 André Schulz
André Schulz Claus Vögele
Claus Vögele- Institute for Health and Behaviour, Integrative Research Unit on Social and Individual Development, University of Luxembourg, Walferdange, Luxembourg
Afferent neural signals are continuously transmitted from visceral organs to the brain. Interoception refers to the processing of visceral-afferent neural signals by the central nervous system, which can finally result in the conscious perception of bodily processes. Interoception can, therefore, be described as a prominent example of information processing on the ascending branch of the brain–body axis. Stress responses involve a complex neuro-behavioral cascade, which is elicited when the organism is confronted with a potentially harmful stimulus. As this stress cascade comprises a range of neural and endocrine pathways, stress can be conceptualized as a communication process on the descending branch of the brain–body axis. Interoception and stress are, therefore, associated via the bi-directional transmission of information on the brain–body axis. It could be argued that excessive and/or enduring activation (e.g., by acute or chronic stress) of neural circuits, which are responsible for successful communication on the brain–body axis, induces malfunction and dysregulation of these information processes. As a consequence, interoceptive signal processing may be altered, resulting in physical symptoms contributing to the development and/or maintenance of body-related mental disorders, which are associated with stress. In the current paper, we summarize findings on psychobiological processes underlying acute and chronic stress and their interaction with interoception. While focusing on the role of the physiological stress axes (hypothalamic-pituitary-adrenocortical axis and autonomic nervous system), psychological factors in acute and chronic stress are also discussed. We propose a positive feedback model involving stress (in particular early life or chronic stress, as well as major adverse events), the dysregulation of physiological stress axes, altered perception of bodily sensations, and the generation of physical symptoms, which may in turn facilitate stress.
Introduction
Interoception, i.e., the perception of bodily processes, plays an important role for symptom generation in body-related mental disorders, such as panic disorder (PD; Ehlers and Breuer, 1996), somatoform disorders (SDs; Bogaerts et al., 2010; Pollatos et al., 2011b), dissociative disorders (Michal et al., 2014; Sedeno et al., 2014; Schulz et al., 2015b), or eating disorders (Pollatos et al., 2008; Herbert and Pollatos, 2014). Stress responses involve a complex neuro-behavioral cascade, which is elicited when the organism is confronted with a potentially harmful stimulus, and includes cognitive (e.g., facilitation of selective attention; Chajut and Algom, 2003), affective (e.g., anxiety or fear; McEwen, 2000), and physiological changes [e.g., activation of the autonomic nervous system (ANS) and the hypothalamic-pituitary-adrenocortical (HPA) axis; Chrousos and Gold, 1992]. Exposure to an acute stressor of limited duration does not normally affect an organism’s health. Nevertheless, the experience of early life stress, chronic stress or major adverse events may represent one of the most important risk factors for mental disorders, cardiovascular diseases, and auto-immune disorders. Previously, it has been proposed that stress alters the central representation of bodily processes (Craig, 2002). Although interoception may be affected differentially across disorders, it is likely that the role of stress in the etiology of these disorders is mediated by interoception. As the knowledge on the interaction of interoception, stress and mental disorders associated with physical symptoms is only fragmentary (e.g., limited to a single interoceptive indicator, to one stress test or one physiological stress axis, or to one mental disorder), the objective of the current review is to summarize and synthesize existing knowledge from both ‘normal’ and ‘dysregulated’ interoception in mental disorders. This review of the existing literature may help to identify yet unanswered questions in this field and our synthesized framework model as developed in this review may stipulate further research in the area.
Structure of this Review
In the current paper we describe the psychobiology of interoception and stress, as well as a framework to demonstrate why both concepts are interconnected and of immediate relevance for health and disease (see The Ascending and the Descending Branch of the Brain-Body Axis). First, we briefly describe both physiological stress axes with specific attention on processes, which may be relevant for the interaction of interoception and stress (see Physiological Stress Axes). We then review the literature on the relationship between interoception and stress, as well as on the role of interoception in mental disorders, which are associated with both dysregulations of physiological stress axes and physical symptoms (see Method and Results). Before we present the results of our literature search, we summarize methodological approaches to assess facets of interoception, which have been used in the relevant literature (see Methodology to Assess Interoception). The summary of findings on interoception and stress (see Synthesis of Findings on Interoception and Stress) is separated into effects of acute and chronic stress on interoception, and into effects of both physiological stress axes on interoception. The synthesis of findings results in a postulation of a framework model incorporating a positive feedback loop that describes the relationship of (chronic) stress, dysregulation of stress axes, altered perception of bodily sensations and physical symptom. Further, we integrate findings on interoception and stress into existing knowledge on the neurobiology of interoception and speculate on the possibly underlying neuro-endocrine signal circuitry (see Neuroendocrine Pathways). Finally, we identify a number of psychological mechanisms (e.g., attention, learning, intuitive decision making), which are known to be affected by interoception and stress and which may play an additional role in the proposed framework model (Psychobiological Mechanisms Involved in Interoception and Stress).
Physiological Stress Axes
Physiological systems to provide resources to cope with the confrontation with a potentially harmful stimulus (e.g., elicitation of a fight-or-flight response) can be sub-divided into two partially independent stress axes: (1) The sympatho-adreno-medullary (SAM) axis, including components of the ANS and (2) the HPA axis (for a detailed review: see Chrousos and Gold, 1992; Chrousos, 2009).
The SAM axis represents a heterogeneous network of neural and endocrine functions, which are interconnected to activate sympathetic processes. The release of corticotropin releasing factor (CRF) as neurotransmitter in the locus coeruleus (LC) leads to the activation of medullary centers, which control the sympathetic nervous system. When activated, sympathetic processes may stimulate two mechanisms with different pathways: (a) a neural pathway via ganglia, which innervate effector organs over mainly noradrenergic synapses and (b) an endocrine pathway that elicits the release of catecholamines (e.g., epinephrine and norepinephrine) into the circulation by the adrenal glands. Circulating catecholamines stimulate effector organs via specific adrenergic receptors (e.g., β1-adrenergic receptors at the myocardium). The LC is part of the central noradrenergic system and connected with structures in the limbic system (e.g., amygdalae) and the frontal cortex. Central α2-adrenergic receptors in the noradrenergic system, mainly located in the LC and the nucleus tractus solitarius (NTS), may down-regulate sympathetic activation and thus represent a negative feedback mechanism in the SAM axis (Isaac, 1980).
The HPA axis involves three consecutive stages: when encountering a stressor, (a) CRF is released from the hypothalamus into the blood circuit, which elicits (b) a release of adrenocorticotropic hormone (ACTH) from the pituitary. Circulating ACTH is registered by (c) the adrenal cortex that releases cortisol in humans, or corticosterone in rodents. Cortisol (or corticosterone) may inhibit the production of both CRF and ACTH and thus constitute a negative feedback loop. Receptors for glucocorticoids, such as cortisol, are distributed over all cells in the entire body. Nuclear mineralocorticoid and glucocorticoid receptors may slowly induce changes in gene transcription over a time frame of hours (De Kloet et al., 1998). Furthermore, glucocorticoids may also elicit rapid, non-genomic effects on cells (mediated via membrane receptors) within several minutes (de Kloet et al., 2008; Groeneweg et al., 2011). As every cell is potentially affected by circulating cortisol, it is a challenge for stress research to understand signaling pathways. Cortisol, for example, has been shown to affect metabolism, immune function and various CNS processes, such as sleep and activity, learning and memory, and attention. Nevertheless, CRF and ACTH may also affect CNS processes, as is evident from their role in anxiety and depression (Arborelius et al., 1999; Strohle et al., 2000). Given the interdependence between CRF, ACTH and cortisol via negative feedback mechanisms, all in vivo relationships between cortisol and psychological processes cannot be solely attributed to cortisol. However, since the majority of findings on HPA axis activity focuses on cortisol effects, the current review will focus on the latter.
It should be noted that the two axes are not fully independent from each other. For instance, circulating CRF may inhibit central noradrenergic processes, while noradrenergic activity inhibits CRF production in the hypothalamus (Chrousos and Gold, 1992). There is evidence for between-individual specificity in stress response patterns (Fahrenberg and Foerster, 1982), which is assumed to play a role for individuals’ vulnerability for certain disorders (Cohen and Hamrick, 2003). This partial independence of the two stress axes may also imply that a response in one stress axis may not necessarily be associated with the other, depending on individual and environmental factors.
The Ascending and the Descending Branch of the Brain–Body Axis
The strong relationship between stress and interoception is illustrated by the fact that both processes reflect the communication between the CNS (i.e., the brain) and the periphery (i.e., the body). On the one hand, interoception describes the processing and perception of internal bodily states, which are transmitted from peripheral organs, presumably via afferent nerve fibers, to the brain. Interoception can, therefore, be seen as an example for ascending information on the brain–body axis. On the other hand, stress, can be seen as a prominent example of communication on the brain–body axis in descending direction (Schulz, 2015), as physiological stress responses affect the activity of peripheral organs, and metabolic and immune functions via neural and endocrine pathways.
It is important to note that ascending and descending information between the brain and the body is continuously exchanged, and not only during interoception and stress. Homeostatic regulation of peripheral physiological processes via neural and endocrine feedback loops typically work without the involvement of higher CNS functions, unless they are working beyond a ‘normal’ range of functioning. If they exceed a certain threshold, they may be pushed into awareness of the organism. A physiological stress response elicits the activation of the SAM and HPA axes, which is far above the ‘normal’ range of homeostatic regulation, a process that has been named ‘allostasis’ (McEwen, 2004). Stress can, therefore, be conceptualized as descending information on the brain–body axis of increased amplitude as compared to a state of homeostasis, which may shift into focus of awareness. Interoception implicates that awareness is focused on bodily functions, even if they work in a normal, homeostatic range.
In summary, the relationship between interoception and stress involves a bi-directional communication on the brain–body axes. The aims of this review are to elucidate (1) the bi-directional communication between the brain and the body and (2) its role in the etiology of mental disorders. Using a systematic literature review we investigated the relationship between descending (stress) and ascending (interoception) transmission on the brain–body axis. We further address the question whether the dysregulation of bi-directional communication on the brain–body axis facilitates the generation of physical sensations. Furthermore, we defined psycholological processes (anxiety, attention, learning, decision making) that have previously been shown to be associated with stress.
Method
A systematic literature review was performed on the relationship between interoception and stress, and between interoception and mental disorders, which are associated with stress and physical symptoms (i.e., depression, PD, SDs, dissociative disorders) using Pubmed/Medline, PsycInfo and PSYNDEX. Primary keywords used were “interoception,” “heartbeat perception/detection,” “visceral perception” and “symptom perception.” To address research objective (1), primary keywords were combined with secondary keywords “stress,” “anxiety,” “autonomic/sympathetic nervous system,” “nor/epinephrine,” “HPA axis” and “cortisol.” To address objective (2), primary keywords were combined with keywords “depression/depressive,” “panic/agoraphobia,” “somatoform,” “fucntional disorder/syndrome/complaint,” “medically unexplained symptoms,” “dissociation/dissociative” and “depersonalization/derealization.” On abstract-based search, studies were identified to contain at least one interoceptive indicator and either an experimental task that is considered inducing stress in a laboratory environment, and/or a player in physiological stress axes, and/or self-reported stress, and/or the inclusion of at least one of the respective mental disorders. Secondary literature as provided by the identified papers was also screened according to these criteria. For supplementary sections addressing the relationship between interoception psychological processes, which are affected by stress (anxiety, attention, learning, decision making), non-exhaustive literature searches were performed. Therefore, primary keywords were combined with secondary keywords dependent on the topic of the respective section. Based on the identified literature, interoceptive research methods are summarized and discussed in Section “Terminology and Definition,” which precedes the integration of the main results. Extraction of literature was performed independently by both authors.
Results
The search on the relationship between interoception and stress resulted in 24 studies, which are exhaustively reported below (see Table 1). Furthermore, we identified nine studies on interoception in depression (one review), 14 studies on interoception in PD (three reviews), 12 studies on interoception and somatoform/functional disorders, and five studies on interoception in depersonalization disorder (DPD; one hypothesis paper). Due to space limitations the search was restricted to studies investigating the relationship between interoception and stress or stress-related processes.
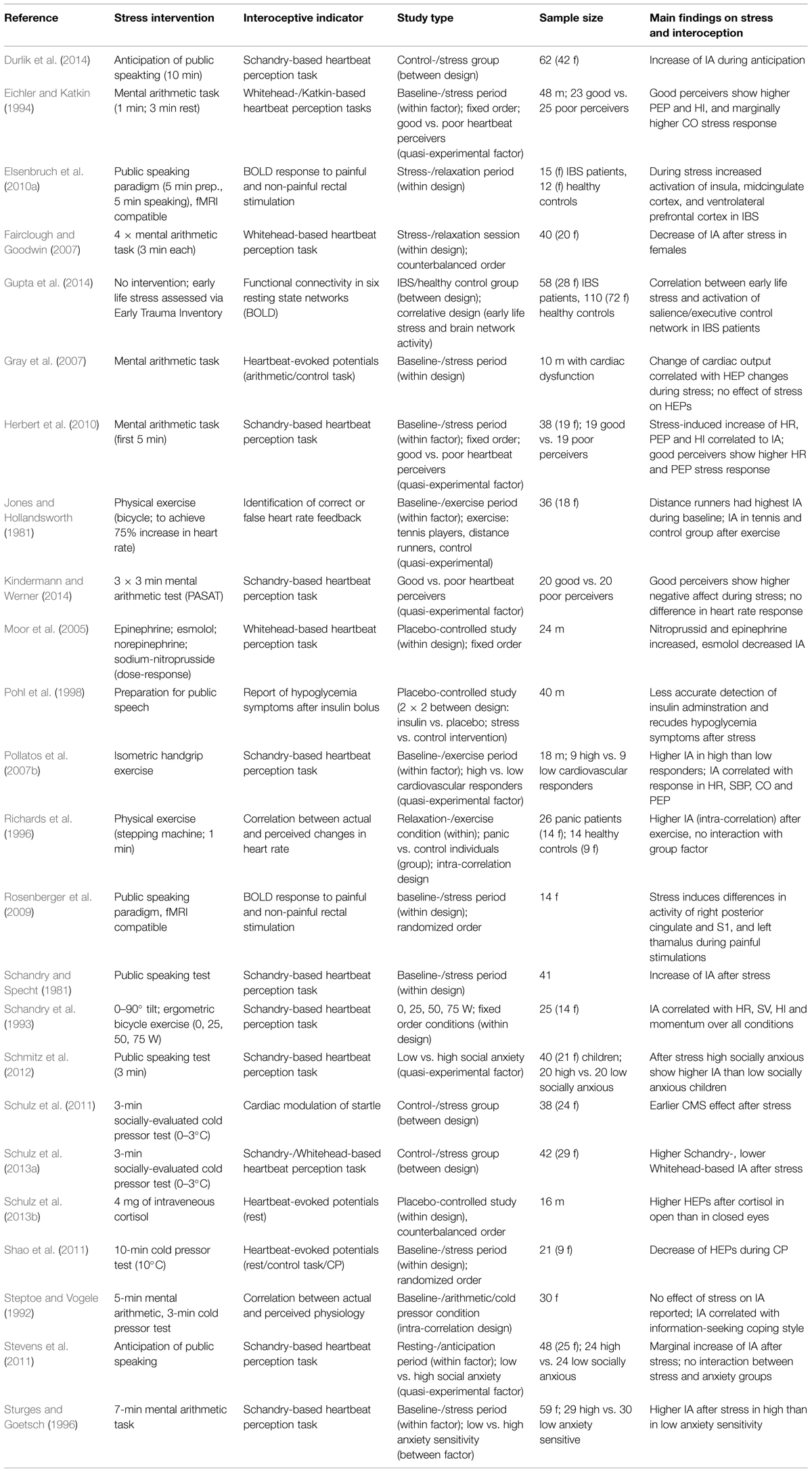
TABLE 1. Summary on empirical research papers addressing acute or chronic stress and interoception.
Terminology and Definition
Sensory information from the body can originate from different sources, i.e., (1) from exteroceptors, e.g., located in the skin, which transmit somatosensory information, (2) from proprioceptors, e.g., receptors in the spindles of skeletal muscles, and (3) from interoceptors, e.g., mechano-, chemo-, thermo-, or metabo-receptors within visceral organs (Schulz, 2015). In fact, signals from interoceptive and somatosensory sources are likely to be integrated in CNS structures to construct a representation of bodily processes (Craig, 2002, 2009; Wiens, 2005). This could explain, for instance, why patients with damaged or degenerated afferent autonomic nerves show impaired perception of visceral sensations (Pauli et al., 1991; Leopold and Schandry, 2001; Schulz et al., 2009a), while largely maintaining their affective experience (Cobos et al., 2004; Heims et al., 2004), which is assumed to be associated with interoception. However, the current review makes the pre-assumption that visceral-afferent signal transmission plays an important, yet not an exclusive role in the central representation of bodily signals.
There has been a debate in the literature on the taxonomy of facets of interoception. Garfinkel and Critchley (2013) have suggested separating the subjective tendency to be focused on interoceptive sensations from the actual accuracy in interoceptive tasks. They call the former ‘interoceptive sensibility’ and the latter ‘interoceptive sensitivity.’ In between, a third level was proposed, which was hypothesized to represent the degree of predictive value between interoceptive sensibility and sensitivity, which was named ‘interoceptive awareness.’ This taxonomy is in partial disagreement with earlier studies, which consider accuracy in heartbeat perception tasks to be an index of ‘interoceptive awareness.’ It is undisputed that awareness is required to perform in heartbeat perception tasks, but empirical evidence clearly shows that ‘awareness’ to interoceptive sensations and performance in heartbeat perception are partially unrelated (Khalsa et al., 2008; Ceunen et al., 2013). Although the differentiation between ‘sensibility’ and ‘sensitivity’ is plausible, in our opinion the measure indexed by interoceptive tasks should be called ‘interoceptive accuracy’ (IA) instead. ‘Interoceptive sensitivity’ originates from signal detection theory and implies the minimum threshold to detect an interoceptive signal from background noise (Farb et al., 2015). In contrast, ‘IA’ refers to the objective performance in counting interoceptive sensations or in discriminating and interoceptive from an exteroceptive sensation (see below; Ceunen et al., 2013). ‘IA’ does not suggest that a specific score reflects ‘sensitivity’ or ‘awareness,’ as some individuals may be hyper-sensitive or show extreme awareness to interoceptive sensations, which leads to misinterpretation of signals and thus to lower accuracy, while others may rarely focus awareness toward interoceptive stimuli, but may be highly accurate in performing heartbeat perception tasks (Schulz, 2015). In a revised version of the theory, performance in heartbeat perception tasks was now called ‘IA’ (Garfinkel et al., 2015), which is also reflected in this review.
The perception of bodily processes involves at least three consecutive stages: (1) afferent neural signals from the body, such as visceral organs, (2) the direction of attention toward bodily sensations, and (3) the evaluation of these signals and their integration into psychological processes. As suggested by Vaitl (1996) these three stages can be further differentiated. For instance, visceral-afferent signals involve the stimulation of interoceptors at the peripheral organ, the neural transmission of these signals from interoceptors to CNS structures, and finally the CNS representation of these signals. The evaluation of bodily sensations and their integration into psychological processes are associated with subjective reports of physical sensations, and an individual learning history concerning these sensations.
Methodology to Assess Interoception
Interoceptive Sensibility
The meta-cognitive tendency (i.e., interoceptive sensibility) to focus on interoceptive sensations is commonly assessed via questionnaires. The majority of existing questionnaires is designed to assess physical symptoms and clinically altered interoception in mental disorders, such as eating disorders (Eating Disorder Inventory-2: Garner, 1991) or SDs (Somatosensory Amplification Scale: Barsky et al., 1990). Instruments for the assessment of ‘normal’ interoception are scarce. Several existing studies have focused on the Body Perception Questionnaire by Porges (1993). However, there are no sufficient estimations for psychometric properties available for this questionnaire. The recently developed Multidimensional Assessment of Interoceptive Awareness (MAIA; Mehling et al., 2012) incorporates a number of interoceptive aspects (noticing, non-distracting, not-worrying, attention regulation, emotional awareness, self-regulation, body-listening, trusting) and is thus designed to study ‘normal’ and clinically dysregulated interoception.
Interoceptive Accuracy
The majority of empirical data on interoception is based on ‘IA’ as assessed by experimental paradigms that require subjective reports of physical sensations. Regarding the cardiovascular system, there are two main categories of tasks: (1) heartbeat counting tasks and (2) heartbeat discrimination tasks, as summarized elsewhere (Schulz, 2015). Heartbeat counting tasks were developed by Schandry (1981), who named the task ‘mental tracking test.’ The task consists of time intervals of different duration (original version: 35, 45, and 55 s), during which participants are instructed to silently count the number of their heartbeats. This number is later compared to the actual number of heartbeats in this interval. The absolute value of the difference between both is divided by the actual number of heartbeats and this ‘inaccuracy’ index is then subtracted from 1, which results in the accuracy score of the heartbeat counting task. It has been repeatedly demonstrated that the accuracy score depends on the wording of the instruction given. The comparison between a standard instruction (“count all heartbeats you feel in the body”) and a strict instruction (“count only those heartbeats about which you are sure of”) suggests that an individual’s knowledge on their heart rate and capacity to accurately estimate the duration of time intervals may be important factors for the accuracy in heartbeat counting tasks (Ehlers et al., 1995; Ehlers and Breuer, 1996). Heartbeat discrimination tasks were originally developed by Brener and Jones (1974) and further elaborated by others (Whitehead et al., 1977; Katkin et al., 1981; Brener and Kluvitse, 1988). In these tasks participants are asked to judge whether a set of consecutive exteroceptive stimuli (e.g., lights, tones, tactile stimuli) appear simultaneously with their own heartbeats (S+ trials) or not (S- trials). The available variants of this task mainly differ in the setup of S- trials, since some present exteroceptive stimuli with a fixed delay to heartbeats, while others simulate a set of artificial stimuli without any relation to the actual heartbeats. Previous research has yielded mixed findings regarding the question whether IA assessed by both tasks families are correlated: some report a moderate positive correlation (Knoll and Hodapp, 1992; Schaefer et al., 2012; Hart et al., 2013), while others did not find any association (Phillips et al., 1999; Schulz et al., 2013a; Michal et al., 2014).
Beyond heartbeat perception tasks, subjective reports are also required for paradigms incorporating the perception of respiratory resistances (Pappens et al., 2013; Tsai et al., 2013; Petersen et al., 2014), of gastric distensions (Rosenberger et al., 2009; Elsenbruch et al., 2010b), and of non-specific skin conductance fluctuations (Andor et al., 2008). In our view, the outcomes of these tasks reflect all stages of interoception at any given point in time (i.e., learning, report, awareness, CNS representation, visceral-afferent signal transmission, and stimulation of interoceptors). We would argue, therefore, that in addition to these measures, methods are required that enable the differentiation of interoception stages, e.g., between those at lower and higher levels of awareness.
Psychophysiological Methods
To date, at least three psychophysiological indicators are available to assess interoceptive signal transmission below the threshold of consciousness: (1) Baroreflex-sensitivity (BRS) quantifies the changes in heart period in responses to changes in arterial blood pressure (Robbe et al., 1987). As this brainstem reflex requires intact baro-afferent signals transmission, BRS is considered an indicator of the integrity of afferent autonomic nerves (Frattola et al., 1997). The disadvantage of this indicator is the fact that it reflects both the afferent and efferent branch of the baroreflex at a time. (2) Using the ‘cardiac modulation of startle’ (CMS) paradigm we could show that visceral-afferent neural signals from the cardiovascular system may affect acoustic startle responses (Schulz et al., 2009a,b,c, 2011). This modulation is largely diminished in individuals with degeneration of afferent autonomic nerves (Schulz et al., 2009a) and may also involve afferent signals from other organ systems (Schachinger et al., 2009; Schulz et al., 2013b, 2015c). The CMS effect can be interpreted as an indicator of visceral-afferent neural signal processing at brainstem level, as it probably only involves brainstem mechanisms and occurs below the threshold of consciousness, (3) viscerally evoked brain potentials are considered indicators for the cortical representation of afferent signals from visceral organs. They can be further differentiated into heartbeat-evoked potentials (HEPs), which are related to the processing of heartbeats (Schandry et al., 1986; Leopold and Schandry, 2001), and respiratory-related potentials that are observed when an inspiratory or expiratory occlusion is presented (Davenport et al., 1986; Chan and Davenport, 2010; von Leupoldt et al., 2010).
Another psychophysiological indicator of interoception is the single trial covariance between event-related brain potentials (N300) and changes in heart period (∼3–4 s after stimulus) in response to feedback in decision making tasks (Mueller et al., 2010a, 2012, 2013). This indicator is limited, however, to the use with decision making paradigms involving feedback. The theoretical background for this effect is provided by Damasio’s (1994) “somatic marker hypothesis” positing that afferent information from visceral organs is integrated in the evaluation of alternatives in decision making. It is plausible that the covariation between cortical and cardiac responses is specifically relevant for decision making paradigms tasks, although it is for future research to show whether this effect also extends to other embodied cognitions.
Synthesis of Findings on Interoception and Stress
Based on the findings from the current review, we propose a conceptual framework that allows for the explanation of the role of the dysregulated association between interoception and stress for the generation of physical symptoms (see Figure 1). In this model we postulate that stress is the initial point of a positive feedback cascade. In case of an acute stressor, the cascade will follow the pathway b–d–e. In particular, acute stress will activate physiological stress axes, such as the release of cortisol and activation of peripheral organs by sympathetic mechanisms. The altered stimulation of peripheral interoceptors and central effects of cortisol and noradrenergic structures will affect the perception of bodily sensations (e.g., perception of tachycardia or positive cardiac inotropy). Alterations in the perception of bodily sensations may temporarily feed into the experience of physical symptom during acute stress, for instance, palpitations, nausea or breathlessness. The perception of these symptoms could then be perceived as stressful and thus contribute to the maintenance of a stress response. However, due to the limited duration of an acute stress response, the cascade will be disrupted after the stress-eliciting stimulus has disappeared. In case of the confrontation with a chronic, early or major life stressor, the dysregulation of physiological stress axes (e.g., chronic hyper-activation of the HPA axis or the SAM axis) is implied and the model will follow the pathway a-c-d-e. Once the manifestation of a stress axis disorder has occurred, the cascade cannot be easily disrupted anymore, even if the initial stressor has disappeared. In detail, if the organism experiences chronic dysregulation of a stress axis, this state may permanently induce altered perception of bodily sensations and contribute to the manifestation of physical symptoms, whose perception consolidates the experience of stress. This model has an intended similarity to the model of somatosensory amplification by Barsky et al. (1988) and Barsky (1992), which emphasizes the importance of positive feedback mechanisms in activation, stress and physical symptoms. In contrast to the model by Barsky et al. (1988) however, the current model makes no assumptions about the role of attention to bodily sensations, although the latter could be conceived of as one of several psychobiological mechanisms. The core assumption in the current model concerns the interrelation of ascending and descending signals on the brain body axis in a positive feedback fashion. The following paragraphs will summarize empirical findings supporting this model.

FIGURE 1. Synthesis of findings on acute and chronic stress, dysregulation of physiological stress axes, altered interoception and the generation of physical symptoms into model comprising a positive feedback loop.
Chronic Stress and Dysregulation of Physiological Stress Axes (Path a)
It is well documented in the literature that prolonged exposure to psychosocial stress, early life adversity and/or major adverse events represent major risk factors for the dysregulation of the physiological stress axes. The regulation of the HPA axis can be differentiated into three different mechanisms (Li-Tempel et al., submited): (1) baseline activity and circadian rhythmicity, such as morning or daily profiles in cortisol level, (2) reactivity of CRF, ACTH, and cortisol in response to acute stress, and (3) (negative) feedback sensitivity, such as the suppression of cortisol release by dexamethasone. In general, similar mechanisms also exist for the SAM axis. In particular, baseline concentrations of catecholamines can be assessed from blood or urine, indirectly from salivary samples via alpha-amylase, or psychophysiological indicators, such as pre-ejection period. The investigation of circadian rhythmicity is limited due to the very short plasma half-life of catecholamines (∼1–3 min.). Furthermore, the same indicators can be investigated in response to an acute stressor. Negative feedback mechanisms in the SAM axis are, for instance, central α2-adrenergic receptors. These receptors are critically involved in brainstem-relayed reflex circuits that regulate the homeostasis of sympathetic and parasympathetic output, e.g., the arterial baroreflex (Sved et al., 1992). As the players involved in SAM axis regulation involve complex neural and endocrine pathways and are, therefore, more heterogeneous, the same is true for their indicators. For example, some parameters, such as low frequency heart rate and blood pressure variability are more sensitive to central sympathetic activation, while others, such as pre-ejection period are indicative of peripheral sympathetic activation (e.g., circulating catecholamines; Schachinger et al., 2001).
Early life and chronic stress, as well as major adverse life events, are consequently associated with dysregulation in all three types of mechanisms and in both stress axes (McEwen, 2000). Regarding HPA axis baseline and circadian rhythmicity, for instance, chronic stress, resulting from caregiving to family members, is reflected in altered cortisol awakening response (CAR; de Vugt et al., 2005), reduced cortisol release during daytime (Bella et al., 2011), and elevated hair cortisol levels (Stalder et al., 2014). In chronic stress associated with peer bullying and victimization reduced CAR was observed (Knack et al., 2011). Aberrant cortisol responses to acute stressors have been also found in family caregivers (De Andres-Garcia et al., 2012) and those exposed to peer victimization (Knack et al., 2011). Feedback sensitivity by dexamethasone suppression is altered in individuals exposed to high work stress, as indicated by self-reported burnout symptomatology (Pruessner et al., 1999), and after major traumatic experiences (Yehuda et al., 1993). Regarding the SAM axis, chronic stress by caregiving is associated with baseline indices of peripheral sympathetic activation (Cacioppo et al., 2000). Work stress as indicated by over-commitment to work is related to lower baseline norepinephrine levels (Wirtz et al., 2008), but increased activity in psychophysiological indicators of peripheral epinephrine circulation (Vrijkotte et al., 2004) and lower central parasympathetic output (Chandola et al., 2010). Furthermore, lower norepinephrine responses to acute stress have been found in individuals high in over-commitment, suggesting high work-related stress (Wirtz et al., 2008). Individuals exposed to early life stress show blunted reactivity to acute stress in indicators reflecting peripheral sympathetic activation (i.e., pre-ejection period; McLaughlin et al., 2014). Finally, there is evidence from animal models of reductions in the expression of α2-adrenergic receptors (which are involved in SNS-down-regulation) in response to chronic psychosocial stress (Flugge et al., 2003). The arterial baroreflex circuit, which is partially mediated by α2-adrenoceptors (Sved et al., 1992), shows reduced functioning under conditions of chronic work stress (Thomas et al., 2004).
Chronic stress and the associated dysregulation of physiological stress responses represent important factors for the development and maintenance of mental disorders, in particular those, which are associated with symptoms of altered perception of bodily sensations (e.g., depression, PD, SDs, dissociative disorders). Nevertheless, the current literature lacks a discussion of the mechanisms underlying the relationship between stress, interoception and physical symptoms. For example, it has been repeatedly suggested that somatic syndromes, e.g., chronic fatigue syndrome (CFS) or fibromyalgia (FMS), are associated with lower cortisol responsiveness or increased feedback sensitivity (Cleare, 2004; Tanriverdi et al., 2007; Wingenfeld et al., 2008; Tak et al., 2011), but only few hypotheses addressing the mechanisms of how cortisol may contribute to the generation of physical symptoms. One such hypothesis addresses immunological changes that are associated with dysregulation of the HPA axis (Fries et al., 2005). It is possible that the modulation of pro- and anti-inflammatory cytokines may contribute to somatic syndromes, such as FMS. Another explanation focuses on the importance of hypocortisolism in CFS and argues that cortisol mobilizes energy resources to overcome daily demands, which are insufficiently available in CFS (Tak and Rosmalen, 2010). However, both explanations are focused on peripheral bodily processes and do not incorporate possible effects of stress on cortical representation and perception of bodily sensations. The possibility that cortisol could also modulate the perception of bodily sensations was first raised by Rief et al. (1998b).
Acute Stress and Interoception (Path b)
Laboratory Stress Tests
A broad variety of methods has been used to induce acute stress in laboratory settings (Steptoe and Vogele, 1991). Mental arithmetic tasks elicit ‘psychological’ stress generally by inducing mental load and specifically by involving central control mechanisms over autonomic processes (Moriguchi et al., 1992; Sloan et al., 1995). Psychosocial stressors that emphasize public speaking challenges, such as the Trier Social Stress Test (TSST), are known to provoke an intense response of the HPA axis (indexed e.g., by an increase of salivary cortisol; (Kirschbaum et al., 1993; Kudielka et al., 2004) and activation of the cardiovascular system by sympathetic mechanisms. Given the TSST’s comparatively long duration of ∼15 min on the one hand, and the very fast acting release and effects of catecholamines (i.e., seconds) on the other hand, specific sympathetic processes involved in the TSST are difficult to disentangle. As the cold pressor test induces stress associated with the experience of ischemic pain, these results in specific cardiovascular response patterns, which can partially be attributed to the experience of pain, but also to the vaso-constricting effect of cold water (Streff et al., 2010). The socially-evaluated cold pressor task (SECPT) attempts to integrate psychosocial and physical stressors, and has been demonstrated to elicit significant increases in saliva cortisol (Schwabe et al., 2008; Lass-Hennemann et al., 2010, 2011; Larra et al., 2014), as well as sympathetic activation, as indicated by an increase of heart rate, and systolic and diastolic blood pressure (Schwabe et al., 2008; Schulz et al., 2011, 2013a) and a decrease of pulse-transit time and BRS (Schulz et al., 2011). To understand possibly diverging results on the impact of acute stressors on indicators of interoceptive signal processing and IA, differences in the methods used to induce stress and to assess interoception have to be taken into account (Steptoe and Vogele, 1992).
Interoceptive Accuracy Assessed with Heartbeat Perception Tasks
Interoceptive accuracy as estimated by Schandry-based heartbeat tracking tasks typically increase under acute stress. While this effect has been observed in anticipation of (Durlik et al., 2014) and after a public speaking test (Schandry and Specht, 1981), one study only found a marginal increase in IA during anticipation (Stevens et al., 2011). The importance of the amplitude of cardiovascular responsiveness during stress for the enhancement of IA is underlined by the positive correlation between IA and responsiveness in heart rate, pre-ejection period and the Heather index in a mental arithmetic task (Herbert et al., 2010). A similar association between cardiovascular responsiveness and IA could also be shown for the heartbeat discrimination task (Eichler and Katkin, 1994). Acute responses to stress include sympathetic effects on the cardiovascular system, and a delayed increase in cortisol secretion, if a psychosocial challenge (e.g., public speaking) is involved. There are also attentional and affective changes in response to stress (see Psychobiological Mechanisms Involved in Interoception and Stress), which may interact with interoception. For instance, individuals with high IA in heartbeat perception show higher negative affect during stress than individuals with low IA (Kindermann and Werner, 2014). Furthermore, it has been shown that IA increases after physical exercise (Jones and Hollandsworth, 1981; Richards et al., 1996), and that the same correlation between cardiovascular reactivity measures (cardiac output, heart rate, systolic blood pressure, Heather index, momentum, stroke volume) to exercise and IA exists (Schandry et al., 1993; Pollatos et al., 2007b). It could be argued, therefore, that peripheral sympathetic activation during stress represents the core factor of this relationship. In contrast to these results using Schandry-based heartbeat tracking tasks, IA as assessed with heartbeat discrimination tasks, is reduced in women performing a mental arithmetic task (Fairclough and Goodwin, 2007). Using the SECPT we could replicate these seemingly contradictory findings, i.e., an increase in IA after stress as assessed by the Schandry-heart beat tracking task and a decrease in IA when using a visual heartbeat discrimination task (Schulz et al., 2013a). As the results obtained in these studies seems to be IA-paradigm dependent the observed differences are most likely associated with task specific characteristics: while heartbeat tracking tasks require participants to only focus their attention on visceral sensations, the heartbeat discrimination task involves the concurrent monitoring of visceral sensations and external signals. According to the competition-of-cues model by Pennebaker and Lightner (1980), interoceptive and exteroceptive signals compete for a limited resource, i.e., attention. As acute stress may narrow attentional resources and favor attention to task-relevant stimuli (Chajut and Algom, 2003; Plessow et al., 2011), the multisensory integration of information as required by the heartbeat discrimination task may be impaired by stress exposure. This assumption is further supported by the observation that after the preparation for public speech, individuals exhibited reduced accuracy in detecting an insulin-induced hypoglycemic state (Pohl et al., 1998), which also requires attention focused on multiple symptoms. In summary, there is evidence for two mechanisms relevant for IA in response to acute stress: (1) peripheral sympathetic activation, which induces an increased stimulation of cardiac interoceptors (e.g., arterial baroreceptors) and intensifies afferent signal transmission from the cardiovascular system, and (2) attention focus on visceral sensations, which may improve the detection of visceral signals, but diminish the integration of signals from other sensory modalities. When both effects are opposed, as e.g., in the heartbeat discrimination task, it appears that attention represents the more important determinant of IA. This dissociation emphasizes the need for the separate assessment of visceral-afferent signal transmission and representation on the one hand, and attention to visceral signals on the other hand.
Psychophysiological Indicators of Interoceptive Signal Processing
While HEPs and CMS may serve as indicators of visceral-afferent signal transmission independent of attention to these signals, empirical data on stress effects on these indicators is scarce. In one study we investigated the impact of the SECPT on CMS (Schulz et al., 2011). After stress exposure we observed CMS to occur earlier within the cardiac cycle (0–200 ms instead of 200–300 ms after the R-wave); yet, contrary to our expectations, there was no amplification of the CMS effect. Nevertheless, as the CMS reflects the intact transmission of afferent neural signals from the cardiovascular system at brainstem level (Schulz et al., 2009a), these findings suggest that the amplitude of representation of afferent signals may be unchanged by stress. Similar findings have also been reported for HEPs, with reduced amplitudes during a long-term, but mild cold pressor test (10-min, 10°C) and a return to baseline levels after the cold pressor test (Shao et al., 2011). This response pattern may be explained by participants focusing their attention during the cold pressor primarily on the pain experience, and thus away from cardiovascular sensations. Nevertheless, after termination of stress exposure, the expected sympathetic response did not affect HEP amplitudes. Gray et al. (2007) investigated the impact of a mental arithmetic task on HEPs in 12 men with cardiac dysfunction, and found a positive relationship between changes in cardiac output and HEPs, but no effect of the stress task on HEP amplitudes. To the best of our knowledge, there is only one study reporting an effect of acute stress on a psychophysiological indicator of interoception: after an MRT-compatible public speaking test, the BOLD response to painful rectal distensions was different in the right posterior cingulate and right somatosensory area, as well as the left thalamus (Rosenberger et al., 2009). Despite the large methodological differences across these studies in terms of laboratory stress tasks employed, organ systems investigated and derivation of interoceptive indicators, it may be speculated that acute stress specifically changes the processing of visceral-afferent signals from the gastrointestinal system. As the majority of studies demonstrate no quantitative change of raw representations of visceral-afferent signals by stress, it is still plausible that stress induces a qualitative change in interoception, as suggested by our findings (Schulz et al., 2011). It can be summarized that, contrary to expectations, current results implicate that there is no main effect of acute laboratory stress on the amplitude of psychophysiological indicators of interoception after the stressor. Possible reasons for the null findings in these studies could involve the limited effectiveness of mental arithmetic and 10°C cold pressor tasks, as well as the limited time frame after stress during which responses were monitored. Given the heterogeneity of mechanisms induced by acute laboratory stress, it seems necessary to determine the differential role of the physiological stress axes on interoception. Table 1 provides an overview of studies addressing the relationship of stress and indicators of interoception.
The Sympatho-Adreno-Medullary Axis and Interoception
As described earlier, reactivity in pre-ejection period or Heather index to laboratory stress, (which may reflect peripheral sympathetic activation) is positively related to heartbeat perception (Eichler and Katkin, 1994; Herbert et al., 2010). However, as neither study derived heartbeat perception scores from a post-stress period, it can only be concluded that individuals with a general tendency for responding with peripheral sympathetic activation show higher IA in heartbeat perception tasks. To elucidate the contribution of both the central and peripheral sympathetic branch to heartbeat perception, the impact of different adrenergic agents on accuracy in a heartbeat discrimination task has been investigated by (Moor et al., 2005). In particular, sodium nitroprusside as α1-adrenergic antagonist, and norepinephrine as α1-agonist, as well as the β1-agonist epinephrine and the β1-antagonist esmolol were employed. Since β1-adrenoreceptors are located in the myocardium and are sensitive to circulating catecholamines, especially epinephrine, selective stimulation or de-stimulation represents a pharmacological model for peripheral sympathetic activity (Schachinger et al., 2001). Epinephrine increased and esmolol decreased IA as assessed by a heartbeat discrimination task as compared to placebo (Moor et al., 2005). This finding suggests that peripheral sympathetic activation enhances IA. Since epinephrine cannot cross the blood–brain barrier, two alternative ways of signal transmission are possible: first, β-adrenergic receptors localized at vagal nerve endings may directly be stimulated by circulating epinephrine (Mravec, 2011). Second, increased cardiac contractility may cause increased stimulation of cardiac interoceptors (e.g., baroreceptors), whose neural signals are transmitted over the nervus glossopharyngeus (Jänig, 2006). α1-adrenoreceptors are primarily located in the vascular musculature. Their stimulation induces an increase in vascular resistance and, therefore, in blood pressure. This information is relayed via the arterial baroreflex circuit in order to decrease heart rate accordingly for the maintenance of blood pressure level. The dis-stimulation of α1-adrenoreceptors causes the opposite effect. This pharmacological design is thus suitable to investigate the selective loading and unloading of arterial baroreceptors and the subsequent central sympathetic activation induced by this baroreceptor stimulation (Schachinger et al., 2001). Baroreceptor unloading by sodium nitroprusside resulted in an increase in IA as compared to placebo, while norepinephrine had no effect on heartbeat perception (Moor et al., 2005), which could be explained by increased central sympathetic output due to baroreceptor unloading. Central α2-adrenoreceptors, which are mainly located in the LC and NTS act as negative feedback mechanism in the central noradrenergic system (Rockhold and Caldwell, 1980). Hence, administration of a α2-adrenergic antagonist causes a concurrent activation of central and peripheral sympathetic mechanisms (Isaac, 1980; Philippsen et al., 2007). In a pilot study we found the α2-antagonist yohimbine to suppress the CMS effect (Schulz et al., 2007). This finding implies that concurrent central and peripheral sympathetic activation combined with increased arousal and vigilance as caused by noradrenergic activation may diminish the central processing of visceral-afferent signals at a low, presumably brainstem-associated level. Our own observations on α2-antagonism and IA suggest reduced IA after the administration of yohimbine. This observation may implicate that central noradrenergic activation, including increased alertness and vigilance, may also impair the cortical processing of visceral-afferent signals. Taken together, it can be concluded that there is a strong positive relationship between peripheral sympathetic activation and cardiac IA, but only a limited association between central sympathetic tone and IA. A strong activation of central noradrenergic mechanisms may even inhibit interoception, although concurrent peripheral activation would suggest a more intense stimulation of interoceptors.
The HPA Axis and Interoception
Cortisol has been found to modulate HEP amplitudes within a timeframe of 1–17 min after infusion (Schulz et al., 2013c). More specifically, 4 mg of cortisol resulted in higher HEPs under open- compared with closed-eyes conditions, i.e., states of high vs. low alertness. This effect could eventually feed into a vicious circle of increased attention focus on physical symptoms, increased anxiety and higher levels of cortisol, and represent a psychobiological mechanism underlying positive feedback models of somatosensory amplification (Barsky et al., 1988). In this study HEPs were assessed during rest, without the conscious perception of heartbeats. The effect of cortisol administration on resting HEPs suggests that cortisol may affect the raw representation of visceral-afferent signals, independent of the conscious perception of heartbeats. Moreover, we did not observe any effect of cortisol on cardiovascular activation. This finding suggests that cortisol may selectively modulate the central, presumably cortical, representation of visceral-afferent signals, while the peripheral origination of these signals remain unaffected. Interestingly, in a complementary study no effect of 1.5 mg cortisol on the CMS was observed within the same time frame (Schulz et al., 2010). Since the CMS is assumed to reflect visceral-afferent signal transmission at brainstem level, one could speculate that the effects of cortisol on visceral-afferent signal relaying are restricted to the cortex. However, it needs to be acknowledged that the dosages of administered cortisol were not identical. Furthermore, the first study covered a time frame of up to 37 min and therefore includes possible genomic effects, while the second study focused on non-genomic effects only. The importance of cortisol for the cortical representation of visceral-afferent signals is further emphasized by the negative relationship between basal cortisol level and HEP amplitudes of r = -0.29 (Schulz et al., 2015b). Despite the fact that this result was based on a mixed sample of healthy individuals and patients with DPD, the correlation was unaffected by the diagnosis, or depression and anxiety scores. It can be summarized that acute cortisol administration tends to increase HEP amplitudes (when eyes are open), presumably via a non-genomic mechanism, while basal cortisol level shows a negative relationship with HEPs. One may speculate that the effect of cortisol on the cortical representation of visceral-afferent signals reverses into long-term cortisol elevations. Interestingly, these opposite effects contrast with findings in pain research: in experimental short-term manipulation of cortisol, an oral administration of 40 mg of cortisol reduces pain sensitivity (Michaux et al., 2012), while cortisol blockade by metyrapone intensifies pain perception (Kuehl et al., 2010). Meanwhile, in chronic pain syndromes, such as FMS, also reduced cortisol baseline levels and hyper-suppression by dexamethasone were observed (Wingenfeld et al., 2007, 2008). This possible dissociation in cortisol relationships between ‘normal’ visceral-afferent neural transmission and pain has to be acknowledged, since interoception and pain processing only partially share neural structures (Craig, 2002). Future research should clarify whether acute cortisol release may selectively favor the representation of ‘normal’ visceral-afferent transmission, which may reverse in long-term increases of cortisol levels. The majority of the existing literature on processes of body, symptom and pain perception concentrates on the role of cortisol, the final product of HPA axis activation. However, it needs to be taken into account that the role of CRF and ACTH on interoceptive processes remains unclear and possible differences in acute and chronic cortisol levels may also be attributed to feedback-induced changes in CRF or ACTH.
Taken together, there is considerable evidence to show that acute stress and players of physiological stress axes may alter the perception of bodily sensations. Based on our model, we propose that acute stress may induce an acute alteration of physical sensations and transient symptoms specific to the stress, which constitute the subjective experience of the stress response (e.g., tachycardia, palpitations, nausea, etc.). However, these symptoms disappear when the stress is of limited duration.
Dysregulation of Physiological Stress Axes and Interoception (Path c)
So far there is only partial and indirect evidence for a direct effect of dysregulated stress axes on altered perception of bodily sensations. Earlier findings suggest that chronic stress in healthy individuals is not related to heartbeat perception accuracy (Schulz et al., 2013a) or sensitivity for gastric stimulations (Rosenberger et al., 2009). The direct pathway between chronic stress and altered perception of bodily sensations was, therefore, omitted in our model. It should be noted, however, that moderately elevated levels of self-reported chronic stress in healthy individuals, as reported in the former studies, are unlikely to be accompanied by a dysregulation of the physiological stress axes.
Our assumption that the dysregulation of the physiological stress axes may induce altered perception of bodily sensations is mainly based on the following observations:
(1) In numerous mental disorders that are accompanied by physical symptoms, altered interoception and dysregulation of physiological stress axes are reported. In detail, (a) individuals with major depression (MD) and depressive symptoms exhibit reduced IA and HEP amplitudes (Dunn et al., 2007; Terhaar et al., 2012). Concerning the activity of the HPA axis, previous research has shown differences between the melancholic and the atypical sub-type (Gold and Chrousos, 2002; O’Keane et al., 2012; Gold, 2015): on the one hand, the atypical sub-type exhibited normal basal cortisol levels and increased dexamethasone-induced feedback sensitivity (Levitan et al., 2002; O’Keane et al., 2005; O’Keane et al., 2012). On the other hand, the more frequent melancholic sub-type is characterized by hypersecretion of CRF (Nemeroff et al., 1984; Wong et al., 2000), and reduced concentration and sensitivity of CRF neurons, resulting in a blunted ACTH response to CRF administration (Gold et al., 1988; Lesch et al., 1988; Ehlert et al., 2001), elevated basal cortisol levels (Gold et al., 1988; Ehlert et al., 2001; Gold, 2015), blunted cortisol responsiveness to acute stressors and reduced feedback sensitivity as provoked by dexamethasone (Gold et al., 1995; Pariante and Lightman, 2008). (b) Patients with PD show increased IA compared to healthy individuals (Ehlers and Breuer, 1992, 1996; Ehlers et al., 1995), whereas a meta-analysis has pointed out that this difference may be due to a minority within the PD group (Willem Van der Does et al., 2000). However, PD patients exhibit an increased coupling of feedback-evoked EEG amplitude and heart rate changes than healthy individuals (Mueller et al., 2014), which has previously been shown to be an indicator of neuro-visceral connectivity (Mueller et al., 2010a). Regarding the physiological stress axes, PD patients do not differ from healthy individuals in baseline morning or diurnal cortisol release or negative feedback sensitivity (Ising et al., 2012), but they show blunted cortisol responses to psychosocial stress (Petrowski et al., 2010, 2013). In terms of SAM axis activity existing studies have yielded mixed findings. While some failed to find differences in indicators of central (e.g., low frequency heart rate variability/HRV) or peripheral sympathetic activation (α-amylase) between PD and healthy controls (Tanaka et al., 2012), others observed reduced central (McCraty et al., 2001), but increased peripheral activation (Marshall et al., 2002). However, it has been proposed that dysfunction in α2-adrenergic regulation of the ANS represents an important neurophysiological correlate of PD (Bremner et al., 1996). (c) In SDs previous studies have yielded mixed findings, which may be partially explained by the heterogeneity of symptomatology collapsed in this diagnostic category. Some studies have failed to find differences in IA between patients with SD and healthy individuals (Barsky et al., 1995; Mussgay et al., 1999; Schaefer et al., 2012), while others observed exaggerated report of bodily sensations (Bogaerts et al., 2010), resulting in an overall decrease of IA (Pollatos et al., 2011a; Weiss et al., 2014). SD patients show both alterations in baseline cortisol levels, such as the CAR (Rief et al., 1998b) or daily profile (Tak et al., 2009; Tak and Rosmalen, 2010), and aberrant cortisol responses to psychosocial stress (Janssens et al., 2012). The question, therefore, is not whether dysregulation of physiological stress axes, altered interoception and physical symptoms are associated, but how they are related and which direction these relationships have.
(2) In pharmacological studies, the acute administration of catecholamines (Moor et al., 2005) or cortisol (Schulz et al., 2013c) affects interoception and interoceptive signal transmission. Despite the fact that chronically altered levels of adrenergic stress hormones or cortisol are not fully comparable to an acute administration, it is unlikely that chronic alterations of these hormones do not affect interoception at all. We interpret these findings as support for the assumed direction of physiological stress axes affecting interoception in our model.
Previous studies have addressed mental disorders with physical symptoms, interoception, and indicators of only one physiological stress axis in the same sample. Pollatos et al. (2011a) found lower IA in patients with SD to be accompanied with decreased autonomic balance, as indicated by low/high frequency HRV ratio. In a study by Ehlers et al. (1995) higher heart rates were observed in panic patients than in healthy individuals, suggesting increased sympathetic tone in patients with PD. As both stress axes may differentially affect interoceptive processes, however, future studies are needed that include indicators of both stress axes at a time. Currently, there is only one study using multiple indicators of both physiological stress axes in the investigation of physical symptoms associated with a mental disorder (Schulz et al., 2015b). Results show an insensitivity of HEPs for attention focused on heartbeats in patients with DPD, but not in healthy individuals. This difference was associated with higher basal level of salivary α-amylase, an indicator for peripheral sympathetic activation (Chatterton et al., 1996; Nater et al., 2005). Furthermore, across DPD patients and healthy individuals there was a negative correlation between basal cortisol level and HEP amplitude (Schulz et al., 2015b). In the same sample IA did not differ between DPD and healthy control individuals (Michal et al., 2014), although differences between DPD and healthy individuals have been reported in another study (Sedeno et al., 2014).
To elucidate the direction of the relationship between dysregulated stress responses and interoception, prospective long-term observations may be required. There is a notable lack of reports on interoception in chronically stressed individuals, who already show a dysregulation of both physiological stress axes, but do not yet fulfill the diagnostic criteria for a mental disorder associated with physical symptoms. To date, there is only one study investigating the relationship of early life stress and brain activity in healthy individuals and patients with irritable bowel syndrome (IBS): Gupta et al. (2014) report altered activation of brain networks associated with pain processing and early life stress in IBS patients. The importance of chronic stress for altered interoception in IBS is further underlined by the fact that IBS patients also show altered activation of brain regions associated with interoception during stress (Elsenbruch et al., 2010a).
The question remains whether there is a direct relationship between the dysregulation of physiological stress axes and the generation of physical symptoms without the mediating effect of interoception. Based on the currently available evidence this question cannot be unequivocally answered. We do not assume a direct pathway for the following reasons: (1) There is currently no model to explain the psychobiological processes connecting the dysregulation of physiological stress axes and the generation of physical symptoms, despite a broad empirical basis showing stress axes dysregulation in mental disorders to be associated with bodily symptoms, e.g., in depression (Holsboer, 2000; Ehlert et al., 2001; Gold and Chrousos, 2002; Pariante and Lightman, 2008), FMS (Tanriverdi et al., 2007; Wingenfeld et al., 2007, 2008); IBS (Chang et al., 2009; Suarez-Hitz et al., 2012), or CFS (Cleare, 2004; Nater et al., 2008). (2) Some direct relationships between stress axes and physical symptoms are even more heterogeneous than those observed between interoception and physical symptoms, e.g., in SD. Previous studies addressing interoception in SD have either produced null findings or shown reduced IA. Meanwhile, studies investigating indicators of stress axes in SD find directly opposing findings, ranging from higher (Rief et al., 1998a; Rief and Barsky, 2005), identical (Rief and Auer, 2000) to reduced cortisol output in SD patients compared to healthy controls (Heim et al., 2000; Tak and Rosmalen, 2010; Janssens et al., 2012). A popular way of explaining these potentially conflicting findings is to emphasize the heterogeneity of symptoms collapsed into SD classification. However, an alternative explanation would be a mediating factor between stress axes and physical symptoms, i.e., altered interoceptive processes.
Interoception and the Generation of Physical Symptoms (Path d)
Many mental disorders that are associated with physical symptoms are characterized by altered interoception, as discussed earlier in this paper. Among those, PD is a prominent example, where the experience of (frightening) physical symptoms is a defining diagnostic criterion. It is, therefore, not surprising that PD has had perhaps the longest standing history in the investigation of interoceptive processes, however, with very mixed results. There are reports of increased IA in panic patients (Ehlers and Breuer, 1992; Ehlers et al., 1995), but also those failing to find any differences in IA between panic patients and healthy individuals (Antony et al., 1995; Van der Does et al., 1997; Wolk et al., 2014). This mixed picture of results gave rise to the assumption that methodological differences (e.g., the precise wording of instructions for the most often employed heart-beat tracking task) between studies may account for these inconsistent findings (Ehlers et al., 1995; Ehlers and Breuer, 1996). Alternatively, the possibility of differences in IA between PD patients has been discussed (i.e., a PD subgroup high in IA; (Ehlers et al., 1995; Ehlers and Breuer, 1996; Van der Does et al., 1997; Willem Van der Does et al., 2000). Furthermore, the question arose whether IA may represent a risk factor for the development of PD symptoms. Supporting this assumption, IA was found to be higher in patients with maintained or relapsed PD and infrequent panic attacks, than in remitted patients (Ehlers, 1995). Moreover, exposure treatment did not change IA (Ehlers et al., 1995).
In contrast to PD, IA has been shown to be lower in SD (Bogaerts et al., 2010; Pollatos et al., 2011a; Weiss et al., 2014), and there also seems to be a positive association between symptom severity and IA impairment (Schaefer et al., 2012). Accordingly, reduced IA could be a risk factor for the development of SD. This hypothesis is supported in a recent study by Schaefer et al. (2014), showing that state symptom perception is reduced after cardiac IA training. Although the course of symptom severity in dependence of IA may suggest the direction of the assumed pathway, the quasi-experimental design of these studies prevents from causal inferences to be drawn. In fact, all of these studies included individuals who had already been diagnosed with a mental disorder at the time of the investigation and, therefore, a pre-morbid indicator of IA was not available. Again, to address this potential shortcoming, prospective studies in the general population or in specific high-risk groups (e.g., exposed to chronic stress) are required.
In addition to SD, reduced IA has also been observed in patients with anorexia nervosa (Pollatos et al., 2008) and obesity (Herbert and Pollatos, 2014), whereas intuitive eating is positively associated with IA (Herbert et al., 2013). The direction of the pathway between interoception and the generation of physical symptoms may depend on the type of mental disorder. As almost all studies in this field are cross-sectional and quasi-experimental in design, it is difficult to come to any conclusions on the direction of this relationship. Beyond clinically relevant eating behavior, the experimental manipulation of eating behavior using short-term food deprivation induces an increase in IA as assessed by heartbeat perception (Herbert et al., 2012) and HEP amplitudes (Schulz et al., 2015a). As some neuroendocrinological parameters, such as sympathetic tone or peptide YY output reverse from hyper- to hypo-activation in long-term fasting, one may argue that eating behavior could serve as a coping mechanism to regulate the perception of bodily sensations. The direction of the relationship between interoception and physical symptoms in eating disorders may, therefore, be reversed in disordered eating (path d) or even bi-directional. It needs to be acknowledged that this interpretation is based on experimentally manipulated eating behavior and its translation to pathological eating behavior (i.e., eating disorders) should be done with caution. In the interest of greater simplicity of our suggested model, we have refrained, however, from including this reversed path d, and acknowledge, therefore, that the current model is limited to physical symptoms and disorders that imply the passive experience of altered bodily sensations (e.g., MD, PD, SD, DPD), and does not apply to physical symptoms that are partially induced by disordered eating behavior.
Physical Symptoms and Stress (Path e)
It is widely accepted that the repeated and enduring experience of physical symptoms is (probably causally) related to the experience of stress, as denoted by the term ‘symptom distress.’ The majority of reported physical ‘somatization’ symptoms involve the experience of pain (Rief et al., 2001). Chronic pain is described in the literature as severe ‘inescapable stress.’ In animal models chronic pain has been shown to induce depressive-like symptoms, which may provide a model for the co-occurrence of chronic stress, pain and depression in humans (Blackburn-Munro and Blackburn-Munro, 2001). In support of this notion, individuals with chronic pain exhibit higher perceived chronic stress and higher cortisol as assessed from hair (Van Uum et al., 2008), an indicator sensitive to chronic stress (Russell et al., 2012). Symptom distress may not necessarily have sufficient quality and intensity to be comparable to severe chronic psychosocial stress, such as caregiving to family members or harassment at working place. Nevertheless, the current model posits that the repeated exposure to physical symptoms may induce stress, thereby increasing and adding to the experience of already existing stress. In SD, symptom distress is largely affected by SD-typical forms of behavior, such as inadequate reassurance or negative interactions between patient and doctor (Rief and Broadbent, 2007). As symptom distress is associated with the automatic negative evaluation of afferent somatosensory signals (Witthoft et al., 2012), it could be hypothesized that specific cognitive styles may contribute to the positive feedback mechanism relating stress and physical symptoms. The correlational design of this study, however, does not allow for any such causal interpretations, i.e., whether automatic negative evaluation is a cause or consequence of symptom distress. In the current model, negative evaluation style may be seen as a moderator of the pathway between physical symptoms and stress. In summary, we propose that via the impact of physical symptoms on stress, the vicious circle between stress and symptoms are completed, which is characteristic of a self-maintaining positive feedback mechanism.
Neuroendocrine Pathways
Comprehensive reviews on neural structures supporting interoceptive signal processing, and original research papers including neuroimaging data are available elsewhere (Cameron, 2001; Craig, 2002; Critchley et al., 2004; Pollatos et al., 2007a,c). In the following we provide a summary of neural pathways that are important for transmitting and processing afferent signals from the body to the brain, and which may also be involved in the experience of stress. The majority of the available literature focuses on interoception of cardiac sensations. As previously described (Schulz et al., 2013c), visceral-afferent neural signals from the cardiovascular system are relayed over the NTS, the major sensory center for visceral-afferent neural signals in the brainstem (Jänig, 2006). The NTS projects onto the parabrachial nucleus and the LC, from where hypothalamic and thalamic nuclei are reached (Cameron, 2001). Cortical structures that process visceral-afferent neural signals include the anterior cingulate cortex (ACC), the frontal cortex, the somatosensory cortex and the right insula (Cameron, 2001; Critchley et al., 2004; Pollatos et al., 2007a,c). In a dipole localization study it was demonstrated that HEPs originate from exactly these four brain areas (Pollatos et al., 2005). When focusing on brain regions that are involved in the elicitation of a stress response, specific attention is paid to the ACC and the right insula. Blood perfusion in both areas is increased after a mental arithmetic task in individuals with high self-reported stress (Wang et al., 2005). The replication of these results suggests that these areas are specifically sensitive to psychological stress in women (Wang et al., 2007). The importance of the ACC in mediating stress responses has been demonstrated in numerous studies: functional connectivity in the ACC is related to cortisol release in response to a combined dexamethasone/CRF administration in healthy individuals (Kiem et al., 2013), while ACC connectivity may be reduced in traumatic stress experience (Kennis et al., 2014). Altered insular activity can be observed in early life (Mueller et al., 2010b) and traumatic stress (Bruce et al., 2012). However, these studies may not disentangle whether the ACC and the insula are involved in the up-regulation of physiological stress axes or if their altered activity and connectivity is a result of altered afferent input from visceral organs due to the dysregulation of stress responses.
Brain regions that are sensitive to the effects of stress hormones, and play a role in interoception, involve the thalamus, and other limbic structures, such as the amygdala and hippocampus, which are important for learning and memory (see below). In particular, blood perfusion in the thalamus is reduced within a time frame of 17 min after cortisol infusion, implying a rapid, non-genomic effect of cortisol (Strelzyk et al., 2012). In contrast, when investigating a later time period, in which genomic and non-genomic mechanisms overlap, cortisol affects activity in the amygdala and hippocampus, whereas no effect on the thalamus could be observed (Lovallo et al., 2010). The thalamus is a major relay center for sensory information that processes and integrates intero- and exteroceptive signals. Cortisol may rapidly affect thalamic activity and could eventually favor the processing of interoceptive signals at the cost of others, while this effect disappears over time. Future studies should clarify if thalamic activity may be chronically altered in HPA axis dysregulation, which may play a role in the perception of physical symptoms.
At receptor level, special attention has been paid to the role of α2-adrenoceptors. As repeatedly demonstrated, early life stress (Caldji et al., 1998; Liu et al., 2000) and chronic stress (Flugge et al., 2003) may impair the expression of α2-receptors in the NTS, which plays an important role in the down-regulation of the sympathetic nervous system (Flugge, 1999) and the sensory relaying of visceral-afferent neural signals (Jänig, 2006). α2-adrenoceptors are involved in the adequate processing of visceral-afferent signals from the cardiovascular (Sved et al., 1992) and the gastrointestinal system (Myers et al., 2005). It could be argued, therefore, that the reduced density and functionality of α2-adrenoceptors under chronic stress may reduce the individual’s capacity for the adequate processing of visceral-afferent signals, which may eventually result in the generation of physical symptoms.
In SD patients there is evidence for altered activity in medullary control mechanisms for the sympathetic nervous system (Laederach-Hofmann et al., 2008), the ACC (Klug et al., 2011) and insula (Gundel et al., 2008). PD may be accompanied by reduced sensitivity of α2-adrenoceptors (Bremner et al., 1996), while reduced gray matter in the insula (Uchida et al., 2008) and altered ACC activity (Asami et al., 2008; Shin et al., 2013) could also be observed. Anorexia nervosa is characterized by changes in insular activity (Oberndorfer et al., 2013; Strigo et al., 2013). We propose that alterations in receptor sensitivity, volume, blood- flow or connectivity of the respective brain regions could be a result of stress system dysregulation and represent an important factor in altered interoception in these disorders.
Taken together, acute stress, the release of stress hormones and chronic stress may affect the processing of visceral-afferent neural signals at different brain levels, which are important for interoception, such as the NTS, the thalamus, the ACC and the insula. It remains for future research to elucidate whether certain mental disorders, which are associated with physical symptoms, can be differentiated by specific neurobiological patterns of dysregulated receptor- and brain area functioning.
Psychobiological Mechanisms Involved in Interoception and Stress
While the present review and the proposed model focus on the role of physiological stress responses and their dysregulation for altered interoception and the generation of physical symptoms, stress responses include psychological processes that are constituent factors of the experience of stress. In the following we, therefore, briefly discuss psychological processes that may be important for interoception and stress, without making specific reference to these in the proposed model.
Anxiety and Interoception
Individual stress responses are accompanied by mood changes that include anxiety or fear. There is indeed such an overlap of concepts, methodologies (including experimental paradigms) and physiological correlates of acute stress and anxiety that it seems all but impossible to differentiate between the two constructs (Stevens et al., 2011; Schmitz et al., 2012; Durlik et al., 2014). At least part of the literature discussed in the Section on “Acute Stress and Interoception (Path b)” could, therefore, also be subsumed under the title “Anxiety and Interoception.” To understand similarities and differences between both concepts, their theoretical definitions have to be evaluated. On the one hand, anxiety is a distinct emotion, which is characterized by a limited duration, an antecedent appraisal of a stimulus, and multiple components including specific affective, behavioral, cognitive, and physiological response patterns (Russell, 2003; Ekkekakis, 2013). Anxiety has a state component, i.e., a transitory condition of anxiety, and a trait component, which describes a relative stable disposition to respond to stimuli with state anxiety (Spielberger and Reheiser, 2004). On the other hand, psychological (dis-)stress is characterized as core affect (component of negative affect), which has a long duration, a variable intensity, it does not require either an antecedent appraisal or an eliciting stimulus, and is restricted to the affective component (Russell, 2003; Ekkekakis, 2013). It has to be acknowledged, however, that this definition may be limited to affective response patterns as to be expected in enduring or chronic stress, but it does not include specific physiological response patterns that are implied by chronic exposure to stress. In stress research, the definition of acute stress is commonly based on a neuro-psycho-endocrine perspective. This perspective emphasizes physiological response patterns, as earlier described, but also includes behavioral, cognitive and affective components of stress, such as increased anxiety (McEwen, 2000). In contrast to the core affect of psychological distress, in research on acute stress as induced by established laboratory stressors, an antecedent appraisal of a specific stimulus exists, which may be an additional overlap with the emotional state of anxiety. Important differences between state anxiety and acute stress would be the broader range of distinct emotions that could be elicited during acute stress (e.g., anxiety, fear, anger, etc.) that imply a more diffuse pattern of behavioral and physiological responses, and the longer duration of an acute stress response. Trait anxiety, anxiety sensitivity, and anxiety disorders are all strongly associated with interoception. For example, healthy individuals with high social anxiety show higher IA than those with low social anxiety (Stevens et al., 2011). Furthermore, anxiety sensitivity may moderate the effects of acute stress on interoception (Sturges and Goetsch, 1996). As argued by Domschke et al. (2010), anxiety sensitivity may increase the vulnerability to anxiety disorders by increasing the perceptual basis for catastrophic interpretations of physical symptoms. Although we do not explicate the role of trait anxiety or anxiety sensitivity for interoception in the current model, it may be speculated that either aspect of anxiety could potentially moderate almost all proposed variables (e.g., the probability and intensity of stress experience, physical symptoms) and pathways, e.g., between physiological stress axes and interoception (path c), between interoception and perception of physical symptoms (d) and between symptoms and stress experience (e).
Attention and Interoception
In models on altered interoception in PD (Ehlers and Breuer, 1996; Willem Van der Does et al., 2000) or SD (Rief and Barsky, 2005; Rief and Broadbent, 2007) attention focused on physical symptoms plays an essential role. While there is largely agreement in the literature on attentional bias to body- and health-related sensations in patients with PD and SD, evidence on the factors or processes contributing to attentional bias (e.g., acute and chronic stress) and, therefore, physical symptom generation, is lacking. Regarding the SAM axis, the central noradrenergic system, including the LC, is of particular importance. Early life or chronic stress may decrease the expression of α2-adrenoceptors in the LC (Caldji et al., 1998; Liu et al., 2000), which are involved in the regulation of the ‘alertness’ component of attention. As described above, α2-adrenoceptors are involved in a negative feedback mechanism that down-regulates sympathetic activation and associated processes, such as alertness and vigilance. Consequently, the acute activation of α2-adrenergic receptors increases vigilance and alertness (Berridge and Foote, 1991), while their deactivation results in chronic hypervigilance and alertness (Aston-Jones et al., 1999; Usher et al., 1999). Regarding the HPA axis, the relationship between stress hormones and attentional processes may be more complex, since cortisol may affect all neurons in the entire brain. As summarized by Erickson et al. (2003), glucocorticoids facilitate focused attention at the cost of irrelevant stimuli and alter the perceptual threshold of external stimuli (Fehm-Wolfsdorf and Nagel, 1996). These effects are probably due to increased availability of norepinephrine, as caused by glucocorticoids (Irwin et al., 1986; Kvetnansky et al., 1993). Future research should clarify whether this change in perceptual threshold may also apply to interoceptive signals. Mental stress tests that have been shown to affect both stress axes, also induce increased selective attention on primary tasks at the expense of cognitive flexibility (Chajut and Algom, 2003; Plessow et al., 2011). The differential effects of stress on IA as assessed by the Schandry- and Whitehead-based heartbeat perception tasks (Schulz et al., 2013a) indicate that different facets of attention are of relevance for the impact of stress on interoception. It is plausible, for example, that focused attention on interoceptive signals is facilitated (Schandry task), while the integration of intero- and exteroceptive signals is attenuated (Whitehead task) during stress. With respect to the proposed model, attention may moderate the relationship between physiological stress axes and interoception (path c), between interoception and the perception of physical symptoms (d), and between symptoms and the experience of stress (e).
The Role of Interoceptive Signal Processing in Stress Effects on Memory and Learning
Stress affects memory at acquisition, consolidation and recall. Interoceptive signal processing may contribute to some of these mechanisms (Schulz, 2015). McGaugh (2000) and Roozendaal et al. (2006) postulated the requirement of two physiological processes for the enhancement of memory consolidation after stress: (1) glucocorticoids that bind on glucocorticoid receptors within the NTS, the LC and the basolateral amygdala (BLA) and (2) peripherally circulating epinephrine, which cannot cross the blood-brain barrier, but activates visceral organs, whose afferent signals are transmitted via the vagus nerve to the NTS and LC. From these structures, via noradrenergic pathways, β-adrenergic synapses in the BLA are activated, which induces the release of cyclic adenosine monophosphate (cAMP) and cAMP-dependent protein kinase. Both substances can enhance memory consolidation. Postsynaptic efficacy of β-adrenergic synapses is increased by glucocorticoids. In animal studies it could be shown that both processes glucocorticoid secretion (Quirarte et al., 1997) and afferent signal transmission from visceral organs (Roozendaal et al., 1999) are necessary for improvement of memory consolidation. In humans, the increase of baro-afferent neural feedback induced by norepinephrine-infusion can improve long-term memory (Moor et al., 2005). Interoceptive neural signals, therefore, play an important role for the stress-induced improvement of memory consolidation. It is plausible that increased visceral-afferent input could also facilitate dysfunctional learning processes that contribute to the generation of physical symptoms, such as the formation of pain memory.
Intuitive Decision Making
The influential somatic marker hypothesis by Damasio (1994) postulates that interoceptive signals are regularly integrated into processes of intuitive decision making. It is assumed that in every context, in which intuitive decisions are made, the outcomes of possible action alternatives are anticipated. This anticipation produces a specific visceral response to be integrated in affective responses to the expected outcome. These responses, either transmitted over somatosensory or visceral-afferent circuits, are called ‘somatic markers.’ Dunn et al. (2006) have provided a comprehensive review of the somatic marker hypothesis. In support of this theory IA positively correlates with performance in decision-making tasks (Werner et al., 2009; Wolk et al., 2014). Stress affects executive functions and thus decision making: cortisol may reduce performance in intuitive decision making tasks both in humans and rodents (van den Bos et al., 2009; Koot et al., 2013). This effect was explained by rapid, presumably non-genomic glucocorticoid effects in the orbitofrontal and insular cortex (Koot et al., 2013), which are also involved in interoceptive signal processing. Furthermore, a quadratic relationship exists between peripheral SAM axis activity and performance in decision making tasks, implying a performance increase in moderate sympathetic activation (Pabst et al., 2013). Again, since catecholamines do not cross the blood–brain barrier, afferent neural signals from visceral organs (e.g., transmitted over the vagus nerve) due to increased stimulation of interoceptors, are likely to be involved in this effect. All effects of peripheral sympathetic activation on brain functions, therefore, require neural signal transmission from visceral organs, which can be considered ‘interoceptive’ signals.
Integration and Outlook
In the earlier introduced model of interoception by Vaitl (1996) different levels of interoception were proposed. The model implies that interoception is a bottom–up process, whose effects at a low hierarchical level consecutively affect higher levels. The current review summarizes the state-of-the-art knowledge on the effects of stress (research objective 1), mainly based on research investigating afferent signals from the cardiovascular system. In Figure 2 we integrate the reported findings on interoception and stress into a model, specifically providing links to different levels of interoception, as suggested by Vaitl (1996). In contrast to the existing model, and based on evidence of dissociations between interoceptive indicators for cortical (HEP; Schulz et al., 2013c) and brainstem processes (CMS; Schulz et al., 2010), in the current model we separated ‘CNS representation’ into cortical and sub-cortical processes. Furthermore, we focus on visceral-afferent signals and do not address the possible contribution of somatosensory information to interoception. As some conditions are associated with selective degeneration of afferent autonomic nerves (e.g., diabetes mellitus), the level ‘afferent neurotransmission’ was newly introduced into the model. Although the same mechanisms (e.g., stress axes) are involved in mediating acute stress responses and chronic stress conditions, the complex interplay between stress hormones, binding sites and feedback regulation requires their independent consideration. Acute peripheral sympathetic activation clearly increases stimulation of cardiac interoceptors (Moor et al., 2005). Given the clear physiological relationship between circulating catecholamines and stimulation of cardiac interoceptors (e.g., arterial baroreceptors), we assume that this relationship persists also in chronic elevations of catecholamine output. To our knowledge, selective effects of stress on afferent neurotransmission or sub-cortical CNS representation concerning interoception are unknown, as of to date. A first study by our group suggests that there is no effect of cortisol on the CMS (Schulz et al., 2010), possibly reflecting brainstem processing of visceral-afferent neural signals (Schulz et al., 2009a). On the one hand, acute administration of cortisol affects the cortical representation of visceral-afferent signals (Schulz et al., 2013c) as indicated by larger HEP amplitudes (when eyes are open). On the other hand, basal cortisol levels are negatively associated with HEPs (Schulz et al., 2015b). The additional finding that HEP sensitivity for attention focused on heartbeats is negatively associated with α-amylase may implicate that repeated stimulation of interoceptors could reduce sensitivity of CNS representation at a cortical level. Central sympathetic activation has particularly strong effects on attention and awareness. The overall activation of the central noradrenergic system (via α2-adrengergic antagonists) facilitates arousal and alertness, which may impair the adequate processing of visceral-afferent signals. This relationship may eventually be translated to chronic states of central sympathetic activation. It has repeatedly been demonstrated that chronic stress (Flugge, 1999) and mental disorders associated with physical symptoms are related to the dysregulation of α2-adrenoceptors (Nutt, 1989; Bremner et al., 1996). We thus assume that the chronically inadequate processing of visceral-afferent signals may reverse in increased awareness of physical sensations, which may facilitate the generation of physical symptoms. The collective effect of sympathetic activation and cortisol on learning and memory is conceptualized as ‘proposed’ effect in this model, since it has so far only been demonstrated for declarative memory (Roozendaal et al., 2004; McGaugh, 2006), although it may also play a role in the formation of memory for physical symptoms. With this model, we hope to stimulate future research on the yet unknown relationships between physiological stress systems and levels of interoceptive signal processing.
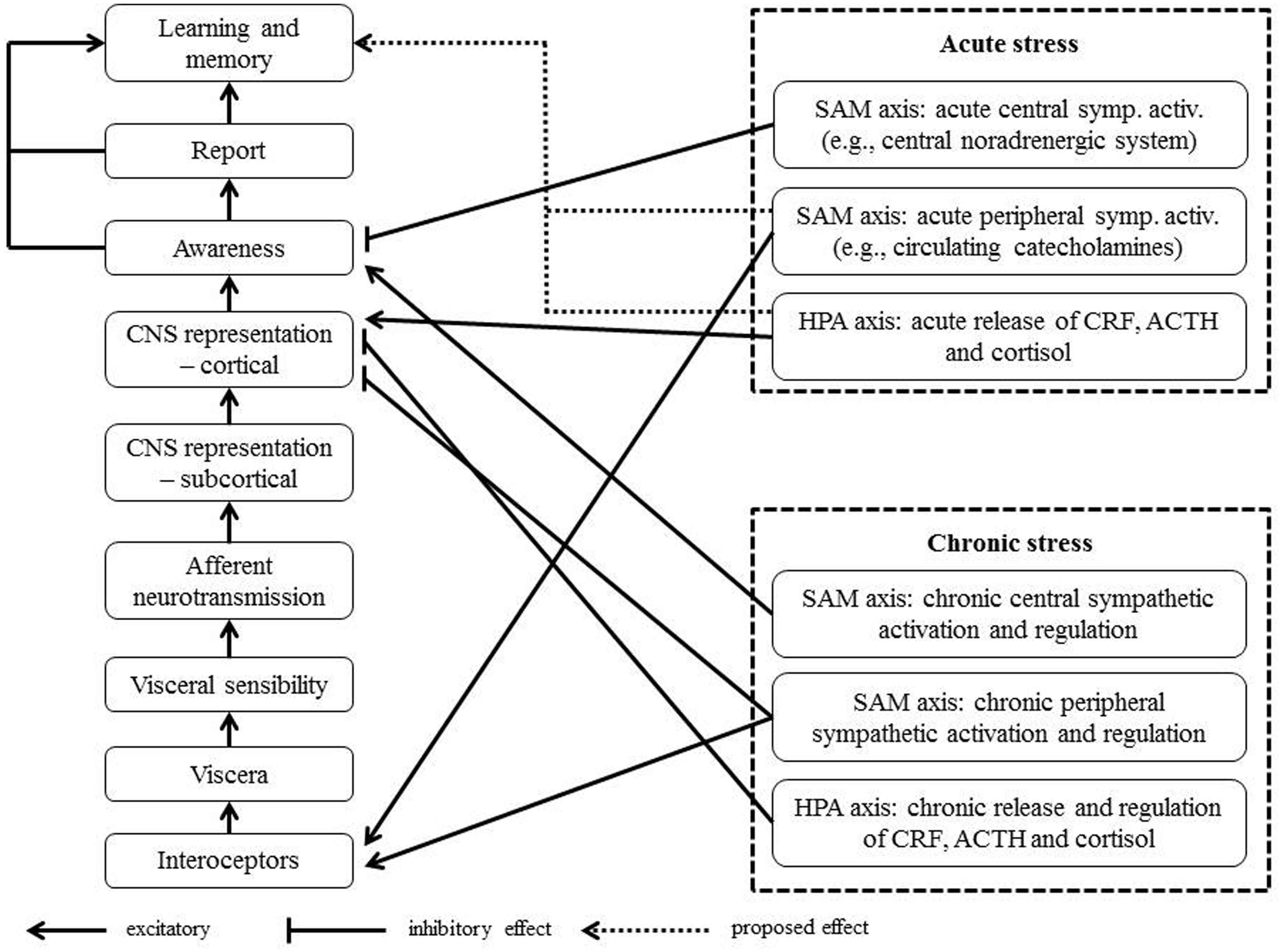
FIGURE 2. Overview of existing findings on interoception and stress based on the model of interoception by Vaitl (1996).
Author Contributions
AS conceived the idea to conduct this review; AS and CV performed literature search; AS and CV interpreted the data and integrated them into the proposed framework models; and AS and CV authored the manuscript.
Conflict of Interest Statement
The Research Office of the University of Luxembourg had no further role in conceptualization of this review, in the collection, analysis and interpretation of data, in the writing of the review, and in the decision to submit the paper for publication. The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work is in parts supported by means of the project “Interoception in medically unexplained symptoms” (INMEDEX) (grant to Dr. André Schulz), funded by the University of Luxembourg.
References
Andor, T., Gerlach, A. L., and Rist, F. (2008). Superior perception of phasic physiological arousal and the detrimental consequences of the conviction to be aroused on worrying and metacognitions in GAD. J. Abnorm. Psychol. 117, 193–205. doi: 10.1037/0021-843X.117.1.193
Antony, M. M., Brown, T. A., Craske, M. G., Barlow, D. H., Mitchell, W. B., and Meadows, E. A. (1995). Accuracy of heartbeat perception in panic disorder, social phobia and nonanxious subjects. J. Anxiety Disord. 5, 355–371. doi: 10.1016/0887-6185(95)00017-I
Arborelius, L., Owens, M. J., Plotsky, P. M., and Nemeroff, C. B. (1999). The role of corticotropin-releasing factor in depression and anxiety disorders. J. Endocrinol. 160, 1–12. doi: 10.1677/joe.0.1600001
Asami, T., Hayano, F., Nakamura, M., Yamasue, H., Uehara, K., Otsuka, T., et al. (2008). Anterior cingulate cortex volume reduction in patients with panic disorder. Psychiatry Clin. Neurosci. 62, 322–330. doi: 10.1111/j.1440-1819.2008.01800.x
Aston-Jones, G., Rajkowski, J., and Cohen, J. (1999). Role of locus coeruleus in attention and behavioral flexibility. Biol. Psychiatry 46, 1309–1320. doi: 10.1016/S0006-3223(99)00140-7
Barsky, A. J. (1992). Amplification, somatization, and the somatoform disorders. Psychosomatics 33, 28–34. doi: 10.1016/S0033-3182(92)72018-0
Barsky, A. J., Brener, J., Coeytaux, R. R., and Cleary, P. D. (1995). Accurate awareness of heartbeat in hypochondriacal and non-hypochondriacal patients. J. Psychosom. Res. 39, 489–497. doi: 10.1016/0022-3999(94)00166-3
Barsky, A. J., Goodson, J. D., Lane, R. S., and Cleary, P. D. (1988). The amplification of somatic symptoms. Psychosom. Med. 50, 510–519. doi: 10.1097/00006842-198809000-00007
Barsky, A. J., Wyshak, G., and Klerman, G. L. (1990). The somatosensory amplification scale and its relationship to hypochondriasis. J. Psychiatr. Res. 24, 323–334. doi: 10.1016/0022-3956(90)90004-A
Bella, G. P., Garcia, M. C., and Spadari-Bratfisch, R. C. (2011). Salivary cortisol, stress, and health in primary caregivers (mothers) of children with cerebral palsy. Psychoneuroendocrinology 36, 834–842. doi: 10.1016/j.psyneuen.2010.11.005
Berridge, C. W., and Foote, S. L. (1991). Effects of locus coeruleus activation on electroencephalographic activity in neocortex and hippocampus. J. Neurosci. 11, 3135–3145.
Blackburn-Munro, G., and Blackburn-Munro, R. E. (2001). Chronic pain, chronic stress and depression: coincidence or consequence? J. Neuroendocrinol. 13, 1009–1023. doi: 10.1046/j.0007-1331.2001.00727.x
Bogaerts, K., Van Eylen, L., Li, W., Bresseleers, J., Van Diest, I., De Peuter, S., et al. (2010). Distorted symptom perception in patients with medically unexplained symptoms. J. Abnorm. Psychol. 119, 226–234. doi: 10.1037/a0017780
Bremner, J. D., Krystal, J. H., Southwick, S. M., and Charney, D. S. (1996). Noradrenergic mechanisms in stress and anxiety: II. Clin. Stud. Synapse 23, 39–51. doi: 10.1002/(SICI)1098-2396(199605)23:1<39::AID-SYN5>3.0.CO;2-I
Brener, J., and Jones, J. M. (1974). Interoceptive discrimination in intact humans: detection of cardiac activity. Physiol. Behav. 13, 763–767. doi: 10.1016/0031-9384(74)90259-5
Brener, J., and Kluvitse, C. (1988). Heartbeat detection: judgments of the simultaneity of external stimuli and heartbeats. Psychophysiology 25, 554–561. doi: 10.1111/j.1469-8986.1988.tb01891.x
Bruce, S. E., Buchholz, K. R., Brown, W. J., Yan, L., Durbin, A., and Sheline, Y. I. (2012). Altered emotional interference processing in the amygdala and insula in women with Post-Traumatic Stress Disorder. Neuroimage Clin. 2, 43–49. doi: 10.1016/j.nicl.2012.11.003
Cacioppo, J. T., Burleson, M. H., Poehlmann, K. M., Malarkey, W. B., Kiecolt-Glaser, J. K., Berntson, G. G., et al. (2000). Autonomic and neuroendocrine responses to mild psychological stressors: effects of chronic stress on older women. Ann. Behav. Med. 22, 140–148. doi: 10.1007/BF02895778
Caldji, C., Tannenbaum, B., Sharma, S., Francis, D., Plotsky, P. M., and Meaney, M. J. (1998). Maternal care during infancy regulates the development of neural systems mediating the expression of fearfulness in the rat. Proc. Natl. Acad. Sci. U.S.A. 95, 5335–5340. doi: 10.1073/pnas.95.9.5335
Cameron, O. G. (2001). Interoception: the inside story–a model for psychosomatic processes. Psychosom. Med. 63, 697–710. doi: 10.1097/00006842-200109000-00001
Ceunen, E., Van Diest, I., and Vlaeyen, J. W. (2013). Accuracy and awareness of perception: related, yet distinct (commentary on Herbert et al., 2012). Biol. Psychol. 92, 426–427. doi: 10.1016/j.biopsycho.2012.09.012
Chajut, E., and Algom, D. (2003). Selective attention improves under stress: implications for theories of social cognition. J. Pers. Soc. Psychol. 85, 231–248. doi: 10.1037/0022-3514.85.2.231
Chan, P. Y., and Davenport, P. W. (2010). Respiratory related evoked potential measures of cerebral cortical respiratory information processing. Biol. Psychol. 84, 4–12. doi: 10.1016/j.biopsycho.2010.02.009
Chandola, T., Heraclides, A., and Kumari, M. (2010). Psychophysiological biomarkers of workplace stressors. Neurosci. Biobehav. Rev. 35, 51–57. doi: 10.1016/j.neubiorev.2009.11.005
Chang, L., Sundaresh, S., Elliott, J., Anton, P. A., Baldi, P., Licudine, A., et al. (2009). Dysregulation of the hypothalamic-pituitary-adrenal (HPA) axis in irritable bowel syndrome. Neurogastroenterol. Motil. 21, 149–159. doi: 10.1111/j.1365-2982.2008.01171.x
Chatterton, R. T. Jr., Vogelsong, K. M., Lu, Y. C., Ellman, A. B., and Hudgens, G. A. (1996). Salivary alpha-amylase as a measure of endogenous adrenergic activity. Clin. Physiol. 16, 433–448. doi: 10.1111/j.1475-097X.1996.tb00731.x
Chrousos, G. P. (2009). Stress and disorders of the stress system. Nat. Rev. Endocrinol. 5, 374–381. doi: 10.1038/nrendo.2009.106
Chrousos, G. P., and Gold, P. W. (1992). The concepts of stress and stress system disorders. Overview of physical and behavioral homeostasis. Jama 267, 1244–1252. doi: 10.1001/jama.1992.03480090092034
Cleare, A. J. (2004). The HPA axis and the genesis of chronic fatigue syndrome. Trends Endocrinol. Metab. 15, 55–59. doi: 10.1016/j.tem.2003.12.002
Cobos, P., Sánchez, M., Pérez, N., and Vila, J. (2004). Effects of spinal cord injuries on the subjective component of emotions. Cogn. Emot. 18, 281–287. doi: 10.1080/02699930244000471
Cohen, S., and Hamrick, N. (2003). Stable individual differences in physiological response to stressors: implications for stress-elicited changes in immune related health. Brain Behav. Immun. 17, 407–414. doi: 10.1016/S0889-1591(03)00110-7
Craig, A. D. (2002). How do you feel? Interoception: the sense of the physiological condition of the body. Nat. Rev. Neurosci. 3, 655–666. doi: 10.1038/nrn894
Craig, A. D. (2009). How do you feel–now? The anterior insula and human awareness. Nat. Rev. Neurosci. 10, 59–70. doi: 10.1038/nrn2555
Critchley, H. D., Wiens, S., Rotshtein, P., Ohman, A., and Dolan, R. J. (2004). Neural systems supporting interoceptive awareness. Nat. Neurosci. 7, 189–195. doi: 10.1038/nn1176
Damasio, A. (1994). Descartes’ Error: Emotion, Reason and the Human Brain. New York: Grosset/Putnam.
Davenport, P. W., Friedman, W. A., Thompson, F. J., and Franzen, O. (1986). Respiratory-related cortical potentials evoked by inspiratory occlusion in humans. J. Appl. Physiol. 60, 1843–1848.
De Andres-Garcia, S., Moya-Albiol, L., and Gonzalez-Bono, E. (2012). Salivary cortisol and immunoglobulin A: responses to stress as predictors of health complaints reported by caregivers of offspring with autistic spectrum disorder. Horm. Behav. 62, 464–474. doi: 10.1016/j.yhbeh.2012.08.003
de Kloet, E. R., Karst, H., and Joels, M. (2008). Corticosteroid hormones in the central stress response: quick-and-slow. Front. Neuroendocrinol. 29:268. doi: 10.1016/j.yfrne.2007.10.002
De Kloet, E. R., Vreugdenhil, E., Oitzl, M. S., and Joels, M. (1998). Brain corticosteroid receptor balance in health and disease. Endocr. Rev. 19, 269–301. doi: 10.1210/er.19.3.269
de Vugt, M. E., Nicolson, N. A., Aalten, P., Lousberg, R., Jolle, J., and Verhey, F. R. (2005). Behavioral problems in dementia patients and salivary cortisol patterns in caregivers. J. Neuropsychiatry Clin. Neurosci. 17, 201–207. doi: 10.1176/jnp.17.2.201
Domschke, K., Stevens, S., Pfleiderer, B., and Gerlach, A. L. (2010). Interoceptive sensitivity in anxiety and anxiety disorders: an overview and integration of neurobiological findings. Clin. Psychol. Rev. 30, 1–11. doi: 10.1016/j.cpr.2009.08.008
Dunn, B. D., Dalgleish, T., and Lawrence, A. D. (2006). The somatic marker hypothesis: a critical evaluation. Neurosci. Biobehav. Rev. 30, 239–271. doi: 10.1016/j.neubiorev.2005.07.001
Dunn, B. D., Dalgleish, T., Ogilvie, A. D., and Lawrence, A. D. (2007). Heartbeat perception in depression. Behav. Res. Ther. 45, 1921–1930. doi: 10.1016/j.brat.2006.09.008
Durlik, C., Brown, G., and Tsakiris, M. (2014). Enhanced interoceptive awareness during anticipation of public speaking is associated with fear of negative evaluation. Cogn. Emot. 28, 530–540. doi: 10.1080/02699931.2013.832654
Ehlers, A. (1995). A 1-year prospective study of panic attacks: clinical course and factors associated with maintenance. J. Abnorm. Psychol. 104, 164–172. doi: 10.1037/0021-843X.104.1.164
Ehlers, A., and Breuer, P. (1992). Increased cardiac awareness in panic disorder. J. Abnorm. Psychol. 101, 371–382. doi: 10.1037/0021-843X.101.3.371
Ehlers, A., and Breuer, P. (1996). How good are patients with panic disorder at perceiving their heartbeats? Biol. Psychol. 42, 165–182. doi: 10.1016/0301-0511(95)05153-8
Ehlers, A., Breuer, P., Dohn, D., and Fiegenbaum, W. (1995). Heartbeat perception and panic disorder: possible explanations for discrepant findings. Behav. Res. Ther. 33, 69–76. doi: 10.1016/0005-7967(94)E0002-Z
Ehlert, U., Gaab, J., and Heinrichs, M. (2001). Psychoneuroendocrinological contributions to the etiology of depression, posttraumatic stress disorder, and stress-related bodily disorders: the role of the hypothalamus-pituitary-adrenal axis. Biol. Psychol. 57, 141–152. doi: 10.1016/S0301-0511(01)00092-8
Eichler, S., and Katkin, E. S. (1994). The relationship between cardiovascular reactivity and heartbeat detection. Psychophysiology 31, 229–234. doi: 10.1111/j.1469-8986.1994.tb02211.x
Ekkekakis, P. (2013). The Measurement of Affect, Mood, and Emotion. Cambridge: Cambridge University Press. doi: 10.1017/CBO9780511820724
Elsenbruch, S., Rosenberger, C., Bingel, U., Forsting, M., Schedlowski, M., and Gizewski, E. R. (2010a). Patients with irritable bowel syndrome have altered emotional modulation of neural responses to visceral stimuli. Gastroenterology 139, 1310–1319. doi: 10.1053/j.gastro.2010.06.054
Elsenbruch, S., Rosenberger, C., Enck, P., Forsting, M., Schedlowski, M., and Gizewski, E. R. (2010b). Affective disturbances modulate the neural processing of visceral pain stimuli in irritable bowel syndrome: an fMRI study. Gut 59, 489–495. doi: 10.1136/gut.2008.175000
Erickson, K., Drevets, W., and Schulkin, J. (2003). Glucocorticoid regulation of diverse cognitive functions in normal and pathological emotional states. Neurosci. Biobehav. Rev. 27, 233–246. doi: 10.1016/S0149-7634(03)00033-2
Fahrenberg, J., and Foerster, F. (1982). Covariation and consistency of activation parameters. Biol. Psychol. 15, 151–169. doi: 10.1016/0301-0511(82)90039-4
Fairclough, S. H., and Goodwin, L. (2007). The effect of psychological stress and relaxation on interoceptive accuracy: implications for symptom perception. J. Psychosom. Res. 62, 289–295. doi: 10.1016/j.jpsychores.2006.10.017
Farb, N. A., Daubenmier, J. J., Price, C. J., Gard, T., Kerr, C. E., Dunn, B. D., et al. (2015).Interoception, contemplative practice, and health. Front. Psychol. 6:763 doi: 10.3389/fpsyg.2015.00763
Fehm-Wolfsdorf, G., and Nagel, D. (1996). Differential effects of glucocorticoids on human auditory perception. Biol. Psychol. 42, 117–130. doi: 10.1016/0301-0511(95)05150-3
Flugge, G. (1999). Effects of cortisol on brain alpha2-adrenoceptors: potential role in stress. Neurosci. Biobehav. Rev. 23, 949–956. doi: 10.1016/S0149-7634(99)00028-7
Flugge, G., Van Kampen, M., Meyer, H., and Fuchs, E. (2003). Alpha2A and alpha2C-adrenoceptor regulation in the brain: alpha2A changes persist after chronic stress. Eur. J. Neurosci. 17, 917–928. doi: 10.1046/j.1460-9568.2003.02510.x
Frattola, A., Parati, G., Gamba, P., Paleari, F., Mauri, G., Di Rienzo, M., et al. (1997). Time and frequency domain estimates of spontaneous baroreflex sensitivity provide early detection of autonomic dysfunction in diabetes mellitus. Diabetologia 40, 1470–1475. doi: 10.1007/s001250050851
Fries, E., Hesse, J., Hellhammer, J., and Hellhammer, D. H. (2005). A new view on hypocortisolism. Psychoneuroendocrinology 30, 1010–1016. doi: 10.1016/j.psyneuen.2005.04.006
Garfinkel, S. N., and Critchley, H. D. (2013). Interoception, emotion and brain: new insights link internal physiology to social behaviour. Commentary on:: “Anterior insular cortex mediates bodily sensibility and social anxiety” by Terasawa et al. (2012). Soc. Cogn. Affect. Neurosci. 8, 231–234. doi: 10.1093/scan/nss140
Garfinkel, S. N., Seth, A. K., Barrett, A. B., Suzuki, K., and Critchley, H. D. (2015). Knowing your own heart: distinguishing interoceptive accuracy frominteroceptive awareness. Biol. Psychol. 104, 65–74. doi: 10.1016/j.biopsycho.2014.11.004
Garner, D. M. (1991). Eating Disorder Inventory-2 Professional Manual. Odessa: Psychological Resources.
Gold, P. W. (2015). The organization of the stress system and its dysregulation in depressive illness. Mol. Psychiatry 20, 32–47. doi: 10.1038/mp.2014.163
Gold, P. W., and Chrousos, G. P. (2002). Organization of the stress system and its dysregulation in melancholic and atypical depression: high vs low CRH/NE states. Mol. Psychiatry 7, 254–275. doi: 10.1038/sj.mp.4001032
Gold, P. W., Goodwin, F. K., and Chrousos, G. P. (1988). Clinical and biochemical manifestations of depression. Relation to the neurobiology of stress (2). New Engl. J. Med. 319, 413–420. doi: 10.1056/NEJM198808183190706
Gold, P. W., Licinio, J., Wong, M. L., and Chrousos, G. P. (1995). Corticotropin releasing hormone in the pathophysiology of melancholic and atypical depression and in the mechanism of action of antidepressant drugs. Ann. N. Y. Acad. Sci. 771, 716–729. doi: 10.1111/j.1749-6632.1995.tb44723.x
Gray, M. A., Taggart, P., Sutton, P. M., Groves, D., Holdright, D. R., Bradbury, D., et al. (2007). A cortical potential reflecting cardiac function. Proc. Natl. Acad. Sci. U.S.A. 104, 6818–6823. doi: 10.1073/pnas.0609509104
Groeneweg, F. L., Karst, H., De Kloet, E. R., and Joels, M. (2011). Rapid non-genomic effects of corticosteroids and their role in the central stress response. J. Endocrinol. 209, 153–167. doi: 10.1530/JOE-10-0472
Gundel, H., Valet, M., Sorg, C., Huber, D., Zimmer, C., Sprenger, T., et al. (2008). Altered cerebral response to noxious heat stimulation in patients with somatoform pain disorder. Pain 137, 413–421. doi: 10.1016/j.pain.2007.10.003
Gupta, A., Kilpatrick, L., Labus, J., Tillisch, K., Braun, A., Hong, J. Y., et al. (2014). Early adverse life events and resting state neural networks in patients with chronic abdominal pain: evidence for sex differences. Psychosom. Med. 76, 404–412. doi: 10.1097/PSY.0000000000000089
Hart, N., Mcgowan, J., Minati, L., and Critchley, H. D. (2013). Emotional regulation and bodily sensation: interoceptive awareness is intact in borderline personality disorder. J. Personal. Disord. 27, 506–518. doi: 10.1521/pedi_2012_26_049
Heim, C., Ehlert, U., and Hellhammer, D. H. (2000). The potential role of hypocortisolism in the pathophysiology of stress-related bodily disorders. Psychoneuroendocrinology 25, 1–35. doi: 10.1016/S0306-4530(99)00035-9
Heims, H. C., Critchley, H. D., Dolan, R., Mathias, C. J., and Cipolotti, L. (2004). Social and motivational functioning is not critically dependent on feedback of autonomic responses: neuropsychological evidence from patients with pure autonomic failure. Neuropsychologia 42, 1979–1988. doi: 10.1016/j.neuropsychologia.2004.06.001
Herbert, B. M., Blechert, J., Hautzinger, M., Matthias, E., and Herbert, C. (2013). Intuitive eating is associated with interoceptive sensitivity. Effects on body mass index. Appetite 70C, 22–30. doi: 10.1016/j.appet.2013.06.082
Herbert, B. M., Herbert, C., Pollatos, O., Weimer, K., Enck, P., Sauer, H., et al. (2012). Effects of short-term food deprivation on interoceptive awareness, feelings and autonomic cardiac activity. Biol. Psychol. 89, 71–79. doi: 10.1016/j.biopsycho.2011.09.004
Herbert, B. M., and Pollatos, O. (2014). Attenuated interoceptive sensitivity in overweight and obese individuals. Eat. Behav. 15, 445–448. doi: 10.1016/j.eatbeh.2014.06.002
Herbert, B. M., Pollatos, O., Flor, H., Enck, P., and Schandry, R. (2010). Cardiac awareness and autonomic cardiac reactivity during emotional picture viewing and mental stress. Psychophysiology 47, 342–354. doi: 10.1111/j.1469-8986.2009.00931.x
Holsboer, F. (2000). The corticosteroid receptor hypothesis of depression. Neuropsychopharmacology 23, 477–501. doi: 10.1016/S0893-133X(00)00159-7
Irwin, J., Ahluwalia, P., Zacharko, R. M., and Anisman, H. (1986). Central norepinephrine and plasma corticosterone following acute and chronic stressors: influence of social isolation and handling. Pharmacol. Biochem. Behav. 24, 1151–1154. doi: 10.1016/0091-3057(86)90471-5
Isaac, L. (1980). Clonidine in the central nervous system: site and mechanism of hypotensive action. J. Cardiovasc. Pharmacol. 2(Suppl. 1), S5–S19. doi: 10.1097/00005344-198000021-00002
Ising, M., Hohne, N., Siebertz, A., Parchmann, A. M., Erhardt, A., and Keck, M. (2012). Stress response regulation in panic disorder. Curr. Pharm. Des. 18, 5675–5684. doi: 10.2174/138161212803530880
Jänig, W. (2006). The Integrative Action of the Autonomic Nervous System. New York: Cambridge University Press. doi: 10.1017/CBO9780511541667
Janssens, K. A., Oldehinkel, A. J., Verhulst, F. C., Hunfeld, J. A., Ormel, J., and Rosmalen, J. G. (2012). Symptom-specific associations between low cortisol responses and functional somatic symptoms: the TRAILS study. Psychoneuroendocrinology 37, 332–340. doi: 10.1016/j.psyneuen.2011.06.016
Jones, G. E., and Hollandsworth, J. G. (1981). Heart rate discrimination before and after exercise-induced augmented cardiac activity. Psychophysiology 18, 252–257. doi: 10.1111/j.1469-8986.1981.tb03029.x
Katkin, E. S., Blascovich, J., and Goldband, S. (1981). Empirical assessment of visceral self-perception: individual and sex differences in the acquisition of heartbeat discrimination. J. Pers. Soc. Psychol. 40, 1095–1101. doi: 10.1037/0022-3514.40.6.1095
Kennis, M., Rademaker, A. R., Van Rooij, S. J., Kahn, R. S., and Geuze, E. (2014). Resting state functional connectivity of the anterior cingulate cortex in veterans with and without post-traumatic stress disorder. Hum. Brain Mapp. 36, 99–109. doi: 10.1002/hbm.22615
Khalsa, S. S., Rudrauf, D., Damasio, A. R., Davidson, R. J., Lutz, A., and Tranel, D. (2008). Interoceptive awareness in experienced meditators. Psychophysiology 45, 671–677. doi: 10.1111/j.1469-8986.2008.00666.x
Kiem, S. A., Andrade, K. C., Spoormaker, V. I., Holsboer, F., Czisch, M., and Samann, P. G. (2013). Resting state functional MRI connectivity predicts hypothalamus-pituitary-axis status in healthy males. Psychoneuroendocrinology 38, 1338–1348. doi: 10.1016/j.psyneuen.2012.11.021
Kindermann, N. K., and Werner, N. S. (2014). Cardiac perception enhances stress experience. J. Psychophysiol. 28, 225–232. doi: 10.1027/0269-8803/a000114
Kirschbaum, C., Pirke, K. M., and Hellhammer, D. H. (1993). The ‘trier social stress test’–a tool for investigating psychobiological stress responses in a laboratory setting. Neuropsychobiology 28, 76–81. doi: 10.1159/000119004
Klug, S., Anderer, P., Saletu-Zyhlarz, G., Freidl, M., Saletu, B., Prause, W., et al. (2011). Dysfunctional pain modulation in somatoform pain disorder patients. Eur. Arch. Psychiatry Clin. Neurosci. 261, 267–275. doi: 10.1007/s00406-010-0148-4
Knack, J. M., Jensen-Campbell, L. A., and Baum, A. (2011). Worse than sticks and stones? Bullying is associated with altered HPA axis functioning and poorer health. Brain Cogn. 77, 183–190. doi: 10.1016/j.bandc.2011.06.011
Knoll, J. F., and Hodapp, V. (1992). A comparison between two methods for assessing heartbeat perception. Psychophysiology 29, 218–222. doi: 10.1111/j.1469-8986.1992.tb01689.x
Koot, S., Baars, A., Hesseling, P., Van Den Bos, R., and Joels, M. (2013). Time-dependent effects of corticosterone on reward-based decision-making in a rodent model of the Iowa Gambling Task. Neuropharmacology 70, 306–315. doi: 10.1016/j.neuropharm.2013.02.008
Kudielka, B. M., Schommer, N. C., Hellhammer, D. H., and Kirschbaum, C. (2004). Acute HPA axis responses, heart rate, and mood changes to psychosocial stress (TSST) in humans at different times of day. Psychoneuroendocrinology 29, 983–992. doi: 10.1016/j.psyneuen.2003.08.009
Kuehl, L. K., Michaux, G. P., Richter, S., Schachinger, H., and Anton, F. (2010). Increased basal mechanical pain sensitivity but decreased perceptual wind-up in a human model of relative hypocortisolism. Pain 149, 539–546. doi: 10.1016/j.pain.2010.03.026
Kvetnansky, R., Fukuhara, K., Pacak, K., Cizza, G., Goldstein, D. S., and Kopin, I. J. (1993). Endogenous glucocorticoids restrain catecholamine synthesis and release at rest and during immobilization stress in rats. Endocrinology 133, 1411–1419. doi: 10.1210/en.133.3.1411
Laederach-Hofmann, K., Ruddel, H., and Mussgay, L. (2008). Pathological baroreceptor sensitivity in patients suffering from somatization disorders: do they correlate with symptoms? Biol. Psychol. 79, 243–249. doi: 10.1016/j.biopsycho.2008.06.003
Larra, M. F., Schulz, A., Schilling, T. M., Ferreira De Sa, D. S., Best, D., Kozik, B., et al. (2014). Heart rate response to post-learning stress predicts memory consolidation. Neurobiol. Learn. Mem. 109C, 74–81. doi: 10.1016/j.nlm.2013.12.004
Lass-Hennemann, J., Deuter, C. E., Kuehl, L. K., Schulz, A., Blumenthal, T. D., and Schachinger, H. (2010). Effects of stress on human mating preferences: stressed individuals prefer dissimilar mates. Proc. Biol. Sci. 277, 2175–2183. doi: 10.1098/rspb.2010.0258
Lass-Hennemann, J., Kuehl, L. K., Schulz, A., Oitzl, M. S., and Schachinger, H. (2011). Stress strengthens memory of first impressions of others’ positive personality traits. PLoS ONE 6:e16389. doi: 10.1371/journal.pone.0016389
Leopold, C., and Schandry, R. (2001). The heartbeat-evoked brain potential in patients suffering from diabetic neuropathy and in healthy control persons. Clin. Neurophysiol. 112, 674–682. doi: 10.1016/S1388-2457(01)00480-1
Lesch, K. P., Widerlov, E., Ekman, R., Laux, G., Schulte, H. M., Pfuller, H., et al. (1988). Delta sleep-inducing peptide response to human corticotropin-releasing hormone (CRH) in major depressive disorder. Comparison with CRH-induced corticotropin and cortisol secretion. Biol. Psychiatry 24, 162–172. doi: 10.1016/0006-3223(88)90271-5
Levitan, R. D., Vaccarino, F. J., Brown, G. M., and Kennedy, S. H. (2002). Low-dose dexamethasone challenge in women with atypical major depression: pilot study. J. Psychiatry Neurosci. 27, 47–51.
Liu, D., Caldji, C., Sharma, S., Plotsky, P. M., and Meaney, M. J. (2000). Influence of neonatal rearing conditions on stress-induced adrenocorticotropin responses and norepinepherine release in the hypothalamic paraventricular nucleus. J. Neuroendocrinol. 12, 5–12. doi: 10.1046/j.1365-2826.2000.00422.x
Lovallo, W. R., Robinson, J. L., Glahn, D. C., and Fox, P. T. (2010). Acute effects of hydrocortisone on the human brain: an fMRI study. Psychoneuroendocrinology 35, 15–20. doi: 10.1016/j.psyneuen.2009.09.010
Marshall, R. D., Blanco, C., Printz, D., Liebowitz, M. R., Klein, D. F., and Coplan, J. (2002). A pilot study of noradrenergic and HPA axis functioning in PTSD vs. panic disorder. Psychiatry Res. 110, 219–230. doi: 10.1016/S0165-1781(02)00126-9
McCraty, R., Atkinson, M., Tomasino, D., and Stuppy, W. P. (2001). Analysis of twenty-four hour heart rate variability in patients with panic disorder. Biol. Psychol. 56, 131–150. doi: 10.1016/S0301-0511(01)00074-6
McEwen, B. S. (2000). The neurobiology of stress: from serendipity to clinical relevance. Brain Res. 886, 172–189. doi: 10.1016/S0006-8993(00)02950-4
McEwen, B. S. (2004). Protection and damage from acute and chronic stress: allostasis and allostatic overload and relevance to the pathophysiology of psychiatric disorders. Ann. N. Y. Acad. Sci. 1032, 1–7. doi: 10.1196/annals.1314.001
McGaugh, J. L. (2000). Memory–a century of consolidation. Science 287, 248–251. doi: 10.1126/science.287.5451.248
McGaugh, J. L. (2006). Make mild moments memorable: add a little arousal. Trends Cogn. Sci. 10, 345–347. doi: 10.1016/j.tics.2006.06.001
McLaughlin, K. A., Sheridan, M. A., Alves, S., and Mendes, W. B. (2014). Child maltreatment and autonomic nervous system reactivity: identifying dysregulated stress reactivity patterns by using the biopsychosocial model of challenge and threat. Psychosom. Med. 76, 538–546. doi: 10.1097/PSY.0000000000000098
Mehling, W. E., Price, C., Daubenmier, J. J., Acree, M., Bartmess, E., and Stewart, A. (2012). The Multidimensional assessment of interoceptive awareness (MAIA). PLoS ONE 7:e48230. doi: 10.1371/journal.pone.0048230
Michal, M., Reuchlein, B., Adler, J., Reiner, I., Beutel, M. E., Vogele, C., et al. (2014). Striking discrepancy of anomalous body experiences with normal interoceptive accuracy in depersonalization-derealization disorder. PLoS ONE 9:e89823. doi: 10.1371/journal.pone.0089823
Michaux, G. P., Magerl, W., Anton, F., and Treede, R. D. (2012). Experimental characterization of the effects of acute stresslike doses of hydrocortisone in human neurogenic hyperalgesia models. Pain 153, 420–428. doi: 10.1016/j.pain.2011.10.043
Moor, T., Mundorff, L., Bohringer, A., Philippsen, C., Langewitz, W., Reino, S. T., et al. (2005). Evidence that baroreflex feedback influences long-term incidental visual memory in men. Neurobiol. Learn. Mem. 84, 168–174. doi: 10.1016/j.nlm.2005.07.003
Moriguchi, A., Otsuka, A., Kohara, K., Mikami, H., Katahira, K., Tsunetoshi, T., et al. (1992). Spectral change in heart rate variability in response to mental arithmetic before and after the beta-adrenoceptor blocker, carteolol. Clin. Auton. Res. 2, 267–270. doi: 10.1007/BF01819547
Mravec, B. (2011). Role of catecholamine-induced activation of vagal afferent pathways in regulation of sympathoadrenal system activity: negative feedback loop of stress response. Endocr. Regul. 45, 37–41.
Mueller, E. M., Evers, E. A., Wacker, J., and Van Der Veen, F. (2012). Acute tryptophan depletion attenuates brain-heart coupling following external feedback. Front. Hum. Neurosci. 6:77. doi: 10.3389/fnhum.2012.00077
Mueller, E. M., Panitz, C., Nestoriuc, Y., Stemmler, G., and Wacker, J. (2014). Panic disorder and serotonin reuptake inhibitors predict coupling of cortical and cardiac activity. Neuropsychopharmacology 39, 507–514. doi: 10.1038/npp.2013.224
Mueller, E. M., Stemmler, G., Hennig, J., and Wacker, J. (2013). 5-HTTLPR and anxiety modulate brain-heart covariation. Psychophysiology 50, 441–453. doi: 10.1111/psyp.12016
Mueller, E. M., Stemmler, G., and Wacker, J. (2010a). Single-trial electroencephalogram predicts cardiac acceleration: a time-lagged P-correlation approach for studying neurovisceral connectivity. Neuroscience 166, 491–500. doi: 10.1016/j.neuroscience.2009.12.051
Mueller, S. C., Maheu, F. S., Dozier, M., Peloso, E., Mandell, D., Leibenluft, E., et al. (2010b). Early-life stress is associated with impairment in cognitive control in adolescence: an fMRI study. Neuropsychologia 48, 3037–3044. doi: 10.1016/j.neuropsychologia.2010.06.013
Mussgay, L., Klinkenberg, N., and Ruddel, H. (1999). Heart beat perception in patients with depressive, somatoform, and personality disorders. J. Psychophysiol. 13, 27–36. doi: 10.1027//0269-8803.13.1.27
Myers, E. A., Banihashemi, L., and Rinaman, L. (2005). The anxiogenic drug yohimbine activates central viscerosensory circuits in rats. J. Comp. Neurol. 492, 426–441. doi: 10.1002/cne.20727
Nater, U. M., Maloney, E., Boneva, R. S., Gurbaxani, B. M., Lin, J. M., Jones, J. F., et al. (2008). Attenuated morning salivary cortisol concentrations in a population-based study of persons with chronic fatigue syndrome and well controls. J. Clin. Endocrinol. Metab. 93, 703–709. doi: 10.1210/jc.2007-1747
Nater, U. M., Rohleder, N., Gaab, J., Berger, S., Jud, A., Kirschbaum, C., et al. (2005). Human salivary alpha-amylase reactivity in a psychosocial stress paradigm. Int. J. Psychophysiol. 55, 333–342. doi: 10.1016/j.ijpsycho.2004.09.009
Nemeroff, C. B., Widerlov, E., Bissette, G., Walleus, H., Karlsson, I., Eklund, K., et al. (1984). Elevated concentrations of CSF corticotropin-releasing factor-like immunoreactivity in depressed patients. Science 226, 1342–1344. doi: 10.1126/science.6334362
Nutt, D. J. (1989). Altered central alpha 2-adrenoceptor sensitivity in panic disorder. Arch. Gen. Psychiatry 46, 165–169. doi: 10.1001/archpsyc.1989.01810020067011
O’Keane, V., Dinan, T. G., Scott, L., and Corcoran, C. (2005). Changes in hypothalamic-pituitary-adrenal axis measures after vagus nerve stimulation therapy in chronic depression. Biol. Psychiatry 58, 963–968. doi: 10.1016/j.biopsych.2005.04.049
O’Keane, V., Frodl, T., and Dinan, T. G. (2012). A review of atypical depression in relation to the course of depression and changes in HPA axis organization. Psychoneuroendocrinology 37, 1589–1599. doi: 10.1016/j.psyneuen.2012.03.009
Oberndorfer, T., Simmons, A., Mccurdy, D., Strigo, I., Matthews, S., Yang, T., et al. (2013). Greater anterior insula activation during anticipation of food images in women recovered from anorexia nervosa versus controls. Psychiatry Res. 214, 132–141. doi: 10.1016/j.pscychresns.2013.06.010
Pabst, S., Brand, M., and Wolf, O. T. (2013). Stress and decision making: a few minutes make all the difference. Behav. Brain Res. 250, 39–45. doi: 10.1016/j.bbr.2013.04.046
Pappens, M., Van Den Bergh, O., Vansteenwegen, D., Ceunen, E., De Peuter, S., and Van Diest, I. (2013). Learning to fear obstructed breathing: comparing interoceptive and exteroceptive cues. Biol. Psychol. 92, 36–42. doi: 10.1016/j.biopsycho.2011.05.004
Pariante, C. M., and Lightman, S. L. (2008). The HPA axis in major depression: classical theories and new developments. Trends Neurosci. 31, 464–468. doi: 10.1016/j.tins.2008.06.006
Pauli, P., Hartl, L., Marquardt, C., Stalmann, H., and Strian, F. (1991). Heartbeat and arrhythmia perception in diabetic autonomic neuropathy. Psychol. Med. 21, 413–421. doi: 10.1017/S0033291700020523
Pennebaker, J. W., and Lightner, J. M. (1980). Competition of internal and external information in an exercise setting. J. Pers. Soc. Psychol. 39, 165–174. doi: 10.1037/0022-3514.39.1.165
Petersen, S., Schroijen, M., Molders, C., Zenker, S., and Van Den Bergh, O. (2014). Categorical Interoception: perceptual organization of sensations from inside. Psychol. Sci. 25, 1059–1066. doi: 10.1177/0956797613519110
Petrowski, K., Herold, U., Joraschky, P., Wittchen, H. U., and Kirschbaum, C. (2010). A striking pattern of cortisol non-responsiveness to psychosocial stress in patients with panic disorder with concurrent normal cortisol awakening responses. Psychoneuroendocrinology 35, 414–421. doi: 10.1016/j.psyneuen.2009.08.003
Petrowski, K., Wintermann, G. B., Schaarschmidt, M., Bornstein, S. R., and Kirschbaum, C. (2013). Blunted salivary and plasma cortisol response in patients with panic disorder under psychosocial stress. Int. J. Psychophysiol. 88, 35–39. doi: 10.1016/j.ijpsycho.2013.01.002
Philippsen, C., Hahn, M., Schwabe, L., Richter, S., Drewe, J., and Schachinger, H. (2007). Cardiovascular reactivity to mental stress is not affected by alpha2-adrenoreceptor activation or inhibition. Psychopharmacology 190, 181–188. doi: 10.1007/s00213-006-0597-7
Phillips, G. C., Jones, G. E., Rieger, E. J., and Snell, J. B. (1999). Effects of the presentation of false heart-rate feedback on the performance of two common heartbeat-detection tasks. Psychophysiology 36, 504–510. doi: 10.1017/S0048577299980071
Plessow, F., Fischer, R., Kirschbaum, C., and Goschke, T. (2011). Inflexibly focused under stress: acute psychosocial stress increases shielding of action goals at the expense of reduced cognitive flexibility with increasing time lag to the stressor. J. Cogn. Neurosci. 23, 3218–3227. doi: 10.1162/jocn_a_00024
Pohl, J., Frenzel, G., Kerner, W., and Fehm-Wolfsdorf, G. (1998). Acute stress modulates symptom awareness and hormonal counterregulation during insulin-induced hypoglycemia in healthy individuals. Int. J. Behav. Med. 5, 89–105. doi: 10.1207/s15327558ijbm0502_1
Pollatos, O., Gramann, K., and Schandry, R. (2007a). Neural systems connecting interoceptive awareness and feelings. Hum. Brain Mapp. 28, 9–18. doi: 10.1002/hbm.20258
Pollatos, O., Herbert, B. M., Kaufmann, C., Auer, D. P., and Schandry, R. (2007b). Interoceptive awareness, anxiety and cardiovascular reactivity to isometric exercise. Int. J. Psychophysiol. 65, 167–173. doi: 10.1016/j.ijpsycho.2007.03.005
Pollatos, O., Schandry, R., Auer, D. P., and Kaufmann, C. (2007c). Brain structures mediating cardiovascular arousal and interoceptive awareness. Brain Res. 1141, 178–187. doi: 10.1016/j.brainres.2007.01.026
Pollatos, O., Herbert, B. M., Wankner, S., Dietel, A., Wachsmuth, C., Henningsen, P., et al. (2011a). Autonomic imbalance is associated with reduced facial recognition in somatoform disorders. J. Psychosom. Res. 71, 232–239. doi: 10.1016/j.jpsychores.2011.03.012
Pollatos, O., Werner, N. S., Duschek, S., Schandry, R., Matthias, E., Traut-Mattausch, E., et al. (2011b). Differential effects of alexithymia subscales on autonomic reactivity and anxiety during social stress. J. Psychosom. Res. 70, 525–533. doi: 10.1016/j.jpsychores.2010.12.003
Pollatos, O., Kirsch, W., and Schandry, R. (2005). Brain structures involved in interoceptive awareness and cardioafferent signal processing: a dipole source localization study. Hum. Brain Mapp. 26, 54–64. doi: 10.1002/hbm.20121
Pollatos, O., Kurz, A. L., Albrecht, J., Schreder, T., Kleemann, A. M., Schopf, V., et al. (2008). Reduced perception of bodily signals in anorexia nervosa. Eat. Behav. 9, 381–388. doi: 10.1016/j.eatbeh.2008.02.001
Porges, S. W. (1993). Body Perception Questionnaire. Laboratory of Developmental Assessment. Baltimore, MD: University of Maryland.
Pruessner, J. C., Hellhammer, D. H., and Kirschbaum, C. (1999). Burnout, perceived stress, and cortisol responses to awakening. Psychosom. Med. 61, 197–204. doi: 10.1097/00006842-199903000-00012
Quirarte, G. L., Roozendaal, B., and Mcgaugh, J. L. (1997). Glucocorticoid enhancement of memory storage involves noradrenergic activation in the basolateral amygdala. Proc. Natl. Acad. Sci. U.S.A. 94, 14048–14053. doi: 10.1073/pnas.94.25.14048
Richards, J., Edgar, L., and Gibbon, P. (1996). Cardiac acuity in panic disorder. Cogn. Ther. Res. 20, 261–376. doi: 10.1007/BF02228039
Rief, W., and Auer, C. (2000). Cortisol and somatization. Biol. Psychol. 53, 13–23. doi: 10.1016/S0301-0511(00)00042-9
Rief, W., and Barsky, A. J. (2005). Psychobiological perspectives on somatoform disorders. Psychoneuroendocrinology 30, 996–1002. doi: 10.1016/j.psyneuen.2005.03.018
Rief, W., and Broadbent, E. (2007). Explaining medically unexplained symptoms-models and mechanisms. Clin. Psychol. Rev. 27, 821–841. doi: 10.1016/j.cpr.2007.07.005
Rief, W., Hessel, A., and Braehler, E. (2001). Somatization symptoms and hypochondriacal features in the general population. Psychosom. Med. 63, 595–602. doi: 10.1097/00006842-200107000-00012
Rief, W., Hiller, W., and Margraf, J. (1998a). Cognitive aspects of hypochondriasis and the somatization syndrome. J. Abnorm. Psychol. 107, 587–595. doi: 10.1037/0021-843X.107.4.587
Rief, W., Shaw, R., and Fichter, M. M. (1998b). Elevated levels of psychophysiological arousal and cortisol in patients with somatization syndrome. Psychosom. Med. 60, 198–203. doi: 10.1097/00006842-199803000-00016
Robbe, H. W., Mulder, L. J., Ruddel, H., Langewitz, W. A., Veldman, J. B., and Mulder, G. (1987). Assessment of baroreceptor reflex sensitivity by means of spectral analysis. Hypertension 10, 538–543. doi: 10.1161/01.HYP.10.5.538
Rockhold, R. W., and Caldwell, R. W. (1980). Cardiovascular effects following clonidine microinjection into the nucleus tractus solitarii of the rat. Neuropharmacology 19, 919–922. doi: 10.1016/0028-3908(80)90094-5
Roozendaal, B., Hahn, L. H., Nathan, S. V., De Quervain, D. J.-F., and Mcgaugh, J. L. (2004). Glucocorticoid effect on memory retrieval require concurrent noradrenergic activity in the hippocampus and basolateral amygdala. J. Neurosci. 34, 8161–8169. doi: 10.1523/JNEUROSCI.2574-04.2004
Roozendaal, B., Okuda, S., De Quervain, D. J., and Mcgaugh, J. L. (2006). Glucocorticoids interact with emotion-induced noradrenergic activation in influencing different memory functions. Neuroscience 138, 901–910. doi: 10.1016/j.neuroscience.2005.07.049
Roozendaal, B., Williams, C. L., and Mcgaugh, J. L. (1999). Glucocorticoid receptor activation in the rat nucleus of the solitary tract facilitates memory consolidation: involvement of the basolateral amygdala. Eur. J. Neurosci. 11, 1317–1323. doi: 10.1046/j.1460-9568.1999.00537.x
Rosenberger, C., Elsenbruch, S., Scholle, A., De Greiff, A., Schedlowski, M., Forsting, M., et al. (2009). Effects of psychological stress on the cerebral processing of visceral stimuli in healthy women. Neurogastroenterol. Motil. 21, e740–e745. doi: 10.1111/j.1365-2982.2009.01295.x
Russell, E., Koren, G., Rieder, M., and Van Uum, S. (2012). Hair cortisol as a biological marker of chronic stress: current status, future directions and unanswered questions. Psychoneuroendocrinology 37, 589–601. doi: 10.1016/j.psyneuen.2011.09.009
Russell, J. A. (2003). Core affect and the psychological construction of emotion. Psychol. Rev. 110, 145–172. doi: 10.1037/0033-295X.110.1.145
Schachinger, H., Degen, L., and Beglinger, C. (2009). Subliminal gastric distension enhances startle [Abstract]. Psychophysiology 46, S17.
Schachinger, H., Weinbacher, M., Kiss, A., Ritz, R., and Langewitz, W. (2001). Cardiovascular indices of peripheral and central sympathetic activation. Psychosom. Med. 63, 788–796. doi: 10.1097/00006842-200109000-00012
Schaefer, M., Egloff, B., Gerlach, A. L., and Witthoft, M. (2014). Improving heartbeat perception in patients with medically unexplained symptoms reduces symptom distress. Biol. Psychol. 101, 69–76. doi: 10.1016/j.biopsycho.2014.05.012
Schaefer, M., Egloff, B., and Witthoft, M. (2012). Is interoceptive awareness really altered in somatoform disorders? Testing competing theories with two paradigms of heartbeat perception. J. Abnorm. Psychol. 121, 719–724. doi: 10.1037/a0028509
Schandry, R. (1981). Heart beat perception and emotional experience. Psychophysiology 18, 483–488. doi: 10.1111/j.1469-8986.1981.tb02486.x
Schandry, R., Bestler, M., and Montoya, P. (1993). On the relation between cardiodynamics and heartbeat perception. Psychophysiology 30, 467–474. doi: 10.1111/j.1469-8986.1993.tb02070.x
Schandry, R., Sparrer, B., and Weitkunat, R. (1986). From the heart to the brain: a study of heartbeat contingent scalp potentials. Int. J. Neurosci. 30, 261–275. doi: 10.3109/00207458608985677
Schandry, R., and Specht, G. (1981). The influence of psychological and physical stress on the perception of heartbeats. [Abstract]. Psychophysiology 18, 154.
Schmitz, J., Blechert, J., Kramer, M., Asbrand, J., and Tuschen-Caffier, B. (2012). Biased perception and interpretation of bodily anxiety symptoms in childhood social anxiety. J. Clin. Child Adolesc. Psychol. 41, 92–102. doi: 10.1080/15374416.2012.632349
Schulz, A. (2015). “Interoception,” in International Encyclopedia of the Social and Behavioral Sciences, 2nd Rev. Edn, ed. J. D. Wright (Amsterdam: Elsevier), 614–620. doi: 10.1016/B978-0-08-097086-8.14145-5
Schulz, A., Ferreira De Sá, D. S., Dierolf, A. M., Lutz, A., Van Dyck, Z., Vögele, C., et al. (2015a). Short-term food deprivation increases amplitudes of heartbeat-evoked potentials. Psychophysiology 52, 695–703. doi: 10.1111/psyp.12388
Schulz, A., Köster, S., Beutel, M. E., Schächinger, H., Vögele, C., Rost, S., et al. (2015b). Altered patterns of heartbeat-evoked potentials in depersonalization/derealization disorder: neurophysiological evidence for impaired cortical representation of bodily signals. Psychosom. Med. 77, 506–516. doi: 10.1097/PSY.0000000000000195
Schulz, A., Van Dyck, Z., Lutz, A., and Vögele, C. (2015c). “Gastrische modulation der Schreckreaktion: eine prä-attentive methode zur abbildung afferenter signale aus dem gastrointestinalen system. [Gastric modulation of startle: a pre-attentive method to assess afferent signals originating from the gastrointestinal system],” in: The Proceeding of 41. Annual Meeting Psychologie und Gehirn [Psychology and Brain], Frankfurt.
Schulz, A., Lass-Hennemann, J., Nees, F., Blumenthal, T. D., Berger, W., and Schachinger, H. (2009a). Cardiac modulation of startle eye blink. Psychophysiology 46, 234–240. doi: 10.1111/j.1469-8986.2008.00768.x
Schulz, A., Lass-Hennemann, J., Richter, S., Romer, S., Blumenthal, T. D., and Schachinger, H. (2009b). Lateralization effects on the cardiac modulation of acoustic startle eye blink. Biol. Psychol. 80, 287–291. doi: 10.1016/j.biopsycho.2008.10.006
Schulz, A., Reichert, C. F., Richter, S., Lass-Hennemann, J., Blumenthal, T. D., and Schachinger, H. (2009c). Cardiac modulation of startle: effects on eye blink and higher cognitive processing. Brain Cogn. 71, 265–271. doi: 10.1016/j.bandc.2009.08.002
Schulz, A., Lass-Hennemann, J., Sutterlin, S., Schächinger, H., and Vögele, C. (2013a). Cold pressor stress induces opposite effects on cardioceptive accuracy dependent on assessment paradigm. Biol. Psychol. 93, 167–174. doi: 10.1016/j.biopsycho.2013.01.007
Schulz, A., Schilling, T. M., and Schachinger, H. (2013b). “Modulation der schreckreaktion durch viszeral-afferente signale aus dem kardialen und respiratorischen system. [Modulation of startle by visceral-afferent signals originating from the cardiovascular and respiratory system],” in The Proceeding of 39. Annual Meeting Psychologie und Gehirn [Psychology and Brain], Würzburg.
Schulz, A., Strelzyk, F., Ferreira De Sa, D. S., Naumann, E., Vögele, C., and Schächinger, H. (2013c). Cortisol rapidly affects amplitudes of heartbeat-evoked brain potentials-Implications for the contribution of stress to an altered perception of physical sensations? Psychoneuroendocrinology 38, 2686–2693. doi: 10.1016/j.psyneuen.2013.06.027
Schulz, A., Philippsen, C., and Schächinger, H. (2007). The influence of central Alpha2-adrenoreceptor agonism and antagonism on the cardiac modulation of acoustic startle. Psychophysiology 44, S29.
Schulz, A., Plein, D. E., Richter, S., Blumenthal, T. D., and Schachinger, H. (2011). Cold pressor stress affects cardiac attenuation of startle. Int. J. Psychophysiol. 79, 385–391. doi: 10.1016/j.ijpsycho.2010.12.008
Schulz, A., Richter, S., and Schächinger, H. (2010). “Evidence that cortisol-induced increase in baroreflex sensitivity of heart rate control is not due to afferent visceral neurotransmission,” in The Proceeding of 7th World Congress on Stress. Leiden.
Schwabe, L., Haddad, L., and Schachinger, H. (2008). HPA axis activation by a socially evaluated cold-pressor test. Psychoneuroendocrinology 33, 890–895. doi: 10.1016/j.psyneuen.2008.03.001
Sedeno, L., Couto, B., Melloni, M., Canales-Johnson, A., Yoris, A., Baez, S., et al. (2014). How do you feel when you can’t feel your body? Interoception, functional connectivity and emotional processing in depersonalization-derealization disorder. PLoS ONE 9:e98769. doi: 10.1371/journal.pone.0098769
Shao, S., Shen, K., Wilder-Smith, E. P., and Li, X. (2011). Effect of pain perception on the heartbeat evoked potential. Clin. Neurophysiol. 122, 1838–1845. doi: 10.1016/j.clinph.2011.02.014
Shin, Y. W., Dzemidzic, M., Jo, H. J., Long, Z., Medlock, C., Dydak, U., et al. (2013). Increased resting-state functional connectivity between the anterior cingulate cortex and the precuneus in panic disorder: resting-state connectivity in panic disorder. J. Affect. Disord. 150, 1091–1095. doi: 10.1016/j.jad.2013.04.026
Sloan, R. P., Shapiro, P. A., Bagiella, E., Gorman, J. M., and Bigger, J. T. Jr. (1995). Temporal stability of heart period variability during a resting baseline and in response to psychological challenge. Psychophysiology 32, 191–196. doi: 10.1111/j.1469-8986.1995.tb03311.x
Spielberger, C. D., and Reheiser, E. C. (2004). “Measuring anxiety, anger, depression, and curiosity as emotional states and personality traits with the STAI, STAXI, and STPI,” in Comprehensive Handbook of Psychological Assessment, eds M. J. Hilsenroth and D. L. Segal (Hoboken: Wiley), 70–86.
Stalder, T., Tietze, A., Steudte, S., Alexander, N., Dettenborn, L., and Kirschbaum, C. (2014). Elevated hair cortisol levels in chronically stressed dementia caregivers. Psychoneuroendocrinology 47, 26–30. doi: 10.1016/j.psyneuen.2014.04.021
Steptoe, A., and Vogele, C. (1991). Methodology of mental stress testing in cardiovascular research. Circulation 83, II14–II24.
Steptoe, A., and Vogele, C. (1992). Individual differences in the perception of bodily sensations: the role of trait anxiety and coping style. Behav. Res. Ther. 30, 597–607. doi: 10.1016/0005-7967(92)90005-2
Stevens, S., Gerlach, A. L., Cludius, B., Silkens, A., Craske, M. G., and Hermann, C. (2011). Heartbeat perception in social anxiety before and during speech anticipation. Behav. Res. Ther. 49, 138–143. doi: 10.1016/j.brat.2010.11.009
Streff, A., Kuehl, L. K., Michaux, G., and Anton, F. (2010). Differential physiological effects during tonic painful hand immersion tests using hot and ice water. Eur. J. Pain 14, 266–272. doi: 10.1016/j.ejpain.2009.05.011
Strelzyk, F., Hermes, M., Naumann, E., Oitzl, M., Walter, C., Busch, H. P., et al. (2012). Tune it down to live it up? Rapid, nongenomic effects of cortisol on the human brain. J. Neurosci. 32, 616–625. doi: 10.1523/JNEUROSCI.2384-11.2012
Strigo, I. A., Matthews, S. C., Simmons, A. N., Oberndorfer, T., Klabunde, M., Reinhardt, L. E., et al. (2013). Altered insula activation during pain anticipation in individuals recovered from anorexia nervosa: evidence of interoceptive dysregulation. Int. J. Eat. Disord. 46, 23–33. doi: 10.1002/eat.22045
Strohle, A., Holsboer, F., and Rupprecht, R. (2000). Increased ACTH concentrations associated with cholecystokinin tetrapeptide-induced panic attacks in patients with panic disorder. Neuropsychopharmacology 22, 251–256. doi: 10.1016/S0893-133X(99)00115-3
Sturges, L. V., and Goetsch, V. L. (1996). Psychophysiological reactivity and heartbeat awareness in anxiety sensitivity. J. Anxiety Disord. 10, 283–294. doi: 10.1016/0887-6185(96)00012-6
Suarez-Hitz, K. A., Otto, B., Bidlingmaier, M., Schwizer, W., Fried, M., and Ehlert, U. (2012). Altered psychobiological responsiveness in women with irritable bowel syndrome. Psychosom. Med. 74, 221–231. doi: 10.1097/PSY.0b013e318244fb82
Sved, A. F., Tsukamoto, K., and Schreihofer, A. M. (1992). Stimulation of alpha 2-adrenergic receptors in nucleus tractus solitarius is required for the baroreceptor reflex. Brain Res. 576, 297–303. doi: 10.1016/0006-8993(92)90693-4
Tak, L. M., Bakker, S. J., and Rosmalen, J. G. (2009). Dysfunction of the hypothalamic-pituitary-adrenal axis and functional somatic symptoms: a longitudinal cohort study in the general population. Psychoneuroendocrinology 34, 869–877. doi: 10.1016/j.psyneuen.2008.12.017
Tak, L. M., Cleare, A. J., Ormel, J., Manoharan, A., Kok, I. C., Wessely, S., et al. (2011). Meta-analysis and meta-regression of hypothalamic-pituitary-adrenal axis activity in functional somatic disorders. Biol. Psychol. 87, 183–194. doi: 10.1016/j.biopsycho.2011.02.002
Tak, L. M., and Rosmalen, J. G. (2010). Dysfunction of stress responsive systems as a risk factor for functional somatic syndromes. J. Psychosom. Res. 68, 461–468. doi: 10.1016/j.jpsychores.2009.12.004
Tanaka, Y., Ishitobi, Y., Maruyama, Y., Kawano, A., Ando, T., Imanaga, J., et al. (2012). Salivary alpha-amylase and cortisol responsiveness following electrical stimulation stress in panic disorder patients. Neurosci. Res. 73, 80–84. doi: 10.1016/j.neures.2012.01.006
Tanriverdi, F., Karaca, Z., Unluhizarci, K., and Kelestimur, F. (2007). The hypothalamo-pituitary-adrenal axis in chronic fatigue syndrome and fibromyalgia syndrome. Stress 10, 13–25. doi: 10.1080/10253890601130823
Terhaar, J., Viola, F. C., Bar, K. J., and Debener, S. (2012). Heartbeat evoked potentials mirror altered body perception in depressed patients. Clin. Neurophysiol. 123, 1950–1957. doi: 10.1016/j.clinph.2012.02.086
Thomas, K. S., Nelesen, R. A., Ziegler, M. G., Bardwell, W. A., and Dimsdale, J. E. (2004). Job strain, ethnicity, and sympathetic nervous system activity. Hypertension 44, 891–896. doi: 10.1161/01.HYP.0000148499.54730.0d
Tsai, H. W., Chan, P. Y., Von Leupoldt, A., and Davenport, P. W. (2013). The impact of emotion on the perception of graded magnitudes of respiratory resistive loads. Biol. Psychol. 93, 220–224. doi: 10.1016/j.biopsycho.2013.02.008
Uchida, R. R., Del-Ben, C. M., Busatto, G. F., Duran, F. L., Guimaraes, F. S., Crippa, J. A., et al. (2008). Regional gray matter abnormalities in panic disorder: a voxel-based morphometry study. Psychiatry Res. 163, 21–29. doi: 10.1016/j.pscychresns.2007.04.015
Usher, M., Cohen, J. D., Servan-Schreiber, D., Rajkowski, J., and Aston-Jones, G. (1999). The role of locus coeruleus in the regulation of cognitive performance. Science 283, 549–554. doi: 10.1126/science.283.5401.549
van den Bos, R., Harteveld, M., and Stoop, H. (2009). Stress and decision-making in humans: performance is related to cortisol reactivity, albeit differently in men and women. Psychoneuroendocrinology 34, 1449–1458. doi: 10.1016/j.psyneuen.2009.04.016
Van der Does, A. J., Van Dyck, R., and Spinhoven, P. (1997). Accurate heartbeat perception in panic disorder: fact and artefact. J. Affect. Disord. 43, 121–130. doi: 10.1016/S0165-0327(96)01414-0
Van Uum, S. H., Sauve, B., Fraser, L. A., Morley-Forster, P., Paul, T. L., and Koren, G. (2008). Elevated content of cortisol in hair of patients with severe chronic pain: a novel biomarker for stress. Stress 11, 483–488. doi: 10.1080/10253890801887388
von Leupoldt, A., Keil, A., Chan, P. Y., Bradley, M. M., Lang, P. J., and Davenport, P. W. (2010). Cortical sources of the respiratory-related evoked potential. Res. Physiol. Neurobiol. 170, 198–201. doi: 10.1016/j.resp.2009.12.006
Vrijkotte, T. G., Van Doornen, L. J., and De Geus, E. J. (2004). Overcommitment to work is associated with changes in cardiac sympathetic regulation. Psychosom. Med. 66, 656–663. doi: 10.1097/01.psy.0000138283.65547.78
Wang, J., Korczykowski, M., Rao, H., Fan, Y., Pluta, J., Gur, R. C., et al. (2007). Gender difference in neural response to psychological stress. Soc. Cogn. Affect. Neurosci. 2, 227–239. doi: 10.1093/scan/nsm018
Wang, J., Rao, H., Wetmore, G. S., Furlan, P. M., Korczykowski, M., Dinges, D. F., et al. (2005). Perfusion functional MRI reveals cerebral blood flow pattern under psychological stress. Proc. Natl. Acad. Sci. U.S.A. 102, 17804–17809. doi: 10.1073/pnas.0503082102
Weiss, S., Sack, M., Henningsen, P., and Pollatos, O. (2014). On the interaction of self-regulation, interoception and pain perception. Psychopathology 47, 377–382. doi: 10.1159/000365107
Werner, N. S., Jung, K., Duschek, S., and Schandry, R. (2009). Enhanced cardiac perception is associated with benefits in decision-making. Psychophysiology 46, 1123–1129. doi: 10.1111/j.1469-8986.2009.00855.x
Whitehead, W. E., Drescher, V. M., Heiman, P., and Blackwell, B. (1977). Relation of heart rate control to heartbeat perception. Biofeedback Self Regul. 2, 371–392. doi: 10.1007/BF00998623
Wiens, S. (2005). Interoception in emotional experience. Curr. Opin. Neurol. 18, 442–447. doi: 10.1097/01.wco.0000168079.92106.99
Willem Van der Does, A. J., Antony, M. M., Ehlers, A., and Barsky, A. J. (2000). Heartbeat perception in panic disorder: a reanalysis. Behav. Res. Ther. 38, 47–62. doi: 10.1016/S0005-7967(98)00184-3
Wingenfeld, K., Heim, C., Schmidt, I., Wagner, D., Meinlschmidt, G., and Hellhammer, D. H. (2008). HPA axis reactivity and lymphocyte glucocorticoid sensitivity in fibromyalgia syndrome and chronic pelvic pain. Psychosom. Med. 70, 65–72. doi: 10.1097/PSY.0b013e31815ff3ce
Wingenfeld, K., Wagner, D., Schmidt, I., Meinlschmidt, G., Hellhammer, D. H., and Heim, C. (2007). The low-dose dexamethasone suppression test in fibromyalgia. J. Psychosom. Res. 62, 85–91. doi: 10.1016/j.jpsychores.2006.06.006
Wirtz, P. H., Siegrist, J., Rimmele, U., and Ehlert, U. (2008). Higher overcommitment to work is associated with lower norepinephrine secretion before and after acute psychosocial stress in men. Psychoneuroendocrinology 33, 92–99. doi: 10.1016/j.psyneuen.2007.10.003
Witthoft, M., Basfeld, C., Steinhoff, M., and Gerlach, A. L. (2012). Can’t suppress this feeling: automatic negative evaluations of somatosensory stimuli are related to the experience of somatic symptom distress. Emotion 12, 640–649. doi: 10.1037/a0024924
Wolk, J., Sutterlin, S., Koch, S., Vogele, C., and Schulz, S. M. (2014). Enhanced cardiac perception predicts impaired performance in the Iowa Gambling Task in patients with panic disorder. Brain Behav. 4, 238–246. doi: 10.1002/brb3.206
Wong, M. L., Kling, M. A., Munson, P. J., Listwak, S., Licinio, J., Prolo, P., et al. (2000). Pronounced and sustained central hypernoradrenergic function in major depression with melancholic features: relation to hypercortisolism and corticotropin-releasing hormone. Proc. Natl. Acad. Sci. U.S.A. 97, 325–330. doi: 10.1073/pnas.97.1.325
Keywords: chronic stress, HPA axis, interoception, SAM axis, somatization, stress disorders, sympathetic nervous system, symptom perception
Citation: Schulz A and Vögele C (2015) Interoception and stress. Front. Psychol. 6:993. doi: 10.3389/fpsyg.2015.00993
Received: 13 January 2015; Accepted: 30 June 2015;
Published: 20 July 2015.
Edited by:
Olga Pollatos, University of Ulm, GermanyReviewed by:
Tony W. Buchanan, Saint Louis University, USAGiorgio Marchetti, Mind, Consciousness, and Language Research Net, Italy
Copyright © 2015 Schulz and Vögele. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: André Schulz, Institute for Health and Behaviour, Integrative Research Unit on Social and Individual Development, University of Luxembourg, 11, Porte des Sciences, L-4366 Esch-sur-Alzette, Luxembourg,YW5kcmUuc2NodWx6QHVuaS5sdQ==