 Jiamin Hao1†
Jiamin Hao1† Zhangchen Zhao1†Xiaoge Fu1Yujiao Zhao1Muhammad Ateeq1Liming Mou2Yong Han3Yangbin Liu4Yongan Yin5
Zhangchen Zhao1†Xiaoge Fu1Yujiao Zhao1Muhammad Ateeq1Liming Mou2Yong Han3Yangbin Liu4Yongan Yin5 Lyudmila Zotova6
Lyudmila Zotova6 Dauren Serikbay6Chunyan Fan7
Dauren Serikbay6Chunyan Fan7 Yin-Gang Hu1,8*Liang Chen1*
Yin-Gang Hu1,8*Liang Chen1*- 1State Key Laboratory of Crop Stress Resistance and High-Efficiency Production and College of Agronomy, Northwest A&F University, Yangling, Shaanxi, China
- 2Dingxi Academy of Agricultural Sciences, Dingxi, Gansu, China
- 3Jiushenghe Seed Industry Co., Ltd, Changji, Xinjiang, China
- 4Yangling Digital Agricultural Technology Co., Ltd, Yangling, Shaanxi, China
- 5Shaanxi Grain and Agriculture Group Co., Ltd., Xi’an, Shaanxi, China
- 6Faculty of Agronomy, S. Seifullin Kazakh Agro Technical Research University, Astana, Kazakhstan
- 7Baoji Academy of Agricultural Sciences, Baoji, China
- 8Institute of Water Saving Agriculture in Arid Regions of China, Northwest A&F University, Yangling, Shaanxi, China
The introduction of dwarfing genes triggered a wave of “green revolution”. A number of wheats dwarfing genes have been reported in previous studies, and only a small fraction of these have been applied to production practices. Therefore, the development of novel dwarfing genes for wheat is of great value. In this study, a novel dwarfing site, Rht-yz, identified in the Yanzhan mutation, is located on chromosome 4B (30-33MB) and its mechanism of action is different from that of Rht-B1b (C-T mutation), but whether it affects the Rht-B1a (TraesCS4B02G043100) or other genes is unclear. Exogenously applied GA3 experiments showed that Rht-yz is one of the gibberellin-insensitive dwarf genes. The effects of the dwarf gene Rht-yz on agronomic traits in wheat were evaluated in the field using Yanzhan, Yanzhan mutations, F2:3 and F3:4 lines. The results showed that Rht-yz improved lodging resistance by reducing plant height, increasing diameter, wall thickness and mechanical strength of the basal stem. In terms of yield traits, Rht-yz had negative effects on tiller number plant-1, biomass plant-1 and yield plant-1, but had no significant effect on harvest index, 1000-kernel weight and spike traits. In addition, Rht-yz significantly increased crude protein, wet gluten and starch content. Therefore, the rational use of the new dwarfing site Rht-yz has potential and value in dwarf wheat breeding.
Introduction
Wheat (Triticum aestivum L.) is an important cereal crop playing outstanding role in feeding the hunger world and global food security. Plant height is one of the main agronomic traits for morphogenesis affecting wheat yield. In the “Green Revolution” of late 20th century, Rht-B1b and Rht-D1b significantly reduced plant height in wheat and increased wheat production (Hedden, 2003). Wheat varieties carrying dwarf genes Rht-B1b and Rht-D1b (Chihoumu and Dharma) rapidly expanded the planting area, and the wheat dwarf gene was gradually used in wheat breeding. To date, more than 25 dwarf genes have been discovered. The mainly used wheat dwarf genes were Rht-B1b, Rht-D1b and Rht8, which limited the improvement in wheat yield potential. Rht-B1b/Rht-D1b reduced plant height by 23%, and Rht-B1b+Rht-D1b reduced plant height by 47% (Richards, 1992), but Rht-B1b and Rht-D1b had certain adverse effects on seedling traits. The dwarf gene Rht-B1b and Rht8 also had a significant effect on the tillers number plant-1 in wheat. Rht-B1b reduced 1000-kernel weight but increased yield to eliminate the negative effect of small biomass (Hayat et al., 2019). Meanwhile, Rht-B1b and Rht-D1b associated with reduction in seed size and protein content (Casebow et al., 2016).
Furthermore, the dwarf gene Rht3 has a three times greater effect on plant height than Rht-B1b, but plants that are too short are not conducive to yield formation (Gale et al., 1985; Flintham et al., 2000; Derkx et al., 2017). Rht5 reduced plant height by 25.84%, but had a negative effect on spike development (Daoura et al., 2014). Rht13 reduces plant height by 21.5% but has a negative effect on internode diameter (Wang et al., 2014). Therefore, many dwarf genes are difficult to be applied in wheat breeding.
Statistics show that about 70% of the wheat varieties released worldwide contain Rht-B1b or/and Rht-D1b (Evans, 2008). During 2012-2018, Rht-D1b was present at a frequency of more than 80% in Uniform Southern Soft Red Winter Wheat Nursery (Hayat et al., 2019). In addition, more than half of breeding lines in SRWW in the southern United States carry Rht-D1b, and approximately 31% of breeding lines carry Rht-B1b (Guedira et al., 2010). Rht-B1b and Rht-D1b were also widely distributed in Chinese wheat, the genetic frequencies of Rht-D1b, Rht8 and Rht-B1b were 45.5%, 46.8% and 24.5%, respectively (Zhang et al., 2006).
However, the widespread distribution of single dwarf source is not beneficial to the sustainable development of wheat breeding, which reduces the diversity of dwarf sources and the safety of wheat production. Wheat dwarf breeding requires diversified dwarf genes. Therefore, it is important to identify new wheat dwarf genes and determine their application value for wheat dwarf breeding. The aim of this study was to identify the novel wheat dwarfing site Rht-yz, and to clarify its effects on wheat agronomic traits by measuring plant height, internode traits, lodging resistance traits, yield and grain traits.
Materials and methods
Materials and field experiment
The Yanzhan 4110 (followed as Yanzhan) and natural dwarf mutants of Yanzhan 4110 (followed as Yanzhan mutation) provided by Dong Puhui’s research group at Henan University of Science and Technology (HAUST), were crossed to produce F2:3 and F3:4 populations, including 15 dwarf lines and 15 tall lines. Field trials were conducted in 2019-2020 and 2020-2021 at the experimental farm of the Institute of Water-Saving Agriculture for Arid Areas of China, Northwest A&F University, Yangling, Shaanxi, China. The experimental design was randomized complete block design (RCBD), and each line was planted in 2.5m2 sized plots (five rows of 2 m long, 25 cm between two rows, and 6.7 cm between two plants). Each plot was repeated three times.
The GA3 treatment was applied separately on both Yanzhan and Yanzhan mutation, and repeated three times. When wheat plants reached at stem elongation (Z31), early heading (Z51), early flowering (Z61), and grain filling (Z71) stages, GA3 solution (100 μM) was sprayed on the surfaces of leaves, culms and young panicles (Zadoks et al., 1974), and the control was applied with distilled water of the same volume. GA3 treatments in field were performed referred to Duan et al. 2020a.
Genomic sequence clone
Genomic DNA was extracted by CTAB method (Clark, 1997). The genomic sequence of Rht-B1b was cloned with primers in Supplementary Table 1, using PCR conditions described. PCR was performed in a Peltier Thermal cycler with a 20μl volume used ApexHF HS DNA Polymerase FL (Accurate Biology, China), and the PCR program was set according to the instructions (temperature was 60°C, extension time was 3min). PCR products were distinguished by 2% agarose gel electrophoresis.
The PCR target band was purified using Universal DNA Purification Kit (Tiangen, China). After adding ‘A’ base at the end, the recovered product was cloned into the T vector (TAKARA pMD18-T Vector Cloning Kit). The vector was transferred into E. coli DH5α competent cells by the 42°C heat shock method, and 950μl of SOC liquid medium was added to the 37°C shaking culture for 45 minutes. The product was plated in AMP-containing LB medium. After overnight incubation at 37°C, individual colonies were detected and sequenced. Primer synthesis and clone sequencing were performed at Tsingke Biology Technique (Beijing, China).
DNA extraction based on bulked segregation analysis
Yanzhan and Yanzhan mutation F2 population used to create tall and dwarf mixed pools. Genomic DNA was extracted from the second leaf below the spike of 20 tall and 20 dwarf plants at the filling stage. Equal amounts of tall plant genomic DNA were mixed into tall mixed pools, and equal amounts of dwarf plants genomic DNA were mixed into dwarf mixed pools. Combined with 660K chip (Affymetrix), SNP typing was detected (CapitalBio Technology, Beijing, China).
Plant height and internode length traits
Plant height and the internode length were investigated at the mature stage. Internode diameter and wall thickness were measured with Vernier scale during the grain filling stage (Du et al., 2018), with five replicates.
With reference to the method of Zhao et al. (2021), the internodes strength tester was used to measure the mechanical strength of the 4th and 5th basal internodes. The center of gravity height and fresh weight of the main stem were also measured. The formula for calculating the lodging resistance index is as follows:
LRI: Lodging Resistance Index; MS: Mechanical Strength (N); CGH: Center of Gravity Height (m); FW: Fresh Weight(g).
Yield traits
The standard and method for yield characters were referred to “Physiological Breeding II: A field Guide to Wheat Phenotyping” (Reynolds, 2012). At maturity, tillers plant-1, spike length, spikelet number spike-1, grains number spike-1, 1000-kernel weight, biomass plant-1, yield plant-1 and harvest index were measured using a random sample of 20 individual plants.
Grain traits
After threshing and drying, grain length, width and diameter were measured using the SC-G automatic seed analyzer (Wanshen, China). Grain quality traits, including water content, crude protein, wet gluten, water absorption, starch, stable time and formation time were measured using a Diode Array 7250 near-infrared reflectance (NIR) spectrometer (Perten Instrument AB, Sweden).
Data analysis
The formula for the difference between dwarf and tall plants as follow:
Data was analyzed using analysis of variance (ANOVA) for a t-test of the difference between the dwarf and tall lines using IBM SPSS Statistics 23.0. * means significant at p< 0.05. ** means significant at p< 0.01, none means not significant at p > 0.05. Oligo 7 was used for primer design. Origin 2018 and Microsoft Office Excel 2016 were used to complete the figures and tables.
Results
Discovery of a novel dwarfing site
Exogenous GA3 treatment of Yanzhan and Yanzhan mutation
Compared to Yanzhan, the height of the mutant was reduced by 22.5 cm (29.8%). At the jointing stage, Yanzhan and Yanzhan mutants were treated with exogenous GA3. The results showed that exogenous GA3 had no significant effect on plant height, length, diameter and wall thickness of the fourth and fifth stems of Yanzhan and Yanzhan mutants (Table 1). This indicated that the type of Yanzhan mutation was gibberellin insensitive and the mutation was not responsive to exogenous gibberellin.
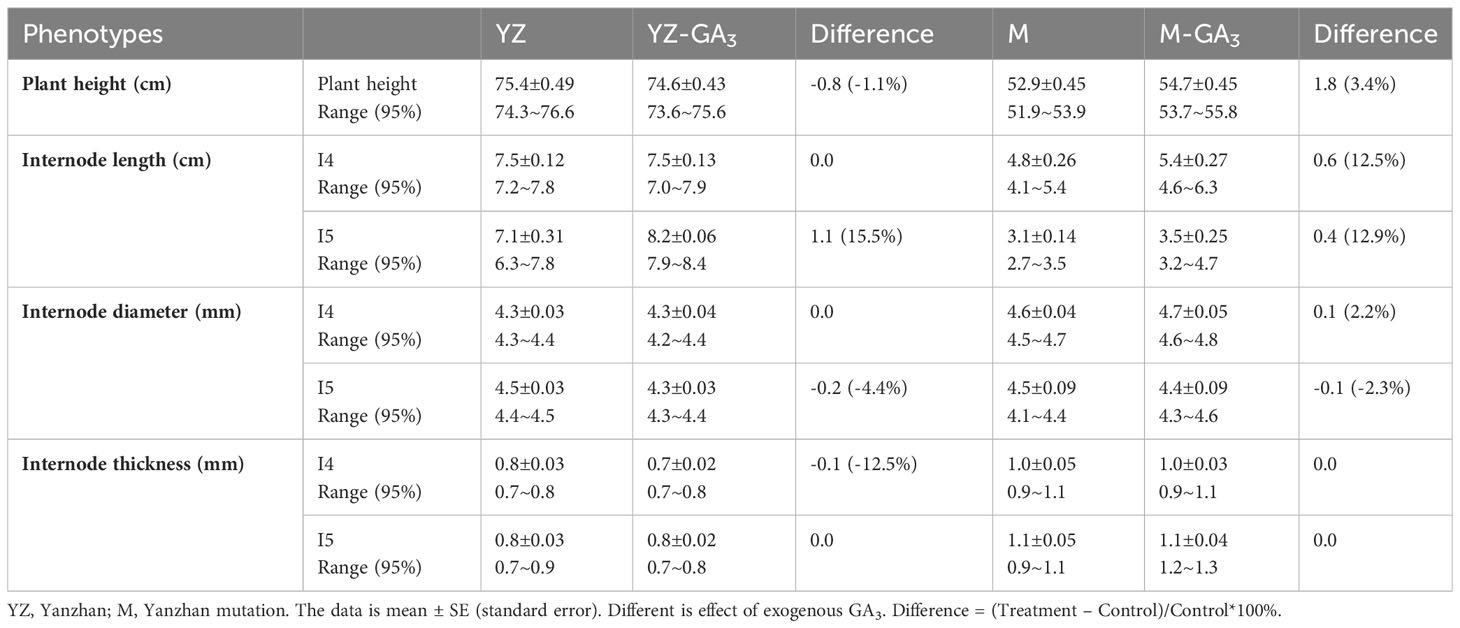
Table 1 Effect of exogenous GA3 treatment on plant height and internodes traits of YZ and M.
BSA (bulked segregation analysis) of F2 population
The homozygous differential SNPs of dwarf and tall bulks were analyzed using BSA and 660K SNP array. The results showed that 92 high quality SNPs were detected, of which 36 (39.1%) were found on chromosome 4B, and the rest were distributed on other chromosomes. And the differential SNPs on chromosome 4B were mainly enriched on the short arm, of which 28 (77.8%) were within 30-33 Mb (Figure 1A). This indicated that the mutation causing dwarf plant height was likely to be located in this interval.
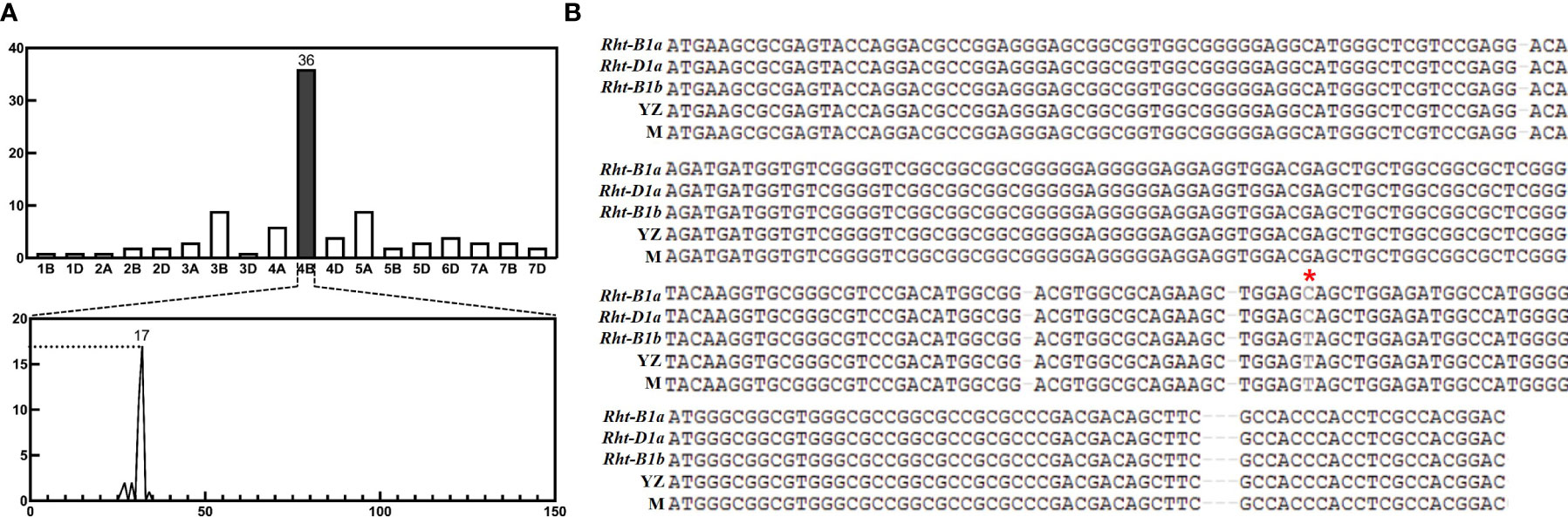
Figure 1 Bulked segregation analysis (BSA) results and sequencing of Rht-yz. (A) Results of BSA analysis in F2 population. The 4B chromosome has 36 SNPs, which concentrates on 30-33MB. (B) Results of Rht-B1b sequence. The * pointss the mutant site of Rht-B1b, which had no different between Yanzhan and mutation lines. YZ, Yanzhan; M, Yanzhan mutation.
Genomic sequence clone of Rht-B1b
The reported dwarfing gene Rht-B1b was found within this interval by searching for high confidence genes in 4BS: 30-33 MB according to IWGSC RefSeq v1.1, in combination with gene annotation (Table 2). In order to determine the genomic sequence of Rht-B1b, primers were designed (Supplementary Table 1). The coding region sequence was cloned using PCR technology and monoclonal sequencing. The sequence alignment revealed that the mutation site of Rht-B1b was the same in Yanzhan and Yanzhan mutation, this indicated that the mechanism of plant height reduction of this dwarfing site may be different from that of Rht-B1b (C-T mutation) (Figure 1B).
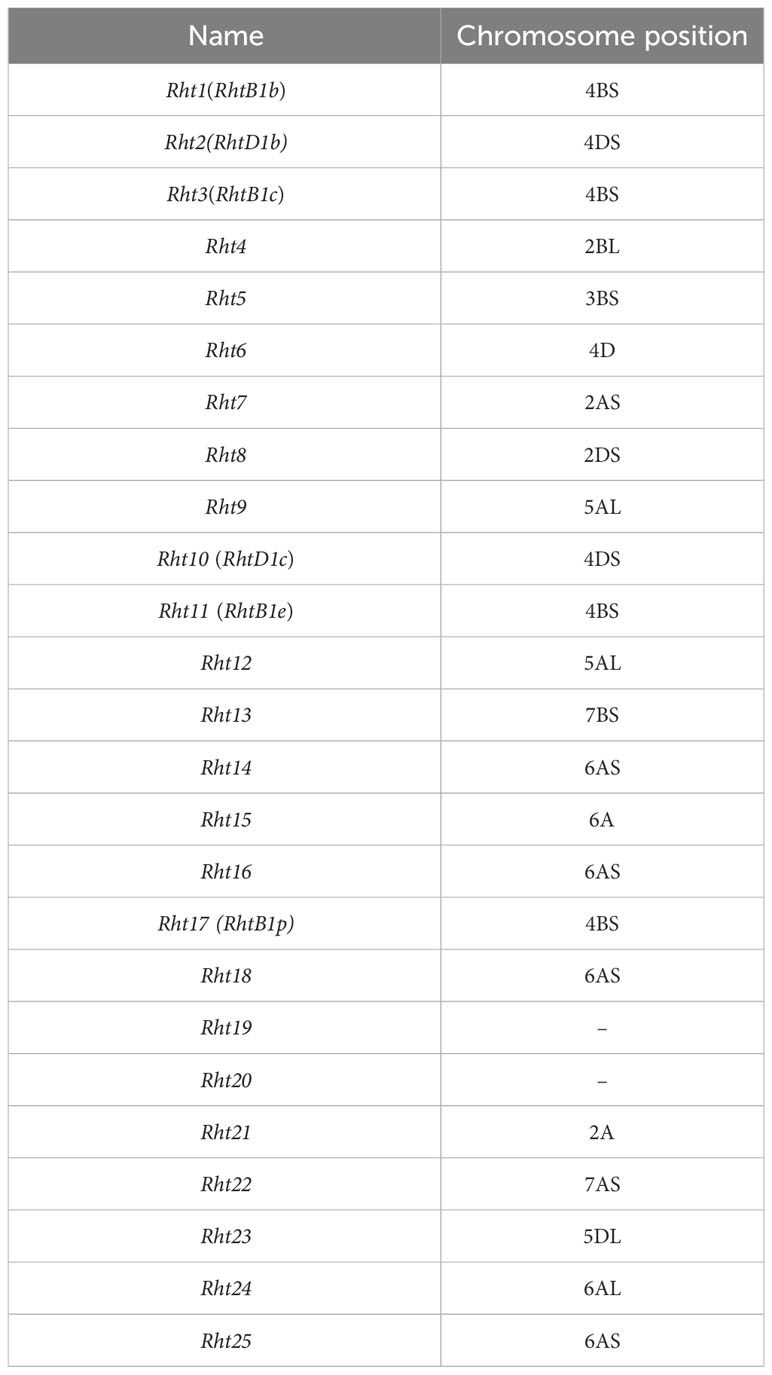
Table 2 Names of dwarf genes and positional information of chromosomes.
Effect of Rht-yz on plant height and internodes traits
Rht-yz significantly reduced plant height. The height of Yanzhan was approximately 75.4 cm, and the height of the Yanzhan mutant carrying Rht-yz was reduced by 22.5 cm (29.8%). In addition, Rht-yz reduced plant height by 36.4% and 33.4% in the F2:3 and F3:4 populations, respectively (Figure 2). Rht-yz also had significant effects on internode length, diameter and thickness in wheat. Both Yanzhan and Yanzhan mutations had 5 internodes, and the Yanzhan mutations had significantly shorter internode lengths compared with Yanzhan (Figures 3A, B), in which the peduncle was significantly shortened by 8.0 cm (31.3%), I2, I3, I4 and I5 were significantly shortened by 3.9 cm (25.8%), 4.5 cm (38.8%), 2.7 cm (36.0%) and 4.0 cm (56.3%), respectively. In addition, compared with Yanzhan the diameters of I2, I3, I4, and I5 with Yanzhan mutations were significantly increased by 0.4 mm (9.8%), 0.4 mm (9.1%), 0.3 mm (7.0%), and 0.2 mm (4.4%), respectively, the thickness of I3, I4 and I5 was significantly increased by 0.2 mm (33.3%), 0.2 mm (25.0%) and 0.3 mm (37.5%) respectively (Figure 3C and Table 3).
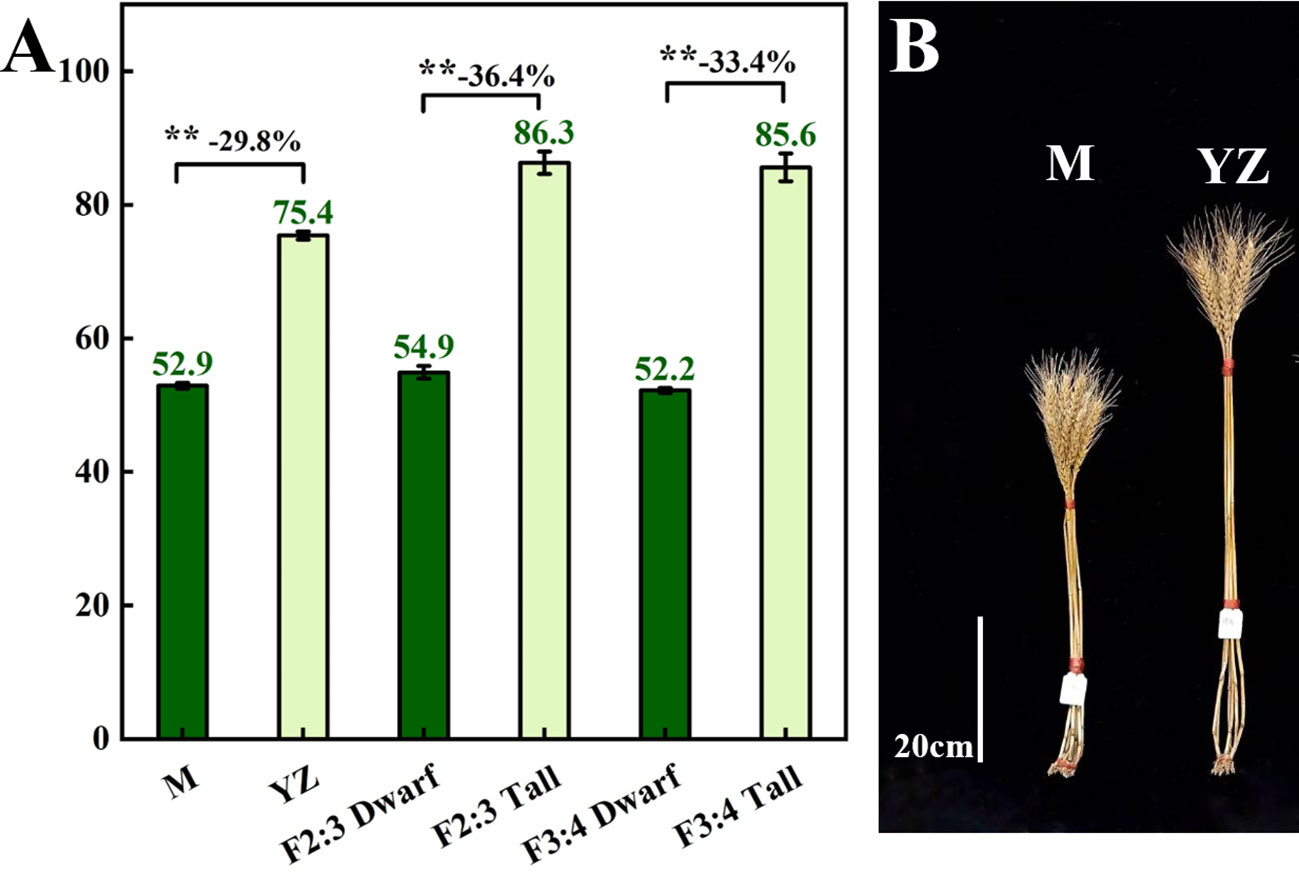
Figure 2 Plant height of Yanzhan mutation population. (A) Difference of plant height between dwarf and tall lines. (B) The mature plant height of Yanzhan and Yanzhan mutation. ** significant at p<0.01.
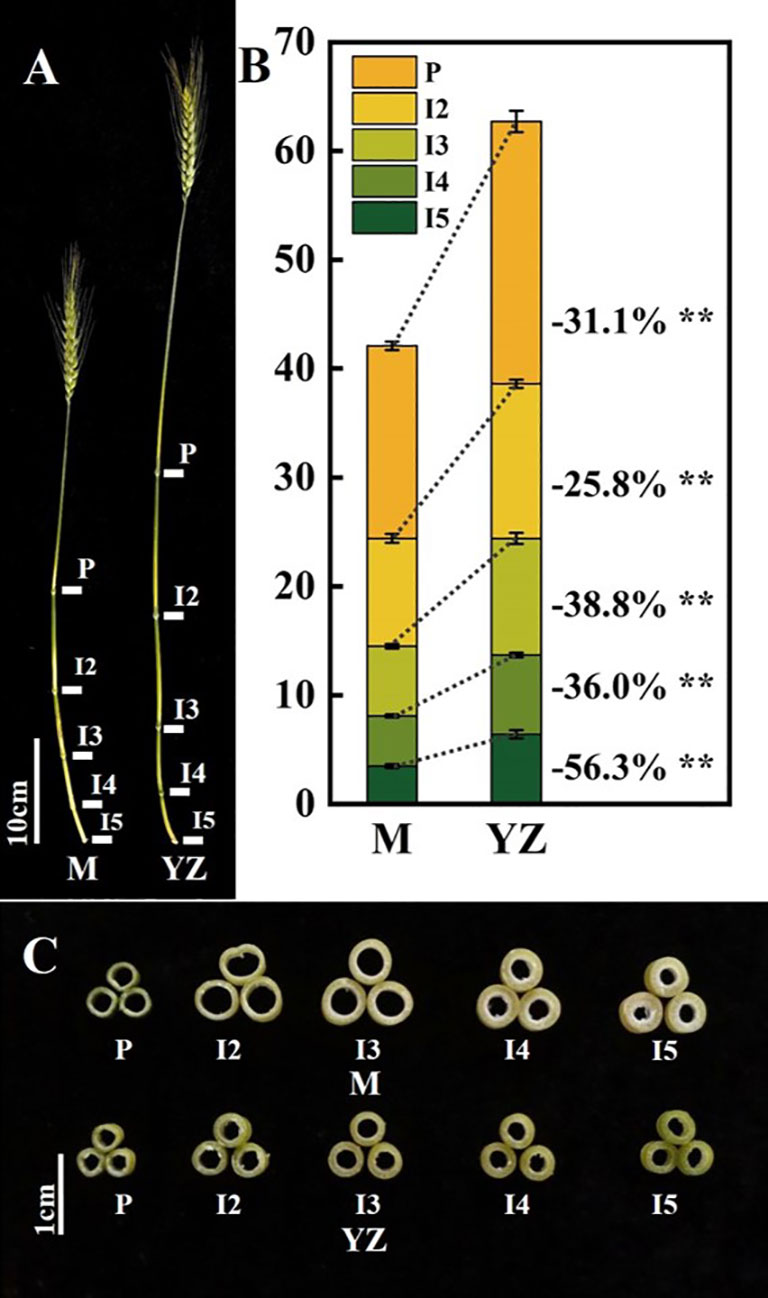
Figure 3 Internodes traits of Yanzhan and Yanzhan mutation at the filling stage. (A) The main stem. (B) Difference of internode length. (C) Cross section of the middle of each internode. ** significant at p<0.01. P, peduncle. I2, I3, I4 and I5, the second internode under spike, the third internode under spike, the fourth internode under spike, the fifth internode under spike.
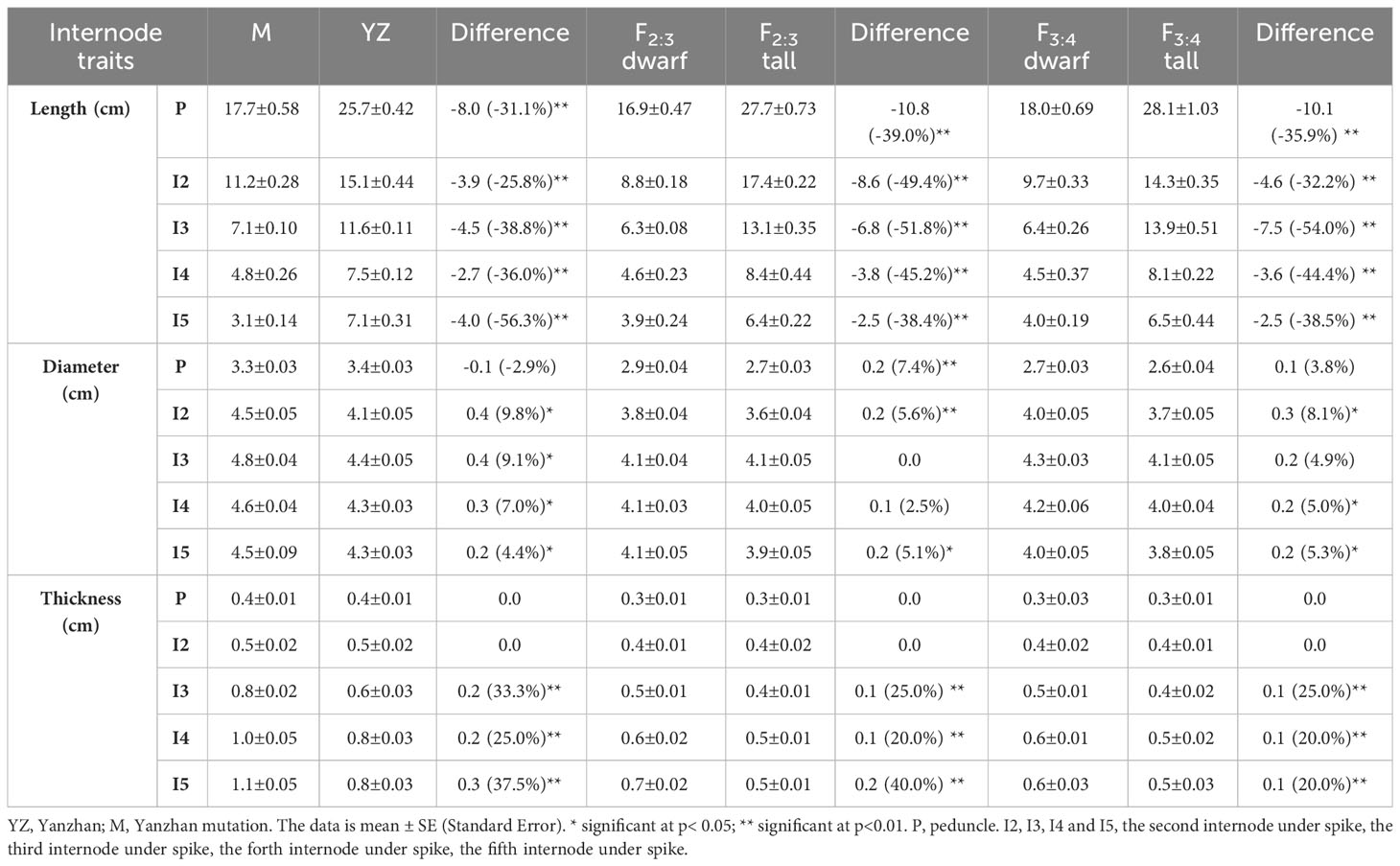
Table 3 Effect of Rht-yz on internode traits.
Similar results were observed in the F2:3 and F3:4 populations, where Rht-yz significantly reduced plant height and internode length. Rht-yz also significantly increased the diameter of I2 and I5 and the wall thickness of I3, I4 and I5 (Table 3).
Effect of Rht-yz on lodging resistant traits
This study focused on the lodging resistance index of the basal stem (I4 and I5), which are most relevant for lodging resistance traits (Figure 4). The results showed that Rht-yz significantly increased the mechanical strength of Yanzhan mutants I4 and I5 by 7.4% and 15.5% respectively. Meanwhile, Rht-yz significantly increased the lodging resistance index of I4 (72.7%) and I5 (90.6%) (Table 4).
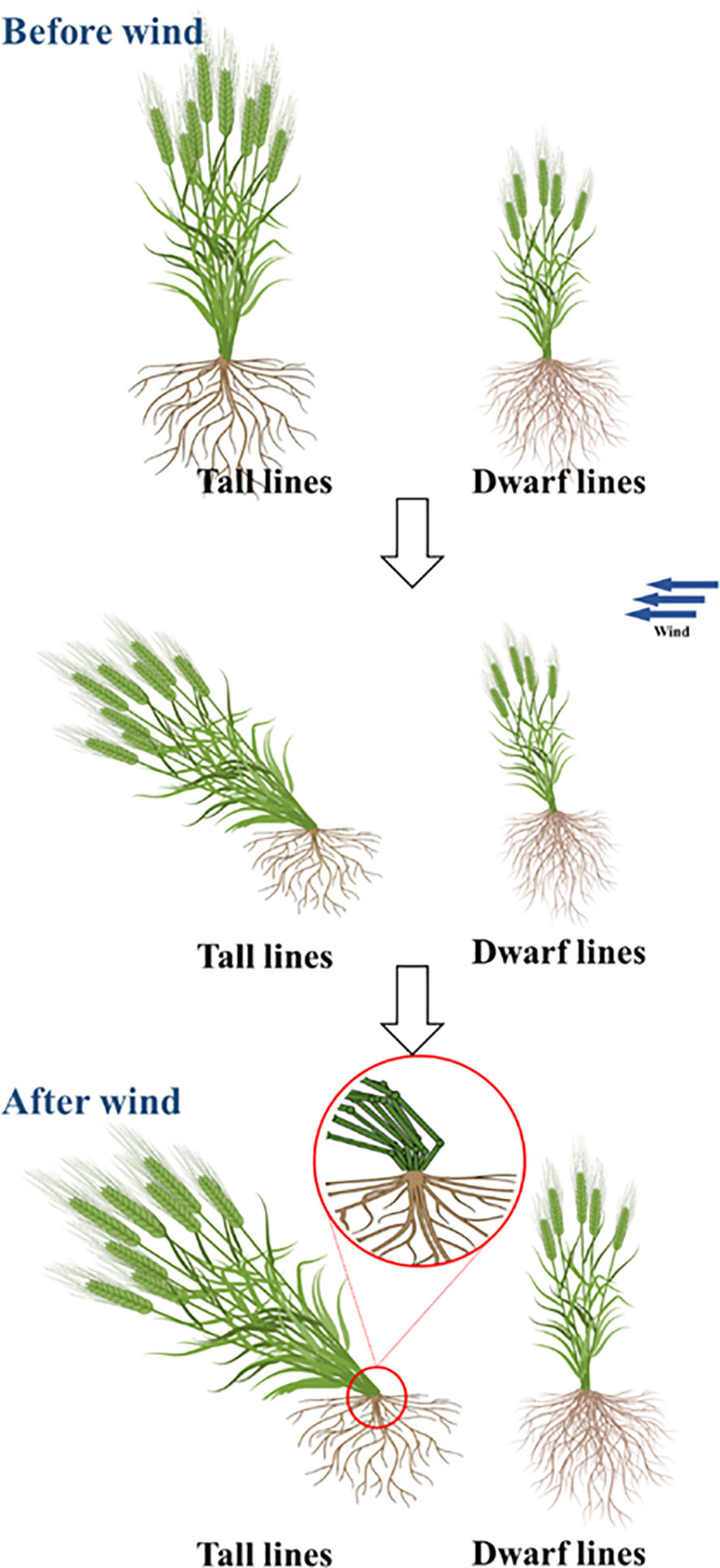
Figure 4 Diagram of lodging resistance of dwarf mutant.
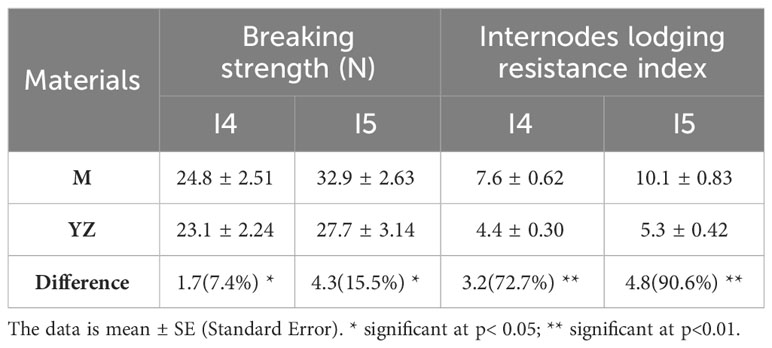
Table 4 Effect of Rht-yz on internodes lodging resistance index of basal internodes.
Effect of Rht-yz on yield and yield component
In the parental materials, F2:3 and F3:4 populations, Rht-yz significantly reduced the tillers plant-1, biomass plant-1 and yield plant-1. In addition, 1000-kernel weight and harvest index of the Yanzhan mutant and Rht-yz dwarf lines were also significantly reduced in the F2:3 population. For spike traits, Rht-yz increased spike length, spikelet number spike-1 and grain number spike-1, but the difference was not significant (Table 5).

Table 5 Effect of Rht-yz on yield traits.
Effect of Rht-yz on grain size and quality
Grain length, width and diameter of the Yanzhan mutation were not significantly different from those of Yanzhan, but Rht-yz reduced grain width by 6.5% and also had a negative effect on grain plumpness (Figure 5).
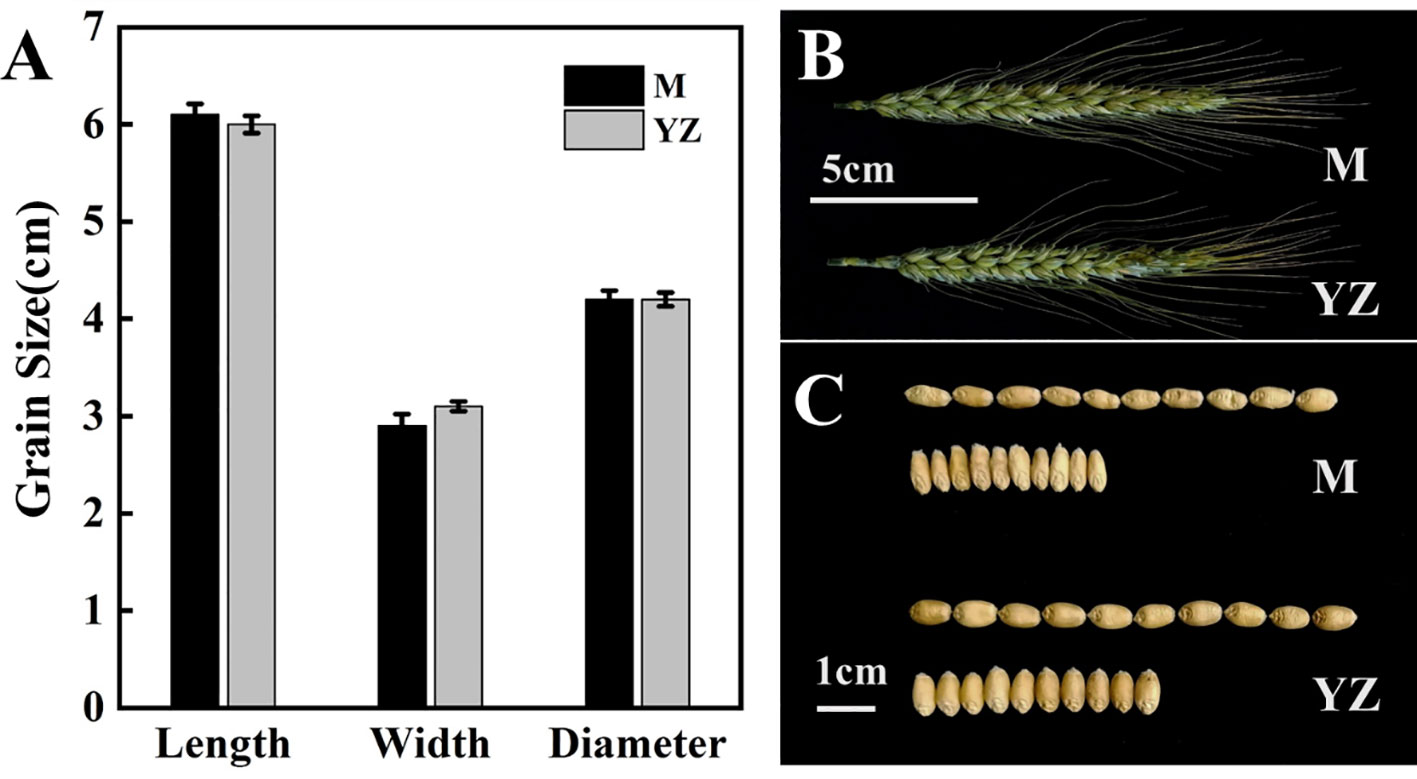
Figure 5 Grain traits of Yanzhan and Yanzhan mutation. (A) Difference of grain length, width and diameter. (B) Spike at the filling stage. (C) Grain photograph.
Rht-yz significantly increased grain crude protein (12.8%), wet gluten (10.0%), starch (6.9%), dough stability time (70.9%) and dough developing time (7.4%), but had no significant effect on water content and water absorption (Table 6).
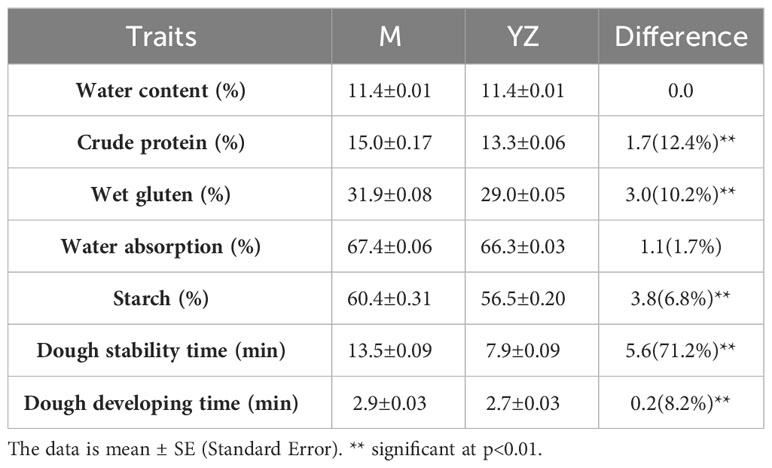
Table 6 Effect of Rht-yz on grain quality.
Discussion
The dwarfing gene solved the lodging problem in wheat breeding and greatly increased wheat yield per unit area (Flintham et al., 2000; Hedden, 2003). However, the negative effects of many dwarfing genes on yield and other traits in wheat have led to difficulties in their application. In this study, a new dwarf wheat mutant was identified and found to carry a new dwarf gene, provisionally named Rht-yz. To clarify the genetic effects of this gene, the plant height and internode length, lodging resistance, yield and grain traits were investigated from Yanzhan, Yanzhan mutations, F2:3 and F3:4 populations.
Discovery of a novel dwarfing site
Rht-B1a is wild-type and encodes a normal DELLA protein. The Rht-B1b and Rht-D1b have single base mutations in the coding region of the DELLA protein (Rht-B1b has a C-T mutation and Rht-D1b has a G-T mutation in the coding region), which ultimately leads to inactivity of the DELLA protein and reduced plant height (Peng et al., 1999; Pearce et al., 2011). The Rht-B1c transcript has a 90bp insertion, while Rht-B1d and Rht-B1e are prematurely terminated by single base (AT) mutations (Pearce et al., 2011), while Rht-B1k has displacement mutations in the VHIID domain to produce abnormal DELLA proteins. In this study, it was found that the number of differential SNPs enriched in the chromosome 4B was higher (36) and that there were 28 differential SNPs in the 30-33 MB interval, forming a differential SNPs enrichment peak. However, the number of differential SNPs was less and the distribution was scattered on other chromosomes, which did not form an obvious differential SNPs enrichment peak. Therefore, we can only obtain one interval (4B: 30-33MB) that regulates plant height, which contains a dwarf gene, Rht-B1b. The mutation site of the Rht-B1b did not differ between the Yanzhan and Yanzhan mutations. Therefore, Rht-yz is a new dwarfing site that regulates plant height and its mechanism of action is different from that of Rht-B1b (C-T mutation), but it is not clear whether it affects the Rht-B1a (TraesCS4B02G043100) or other genes.
Rht-yz improved the lodging resistance in wheat
Improving lodging resistance in wheat is one of the most important factors in increasing wheat yields. Many pre-Green Revolution wheat varieties were replaced due to lodging resistance defects. The wheat dwarf genes Rht-B1b and Rht-D1b reduced plant height by 23%, Rht-B1b+Rht-D1b reduced plant height by 50% (Butler et al., 2005). The dwarfing genes Rht8 and Rht12 reduced plant height by 8.0% and 40%, respectively (Tang et al., 2009; Chen et al., 2013). In this study, Rht-yz reduced plant height by approximately 29.8%, which was a moderate strength effect.
Studies have shown that the length of I4 and I5 is significantly correlated with lodging resistance in wheat (Kashiwagi et al., 2005), and lodging resistance is positively correlated with internode diameter, thickness and fullness (Zhao et al., 2021). Studies have shown that Rht18 increased basal internode strength (Yang et al., 2015), while Rht13 decreased basal internode diameter (Wang et al., 2014). In addition, the height of center of gravity is also an important factor in the lodging resistance index. In this study, Rht-yz mainly improved the lodging resistance by reducing the plant height and increasing the diameter, wall thickness and mechanical strength of the basal internode, which has important application value for dwarf breeding in wheat.
Rht-yz had negative effect on part of the yield components
The study reported that Rht3 increased tiller number (Zhang et al., 1995), Rht5 reduced spikelet number and grain number spike-1 in two different genetic backgrounds (Daoura et al., 2014), and Rht8 had little effect on yield traits (Rebetzke et al., 2012). In this study, Rht-yz had a significant negative effect on tiller number plant-1, biomass plant-1 and yield plant-1, but no significant effect on harvest index.
An unexpected disease (wheat stripe rust) during the 2019-2020 growing season led to a yield reduction in the F2:3 population. During the 2020-2021 growing season, no signs of disease were observed. Therefore, the effect of F3:4 on yield traits is closer to the actual effect of Rht-yz. The growth period of the Yanzhan mutation was 5-7 days later than that of Yanzhan, suggesting that Rht-yz may be involved in regulating the growth and development of wheat. Studies have also shown that Rht4+Rht-B1b has higher yield and harvest index under different moisture conditions (Liu et al., 2017). Rht5 reduced wheat yield, but Ppd-D1 effectively eliminated the negative effects of Rht5 on yield and growth by advancing the growth period (Chen et al., 2018). These studies remind us that the negative effects of Rht-yz on yield can also be eliminated by introducing other new genes.
Rht-yz improved wheat grain quality
Protein content, starch content and gluten content in wheat have an important impact on the promotion and application of wheat varieties. The quality of flour is mainly determined by protein and starch content (Ahmed, 2015). The dwarf gene Rht-B1c significantly increased the protein content, Rht14 had a negative effect on quality traits (Duan et al., 2020b), and Rht15 increased the protein and wet gluten content (Zhao et al., 2021). In this study, Rht-yz significantly increased crude protein, wet gluten and starch content. Therefore, Rht-yz is of great significance for improving the grain quality in wheat.
Conclusions
In this study, a new dwarfing site, Rht-yz, was identified in wild mutants that could improve lodging resistance by reducing plant height, increasing basal internode diameter, wall thickness and mechanical strength, and had a positive effect on grain quality traits. In addition, Rht-yz had a negative effect on some yield traits. Therefore, the rational use of the dwarf gene Rht-yz has some utility for lodging resistance breeding in wheat, but attention should be paid to improving its negative effects.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Author contributions
JH: Investigation, Writing – original draft, Writing – review & editing. ZZ: Investigation, Writing – review & editing. XF: Investigation, Writing – review & editing. YZ: Investigation, Writing – review & editing. MA: Writing – review & editing. LM: Data curation, Writing – review & editing. YH: Data curation, Writing – review & editing. YL: Data curation, Writing – review & editing. YY: Data curation, Writing – review & editing. LZ: Data curation, Writing – review & editing. DS: Data curation, Writing – review & editing. CF: Writing – review & editing, Data curation. Y-GH: Funding acquisition, Supervision, Writing – review & editing. LC: Funding acquisition, Supervision, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was financially supported by the Basic Research Program of natural sciences of Shaanxi province (2023-JC-ZD-08), ANSO Joint Research and cooperation project (ANSO-CR-KP-2022-05), the National Natural Science Foundation of China (32171991), Key Research and Development Program of Shaanxi (2022NY-189).
Conflict of interest
Author YH was employed by the company Jiushenghe Seed Industry Co., Ltd. Author YL was employed by the company Yangling Digital Agricultural Technology Co., Ltd. Author YY was employed by the company Shaanxi Grain and Agriculture Group Co., Ltd.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2024.1338425/full#supplementary-material
References
Ahmed, M. (2015). Response of spring wheat (Triticum aestivum L.) quality traits and yield to sowing date. PloS One 10 (4), e0126097. doi: 10.1371/journal.pone.0126097
Butler, J. D., Byrne, P. F., Mohammadi, V., Chapman, P. L., Haley, S. D. (2005). Agronomic performance of alleles in a spring wheat population across a range of moisture levels. Crop Sci. 45, 939. doi: 10.2135/cropsci2004.0323
Casebow, R., Hadley, C., Uppal, R., Addisu, M., Gooding, M. (2016). Reduced height (Rht) alleles affect wheat grain quality. PloS One 11, e0156056. doi: 10.1371/journal.pone.0156056
Chen, L., Phillips, A. L., Condon, A. G., Parry, M. A. J., Hu, Y.-G. (2013). GA-responsive dwarfing gene Rht12 affects the developmental and agronomic traits in common bread wheat. PloS One 8, e62285. doi: 10.1371/journal.pone.0062285
Chen, L., Yang, Y., Cui, C., Lu, Q., Du, Y., Su, R., et al. (2018). Effects of Vrn-B1 and Ppd-D1 on developmental and agronomic traits in Rht5 dwarf plants of bread wheat. Field Crop Res. 219, 24–32. doi: 10.1016/j.fcr.2018.01.022
Clark, M. S. (1997). Plant Molecular Biology — A Laboratory Manual || Genomic DNA Isolation, Southern Blotting and Hybridization. Springer, Berlin, Heidelberg: Springer, 3–53. doi: 10.1007/978-3-642-87873-2
Daoura, B. G., Chen, L., Du, Y., Hu, Y. G. (2014). Genetic effects of dwarfing gene Rht5 on agronomic traits in common wheat ( Triticum aestivum L.) and QTL analysis on its linked traits. Field Crops Res. 156, 22–29. doi: 10.1016/j.fcr.2013.10.007
Derkx, A. P., Harding, C., Miraghazadeh, A., Chandler, P. M. (2017). Overgrowth (Della) mutants of wheat: development, growth and yield of intragenic suppressors of the Rht-B1c dwarfing gene. Funct. Plant Biol. 44, 525. doi: 10.1071/FP16262
Du, Y., Liang, C., Wang, Y., Yang, Z., Saeed, I., Daoura, B. G., et al. (2018). The combination of dwarfing genes Rht4 and Rht8 reduced plant height, improved yield traits of rainfed bread wheat ( Triticum aestivum L.). Field Crops Res. 215, 149–155. doi: 10.1016/j.fcr.2017.10.015
Duan, S., Zhao, Z., Qiao, Y., Cue, C., Condon, A. G., Chen, L., et al. (2020a). Vigorous responsiveness of dwarf gene Rht14 to exogenous GA3 evaluated on important morphological and agronomic traits in durum wheat. Agron. J. 112 (6), 5033–5044. doi: 10.1002/agj2.20409
Duan, S., Zhao, Z., Qiao, Y., Cui, C., Morgunov, A., Condon, A. G., et al. (2020b). GAR dwarf gene Rht14 reduced plant height and affected agronomic traits in durum wheat (Triticum durum) - ScienceDirect. Field Crops. Res. 248. doi: 10.1016/j.fcr.2020.107721
Flintham, J. E., Angus, W. J., Gale, M. (2000). Heterosis, overdominance for grain yield, and alpha-amylase activity in F1 hybrids between near-isogenic Rht dwarf and tall wheats. J. Agric. Sci. 129, 371–378. doi: 10.1017/S0021859697004899
Gale, M. D., Youssefian, S., Russell, G. E. (1985). “Dwarfing genes in wheat.” In Progress in Plant Breeding–1 (Elsevier), pp. 1–35. doi: 10.1016/b978-0-407-00780-2.50005-9
Guedira, M., Brown-Guedira, G., Sanford, D. V., Sneller, C., Souza, E., Marshall, D. (2010). Distribution of rht genes in modern and historic winter wheat cultivars from the eastern and central USA. Crop Sci. 50, 1811–1822. doi: 10.2135/cropsci2009.10.0626
Hayat, H., Mason, R. E., Lozada, D. N., Acuna, A., Miller, R. G. (2019). Effects of allelic variation at Rht-B1 and Rht-D1 on grain yield and agronomic traits of southern US soft red winter wheat. Euphytica 215, 172. doi: 10.1007/s10681-019-2478-2
Hedden, P. (2003). The genes of the green revolution. Trends Genet. 19, 5–9. doi: 10.1016/s0168-9525(02)00009-4
Kashiwagi, T., Sasaki, H., Ishimaru, K. (2005). Factors responsible for decreasing sturdiness of the lower part in lodging of rice (Oryza sativa L.). Plant Product. Sci. 8, 166–172. doi: 10.1626/pps.8.166
Liu, Y., Zhang, J., Hu, Y. G., Chen, J. (2017). Dwarfing genes Rht4 and Rht-B1b affect plant height and key agronomic traits in common wheat under two water regimes. Field Crops Res. 204, 242–248. doi: 10.1016/j.fcr.2017.01.020
Pearce, S., Saville, R., Vaughan, S. P., Chandler, P. M., Wilhelm, E. P., Korolev, A., et al. (2011). Molecular characterisation of Rht1 dwarfing genes in hexaploid wheat (Triticum aestivum). Plant Physiol. 157 (4), 1820–31. doi: 10.1104/pp.111.183657
Peng, J., Richards, D. E., Hartley, N. M., Murphy, G. P., Devos, K. M., Flintham, J. E., et al. (1999). "Green revolution" genes encode mutant gibberellin response modulators. Nature 400, 256. doi: 10.1038/22307
Rebetzke, G. J., Ellis, M. H., Bonnett, D. G., Mickelson, B., Condon, A. G., Richards, R. A. (2012). Height reduction and agronomic performance for selected gibberellin-responsive dwarfing genes in bread wheat (Triticum aestivum L.). Field Crops Res. 126, 87–96. doi: 10.1016/j.fcr.2011.09.022
Reynolds, M. (2012). Physiological Breeding II: A field Guide to Wheat Phenotyping, Physiological Breeding II: A field Guide to Wheat Phenotyping. Mexico: CIMMYT.
Richards, R. A. (1992). The effect of dwarfing genes in spring wheat in dry environments. I. Agronomic characteristics. Aust. J. Agric. Res. 43, 517–527. doi: 10.1071/AR9920517
Tang, N., Jiang, Y., He, B.-R., Hu, Y.-G. (2009). The effects of dwarfing genes (Rht-B1b, rht-D1b, and rht8) with different sensitivity to GA_3 on the coleoptile length and plant height of wheat. Agric. Sci. China. 9, 11. doi: 10.1016/S1671-2927(08)60310-7
Wang, Y., Liang, C., Du, Y., Yang, Z., Condon, A. G., Hu, Y. G. (2014). Genetic effect of dwarfing gene Rht13 compared with Rht-D1b on plant height and some agronomic traits in common wheat (Triticum aestivum L.). Field Crops Res. 162, 39–47. doi: 10.1016/j.fcr.2014.03.014
Yang, Z.Y., Zheng, J.C., Liu, C.Y., Wang, Y.S., Condon, A.G., Chen, Y.F., et al. (2015). Effects of the GA-responsive dwarfing gene Rht18 from tetraploid wheat on agronomic traits of common wheat. Field Crops Res. 183, 92–101. doi: 10.1016/j.fcr.2015.07.028
Zadoks, J. C., Chang, T. T., Konzak, F. C. (1974). A decimal code for the growth stages of cereals. Weed Res. 14, 415–421. doi: 10.1111/j.1365-3180.1974.tb01084.x
Zhang, R., Lu, J., Ma, G., Wu, Y., Min, F., Zhao, Y. (1995). Effects of rht3 gene and 4B-chromosome with rht3 on photosynthetic carbon-assimilation of wheat. Acta GENE. SINICA. 22, 264–271.
Zhang, X., Yang, S., Zhou, Y., He, Z., Xia, X. (2006). Distribution of the Rht-B1b, Rht-D1b and Rht8 reduced height genes in autumn-sown Chinese wheats detected by molecular markers. Euphytica 152, 109–116. doi: 10.1007/s10681-006-9184-6
Keywords: dwarf gene, plant height, lodging resistance, wheat, agronomic traits
Citation: Hao J, Zhao Z, Fu X, Zhao Y, Ateeq M, Mou L, Han Y, Liu Y, Yin Y, Zotova L, Serikbay D, Fan C, Hu Y-G and Chen L (2024) Effect of a novel dwarfing mutant site on chromosome 4B on agronomic traits in common wheat. Front. Plant Sci. 15:1338425. doi: 10.3389/fpls.2024.1338425
Received: 14 November 2023; Accepted: 15 February 2024;
Published: 20 March 2024.
Edited by:
Thomas Miedaner, University of Hohenheim, GermanyReviewed by:
Vikas Gupta, Indian Institute of Wheat and Barley Research (ICAR), IndiaPaul Nicholson, John Innes Centre, United Kingdom
Copyright © 2024 Hao, Zhao, Fu, Zhao, Ateeq, Mou, Han, Liu, Yin, Zotova, Serikbay, Fan, Hu and Chen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yin-Gang Hu, aHV5aW5nYW5nQG53c3VhZi5lZHUuY24=; Liang Chen, Y2hlbmxpYW5nOTExN0Bud3N1YWYuZWR1LmNu
†These authors have contributed equally to this work