
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Plant Sci. , 20 March 2025
Sec. Functional Plant Ecology
Volume 16 - 2025 | https://doi.org/10.3389/fpls.2025.1503627
 Feng Zhu1,2,3
Feng Zhu1,2,3 Jing Yuan1,3*Zeying Hou1,3Xia Guo3,4Wanxue Liao1,3
Jing Yuan1,3*Zeying Hou1,3Xia Guo3,4Wanxue Liao1,3 Shenglin Yang1,2,3
Shenglin Yang1,2,3 Zhaosheng Chu1,2,3*
Zhaosheng Chu1,2,3*The littoral width of lakeshores is crucial for maintaining and promoting plant diversity. However, it remains unclear how changes in seasonal water level affect littoral widths by regulating plant diversity and soil nutrient content. This study selected three elevation ranges in the lakeshore of Erhai: supralittoral, eulittoral, and infralittoral. We explored the effects of hydrological changes on littoral widths and their potential relationships by analyzing seasonal differences in plant communities and soil physicochemical properties during an extremely drought year. Our results indicated that the most significant seasonal differences in diversity indices, biomass, and soil physicochemical properties were observed in the eulittoral, followed by the infralittoral and supralittoral. The niche breadths of perennials was significantly decreased by 44.4% and the width of the eulittoral was significantly decreased by 48.6% during the winter. Generalized Additive Models (GAMs) were applied to analyze the elevation distribution ranges of dominant species. The results revealed that species with monotonically increasing distributions had the widest niche breadths, followed by symmetric unimodal species, while monotonically decreasing species exhibited the narrowest. Structural equation modeling revealed a positive and significant correlation between flooding days and soil water content and pH, and a negative correlation with plant parameters (species number, biomass, and coverage). Moreover, plant parameters showed a significant positive correlation with plant diversity. Importantly, plant diversity and soil nutrients were significantly positively correlated with littoral widths, suggesting their key roles in influencing littoral widths. This study highlights the significant impact of hydrological seasonal changes on the littoral widths of lakeshore zones, providing valuable guidance for managing wetland water levels in response to extreme drought events.
The lakeshore zone is a transitional area between aquatic and terrestrial ecosystems and is a crucial component of the lake ecosystem (Ostendorp, 2004). It serves as a terrestrial-lake ecotone, being a sensitive area susceptible to water level fluctuations, and is also critical for maintaining biodiversity (Wang et al., 2022; Zhang et al., 2015). Lakeshore plants play a crucial role as a primary producer in preventing soil erosion, intercepting pollutants, and providing diverse habitats (Li et al., 2019; Zhang et al., 2021). Seasonal water level changes significantly influence the distribution of lakeshore plants, exhibiting distinct vertical distribution characteristics (Gaberscik et al., 2018; Zheng et al., 2022). Aquatic plants in the infralittoral are adapted to long-term flooding (Baschuk et al., 2012), hygrophytes in the eulittoral are adapted to intermittent flooding (Garssen et al., 2017; Ojdanic et al., 2023), and xerophytes in the supralittoral are adapted to drought conditions (Stroh et al., 2008).
Water level changes are one of the driving factors shaping the landscape patterns of wetlands and a determining factor influencing the spatiotemporal distribution of plant communities (Gathman et al., 2005; Hu et al., 2018). Wetland hydrological processes significantly influence the composition, diversity, distribution width, and area of plant communities (Gaberscik et al., 2018; Wang et al., 2015; You et al., 2015; Zheng et al., 2022). Such as long-time droughts or floods may lead to decreased numbers and diversity of plant species in wetlands, or even the formation of a single dominant species community (Gaberscik et al., 2020; Wassens et al., 2017). Secondly, the frequency and duration of wet-dry alternations significantly affect the growth and physiological characteristics of wetland plants, with frequent alternations increasing plant community diversity (Pollock et al., 1998; Zhang et al., 2022). In addition, fluctuations in the highest and lowest water levels determine the distribution width of plant communities and influence the habitat range of different plant communities (Chapin and Paige, 2013). Water level fluctuations directly or indirectly influence the seed germination and reproductive success of lakeshore plants by affecting soil water and nutrient content (Fu et al., 2018; Zhao et al., 2021). Therefore, wetland plant community structure and growth may vary significantly across elevation zones under different water level gradients.
The response of plant species to environmental gradients reflects adaptive adjustments in their niche (Hutchinson, 1957), which have received extensive attention in wetland research and management. The niche breadth of a species determines its ability to utilize different environmental gradients, thereby influencing its distribution range and competitive ability (Costa et al., 2018). The hydrology, soil, and plants are three important components of wetland ecosystems that interact and influence each other (Feng et al., 2020; Zhang et al., 2022). The constrained spatial extent of lakeshore zones results in the distribution and niche breadth of plants being particularly sensitive to variations in soil physicochemical properties, water levels, and elevation (Lou et al., 2018; Wang et al., 2022; Zheng et al., 2022). For example, larger inter-annual differences in water levels can alter wetland soil moisture and nutrient availability, thereby affecting plant diversity (Shen et al., 2020). Lakeshore zones are ideal habitats for studying changes in plant ecological behavior, as the significant environmental gradients and diverse habitats make the response of plant communities to environmental changes more intuitive and easier to measure (Chen et al., 2020; Duval et al., 2012). However, knowledge is limited on the relationship between plant niche breadth at different elevation ranges in the lakeshore zone and environmental gradients, especially in plateau lakes.
Lake Erhai, the second largest plateau lake in the Yunnan Province of China, serves multiple functions including agricultural irrigation, climate regulation, tourism and water supply (Wang et al., 2023). In recent years, due to eutrophication and the rapid development of agriculture and tourism, the stability of the Erhai lakeshore ecosystem has been seriously damaged (Li et al., 2020). In 2023, Lake Erhai experienced an extreme drought, leading to the revision of the statutory minimum operating water level from 1964.30 m to 1964.10 m (DBAPPG, 2023). Although many studies have focused on the effects of water level fluctuations on aquatic plant communities in Lake Erhai (Wen et al., 2023; Wu et al., 2023; Zhu et al., 2018), there is still a lack of research on the effects of plant communities in different elevation zones along the lakeshore. Effective water level management requires a comprehensive understanding of how water level fluctuations affect plant community distribution ranges and structure in the Erhai lakeshore zones.
This study conducted comprehensive surveys and analyses of plants and soils in three elevation zones along the lakeshore of Lake Erhai during the summer and winter of 2023, and collected daily water level data provided by the Erhai Administration Bureau. The questions addressed in this study are: 1) to clarify the distribution characteristics and dynamics patterns of plant communities across different elevation zones of the lakeshore; 2) to compare summer and winter differences in the niche breadths of dominant species and their responses to elevation; and 3) to reveal the relationships among hydrology, plants, and soil, and the mechanisms influencing littoral widths. This study aims to demonstrate how seasonal water level fluctuations affect plant diversity and littoral widths in the lakeshore zone, providing a theoretical basis for effective water level management to improve the total plant diversity of the Lake Erhai.
Lake Erhai (25°36′~25°58′ N, 100°06′~100°18′ E), located in Yunnan Province, China, is a faulted freshwater lake formed by crustal movement (Figure 1). The lake covers an area of 252 km², with average annual temperatures of 15.1°C. It has distinct wet and dry seasons, but precipitation is unevenly distributed, with more than 85% occurring during the rainy season from May to October (approximately 870 mm). A total of 117 tributaries flow into Lake Erhai, with legally authorized maximum and minimum operating water levels of 1966.0 m and 1964.3 m, respectively (Gong et al., 2023). Under the combined influence of reduced precipitation and artificial regulation since 2004, the time lag between water level and precipitation in Lake Erhai has become longer, with longer time intervals between the highest and lowest water levels, consequently altering the inundation time in the lakeshore zone (Wen et al., 2021). Lake Erhai is an important ecological area in China, characterized by high plant diversity and coverage, and rich biodiversity. The Cangshan and Erhai Nature Reserves were upgraded to national nature reserve in 1993 (Wang et al., 2023).
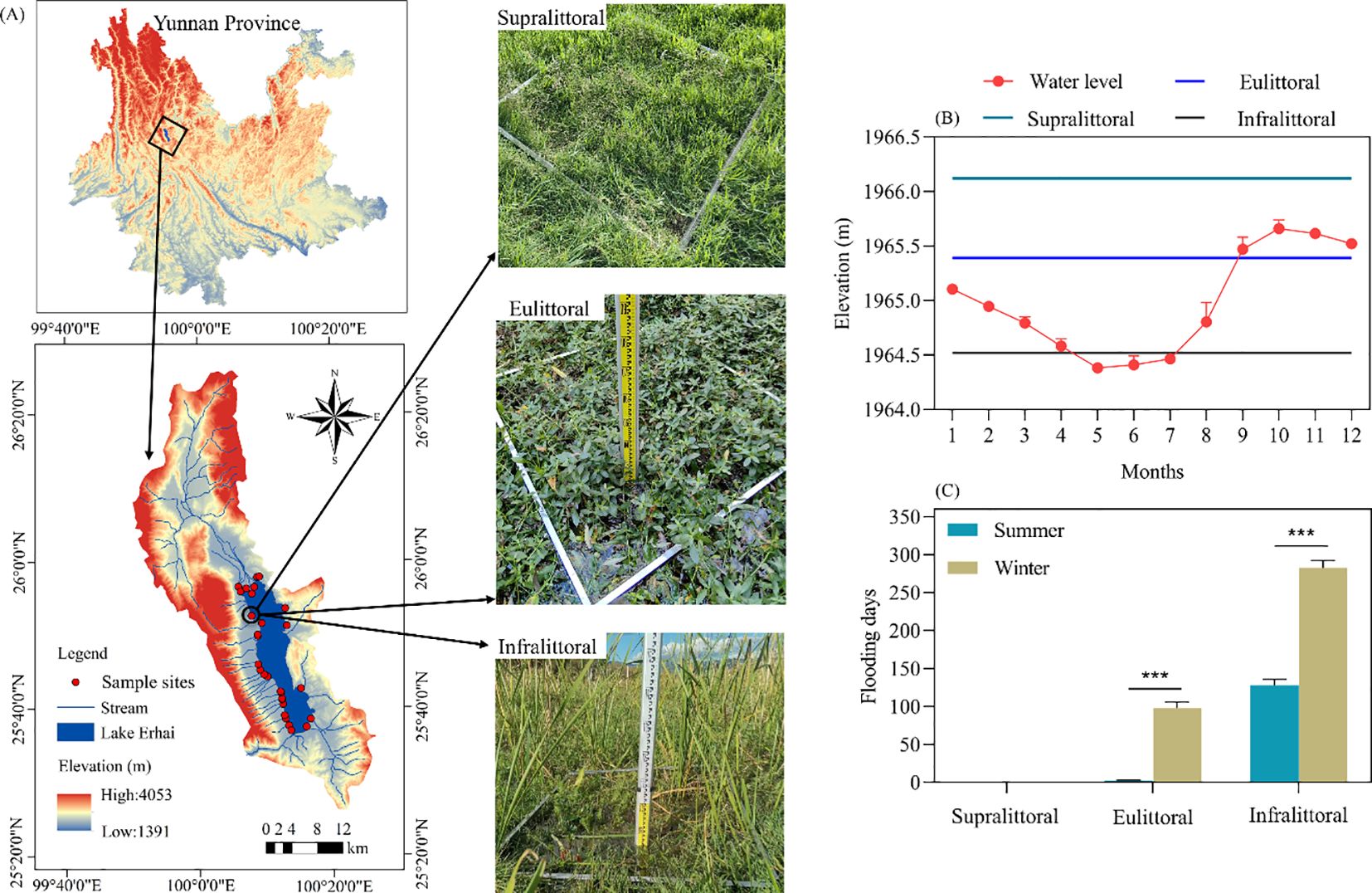
Figure 1. (A) Map of the study area and distribution of sample sites. (B) The monthly average water level of Lake Erhai in 2023 and the average elevations of the three elevation zones. (C) The difference in flooding days between summer and winter at three elevation zones. Values are means ± standard error. The significant differences are indicated by *** p < 0.001.
This study conducted plant surveys and soil sampling along the lakeshore zone of Lake Erhai in the summer (mid-July) and winter (mid-December) of 2023. We selected 29 fixed sample sites based on their accessibility and the distinct, high coverage of dominant plant communities. Each site was vertically divided into three zones based on the number of days of submergence: supralittoral (1965.77m-1967.37m), eulittoral (1965.06m-1965.74m), and infralittoral (1964.30m-1965.06m). In 2023, the average number of flooding days was 0 in the supralittoral, 98 in the eulittoral, and 283 in the infralittoral (Figure 1C). Three parallel quadrats (1m×1m) were randomly set up in each zone, and plant name, height, coverage, biomass (fresh weight), and numbers were recorded within each quadrat. The geographical coordinates of each quadrat were recorded using a portable GPS locator and combined with spray paint and red string markers for reference.
After harvesting the aboveground biomass in the quadrats using a sickle, three soil cores (0-20 cm) were collected diagonally using a soil auger. The soil samples from each zone were thoroughly mixed to form a composite sample and transported back to the laboratory for physicochemical properties analysis. The relative elevation above the water surface in each zone was measured using a level instrument and combined with the water level recorded on the survey day to calculate the elevation of each zone (Shen et al., 2019).
The soil samples were divided into three sub-samples for different analyses. One sub-sample was used to determine soil water content (SW) using fresh soil; one sub-sample was refrigerated at 4°C to determine ammonium nitrogen (NH4+-N) and nitrate nitrogen (NO3−-N); and the other sub-sample was freeze-dried, grind, and preserved by passing it through a 0.149-mm nylon sieve to determine total phosphorus (TP), C/N ratio, total nitrogen (TN), soil organic matter (SOM), and pH.
SW was determined using the thermostat drying method: dried at 105°C for 24 hours. pH was measured using a pH meter in a mixture with a soil-water ratio of 1:2.5. NH4+-N, NO3−-N and TP were determined by UV/visible spectrophotometer (UV-1900i). TN (%) and C/N ratio were determined with an elemental analyzer (Vario Macro Cube, Germany). SOM was analyzed by the potassium dichromate volumetric method under externally heated conditions (Ji, 2005).
At the species level, the seasonal variation of the top 30 dominant species were calculated based on the ordering of importance values (IV). At the diversity level, assessed by Patrick index (R), Shannon-Wiener index (H), Simpson Index (D), Pielou Evenness index (E), and dominance index (λ). The equations for all indices were calculated as follows Equations 1–6:
where S is the total number of species (species richness) recorded in each zone and Pi represents the relative abundance of ith species in each zone.
Plant species niche breadth and littoral widths were calculated using the Levins method from package spaa. Meanwhile, Generalized Additive Models (GAMs) were constructed to understand the distribution ranges of plants, and to assess plant responses to elevation changes. GAMs are more flexible than generalized linear models, to explore nonlinear relationships between independent and dependent variables, and perform well in spatial prediction (Elith et al., 2006). GAMs were performed using the mgcv package, employing a Gaussian distribution and nonlinear fitting of species abundance and elevation through a smoothing function. The smoothing parameter was automatically selected by generalized cross validation (GCV), which obtained a low GCV value, indicating a good fit of the model (Guisan et al., 2002). The spaa and mgcv packages were both used for analysis in R (version 4.3.2).
Principal component analysis (PCA) was performed to assess correlations between components and identify the main components associated with littoral widths. PCA is an unsupervised method that identifies principal components capturing the maximum variance in the dataset without predefined explanatory variable relationships (Lever et al., 2017). Structural equation modeling (SEM) was employed to explore the relationships among flooding days, soil nutrients, SW, soil pH, plant diversity, plant parameters (R, biomass, coverage), and littoral widths. SEM is a multivariate statistical analysis method that allows for the simultaneous examination of both direct and indirect relationships among multiple observed variables, thereby revealing hidden structural patterns within complex systems. For SEM construction, we ensured the key assumptions of the linearity of relationships among variables and sufficient sample size were met. Model fitting was completed by removing the observed variables based on the modification indices and confirming that the key fit metrics met the thresholds.
All data tested for significance were assessed for variance homogeneity and normal distribution, and data satisfied with normal distribution were tested using the independent sample t-test; otherwise, the Mann-Whitney U-test was used. All significance tests for the data were conducted using IBM SPSS 27.0 software. PCA and mapping were performed using Origin2023b and ArcMap 10.8. SEM was constructed and analyzed using IBM SPSS Amos 28 software.
A total of 110 species belonging to 40 families and 82 genera were recorded in the summer, with the highest species richness in the order of Poaceae (N=23, 20.9%), Asteraceae (N=16, 14.5%), Polygonaceae (N=7, 6.4%), and Cyperaceae (N=6, 5.5%). A total of 71 species belonging to 30 families and 58 genera were recorded in the winter, with the highest species richness in the order of Poaceae (N=18, 25.4%), Asteraceae (N=10, 14.1%), Polygonaceae (N=5, 7.0%), and Fabaceae (N=4, 5.6%). All species surveyed in this study were listed in Appendix 1.
Compared to summer, the importance values (IV) of dominant species showed seasonal differences across the three zones during winter (Figure 2). In the supralittoral, 7 dominant species showed an increase in IV, with Trifolium repens exhibiting the highest increase of 416.9%, while 10 species showed a decrease, with Cynodon dactylon exhibiting the highest decrease of 31.8%. In the eulittoral, 11 dominant species showed an increase in IV, with Alternanthera philoxeroides exhibiting the highest increase of 66.5%, while 13 species showed a decrease, with Ageratina adenophora exhibiting the highest decrease of 62.3%. In the infralittoral, both increased and decreased in IV were 11 dominant species, with Phragmites karka exhibiting the highest increase of 100% and Phragmites australis exhibiting the highest decrease of 40.2%.
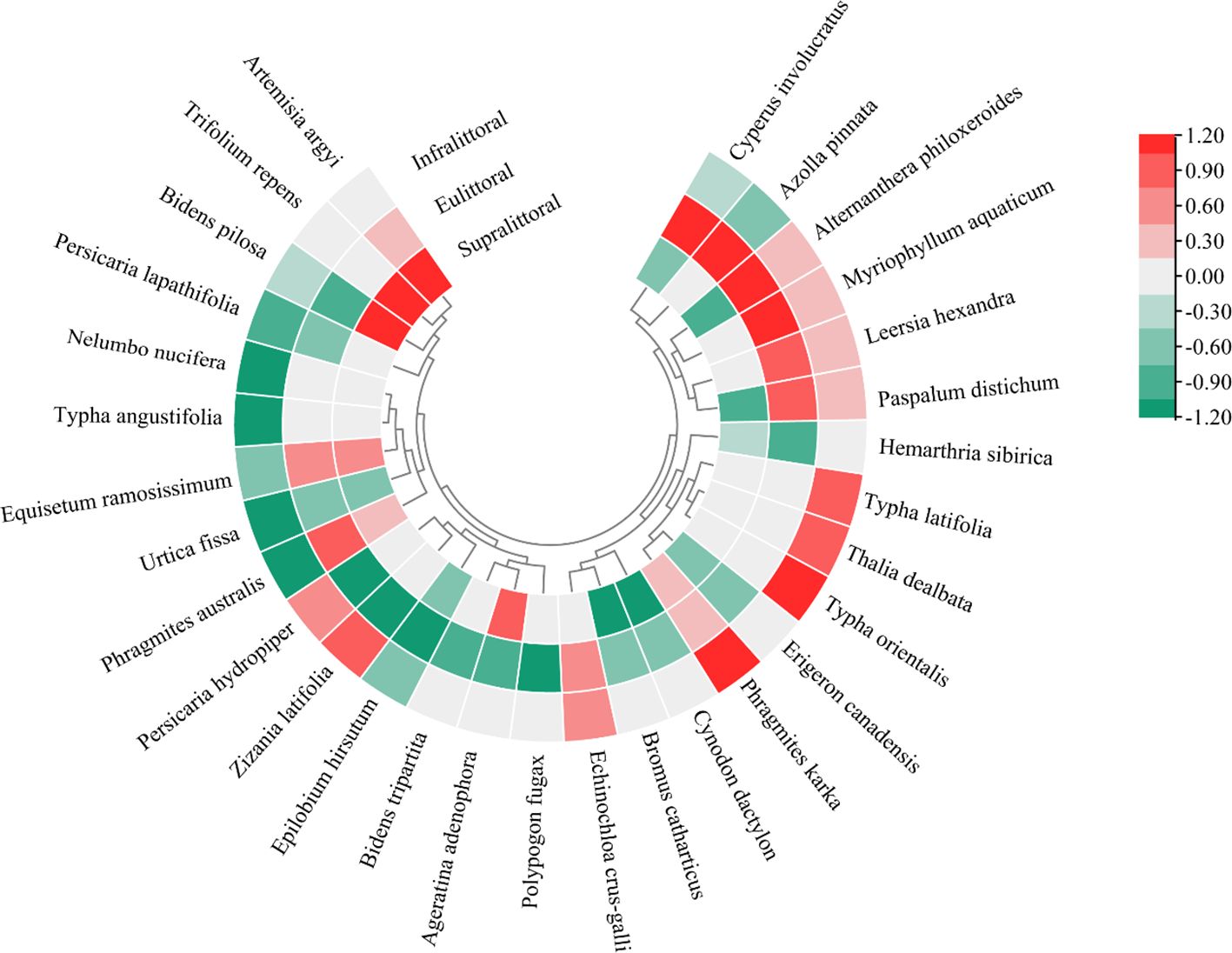
Figure 2. The seasonal differences in the importance values of the top 30 dominant species. Red indicates an increased value and green a decreased value.
Compared to summer, the diversity indices showed seasonal differences across the three zones during winter (Figure 3). Specifically, in the supralittoral, the Evenness index increased significantly by 31.0%. In the eulittoral, the Dominance index and Evenness index increased significantly by 46.3% and 64.5%, respectively, while the Simpson index, Shannon-Wiener index, and species richness decreased significantly by 48.3%, 56.8%, and 67.3%. In the infralittoral, the Shannon-Wiener index and species richness decreased significantly by 40.4% and 50.8%, respectively, while the Evenness index increased significantly by 43.0%. In summary, seasonal differences in diversity indices were most significant in the eulittoral, followed by the infralittoral and supralittoral.
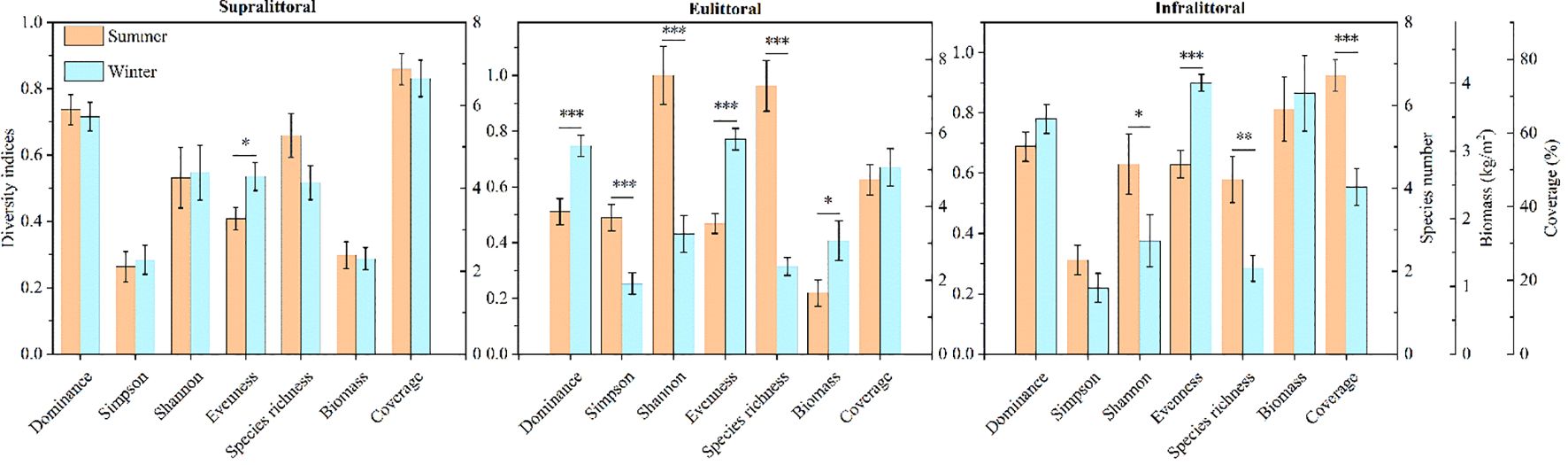
Figure 3. The seasonal differences in diversity indices, biomass and coverage across the three zones. Values are means ± SE. *P < 0.05, **P < 0.01, ***P < 0.001.
The biomass did not differ seasonally in the supralittoral and infralittoral, but increased significantly by 85.9% in the eulittoral. The coverage did not differ seasonally in the supralittoral and eulittoral, but decreased significantly by 40.1% in the infralittoral.
Seasonal comparisons of niche breadths showed significant differences among species (Figure 4A). The niche breadth of Alternanthera philoxeroides was largest in summer (21.39), but only 9.39 (decreased by 56.1%) in winter. This significant decrease indicates that rising water levels may be limiting the habitat of this species, affecting its dispersal and competitive ability. Similarly, other perennial herbaceous plants with high niche breadths in summer and significant decreases in winter include Cynodon dactylon, Phragmites australis, Urtica fissa, and Epilobium hirsutum. However, there was no significant seasonal difference in the niche breadth of the helophytes Zizania latifolia. This indicates that the habitat of this species is not limited by changes in water level and that it has strong adaptability. In summary, the mean niche breadth for perennial plants was 5.45 in the summer and 3.03 in the winter, showing a significant decrease of 44.4%. Most annual plants, such as Solanum nigrum, Polypogon fugax, and Rorippa palustris, showed a 100% decrease in niche breadth during winter.
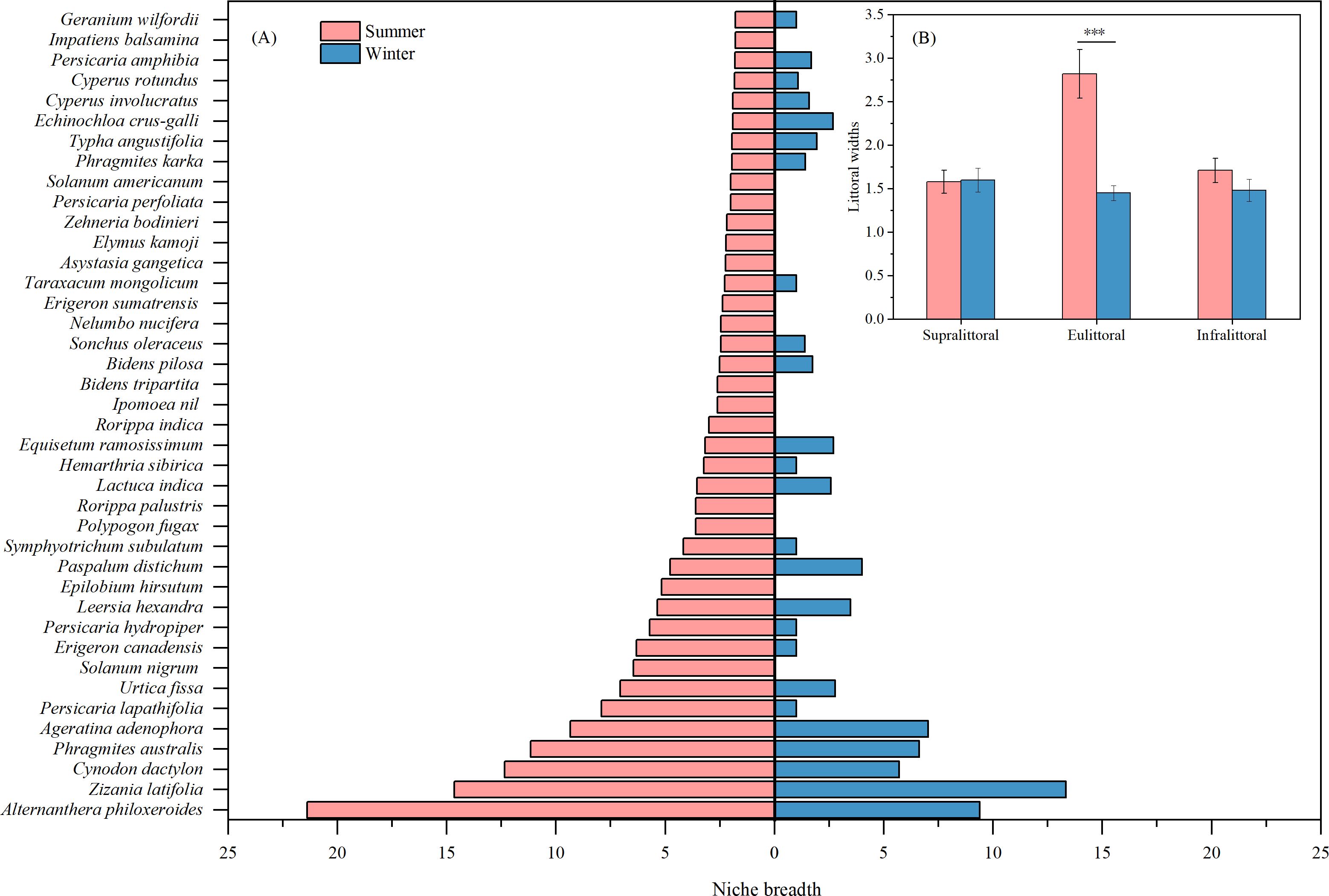
Figure 4. Seasonal differences in species niche breadths (A) and littoral widths (B). Values are means ± SE. ***P < 0.001.
There were no significant seasonal differences in littoral widths between the supralittoral and infralittoral (Figure 4B), suggesting that species composition and habitat conditions are relatively stable in both zones. However, the eulittoral exhibited a significant decrease in littoral width of 48.6% during winter (Figure 4B), probably caused by rising water levels that narrowed the habitat range in this zone.
The GAMs fitting was successful for the six dominant species (all P < 0.01, Table 1; Figure 5), exhibiting different elevation response curves. The response curves of Alternanthera philoxeroides and Leersia hexandra decreased with increasing elevation. Conversely, the response curve of Cynodon dactylon increased with increasing elevation. One species (Zizania latifolia) exhibited a monotonically decreasing response curve. Two species (Phragmites australis and Ageratina adenophora) exhibited symmetrical unimodal response curves.
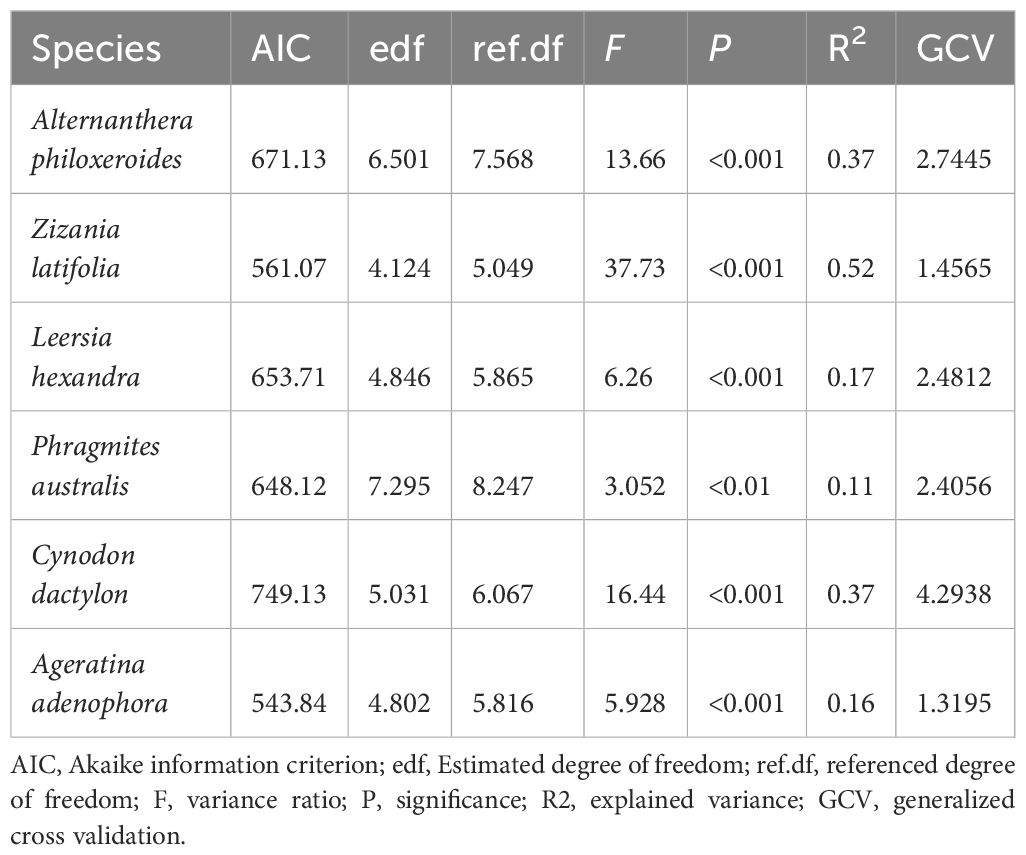
Table 1. Statistical parameters of the response curves (GAMs) to elevation for the top 6 species.
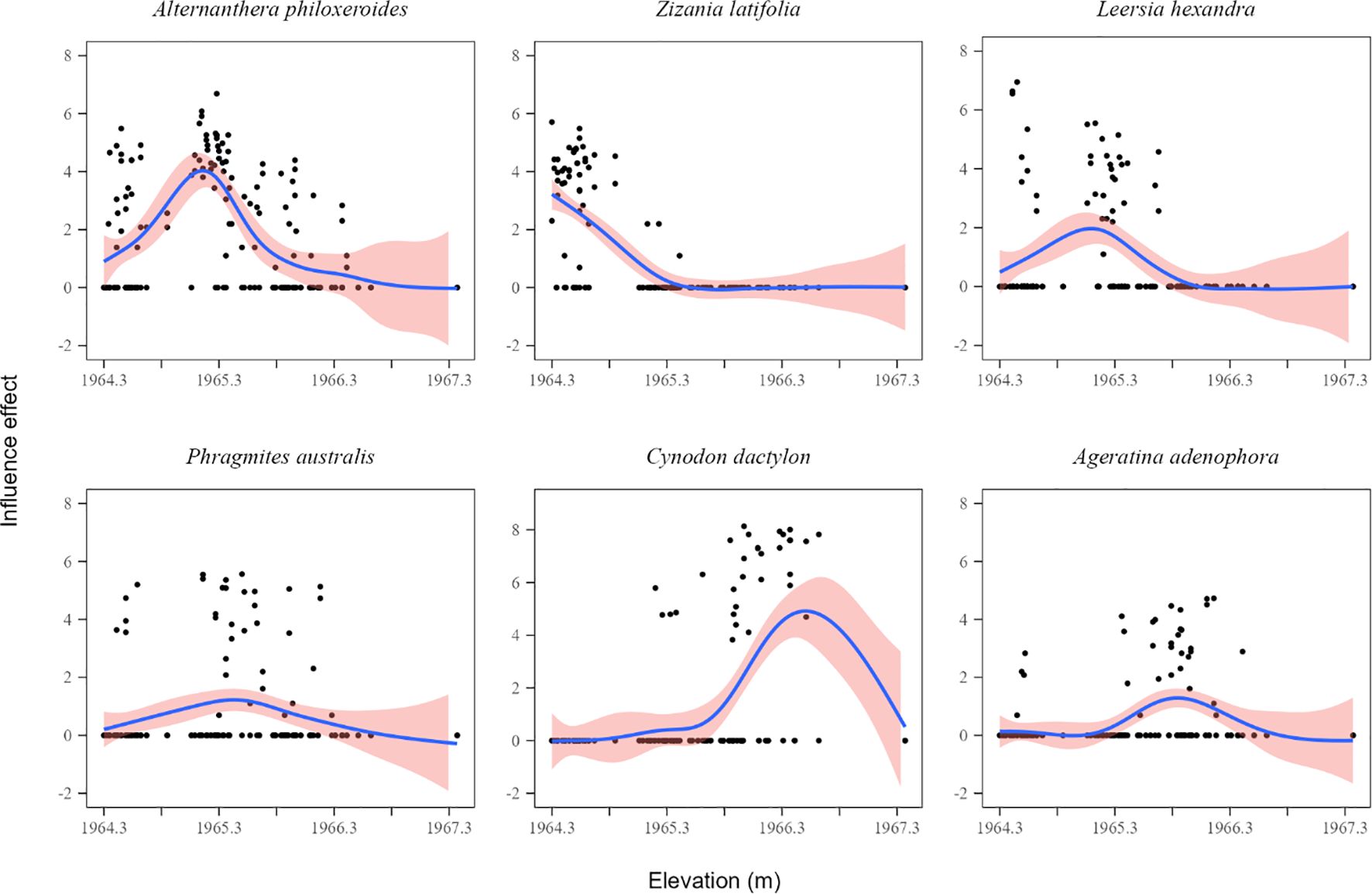
Figure 5. Response curves to elevation for the top 6 species. The red shaded areas indicate 95% confidence intervals.
There were significant seasonal changes in soil physico-chemical properties, but the differences varied across zones during winter (Figure 6). Specifically, in the supralittoral, SW, NO3−-N, and the C/N ratio increased significantly, while pH and TN decreased significantly, and there were no significant differences in NH4+-N, SOM, and TP. In the eulittoral, SW, NO3−-N, and the C/N ratio increased significantly, while pH, TP, and TN decreased significantly, and there were no significant differences in NH4+-N and SOM. In the infralittoral, SW and NH4+-N increased significantly, while pH decreased significantly, and there were no significant differences in NO3−-N, SOM, TP, TN, and the C/N ratio. In summary, the seasonal differences in soil physico-chemical properties were most significant in the eulittoral, followed by the supralittoral and infralittoral. SW and pH exhibited significant seasonal differences in all three zones.
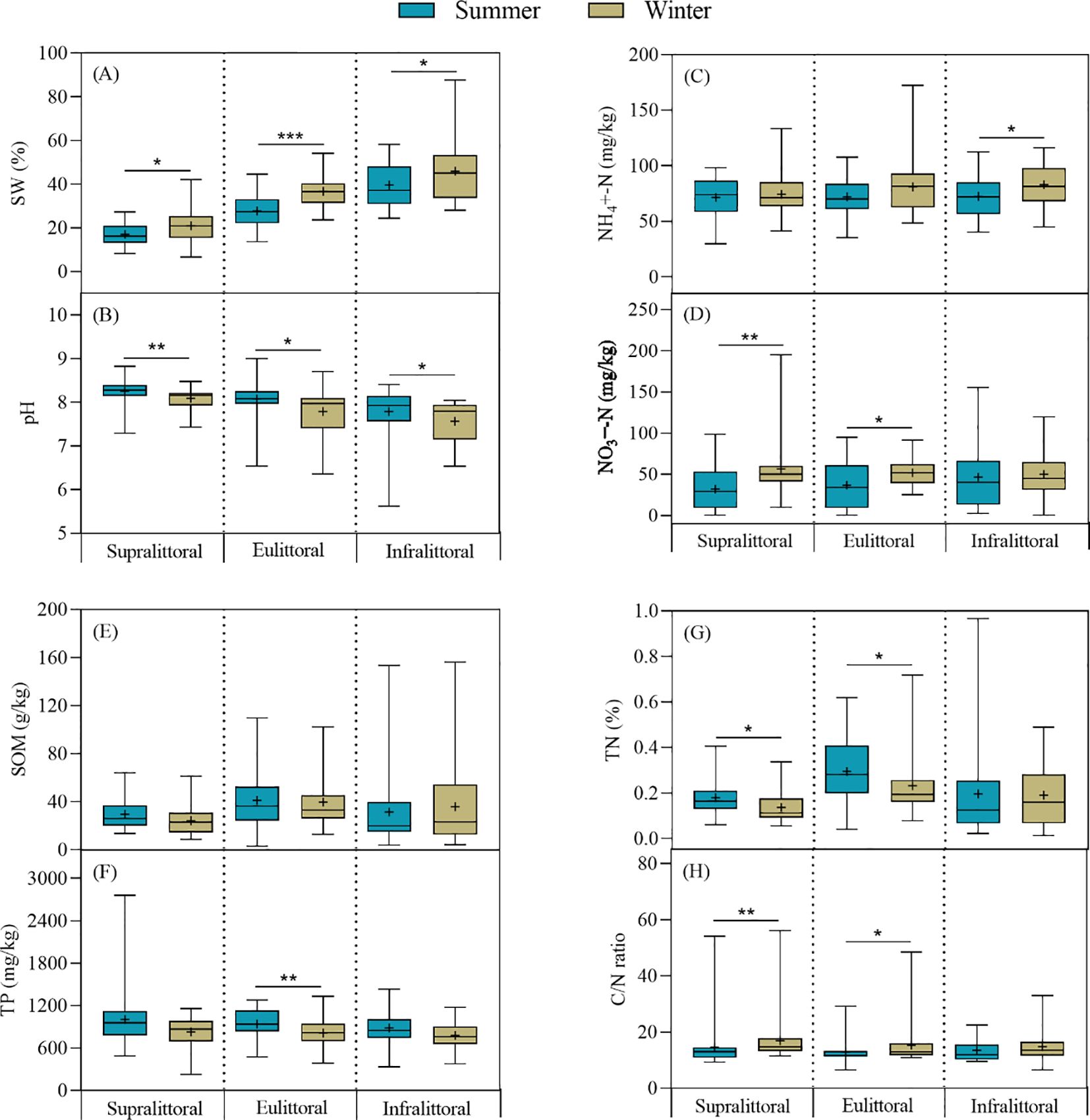
Figure 6. Seasonal differences in soil physico-chemical properties across three zones. (A) soil water content (SW), (B) pH, (C) ammonium nitrogen (NH4+-N), (D) nitrate nitrogen (NO3−-N), (E) soil organic matter (SOM), (F) total phosphorus (TP), (G) total nitrogen (TN), (H) C/N ratio. Values are mean ± SE, + indicates the mean. Note: * P < 0.05, ** P < 0.01, *** P < 0.001.
Principal component analysis determined the relationship among littoral widths, soil physico-chemical properties, flooding days, plant diversity index, coverage, and biomass (Figure 7). The analysis results showed that the eigenvalues of the first principal component (PC1) and the second principal component (PC2) were 4.60 and 3.21, respectively, explaining 27.0% and 18.9% of the total variability. The eigenvectors of the PC1 in descending order were Shannon-Wiener index (0.43), Simpson index (0.41), species richness (0.40), TP (0.14), TN (0.10), and SOM (0.08), all of which were positively correlated with littoral widths. The eigenvectors of the PC2 in descending order were SW (0.46), FD (0.32), biomass (0.25), E (0.23), and NH4+-N (0.15), all of which were negatively correlated with littoral widths.
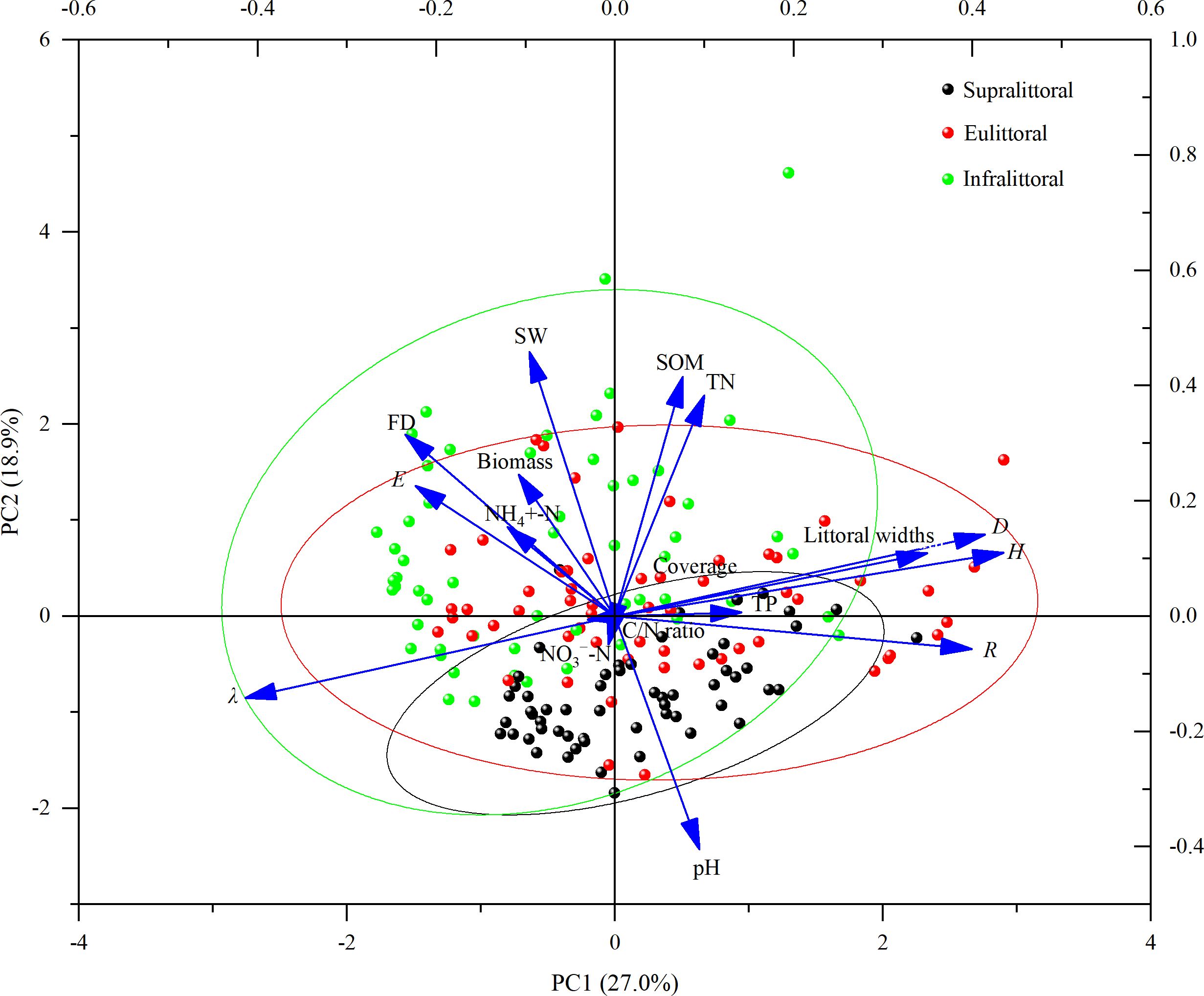
Figure 7. Principal component analysis of littoral widths with soil physico-chemical properties, plant diversity index, coverage, biomass, and flooding days (FD). Ellipses indicate 95% confidence intervals. SW, soil water content; NH4+-N, ammonium nitrogen; NO3−-N, nitrate nitrogen; SOM, soil organic matter; TP, total phosphorus; TN, total nitrogen; H, Shannon-Wiener index; D, Simpson Index; E, Pielou Evenness index; λ, dominance index; R, species richness.
Structural equation modeling path analyses revealed significant correlations among hydrology, soil, and plant variables, and that each variable was closely related to changes in littoral widths (Figure 8). The flooding days (FD) had a significant positive correlation with soil water content (SW) and pH, indicating that higher FD increased SW and regulated soil pH. FD had a significant negative correlation with plant parameters, indicating that changes in FD may have an important impact on plant productivity, coverage, and species richness. Moreover, plant parameters showed a significant positive correlation with plant diversity. Both soil nutrients and plant diversity exhibited a significant positive correlation with littoral widths, while FD, SW, pH, and plant parameters had no significant impact on littoral widths. In summary, FD directly affects soil (SW and pH) and plant parameters, while littoral widths were directly affected by soil nutrients and plant diversity.
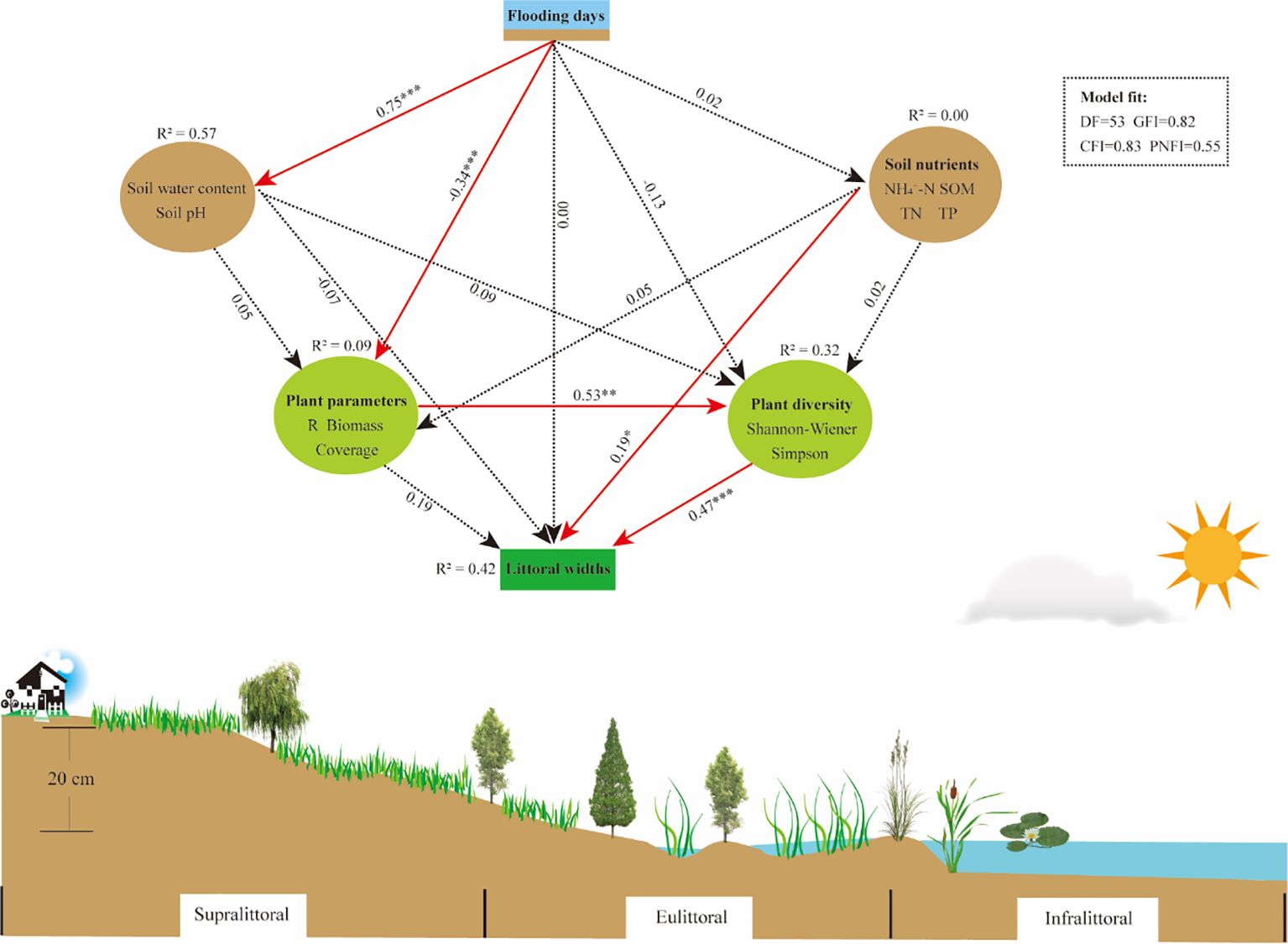
Figure 8. Structural equation modeling (SEM) reveals the relationship of flooding days, soil (water content, pH, nutrients), and plants (parameters, diversity) to littoral widths. Solid red arrows and dashed black arrows indicate significant and nonsignificant effects, respectively. The numbers beside the arrows represent standardized path coefficients; *P < 0.05, **P < 0.01, ***P < 0.001 indicate varying levels of significance. R2 values indicate the total variance explained for each variable. NH4+-N, ammonium nitrogen; SOM, soil organic matter; TN, total nitrogen; TP, total phosphorus.
Hydrological gradient changes directly determine the composition and range of plant communities across different elevations in the lakeshore zone (Holmquist et al., 2021; Zheng et al., 2019). We found that the importance values of hydrophytes and helophytes increased in the eulittoral and infralittoral under high water levels, while mesophytes decreased (Figure 2). This may be attributed to the frequent alternation of water levels in both zones, causing edge effects and changes in plant community structure and composition. Heterogeneity in lakeshore soil physicochemical properties under low-water conditions promoted microhabitat diversity, and hydrophytes and mesophytes coexisted by reducing direct competition (Zhang et al., 2022). A bigger challenge for these plants is to tolerate anoxic conditions in the substrate and supply oxygen to the roots during the high water. Hydrophytes and helophytes have well-developed root systems that efficiently absorb soil nutrients, allowing them to survive in flooded environments (Yang et al., 2024). Growth and biomass accumulation of the flood- and drought-tolerant invasive Alternanthera philoxeroides were promoted by adequate sunlight and water availability (Peng et al., 2021). This explains its increased importance value and biomass in the eulittoral (Figures 2, 3B), with a mean biomass of 0.33 kg/m2 in summer and 0.79 kg/m2 in winter.
Significant seasonal differences were observed in diversity indices, biomass, and coverage in both the eulittoral and infralittoral, which are susceptible to water level fluctuations (Figure 3). Longer flooding durations, as an abiotic stressor, inhibit the survival of flood-intolerant species, thereby reducing species richness and diversity (Casanova and Brock, 2000; Huang et al., 2021). Previous studies suggest that rising water levels regulate competition in eulittoral plant communities, favoring flood-tolerant plants and driving species homogenization (Garssen et al., 2017), which is consistent with our findings. Flood-adapted species occupy broader niches, and their increased relative abundance may enhance evenness (Altenfelder et al., 2016). Supralittoral plant communities maintain stable species composition and diversity, possibly because they are unaffected by flooding (Figure 3A).
Plants adjust their niche to optimize resource use (Carscadden et al., 2020), with niche breadth being positively correlated with their adaptability and tolerance to the environment (Costa et al., 2018). Wetland plants exhibit distinct adaptive strategies across hydrological gradients, with niche breadth variations reflecting species-specific hydrologic responses (Lou et al., 2018). In this study, the niche breadth of dominant species decreased in winter (Figure 4A), suggesting that rising water levels reduced their resource utilization capacity. The invasive Alternanthera philoxeroides exhibits the highest niche breadth, suggesting it is a generalist species. However, high water levels can inhibit its regeneration and dispersal (Zhang et al., 2024). Helophytes such as Zizania latifolia are distributed at lower elevations, and their niche breadths are almost unaffected by water level changes (Figures 4A, 5). High seed production and dispersal capacity enable Cynodon dactylon to quickly colonize and expand in receding environments (Li et al., 2023). Therefore, rising water levels may narrow its niche breadth.
Littoral width determines plant survival space and resource utilization, and is significantly influenced by water level changes. Rising water levels inundate low-elevation zones, driving vegetation migration upland and reducing primary distribution ranges (Leyer, 2005). Declining water levels expand plant-growing space in the lakeshore zone, increasing habitat diversity and availability (Dai et al., 2019). Our results indicated that rising water levels significantly decreased the eulittoral width but had no effect on the infralittoral and supralittoral (Figure 4B).
Littoral width differences are explained by seasonal variations in flooding days (Figure 1C): (1) Eulittoral plants are highly sensitive to periodic flooding and exposure, and prolonged inundation leads to the disappearance of flood-intolerant species (Garssen et al., 2015; Ye et al., 2020), while combined low-temperature and flooding stress restricts growth and niche breadth. (2) The supralittoral at higher elevations is less sensitive to water level fluctuations, providing stable conditions for plant survival and growth. (3) The aerenchyma and specialized leaves of helophytes and floating plants facilitate survival in hypoxic and fluctuating aquatic environments (Björn et al., 2022; Chen et al., 2002; Venter et al., 2017). Stable niche breadths were maintained by infralittoral plants despite increased flooding days, with growth likely constrained by water nutrients, water exchange, and sunlight conditions (Jin et al., 2024; Wu et al., 2023).
Seasonal variations in flooding duration caused significant changes in plants, soils, and littoral widths in the lakeshore zone. We observed that plant diversity indices were positively correlated with littoral widths (Figures 7, 8), suggesting that higher plant diversity contributes to expanding niche breadth. Plant communities with high diversity reduce interspecific competition through niche differentiation, promoting efficient resource partitioning and enhancing community stability (McKane et al., 2002). Littoral width is a metric for predicting range size and is positively correlated with environmental tolerance breadth (Slatyer et al., 2013). A broader width indicates that plant communities can adapt to a wider range of environmental conditions, with high ecological resilience and competitiveness.
Subtle water level fluctuations alter wetland plant distribution and ecological processes (Liu et al., 2020; Qin et al., 2017), and similarly impact habitat conditions such as soil redox potential, microbial activity, and oxygen availability, which in turn influence plant growth and community composition (Hájek et al., 2013; Huang et al., 2023). Nutrient availability gradients alter plant diversity and productivity, influencing habitat ranges for different vegetation types (Zhang et al., 2022). For example, organic-rich soils foster aquatic plant growth (Silveira and Thomaz, 2015), while xerophytes dominate in low-nutrient soils (Fan et al., 2019). Therefore, flood duration influences littoral width by altering soil properties, plant diversity and niche breadth, and promoting plant community environmental adaptations.
Our results suggest that high water levels reduced the niche breadth of most dominant perennial plants and narrowed the eulittoral width, while littoral widths were positively correlated with plant diversity and soil nutrients. Based on our findings and management needs, we recommend: 1) Regulate water levels to stabilize eulittoral habitat and promote plant diversity; 2) Optimize plant community vertical structure across elevation gradients to maintain plant diversity under water level fluctuations; 3) Develop appropriate water level thresholds to avoid extreme drought-flood events that threaten lakeshore plants.
Water level fluctuations not only shape the vertical structure of wetland plants, but are also a crucial factor influencing plant community composition and diversity. We investigated the seasonal variations in plant diversity, coverage, aboveground biomass, and soil physicochemical properties across three elevation ranges in the Erhai lakeshore zone, and explored the influence of hydrological gradient changes on littoral widths as well as the potential relationships. We found that the most significant seasonal differences in diversity indices, biomass, littoral widths, and soil physicochemical properties were observed in the eulittoral, followed by the infralittoral and supralittoral. Soil nutrients and plant diversity are the main influences on littoral widths. In addition, species response curves provide further insight into the elevation range of plant distribution in the lakeshore zone, which helps to capture the response of plant niche breadth to changes in the hydrologic gradient. These findings deepen our understanding that seasonal hydrologic changes affect plant communities and littoral widths in the lakeshore zone, and provide crucial guidance for developing effective wetland water level management in response to extreme drought events.
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding authors.
FZ: Conceptualization, Formal Analysis, Investigation, Writing – original draft, Writing – review & editing. JY: Conceptualization, Investigation, Writing – review & editing. ZH: Investigation, Methodology, Software, Writing – review & editing. XG: Investigation, Resources, Writing – review & editing. WL: Data curation, Investigation, Writing – review & editing. SY: Formal Analysis, Validation, Writing – review & editing. ZC: Conceptualization, Funding acquisition, Supervision, Writing – review & editing.
The author(s) declare that financial support was received for the research and/or publication of this article. This research was funded by the Basic Scientific Research Service Funds of Central Level Public Welfare Research Institutes, 2024YSKY-18.
We thank the staff of Erhai Administration Bureau for coordination of field work; Renxin Xu for help in the field investigation; Qiu Yu, and Bei Du for their assistance in the field work.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Altenfelder, S., Schmitz, M., Poschlod, P., Kollmann, J., Albrecht, H. (2016). Managing plant species diversity under fluctuating wetland conditions: The case of temporarily flooded depressions. Wetl Ecol. Manage. 24, 597–608. doi: 10.1007/s11273-016-9490-2
Baschuk, M. S., Ervin, M. D., Clark, W. R., Armstrong, L. M., Wrubleski, D. A., Goldsborough, G. L. (2012). Using satellite imagery to assess macrophyte response to water-level manipulations in the saskatchewan river delta, manitoba. Wetlands 32, 1091–1102. doi: 10.1007/s13157-012-0339-z
Björn, L. O., Middleton, B. A., Germ, M., Gaberscik, A. (2022). Ventilation systems in wetland plant species. Diversity-Basel 14. doi: 10.3390/d14070517
Carscadden, K. A., Emery, N. C., Arnillas, C. A., Cadotte, M. W., Afkhami, M. E., Gravel, D., et al. (2020). Niche breadth: Causes and consequences for ecology, evolution, and conservation. Q Rev Biol. 95, 179–214. doi: 10.1086/710388
Casanova, M. T., Brock, M. A. (2000). How do depth, duration and frequency of flooding influence the establishment of wetland plant communities? Plant Ecol. 147, 237–250. doi: 10.1023/a:1009875226637
Chapin, D. M., Paige, D. K. (2013). Response of delta vegetation to water level changes in a regulated mountain lake, washington state, USA. Wetlands 33, 431–444. doi: 10.1007/s13157-013-0401-5
Chen, H. J., Qualls, R. G., Miller, G. C. (2002). Adaptive responses of lepidium latifolium to soil flooding: Biomass allocation, adventitious rooting, aerenchyma formation and ethylene production. Environ. Exp. Bot. 48, 119–128. doi: 10.1016/s0098-8472(02)00018-7
Chen, Y. S., Stagg, C. L., Cai, Y. J., Lü, X. T., Wang, X. L., Shen, R. C., et al. (2020). Scaling responses of leaf nutrient stoichiometry to the lakeshore flooding duration gradient across different organizational levels. Sci. Total Environ. 740. doi: 10.1016/j.scitotenv.2020.139740
Costa, D. S., Gerschlauer, F., Kiese, R., Fischer, M., Kleyer, M., Hemp, A. (2018). Plant niche breadths along environmental gradients and their relationship to plant functional traits. Divers. Distrib. 24, 1869–1882. doi: 10.1111/ddi.12815
Dai, X., Wan, R. R., Yang, G. S., Wang, X. L., Xu, L. G., Li, Y. Y., et al. (2019). Impact of seasonal water-level fluctuations on autumn vegetation in poyang lake wetland, China. Front. Earth Sci. 13, 398–409. doi: 10.1007/s11707-018-0731-y
Dali Bai Autonomous Prefecture People’s Government (DBAPPG) (2023).Proposal for deliberation on the minimum operating water level of Erhai Lake in the special year. Available online at: https://www.dali.gov.cn/dlrmzf/c101530/202306/03cfc4fc80c6435ab5e7343e6c9e3cf0.shtml (Accessed (accessed 2 June 2023)).
Duval, T. P., Waddington, J. M., Branfireun, B. A. (2012). Hydrological and biogeochemical controls on plant species distribution within calcareous fens. Ecohydrology 5, 73–89. doi: 10.1002/eco.202
Elith, J., Graham, C. H., Anderson, R. P., Dudík, M., Ferrier, S., Guisan, A., et al. (2006). Novel methods improve prediction of species’ distributions from occurrence data. Ecography 29, 129–151. doi: 10.1111/j.2006.0906-7590.04596.x
Fan, H. X., Xu, L. G., Wang, X. L., Jiang, J. H., Feng, W. J., You, H. L. (2019). Relationship between vegetation community distribution patterns and environmental factors in typical wetlands of Poyang lake, China. Wetlands 39, S75–S87. doi: 10.1007/s13157-017-0903-7
Feng, W. J., Santonja, M., Bragazza, L., Buttler, A. (2020). Shift in plant-soil interactions along a lakeshore hydrological gradient. Sci. Total Environ. 742. doi: 10.1016/j.scitotenv.2020.140254
Fu, H., Lou, Q., Dai, T. T., Yuan, G. X., Huang, Z. R., Ge, D. B., et al. (2018). Hydrological gradients and functional diversity of plants drive ecosystem processes in Poyang lake wetland. Ecohydrology 11. doi: 10.1002/eco.1950
Gaberscik, A., Grasic, M., Abram, D., Zelnik, I. (2020). Water level fluctuations and air temperatures affect common reed habitus and productivity in an intermittent wetland ecosystem. Water 12. doi: 10.3390/w12102806
Gaberscik, A., Krek, J. L., Zelnik, I. (2018). Habitat diversity along a hydrological gradient in a complex wetland results in high plant species diversity. Ecol. Eng. 118, 84–92. doi: 10.1016/j.ecoleng.2018.04.017
Garssen, A. G., Baattrup-Pedersen, A., Riis, T., Raven, B. M., Hoffman, C. C., Verhoeven, J. T. A., et al. (2017). Effects of increased flooding on riparian vegetation: Field experiments simulating climate change along five European lowland streams. Glob. Chang. Biol. 23, 3052–3063. doi: 10.1111/gcb.13687
Garssen, A. G., Baattrup-Pedersen, A., Voesenek, L., Verhoeven, J. T. A., Soons, M. B. (2015). Riparian plant community responses to increased flooding: A meta-analysis. Glob. Chang. Biol. 21, 2881–2890. doi: 10.1111/gcb.12921
Gathman, J. P., Albert, D. A., Burton, T. M. (2005). Rapid plant community response to a water level peak in northern lake huron coastal wetlands. J. Gt Lakes Res. 31, 160–170. doi: 10.1016/s0380-1330(05)70296-3
Gong, F., Luo, L., Li, H., Chen, L., Zhang, R., Wu, G., et al. (2023). Quantitative assessment of water quality improvement by reducing external loadings at lake Erhai, southwest China. Int. J. Environ. Res. Public Health 20. doi: 10.3390/ijerph20065038
Guisan, A., Edwards, T. C., Hastie, T. (2002). Generalized linear and generalized additive models in studies of species distributions: Setting the scene. Ecol. Model. 157, 89–100. doi: 10.1016/s0304-3800(02)00204-1
Hájek, M., Hájková, P., Kocí, M., Jirousek, M., Mikulásková, E., Kintrová, K. (2013). Do we need soil moisture measurements in the vegetation-environment studies in wetlands? J. Veg. Sci. 24, 127–137. doi: 10.1111/j.1654-1103.2012.01440.x
Holmquist, J. R., SChile-Beers, L., Buffington, K., Lu, M., Mozdzer, T. J., Riera, J., et al. (2021). Scalability and performance tradeoffs in quantifying relationships between elevation and tidal wetland plant communities. Mar. Ecol. Prog. Ser. 666, 57–72. doi: 10.3354/meps13683
Hu, J. Y., Xie, Y. H., Tang, Y., Li, F., Zou, Y. A. (2018). Changes of vegetation distribution in the east Dongting lake after the operation of the three Gorges dam, China. Front. Plant Sci. 9. doi: 10.3389/fpls.2018.00582
Huang, Y., Chen, X. S., Li, F., Hou, Z. Y., Li, X., Zeng, J., et al. (2021). Community trait responses of three dominant macrophytes to variations in flooding during 2011-2019 in a yangtze river-connected floodplain wetland (Dongting lake, China). Front. Plant Sci. 12. doi: 10.3389/fpls.2021.604677
Huang, X. Y., Li, Y. X., Lin, H. Y., Wen, X. T., Liu, J., Yuan, Z. F., et al. (2023). Flooding dominates soil microbial carbon and phosphorus limitations in Poyang lake wetland, China. Catena 232. doi: 10.1016/j.catena.2023.107468
Hutchinson, G. E. (1957). Population studies: Animal ecology and demography. Cold Spring Harb. Symp. Quant. Biol. 22, 415–427. doi: 10.1101/sqb.1957.022.01.039
Ji, T. (2005). Comparison on determining the organic matter contents in the soils by different heating methods in the potassium dichromate-volumetric method. Acta Agricult. Zhejiangensis. 17, 311–313.
Jin, L., Wu, Q. H., Xie, S. J., Chen, W. W., Duan, C. Q., Sun, C. Q., et al. (2024). Phosphorus stoichiometric homeostasis of submerged macrophytes and associations with interspecific interactions and community stability in Erhai lake, China. Water Res. 256. doi: 10.1016/j.watres.2024.121575
Lever, J., Krzywinski, M., Atman, N. (2017). Points of significance: Principal component analysis. Nat. Meth. 14, 641–642. doi: 10.1038/nmeth.4346
Leyer, I. (2005). Predicting plant species’ responses to river regulation: The role of water level fluctuations. J. Appl. Ecol. 42, 239–250. doi: 10.1111/j.1365-2664.2005.01009.x
Li, J. H., Bai, Y., Alatalo, J. M. (2020). Impacts of rural tourism-driven land use change on ecosystems services provision in erhai lake basin, China. Ecosyst. Serv. 42. doi: 10.1016/j.ecoser.2020.101081
Li, D., Chu, Z. S., Huang, M. S., Zheng, B. H. (2019). Multiphasic assessment of effects of design configuration on nutrient removal in storing multiple-pond constructed wetlands. Bioresour. Technol. 290. doi: 10.1016/j.biortech.2019.121748
Li, X. L., Li, S. Z., Xie, Y. W., Wei, Z. H., Li, Z. L. (2023). What drives the morphological traits of stress-tolerant plant cynodon dactylon in a riparian zone of the three gorges reservoir, China. Water 15. doi: 10.3390/w15183183
Liu, Q., Liu, J. L., Liu, H. F., Liang, L. Q., Cai, Y. P., Wang, X., et al. (2020). Vegetation dynamics under water-level fluctuations: Implications for wetland restoration. J. Hydrol. 581. doi: 10.1016/j.jhydrol.2019.124418
Lou, Y. J., Gao, C. Y., Pan, Y. W., Xue, Z. S., Liu, Y., Tang, Z. H., et al. (2018). Niche modelling of marsh plants based on occurrence and abundance data. Sci. Total Environ. 616, 198–207. doi: 10.1016/j.scitotenv.2017.10.300
McKane, R. B., Johnson, L. C., Shaver, G. R., Nadelhoffer, K. J., Rastetter, E. B., Fry, B., et al. (2002). Resource-based niches provide a basis for plant species diversity and dominance in arctic tundra. Nature 415, 68–71. doi: 10.1038/415068a
Ojdanic, N., Holcar, M., Golob, A., Gaberscik, A. (2023). Environmental extremes affect productivity and habitus of common reed in intermittent wetland. Ecol. Eng. 189. doi: 10.1016/j.ecoleng.2023.106911
Ostendorp, W. (2004). New approaches to integrated quality assessment of lakeshores. Limnologica 34, 160–166. doi: 10.1016/s0075-9511(04)80036-7
Peng, X. M., Yang, Y. F., Yan, X. M., Li, H. Y. (2021). The effects of water control on the survival and growth of alternanthera philoxeroides in the vegetative reproduction and seedling stages. Sci. Rep. 11. doi: 10.1038/s41598-021-92674-2
Pollock, M. M., Naiman, R. J., Hanley, T. A. (1998). Plant species richness in riparian wetlands - a test of biodiversity theory. Ecology 79, 94–105.
Qin, L., Jiang, M., Tian, W., Zhang, J., Zhu, W. H. (2017). Effects of wetland vegetation on soil microbial composition: A case study in Tumen river basin, northeast China. Chin. Geogr. Sci. 27, 239–247. doi: 10.1007/s11769-017-0853-2
Shen, R. C., Lan, Z. C., Chen, Y. S., Leng, F., Jin, B. S., Fang, C. M., et al. (2019). The effects of flooding regimes and soil nutrients on lakeshore plant diversity in a pristine lake and a human managed lake in subtropical China. J. Freshw. Ecol. 34, 757–769. doi: 10.1080/02705060.2019.1687340
Shen, R. C., Lan, Z. C., Huang, X. Y., Chen, Y. S., Hu, Q. W., Fang, C. M., et al. (2020). Soil and plant characteristics during two hydrologically contrasting years at the lakeshore wetland of Poyang lake, China. J. Soils Sediments. 20, 3368–3379. doi: 10.1007/s11368-020-02638-8
Silveira, M. J., Thomaz, S. M. (2015). Growth of a native versus an invasive submerged aquatic macrophyte differs in relation to mud and organic matter concentrations in sediment. Aquat. Bot. 124, 85–91. doi: 10.1016/j.aquabot.2015.03.004
Slatyer, R. A., Hirst, M., Sexton, J. P. (2013). Niche breadth predicts geographical range size: A general ecological pattern. Ecol. Lett. 16, 1104–1114. doi: 10.1111/ele.12140
Stroh, C. L., De Steven, D., Guntenspergen, G. R. (2008). Effect of climate fluctuations on long-term vegetation dynamics in Carolina bay wetlands. Wetlands 28, 17–27. doi: 10.1672/06-117.1
Venter, N., Cowie, B. W., Witkowski, E. T. F., Snow, G. C., Byrne, M. J. (2017). The amphibious invader: Rooted water hyacinth’s morphological and physiological strategy to survive stranding and drought events. Aquat. Bot. 143, 41–48. doi: 10.1016/j.aquabot.2017.09.004
Wang, Z. J., Gong, H. L., Zhang, J. (2015). Receding water line and interspecific competition determines plant community composition and diversity in wetlands in Beijing. PloS One 10. doi: 10.1371/journal.pone.0124156
Wang, J., Song, Y. H., Ge, B. C., Zhou, Y. (2023). Dynamic spatiotemporal land use evolution in China’s plateau lake basins in response to landscape ecological sensitivity. Sustainability 15. doi: 10.3390/su152015020
Wang, X., Wang, H. L., Wang, H. Y., Guo, W. X., Zhai, H. Y., Zhang, X. K. (2022). Responses of lakeshore herbaceous plant guilds to altered water level fluctuations in Yangtze floodplain lakes, China. Ecol. Indic. 145. doi: 10.1016/j.ecolind.2022.109714
Wassens, S., Ning, N., Hardwick, L., Bino, G., Maguire, J. (2017). Long-term changes in freshwater aquatic plant communities following extreme drought. Hydrobiologia 799, 233–247. doi: 10.1007/s10750-017-3219-y
Wen, J. H., Li, B. Y., Xiao, H. Y., Gong, C. Y., Gao, A. G., Wang, Y. H., et al. (2023). Floating mat formation makes zizania latifolia more competitive under the conditions of continuous significant water level rise. Plants-Basel 12. doi: 10.3390/plants12051193
Wen, Z. H., Ma, Y. W., Wang, H., Cao, Y., Yuan, C. B., Ren, W. J., et al. (2021). Water level regulation for eco-social services under climate change in Erhai lake over the past 68 years in China. Front. Environ. Sci. 9. doi: 10.3389/fenvs.2021.697694
Wu, Q. H., Xie, M. L., Jin, L., Dong, H. J., Yuan, D. Y., Yang, T. M., et al. (2023). Water exchange unevenness alters the species dominance and community composition of submerged macrophytes in Erhai lake and the potential mechanisms revealed by laboratory experiment. Sci. Total Environ. 882. doi: 10.1016/j.scitotenv.2023.163624
Yang, S. L., Yuan, Z., Ye, B. B., Zhu, F., Chu, Z. S., Liu, X. W. (2024). Impacts of landscape pattern on plants diversity and richness of 20 restored wetlands in Chaohu lakeside of China. Sci. Total Environ. 906. doi: 10.1016/j.scitotenv.2023.167649
Ye, C., Butler, O. M., Chen, C. R., Liu, W. Z., Du, M., Zhang, Q. F. (2020). Shifts in characteristics of the plant-soil system associated with flooding and revegetation in the riparian zone of three gorges reservoir, China. Geoderma 361. doi: 10.1016/j.geoderma.2019.114015
You, H. L., Xu, L. G., Liu, G. L., Wang, X. L., Wu, Y. M., Jiang, J. H. (2015). Effects of inter-annual water level fluctuations on vegetation evolution in typical wetlands of Poyang lake, China. Wetlands 35, 931–943. doi: 10.1007/s13157-015-0684-9
Zhang, H. J., Huang, B., Huang, Y. T., Liu, J. (2024). High water levels and low light co-inhibit colonization of fragments from the clonal invader alternanthera philoxeroides. Aquat. Bot. 192. doi: 10.1016/j.aquabot.2024.103749
Zhang, X. K., Liu, X. Q., Wang, H. Z. (2015). Effects of water level fluctuations on lakeshore vegetation of three subtropical floodplain lakes, China. Hydrobiologia 747, 43–52. doi: 10.1007/s10750-014-2121-0
Zhang, Q. J., Wang, Z. S., Xia, S. X., Zhang, G. S., Li, S. X., Yu, D. K., et al. (2022). Hydrologic-induced concentrated soil nutrients and improved plant growth increased carbon storage in a floodplain wetland over wet-dry alternating zones. Sci. Total Environ. 822. doi: 10.1016/j.scitotenv.2022.153512
Zhang, S. Q., Zhang, P. Y., Pan, B. H., Zou, Y., Xie, Y. H., Zhu, F., et al. (2021). Wetland restoration in the east Dongting lake effectively increased waterbird diversity by improving habitat quality. Glob. Ecol. Conserv. 27. doi: 10.1016/j.gecco.2021.e01535
Zhao, Y. T., Wang, G. D., Zhao, M. L., Wang, M., Xue, Z. S., Liu, B., et al. (2021). Seed limitation and saline-alkaline stress restrict wetland restoration potential in the Songnen plain, Northeastern China. Ecol. Indic. 129. doi: 10.1016/j.ecolind.2021.107998
Zheng, X. J., Fu, J., Ramamonjisoa, N., Zhu, W. H., He, C. G., Lu, C. Y. (2019). Relationship between wetland plant communities and environmental factors in the tumen river basin in northeast China. Sustainability 11. doi: 10.3390/su11061559
Zheng, P. R., Li, C. H., Ye, C., Wang, H., Wei, W. W., Zheng, Y., et al. (2022). Characteristic and affecting factors of wetland herbs’ distribution in the radiant belt toward land of lake-terrestrial ecotone in Tibet, China. Environ. Sci. Eur. 34. doi: 10.1186/s12302-022-00593-4
Zhu, G. R., Yuan, C. B., Di, G. L., Zhang, M., Ni, L. Y., Cao, T., et al. (2018). Morphological and biomechanical response to eutrophication and hydrodynamic stresses. Sci. Total Environ. 622, 421–435. doi: 10.1016/j.scitotenv.2017.11.322
Keywords: littoral widths, plant diversity, soil nutrient, hydrological seasonal changes, niche breadth
Citation: Zhu F, Yuan J, Hou Z, Guo X, Liao W, Yang S and Chu Z (2025) Seasonal water level changes affect plant diversity and littoral widths at different elevation zones in the Erhai Lake. Front. Plant Sci. 16:1503627. doi: 10.3389/fpls.2025.1503627
Received: 30 September 2024; Accepted: 03 March 2025;
Published: 20 March 2025.
Edited by:
Xin-Sheng Chen, Anhui University, ChinaReviewed by:
Igor Zelnik, University of Ljubljana, SloveniaCopyright © 2025 Zhu, Yuan, Hou, Guo, Liao, Yang and Chu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jing Yuan, eHl2eS04OTQ1QDE2My5jb20=; Zhaosheng Chu, Y2h1enNzY2lAeWVhaC5uZXQ=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.