 Haixia Guo
Haixia Guo Sheng Luo1
Sheng Luo1 Yike Li
Yike Li Xie Wang
Xie Wang
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Plant Sci. , 17 March 2025
Sec. Plant Nutrition
Volume 16 - 2025 | https://doi.org/10.3389/fpls.2025.1520936
Introduction: Stoichiometric homeostasis is an important strategy used by plants to function optimally in changing environments.
Methods: In order to investigate whether plants under stricter resource restrictions exhibit stronger homeostasis, this study took M. alba inhabiting in a dry-hot valley as the research subject.
Results: The stoichiometry of M. alba leaves, their variations in response to altitude and slope, and their correlations with soil were analyzed. The results showed that soil nutrient levels were higher on the shady slope compared to the sunny slope, and responded differently to altitude on the two slopes. On the sunny slope, soil carbon (C) content increased significantly with altitude, whereas on the shady slope, soil phosphorus (P) content decreased with increasing altitude. The C: N and C: P ratios of the soil were lower than the average in China. The C: N and C: P ratios of M. alba leaves were lower than those of global and Chinese forest ecosystems. The N: P ratio of M. alba leaves was < 14. However, no significant correlation was observed between M. alba leaves and soil C, N, P, or stoichiometric characteristics. The changes in C, N, and P and their ratios in M. alba leaves did not correspond with those in the soil. M. alba exhibited "strict homeostasis" on both sunny and shady slopes.
Discussion: The results suggest that M. alba's growth is limited by nutrients availability, particularly nitrogen. The strict stoichiometric homeostasis is an adaptation strategy for M. alba in dry-hot valleys to alleviate nutrient limitations, which leads to a decoupling of ecological stoichiometry between M. alba leaves and soil.
Ecological stoichiometry examines the balance of multiple chemical elements in ecological interactions, with a focus on C, N, and P (Elser et al., 2000, 2010; Fan et al., 2015). It serves as a powerful tool for understanding plant adaptation strategies to changing environments and detecting nutrient limitations (Güsewell, 2004; Zhou et al., 2024). To function optimally, organisms must maintain a relatively fixed C:N:P ratio, a concept known as stoichiometric homeostasis (Sterner and Elser, 2002; Güsewell, 2004). This homeostatic balance exists at both the individual and community levels (Bertrand et al., 2019). The concept of stoichiometric homeostasis was first introduced by Redfield in 1958, who discovered that the C:N:P ratio of plankton remained consistently at 106:16:1 (Redfield, 1958). Since then, stoichiometric homeostasis has been observed in various organisms, including microbes, forests, and herbaceous plants (Makino et al., 2003; Zhang et al., 2018). However, plant stoichiometry often changes in response to environmental fluctuations, as plants primarily obtain their nutrients from the soil (Reich and Oleksyn, 2004). Wirtz and Kerimoglu (2016) termed this variation as stoichiometric flexibility, identifying it is a strategy autotrophs use to optimize resource utilization under nutrient-limited conditions Stoichiometric flexibility is influenced by various factors, including an organism’s nature, climate, altitude, nutrient availability, intensity of perturbation, and geographic range size (Elser et al., 2010; Sistla and Schimel, 2012; Bertrand et al., 2019). To quantify an organism’s ability to maintain stoichiometric homeostasis or its range of stoichiometric flexibility, Sterner and Elser (2002) proposed a continuously variable regulation parameter (H), which was found to vary significantly among different organisms (Elser et al., 2010). Numerous studies have explored stoichiometric flexibility across various organisms and levels, leading to some generalizable hypotheses. For instance, stoichiometric homeostasis tends to be stricter at higher trophic levels compared than at lower ones (Hessen et al., 2004; Sistla and Schimel, 2012) and increases with scale (Sistla and Schimel, 2012). Moreover, autotrophs exhibit greater variability in stoichiometric ratios than heterotrophs across the food web (Hessen et al., 2004; Persson et al., 2010; Bertrand et al., 2019).
One of the main factors influencing stoichiometric flexibility is the shift in nutrient limitation (Güsewell, 2004; Sistla and Schimel, 2012). In theory, stoichiometric homeostasis is a strategy plants use to mitigate resource limitations (Rastetter and Shaver, 1992; Wirtz and Kerimoglu, 2016). From this perspective, plants in nutrient-limited environments are expected to maintain stricter stoichiometric homeostasis (Güsewell, 2004). To achieve this, they employ various adaptive strategies. An excessively strong homeostatic mechanism could even lead to the decoupling of plants from soil. Some researchers have found that soil nutrients influence plant stoichiometric homeostasis, with plants experiencing stricter resource limitations exhibiting stronger homeostasis (Yu et al., 2011; Chen et al., 2016; Su and Shangguan, 2022). Han et al. (2011) proposed the Stability of Limiting Elements Hypothesis, which suggests that variability and environmental sensitivity are lowest for elements that are most limiting in nature, indicating that plant stoichiometric homeostasis varies with nutrient limitation. Chen et al. (2016) found that during the ecological restoration, as soil nutrients improved, the stoichiometry of D. dichotoma shifted from strong stoichiometric homeostasis in the early stage to weak stoichiometric homeostasis in the later stage. This finding also suggests that plants exhibit stronger internal homeostasis in more nutrient-restricted environments. However, this hypothesis remains unconfirmed, as the studies mentioned above did not directly address the flexibility of plants’ stoichiometric ratios are in resource-constrained environments.
The dry-hot river valley is a distinct type of river valley characterized by high temperatures, aridity, and low air humidity (Ya et al., 2004). In China, these valleys are primarily found in Yunnan, northwestern Taiwan, southwestern Hainan, and southwestern Sichuan. Within Sichuan Province, they are mainly located along the Jinsha, Yalong, and Dadu rivers in southern Ganzi, as well as the Jinsha River in Panzhihua and Liangshan (Zheng, 2010). Plant growth in these valleys is severely limited by high temperatures, arid conditions, poor soil fertility, and severe soil erosion (Shoukang et al., 2022). To optimize their fitness, plants in dry-hot river valley have evolved specific stoichiometric strategies to adapt to this environment. They typically exhibit higher leaf N and P contents and lower C:N and C:P ratios (Hong-bo et al., 2021). However, previous research has primarily focused on nutrient content and ratios, with no studies to date investigating the homeostatic stoichiometric features of plants in this region. It remains unclear whether plants enhance their adaptability through strong homeostasis. Furthermore, numerous studies have indicated that soil fertility in dry-hot valleys increases with precipitation and soil nutrient content along an elevation gradient (Lei Shan-Yu et al., 2022; Chang-ming et al., 2023) and is higher on shaded slopes compared to sunny slopes (Shoukang et al., 2022; Chang-ming et al., 2024). However, it remains uncertain whether these variations in nutrient status influence the homeostasis of plants in dry-hot valleys.
Morus alba is a common economic timber species in China’s dry-hot valleys, valued for both its fruit and leaves (Batiha et al., 2023). It plays a crucial role in soil and water conservation and soil improvement, exhibiting a rapid growth rate, strong drought resistance, and high environmental adaptability (Jianfeng et al., 2016; Xie et al., 2024). As a result, it has been widely planted throughout the Jinsha River valley since the 1990s (Mingqin, 1996; Sheng et al., 1999). This study examines M. alba by analyzing the stoichiometric characteristics of its leaves, its variation with altitude, differences between sunny and shady slopes, and correlations with soil. The objective is to determine whether plants in dry-hot valleys exhibit strong stoichiometric homeostasis and whether this homeostasis varies with altitude and slope.
The study area is located in Yanbian County, Panzhihua City, Sichuan Province, China (101.52°∼101.53°E, 26.95°∼26.96°N), within the dry-hot river valley at the junction of Sichuan and Yunnan provinces (Figure 1). This region experiences a typical South Asian tropical dry-hot valley climate, with an annual average temperature was 19.2°C. The three main soil types are bauxite, red soil, and yellow-red soil.
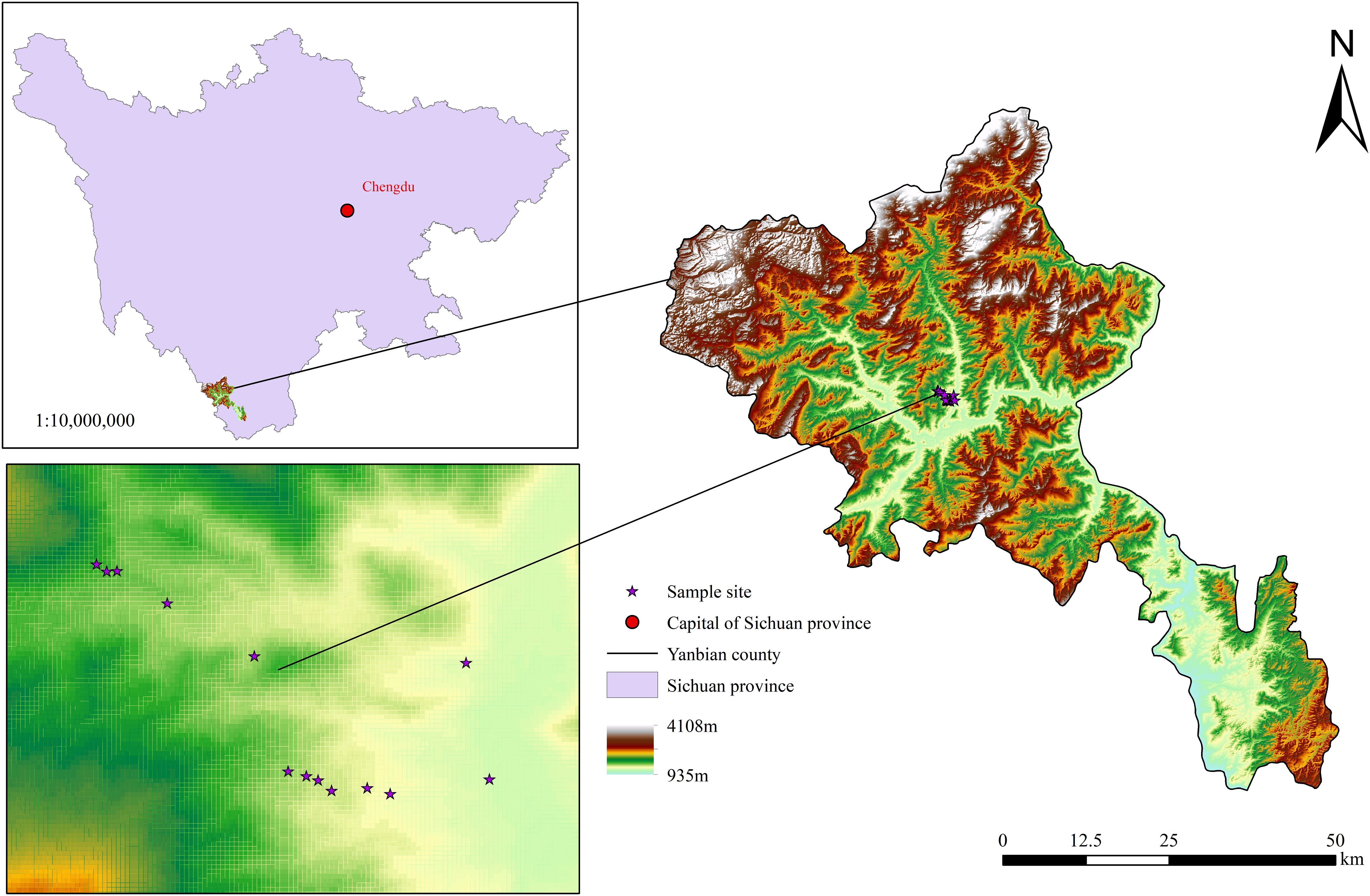
Figure 1. The study area.
The predominant variety of M. alba cultivated in this region is Yunsang No. 2, which is used for both fruit and leaf production. In the study area, M. alba is primarily found at altitudes between 1,000 and 2,000 m asl, with most trees concentrated in the mulberry forest at 1,200–1,500 m asl. At altitudes of 1,000∼1,200 and 1,500∼2,000 m asl, M. alba was observed to be scattered along roadsides, ditches, and near houses.
All samples were collected in June 2023 from eight sampling sites in mulberry orchards that had been established for more than 3 years, at altitudes ranging from 1,200 to 1,500 m asl. Sampling was conducted on both sunny and shady slopes (Figure 1). The mulberry trees had trunk diameters of 6–10 cm, with row spacing of 1.5–2 and plant spacing of 0.6–1 m.
At each sampling site, three 10 m2 × 10 m2 sampling plots were randomly selected. Withon each plot, three 1 m2 × 1 m2 quadrats were placed along the diagonal (at both ends and the midpoint). In each quadrat, five soil samples were collected from the center and corners at a depth of 0 to 25 cm using a soil drill, then combined into a single composite sample. Similarly, healthy M. alba leaf samples were collected from each quadrat.
M. alba leaves were oven-dried at 70°C for 48 h and then powdered. Soil samples were air-dried and subsequently ground. The ground samples were used to determine organic carbon content, total nitrogen content, and total phosphorus content using the potassium dichromate oxidation external heating method, the micro-Kjeldahl method, and the ammonium molybdate method, respectively. Measurement results are expressed as nutrient content per unit mass (g kg−1).
Statistical analyses were performed using R (version 4.4.1) and SPSS 22.0 (SPSS Inc, Chicago, USA). Variance decomposition was conducted to assess the relative effects of altitude, slope, and their interactions using the “vegan” package in R. Nutrient levels and stoichiometric ratios between sunny and shady slopes were compared using a t-test. To identify trends in nutrient contents and stoichiometric ratios at different altitudes, linear curve fitting was applied to examine the link between C, N, and P contents, stoichiometric ratios, and altitude. Spearman correlation analysis was used to assess the relationships between the stoichiometric ratios and C, N, and P contents.
The distribution ranges of difference values (D-values) for nutrient contents and stoichiometric ratios between M. alba leaves and soil were analyzed to determine if M. alba leaves maintained synchrony with soil. Firstly, the nutrient concentrations and stoichiometric ratios of mulberry leaves and soil were standardized as follows:
Subsequently, the difference between the two standardized values was calculated (e.g., leaf C − soil C). If the distribution of D-values remained within the 95% confidence interval of its mean, it indicated that changes in leaves were synchronized with those in the soil. Conversely, if the distribution exceeded the 95% confidence interval, it was assumed that leaf changes were not synchronized with soil changes.
The strength of plant stoichiometric homeostasis was analyzed on a log–log scale using the model: log(y) = log(c) + (1/H)log(x), where y represents the content of C, N, or P, or the ratios of C:N, C:P, or N:P in leaves; x represents the content of C, N, or P, or the ratios of C:N, C:P, or N:P in soil; and c is a constant. Values for H and c are determined through regression analysis of the relationship between y and x. The value of 1/H was derived from the regression slope between log x and log y, ranging from 0.00 to 1.00. To assess stoichiometric homeostasis, one-tailed tests with α = 0.10 were conducted. If the regression relationship was not significant (p > 0.10), the plant was classified as “strictly homeostatic”. However, if the regression relationship was significant (p < 0.10), stoichiometric homeostasis was categorized into four levels: homeostatic (0 < 1/H < 0.25), weakly homeostatic (0.25 < 1/H < 0.5), weakly plastic (0.5 < 1/H < 0.75), and plastic (1/H > 0.75) (Figure 2).
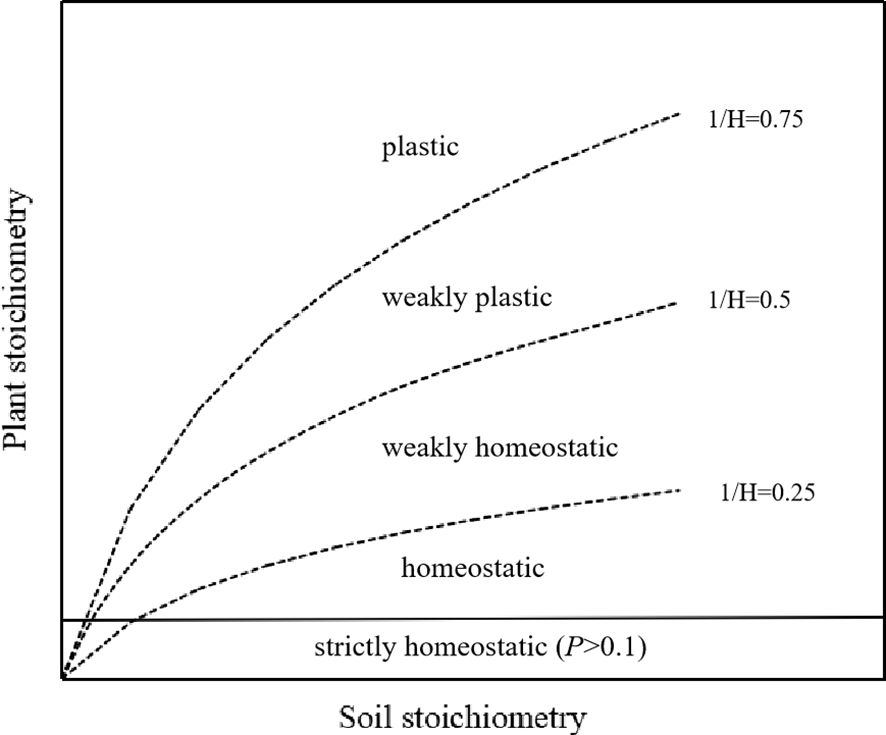
Figure 2. Potential patterns relating soil to plant stoichiometry (adapted from Sterner and Elser, 2002; Zhou et al., 2024).
All figures in this paper were created using Origin 2024 (OriginLab Corporation, Northampton, MA, USA).
The soil had significantly lower C, N, and P contents, as well as C:N, C:P, and N:P ratios, compared to M. alba leaves (p < 0.05). Moreover, the coefficients of variation for soil C, N, and P contents, as well as C:N, C:P, and N:P ratios, were higher than those for M. alba leaves (Table 1).
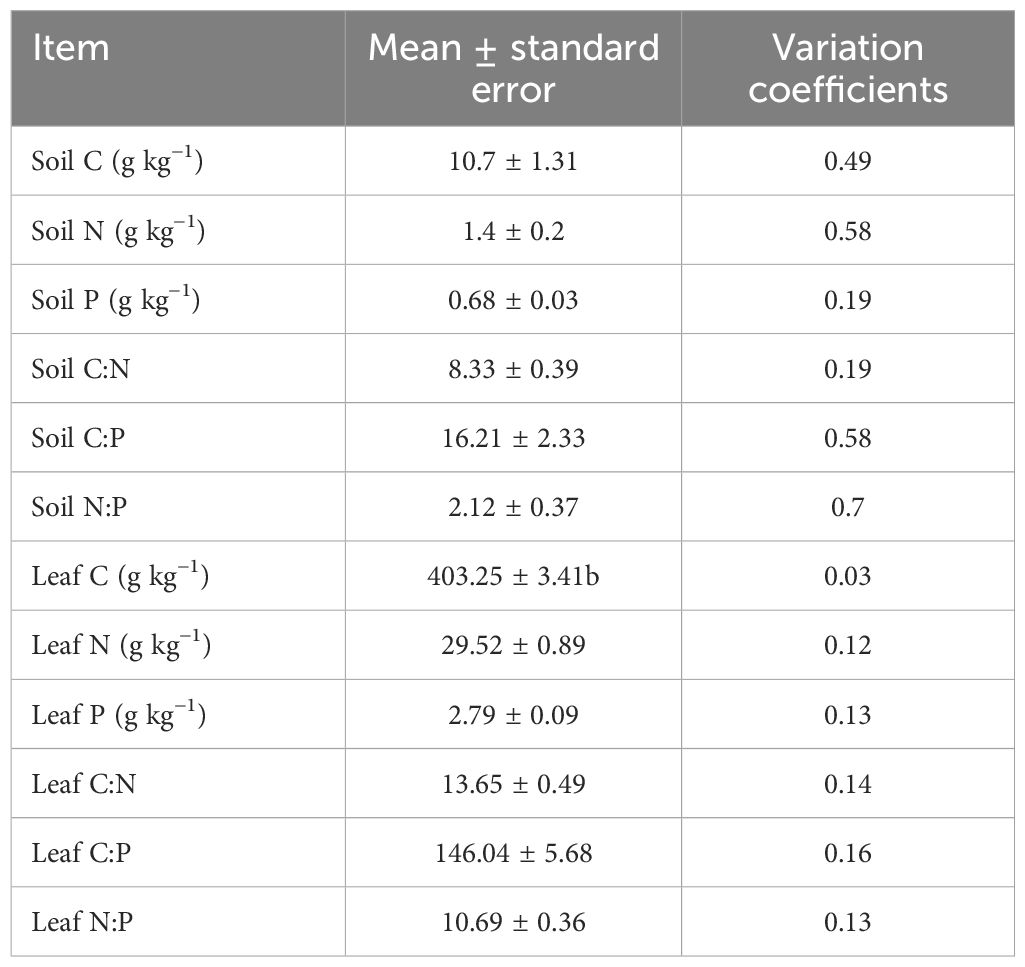
Table 1. Variation coefficients for stoichiometric ratios and C, N, and P contents in soil and M. alba leaves.
Slope significantly influenced soil C, N, C:N, C:P, and N:P (p < 0.01), whereas altitude had a significant effect on soil P (p < 0.01). Additionally, soil C:P was influenced by both slope and altitude. Variance decomposition analysis indicated that slope was the primary factor influencing soil C, N, C:N, C:P, and N:P. For soil C, N, and N:P, the order of influence magnitude was slope > interaction effects > altitude. For soil C:N and C:P, the order of influence magnitude was slope > altitude > interaction effects. The dominant factor affecting soil P was slope (Figure 3).
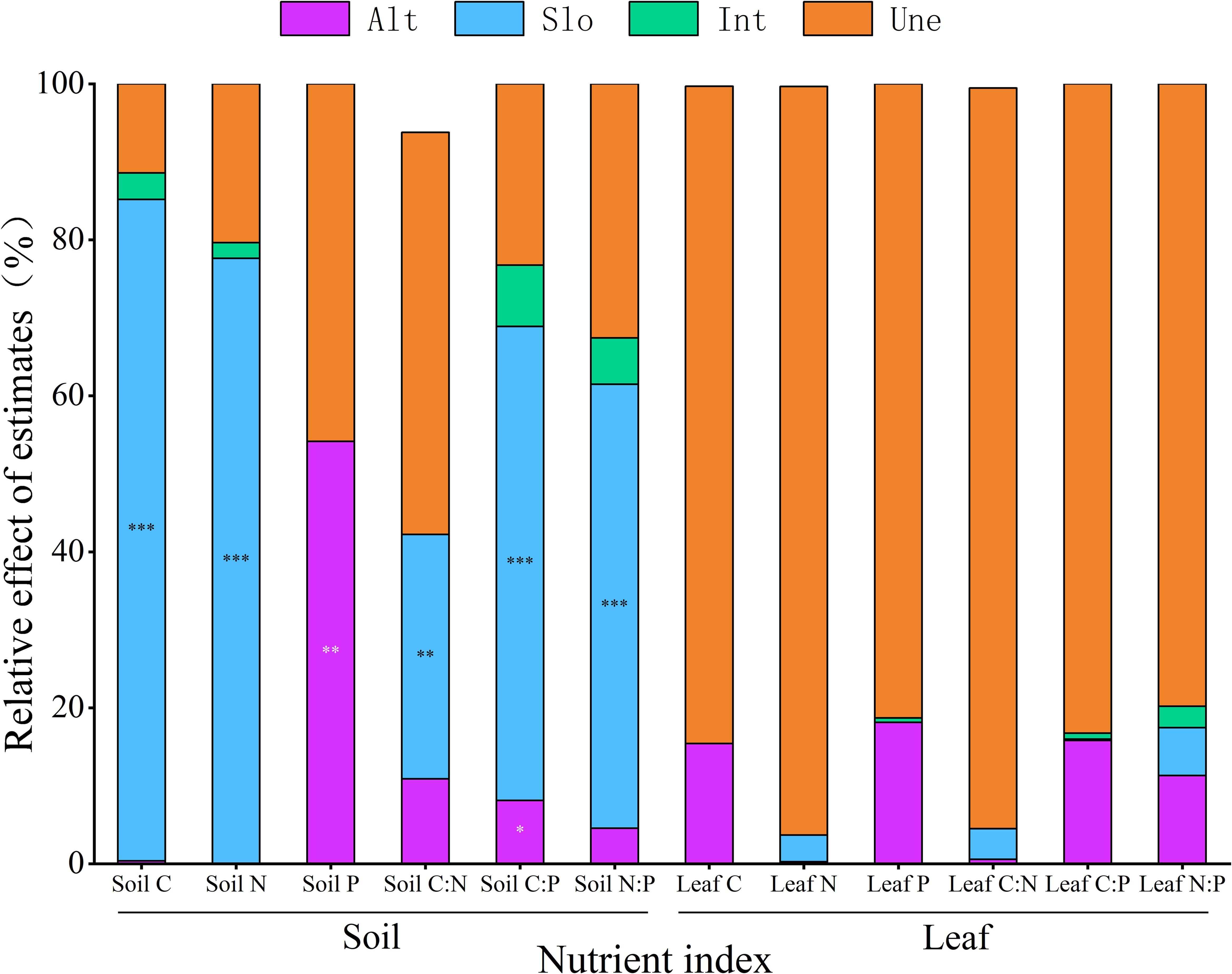
Figure 3. The effects of altitude and slope and their interactions on C, N, and P contents and stoichiometric ratios of soil and M. alba leaves. *p < 0.05; **p < 0.01; ***p < 0.001. Alt, altitude; Slo, slope; Int, interactions; Une, unexplained.
The nutrient content and stoichiometric ratios of the soil differed significantly between the shady and sunny slopes (p < 0.05). The shady slope had a higher concentration of carbon (15.47 g·kg−1) than the sunny slope (5.93 g·kg−1). Similarly, nitrogen content was significantly higher on the shady slope (2.10 g·kg−1) compared to the sunny slope (0.69 g·kg−1). In contrast, the difference in phosphorus content between the two slopes was not significant (p > 0.05), with the shady slope having a slightly higher concentration (0.69 g·kg−1) than the sunny slope (0.68 g·kg−1). The C:P and N:P ratios were significantly higher on the shady slope than on the sunny slope (p < 0.05), whereas the C:N ratio was lower (p < 0.05) (Figure 4a).
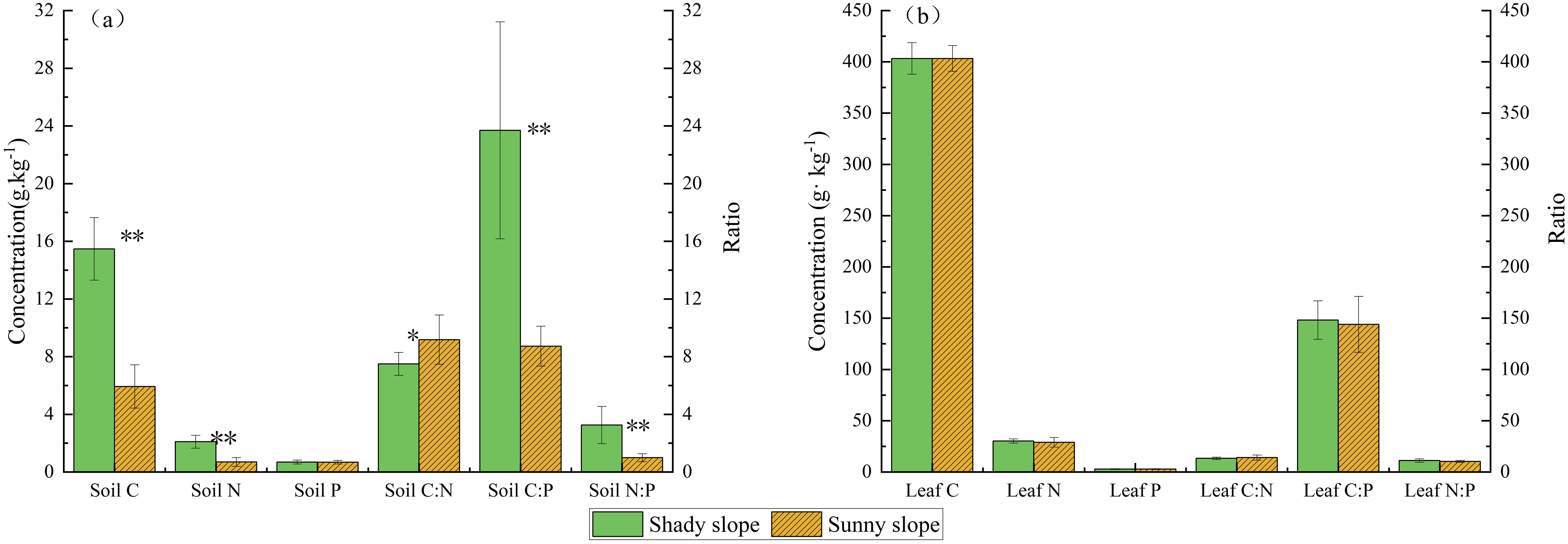
Figure 4. C, N, and P contents and stoichiometric ratios in soil (a) and M. alba leaves (b) on sunny and shady slopes. *p < 0.05; **p < 0.01;.
However, no significant difference was found in the stoichiometric ratios or the C, N, and P contents of M. alba leaves between the shady and sunny slopes (p > 0.05) (Figure 4b).
The response of nutrient contents and stoichiometric ratios in soil and M. alba leaves to altitude varied significantly between sunny and shady slopes (Table 2).
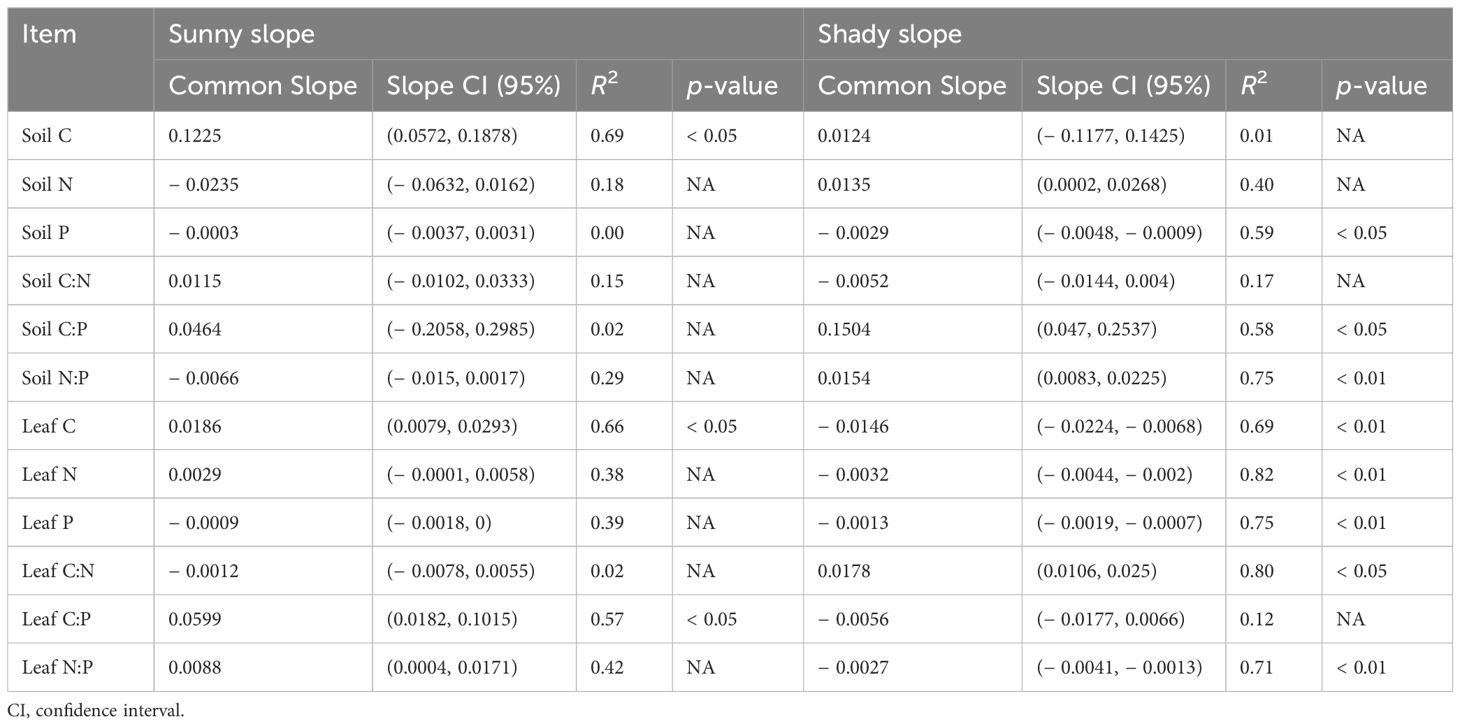
Table 2. Curve fitting of nutrient contents, stoichiometric ratios, and altitude.
On the sunny slope, only soil C showed a significant linear increase with altitude (p < 0.05). In contrast, on the shady slope, soil P decreased linearly with altitude, while soil C:P and N:P increased (p < 0.05).
For M. alba, leaf C scaled linearly and positively with altitude on the sunny slope (slope = 0.0186, R2 = 0.66, p < 0.05) but negatively on the shady slope (slope = − 0.0146, R2 = 0.69, p < 0.01). Leaf N, P, and N:P decreased linearly with increasing altitude on the shady slope (p < 0.05) but showed no significant change on the sunny slope (p > 0.05). Similarly, leaf C:N increased linearly with altitude on the shady slope (p < 0.05) but remained relatively constant on the sunny slope (p > 0.05). In contrast, leaf C:P showed a positive linear relationship with altitude on the sunny slope (slope = 0.0599, R2 = 00.57, p < 0.01), but no significant linear relationship was found on the shady slope (p > 0.05).
A positive correlation between was found between soil C and N (r = 0.98, p < 0.05). Additionally, both soil C and N were positively correlated with soil C:P and N:P but negatively correlated with soil C:N (p < 0.05). However, the correlations among leaf C, N, and P and leaf stoichiometric ratios were all found to be insignificant (p > 0.05). Leaf N was negatively correlated with leaf C:N, but positively correlated with leaf N:P (p < 0.05). Similarly, leaf P showed a negative correlation with leaf C:P but a positive correlation with N:P (p < 0.05). Furthermore, no significant correlation was found between soil and leaf (p > 0.05) (Figure 5).
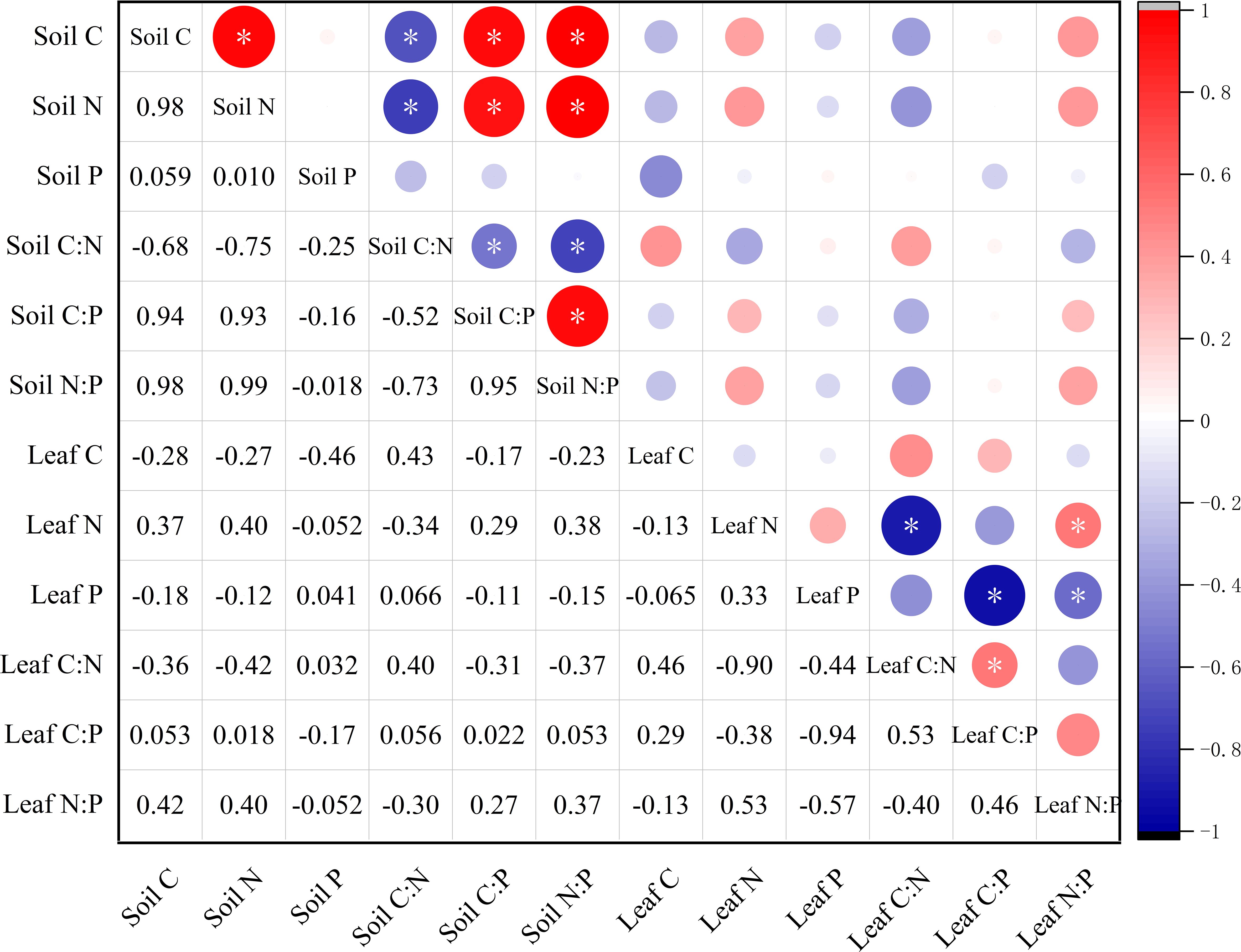
Figure 5. Correlations among C, N, and P contents and stoichiometric ratios in soil and M. alba leaves. *p < 0.05.
On both shady and sunny slopes, the D-values in nutrient contents and stoichiometric ratios between M. alba leaves and soil did not stabilize within a specific range (95% confidence interval of the means). These results suggested that the nutrient contents and stoichiometric ratios of M. alba leaves did not correspond to changes in soil (Figure 6).
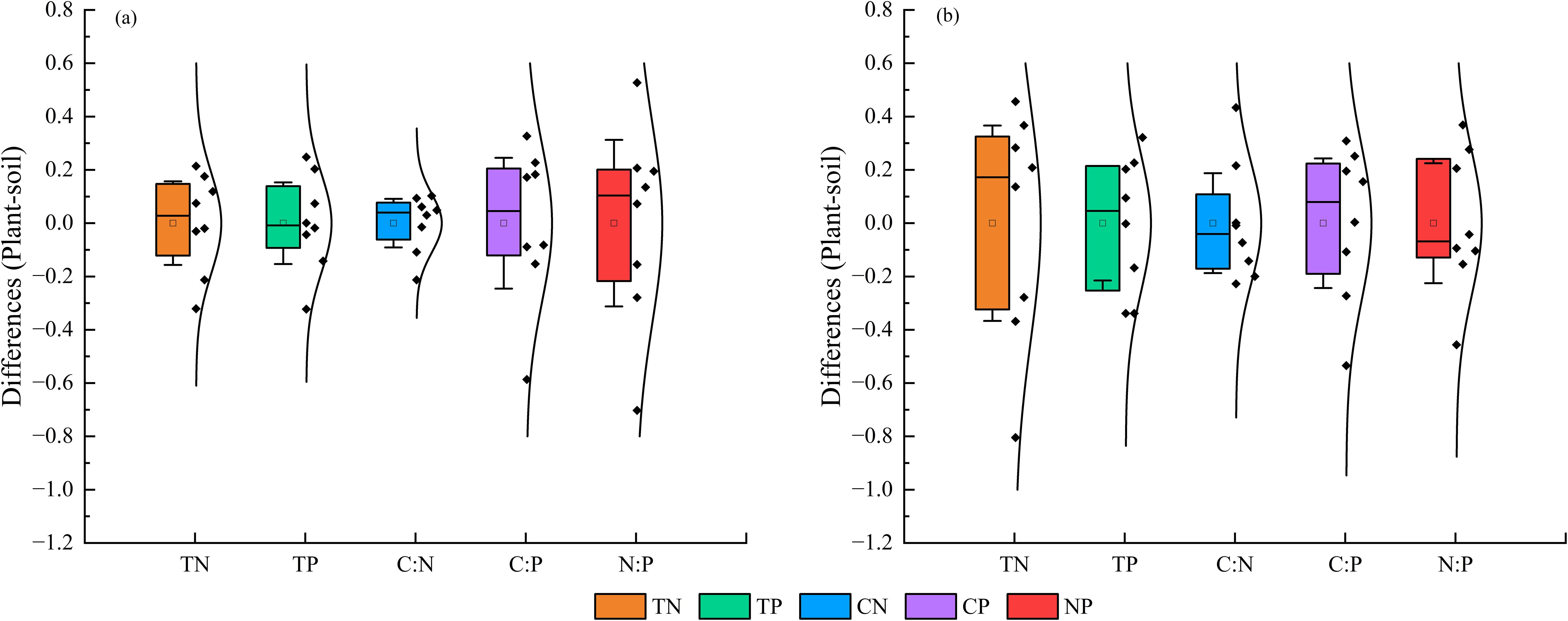
Figure 6. D-values of nutrient contents and stoichiometric ratios between M. alba leaves and soil on shady (a) and sunny (b) slopes. The bars above and below the box represent the 95% confidence intervals of the mean.
In this study, the regression relationships between M. alba leaves and soil on both shady and sunny slopes were not significant (p > 0.10). Therefore, M. alba should be classified as strictly homeostatic (Table 3).
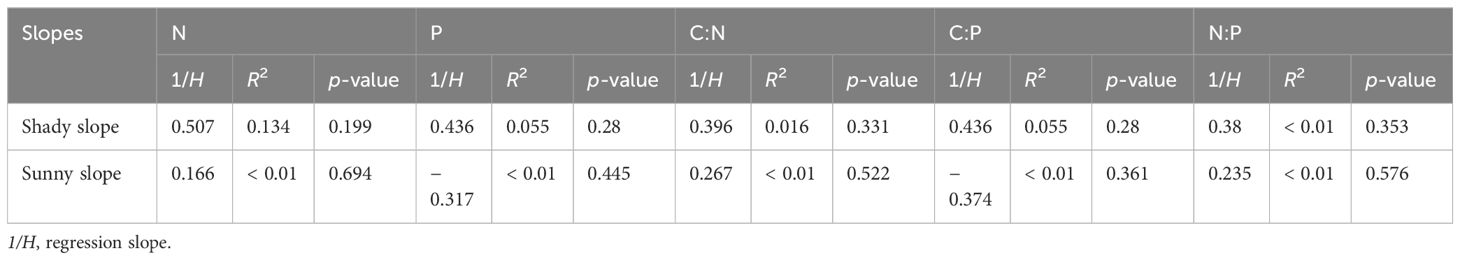
Table 3. Homeostasis coefficients (1/H) between sunny and shady slopes.
The dry-hot valley has traditionally been regarded as a nutrient-poor and water-scarce environment, unsuitable for plant growth (Ya et al., 2004; Duan et al., 2017). However, in this study area, the average soil organic carbon, total soil nitrogen, and total phosphorus were 10.7, 1.4, and 0.68g·kg−1, respectively. These values indicate that the soil nutrient level in the study area is at a medium level compared to the national average (Fangyan et al., 2007; Tian et al., 2010; Zi-qi et al., 2022). Further analysis revealed that the soil nutrient level on the sunny slope was lower, consistent with most studies on dry-hot valleys (Guo, 2014; Guan, 2022; Hang et al., 2022; Yiting et al., 2023), whereas the nutrient level on the shady slope was at a medium level. These findings highlight the importance of distinguishing between the two slopes in future research.
Soil stoichiometry is a useful tool for understanding the cycling of elements in soil (Klemmedson and Wienhold, 1992; Zechmeister-Boltenstern et al., 2015). Soil C:N and C:P ratios reflect the rates of soil organic matter (SOM) decomposition, nutrient mineralization or immobilization, and plant nutrient limitation. A lower C:N ratio indicates a faster SOM mineralization rate, with the cumulative SOM rate being lower than the decomposition rate (Enwezor, 1976; Bui and Henderson, 2013; Pan et al., 2024). Although less frequently used, the soil C:P ratio also serves as a useful indicator of the source/nature of organic matter, with a lower C:P suggesting a higher SOM mineralization rate (Bui and Henderson, 2013). In this study, the C:N ratio in the dry-hot valley (8.33) was lower than the Chinese national average (11.9) (Tian et al., 2010; Pan et al., 2024) and findings from other studies on dry-hot valleys (Changming et al., 2022; Yiting et al., 2023; Zhifeng et al., 2024). Similarly, the C:P ratio in the study area (16.21) was significantly lower than the Chinese national average (61) (Tian et al., 2010; Pan et al., 2024) and findings from other studies on dry-hot valleys (Changming et al., 2022; Yiting et al., 2023; Zhifeng et al., 2024). These results suggest that SOM mineralization in the study area was higher than the Chinese national average and other reported dry-hot valley studies (Tian et al., 2010; Bui and Henderson, 2013).
Nitrogen and phosphorus are essential nutrients for plant growth and often serve as limiting factors in terrestrial ecosystems (Reich and Oleksyn, 2004; Yanan et al., 2014; Zhang et al., 2018). The N:P ratio in plant tissues is a widely used indicator of nutrient limitation, with values > 16 suggesting phosphorus limitation and values < 14 indicating nitrogen limitation (Güsewell, 2004; Han et al., 2005; Bertrand et al., 2019). In this study, no significant difference was observed in the N:P ratio of M. alba leaves between the sunny and shady slopes in the dry-hot valley, with an average of 10.28 on the sunny slope and 11.10 on the shady slope—both below 14. These findings suggest that M. alba growth is limited by nitrogen. The C:N and C:P ratios of plant leaves reflect nitrogen and phosphorus use efficiency as well as the plant’s capacity for carbon fixation (Sun et al., 2019; Pan et al., 2024). In this study, the C:N and C:P ratios of M. alba leaves in the dry-hot valley were 13.65 and 146.04, respectively, which were lower than the value reported for global (37.1, 469.2) and Chinese forest ecosystems (28.5, 513.0) (Han et al., 2005). This indicates that the plant’s nitrogen and phosphorus utilization efficiency, as well as its carbon fixation capacity, are lower in this study area compared to broader global and Chinese ecosystems (Sun et al., 2019). Additionally, reducing the C:N and C:P ratios is a known adaptive strategy that plants use to cope with resource-limited environments (Elser et al., 1996, 2000). Some species in dry-hot valleys have been observed to adopt this strategy (Lin et al., 2019; Jingwen et al., 2020; Hang et al., 2022), and M. alba appears to employ a similar mechanism to enhance its growth under these conditions.
Slope aspect significantly influences soil nutrient levels by regulating water and energy availability and energy input, thereby shaping local abiotic and biotic environments (Bale et al., 1998; Yang et al., 2020; Qin et al., 2021). This is consistent with the findings of this study, where the primary effects on soil organic carbon, soil nitrogen, C:N, C:P, and N:P were attributed to slope differences. Previous studies have also shown that shady slopes, characterized by lower temperatures, reduced solar radiation, smaller temperature fluctuations, and higher topsoil water retention, promote organic matter accumulation, leading to more fertile compared to sunny slopes (Sharma et al., 2010; Zhaoyang et al., 2019; Tan et al., 2020; Yang et al., 2020; Liu et al., 2024). Research conducted in the dry-hot valleys of the Jinsha River (Chang-ming et al., 2024) and Minjiang River (Nan, 2016; Yang et al., 2020) similarly indicates that soil fertility is higher on shady slopes, which is consistent with the results of this study.
In mountainous areas, altitude is the primary factor driving spatial heterogeneity. Soil nutrient levels and stoichiometry vary significantly between higher and lower altitudes due to differences in climatic conditions, precipitation patterns, vegetation, and microbiome composition (Jeyakumar et al., 2020; Chang-ming et al., 2023). In this study, altitude was identified as the main factor influencing soil phosphorus content and the C:P ratio. Numerous studies have demonstrated that in dry-hot valleys, increasing altitude results in reduced dry-hot winds and higher precipitation, leading to a steady accumulation of soil organic carbon, nitrogen, and phosphorus, and ultimately enhancing soil fertility (Chunming et al., 2003; Mullen, 2011; Peng et al., 2011). However, previous studies have not thoroughly examined how soil nutrient changes with altitude differ between sunny and shady slopes. In this study, we found that soil nutrient levels responded differently to altitude depending on slope aspect. On sunny slopes, soil organic carbon exhibited a significant increasing trend with altitude, aligning with findings from other research in dry-hot valleys (Chunming et al., 2003; Mullen, 2011; Peng et al., 2011), In contrast, on shady slopes, soli phosphorus decreased with increasing altitude, indicating a reduction in soil nutrient levels. The levels of soil carbon, nitrogen, and phosphorus are influenced by various processes, including nutrient input, mineralization, immobilization, and leaching (Johnson et al., 1998; Wang et al., 2001; Soon and Arshad, 2002). As altitude increases, higher precipitation promotes vegetation growth and enhances soil organic matter accumulation (Chang-ming et al., 2024). However, unlike soil carbon and nitrogen, soil phosphorus primarily originates from the parent material rather than SOM (Tian et al., 2010). At higher altitudes, increased precipitation may lead to greater nutrient leaching, which in turn reduces soil phosphorus levels (Maojie, 2011; Liu et al., 2019). A similar decline in soil phosphorus with increasing altitude has also been observed in other studies on dry-hot valleys (Xueju, 2005; Zhen-heng and Yuan-bo, 2018). Similarly, another study found that soil nutrient levels responded differently to altitude between sunny and shady slopes, with soil nitrogen increasing with altitude on sunny slopes but decreasing on shady slopes (Zhen-heng and Yuan-bo, 2018).
Stoichiometric homeostasis is a key parameter in ecological stoichiometry (Zhou et al., 2024). Plants with strong stoichiometric homeostasis are relatively conservative in nutrient use, whereas those with weaker homeostasis can flexibly use nutrients (Yu et al., 2010). Thus, the level of stoichiometric homeostasis reflects plant ecological adaptation mechanisms (Wirtz and Kerimoglu, 2016; Peng et al., 2017). In this study, M. alba exhibited strong homeostasis, as no significant correlation was found between the nutrient contents and stoichiometry ratios of M. alba leaves and soil. Additionally, M. alba leaves and soil responded differently to slope aspects and altitude. Moreover, the differences in nutrient content and stoichiometry between M. alba and soil were not constrained within a specific range. The 1/H calculation results indicated that M. alba was strictly homeostatic. These findings clearly support our expectation that plants in resource-limited environments, such as dry-hot valleys, exhibit strong stoichiometric homeostasis. Plants employ various metabolic and physiological mechanisms to maintain stable nutrient levels when nutrients limit their growth. The level of homeostatic flexibility largely depends on how effectively they use these limited resources (Hessen et al., 2004; Peng et al., 2017; Hong-bo et al., 2021; Su and Shangguan, 2022). Previous studies have reported that plants in dry-hot valleys can maintain stoichiometric stability by increasing nutrient absorption and resorption, which may lead to a decoupling of plant and soil stoichiometry (Jingwen et al., 2020). Plants with stronger stoichiometric homeostasis are better adapted to environmental changes (Chen et al., 2016; Peng et al., 2017; Wei et al., 2021). Our study demonstrated that M. alba, with its strict stoichiometric homeostasis, is well-equipped to thrive in a dry-hot environment.
The results of this study support the expectation that plants in dry-hot valleys exhibit strong stoichiometric homeostasis to cope with resource-limiting environments. Although the total nutrient level in the study area was at a medium level compared to the Chinese national average, the growth of M. alba was limited by nutrient availability, particularly nitrogen. M. alba maintained strict stoichiometric homeostasis on both sunny and shady slopes, despite significantly better nutrient conditions on the shady slope. This strict stoichiometric homeostasis represents an adaptive strategy of M. alba to the dry-hot valley, and its strength led to a decoupling of nutrient content and stoichiometry between M. alba leaves and the soil.
The datasets presented in this article are not readily available due to restriction. Requests to access the datasets should be directed to OTkwMjIxQGNkbnUuZWR1LmNu.
HG: Writing – original draft. SL: Data curation, Writing – original draft. SC: Methodology, Writing – original draft. YL: Software, Writing – original draft. JZ: Resources, Writing – review & editing. GC: Resources, Writing – review & editing. XW: Supervision, Writing – review & editing.
The author(s) declare that financial support was received for the research and/or publication of this article. This research was funded by the Sichuan Science and Technology Program (2023NSFSC0761), the China Agriculture Research System, the earmarked fund for CARS-18,and the Chengdu Normal University Research Program (CS19ZDZ02).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Bale, C. L., Williams, J. B., Charley, J. L. (1998). The impact of aspect on forest structure and floristics in some Eastern Australian sites. For. Ecol. Manage. 110, 363–377. doi: 10.1016/s0378-1127(98)00300-4
Batiha, G. E.-S., Al-Snafi, A. E., Thuwaini, M. M., Teibo, J. O., Shaheen, H. M., Akomolafe, A. P., et al. (2023). Morus alba: a comprehensive phytochemical and pharmacological review. Naunyn-schmiedeberg’s Arch. Pharmacol. 396, 1399–1413. doi: 10.1007/s00210-023-02434-4
Bertrand, I., Viaud, V., Daufresne, T., Pellerin, S., Recous, S. (2019). Stoichiometry constraints challenge the potential of agroecological practices for the soil C storage. A review. Agron. Sustain. Dev. 39, 16. doi: 10.1007/s13593-019-0599-6
Bui, E. N., Henderson, B. L. (2013). C:N:P stoichiometry in Australian soils with respect to vegetation and environmental factors. Plant Soil 373, 553–568. doi: 10.1007/s11104-013-1823-9
Chang-ming, R., Guo-Yong, T., Shou-Kang, D., Chun-Hua, Z., Chao-Rong, O., Kong, L., et al. (2023). Stoichiometry of soil carbon,nitrogen,phosphorus and enzyme activities at various elevations in the Dry⁃Hot Valley of the Jinsha River. Southwest China J. Agric. Sci. 36, 2464–2472. doi: 10.16213/j.cnki.scjas.2023.11.018
Chang-ming, R., Meng, W., Yong-Yu, S., Shou-Kang, D., Guo-Yong, T., Chun-hua, Z., et al. (2024). Characteristics of soil fertility and the influencing factors in the hot-dry valley of jinsha river. Chin. J. Soil Sci. 55, 93–101. doi: 10.19336/j.cnki.trtb.2022092904
Changming, R., Shoukang, D., Yongyu, S., Chunhua, Z., Chaorong, O., Meng, W., et al. (2022). Stoichiometry of soil carbon,Nitrogen and phosphorus and soil enzyme activity at various reaches of the dry-hot valley of jinsha river. Res. Environ. Sci. 35, 2486–2494. doi: 10.13198/j.issn.1001-6929.2022.07.21
Chen, Z. Q., Chen, Z. B., Yan, X. Y., Bai, L. Y. (2016). Stoichiometric mechanisms of Dicranopteris dichotoma growth and resistance to nutrient limitation in the Zhuxi watershed in the red soil hilly region of China. Plant Soil 398, 367–379. doi: 10.1007/s11104-015-2670-7
Chunming, W., Weikai, B., Jianzhong, C., Hui, S., Jiasui, X. (2003). Profile characteristics and nutrients of dry cinnamon soils in the dry valley of the Upper Minjiang River. Chin. J. Appl. Environ. Biol. 9, 230–234. Available online at: https://cstr.cn/32241.14.CJAEBiology.998.
Duan, X. W., Han, X., Hu, J. M., Feng, D. T., Rong, L. (2017). A novel model to assess soil productivity in the dry-hot valleys of China. J. Mountain Sci. 14, 705–715. doi: 10.1007/s11629-016-4147-4
Elser, J. J., Dobberfuhl, D. R., Mackay, N. A., Schampel, J. H. J. B. (1996). Organism size, life history, and N: P stoichiometry: toward a unified view of cellular and ecosystem processes. BioScience 46, 674–684. doi: 10.2307/1312897
Elser, J. J., Fagan, W. F., Kerkhoff, A. J., Swenson, N. G., Enquist, B. J. (2010). Biological stoichiometry of plant production: metabolism, scaling and ecological response to global change. New Phytol. 186, 593–608. doi: 10.1111/j.1469-8137.2010.03214.x
Elser, J. J., Sterner, R. W., Gorokhova, E., Fagan, W. F., Markow, T. A., Cotner, J. B., et al. (2000). Biological stoichiometry from genes to ecosystems. Ecol. Lett. 3, 540–550. doi: 10.1046/j.1461-0248.2000.00185.x
Enwezor, W. O. (1976). The mineralization of nitrogen and phosphorus in organic materials of varying C:N and C:P ratios. Plant Soil 44, 237–240. doi: 10.1007/BF00016972
Fan, H. B., Wu, J. P., Liu, W. F., Yuan, Y. H., Hu, L., Cai, Q. K. (2015). Linkages of plant and soil C:N:P stoichiometry and their relationships to forest growth in subtropical plantations. Plant Soil 392, 127–138. doi: 10.1007/s11104-015-2444-2
Fangyan, L., Hua, Z., Jipu, S., Xiaoming, C. (2007). Characteristics of plant communities and their soil fertilities in dry-hot valley of yuanjiang county, Yunnan, China. Chin. J. Appl. Environ. Biol. 13, 782–787. Available online at: https://qikan.cqvip.com/Qikan/Article/Detail?id=26228403.
Guan, J. (2022). Comparative study on soil nitrogen transformation between dry-hot valley and non-dry-hot valley (Kunming, China: Yunnan University). Master.
Guo, H. (2014). Spatial pattern of carbon and nitrogen at the core area of the arid valley in the upper Minjiang River (Chengdu, China: Sichuan Agricultural University). Master.
Güsewell, S. (2004). N:P ratios in terrestrial plants: variation and functional significance. New Phytol. 164, 243–266. doi: 10.1111/j.1469-8137.2004.01192.x
Han, W. X., Fang, J. Y., Guo, D. L., Zhang, Y. (2005). Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytol. 168, 377–385. doi: 10.1111/j.1469-8137.2005.01530.x
Han, W. X., Fang, J. Y., Reich, P. B., Woodward, F. I., Wang, Z. H. (2011). Biogeography and variability of eleven mineral elements in plant leaves across gradients of climate, soil and plant functional type in China. Ecol. Lett. 14, 788–796. doi: 10.1111/j.1461-0248.2011.01641.x
Hang, Y., Ruoyun, G., Wenjia, Y., Liusheng, Y., Songyang, L., Yongming, L., et al. (2022). Carbon, nitrogen, and phosphorus contents of leaf, root, and soil and their relationships in dominant herbaceous plants in dry-hot valley. Chin. J. Appl. Environ. Biol. 28, 727–735. doi: 10.19675/j.cnki.1006-687x.2021.01008
Hessen, D. O., Agren, G. I., Anderson, T. R., Elser, J. J., De Ruiter, P. C. (2004). Carbon, sequestration in ecosystems: The role of stoichiometry. Ecology 85, 1179–1192. doi: 10.1890/02-0251
Hong-bo, L., Yun-Lei, X., Zhi-Xiang, Y., Zheng-Ping, L., Yun-Xiang, D., Huan-cheng, M. (2021). Eco-stoichiometric characteristics of C, N and P in the leaves of typical plants in dry-hot valley of Jinsha river. J. Northwest Forestry Univ. 36, 10–16. doi: 10.3969/j.issn.1001-7461.2021.03.02
Jeyakumar, S. P., Dash, B., Singh, A. K., Suyal, D. C., Soni, R. (2020). “Nutrient Cycling at Higher Altitudes,” in Microbiological Advancements for Higher Altitude Agro-Ecosystems & Sustainability. Eds. Goel, R., Soni, R., Suyal., D. C. (Springer Singapore, Singapore), 293–305.
Jianfeng, Z., Baoping, S., Hubo, G., Yan, H., Li, Z., Haojie, S. (2016). Evaluation on effects of soil improvement for six typical forest types in dry-hot valley. Sci. Soil Water Conserv. 14, 95–102. doi: 10.16843/j.sswc.2016.02.013
Jingwen, H., Ying, L., Hang, Y., Jianzhao, W., Yu, C., Yongming, L., et al. (2020). Nutrient reabsorption efficiency of dominant shrubs in dry-hot valley and its C∶N∶P stoichiometry. J. Beijing Forestry Univ. 42, 18–26. doi: 10.12171/j.1000-1522.20190185
Johnson, D. W., Hanson, P. J., Todd, D. E., Susfalk, R. B., Trettin, C. F. (1998). Precipitation change and soil leaching: Field results and simulations from Walker Branch Watershed, Tennessee. Water Air Soil pollut. 105, 251–262. doi: 10.1023/a:1005039711434
Klemmedson, J. O., Wienhold, B. J. (1992). Nitrogen mineralization in soils of a chaparral watershed in arizona. Soil Sci. Soc. America J. 56, 1629–1634. doi: 10.2136/sssaj1992.03615995005600050049x
Lei Shan-Yu, G. Y.-F., Zhang, Y.-J., Ye, T.-H., Yang, P., Yu, W., Yan, F.-F., et al. (2022). Community Structure Analysis of nirS Denitrifying Bacteria in Farmland Soil at Different Altitudes in the Dry-hot Valley of Panzhihua. Chin. J. Soil Sci. 6), 1368–1375. doi: 10.19336/j.cnki.trtb.2021060204
Lin, Y., Chen, A., Yan, S., Rafay, L., Du, K., Wang, D., et al. (2019). Available soil nutrients and water content affect leaf nutrient concentrations and stoichiometry at different ages of Leucaena leucocephala forests in dry-hot valley. J. soils sediments 19, 511–521. doi: 10.1007/s11368-018-2029-9
Liu, J., Bi, X. Q., Ma, M. T., Jiang, L. H., Du, L. F., Li, S. J., et al. (2019). Precipitation and irrigation dominate soil water leaching in cropland in Northern China. Agric. Water Manage. 211, 165–171. doi: 10.1016/j.agwat.2018.09.053
Liu, Q., Chen, Z. M., Wang, S. L., Liang, T. T., Gao, Z. Y., Gao, Z. Y., et al. (2024). Changes in soil hydrological retention properties and controlling factors on shaded and sunny slopes in semi-arid alpine woodlands. Forests 15, 13. doi: 10.3390/f15071136
Makino, W., Cotner, J. B., Sterner, R. W., Elser, J. J. (2003). Are bacteria more like plants or animals?: Growth rate and resource dependence of bacterial C: N: P stoichiometry. Funct. Ecol. 17, 121–130. doi: 10.1046/j.1365-2435.2003.00712.x
Maojie, T. (2011). Spatial differentiation of soil properties in the dry valley along a river’s length of the hengduan mountains. Res. Soil Water Conserv. 18, 19–23. Available online at: http://stbcyj.paperonce.org/oa/DArticle.aspx?type=view&id=20110305.
Mingqin, O. (1996). The main measures for high-yield sericulture along the Jinsha River. Sichuan sericulture 02, 39–40. Available online at: https://kns.cnki.net/kcms2/article/abstract?v=SmerkCJHUJmiADZQikKvgbz7SBaIAEBeDIgPUp30Gwc2Td2sOzAtL31Wu3M7slFTqLZPPyqw7T-3iOJimxv7dcFt7Uunf9sx5ysfXYZ9OY4KRCYtEWkC761-R1lP6_lIjpj0VUTg0Q2NVOv9qYH1cam1uBFU508rmGsNcUU3a3I3zEJzu96TgJ0LeDDG_O6I&uniplatform=NZKPT&language=CHS.
Mullen, R. W. (2011). “Nutrient Cycling in Soils: Nitrogen,” in Soil Management: Building a Stable Base for Agriculture. Eds. Hatfield, J. L., Sauer, T. J. (Amer Soc Agronomy, Madison), 67–78.
Nan, L. (2016). Spatial heterogeneity of soil nutrient in the dry valley of Minjiang River (Chengdu, China: Bachelor Bachelor, Sichuan Normal University).
Pan, L. D., Shi, D. M., Jiang, G. Y., Xu, Y. (2024). Impacts of different management measures on soil nutrients and stoichiometric characteristics for sloping farmland under erosive environments in the Three Gorges Reservoir Area, China. Soil Tillage Res. 244, 15. doi: 10.1016/j.still.2024.106173
Peng, L., Zhanbin, L., Yu, Z. (2011). Effect of different elevation on soil physical-chemical properties and erodibility in dry-hot valley. Bull. Soil Water Conserv. 31, 103–107. doi: 10.13961/j.cnki.stbctb.2011.04.034
Peng, Y. F., Li, F., Zhou, G. Y., Fang, K., Zhang, D. Y., Li, C. B., et al. (2017). Linkages of plant stoichiometry to ecosystem production and carbon fluxes with increasing nitrogen inputs in an alpine steppe. Global Change Biol. 23, 5249–5259. doi: 10.1111/gcb.13789
Persson, J., Fink, P., Goto, A., Hood, J. M., Jonas, J., Kat, S. (2010). To be or not to be what you eat: regulation of stoichiometric homeostasis among autotrophs and heterotrophs. Oikos 119, 741–751. doi: 10.1111/j.1600-0706.2009.18545.x
Qin, Y. Y., Feng, Q., Adamowski, J. F., Zhu, M., Zhang, X. F. (2021). Community level response of leaf stoichiometry to slope aspect in a montane environment: A case study from the Central Qilian Mountains, China. Global Ecol. Conserv. 28, 10. doi: 10.1016/j.gecco.2021.e01703
Rastetter, E. B., Shaver, G. R. (1992). A model of multiple-element limitation for acclimating vegetation. Ecology 73, 1157–1174. doi: 10.2307/1940666
Redfield, A. C. (1958). The biological control of chemical factors in the environment. Am. Scientist 46, 205–221. Available online at: https://www.webofscience.com/wos/woscc/full-record/WOS:A1958WE06000001.
Reich, P. B., Oleksyn, J. (2004). Global patterns of plant leaf N and P in relation to temperature and latitude. Proc. Natl. Acad. Sci. United States America 101, 11001–11006. doi: 10.1073/pnas.0403588101
Sharma, C. M., Baduni, N. P., Gairola, S., Ghildiyal, S. K., Suyal, S. (2010). Effects of slope aspects on forest compositions, community structures and soil properties in natural temperate forests of Garhwal Himalaya. J. Forestry Res. 21, 331–337. doi: 10.1007/s11676-010-0079-y
Sheng, B., Shiqun, H., Gang, L., Xiaofen, W. (1999). Discussion on the “Three highs” Sericulture in the jinsha river dry and hot valley area. Sichuan sericulture 03, 40–42. Available online at: https://kns.cnki.net/kcms2/article/abstract?v=SmerkCJHUJlG5szUPJMt1jWOc0cCzYlvsVQpe6mD0C1XT1wX0XbpjlNw5bIoRR3va-sqrDVZ0W9Ecp9foTGPPIy2Sn9SO6ZlocKEoCigTeeJS5G61t7X5NuY3tgXKLVQY7egC7ciJro7BLwpeFIfhSGqFyGBGRwuR3WgqN4WPZTnyCc8_6_UCxf33o7e_I6W&uniplatform=NZKPT&language=CHS.
Shoukang, D., Guoyong, T., Yungen, L., Chenyu, L., Yingjie, X., Changming, R., et al. (2022). Plant diversity in various sections of Jinsha River dry-hot valley under different site environments. J. Zhejiang A&F Univ. 39, 742–749. doi: 10.11833/j.issn.2095-0756.20210572
Sistla, S. A., Schimel, J. P. (2012). Stoichiometric flexibility as a regulator of carbon and nutrient cycling in terrestrial ecosystems under change. New Phytol. 196, 68–78. doi: 10.1111/j.1469-8137.2012.04234.x
Soon, Y. K., Arshad, M. A. (2002). Comparison of the decomposition and N and P mineralization of canola, pea and wheat residues. Biol. Fertility Soils 36, 10–17. doi: 10.1007/s00374-002-0518-9
Sterner, R. W., Elser, J. J. (2002). Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere. (Princeton: Princeton University Press).
Su, B. Q., Shangguan, Z. P. (2022). Stoichiometric homeostasis in response to variable water and nutrient supply in a Robinia pseudoacacia plant-soil system. J. Plant Ecol. 15, 991–1006. doi: 10.1093/jpe/rtac011
Sun, J. N., Gao, P., Li, C., Wang, R. J., Niu, X., Wang, B. (2019). Ecological stoichiometry characteristics of the leaf-litter-soil continuum of Quercus acutissima Carr. and Pinus densiflora Sieb. in Northern China. Environ. Earth Sci. 78, 20.1–20.13. doi: 10.1007/s12665-018-8012-3
Tan, G., Jiangrong, L., Jie, L., Weilie, Z., Jiarui, C. (2020). Soil nutrient and fertility of different slope directions in the Abies georgei var. smithii forest in Sejila Mountain. Acta Ecologica Sin. 40, 1331–1341. doi: 10.5846/stxb201812062674
Tian, H., Chen, G., Zhang, C., Melillo, J. M., Hall, C. A. (2010). Pattern and variation of C: N: P ratios in China’s soils: a synthesis of observational data. Biogeochemistry 98, 139–151. doi: 10.1007/s10533-009-9382-0
Wang, W. J., Chalk, P. M., Chen, D., Smith, C. J. (2001). Nitrogen mineralisation, immobilisation and loss, and their role in determining differences in net nitrogen production during waterlogged and aerobic incubation of soils. Soil Biol. Biochem. 33, 1305–1315. doi: 10.1016/s0038-0717(01)00034-7
Wei, Y. J., Dang, X. H., Wang, J., Gao, J. L., Gao, Y. (2021). Response of C:N:P in the plant-soil system and stoichiometric homeostasis of Nitraria tangutorum leaves in the oasis-desert ecotone, Northwest China. J. Arid Land 13, 934–946. doi: 10.1007/s40333-021-0019-z
Wirtz, K. W., Kerimoglu, O. (2016). Autotrophic stoichiometry emerging from optimality and variable co-limitation. Front. Ecol. Evol. 4. doi: 10.3389/fevo.2016.00131
Xie, W., Aichun, Z., Guantao, C., Jianhua, Z. (2024). National ecological mulberry industry development layout based on soil conservation. Newslett. Sericultural Sci. 44, 61–67. Available online at: https://kns.cnki.net/kcms2/article/abstract?v=SmerkCJHUJnOBLV4O4vsooBGgXTFBoP8Xwb7ZNmMo7Ehhz8v1G1frfdhG7N1SmR9Xpcx7Tqbqbdd0S6abY2JZzXj1egH0C4RXA9G3hgr5ftf-Czr7Gr3cG4BFVQQ98rH6e4ZJBIQMfSGsMJvB-4KI_j-KqXC3-yWI7YLs2vHNaEK1G-WHb5eUl9ml9wbnwOp&uniplatform=NZKPT&language=CHS.
Xueju, H. (2005). Spatial variability of soil quality in the dry valley along a river’s length of Hengduan Mountains (Chengdu, China: Sichuan University). Master.
Ya, T., Jiasui, X., Hui, S. (2004). Revisiting sustainable development of dry valleys in Hengduan Mountains Region. J. Mountain Sci. 1, 38–45. doi: 10.1007/BF02919358
Yanan, C., Lusha, M., Xiangru, Z., Jiajia, Y., Shaoshan, A. (2014). Ecological stoichiometry characteristics of leaf litter of Robinia pseudoacacia in the Loess Plateau of Shaanxi Province. Acta Ecologica Sin. 34, 4412–4422. doi: 10.5846/stxb201306101562
Yang, J., El-Kassaby, Y. A., Guan, W. (2020). The effect of slope aspect on vegetation attributes in a mountainous dry valley, Southwest China. Sci. Rep. 10, 16465. doi: 10.1038/s41598-020-73496-0
Yiting, G., Siying, Z., Ying, L., Wanling, S., Yongming, L. (2023). Nutrient distribution and ecological stoichiometry characteristics of shrub-grass layer vegetation and soil in the Jiangjia Gully,China. Mountain Res. 41, 824–835. doi: 10.16089/j.cnki.1008-2786.000790
Yu, Q., Chen, Q. S., Elser, J. J., He, N. P., Wu, H. H., Zhang, G. M., et al. (2010). Linking stoichiometric homoeostasis with ecosystem structure, functioning and stability. Ecol. Lett. 13, 1390–1399. doi: 10.1111/j.1461-0248.2010.01532.x
Yu, Q. A., Elser, J. J., He, N. P., Wu, H. H., Chen, Q. S., Zhang, G. M., et al. (2011). Stoichiometric homeostasis of vascular plants in the Inner Mongolia grassland. Oecologia 166, 1–10. doi: 10.1007/s00442-010-1902-z
Zechmeister-Boltenstern, S., Keiblinger, K. M., Mooshammer, M., Peñuelas, J., Richter, A., Sardans, J., et al. (2015). The application of ecological stoichiometry to plant-microbial-soil organic matter transformations. Ecol. Monogr. 85, 133–155. doi: 10.1890/14-0777.1
Zhang, J. H., Zhao, N., Liu, C. C., Yang, H., Li, M. L., Yu, G. R., et al. (2018). C:N:P stoichiometry in China’s forests: From organs to ecosystems. Funct. Ecol. 32, 50–60. doi: 10.1111/1365-2435.12979
Zhaoyang, Z., Juan, Q., Hongzhang, S., Xinfeng, Y., Liqiang, L. (2019). Effects of microtopography on germination layer soil factors in Armeniaca vulgaris Lam. in Daxigou. Acta Ecologica Sin. 39, 2168–2179. doi: 10.5846/stxb201803060446
Zheng, Y. (2010). Study on features of soil quality in dry-hot valley of Jinsha river (Xi’an, China: Xi’an University of Science and Technology).
Zhen-heng, Z., Yuan-bo, G. (2018). Study of soil nitrogen heterogeneity among different slopes in the semi-arid areas of upper reaches of the minjiang river. J. Anhui Agric. Sci. 46, 98–102. doi: 10.13989/j.cnki.0517-6611.2018.23.031
Zhifeng, L., Feng, G., Ping, Z., Hanbiao, O., Junxue, H., Shuoxing, W., et al. (2024). Characteristics of different forest stands of community diversity and soil physicochemical properties and enzyme activities in albizia odoratissima forests in the dry and hot river valleys of guangxi. Chin. J. Trop. Agric. 44, 31–39. doi: 10.12008/j.issn.1009-2196.2024.10.005
Zhou, A., Ge, B., Chen, S., Kang, D., Wu, J., Zheng, W., et al. (2024). Leaf ecological stoichiometry and anatomical structural adaptation mechanisms of Quercus sect. Heterobalanus in southeastern Qinghai–Tibet Plateau. BMC Plant Biol. 24, 325. doi: 10.1186/s12870-024-05010-x
Keywords: Morus alba, soil, slope, altitude, stoichiometric homeostasis
Citation: Guo H, Luo S, Chen S, Li Y, Zhang J, Chen G and Wang X (2025) Stoichiometric homeostasis of Morus alba in the dry-hot valley. Front. Plant Sci. 16:1520936. doi: 10.3389/fpls.2025.1520936
Received: 31 October 2024; Accepted: 11 February 2025;
Published: 17 March 2025.
Edited by:
Muthusamy Ramakrishnan, Nanjing Forestry University, ChinaReviewed by:
Weiguo Zhao, Jiangsu University of Science and Technology, ChinaCopyright © 2025 Guo, Luo, Chen, Li, Zhang, Chen and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xie Wang, d2FuZ3hpZWNoYW5nZGVAaG90bWFpbC5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.