systematic review. Psychol. Aesthet. Creat. Arts 11:59. doi: 10.1037/aca0000079
Puts, D. A. (2010). Beauty and the beast: mechanisms of sexual selection in humans. Evol. Hum. Behav. 31, 157–175. doi: 10.1016/j.evolhumbehav.2010.02.005
Puts, D. (2016). Human sexual selection. Curr. Opin. Psychol. 7, 28–32. doi: 10.1016/j.copsyc.2015.07.011
Rabazo-Rodríguez, A. M., Modesto-Mata, M., Bermejo, L., and García-Díez, M. (2017). New data on the sexual dimorphism of the hand stencils in El Castillo cave (Cantabria, Spain). J. Archaeol. Sci. Rep. 14, 374–381. doi: 10.1016/j.jasrep.2017.06.022
Rantala, M. J., Luoto, S., Borráz-León, J. I., and Krams, I. (2021). Bipolar disorder: an evolutionary psychoneuroimmunological approach. Neurosci. Biobehav. Rev. 122, 28–37. doi: 10.1016/j.neubiorev.2020.12.031
Rantala, M. J., Luoto, S., Borráz-León, J. I., and Krams, I. (2022). Schizophrenia: the new etiological synthesis. Neurosci. Biobehav. Rev. 1-2, 104894. doi: 10.1016/j.neubiorev.2022.104894
Rantala, M. J., Luoto, S., Krama, T., and Krams, I. (2019). Eating disorders: an evolutionary psychoneuroimmunological approach. Front. Psychol. 10:2200. doi: 10.3389/fpsyg.2019.02200
Reader, S. M., and Laland, K. N. (2001). Primate innovation: sex, age and social rank differences. Int. J. Primatol. 22, 787–805. doi: 10.1023/A:1012069500899
Reader, S. M., Morand-Ferron, J., and Flynn, E. (2016). Animal and human innovation: novel problems and novel solutions. Trans. R. Soc. B Biol. Sci. 371:20150182. doi: 10.1098/rstb.2015.0182
Reuter, M., Roth, S., Holve, K., and Hennig, J. (2006). Identification of first candidate genes for creativity: a pilot study. Brain Res. 1069, 190–197. doi: 10.1016/j.brainres.2005.11.046
Roeling, M. P., Willemsen, G., and Boomsma, D. I. (2017). Heritability of working in a creative profession. Behav. Genet. 47, 298–304. doi: 10.1007/s10519-016-9832-0
Rosenthal, G. G., and Ryan, M. J. (2022). Sexual selection and the ascent of women: mate choice research since Darwin. Science 375:eabi6308. doi: 10.1126/science.abi6308
Runco, M. A., and Jaeger, G. J. (2012). The standard definition of creativity. Creat. Res. J. 24, 92–96. doi: 10.1080/10400419.2012.650092
Runco, M. A., Noble, E. P., Reiter-Palmon, R., Acar, S., Ritchie, T., and Yurkovich, J. M. (2011). The genetic basis of creativity and ideational fluency. Creat. Res. J. 23, 376–380. doi: 10.1080/10400419.2011.621859
Ryan, R. M., and Deci, E. L. (2017). Self-determination Theory: Basic Psychological Needs in Motivation Development and Wellness. New York, NY: Guilford Press.
Said-Metwaly, S., Van den Noortgate, W., and Kyndt, E. (2017). Approaches to measuring creativity: a systematic literature review. Creativity. Theories–Res.–Appl. 4, 238–275. doi: 10.1515/ctra-2017-0013
Sanz, C., Call, J., and Morgan, D. (2009). Design complexity in termite-fishing tools of chimpanzees (pan troglodytes). Biol. Lett. 5, 293–296. doi: 10.1098/rsbl.2008.0786
Savage, P. E., Brown, S., Sakai, E., and Currie, T. E. (2015). Statistical universals reveal the structures and functions of human music. Proc. Natl. Acad. Sci. U. S. A. 112, 8987–8992. doi: 10.1073/pnas.1414495112
Savage, P. E., Loui, P., Tarr, B., Schachner, A., Glowacki, L., Mithen, S., et al. (2021). Music as a coevolved system for social bonding. Behav. Brain Sci. 44:E59. doi: 10.1017/S0140525X20000333
Savi, A. O., van der Maas, H. L. J., Maris, G. K. J., and Marsman, M. (2020). Mitochondrial functioning≠ general intelligence. J. Intelligence 8:20. doi: 10.3390/jintelligence8020020
Schmitt, D. P. (2005). Sociosexuality from Argentina to Zimbabwe: a 48-nation study of sex, culture, and strategies of human mating. Behav. Brain Sci. 28, 247–275. doi: 10.1017/S0140525X05000051
Schmitt, D. P., Long, A. E., McPhearson, A., O’Brien, K., Remmert, B., and Shah, S. H. (2017). Personality and gender differences in global perspective. Int. J. Psychol. 52, 45–56. doi: 10.1002/ijop.12265
Schmitt, D. P., and Pilcher, J. J. (2004). Evaluating evidence of psychological adaptation: how do we know one when we see one? Psychol. Sci. 15, 643–649. doi: 10.1111/j.0956-7976.2004.00734.x
Shaner, A., Miller, G., and Mintz, J. (2004). Schizophrenia as one extreme of a sexually selected fitness indicator. Schizophr. Res. 70, 101–109. doi: 10.1016/j.schres.2003.09.014
Shaw, R. C., MacKinlay, R. D., Clayton, N. S., and Burns, K. C. (2019). Memory performance influences male reproductive success in a wild bird. Curr. Biol. 29, 1498–1502. doi: 10.1016/j.cub.2019.03.027
Shizgal, P. (2001). “Motivation,” in The MIT Encyclopedia of the Cognitive Sciences. eds. R. Wilson and F. Keil (Cambridge, MA: MIT Press), 566–568.
Shuker, D. M., and Kvarnemo, C. (2021). The definition of sexual selection. Behav. Ecol. 32, 781–794. doi: 10.1093/beheco/arab055
Silvia, P. J. (2008). Creativity and intelligence revisited: a latent variable analysis of Wallach and Kogan. Creat. Res. J. 20, 34–39. doi: 10.1080/10400410701841807
Silvia, P. J., and Beaty, R. E. (2012). Making creative metaphors: the importance of fluid intelligence for creative thought. Intelligence 40, 343–351. doi: 10.1016/j.intell.2012.02.005
Silvia, P. J., and Nusbaum, E. C. (2013). Verbal fluency and creativity: general and specific contributions of broad retrieval ability (gr) factor to divergente thinking. Intelligence 41, 328–340. doi: 10.1016/j.intell.2013.05.004
Sluming, V. A., and Manning, J. T. (2000). Second to fourth digit ratio in elite musicians: evidence for musical ability as an honest signal of male fitness. Evol. Hum. Behav. 21, 1–9. doi: 10.1016/S1090-5138(99)00026-4
Smith, D., Schlaepfer, P., Major, K., Dyble, M., Page, A. E., Thompson, J., et al. (2017). Cooperation and the evolution of hunter-gatherer storytelling. Nat. Commun. 8, 1–9. doi: 10.1038/s41467-017-02036-8
Snow, D. R. (2013). Sexual dimorphism in European upper Paleolithic cave art. Am. Antiq. 78, 746–761. doi: 10.7183/0002-7316.78.4.746
Somers, M., Sommer, I. E., Boks, M. P., and Kahn, R. S. (2009). Hand-preference and population schizotypy: a meta-analysis. Schizophr. Res. 108, 25–32. doi: 10.1016/j.schres.2008.11.010
Souza, A. L., Conroy-Beam, D., and Buss, D. M. (2016). Mate preferences in Brazil: evolved desires and cultural evolution over three decades. Personal. Individ. Differ. 95, 45–49. doi: 10.1016/j.paid.2016.01.053
Spencer, K. A., Buchanan, K. L., Leitner, S., Goldsmith, A. R., and Catchpole, C. K. (2005). Parasites affect song complexity and neural development in a songbird. Proc. R. Soc. B Biol. Sci. 272, 2037–2043. doi: 10.1098/rspb.2005.3188
Spikins, P., Scott, C., and Wright, B. (2018). How do we explain ‘autistic traits’ in European upper Palaeolithic art? Open Archaeol. 4, 262–279. doi: 10.1515/opar-2018-0016
Spritzer, M. D., Solomon, N. G., and Meikle, D. B. (2005). Influence of scramble competition for mates upon the spatial ability of male meadow voles. Anim. Behav. 69, 375–386. doi: 10.1016/j.anbehav.2004.03.015
Stephen, I., and Luoto, S. (2021). “Physical cues of partner quality,” in The Oxford Handbook of Evolutionary Psychology and Romantic Relationships. eds. J. Mogilski and T. Shackelford (Oxford: Oxford University Press).
Stern, J., Kordsmeyer, T. L., and Penke, L. (2021). A longitudinal evaluation of ovulatory cycle shifts in women’s mate attraction and preferences. Horm. Behav. 128:104916. doi: 10.1016/j.yhbeh.2020.104916
Stewart-Williams, S., and Thomas, A. G. (2013). The ape that thought it was a peacock: does evolutionary psychology exaggerate human sex differences? Psychol. Inq. 24, 137–168. doi: 10.1080/1047840X.2013.804899
Takeuchi, H., Kimura, R., Tomita, H., Taki, Y., Kikuchi, Y., Ono, C., et al. (2021). Polygenic risk score for bipolar disorder associates with divergent thinking and brain structures in the prefrontal cortex. Hum. Brain Mapp. 42, 6028–6037. doi: 10.1002/hbm.25667
Tan, Y. T., McPherson, G. E., Peretz, I., Berkovic, S. F., and Wilson, S. J. (2014). The genetic basis of music ability. Front. Psychol. 5:658. doi: 10.3389/fpsyg.2014.00658
Taylor, A. H. (2014). Corvid cognition. Wiley Interdiscip. Rev. Cogn. Sci. 5, 361–372. doi: 10.1002/wcs.1286
Taylor, C. L. (2017). Creativity and mood disorder: a systematic review and meta-analysis. Perspect. Psychol. Sci. 12, 1040–1076. doi: 10.1177/1745691617699653
Taylor, C. L., and Barbot, B. (2021). Gender differences in creativity: examining the greater male variability hypothesis in different domains and tasks. Personal. Individ. Differ. 174:110661. doi: 10.1016/j.paid.2021.110661
Thavarajah, N. K., Tickle, P. G., Nudds, R. L., and Codd, J. R. (2016). The peacock train does not handicap cursorial locomotor performance. Sci. Rep. 6, 1–6. doi: 10.1038/srep36512
Thomas, A. G., Armstrong, S. L., Stewart-Williams, S., and Jones, B. C. (2021). Current fertility status does not predict sociosexual attitudes and desires in normally ovulating women. Evol. Psychol. 19, 1–7. doi: 10.1177/1474704920976318
Tinbergen, N. (1963). On aims and methods of ethology. Z. Tierpsychol. 20, 410–433. doi: 10.1111/j.1439-0310.1963.tb01161.x
Trevarthen, C. (1999). Musicality and the intrinsic motive pulse: evidence from human psychobiology and infant communication. Music. Sci. 3, 155–215. doi: 10.1177/10298649000030S109
Trivers, R. L. (1972). “Parental investment and sexual selection,” in Sexual Selection and the Descent of Man (Chicago, IL: Aldine), 136–179.
Ujma, P. P., and Kovacs, K. (2020). The mitochondrial theory of g is incompatible with genetic evidence and does not explain statistical phenomena. J. Intelligence 8:27. doi: 10.3390/jintelligence8030027
Valentova, J. V., Mafra, A. L., and Varella, M. A. C. (2022). Enhancing the evolutionary science of self-presentation modification. Arch. Sex. Behav. 51, 79–84. doi: 10.1007/s10508-021-01975-0
van der Linden, D., Dutton, E., and Madison, G. (2020). National-level indicators of androgens are related to the global distribution of scientific productivity and science Nobel prizes. J. Creat. Behav. 54, 134–149. doi: 10.1002/jocb.351
van Schaik, C. P., Burkart, J., Damerius, L., Forss, S. I. F., Koops, K., Van Noordwijk, M. A., et al. (2016). The reluctant innovator: orangutans and the phylogeny of creativity. Philos. Trans. R. Soc. B: Biol. Sci. 371:20150183. doi: 10.1098/rstb.2015.0183
Varella, M. A. C. (2021). Evolved features of artistic motivation: analysing a Brazilian database spanning three decades. Front. Psychol. doi: 10.3389/fpsyg.2021.769915
Varella, M. A. C., de Souza, A. A. L., and Ferreira, J. H. B. P. (2011). Evolutionary aesthetics and sexual selection in the evolution of rock art aesthetics [with comments]. Rock Art Res. J. Aust. Rock Art Res. Assoc. (AURA) 28, 153–186.
Varella, M. A. C., de Souza, A. A. L., and Ferreira, J. H. B. P. (2012). Considering both proximal and distal explanations for (rock) art production and appreciation as fruitful. Rock Art Res. 29, 227–229.
Varella, M. A. C., Ferreira, J. H. B. P., Cosentino, L. A. M., Ottoni, E., and Bussab, V. S. R. (2010). Sex differences in aspects of musicality in a Brazilian sample: adaptative hypotheses. Cognition Musical Arts 4, 10–16.
Varella, M. A. C., Ferreira, J. H. B. P., and de Souza, A. A. L. (2014). The role of male and female in reproduction, and understanding of sexual selection when applied to human artistic propensities. Rock Art Res. 31, 239–243.
Varella, M. A. C., Santos, I. B. C. D., Ferreira, J. H. B. P., and Bussab, V. S. R. (2013). Misunderstandings in applying evolution to human mind and behavior and its causes: a systematic review. EvoS J: J. Evol. Stud. Consortium 5, 81–107.
Varella, M. A. C., Štěrbová, Z., Bártová, K., Fisher, M. L., and Valentova, J. V. (2022). Evolution of artistic and athletic propensities: testing of intersexual selection and intrasexual competition. Front. Psychol. 3892:925862. doi: 10.3389/fpsyg.2022.925862
Varella, M. A. C., Valentova, J. V., and Fernández, A. M. (2017). “Evolution of artistic and aesthetic propensities through female competitive ornamentation,” in The Oxford Handbook of Female Competition. ed. M. L. Fisher (New York: Oxford University Press), 757–783.
Vartanian, O., Wertz, C. J., Flores, R. A., Beatty, E. L., Smith, I., Blackler, K., et al. (2018). Structural correlates of openness and intellect: implications for the contribution of personality to creativity. Hum. Brain Mapp. 39, 2987–2996. doi: 10.1002/hbm.24054
Velázquez, J. A., Segal, N. L., and Horwitz, B. N. (2015). Genetic and environmental influences on applied creativity: a reared-apart twin study. Personal. Individ. Differ. 75, 141–146. doi: 10.1016/j.paid.2014.11.014
Verpooten, J., and Nelissen, M. (2012). “Sensory exploitation: underestimated in the evolution of art as once in sexual selection theory?” in Philosophy of Behavioral Biology (Boston Stu). eds. K. Plaisance and T. Reydon (Dordrecht: Springer).
Verweij, K. J. H., Burri, A. V., and Zietsch, B. P. (2014). Testing the prediction from sexual selection of a positive genetic correlation between human mate preferences and corresponding traits. Evol. Hum. Behav. 35, 497–501. doi: 10.1016/j.evolhumbehav.2014.06.009
Vinkhuyzen, A. A., Van der Sluis, S., Posthuma, D., and Boomsma, D. I. (2009). The heritability of aptitude and exceptional talent across different domains in adolescents and young adults. Behav. Genet. 39, 380–392. doi: 10.1007/s10519-009-9260-5
Walter, K. V., Conroy-Beam, D., Buss, D. M., Asao, K., Sorokowska, A., Sorokowski, P., et al. (2020). Sex differences in mate preferences across 45 countries: a large-scale replication. Psychol. Sci. 31, 408–423. doi: 10.1177/0956797620904154
Walter, K. V., Conroy-Beam, D., Buss, D. M., Asao, K., Sorokowska, A., Sorokowski, P., et al. (2021). Sex differences in human mate preferences vary across sex ratios. Proc. R. Soc. B 288:20211115. doi: 10.1098/rspb.2021.1115
Wang, L., Long, H., Plucker, J. A., Wang, Q., Xu, X., and Pang, W. (2018). High schizotypal individuals are more creative? The mediation roles of overinclusive thinking and cognitive inhibition. Front. Psychol. 9:1766. doi: 10.3389/fpsyg.2018.01766
Watanabe, S. (2013). “Animal aesthetics from the perspective of comparative cognition,” in The Science of the Mind. Emotions of Animals and Humans: Comparative Perspectives. eds. S. Watanabe and S. Kuczaj (New York, NY: Springer Science + Business Media), 129–162.
Watkins, C. D. (2017). Creating beauty: creativity compensates for low physical attractiveness when individuals assess the attractiveness of social and romantic partners. R. Soc. Open Sci. 4:160955. doi: 10.1098/rsos.160955
Weisberg, Y. J., DeYoung, C. G., and Hirsh, J. B. (2011). Gender differences in personality across the ten aspects of the big five. Front. Psychol. 2:178. doi: 10.3389/fpsyg.2011.00178
Westphal-Fitch, G., and Fitch, W. T. (2018). Bioaesthetics: the evolution of aesthetic cognition in humans and other animals. Prog. Brain Res. 237, 3–24. doi: 10.1016/bs.pbr.2018.03.003
White, J., Lorenz, H., Perilloux, C., and Lee, A. (2018). Creative casanovas: mating strategy predicts using—but not preferring—atypical flirting tactics. Evol. Psychol. Sci. 4, 443–455. doi: 10.1007/s40806-018-0155-7
Winegard, B., Winegard, B., and Geary, D. C. (2018). The status competition model of cultural production. Evol. Psychol. Sci. 4, 351–371. doi: 10.1007/s40806-018-0147-7
Woodley of Menie, M. A. (2015). How fragile is our intellect? Estimating losses in general intelligence due to both selection and mutation accumulation. Personal. Individ. Differ. 75, 80–84. doi: 10.1016/j.paid.2014.10.047
Woodley of Menie, M. A., and Kanazawa, S. (2017). Paternal age negatively predicts offspring physical attractiveness in two, large, nationally representative datasets. Personal. Individ. Differ. 106, 217–221. doi: 10.1016/j.paid.2016.11.003
Wright, D. B., Eaton, A. A., and Skagerberg, E. (2015). Occupational segregation and psychological gender differences: how empathizing and systemizing help explain the distribution of men and women into (some) occupations. J. Res. Pers. 54, 30–39. doi: 10.1016/j.jrp.2014.06.004
Zabelina, D. L., Colzato, L., Beeman, M., and Hommel, B. (2016). Dopamine and the creative mind: individual differences in creativity are predicted by interactions between dopamine genes DAT and COMT. PLoS One 11:e0146768. doi: 10.1371/journal.pone.0146768
Zahavi, A. (1975). Mate selection—a selection for a handicap. J. Theor. Biol. 53, 205–214. doi: 10.1016/0022-5193(75)90111-3
Zahavi, A., and Zahavi, A. (1997). The Handicap Principle: A Missing Piece of Darwin’s Puzzle, Oxford, UK: Oxford University Press.
Zhang, L., Lee, A. J., DeBruine, L. M., and Jones, B. C. (2019). Are sex differences in preferences for physical attractiveness and good earning capacity in potential mates smaller in countries with greater gender equality? Evol. Psychol. 17, 1–6. doi: 10.1177/1474704919852921
Keywords: creativity, intelligence, sexual selection, proximate, ultimate, mating, ornament, WEIRD
Citation: Novaes FC and Natividade JC (2023) The sexual selection of creativity: A nomological approach. Front. Psychol. 13:874261. doi: 10.3389/fpsyg.2022.874261
Edited by:
Marco Antonio Correa Varella, University of São Paulo, BrazilReviewed by:
Aaron Kozbelt, The City University of New York, United StatesCopyright © 2023 Novaes and Natividade. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Felipe Carvalho Novaes, ZmVsaXBlY2FydmFsaG8ubkBnbWFpbC5jb20=; Jean Carlos Natividade, amVhbm5hdGl2aWRhZGVAZ21haWwuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.