 Hui Li1,2
Hui Li1,2 Yaoyao Zhao1,2
Yaoyao Zhao1,2 Xiaohang Weng1,2Yongbin Zhou3Yan Huo4
Xiaohang Weng1,2Yongbin Zhou3Yan Huo4 Songzhu Zhang1,2
Songzhu Zhang1,2 Liying Liu1,2
Liying Liu1,2 Jiubo Pei5*
Jiubo Pei5*- 1College of Forestry, Shenyang Agricultural University, Shenyang, Liaoning, China
- 2Research Station of Liaohe-River Plain Forest Ecosystem, Chinese Forest Ecosystem Research Network (CFERN), Shenyang Agricultural University, Changtu, China
- 3Institute of Modern Agricultural Research, Dalian University, Dalian, China
- 4Liaoning Dryland Agriculture and Forestry Research Institute, Chaoyang, Liaoning, China
- 5College of Land and Environment, Shenyang Agricultural University, Shenyang, Liaoning, China
Introduction: Pinus tabuliformis as a crucial afforestation species in semi-arid regions, faces issues such as the reduction of plantations. Calcium plays a significant role in alleviating drought stress and promoting nutrient uptake in plants.
Methods: Utilizing a pot experiment approach, seedlings were treated with exogenous calcium at five concentrations (0, 50, 100, 200, and 400 mg•kg-1). The nutrient content of the plants and soil was measured, and their ecological stoichiometric characteristics and internal stability were analyzed. This was followed by a series of related studies.
Results: As the concentration of calcium increases, the contents of carbon, nitrogen, phosphorus, and potassium in various organs and the whole plant exhibit a trend of first increasing and then decreasing, peaking at calcium treatment of 50-100 mg•kg-1. Concurrently, the calcium concentration in plant organs and the entire plant gradually increases with the availability of calcium in the soil. The addition of exogenous calcium has a certain impact on the ecological stoichiometric ratios (C:N, C:P, N:P) of Pinus tabuliformis seedlings' leaves, stems, roots, and the whole plant, exhibiting distinct variation characteristics. At calcium concentrations of 50-100 mg•kg-1, the ratios of C:N and C:P are relatively lower. Under calcium concentrations of 0, 50, and 100 mg•kg-1, soil calcium shows a positive correlation with the total carbon (TC), total nitrogen (TN), total phosphorus (TP), total potassium (TK), and calcium contents in leaves, stems, roots, and the entire plant. However, at calcium concentrations of 200 and 400 mg•kg-1, soil calcium exhibits a significant positive correlation with the calcium content in leaves, stems, roots, and the entire plant, and a significant negative correlation with the total phosphorus, total nitrogen, total phosphorus, and total potassium contents. After the addition of exogenous calcium at different concentrations, most stoichiometric indices of various organs of Pinus tabuliformis seedlings demonstrate strong balance.
Discussion: Calcium, as an essential structural component and second messenger, regulates the nutrient uptake and utilization in plants, influencing the stoichiometry. However, both low and high concentrations of calcium can be detrimental to plant growth by disrupting nutrient metabolism and internal structures. Consequently, there exists an optimal calcium concentration for nutrient absorption.
1 Introduction
The semi-arid region is characterized by a dry climate, scarce precipitation, and an extremely fragile ecological environment. Planted forests play a crucial role in this area by helping to prevent winds, fix sands, and maintain soil and water (Barroso et al., 2022). Pinus tabuliformis is one of the main afforestation species in semi-arid regions of China and is also the main component of the forests in the western part of Liaoning province. Its exceptional drought-resistant and indigenous characteristics have also made it useful in other projects to prevent desertification and control sandstorms, such as the “Three-North Protected Forest Program” and the “Returning Cultivated Land to Forests Project” (Zhu and Zheng, 2019; Zeng et al., 2020). Despite its ecological significance, Pinus tabuliformis has faced significant challenges in recent decades, experiencing serious decline and mortality issues. Furthermore, plantation forests have also triggered other aspects of ecological crises due to monoculture stand structure and poor stand stability, including declining land productivity, increasing pests and diseases incidence, and declining carbon storage (Liu et al., 2019; Li et al., 2020; Zhang et al., 2021). Therefore, scientific and systematic research on the environmental adaptation of plantation forest growth in semi-arid areas, elimination and mitigation of constraints leading to the decline of plantation forests have become the key to solving the problem of healthy development of plantation forests in the region, and also the way to the development of healthy and sustainable management of plantation forests in the region.
The defining element that migrates within the soil of semi-arid regions is calcium. In these areas, the soil is predominantly sandy and nutrient-poor, exhibiting a deficiency in calcium. While on the other hand, due to the seasonal fluctuations in rainfall and drought typical of semi-arid environments, Ca2+ leaching within the soil can precipitate at certain depths, forming a layer of calcium accumulation that influences the soil’s physical, chemical, and biological characteristics (Arkley, 1963; Certini and Scalenghe, 2006; Ermakov et al., 2020; Lawrence et al., 2020; Sun et al., 2021). Calcium plays a pivotal role in plant growth, moderate levels of calcium can mitigate drought stress and foster growth, while a deficiency can inhibit growth (Aras et al., 2021; Li et al., 2022a, b). Studies in Pinus massoniana have shown that calcium deficiency significantly inhibits seedling growth and development and impairs photosynthesis processes (Hu et al., 2022). Therefore, from the perspective of element utilization in soil-forest vegetation systems, calcium can be considered an indirect influencing factor on forest vegetation growth. Based on this, in environments where vegetation growth in semi-arid regions is affected by both calcium deficiency and abundance, it is necessary to conduct in-depth research on the different effects and mechanisms of calcium regulation in artificial forests in this region. This is of significant importance for studying the mechanisms of decline in artificial forests in semi-arid regions and addressing ecological issues and research needs in artificial forest cultivation in these areas.
Since the publication of Sterner and Elser (2002), the field of ecological stoichiometry has matured considerably over time and has emerged as a new approach to exploring the effects of environmental nutrient supply fluctuations on nutrient element limitations, population dynamics, and ecosystem functioning (Qi et al., 2020; Li et al., 2023b). It primarily investigates the coupled relationships among the essential nutrients for plant growth, C, N, P, and K, reflecting the balance of resource acquisition by plants and their growth strategies (Liao et al., 2023; Wei et al., 2021a). The ecological stoichiometric characteristics of plant C:N:P can indicate the patterns of C accumulation and N, P limitations in the ecosystem they inhabit (Elser et al., 2010). Nutrient elements in plants are inevitably intertwined with soil nutrients, and measuring nutrients in the soil is a direct method to explore soil nutrient supply capacity (Chapin et al., 1986). Soils and their nutrients provide a favorable growth environment for plant growth, while plants, in turn, impact the structure of soil nutrients. The interaction between plants and soil constitutes the foundation of plant-soil feedback (Liu et al., 2023). Ecological stoichiometry is also considered an effective new approach to exploring soil-plant interactions and the cycling characteristics of ecosystem elements (Wu et al., 2023).
Previous studies on the effects of exogenous nutrient addition on the stoichiometric characteristics of plants have focused on elements such as silicon and nitrogen, but there is a paucity of research on the impacts of exogenous calcium addition (Long et al., 2018; Xu et al., 2021). Past studies on Pinus tabuliformis have primarily examined aspects like forest management, water regulation, and site quality, while the response of Pinus tabuliformis to soil calcium and the underlying physiological regulation mechanisms remain underexplored. Therefore, this study used Pinus tabuliformis seedlings as the research subject, applying different exogenous calcium concentration treatments. Targeting the calcium-deficient and calcium-rich soil environments typical of the semi-arid regions of the Three-North Shelter Forest in China, the study investigated the stoichiometric characteristics of different organs of Pinus tabuliformis seedlings. The aim was to identify the optimal calcium concentration for Pinus tabuliformis growth, further complement the research on the impacts of calcium on forest stands, and provide scientific theoretical support for the rational cultivation and healthy, sustainable management of artificial forests in semi-arid regions from the perspective of nutrient absorption.
2 Materials and methods
2.1 Experimental design
The experiment was conducted at No.37 experimental shed in Beishan Research Base of Shenyang Agricultural University. Three-year-old Pinus tabuliformis seedlings with consistent growth were chosen as the test materials. The collected sandy soil was carefully sieved to eliminate stones and impurities. Then, 3 kg of the air-dried soil weight was precisely weighed and mixed with 2 kg of 60-mesh quartz sand to create the potting soil. After thoroughly mixing the substrate, the soil was rinsed with a diluted hydrochloric acid solution to eliminate any potential Ca2+ ions before being filled into the pots. The pH of the soil-sand mixture after the hydrochloric acid wash was measured at 6.3. The seedlings, along with nutrient pots, were planted in plastic pots with an inner diameter of 23.8 cm, a depth of 24.8 cm, and a volume of approximately 11.36 L, in June 2019. Each pot received one seedling, and a plastic tray was placed beneath the pot to prevent water loss. After a two-week period of seedling acclimatization, a calcium-deficient standard nutrient solution was uniformly added once. This nutrient solution was prepared using ultrapure water according to Xie (2014) recipe for sand cultivation nutrient solution. The pH of the nutrient solution was adjusted to a range of 5-6 using NaOH. The components of the nutrient solution included 5 ml/L KNO3, 5 ml/L MgSO4, 5 ml/L KH2PO4, 5 ml/L NaNO3, and 5 ml/L EATA-Fe. Trace elements such as H3BO3, MnCl2, CuSO4, ZnSO4, and H2MoO4 were also incorporated into the nutrient solution.
In this experiment, five different treatments were set up and each treatment was replicated six times. Five calcium concentration levels of 0, 50, 100, 200 and 400 mg·kg-1 were set according to Paiva et al., 1998 and the condition that the local water-soluble calcium content ranged from 14.66 ~ 299.02 mg·kg-1 (Zhou and Zou, 2017). Treatments were applied in July and calcium was supplied by anhydrous CaCl2.To ensure the even distribution of CaCl2 in the soil, the CaCl2 solution was divided into aliquots and then irrigated into the soil. The experimental sandy soil was sourced from the understory soil of Pinus tabuliformis forest in Fujia Forest Farm, Changtu County, Tieling City, Liaoning Province. The soil samples were randomly collected from multiple points under standard standing conditions, with a collection depth ranging from 0-40 cm. Throughout the testing period, measures were taken to closely monitor the experimental phenomena. Adequate moisture was maintained by using deionized water, and necessary care, including loosening and weeding, was undertaken to ensure the smooth progress of the experiment.
2.2 Determination indicators and methods
In June of the following year, seedlings and soil samples were harvested for analysis. The seedlings were carefully washed and separated into leaves, stems, and roots. They were then heated at 105 °C for 0.5 hours and subsequently dried at 65 °C until a constant weight was achieved. The dry weight of each plant part was measured using a precision balance with a sensitivity of 1/1000. The individual dry weights of the leaves, stems, and roots were combined to obtain the total biomass of the seedlings. The pulverized leaves, stems, and roots were further crushed using a ball mill until they could pass through a 100-mesh sieve. These samples were then stored in small envelopes under dry conditions. As for the soil samples, they were air-dried, filtered, and then crushed and ground using a pulverizer. The final samples were stored in self-sealing bags and kept dry (Weng et al., 2022).
The determination of total carbon (TC) and total nitrogen (TN) contents in the plant leaves, stems, and roots was conducted using an elemental analyzer following the method described by Ding et al. (2022). The total phosphorus (TP) content in the plant tissues was determined using the acid-soluble molybdenum-antimony colorimetric method, as outlined by da Silveira Sousa Junior et al. (2022). The total potassium (TK) and calcium (Ca) contents in the plant tissues were determined using the acid-soluble-flame photometric method, following the procedure described by Sarto et al. (2019). For the soil samples, the total carbon (TC) and total nitrogen (TN) contents were determined using an elemental analyzer. The soil’s total phosphorus (TP) content was determined through acid solubility-molybdenum-antimony colorimetry. The soil available potassium (AK) content was determined using the NH4OAc extraction-flame photometric method, as described by Yin et al. (2021). The soil water-soluble calcium and soil-exchangeable calcium contents were determined following the method developed by Zhou Wei and Lin Bao, as mentioned by He et al. (2012).
2.3 Statistical analysis
The chemical stoichiometric characteristics of Pinus tabuliformis seedlings were subjected to Pearson correlation analysis with soil. When the test results were significant (p<0.05), an internal homeostasis model was introduced for further quantitative analysis of the stoichiometric characteristics of seedling leaves, stems, roots, and whole plants in response to environmental changes concerning the content and ratios of C, N, P, K, and Ca elements. The equation of the internal homeostasis model is: (Sterner and Elser, 2002). In this equation, y represents the nutrient elements C, N, P, K, and Ca content and ratios in different plant organs, x represents the corresponding nutrient element content and ratios in the soil environment, c is a constant, and H is the internal homeostasis index. The strength of internal homeostasis is measured by 1/H (Hood and Sterner, 2010). If the fitting equation does not reach a significant level (p≥0.05) or 1/H ≤ 0, it is strictly homeostatic; 0<1/H<0.25, it is homeostatic; 0.25<1/H<0.5, it is weak homeostatic; 0.5<1/H<0.75, it is weak responsive; 1/H >0.75, it is responsive (Persson et al., 2010).
Basic data organization was done using Microsoft Excel 2019 software, significance testing (p<0.05) was performed using SPSS 19, redundancy analysis and plotting were conducted using Canoco 5 software, and plotting was carried out using GraphPad Prism 8.0.2.
3 Results
3.1 Effects of exogenous calcium addition on the nutrient content of various organs and the distribution characteristics of nutrient content in Pinus tabuliformis seedlings
As shown in Figures 1A-E, the exogenous calcium addition significantly influenced the contents of TC, TN, TP, TK, and Ca in the leaves, stems, roots, and whole seedlings of Pinus tabuliformis (p<0.05). The contents of TC, TN, TP, and TK in different organs and the whole seedling showed a trend of first increasing and then decreasing with the increase of calcium concentration, while the Ca content gradually increased.
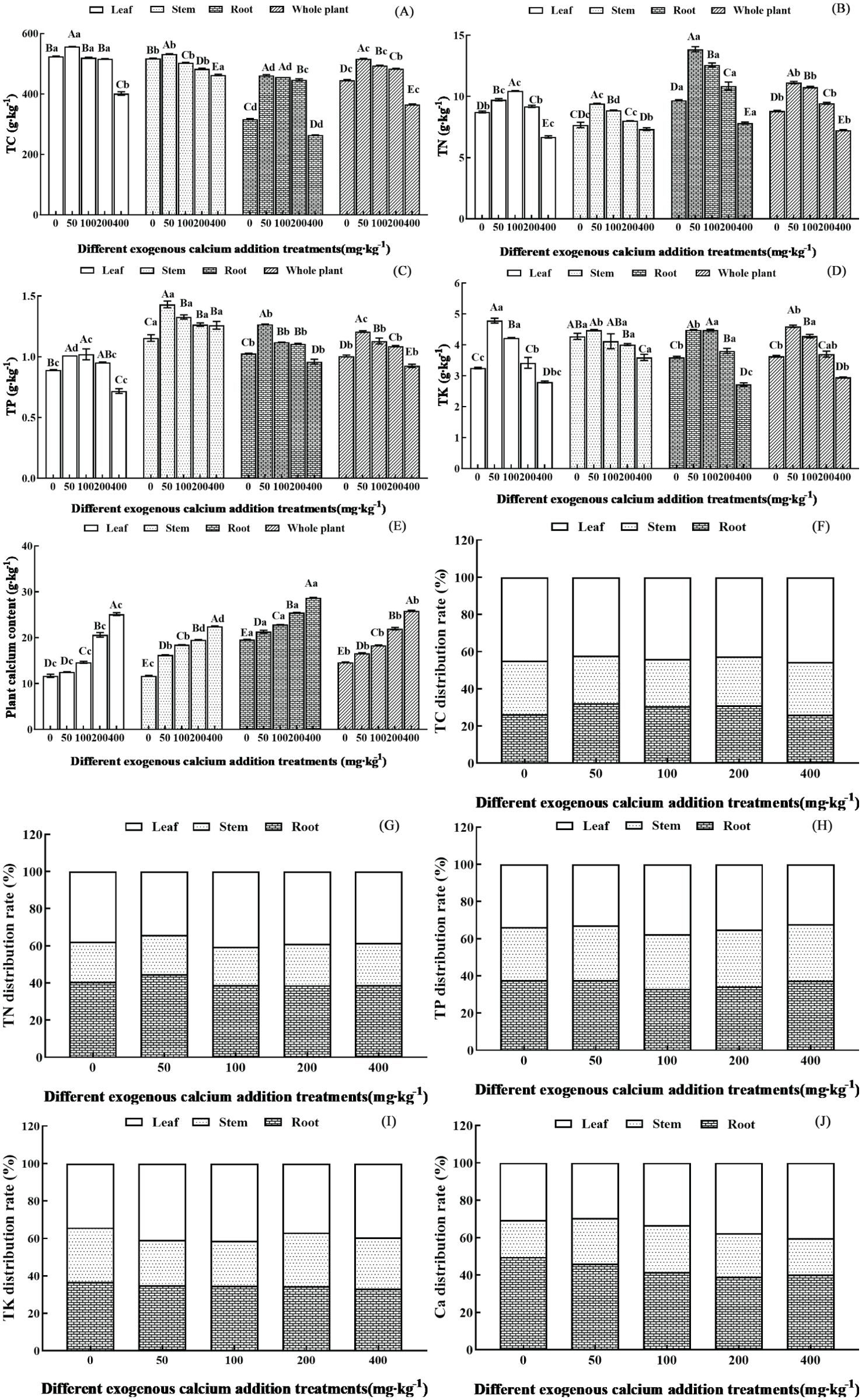
Figure 1. Nutrient content and distribution characteristics of different organs of Pinus tabuliformis seedlings under exogenous calcium supplementation. Capital letters indicated the difference between different calcium concentrations in leaves, stems, roots and whole plants (p<0.05), while lower letters indicated the difference between leaves, stems, roots and whole plants at the same concentration (p<0.05). (A) TC (total carbon); (B) TN(total nitrogen); (C) TP (total phosphorus); (D) TK (total potassium); (E) Plant calcium content; (F) TC distribution rate; (G) TN distribution rate; (H) TP distribution rate; (I) TK distribution rate; (J) Ca distribution rate.
The TC content in the leaves, stems, and whole plants reached the maximum value under the treatment of 50 mg·kg-1 calcium, increasing by 6.18%, 2.85%, and 15.82% respectively compared to the control without calcium; while the root content reached the highest at calcium concentrations of 50-100 mg·kg-1, which increased by 44.63% compared to the control without calcium. The TN and TP content in the leaves significantly exceeded other treatments at 100 mg·kg-1, increasing by 19.69% and 14.03%, respectively, compared to the control without calcium; while the stem, root, whole plant was significantly higher at 50 mg·kg-1, with increases of 22.75% and 23.91%, 43.08% and 23.41%, 26.25% and 19.95% respectively, when compared to no calcium application (p< 0.05). The TK content in the leaves, stems, and whole plants reached its peak at a calcium treatment of 50 mg·kg-1, with increases of 47.27%, 4.75%, and 26.66% respectively, compared to no calcium, while the TK content in the roots peaked at a calcium treatment of 100 mg·kg-1, showing an increase of 24.60% over the absence of calcium. Subsequently, with the increase of calcium concentration, the TC, TN, TP, and TK contents in each organ and the whole plant showed a decreasing trend, and when the calcium concentration reached 400 mg·kg-1, they all significantly decreased compared to the control without calcium (except for the TP content in stems) (p< 0.05). The calcium content in each organ and the whole plant gradually increased with the increase of exogenous calcium concentration. When the calcium concentration increased to 400 mg·kg-1, the calcium contents of the leaves, stems, roots, and whole plants significantly increased by 115.01%, 92.61%, 46.76%, and 76.87%, respectively, compared to the control without calcium (p< 0.05).
Overall, the contents of TC, TN, TP, TK, and Ca in Pinus tabuliformis seedlings exhibited the order of TC > Ca > TN > TK > TP. The contents of other elements except for calcium reached the maximum value when the calcium concentration was between 50 and 100 mg·kg-1.
The nutrient content (unit: g) distribution characteristics of leaves, stems, and roots are shown in Figures 1F-J. Under the same calcium concentration, significant differences were observed in the contents of TC, TN, TP, TK, and Ca in different organs of Pinus tabuliformis (p< 0.05). The content of TC and TK was characterized by leaves > roots > stems, while the content of TN, TP, and Ca was characterized by roots > leaves > stems. In terms of the average proportion of each element in different organs, C and K elements were most distributed in leaves, accounting for 43.88% and 38.51% respectively, while N, P, and Ca elements were most distributed in roots, accounting for 40.40%, 36.07%, and 43.33%, respectively.
3.2 Effect of exogenous calcium addition on ecological stoichiometric ratios of various organs of Pinus tabuliformis seedlings
The ecological stoichiometric ratios (C:N, C:P, N:P) of different organs of Pinus tabuliformis seedlings under varying calcium concentrations are shown in Figures 2A-C. The addition of exogenous calcium significantly affected the leaf C:N, C:P, and N:P (except N:P) (p<0.05), presenting a “decreasing-increasing”, “decreasing-increasing”, and “increasing-decreasing” trend, respectively, with the gradual increase in added calcium concentration. The leaf C:N and C:P reached their minimum values at 100 mg·kg-1 calcium treatment. The calcium addition also significantly affected the stem C:N, C:P, and N:P (except N:P) (p<0.05), showing a “decreasing-increasing”, “decreasing”, and “decreasing” trend, respectively, with the gradual increase in added calcium concentration. The stem C:N was significantly reduced and reached the minimum at 50-100 mg·kg-1 calcium treatment compared to the control. The root C:N, C:P, and N:P were significantly affected by calcium addition (p<0.05), all exhibiting an “increasing-decreasing” trend with increasing calcium concentration, with the ratios ranging from 32.10-42.78, 267.25-412.12, and 8.01-11.51, respectively. The whole plant C:N, C:P, and N:P were also significantly affected by calcium (p<0.05), showing a “decreasing-increasing”, “decreasing”, and “increasing-decreasing” trend with increasing calcium concentration, respectively. For the whole plant, the C:N was significantly reduced and reached the minimum at 100 mg·kg-1 calcium, and the N:P was significantly increased and reached the maximum at 50-100 mg·kg-1 calcium compared to the control.
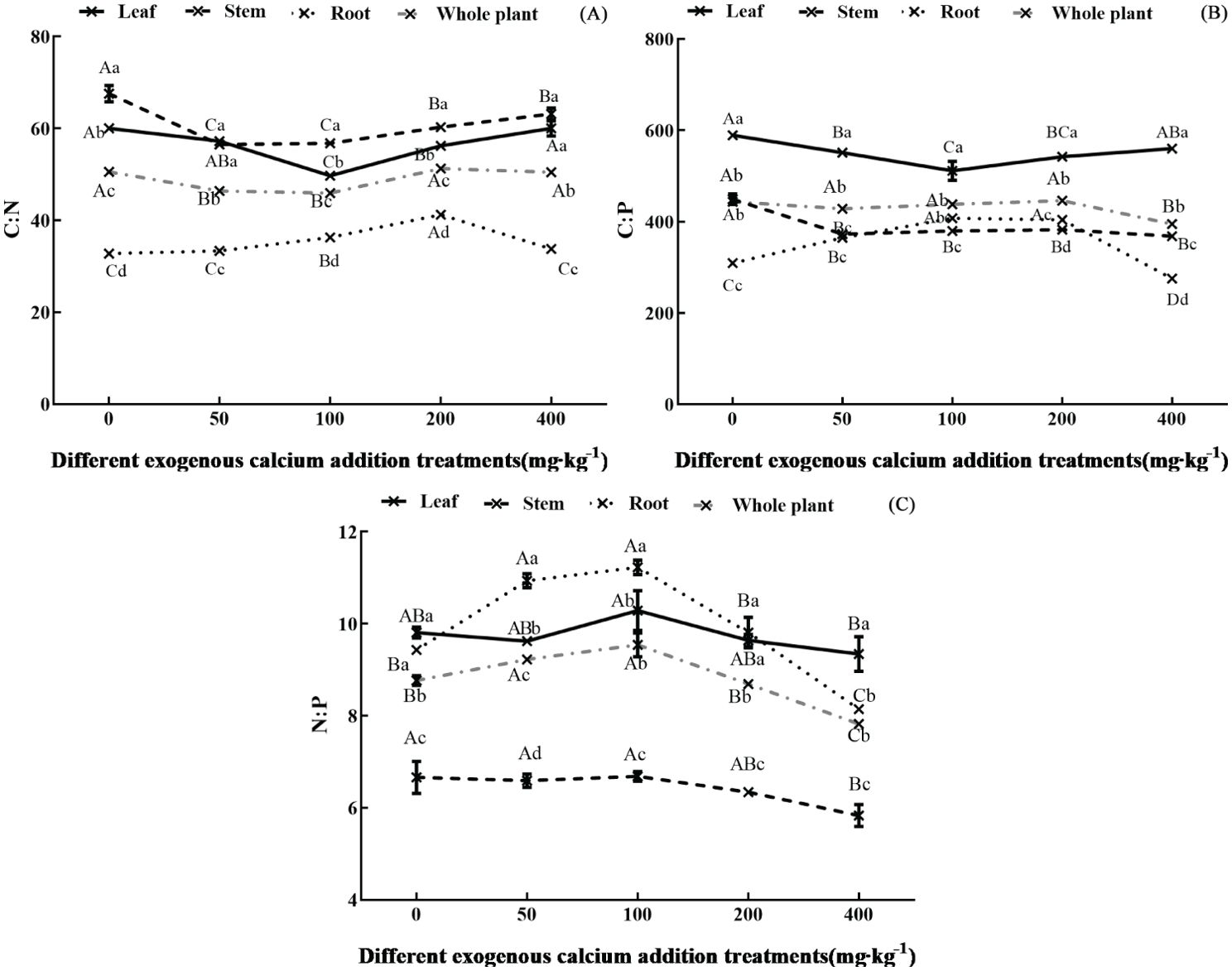
Figure 2. Ecological stoichiometric ratio of organs of Pinus tabuliformis seedlings under exogenous calcium supplementation. Capital letters indicated the difference between different calcium concentrations in leaves, stems, roots and whole plants (p<0.05), while lower letters indicated the difference between leaves, stems, roots and whole plants at the same concentration (p<0.05). (A) C:N; (B) C:P; (C) N:P.
The stoichiometric ratios differed among organs, with the overall trend being stem > leaf > whole plant > root for C:N, leaf > whole plant > stem > root for C:P, and leaf, stem, root, and whole plant showing little difference for N:P.
3.3 Effect of exogenous calcium on soil nutrient content and ecological stoichiometric ratios of Pinus tabuliformis seedlings
Table 1 presents the values of soil nutrient contents and stoichiometric ratios of Pinus tabuliformis seedlings under different calcium concentrations. With the increase of calcium concentration, soil contents of TC, TN, TP, AK, C:N, C:P, and N:P showed no significant differences (p>0.05), while soil water-soluble calcium and exchangeable calcium exhibited significant differences and showed a trend of gradually increasing content (p<0.05).
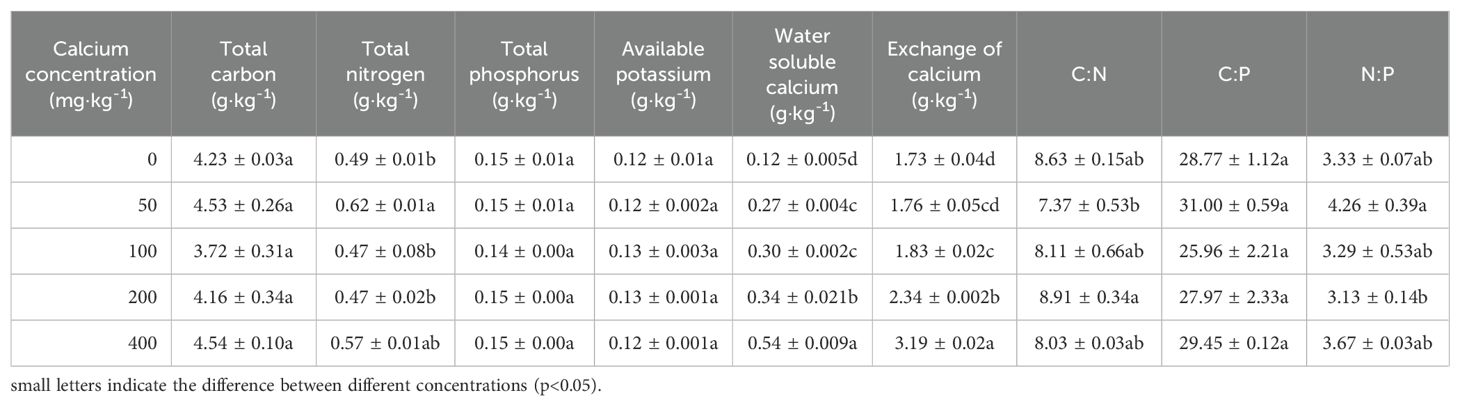
Table 1. Soil elemental contents and ecological stoichiometric ratios of Pinus tabuliformis seedlings under exogenous calcium addition.
After the application of exogenous calcium, the soil nutrient contents changed as follows: TC (3.18 g·kg-1 to 4.98 g·kg-1), TN (0.34 g·kg-1 to 0.63 g·kg-1), TP (0.13 g·kg-1 to 0.17 g·kg-1), AK (0.10 g·kg-1 to 0.14 g·kg-1), water-soluble calcium (0.11 g·kg-1 to 0.55 g·kg-1), and exchangeable calcium (1.66 g·kg-1 to 3.21 g·kg-1). Without adding exogenous calcium, the content of water-soluble calcium and exchangeable calcium was 0.12 g·kg-1 and 1.73 g·kg-1 respectively. As the calcium concentration gradually increased, their content significantly increased by 129.21% and 1.75%, 149.58% and 6.17%, 190.32% and 35.82%, 353.03% and 84.56% (p<0.05) respectively.
After the application of exogenous calcium, the range of soil C:N ratios changed from 6.46 to 9.48, C:P ratios changed from 22.13 to 32.07, and N:P ratios changed from 2.37 to 4.96. Specifically, at a calcium concentration of 200 mg·kg-1, the soil C:N ratio was maximum (8.91), and at 50 mg·kg-1, it was minimum (7.37); at a calcium concentration of 50 mg·kg-1, the soil C:P ratio was maximum (31.00), and at 100 mg·kg-1, it was minimum (25.96); at a calcium concentration of 50 mg·kg-1, the soil N:P ratio was maximum, at 4.26.
3.4 Effect of exogenous calcium on the correlation between seedling-soil ecological stoichiometric traits of Pinus tabuliformis seedlings
The Table 2 shows the correlation coefficients between the nutrient content and stoichiometric ratios of leaves, stems, roots, and whole seedlings of Pinus tabuliformis and the soil. The analysis indicates that the chemical stoichiometric characteristics of Pinus tabuliformis seedlings are correlated with the soil.
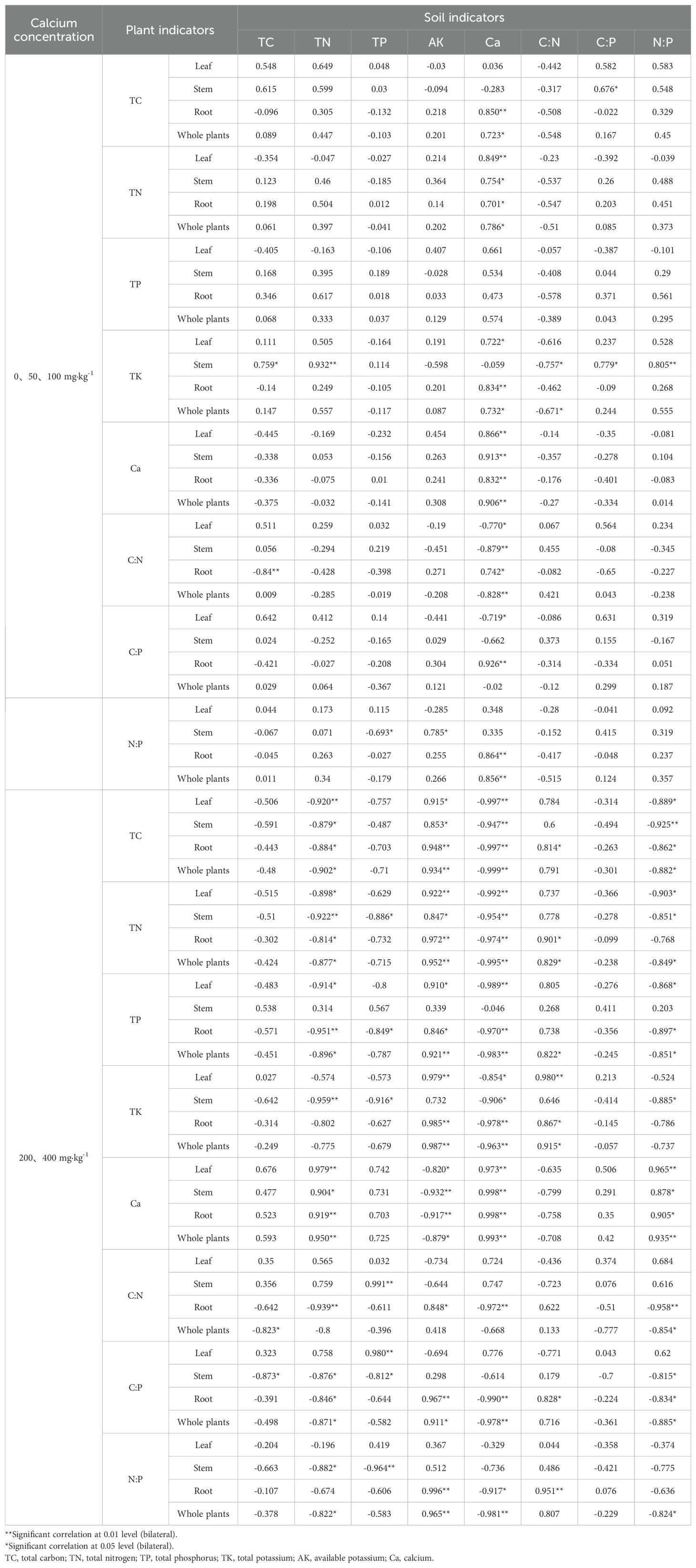
Table 2. Correlation coefficient between nutrient content and stoichiometric ratio of leaves, stems, roots and whole plants of Pinus tabuliformis seedlings and soil.
At calcium concentrations of 0, 50, and 100 mg·kg-1, soil Ca is mostly positively correlated with the nutrient (TC, TN, TP, TK, Ca) contents in leaves, stems, roots, and whole seedlings, and negatively correlated with the C:N and C:P ratios in leaves, stems, and whole seedlings, but positively correlated with the N:P ratios in leaves, stems, roots, and whole seedlings. At calcium concentrations of 200 and 400 mg·kg-1, soil Ca is positively correlated with the Ca contents in leaves, stems, roots, and whole seedlings, but negatively correlated with the TC, TN, TP, and TK contents in leaves, stems, roots, and whole seedlings, and negatively correlated with the C:N, C:P, and N:P ratios in roots, but the correlations with the C:N, C:P, and N:P ratios in leaves and stems are not significant.
The homeostasis index (1/H) calculations show that at calcium concentrations of 0, 50, and 100 mg·kg-1, the Ca in leaves, stems, roots, and whole seedlings exhibits “sensitive” homeostasis. At calcium concentrations of 200 and 400 mg·kg-1, the N in leaves, stems, roots, and whole seedlings, and the P in roots exhibit “strict homeostasis”, while the K in leaves, roots, and whole seedlings exhibits “sensitive” homeostasis, and the Ca in leaves, stems, roots, and whole seedlings exhibits “weakly sensitive” to “weakly homeostatic” homeostasis. The whole seedling N:P ratio also exhibits “strict homeostasis”, and the other elements in different organs generally exhibit “strict homeostasis”.
3.5 Response of ecological stoichiometric characteristics of Pinus tabuliformis seedlings to environmental factors under exogenous calcium supplementation
As shown in the redundancy analysis (RDA) two-dimensional ordination diagram (Figure 3), the first two axes effectively demonstrate the relationship between soil factors and the stoichiometric characteristics of leaves, stems, roots, and whole plants of oil pine seedlings. The first two axes of Figure 3 (a, b, c, and d) explain a significant portion of the variation in ecological stoichiometric characteristics. Specifically, they account for 77.19%, 82.96%, 80.85%, and 86.67% of the variation in the characteristics of leaves, stems, roots, and whole plants of Pinus tabuliformis seedlings, respectively. Furthermore, an analysis was conducted to evaluate the importance of soil environmental factors in contributing to the chemometric traits of the seedlings (see Table 3). The findings indicate that each soil factor has a varying contribution to the different chemometric traits of Pinus tabuliformis seedlings. Broadly speaking, the stoichiometric characteristics of Pinus tabuliformis seedlings are primarily influenced by the exchange calcium and water-soluble calcium, which have reached a highly significant level (p< 0.01). Additionally, the stoichiometric characteristics of Pinus tabuliformis leaves and stems are also influenced by soil TN and AK and N:P at a highly significant level (p< 0.01), with contribution rates of 10.3%, 6.8%, and 6.3%, respectively. The remaining soil factors have lower contribution rates.
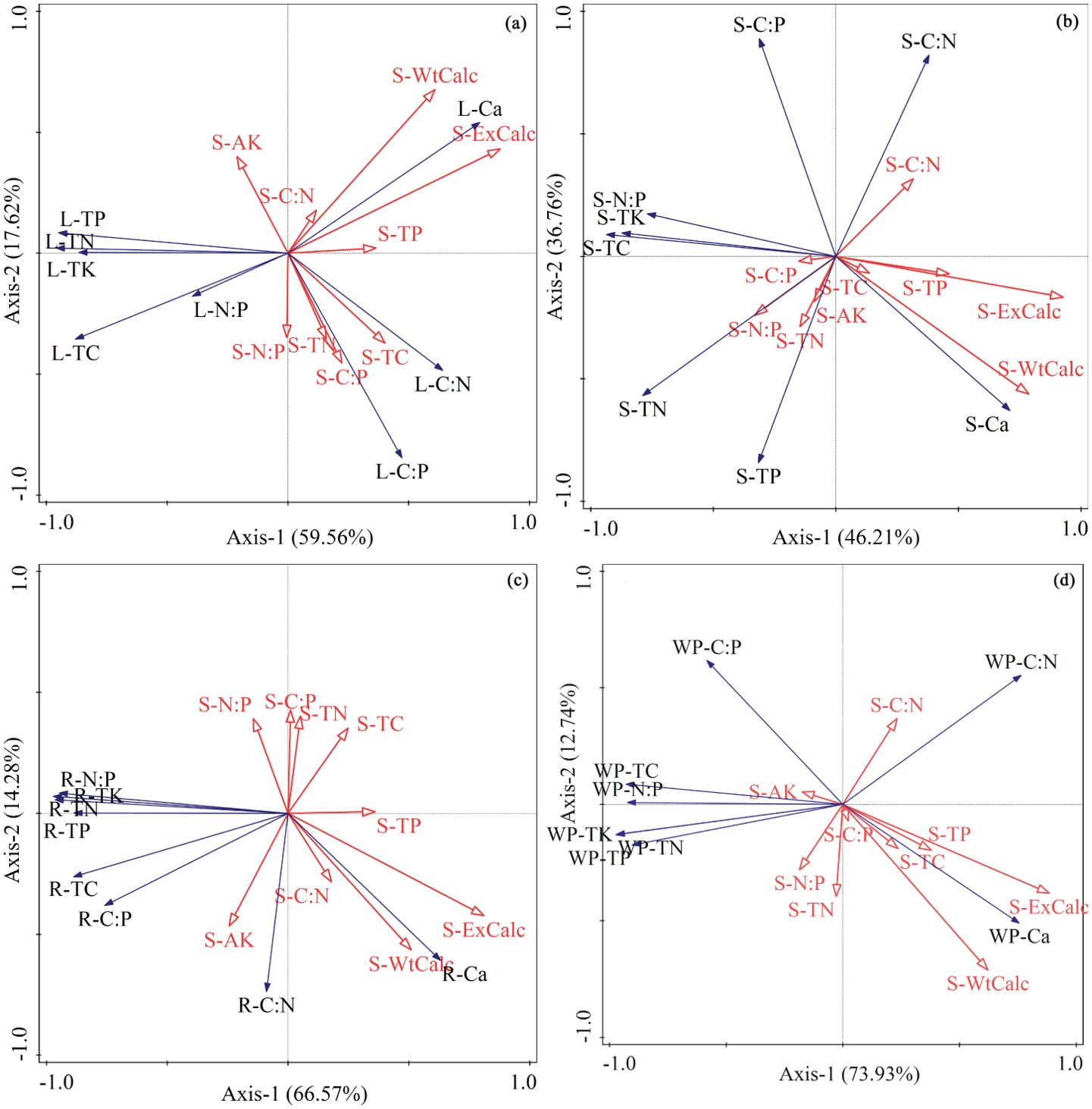
Figure 3. Two-dimensional ordination diagram of redundancy analysis of ecological stoichiometric characteristics and soil factors of Pinus tabuliformis seedlings. The blue solid arrow represents the elemental indicators of each organ, while the red hollow arrow represents soil factors. (A) Leaf; (B) Stem; (C) Root; (D) Whole plant; TC, total carbon; TN, total nitrogen; TP, total phosphorus; TK, total potassium; Ca, calcium; AK, available potassium; ExCalc, Exchange calcium; WtCalc, Water-soluble calcium.
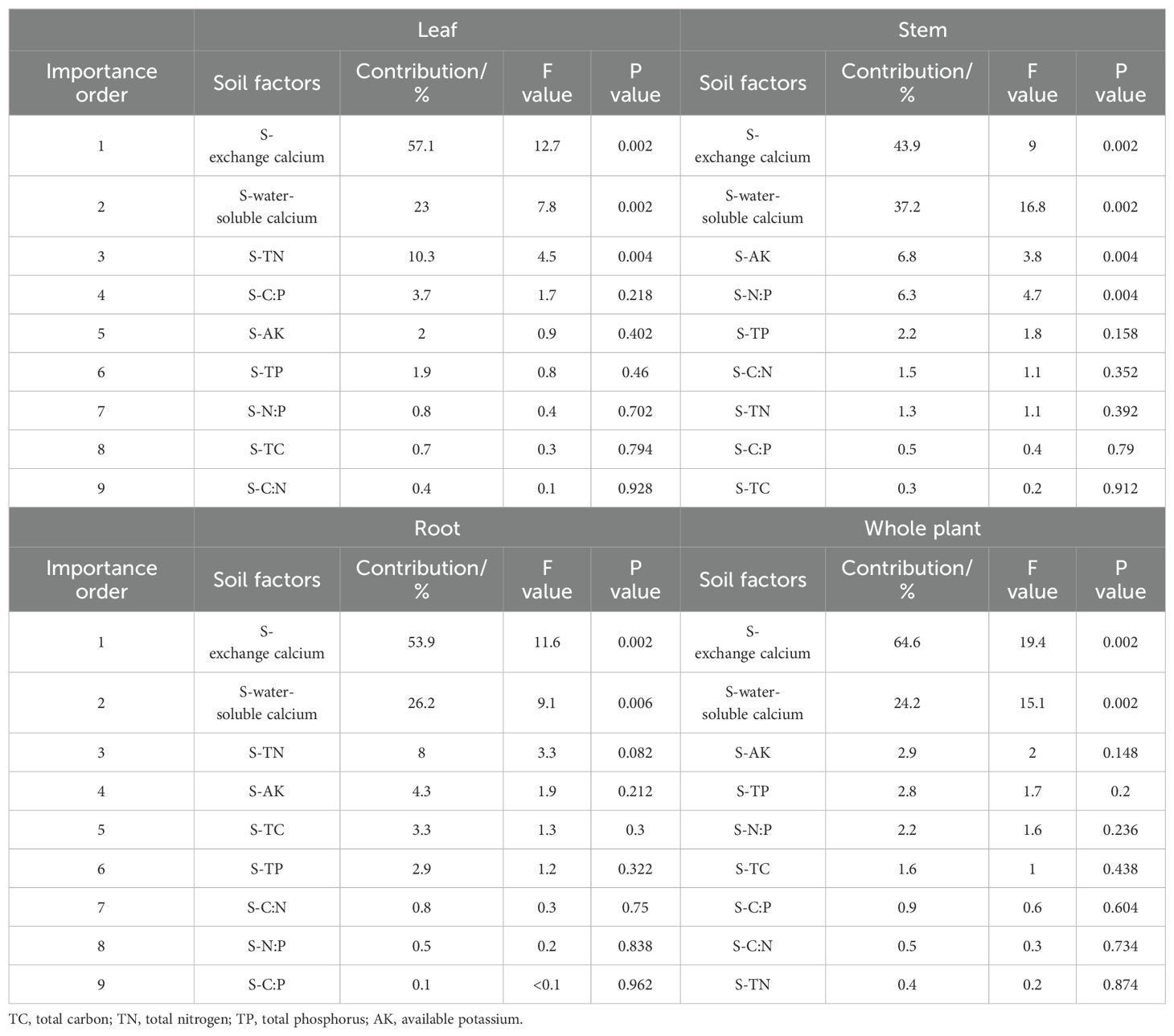
Table 3. The importance order and significance test results of environmental factors of Pinus tabuliformis seedlings.
4 Discussion
4.1 Effects of exogenous calcium on nutrient uptake and nutrient partitioning in Pinus tabuliformis seedlings
Calcium in plants serves not only as a structural component influencing cell wall rigidity, maintaining the stability and permeability of cell membranes, but also as a “second messenger” for intra- and extracellular signaling, responding to various biotic and abiotic stresses, and playing crucial roles in promoting plant growth and nutrient uptake (Liang and Zhang, 2018; Tian et al., 2020; Weng et al., 2022). Previous studies on tall fescue and poplar have shown that the addition of exogenous calcium can promote the absorption and accumulation of C, N, P, K, and Ca by plant organs, and there exists an optimal calcium concentration that maximizes the accumulation of various nutrients (Wang et al., 2017; Weng et al., 2022). Consistent with these findings, the results of this experiment demonstrate that the content of TC, TN, TP, TK, and Ca in different organs of pine seedlings significantly increased after the addition of exogenous calcium, reaching maximum values at calcium concentrations of 50-100 mg·kg-1. However, when the calcium concentration exceeds 400 mg·kg-1, its nutrient uptake effects are inhibited. This may be attributed to the excessive calcium disrupting normal biochemical and nutritional metabolism in plants, with high concentrations of calcium ions even causing cellular toxicity and abnormal plant development (Li et al., 2023a; Guo et al., 2021).
Calcium utilization efficiency calculated by Chapin’s index showed that the calcium utilization efficiency gradually decreased with the increase of added calcium concentration, which was 85.54% when no calcium was applied, and decreased to 39.78% when the calcium concentration was increased to 400 mg·kg-1, which may be due to the abundant supply of calcium nutrient in the soil, and the plant adapted to the environment by reducing the efficiency of the utilization of this nutrient to form the self-regulation mechanism mainly (Chapin, 1980; Liu et al., 2022).The elemental contents of different organs also reflect the physiological activities of plants and their adaptive strategies to the environment. In this study, based on the average proportion of various organs, the following results were obtained: in Pinus tabuliformis seedlings, the highest contents of elements C and K were found in the leaves, while elements N, P, and Ca were most abundant in the roots. When calcium was not applied, the total carbon (TC) content in the leaves of Pinus tabuliformis seedlings was 524.46 g·kg-1, higher than the global average of 464 g·kg-1 for plant leaves, with little difference compared to stems (517.37 g·kg-1) and higher than roots (317.08 g·kg-1) (Elser et al., 2000). This indicates that the ability of carbon fixation in the leaves of Pinus tabuliformis seedlings is stronger, resulting in the accumulation of more organic matter, while the storage capacity of roots is weaker. The main reason is that assimilates accumulate more in the leaves and are transported to the stems through conducting tissues, resulting in higher carbon content in stems and leaves, whereas roots mainly exchange water and inorganic salts and cannot photosynthesize (Fan et al., 2019; Huo et al., 2022). Moharam Zadeh et al. (2010) also observed the phenomenon of “most of the plant’s total carbon being stored in the leaves and stems” when evaluating carbon storage in tamarisk. Leaf, stem, and root C contents increased significantly after the application of suitable concentrations of exogenous calcium, which is consistent with the findings of Weng et al. (2022). This may be due to the fact that the applied calcium improved the photosynthetic capacity of the leaves, which in turn increased the accumulation of C in the plant (Song et al., 2020).
4.2 Effect of exogenous calcium on ecological stoichiometric ratios of Pinus tabuliformis seedlings
The Growth Rate Hypothesis (GRH) suggests that variations in the C:N:P ratio of organisms are mainly determined by changes in the phosphorus content of the biota. Plants with high growth rates require more phosphorus allocated to ribosomes for rapid protein synthesis to support rapid growth, hence they typically have lower C:N and C:P ratios (Reef et al., 2010; Elser et al., 2007). The changing trends of C:N and C:P ratios in Pinus tabuliformis seedlings in this study indicate that as calcium concentration increases, the growth rate of Pinus tabuliformis seedling roots initially slows down and then accelerates, showing an increasing trend when calcium concentration reaches 400 mg·kg-1. The growth rates of leaves, stems, and whole plants also exhibit an accelerating or initially fast and then slow trend. This might be an adaptation to changes in the growth environment. Previous studies by our research group found that when the exogenous calcium concentration exceeds 400 mg·kg-1, the Fv/Fm value of Pinus tabuliformis seedlings drops below 0.8, indicating that the plants are under stress and growth and development are severely constrained. Therefore, when calcium concentration is too high, seedlings allocate more nutrients to root growth to avoid stress (Li et al., 2023a). Furthermore, the C:N ratio in Pinus tabuliformis seedlings in this study shows a hierarchy of stem (60.86) > leaf (56.65) > whole plant (48.91) > root (35.47), while the C:P ratio shows a hierarchy of leaf (550.38) > whole plant (429.81) > stem (390.05) > root (351.79), all of which are higher than the global average levels of C:N and C:P in plants (22.5 and 232.0) (Elser et al., 2000). This indicates that Pinus tabuliformis seedlings have good carbon storage capacity, as well as efficient N and P element utilization, reflecting a relative scarcity of N and P elements required for the growth and development of Pinus tabuliformis seedlings in the region (Wei et al., 2021b; Isanta-Navarro et al., 2022). This is consistent with the assertion by Zhang et al. (2018) that the soils in the western Liaoning region are impoverished, with N and P contents below the global average levels. Moreover, the fact that leaf and stem C:N and C:P ratios are higher than those of the roots suggests that the roots have a faster growth rate, which is advantageous for more comprehensive nutrient absorption from deeper soil layers (Ma et al., 2017).
As the exogenous calcium concentration increased, there was no significant difference in the N:P ratio of Pinus tabuliformis leaves and stems, but the N:P ratio of roots and the whole plant showed a significant “increase-decrease” trend, which also implies that plants will adjust their stoichiometry to adapt to environmental changes. The N:P ratio within plant tissues reflects environmental factors, especially the nutrient supply from the soil for plant growth, and it can clarify which factors limit plant productivity (Aerts and Chapin, 1999; Güsewell, 2004). Yu et al. (2014) found that when the N:P ratio of needles is< 14, plant growth is limited by N; when N:P = 14-16, plant growth is co-limited by N and P; and when N:P > 16, plant growth is limited by P. In this study, the N:P ratio of Pinus tabuliformis seedlings was less than 14, indicating that growth was limited by soil N. Within the calcium concentration range of 50-100 mg·kg-1, the N:P ratio was relatively large, indicating that the availability of soil N was higher and the degree of N limitation was relatively light, and as the calcium concentration increased, the limiting effect strengthened, which also indicates that the calcium concentration of 50-100 mg·kg-1 was more suitable for the growth of Pinus tabuliformis. However, the factors affecting plant N:P values are complex, and it is unreliable to determine the limiting factors in the plant growth process based on a single indicator. Therefore, soil environmental factors should be introduced as auxiliary comprehensive evaluation (Güsewell, 2004). The results of redundancy analysis show that each soil factor has different contribution rates to the various stoichiometric characteristics of Pinus tabuliformis seedlings. And previous studies have shown that the absorption of C and other nutrients (P, K, Ca, and Mg) is doubly regulated by stoichiometry and nutrient limitation control, and the nutrient absorption of coniferous trees mainly depends on stoichiometric control rather than nutrient limitation control (Sun et al., 2023).
The stoichiometric steady state in ecology provides a more accurate representation of the physiological and biochemical adaptation of organisms to environmental changes. Research results indicate that there are differences in the elemental dynamic balance indicators and stoichiometric ratios of leaves, stems, roots, and whole plants. This is similar to the findings of Jia et al. (2023), suggesting a trade-off between nutrient uptake and allocation, indicating that the stoichiometric characteristics of plants in soil differ among plant tissues and elemental types. Apart from Ca and some K elements, the internal steady-state model equations did not significantly simulate most indicators (p > 0.05) or 1/H< 0, indicating a strong ecological stoichiometric balance with no clear trend in response to changes in soil nutrient environments. This is consistent with the research results of Su et al. (2022); You (2022), and Wang et al. (2022), who observed “strict internal stability” in plants, likely reflecting more on species-specific traits or genetic features (Tao and Zhang, 2015; Erfan et al., 2022). Various factors influence the dynamic balance of ecological stoichiometry, such as soil environmental factors, plant structural characteristics, geography and climate, external disturbances, and human activities (Bai et al., 2019; Zhang et al., 2019). Therefore, continuous monitoring and management of the long-term effects of exogenous calcium on the soil are necessary.
5 Conclusion
A well-regulated application of calcium can significantly enhance overall nutrient uptake and accumulation within plants, encompassing their roots, stems, and leaves. The impact of exogenous calcium on plant ecological stoichiometry is greater than that of the soil. This external calcium supplementation facilitates enhanced nutrient absorption and regulates the inter-organ transport of nutrients. Its effectiveness peaks within a specific range, typically between 50-100 mg·kg-1. However, excessive exogenous calcium inhibits the absorption of nutrient elements in various plant organs, indicating an adverse calcium stress effect on Pinus tabuliformis seedlings. Furthermore, different plant organs exhibit varying stabilities in response to changes in soil conditions, with most indicators demonstrating robust internal stability. Therefore, there exists an optimal calcium concentration for the nutrient uptake and stoichiometric balance of Pinus tabuliformis seedlings. The results of this study provide a theoretical basis for future research on Pinus tabuliformis plantation forests.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
HL: Writing – review & editing. YZ: Writing – original draft. XW: Data curation, Writing – review & editing. YZ: Resources, Writing – review & editing. YH: Supervision, Writing – review & editing. SZ: Supervision, Writing – review & editing. LL: Supervision, Writing – review & editing. JP: Project administration, Supervision, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This research was supported by the Natural Science Foundation of China-Research (No. 3170030160, No. 41450007, No.31800364, No.31400611) and the Doctoral research start-up fund (No.880416020).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Aerts, R., Chapin, III, F. S. (1999). “The mineral nutrition of wild plants revisited: a re-evaluation of processes and patterns,” in Advances in ecological research, vol. 30, 1–67. Academic Press
Aras, S., Keles, H., Bozkurt, E. (2021). Physiological and histological responses of peach plants grafted onto different rootstocks under calcium deficiency conditions. Scient. Horticult. 281, 109967. doi: 10.1016/j.scienta.2021.109967
Arkley, R. J. (1963). Calculation of carbonate and water movement in soil from climatic data. Soil Sci. 96, 239–248. doi: 10.1097/00010694-196310000-00003
Bai, X., Wang, B., An, S., Zeng, Q., Zhang, H. (2019). Response of forest species to c:n:p in the plant-litter-soil system and stoichiometric homeostasis of plant tissues during afforestation on the loess plateau, China. Catena 183, 104186. doi: 10.1016/j.catena.2019.104186
Barroso, G. M., Duque, T. S., Custódio, I. G., Leite, M. L. F., Corrêa, J. M., Silva, D. V., et al. (2022). Expansion of planted forests: The risk of pesticides mixtures. Forests 13, 50. doi: 10.3390/f13010050
Certini, G., Scalenghe, R. (2006). “Soil formation on Earth and beyond: the role of additional soil forming factors,” in Soils. Basic Concepts and Future Challenges, 193–210, Cambridge University Press.
Chapin, III FS (1980). The mineral nutrition of wild plants. Annu. Rev. Ecol. Systematics 1, 233–260. doi: 10.1146/annurev.es.11.110180.001313
Chapin, F. S., Vitousek, P. M., Vancleve, K. (1986). The nature of nutrient limitation in plant-communities. Am. Nat. 127, 48–58. doi: 10.1086/284466
da Silveira Sousa Junior, G., Hurtado, A. C., de Souza Junior, J. P., de Mello Prado, R., de Cássia Piccolo, M., Dos Santos, D. M. M. (2022). Beneficial role of silicon on regulating C, N, and P stoichiometric homeostasis and the growth of sugarcane seedlings under aluminum toxicity. J. Soil Sci. Plant Nutr. 22, 4138–4152. doi: 10.1007/s42729-022-01013-5
Ding, D., Arif, M., Liu, M., Li, J., Hu, X., Geng, Q. (2022). Plant-soil interactions and C:N:P stoichiometric homeostasis of plant organs in riparian plantation. Front. Plant Sci. 13. doi: 10.3389/fpls.2022.979023
Elser, J. J., Bracken, M. E. S., Cleland, E. E., Gruner, D. S., Harpole, W. S., Hillebrand, H., et al. (2007). Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 10, 1135–1142. doi: 10.1111/j.1461-0248.2007.01113.x
Elser, J. J., Fagan, W. F., Denno, R. F., Dobberfuhl, D. R., Folarin, A., Huberty, A., et al. (2000). Nutritional constraints in terrestrial and freshwater foodwebs. Nature 408, 578–580. doi: 10.1038/35046058
Elser, J. J., Fagan, W. F., Kerkhof, A. J., Swenson, N. G., Enquist, B. J. (2010). Biological stoichiometry of plant production: metabolism, scaling and ecological response to global change. New Phytol. 186, 593–608. doi: 10.1111/j.1469-8137.2010.03214.x
Erfan, A., Li, J., Zhuang, W. (2022). Relationship between habitat soil factor and stoichiometric characteristics of two kinds of desert leguminous plants. Acta Botanica Boreali-Occidentalia Sin. 42, 1384–1395. doi: 10.7606/j.issn.1000-4025.2022.08.138
Ermakov, V., Bech, J., Gulyaeva, U., Tyutikov, S., Safonov, V., Danilova, V., et al. (2020). Relationship of the mobile forms of calcium and strontium in soils with their accumulation in meadow plants in the area of Kashin-Beck endemia. Env. Geochem. Health 42, 159–171. doi: 10.1007/s10653-019-00323-5
Fan, Y., Pan, Y. L., Chen, Z. W., Lin, H., Xu, R., Wu, C. Z., et al. (2019). C:N: P stoichiometry in roots, stems, and leaves of four mangrove species. Chin. J. Ecol. 38, 1041–1048. doi: 10.13292/j.1000−4890.201904.015
Guo, Y., Liu, Y., Zhang, Y., Liu, J., Gul, Z., Guo, X. R., et al. (2021). Effects of exogenous calcium on adaptive growth, photosynthesis, ion homeostasis and phenolics of gleditsia sinensis lam. plants under salt stress. Agriculture 11, 978. doi: 10.3390/agriculture11100978
Güsewell, S. (2004). N: P ratios in terrestrial plants: variation and functional significance. New Phytol. 164, 243–266. doi: 10.1111/j.1469-8137.2004.01192.x
He, C., Wang, F., Li, Y., Li, Q., Lin, C. (2012). Effects of the combined application of organic fertilizer and calcium fertilizers on characteristics of the calcium leaching, migration and transformation in the AeolianSandy soil of coastal farming. Fujian Agric. J. 27, 533–538. doi: 10.19303/j.issn.1008-0384.2012.05.015
Hood, J. M., Sterner, R. W. (2010). Diet mixing: do animals integrate growth or resources across temporal heterogeneity? Am. Nat. 176, 651–663. doi: 10.1086/656489
Hu, W., Liu, J., Liu, T., Zhu, C., Wu, F., Jiang, C., et al. (2022). Exogenous calcium regulates the growth and development of Pinus massoniana detecting by physiological, proteomic, and calcium-related genes expression analysis. Plant Physiol. Biochem. 196, 1122–1136. doi: 10.22541/au.166029509.94039896/v1
Huo, H., Cao, X., Liu, J., Wang, F., Qi, R., Zhao, Y., et al. (2022). Stoichiometric characteristics of C, N, P, and K in different organs of two secondary forests in Bailongjiang, Gannan. J. Northwest Forestry University. 37, 64−72. doi: 10.3969/j.issn.1001-7461.2022.04.0
Isanta-Navarro, J., Prater, C., Peoples, L. M., Loladze, I., Phan, T., Jeyasingh, P. D., et al. (2022). Revisiting the growth rate hypothesis: Towards a holistic stoichiometric understanding of growth. Ecol. Lett. 25, 2324–2339. doi: 10.1111/ele.14096
Jia, T., Yi, M., Chen, F., Lai, M., Jin, C., Nie, Z., et al. (2023). Response of C: N: P Stoichiometry to phosphorus addition and homeostasis of plant tissues in a Subtropical Slash Pine Plantation. Forests 14, 1355. doi: 10.3390/f14071355
Lawrence, G. B., Scanga, S. E., Sabo, R. D. (2020). Recovery of soils from acidic deposition may exacerbate nitrogen export from forested watersheds. J. Geophys. Res.-Biogeosci. 125, e2019JG005036. doi: 10.1029/2019JG005036
Li, W., Ali, I., Han, X., Ullah, S., Yang, M. (2023b). Soil C, N, P, K and enzymes stoichiometry of an endangered tree species, Parashorea chinensis of different stand ages unveiled soil nutrient limitation factors. Forests 14, 624. doi: 10.3390/f14030624
Li, M., Fang, L., Duan, C., Cao, Y., Yin, H., Ning, Q., et al. (2020). Greater risk of hydraulic failure due to increased drought threatens pine plantations in Horqin Sandy land of northern China. For. Ecol. Manage. 461, 117980. doi: 10.1016/j.foreco.2020.117980
Li, H., Huo, Y., Weng, X., Zhou, Y., Sun, Y., Zhang, G., et al. (2022a). Regulation of the growth of Mongolian pine (Pinus sylvestris var. mongolica) by calcium-water coupling in a semiarid region. Ecol. Indic. 137, 108736. doi: 10.1016/j.ecolind.2022.108736
Li, Z., Liu, Z., Yue, Z., Wang, J., Jin, L., Xu, Z., et al. (2022b). Application of exogenous silicon for alleviating photosynthetic inhibition in tomato seedlings under low– calcium stress. Int. J. Mol. Sci. 23, 13526. doi: 10.3390/ijms232113526
Li, H., Zhao, Y., Weng, X., Zhou, Y., Zhang, S., Liu, L., et al. (2023a). The most suitable calcium concentration for growth varies among different tree species—Taking Pinus tabuliformis, Pinus sylvestris var. mongolica, Populus, and Morus alba as examples. Forests 14, 1437. doi: 10.3390/f14071437
Liang, C., Zhang, B. (2018). Effect of exogenous calcium on growth, nutrients uptake and plasma membrane h+ -ATPase and Ca2+-ATPase activities in soybean (Glycine max) seedlings under simulated acid rain stress. Ecotoxicology Environ. Saf. 165, 261–269. doi: 10.1016/j.ecoenv.2018.09.019
Liao, J. J., Dou, Y. X., An, S. S. (2023). Plant community productivity is associated with multiple ecological stoichiometry in restoration grasslands. Ecol. Eng. 187, 106845. doi: 10.1016/j.ecoleng.2022.106845
Liu, S., An, H., Zhang, X., Du, Z., Liu, X. (2022). Effects of nitrogen and phosphorous addition on the ecological stoichiometry of plant-litter-soil in desert grassland. Acta Ecologica Sin. 42, 8773–8783. doi: 10.5846/stxb202110293043
Liu, N., Bao, G., Bao, M. (2019). Response characteristics of Chinese pine (Pinus tabulaeformis carr.) radial growth to climate and drought variability reconstruction in western Liaoning, northeast China. Forests 10, 752. doi: 10.3390/f10090752
Liu, R., Yao, Y., Guo, Z., Li, Q., Zhang, S. (2023). Plantation rhizosphere soil microbes promote soil−plant phosphorus feedback on the Tibetan Plateau. Plant Soil. 501, 191–209. doi: 10.1007/s11104-023-05939-2
Long, M., Guo, L., Li, J., Yu, C., Hu, T., Yue, J., et al. (2018). Effects of water and exogenous Si on element concentrations and ecological stoichiometry of plantain (Plantago lanceolata L.). J. Plant Nutr. 41, 1263–1275. doi: 10.1080/01904167.2018.1443128
Ma, F., Xu, T. T., Liu, J. L., Xiao, G. J., Li, M., Bi, J. T., et al. (2017). Variations in carbon, nitrogen and phosphorus stoichiometry of Caragana liouana originated from nine provenances in a common garden. Acta Bot. Boreal. -Occident. Sin. 37, 1381–1389. doi: 10.7606/j.issn.1000-4025.2017.07.1381
Moharam Zadeh, M., Feyznia, S., Jafari, M., Zehtabian, G. H., Akbarpoor, M. B., Seyed Mosavi, M. S., et al. (2010). “The amount of carbon stored in Tamarix aphylla,” in First National Conference of Iranian natural resources research, October 2010 (in Persian).
Paiva, E. A. S., Sampaio, R. A., Martinez, H. E. P. (1998). Composition and quality of tomato fruit cultivated in nutrient solutions containing different calcium concentrations. J. Plant Nutr. 21, 2653–2661. doi: 10.1080/01904169809365595
Persson, J., Fink, P., Goto, A., Hood, J. M., Jonas, J., Kato, S. (2010). To be or not to be what you eat: Regulation of stoichiometric homeostasis among autotrophs and heterotrophs. Oikos 119, 741–751. doi: 10.1111/j.1600-0706.2010.18545.x
Qi, K., Pang, X., Yang, B., Bao, W. (2020). Soil carbon, nitrogen and phosphorus ecological stoichiometry shifts with tree species in subalpine plantations. PeerJ 8, e9702. doi: 10.7717/peerj.9702
Reef, R., Ball, M. C., Feller, I. C., Lovelock, C. E. (2010). Relationship among RNA : DNA ratio, growth and elemental stoichiometry in mangrove trees. Funct. Ecol. 24, 1064–1071. doi: 10.1111/j.1365-2435.2010.01722.x
Sarto, M. V. M., do Carmo Lana, M., Rampim, L., Rosset, J. S., Sarto, J. R. W., Bassegio, D. (2019). Effects of calcium and magnesium silicate on the absorption of silicon and nutrients in wheat. Semina: Ciências Agrárias. 40, 67–80. doi: 10.5433/1679-0359.2019v40n1p67
Song, Q., Liu, Y., Pang, J., Yong, J. W. H., Chen, Y., Bai, C., et al. (2020). Supplementary calcium restores peanut (Arachis hypogaea) growth and photosynthetic capacity under low nocturnal temperature. Front. Plant Sci. 10. doi: 10.3389/fpls.2019.01637
Sterner, R. W., Elser, J. J. (2002). Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere. Princeton (Oxford: Princeton University Press).
Su, Z., Su, B., Mao, S., Shangguan, Z. (2022). Leaf C:N:P stoichiometric homeostasis of a Robinia pseudoacacia plantation on the Loess Plateau. J. For. Res. 34, 929–937. doi: 10.1007/s11676-022-01541-x
Sun, X. B., Li, D. J., Lü, X. T., Fang, Y. T., Ma, Z. L., Wang, Z. C., et al. (2023). Widespread controls of leaf nutrient resorption by nutrient limitation and stoichiometry. Funct. Ecol. 37, 1653–1662. doi: 10.1111/1365-2435.14318
Tao, Y., Zhang, Y. (2015). Leaf and soil stoichiometry of four herbs in the Gurbantunggut Desert, China. Chinese. J. App. Ecol. 26, 659–665. doi: 10.13287/j.1001-9332.20150106.007
Tian, W., Wang, C., Gao, Q., Li, L., Luan, S. (2020). Calcium spikes, waves and oscillations in plant development and biotic interactions. Nat. Plants. 6, 1–10. doi: 10.1079/9780851995120.0001
Wang, G. Y., Bi, A. Y., Amombo, E., Li, H. Y., Zhang, L., Cheng, C., et al. (2017). Exogenous calcium enhances the photosystem II photochemistry response in salt stressed tall fescue. Front. Plant Sci. 8. doi: 10.3389/fpls.2017.02032
Wang, Y., Chen, Y., Sun, Y., Zhao, M., Xue, W., Liu, L. (2022). Seasonal variation of plant organ-litter-soil stoichiometry in Pinus tabulaeformis plantation in loess hilly region. J. Soil Water Conserv. 36, 350–356. doi: 10.13870/j.cnki.stbcxb.2022.04.043
Wei, Y., Dang, X., Wang, J., Gao, J., Gao, Y. (2021a). Response of C: N: P in the plant-soil system and stoichiometric homeostasis of Nitraria tangutorum leaves in the oasis-desert ecotone, Northwest China. J. Arid Land. 13, 934–946. doi: 10.1007/s40333-021-0019-z
Wei, Y., Wang, J., Dang, X., Han, Y., Gao, Y., Li, P., et al. (2021b). Contents and stoichiometric characteristics of C, N, P and K in leaves process of Nitraria tangutorum succession. J. Cent. South Univ. Forestry Technol. 41, 102–110,139. doi: 10.14067/j.cnki.1673-923x.2021.10.012
Weng, X., Li, H., Ren, C., Zhou, Y., Zhu, W., Zhang, S., et al. (2022). Calcium regulates growth and nutrient absorption in poplar seedlings. Front. Plant Sci. 13. doi: 10.3389/fpls.2022.887098
Wu, X., Wang, X., Wang, P., Gu, Y., Li, Y. (2023). Effects of groundwater depth on ecological stoichiometric characteristics of assimilated branches and soil of two desert plants. Front. Plant Sci. 14. doi: 10.3389/fpls.2023.1225907
Xu, L., Xing, A., Du, E., Shen, H., Yan, Z., Jiang, L., et al. (2021). Effects of nitrogen addition on leaf nutrient stoichiometry in an old-growth boreal forest. Ecosphere 12, e03335. doi: 10.1002/ecs2.3335
Yin, Y., Li, Q., Du, H. (2021). Near-natural transformation of Pinus tabuliformis better improve soil nutrients and soil microbial community. Peer J. 9. doi: 10.7717/peerj.12098
You, H. M. (2022). Effects of N addition on carbon and nitrogen stoichiometry and homeostasis characteristics of the Kandelia obovata plants-soil-microbial biomass system. Chin. J. Ecol. 41, 1909–1915. doi: 10.13292/j.1000-4890.202210.024
Yu, Y., Peng, W., Song, T., Zeng, F., Wang, K., Wen, L., et al. (2014). Stoichiometric characteristics of plant and soil C, N and P in different forest types in depressions between karst hills, Southwest China. Chin. J. Appl. Ecol. 25, 947–954. doi: 10.13287/j.1001-9332.2014.0068
Zeng, X., Wei, C., Liu, X., Zhang, L. (2020). Qinghai spruce (Picea crassifolia) and Chinese pine (Pinus tabuliformis) show high vulnerability and similar resilience to early-growing-season drought in the Helan Mountains, China. Ecol. Indic. 110. doi: 10.1016/j.ecolind.2019.105871
Zhang, J., Fu, B., Stafford Smith, M., Wang, S., Zhao, W. (2021). Improve forest restoration initiatives to meet Sustainable Development Goal 15. Nat. Ecol. Evol. 5, 10–13. doi: 10.1038/s41559-020-01332-9
Zhang, T., Li, Y., Han, F., Zhang, S., Zhou, Y. (2018). C: N: P stoichiometry of Pinus tabuliformis leaf and soil in two mixed stands in western Liaoning Province. Chin. J. Ecol. 37, 3061−3067. doi: 10.13292/j.1000-4890.201810.015
Zhang, W., Liu, W. C., Xu, M. P., Deng, J., Han, X. H., Ren, G. X., et al. (2019). Response of forest growth to C:N:P stoichiometry in plants and soils during Robinia pseudoacacia afforestation on the Loess Plateau, China. Geoderma 337, 280–289. doi: 10.1016/j.geoderma.2018.09.042
Zhou, Y., Zou, X. (2017). From matching site with trees towards matching calcium with trees. J. Nanjing Forestry Univ. (Natural Sci. Edition) 41, 1–8. doi: 10.3969/j.issn.1000-2006.2017.02.001
Keywords: semi-arid region, exogenous calcium, Pinus tabuliformis, ecological stoichiometry, three north protected forests
Citation: Li H, Zhao Y, Weng X, Zhou Y, Huo Y, Zhang S, Liu L and Pei J (2024) Effects of exogenous calcium additions on the ecological stoichiometric characteristics of various organs and soil nutrients and their internal stability in Pinus tabuliformis. Front. Plant Sci. 15:1428011. doi: 10.3389/fpls.2024.1428011
Received: 05 May 2024; Accepted: 16 August 2024;
Published: 05 September 2024.
Edited by:
Anoop Kumar Srivastava, Central Citrus Research Institute (ICAR), IndiaReviewed by:
Li Xiangyi, Chinese Academy of Sciences (CAS), ChinaSeyed Majid Mousavi, Soil & Water Research Institute, Iran
Copyright © 2024 Li, Zhao, Weng, Zhou, Huo, Zhang, Liu and Pei. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jiubo Pei, cGVpaml1Ym9Ac3lhdS5lZHUuY24=