
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Physiol., 04 June 2024
Sec. Autonomic Neuroscience
Volume 15 - 2024 | https://doi.org/10.3389/fphys.2024.1422927
This article is part of the Research TopicCardio-Respiratory-Brain Integrative Physiology: Interactions, Mechanisms, and Methods for AssessmentView all 9 articles
 Pablo Alvarez-Araos1,2Sergio Jiménez2
Pablo Alvarez-Araos1,2Sergio Jiménez2 Camila Salazar-Ardiles1
Camila Salazar-Ardiles1 Cristian Núñez-Espinosa3,4
Cristian Núñez-Espinosa3,4 Valeria Paez1,5
Valeria Paez1,5 Maria Rodriguez-Fernandez5
Maria Rodriguez-Fernandez5 Antoine Raberin6
Antoine Raberin6 Gregoire P. Millet6
Gregoire P. Millet6 Rodrigo Iturriaga1,7
Rodrigo Iturriaga1,7 David C. Andrade1*
David C. Andrade1*The hypoxic chemoreflex and the arterial baroreflex are implicated in the ventilatory response to exercise. It is well known that long-term exercise training increases parasympathetic and decreases sympathetic tone, both processes influenced by the arterial baroreflex and hypoxic chemoreflex function. Hypobaric hypoxia (i.e., high altitude [HA]) markedly reduces exercise capacity associated with autonomic reflexes. Indeed, a reduced exercise capacity has been found, paralleled by a baroreflex-related parasympathetic withdrawal and a pronounced chemoreflex potentiation. Additionally, it is well known that the baroreflex and chemoreflex interact, and during activation by hypoxia, the chemoreflex is predominant over the baroreflex. Thus, the baroreflex function impairment may likely facilitate the exercise deterioration through the reduction of parasympathetic tone following acute HA exposure, secondary to the chemoreflex activation. Therefore, the main goal of this review is to describe the main physiological mechanisms controlling baro- and chemoreflex function and their role in exercise capacity during HA exposure.
High altitude (HA), characterized by reduced barometric and ambient O2 pressure, is challenging for human physiology (Huey, 2002; Windsor and Rodway, 2007; Mallet et al., 2021). Therefore, determining short- and long-term physiological adaptations to HA is fundamental. Chronic adaptations to HA involve maintaining oxygen (O2) supply to several tissues (Beall, 2007; Subudhi et al., 2014; Murray et al., 2018; Mallet et al., 2023). Altitude-born residents show physiological adaptations to cope with the hypoxia, such as a rise in minute ventilation at rest, high total lung capacity, lung diffusion, and better exercise performance than low-lander non-native residents (Greksa et al., 1988; Beall, 2007; Huerta-Sanchez et al., 2014; Simonson et al., 2015); however, not all high-altitude residents (i.e., Tibetans and Andeans) display the same adaptations (Beall, 2000; Beall, 2006; Bigham et al., 2013). Indeed, under similar hypobaric-hypoxic stress, Tibetans showed a one-half standard deviation higher resting ventilation and O2 saturation compared to Andeans; however, Andeans depict a higher hemoglobin concentration than Tibetans (Beall, 2006). Interestingly, Tibetans enhanced the hypoxic ventilatory response compared to the Andean population (Beall, 2000).
For non-residents, short-term acclimatization to HA includes an increment in arterial blood pressure, heart rate, metabolic demand, basal ventilation, autonomic alterations characterized by sympathoexcitation and parasympathetic withdrawal, and decreased exercise performance (Bartsch and Saltin, 2008; Naeije, 2010; Mallet et al., 2021). Recently, we found that the parasympathetic arm of the baroreflex (BR) pathway, measured through the phenylephrine-dependent increase of heart rate (HR), was reduced following acute exposure to HA (Beltran et al., 2020). In addition, a hypoxic-dependent potentiation of the chemoreflex was found, which modulates ventilation and autonomic functions in normoxia and during a hypoxic insult (Mahamed and Duffin, 2001). Therefore, it is likely that BR and chemoreflex pathways may interact at the central nervous system level (Somers et al., 1991; Katayama et al., 2019) during HA exposure. This idea is supported by the fact that peripheral carotid body (CB) denervation increases hypoxic-dependent BR range decrement in rats subjected to chronic intermittent hypoxia (Del Rio et al., 2015; Del Rio et al., 2016). In addition, (Kronsbein et al., 2020) also found that the buffering BR activity decreased during chemoreflex activation in normal human subjects (Kronsbein et al., 2020). Therefore, it is reasonable that the decreased BR-dependent parasympathetic activity found during acute exposure to HA (Beltran et al., 2020) results from a chemoreflex activation that may affect exercise performance during HA. (Machhada et al., 2017) found that optogenetic activation of the parasympathetic tone improved exercise capacity, producing a mimic of exercise training (Machhada et al., 2017). Additionally, they found that a decreased vagal control elicited using chemogenetics impairs exercise performance in rats (Machhada et al., 2017). It is also well-established that exposure to hypoxia alters sympathetic activation at rest (Perini et al., 1996; Sevre et al., 2001), which is related to decreased endurance performance (Schmitt et al., 2008). Hypoxia also alters the post-exercise parasympathetic reactivation, impairing recovery (Al Haddad et al., 2012; Fornasiero et al., 2018).
Exposure to HA negatively impacts alveolar pO2 and, consequently, arterial pO2, which has been linked to a marked decrease in exercise capacity (Naeije, 2010). The reduction in maximal O2 uptake (VO2max) is estimated at 6%–7% per 1,000 m increasing altitude (range 4.6%–7.5%) (Wehrlin and Hallen, 2006). However, an altered chemoreflex and BR function may contribute to decreased exercise performance, independent of environmental conditions (Pijacka et al., 2018; Eugenin et al., 2020). Therefore, considering the dependence of exercise performance on vagal activity (Machhada et al., 2017), HA-related chemoreflex activation (Iturriaga and Alcayaga, 2004), and decreased BR-dependent parasympathetic drive (Beltran et al., 2020), it is plausible to propose that the interaction between the chemoreflex and the BR may modulate the cardiorespiratory fitness through parasympathetic control, secondary to the decrease oxygen pressure due to HA. Thus, in the present review, we will analyze and discuss the role played by the chemoreflex and baroreflex and their effects on physical capacity during HA exposure.
The BR is a homeostatic mechanism that maintains the cardiac output at normal levels through negative feedback in the brainstem, modulating heart rate and arterial blood pressure by controlling sympathetic and parasympathetic activities (Thrasher, 2002). Thus, an increment in arterial blood pressure produces stimulation of the baroreceptors located in the aortic arch and carotid bifurcation, increasing parasympathetic cardiac response and attenuating sympathetic outflow to the heart and peripheral vessels, triggering a decrease in HR, cardiac contractility, and peripheral resistance. Conversely, decreased arterial blood pressure reduces the neural discharges of the baroreceptors, leading to increased sympathetic drive, vasoconstriction, and hypertension, producing a positive chronotropic response and increased cardiac contractility. Mechanistically, the BR is initiated in the baroreceptors, which are mechanically sensitive nerve endings and found in the aortic arch and the carotid bifurcation (Persson et al., 1988). The aortic baroreceptor afferent nerve fibers are conveyed in the aortic depressor nerve, with their soma located in the nodose ganglion (NG).
On the other hand, the carotid baroreceptors are found in the carotid sinus, with the soma lying in the petrosal ganglion (PG) (Fadel et al., 2003; Kougias et al., 2010; Lau et al., 2016). The arterial pressure stretches the carotid sinus of the aortic arch, inducing a rise of cytosolic Ca2+. Nevertheless, the cytosolic Ca2+ was higher in aortic baroreceptor neurons than in carotid baroreceptor neurons, suggesting aortic baroreceptors are more sensitive to arterial blood pressure changes than carotid baroreceptors (Lau et al., 2016). Indeed, it has been shown that, through baroreceptor nerve activity in vivo, aortic depressor nerve discharge was increased compared to the carotid sinus nerve activity (afferent activity), a similar change in blood pressure in rats (Lau et al., 2016).
Cardiac sympathetic mechanism. The baroreflex-dependent sympathetic activation (HR increment) is mediated by norepinephrine (NE) releases, which bind to the β-adrenergic receptors activating Na+ channels (Kaupp and Seifert, 2001; Lakatta and DiFrancesco, 2009). In addition, the β-adrenergic receptor controls intracellular Ca2+ control (Fadel et al., 2003) through cyclic adenosine 3′,5′-monophosphate (cAMP)/cAMP-dependent protein kinase (PKA) signaling (Gray et al., 1998). PKA phosphorylation is mediated by the A-kinase-anchoring protein (AKAP-15/18), which interacts with the intracellular domain of the channel and brings the PKA to its binding site (Sampson and Kass, 2010). A similar process occurs in the sarcoplasmic reticulum, where AKAP-6 interacts with the ryanodine channels and recruits the PKA site, increasing the release of intracellular [Ca2+], which contributes to increased HR and contractility rate (Lakatta and DiFrancesco, 2009). For instance, in the NE-mediated chronotropic response, there must be a fast removal of [Ca2+]i, which is performed by mitigating the inhibition of the Ca2+ATPasa pump by phosphorylation of the phospholamban protein (Marx et al., 2002).
Cardiac parasympathetic mechanism. The HR reduction is mediated by muscarinic receptors (M2), acetylcholine-dependent receptors (Kaupp and Seifert, 2001; Thrasher, 2002). Muscarinic receptors are expressed in the sinoatrial, atrioventricular, and cardiomyocyte T-tubules system (Kaupp and Seifert, 2001). The activation of the M2 receptor is mediated by a G protein-coupled receptor, which rectifies K+ conductance and decreases cAMP, reducing PKA activation (Swynghedauw, 1999). All these produce a longer duration of atrial action potentials and consequently decrease the HR and contractility rate of the heart (Olshansky et al., 2008). In addition, M2 receptors activate nitric oxide synthase (NOS) via guanylate cyclase, inhibiting L-type Ca2+ channels (Olshansky et al., 2008) and slowing the entry of Ca2+ into the intracellular medium, which contributes to decreasing the contractility rate of the heart (Swynghedauw, 1999; Olshansky et al., 2008).
Previously, we found a baroreflex-mediated parasympathetic withdrawal during HA exposure. Indeed, we observed a diminished bradycardic response to phenylephrine and a decreased power spectral density at a high-frequency component (parasympathetic drive) of the time-varying heart rate variability, evidencing a marked decrease in vagal outflow (Beltran et al., 2020). The autonomic control of physical performance is of such relevance that decreased parasympathetic drive triggers performance impairment, while increased parasympathetic control promotes a mimicry of exercise training, improving physical performance in rats (Machhada et al., 2017).
The carotid body (CB) is the main peripheral oxygen chemoreceptor, composed of chemoreceptors (glomus or type I cells) and sustentacular clusters type II cells (Iturriaga and Alcayaga, 2004; Prabhakar, 2006; Iturriaga et al., 2021b). The CB type I cells respond to a wide variety of stimuli, such as changes in arterial levels of pO2, carbon dioxide pressure (pCO2), pH, blood flow, glucose, temperature, osmolarity, and insulin; therefore, they are considered polymodal receptors (Gonzalez et al., 1994; Ding et al., 2011; Iturriaga et al., 2021b). In response to low pO2 and high pCO2-H+, type I cells are activated by inhibiting O2-sensitive K+ channels. Further, the hypoxia-dependent production of gasotransmitters (NO, CO, H2S) also regulates ion channel activity in the CB (Iturriaga et al., 2021a). The intracellular pathways related to the neurotransmitter release are AMP-activated protein kinases and PKC, as well as reactive oxygen species in the CB type I cells, promoting the release of neurotransmitters such as acetylcholine (Ach) and adenosine triphosphate (ATP) that interact with receptors in the nerve terminal of petrosal sensory neurons that project through the carotid sinus nerve to the nucleus tractus solitarii (NTS) (Iturriaga and Alcayaga, 2004). In addition, the type I cells also release several molecules, which serve as excitatory or inhibitory modulators of CB chemosensory transduction (i.e., NO, histamine, and Ang II) (Iturriaga and Alcayaga, 2004; Del Rio et al., 2008).
Briefly, a reduction in pO2 in the arterial blood is detected by primary O2 sensors, type I carotid body cells, which rapidly communicate with potassium (K+) channels, leading to the closure of these channels. In turn, via membrane depolarization and increases in intracellular [Ca2+] concentration, the release of neurotransmitters (i.e., Ach and ATP) leads to excitation of the afferent nerve that runs in the carotid nerve sinus up to the respiratory centers in the brain stem (Teppema and Dahan, 2010).
Additionally, it has been evidenced that the CBs are related to exercise capacity in physiological and pathophysiological conditions (Honda et al., 1979; Honda, 1985; Andrade et al., 2021a). CB denervation or resection and exposure to 100% O2 (resulting in decreased CB activity) reduce ventilatory responses in exercise and markedly diminish exercise capacity in humans and animals. Indeed, hyperoxic gas applied during ventilatory threshold 2 (VT2) decreases pulmonary ventilation in humans, suggesting that the CB contributes at least in part to increasing ventilation at VT2 during incremental exercise (Masuda et al., 1988). Further, Honda et al. (1979) showed that CB resection in asthma patients decreases the respiratory response to exercise compared to patients with intact CBs (Honda et al., 1979). Along with this, we showed that CB resection promotes a phenotype shift from heart failure tolerant to physical exercise animals to intolerants (Andrade et al., 2021b). All this evidence strongly suggests the pivotal role of CB peripheral chemoreceptors in ventilatory response to physical exercise as well as training-dependent adaptations.
Figure 1 depicts the neural control of chemoreflex function. During hypobaric hypoxic environments, chemoreceptor activation promotes ventilatory acclimatization and sympathoexcitation (Vizek et al., 1987; Schultz and Sun, 2000). The first central integration of sensory information from peripheral chemoreceptor and baroreceptor inputs occurs in the commissural and medial divisions of the nucleus of the solitary tract (cNTS and mNTS, respectively) (Claps and Torrealba, 1988; Finley and Katz, 1992). The cNTS and mNTS neurons integrate and project to other autonomic and respiratory regions (i.e., rostral ventrolateral medulla [RVLM], caudal ventrolateral medulla [CVLM], and the central pattern generator [CPG]) (Ponikowski et al., 2001; Rosin et al., 2006; Smith et al., 2010; Diaz et al., 2020). King et al. (2012) showed that acute-hypoxic stimulus increments the activation of cNTS catecholaminergic neurons (King et al., 2012). In addition, it has been found that sustained hypoxia, similar to HA exposure, enhances NTS glutamatergic synaptic transmission after 1 day and augments glutamate (Glu) receptor expression after 7 days (Zhang et al., 2009; Pamenter et al., 2014; Accorsi-Mendonça et al., 2015; Accorsi-Mendonça et al., 2019). Other regions sensitive to hypoxia are RVLM and CVLM (King et al., 2013; Boychuk et al., 2012; D'Agostino et al., 2001); nevertheless, it has been demonstrated that the activation of RVLM is CB-dependent and not a direct hypoxic effect (Del Rio et al., 2013). Interestingly, we found BR-dependent autonomic control impairment during HA exposure (3,290 m) (Beltran et al., 2020); however, whether it depends on the chemoreflex activation with their respective neural autonomic nuclei activation has not been demonstrated. Additionally, although considering that autonomic control and chemoreceptors are critically essential to maintaining cardiorespiratory fitness during exercise, there is no comprehensive evidence depicting whether, during HA or normobaric hypoxia, the exercise capacity impairment is related to BR-dependent parasympathetic withdrawal, secondary to a chemoreflex enhancement.
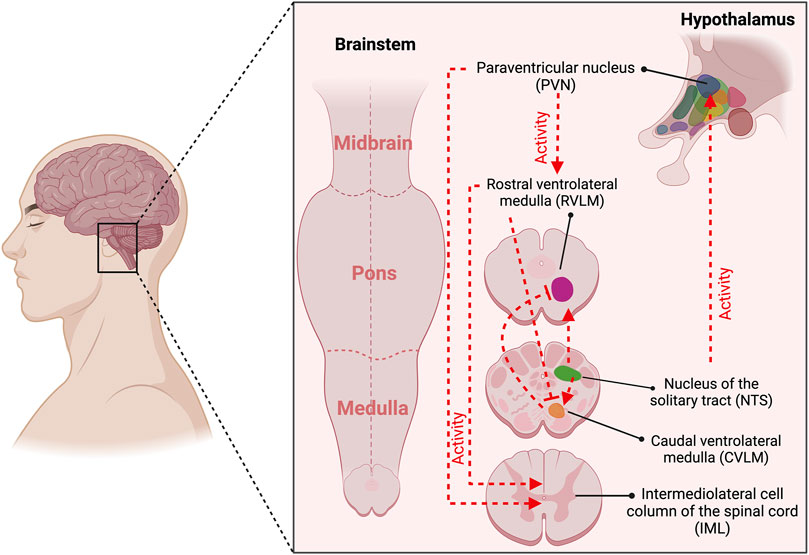
Figure 1. Central command is associated with the activation of baroreceptors and chemoreceptors. The first central integration of sensory information from peripheral chemoreceptor and baroreceptor inputs occurs in the commissural and medial divisions of the nucleus of the solitary tract (NTS). The NTS neurons integrate and project to other cardiac autonomic and respiratory regions (i.e., rostral ventrolateral medulla [RVLM], caudal ventrolateral medulla [CVLM], and the paraventricular nucleus (PVN). Baroreceptor stimulation promotes increased synaptic activity from NTS to CVLM, which projects a gamma-aminobutyric acid (GABA)-mediated activity to RVLM, reducing sympathetic drive. Chemoreceptor activation increases synaptic transmission from NTS-RVLM-IML, raising sympathetic drive. Besides, NTS-mediated chemoreceptor activation increases their activity to PVN and subsequently to RVLM and IML, promoting sympathetic activation. Created with BioRender.com.
The cardiovagal baroreflex is challenged in numerous conditions, such as during altitude exposure (Bourdillon et al., 2018a; Hermand et al., 2021; Bourdillon et al., 2023). However, understanding the underlying mechanisms of the effect of HA on BR remains limited (Olshansky et al., 2008; Beltran et al., 2020). Interestingly, chronic intermittent hypoxia (Swynghedauw, 1999; Freet et al., 2013), high altitude (Bourdillon et al., 2017a; Bourdillon et al., 2018a; Beltran et al., 2020; Bourdillon et al., 2023), and suffocation (Gu et al., 2007) promote similar physiological responses characterized by modified autonomic control assessed through heart rate variability (HRV) (sympathoexcitation and parasympathetic withdrawal) and, moreover, a marked decrease of BR assessed through sequence methods. Therefore, it is possible to suggest that the evidence at high altitude and chronic intermittent hypoxia could be, in part, comparable. Indeed, of interest is that the influence of barometric pressure seems negligible since no differences in the decreased baroreflex sensitivity were found between normobaric hypoxia and hypobaric hypoxia (Bourdillon et al., 2017b). It has been shown that after 30 days of chronic intermittent hypoxia exposure, which promotes hypertension (from the second day of hypoxia), there is a decrease in BR function and an increase in sympathetic outflow in conscious rats (Lai et al., 2006; Zoccal et al., 2009; Freet et al., 2013; Del Rio et al., 2016). However, the evidence is controversial, and no changes in BR-dependent splanchnic nerve activity after chronic intermittent hypoxia have been found (Prabhakar et al., 2012). Similarly, rats exposed to 7 days of intermittent hypoxia do not show any effects on BR function (Faulhaber et al., 2012). Moreover, the controversy is such that Zoccal et al. (2009), using heart-brain stem preparation, reported increased BR function in juvenile rats subjected to chronic intermittent hypoxia (Zoccal et al., 2009). Independent of the controversy, it has been proposed that hypoxic-dependent BR depression could be due to a reduction in the carotid baroreceptor activity rather than a direct effect on brainstem autonomic nuclei (Lesske et al., 1997). Nevertheless, rats exposed to chronic intermittent hypoxia improved BR sensitivity after CB denervation, even though the animals were still hypoxic, suggesting that the chemoreflex could be hierarchically superior to the BR (Del Rio et al., 2016).
During HA exposure, the human body responds at several levels, from cellular to whole-body, encompassing early and late responses. Early responses to HA involve ventilatory and circulatory adjustments, increased basal ventilation, autonomic imbalance marked by elevated release of epinephrine and norepinephrine, changes in acid-base balance, decreased glomerular filtration rate, and impaired pO2 and pCO2 exchange, and as a consequence a reduced exercise performance (Beall, 2007; Naeije, 2010; Farias et al., 2013; Huerta-Sanchez et al., 2014; Simonson et al., 2015; Mallet et al., 2021; Mallet et al., 2023). Late responses are characterized by gene overexpression, including hypoxic inducible factor [HIF]1α, HIF2α, vascular endothelial growth factor [VEGF], and VEGF receptor. Additionally, there is an increase in erythropoietin and hemoglobin, which are concomitant with an increase in hematocrit, leading to elevated blood viscosity. Other late responses include the activation of the renin–angiotensin–aldosterone system and alterations in intra- and extra-vascular fluid distribution (Beall, 2007; Naeije, 2010; Farias et al., 2013; Huerta-Sanchez et al., 2014; Simonson et al., 2015). Interestingly, most early responses are CB-mediated (Iturriaga et al., 2021b; Arce-Álvarez et al., 2022) (Figure 2). Regarding CB-mediated hypoxic ventilatory response, this exhibits several phases: i) initial short-term hyperpnea (Eldridge, 1974; Badr et al., 1992); ii) with continued hypoxic stimulus, there is time-dependent hyperventilation and sympathoexcitation that may persist for several hours or days (Wang et al., 2008); iii) if hyperventilation and increased sympathetic drive persist, it is possible to observe the decline of ventilation and not sympathoexcitation after several days of hypoxia (Hansen and Sander, 2003; Dempsey et al., 2014). Although peripheral chemoreceptor stimulation affects several organs in the body, most changes over the first hours of permanence in hypoxia occur at the level of the respiratory and cardiovascular systems (Bartsch and Gibbs, 2007). Notably, hypoxic-dependent local vasodilation is inhibited by CB (sympathetic)-mediated vasoconstriction in more metabolically activated organs (i.e., skeletal muscles during exercise) (Kumar and Bin-Jaliah, 2007). Vascular hyperactivity promotes two main effects: increment of arterial blood pressure and redistribution of blood flow, both relevant phenomena to physical fitness (Heistad and Abboud, 1980; Kumar and Bin-Jaliah, 2007; Dempsey and Smith, 2014). Accordingly, as was mentioned, chemoreflex activation promotes several physiological changes, which have been associated with homeostatic functions to meet metabolic demand. Then, considering that physical exercise triggers a greater metabolic requirement, it is possible to propose that CB may partially elicit cardiorespiratory and vascular responses by altering the BR function during HA exposure (Figure 2).
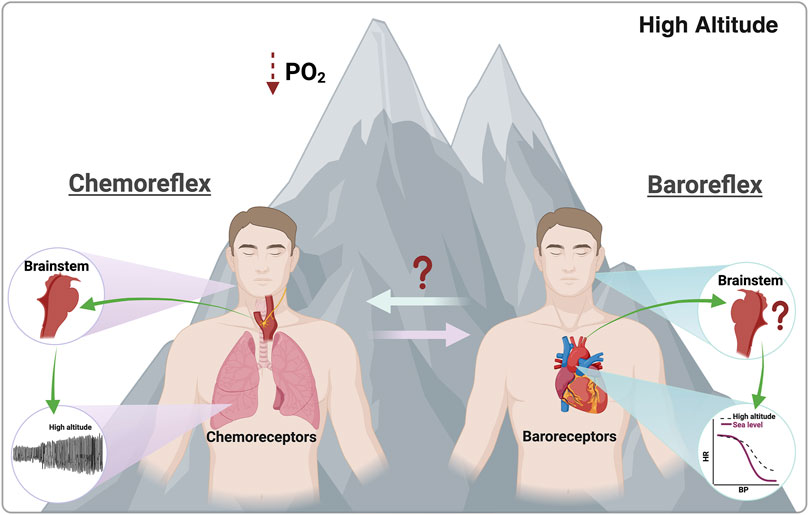
Figure 2. Effects promoted by chemoreflex and baroreflex activation. During high-altitude exposure, peripheral chemoreceptors are activated due to a PO2 reduction. Contrarily, during this environmental insult, the parasympathetic-dependent baroreflex control is reduced due to an overall autonomic control impairment. Notably, there is no evidence of the effect of high-altitude exposure on baroreflex-dependent central command and baroreceptors or the possible interaction between baroreflex and chemoreflex function. Created with BioRender.com.
The BR regulates hemodynamics during exercise (Fukuma et al., 2012; Dipla et al., 2013), which is dependent on age (Grassi et al., 2004; Fukuma et al., 2012), sex (Fukuma et al., 2012), and body fat distribution (Laterza et al., 2007). Further, decreased BR sensitivity has been associated with increased cardiovascular risk, cardiac electrical instability, and orthostatic intolerance (Fukuma et al., 2012). Otherwise, a normal BR function would ensure an appropriate cardiovascular response during exercise, regulating cardiac output (Fukuma et al., 2012). Conversely, an inappropriate high dose of exercise was associated with decreased baroreflex sensitivity, which may be used to diagnose overtraining (Bourdillon et al., 2018b). In physiological conditions, exercise training, similar to hypoxia, can promote resetting the BR function (Halliwill et al., 2003), shifting the BR to operate at higher blood pressure. In contrast, exercise training can induce robust effects on BR control in pathophysiological states, increasing the range, slope, and gain. However, only 25% of hypertensive subjects under chronic aerobic training could stabilize blood pressure, which could be associated with chronic morphological changes and not necessarily BR sensitivity (Liu et al., 2012). Indeed, it has been shown that exercise training can reduce sympathetic activity and increase BR-dependent parasympathetic function, determined through the sequence method, in an animal model of heart failure (Liu et al., 2000; Andrade et al., 2017). In addition, improvement in BR gain after an aerobic training program has been reported in pre-hypertensive and hypertensive subjects (Bertagnolli et al., 2008; Fisher et al., 2012; Liu et al., 2012). The effects of exercise training on BR are limited to cardiovascular diseases, with no demonstrated impact on metabolic disorders, such as diabetes mellitus. Indeed, Dipla et al. (2013) showed no significant association between changes in BR sensitivity in patients suffering from diabetes mellitus after applying a training program (Dipla et al., 2013). The evidence is controversial, and no effect of exercise training on BR has been found. Indeed, carotid BR sensitivity has remained unchanged after an aerobic training program compared with control subjects (Goldberg et al., 2012). Despite the controversy, the prevailing evidence indicates that exercise training significantly improves BR gain and triggers a resetting of the BR control. Nevertheless, it remains unclear whether the deterioration of cardiac BR gain during HA (Beltran et al., 2020) can be improved or remains unchanged compared to sea level following an exercise training intervention. Accordingly, further research is needed to address whether exercise training could be a feasible strategy to counteract the reduction of HA-induced BR gain.
CB is essential to maintain eupneic ventilation (Olson et al., 1988). However, although the evidence suggests that CB could be pivotal in cardiorespiratory response to exercise, apparently, the exercise did not modify CB chemoreflex, except in pathophysiological conditions (Andrade et al., 2018a; Wan et al., 2023). Thus, when CB-dependent hyperreflexia in autonomic-related diseases (i.e., heart failure, hypertension, etc.) is observed, exercise training reduces CB-dependent sympathetic overdrive and hypoxic ventilatory response (Schultz and Sun, 2000; Schultz et al., 2015). One of the first observations showing the relevance of CB chemoreceptors during exercise was made by Weil et al. (1972). They found that the hypoxic breathing response increased during moderate exercise compared to a resting condition, suggesting a potential association with the cardiorespiratory response to physical effort (Weil et al., 1972). Interestingly, peripheral chemoreceptor activation, similar to physical exercise, induces sympathoexcitation, promoting vasoconstriction at several levels, such as skeletal muscles and renal and mesenteric vascular beds (Seals et al., 1991; Gonzalez et al., 1994; Buckwalter and Clifford, 1999). Although vasoconstriction itself may limit blood flow to the muscle during exercise (Joyner et al., 1992), it has been proposed that exercise-dependent sympathoexcitation helps to distribute the blood flow to the active muscles according to their metabolic demand (Buckwalter and Clifford, 1999; Stickland et al., 2007). Even though sympathoexcitation during exercise is well-established, no conclusive evidence defines the principal sensor during exercise. It has been proposed that metaboreflex, exercise pressor reflex (constitute of metabo- and mechano-reflex), baroreflex, and chemoreflex are involved in cardiopulmonary response to exercise (Wan et al., 2023). Indeed, dopamine-dependent inhibition of chemoreflex decreases α-adrenoreceptor-mediated vasoconstriction, which augments blood flow to human active muscles (Stickland et al., 2011). Another mechanism that could increase ventilation during exercise is the increase in temperature. Nevertheless, CB is not essential in hyperventilation associated with exercise-induced hyperthermia (Daniłowicz-Szymanowicz et al., 2010; Fujii et al., 2019). Fujii et al. (2019) showed that the decrease in temperature and exposure to 100% O2 does not differentiate the ventilatory response to an incremental exercise, suggesting that peripheral chemoreceptors are irrelevant to exercise-induced hyperthermia in humans. The evidence indicates that the peripheral chemoreflex is involved in the ventilatory response to exercise. Nevertheless, there is no evidence whether the cardiorespiratory fitness deterioration during hypobaric hypoxia is due to altered chemoreflex function and whether it could be through an interaction with the BR.
It is well known that during physical activity, there is an increase in energy demand and, consequently, in cardiopulmonary and O2 uptake to meet the energy requirements (Hill and Lupton, 1923). In normoxia, during the initial phase of an incremental exercise, there is a cardiodynamic response, principally characterized by an increase in HR and stroke volume, with little contribution from ventilation to O2 consumption (Housh et al., 1991; McLellan and Cheung, 1992; Pringle and Jones, 2002; Whipp et al., 2005). It has been proposed that this cardiodynamic response is “associated” with the exercise pressor reflex (type III and IV afferent feedback fibers in muscles) and not with BR and chemoreflex, triggering an increase in ionotropic and chronotropic heart responses mediated by the activation of the sympathetic system (McCloskey and Mitchell, 1972; Amann et al., 2010; Tocco et al., 2015). At ventilatory threshold 1, there is a significant increase in pulmonary ventilation, which occurs in tandem with an increase in HR and lactate (Hofmann and Tschakert, 2017). Then, at ventilatory threshold 2, there is an abrupt increase in pulmonary ventilation and a significant increment of systemic lactate concentration (>4 mM) (Hofmann and Tschakert, 2017). It is currently under discussion whether chemoreceptors can detect lactate; therefore, the ventilatory response during exercise could be related to CB activation (Chang et al., 2015; Torres-Torrelo et al., 2021). Nevertheless, it has been shown that CB chemoreceptor cells do not respond to lactate in Wistar Kyoto rats (Spiller et al., 2021). In summary, during an incremental exercise, there is a complex, highly coordinated physiological mechanism encompassing pulmonary, cardiovascular, metabolic, and autonomic responses that enable the proper delivery of O2 to the active tissues. However, whether these mechanisms are modified during short-term HA exposure, which may be associated with chemo-baroreflex interaction, is not known yet.
As mentioned before, under resting conditions, HA promotes an increase in minute ventilation and enhances cardiac output, ensuring an adequate oxygen supply to the tissues (Klausen et al., 1966; Naeije et al., 1982; Cremona et al., 2002). It has been shown that a decrease of 12% in the inspired fraction of O2 can promote an increase of ∼22% in cardiac output, which was explained mainly by the increase of HR (∼18%) in healthy subjects (Naeije et al., 1982), decreasing the HR reserve. In addition, concomitant to cardiopulmonary adjustment, there is a marked autonomic control impairment characterized by a BR-dependent parasympathetic withdrawal during HA exposure (Naeije et al., 1982; Beltran et al., 2020). Regarding exercise at HA, it is well-established that the VO2max, exercise performance, and functional capacity are markedly reduced during HA exposure (Dempsey et al., 1972; Maher et al., 1974; Young et al., 1996; Fulco et al., 1998; Bassett and Howley, 2000; Millet et al., 2010; Andrade et al., 2018b; Burtscher et al., 2018). Indeed, highly trained running athletes evidenced a small but significant aerobic performance deterioration, even at 540 m (Fulco et al., 1998) or 580 m (Gore et al., 1996). Alexander et al. (1983) also found evidence that the VO2max is affected by HA, observing a decrease of 25% in maximum aerobic capacity at an altitude of 3,100 m (Alexander and Grover, 1983). Similarly, we found that aerobic time-trial performance decreased by ∼25% during acute exposure to 3,350 m in healthy individuals (Andrade et al., 2018b). Moreover, VO2max is expected to decrease by 0.9% per every 100 m over 1,100 m above sea level (Vogt and Hoppeler, 2010). All these phenomena have been attributed directly to the decrease in partial pressure and arterial SpO2 resulting from lower barometric pressure at HA, affecting the oxygen supply to active organs (Wagner, 2010). Slight differences have been reported between normobaric and hypobaric hypoxia (Millet and Debevec, 2020). However, the resting and exercise adjustments at HA suggest that autonomic control (chemoreflex and baroreflex) could play an important role in exercise performance during short-term HA exposure. Nevertheless, it is worth mentioning that there is no evidence indicating a chemo-baro interaction during HA exposure that could explain the reduction in exercise performance independent of reduced partial O2 pressure (Figure 3).
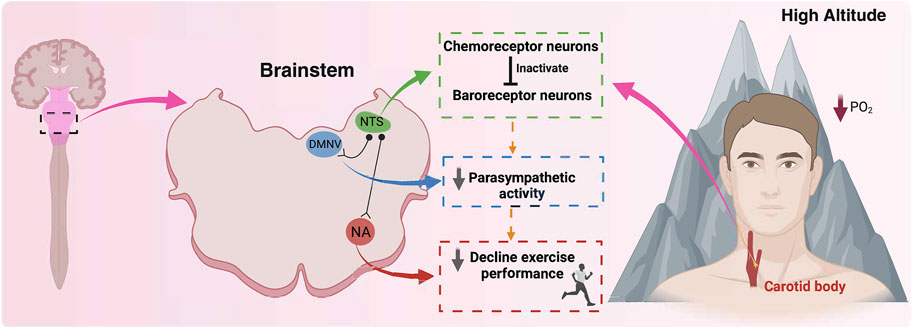
Figure 3. A hypothetical proposal related to the role of chemoreceptor and baroreceptor interaction and their effects on exercise performance at high altitudes. During high altitude exposure, there is an activation of the carotid body (CB) peripheral chemoreceptors, which at the brainstem level, specifically in the nucleus of the tractus solitarii (NTS) activate chemoreceptor neurons. At NTS, chemoreceptor neurons inhibit baroreceptor neurons, reducing their activity to parasympathetic neurons, such as the nucleus ambiguous (NA) and dorsal motor nucleus of the vagus (DMNV), consequently reducing baroreflex control and parasympathetic drive. Finally, the reduction of the vagal control negatively impacts exercise performance at high altitudes. Therefore, we propose that chemoreceptor neurons inactivate baroreceptor neurons, reducing parasympathetic drive and contributing to a decline in exercise performance at high altitudes, secondary to a reduction in barometric pressure. Created with BioRender.com.
Chemoreflex and baroreflex responses and interaction are modified during HA exposure, leading to a marked deterioration in exercise performance. Additionally, it has been determined that parasympathetic control is a determinant of exercise capacity. Therefore, it is reasonable to suggest that further research should focus on the role of chemoreflex and baroreflex-dependent vagal deterioration in the impairment of physical effort during HA exposure. It would be necessary to determine if the brainstem nuclei interact with the BR and the chemoreflex pathways, and their physiological consequences on exercise performance during hypobaric hypoxia. Finally, considering the critical importance of physical capacity at HA in several human activities (i.e., tourism, sports, border security, and mining) and the functional capacity in several chemoreflex/baroreflex-altered pathophysiological states, further research should not only elucidate the roles of chemoreflex and baroreflex separately but also explore potential pharmacological and non-pharmacological strategies to modify them as critical nodal points. This research should be broad in scope, focusing on physical performance and daily activities at high altitudes.
Hypoxic environments, while capable of harboring life, are inhospitable places where organisms struggle to survive. HA houses several human activities, such as tourism, sports, border security, and mining, which justify the research from basic to clinical science. Chemoreceptors and baroreceptors pathways share some brainstem neural nuclei, which are activated during hypoxia, making them susceptible to intervention and improving human whole-body response to hypoxia. Indeed, the present review focuses on chemoreflex and baroreflex interaction and their possible role in exercise capacity during HA exposure. The evidence suggests that an alteration of the chemoreflex could precede baroreflex-dependent parasympathetic withdrawal, resulting in a secondary impact on exercise performance at HA; however, this hypothesis has not yet been elucidated. Thus, our manuscript summarizes the literature and proposes new hypotheses that need to be addressed in the future.
PA-A: Validation, Visualization, Writing–original draft. SJ: Validation, Visualization, Writing–review and editing. CS-A: Validation, Visualization, Writing–review and editing. CN-E: Validation, Writing–review and editing. VP: Validation, Writing–review and editing. MR-F: Validation, Writing–review and editing. AR: Validation, Writing–review and editing. GM: Validation, Writing–review and editing. RI: Validation, Visualization, Writing–review and editing. DA: Conceptualization, Supervision, Validation, Visualization, Writing–review and editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This study was supported by the “Agencia Nacional de Investigación y Desarrollo (ANID)”, through Fondecyt de Iniciación #11220870 and Anillo ACT210083. MR-F. was supported by Fondecyt Regular #1230844 and Millennium Science Initiative Program ICN2021_004. RI was supported by Fondecyt Regular #1211443. DA received funding from Minera Escondida Ltda, through MEL2203. The funder was not involved in the study design, collection, analysis, interpretation of data, the writing of this article, or the decision to submit it for publication.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Accorsi-Mendonça D., Almado C. E. L., Bonagamba L. G. H., Castania J. A., Moraes D. J. A., Machado B. H. (2015). Enhanced firing in NTS induced by short-term sustained hypoxia is modulated by Glia-neuron interaction. J. Neurosci. official J. Soc. Neurosci. 35 (17), 6903–6917. doi:10.1523/JNEUROSCI.4598-14.2015
Accorsi-Mendonça D., Bonagamba L. G. H., Machado B. H. (2019). Astrocytic modulation of glutamatergic synaptic transmission is reduced in NTS of rats submitted to short-term sustained hypoxia. J. neurophysiology 212 (5), 1822–1830. doi:10.1152/jn.00279.2018
Alexander J. K., Grover R. F. (1983). Mechanism of reduced cardiac stroke volume at high altitude. Clin. Cardiol. 6 (6), 301–303. doi:10.1002/clc.4960060612
Al Haddad H., Mendez-Villanueva A., Bourdon P. C., Buchheit M. (2012). Effect of acute hypoxia on post-exercise parasympathetic reactivation in healthy men. Front. Physiol. 3, 289. doi:10.3389/fphys.2012.00289
Amann M., Blain G. M., Proctor L. T., Sebranek J. J., Pegelow D. F., Dempsey J. A. (2010). Group III and IV muscle afferents contribute to ventilatory and cardiovascular response to rhythmic exercise in humans. J. Appl. Physiol. (1985) 109 (4), 966–976. doi:10.1152/japplphysiol.00462.2010
Andrade D. C., Arce-Alvarez A., Toledo C., Díaz H. S., Lucero C., Quintanilla R. A., et al. (2018b). Revisiting the physiological effects of exercise training on autonomic regulation and chemoreflex control in heart failure: does ejection fraction matter? Am. J. physiology. Heart circulatory physiology 314 (3), H464–H474. doi:10.1152/ajpheart.00407.2017
Andrade D. C., Arce-Alvarez A., Toledo C., Díaz H. S., Lucero C., Schultz H. D., et al. (2017). Exercise training improves cardiac autonomic control, cardiac function, and arrhythmogenesis in rats with preserved-ejection fraction heart failure. J. Appl. Physiol. (1985) 123 (3), 567–577. doi:10.1152/japplphysiol.00189.2017
Andrade D. C., Díaz-Jara E., Toledo C., Schwarz K. G., Pereyra K. V., Díaz H. S., et al. (2021a). Exercise intolerance in volume overload heart failure is associated with low carotid body mediated chemoreflex drive. Sci. Rep. 11 (1), 14458. doi:10.1038/s41598-021-93791-8
Andrade D. C., Díaz-Jara E., Toledo C., Schwarz K. G., Pereyra K. V., Díaz H. S., et al. (2021b). Exercise intolerance in volume overload heart failure is associated with low carotid body mediated chemoreflex drive. Sci. Rep. 11 (1), 14458. doi:10.1038/s41598-021-93791-8
Andrade D. C., Iturriaga R., Jeton F., Alcayaga J., Voituron N., Del Rio R. (2018a). Acute effects of systemic erythropoietin injections on carotid body chemosensory activity following hypoxic and hypercapnic stimulation. Adv. Exp. Med. Biol. 1071, 95–102. doi:10.1007/978-3-319-91137-3_12
Arce-Álvarez A., Salazar-Ardiles C., Cornejo C., Paez V., Vásquez-Muñoz M., Stillner-Vilches K., et al. (2022). Chemoreflex control as the cornerstone in immersion water sports: possible role on breath-hold. Front. physiology 13, 894921. doi:10.3389/fphys.2022.894921
Badr M. S., Skatrud J. B., Dempsey J. A. (1992). Determinants of poststimulus potentiation in humans during NREM sleep. J. Appl. Physiol. (1985) 73 (5), 1958–1971. doi:10.1152/jappl.1992.73.5.1958
Bartsch P., Gibbs J. S. (2007). Effect of altitude on the heart and the lungs. Circulation 116 (19), 2191–2202. doi:10.1161/CIRCULATIONAHA.106.650796
Bartsch P., Saltin B. (2008). General introduction to altitude adaptation and mountain sickness. Scand. J. Med. Sci. Sports 18 (Suppl. 1), 1–10. doi:10.1111/j.1600-0838.2008.00827.x
Bassett D. R., Howley E. T. (2000). Limiting factors for maximum oxygen uptake and determinants of endurance performance. Med. Sci. Sports Exerc 32 (1), 70–84. doi:10.1097/00005768-200001000-00012
Beall C. M. (2000). Tibetan and Andean contrasts in adaptation to high-altitude hypoxia. Adv. Exp. Med. Biol. 475, 63–74. doi:10.1007/0-306-46825-5_7
Beall C. M. (2006). Andean, Tibetan, and Ethiopian patterns of adaptation to high-altitude hypoxia. Integr. Comp. Biol. 46 (1), 18–24. doi:10.1093/icb/icj004
Beall C. M. (2007). Two routes to functional adaptation: Tibetan and Andean high-altitude natives. Proc. Natl. Acad. Sci. U. S. A. 104 (Suppl. 1), 8655–8660. doi:10.1073/pnas.0701985104
Beltran A. R., Arce-Álvarez A., Ramirez-Campillo R., Vásquez-Muñoz M., von Igel M., Ramírez M. A., et al. (2020). Baroreflex modulation during acute high-altitude exposure in rats. Front. Physiol. 11, 1049. doi:10.3389/fphys.2020.01049
Bertagnolli M., Schenkel P. C., Campos C., Mostarda C. T., Casarini D. E., Belló-Klein A., et al. (2008). Exercise training reduces sympathetic modulation on cardiovascular system and cardiac oxidative stress in spontaneously hypertensive rats. Am. J. Hypertens. 21 (11), 1188–1193. doi:10.1038/ajh.2008.270
Bigham A. W., Wilson M. J., Julian C. G., Kiyamu M., Vargas E., Leon-Velarde F., et al. (2013). Andean and Tibetan patterns of adaptation to high altitude. Am. J. Hum. Biol. 25 (2), 190–197. doi:10.1002/ajhb.22358
Bourdillon N., Aebi M. R., Kayser B., Bron D., Millet G. P. (2023). Both hypoxia and hypobaria impair baroreflex sensitivity but through different mechanisms. Int. J. sports Med. 44 (3), 177–183. doi:10.1055/a-1960-3407
Bourdillon N., Saugy J., Schmitt L., Rupp T., Yazdani S., Vesin J. M., et al. (2017a). Acute and chronic changes in baroreflex sensitivity in hypobaric vs. normobaric hypoxia. Eur. J. Appl. Physiol. 117 (12), 2401–2407. doi:10.1007/s00421-017-3726-6
Bourdillon N., Saugy J., Schmitt L., Rupp T., Yazdani S., Vesin J. M., et al. (2017b). Acute and chronic changes in baroreflex sensitivity in hypobaric vs. normobaric hypoxia. Eur. J. Appl. Physiology 117, 2401–2407. doi:10.1007/s00421-017-3726-6
Bourdillon N., Yazdani S., Nilchian M., Mariano A., Vesin J. M., Millet G. P. (2018b). Overload blunts baroreflex only in overreached athletes. J. Sci. Med. Sport 21 (9), 941–949. doi:10.1016/j.jsams.2018.01.008
Bourdillon N., Yazdani S., Subudhi A. W., Lovering A. T., Roach R. C., Vesin J. M., et al. (2018a). AltitudeOmics: baroreflex sensitivity during acclimatization to 5,260 m. Front. Physiol. 9, 767. doi:10.3389/fphys.2018.00767
Boychuk C. R., Woerman A. L., Mendelowitz D. (2012). Modulation of bulbospinal rostral ventral lateral medulla neurons by hypoxia/hypercapnia but not medullary respiratory activity. Hypertension 60 (6), 1491–1497. doi:10.1161/HYPERTENSIONAHA.112.197954
Buckwalter J. B., Clifford P. S. (1999). alpha-adrenergic vasoconstriction in active skeletal muscles during dynamic exercise. Am. J. Physiol. 277 (1), H33–H39. doi:10.1152/ajpheart.1999.277.1.H33
Burtscher M., Niedermeier M., Burtscher J., Pesta D., Suchy J., Strasser B. (2018). Preparation for endurance competitions at altitude: physiological, psychological, dietary and coaching aspects. A narrative review. Front. physiology 9, 1504. doi:10.3389/fphys.2018.01504
Chang A. J., Ortega F. E., Riegler J., Madison D. V., Krasnow M. A. (2015). Oxygen regulation of breathing through an olfactory receptor activated by lactate. Nature 527 (7577), 240–244. doi:10.1038/nature15721
Claps A., Torrealba F. (1988). The carotid body connections: a WGA-HRP study in the cat. Brain Res. 455 (1), 123–133. doi:10.1016/0006-8993(88)90121-7
Cremona G., Asnaghi R., Baderna P., Brunetto A., Brutsaert T., Cavallaro C., et al. (2002). Pulmonary extravascular fluid accumulation in recreational climbers: a prospective study. Lancet 359 (9303), 303–309. doi:10.1016/s0140-6736(02)07496-2
D’Agostino D., Hoang-Le T., Mazza E., Neubauer J., Ritucci N., Sobota J., et al. (2001). Hypoxic chemosensitivity of the cardiorespiratory regions of the rostral ventrolateral medulla (RVLM). Respir. Res. 2 (1), 5.2. doi:10.1186/rr115
Daniłowicz-Szymanowicz L., Raczak G., Szwoch M., Ratkowski W., Toruński A. (2010). The effect of anaerobic and aerobic tests on autonomic nervous system activity in healthy young athletes. Biol. sport 27 (1), 65–69. doi:10.5604/20831862.907954
Del Rio R., Andrade D. C., Lucero C., Arias P., Iturriaga R. (2016). Carotid body ablation abrogates hypertension and autonomic alterations induced by intermittent hypoxia in rats. Hypertension 68 (2), 436–445. doi:10.1161/HYPERTENSIONAHA.116.07255
Del Rio R., Andrade D. C., Marcus N. J., Schultz H. D. (2015). Selective carotid body ablation in experimental heart failure: a new therapeutic tool to improve cardiorespiratory control. Exp. Physiol. 100 (2), 136–142. doi:10.1113/expphysiol.2014.079566
Del Rio R., Marcus N. J., Schultz H. D. (2013). Carotid chemoreceptor ablation improves survival in heart failure: rescuing autonomic control of cardiorespiratory function. J. Am. Coll. Cardiol. 62 (25), 2422–2430. doi:10.1016/j.jacc.2013.07.079
Del Rio R., Moya E. A., Koenig C. S., Fujiwara K., Alcayaga J., Iturriaga R. (2008). Modulatory effects of histamine on cat carotid body chemoreception. Respir. Physiol. Neurobiol. 164 (3), 401–410. doi:10.1016/j.resp.2008.09.005
Dempsey J. A., Forster H. V., Birnbaum M. L., Reddan W. G., Thoden J., Grover R. F., et al. (1972). Control of exercise hyperpnea under varying durations of exposure to moderate hypoxia. Respir. Physiol. 16 (2), 213–231. doi:10.1016/0034-5687(72)90052-7
Dempsey J. A., Powell F. L., Bisgard G. E., Blain G. M., Poulin M. J., Smith C. A. (2014). Role of chemoreception in cardiorespiratory acclimatization to, and deacclimatization from, hypoxia. J. Appl. Physiol. (1985) 116 (7), 858–866. doi:10.1152/japplphysiol.01126.2013
Dempsey J. A., Smith C. A. (2014). Pathophysiology of human ventilatory control. Eur. Respir. J. 44 (2), 495–512. doi:10.1183/09031936.00048514
Diaz H. S., Toledo C., Andrade D. C., Marcus N. J., Del Rio R. (2020). Neuroinflammation in heart failure: new insights for an old disease. J. Physiol. 598 (1), 33–59. doi:10.1113/JP278864
Ding Y., Li Y. L., Schultz H. D. (2011). Role of blood flow in carotid body chemoreflex function in heart failure. J. Physiol. 589 (Pt 1), 245–258. doi:10.1113/jphysiol.2010.200584
Dipla K., Papadopoulos S., Zafeiridis A., Kyparos A., Nikolaidis M. G., Vrabas I. S. (2013). Determinants of muscle metaboreflex and involvement of baroreflex in boys and young men. Eur. J. Appl. Physiol. 113 (4), 827–838. doi:10.1007/s00421-012-2493-7
Eldridge F. L. (1974). Central neural respiratory stimulatory effect of active respiration. J. Appl. Physiol. 37 (5), 723–735. doi:10.1152/jappl.1974.37.5.723
Eugenin J., Larrain C., Zapata P. (2020). Plasticity of cardiovascular chemoreflexes after prolonged unilateral carotid body denervation: implications for its therapeutic use. Am. J. Physiol. Heart Circ. Physiol. 318 (5), H1325–H1336. doi:10.1152/ajpheart.00451.2019
Fadel P. J., Stromstad M., Wray D. W., Smith S. A., Raven P. B., Secher N. H. (2003). New insights into differential baroreflex control of heart rate in humans. Am. J. physiology. Heart circulatory physiology 284 (2), H735–H743. doi:10.1152/ajpheart.00246.2002
Farias J. G., Jimenez D., Osorio J., Zepeda A. B., Figueroa C. A., Pulgar V. M. (2013). Acclimatization to chronic intermittent hypoxia in mine workers: a challenge to mountain medicine in Chile. Biol. Res. 46 (1), 59–67. doi:10.4067/S0716-97602013000100009
Faulhaber M., Dünnwald T., Gatterer H., Bernardi L., Burtscher M. (2012). Metabolic adaptations may counteract ventilatory adaptations of intermittent hypoxic exposure during submaximal exercise at altitudes up to 4000 m. PloS one 7 (11), e49953. doi:10.1371/journal.pone.0049953
Finley J. C., Katz D. M. (1992). The central organization of carotid body afferent projections to the brainstem of the rat. Brain Res. 572 (1-2), 108–116. doi:10.1016/0006-8993(92)90458-l
Fisher J. P., Kim A., Hartwich D., Fadel P. J. (2012). New insights into the effects of age and sex on arterial baroreflex function at rest and during dynamic exercise in humans. Aut. Neurosci. basic & Clin. 172 (1-2), 13–22. doi:10.1016/j.autneu.2012.10.013
Fornasiero A., Savoldelli A., Skafidas S., Stella F., Bortolan L., Boccia G., et al. (2018). Delayed parasympathetic reactivation and sympathetic withdrawal following maximal cardiopulmonary exercise testing (CPET) in hypoxia. Eur. J. Appl. Physiol. 118 (10), 2189–2201. doi:10.1007/s00421-018-3945-5
Freet C. S., Stoner J. F., Tang X. (2013). Baroreflex and chemoreflex controls of sympathetic activity following intermittent hypoxia. Auton. Neurosci. 174 (1-2), 8–14. doi:10.1016/j.autneu.2012.12.005
Fujii N., Kashihara M., Kenny G. P., Honda Y., Fujimoto T., Cao Y., et al. (2019). Carotid chemoreceptors have a limited role in mediating the hyperthermia-induced hyperventilation in exercising humans. J. Appl. physiology (Bethesda, Md. 1985) 126 (2), 305–313. doi:10.1152/japplphysiol.00562.2018
Fukuma N., Kato K., Munakata K., Hayashi H., Kato Y., Aisu N., et al. (2012). Baroreflex mechanisms and response to exercise in patients with heart disease. Clin. Physiol. Funct. Imaging 32 (4), 305–309. doi:10.1111/j.1475-097X.2012.01127.x
Fulco C. S., Rock P. B., Cymerman A. (1998). Maximal and submaximal exercise performance at altitude. Aviat. space, Environ. Med. 69 (8), 793–801.
Goldberg M. J., Boutcher S. H., Boutcher Y. N. (2012). The effect of 4 weeks of aerobic exercise on vascular and baroreflex function of young men with a family history of hypertension. J. Hum. Hypertens. 26 (11), 644–649. doi:10.1038/jhh.2011.95
Gonzalez C., Almaraz L., Obeso A., Rigual R. (1994). Carotid body chemoreceptors: from natural stimuli to sensory discharges. Physiol. Rev. 74 (4), 829–898. doi:10.1152/physrev.1994.74.4.829
Gore C. J., Hahn A. G., Scroop G. C., Watson D. B., Norton K. I., Wood R. J., et al. (1996). Increased arterial desaturation in trained cyclists during maximal exercise at 580 m altitude. J. Appl. physiology (Bethesda, Md. 1985) 80 (6), 2204–2210. doi:10.1152/jappl.1996.80.6.2204
Grassi G., Dell’Oro R., Facchini A., Quarti Trevano F., Bolla G. B., Mancia G. (2004). Effect of central and peripheral body fat distribution on sympathetic and baroreflex function in obese normotensives. J. Hypertens. 22 (12), 2363–2369. doi:10.1097/00004872-200412000-00019
Gray P. C., Scott J. D., Catterall W. A. (1998). Regulation of ion channels by cAMP-dependent protein kinase and A-kinase anchoring proteins. Curr. Opin. Neurobiol. 8 (3), 330–334. doi:10.1016/s0959-4388(98)80057-3
Greksa L. P., Spielvogel H., Paz-Zamora M., Caceres E., Paredes-Fernández L. (1988). Effect of altitude on the lung function of high altitude residents of European ancestry. Am. J. Phys. Anthropol. 75 (1), 77–85. doi:10.1002/ajpa.1330750109
Gu H., Lin M., Liu J., Gozal D., Scrogin K. E., Wurster R., et al. (2007). Selective impairment of central mediation of baroreflex in anesthetized young adult Fischer 344 rats after chronic intermittent hypoxia. Am. J. Physiol. Heart Circ. Physiol. 293 (5), H2809–H2818. doi:10.1152/ajpheart.00358.2007
Halliwill J. R., Morgan B. J., Charkoudian N. (2003). Peripheral chemoreflex and baroreflex interactions in cardiovascular regulation in humans. J. Physiol. 552 (Pt 1), 295–302. doi:10.1113/jphysiol.2003.050708
Hansen J., Sander M. (2003). Sympathetic neural overactivity in healthy humans after prolonged exposure to hypobaric hypoxia. J. Physiol. 546 (Pt 3), 921–929. doi:10.1113/jphysiol.2002.031765
Heistad D. D., Abboud F. M. (1980). Dickinson W. Richards lecture: circulatory adjustments to hypoxia. Circulation 61 (3), 463–470. doi:10.1161/01.cir.61.3.463
Hermand E., Lhuissier F. J., Pichon A., Voituron N., Richalet J. P. (2021). Exercising in hypoxia and other stimuli: heart rate variability and ventilatory oscillations. Life (Basel) 11 (7), 625. doi:10.3390/life11070625
Hill A. V., Lupton H. (1923). Muscular exercise, lactic acid, and the supply and utilization of oxygen. QJM Mon. J. Assoc. Physicians 16 (62), 135–171. doi:10.1093/qjmed/os-16.62.135
Hofmann P., Tschakert G. (2017). Intensity- and duration-based options to regulate endurance training. Front. Physiol. 8, 337. doi:10.3389/fphys.2017.00337
Honda Y. (1985). Role of carotid chemoreceptors in control of breathing at rest and in exercise: studies on human subjects with bilateral carotid body resection. Jpn. J. physiology 35 (4), 535–544. doi:10.2170/jjphysiol.35.535
Honda Y., Watanabe S., Hashizume I., Satomura Y., Hata N., Sakakibara Y., et al. (1979). Hypoxic chemosensitivity in asthmatic patients two decades after carotid body resection. J. Appl. Physiol. Respir. Environ. Exerc Physiol. 46 (4), 632–638. doi:10.1152/jappl.1979.46.4.632
Housh T. J., Devries H. A., Housh D. J., Tichy M. W., Smyth K. D., Tichy A. M. (1991). The relationship between critical power and the onset of blood lactate accumulation. J. sports Med. Phys. Fit. 31 (1), 31–36.
Huerta-Sanchez E., Jin X., Asan , Bianba Z., Peter B. M., Vinckenbosch N., et al. (2014). Altitude adaptation in Tibetans caused by introgression of Denisovan-like DNA. Nature 512 (7513), 194–197. doi:10.1038/nature13408
Huey R. B. (2002). High altitude: an exploration of human adaptation. Integr. Comp. Biol. 42 (4), 910. doi:10.1093/icb/42.4.910
Iturriaga R., Alcayaga J. (2004). Neurotransmission in the carotid body: transmitters and modulators between glomus cells and petrosal ganglion nerve terminals. Brain Res. Rev. 47 (1-3), 46–53. doi:10.1016/j.brainresrev.2004.05.007
Iturriaga R., Alcayaga J., Chapleau M., Somers V. (2021a). Carotid body chemoreceptors: physiology, pathology, and implications for health and disease. Physiol. Rev. 101, 1177–1235. doi:10.1152/physrev.00039.2019
Iturriaga R., Alcayaga J., Chapleau M. W., Somers V. K. (2021b). Carotid body chemoreceptors: physiology, pathology, and implications for health and disease. Physiol. Rev. 101 (3), 1177–1235. doi:10.1152/physrev.00039.2019
Joyner M. J., Nauss L. A., Warner M. A., Warner D. O. (1992). Sympathetic modulation of blood flow and O2 uptake in rhythmically contracting human forearm muscles. Am. J. Physiol. 263 (4 Pt 2), H1078–H1083. doi:10.1152/ajpheart.1992.263.4.H1078
Katayama P. L., Castania J. A., Fazan R., Salgado H. C. (2019). Interaction between baroreflex and chemoreflex in the cardiorespiratory responses to stimulation of the carotid sinus/nerve in conscious rats. Aut. Neurosci. basic & Clin. 216, 17–24. doi:10.1016/j.autneu.2018.12.001
Kaupp U. B., Seifert R. (2001). Molecular diversity of pacemaker ion channels. Annu. Rev. Physiol. 63, 235–257. doi:10.1146/annurev.physiol.63.1.235
King T. L., Heesch C. M., Clark C. G., Kline D. D., Hasser E. M. (2012). Hypoxia activates nucleus tractus solitarii neurons projecting to the paraventricular nucleus of the hypothalamus. Am. J. Physiol. Regul. Integr. Comp. Physiol. 302 (10), R1219–R1232. doi:10.1152/ajpregu.00028.2012
King T. L., Kline D. D., Ruyle B. C., Heesch C. M., Hasser E. M. (2013). Acute systemic hypoxia activates hypothalamic paraventricular nucleus-projecting catecholaminergic neurons in the caudal ventrolateral medulla. Am. J. Physiol. Regul. Integr. Comp. Physiol. 305 (10), R1112–R1123. doi:10.1152/ajpregu.00280.2013
Klausen K., Robinson S., Micahel E. D., Myhre L. G. (1966). Effect of high altitude on maximal working capacity. J. Appl. Physiol. 21 (4), 1191–1194. doi:10.1152/jappl.1966.21.4.1191
Kougias P., Weakley S. M., Yao Q., Lin P. H., Chen C. (2010). Arterial baroreceptors in the management of systemic hypertension. Med. Sci. Monit. 16 (1), RA1–8.
Kronsbein H., Gerlach D. A., Heusser K., Hoff A., Hoffmann F., Diedrich A., et al. (2020). Testing individual baroreflex responses to hypoxia-induced peripheral chemoreflex stimulation. Clin. Auton. Res. 30 (6), 531–540. doi:10.1007/s10286-019-00660-6
Kumar P., Bin-Jaliah I. (2007). Adequate stimuli of the carotid body: more than an oxygen sensor? Respir. physiology Neurobiol. 157 (1), 12–21. doi:10.1016/j.resp.2007.01.007
Lai C. J., Yang C. C. H., Hsu Y. Y., Lin Y. N., Kuo T. B. J. (2006). Enhanced sympathetic outflow and decreased baroreflex sensitivity are associated with intermittent hypoxia-induced systemic hypertension in conscious rats. J. Appl. Physiol. (1985) 100 (6), 1974–1982. doi:10.1152/japplphysiol.01051.2005
Lakatta E. G., DiFrancesco D. (2009). What keeps us ticking: a funny current, a calcium clock, or both? J. Mol. Cell. Cardiol. 47 (2), 157–170. doi:10.1016/j.yjmcc.2009.03.022
Laterza M. C., de Matos L. D. N. J., Trombetta I. C., Braga A. M. W., Roveda F., Alves M. J. N. N., et al. (2007). Exercise training restores baroreflex sensitivity in never-treated hypertensive patients. Hypertension 49 (6), 1298–1306. doi:10.1161/HYPERTENSIONAHA.106.085548
Lau E.O.-C., Lo C. Y., Yao Y., Mak A. F. T., Jiang L., Huang Y., et al. (2016). Aortic baroreceptors display higher mechanosensitivity than carotid baroreceptors. Front. physiology 7, 384. doi:10.3389/fphys.2016.00384
Lesske J., Fletcher E. C., Bao G., Unger T. (1997). Hypertension caused by chronic intermittent hypoxia--influence of chemoreceptors and sympathetic nervous system. J. Hypertens. 15 (12 Pt 2), 1593–1603. doi:10.1097/00004872-199715120-00060
Liu J. L., Irvine S., Reid I. A., Patel K. P., Zucker I. H. (2000). Chronic exercise reduces sympathetic nerve activity in rabbits with pacing-induced heart failure: a role for angiotensin II. Circulation 102 (15), 1854–1862. doi:10.1161/01.cir.102.15.1854
Liu S., Goodman J., Nolan R., Lacombe S., Thomas S. G. (2012). Blood pressure responses to acute and chronic exercise are related in prehypertension. Med. Sci. Sports Exerc 44 (9), 1644–1652. doi:10.1249/MSS.0b013e31825408fb
Machhada A., Trapp S., Marina N., Stephens R. C. M., Whittle J., Lythgoe M. F., et al. (2017). Vagal determinants of exercise capacity. Nat. Commun. 8, 15097. doi:10.1038/ncomms15097
Mahamed S., Duffin J. (2001). Repeated hypoxic exposures change respiratory chemoreflex control in humans. J. Physiol. 534 (Pt. 2), 595–603. doi:10.1111/j.1469-7793.2001.00595.x
Maher J. T., Jones L. G., Hartley L. H. (1974). Effects of high-altitude exposure on submaximal endurance capacity of men. J. Appl. Physiol. 37 (6), 895–898. doi:10.1152/jappl.1974.37.6.895
Mallet R. T., Burtscher J., Pialoux V., Pasha Q., Ahmad Y., Millet G. P., et al. (2023). Molecular mechanisms of high-altitude acclimatization. Int. J. Mol. Sci. 24 (2), 1698. doi:10.3390/ijms24021698
Mallet R. T., Burtscher J., Richalet J. P., Millet G. P., Burtscher M. (2021). Impact of high altitude on cardiovascular health: current perspectives. Vasc. Health Risk Manag. 17, 317–335. doi:10.2147/VHRM.S294121
Marx S. O., Kurokawa J., Reiken S., Motoike H., D’Armiento J., Marks A. R., et al. (2002). Requirement of a macromolecular signaling complex for beta adrenergic receptor modulation of the KCNQ1-KCNE1 potassium channel. Science 295 (5554), 496–499. doi:10.1126/science.1066843
Masuda A., Paulev P. E., Sakakibara Y., Ahn B., Takaishi S., Pokorski M., et al. (1988). Estimation of peripheral chemoreceptor contribution to exercise hyperpnea in man. Jpn. J. Physiol. 38 (5), 607–618. doi:10.2170/jjphysiol.38.607
McCloskey D. I., Mitchell J. H. (1972). Reflex cardiovascular and respiratory responses originating in exercising muscle. J. Physiol. 224 (1), 173–186. doi:10.1113/jphysiol.1972.sp009887
McLellan T. M., Cheung K. S. (1992). A comparative evaluation of the individual anaerobic threshold and the critical power. Med. Sci. Sports Exerc 24 (5), 543–550. doi:10.1249/00005768-199205000-00008
Millet G. P., Debevec T. (2020). CrossTalk proposal: barometric pressure, independent of P O 2, is the forgotten parameter in altitude physiology and mountain medicine. J. physiology 598 (5), 893–896. doi:10.1113/JP278673
Millet G. P., Roels B., Schmitt L., Woorons X., Richalet J. P. (2010). Combining hypoxic methods for peak performance. Sports Med. 40 (1), 1–25. doi:10.2165/11317920-000000000-00000
Murray A. J., Montgomery H. E., Feelisch M., Grocott M. P. W., Martin D. S. (2018). Metabolic adjustment to high-altitude hypoxia: from genetic signals to physiological implications. Biochem. Soc. Trans. 46 (3), 599–607. doi:10.1042/BST20170502
Naeije R. (2010). Physiological adaptation of the cardiovascular system to high altitude. Prog. Cardiovasc Dis. 52 (6), 456–466. doi:10.1016/j.pcad.2010.03.004
Naeije R., Melot C., Mols P., Hallemans R. (1982). Effects of vasodilators on hypoxic pulmonary vasoconstriction in normal man. Chest 82 (4), 404–410. doi:10.1378/chest.82.4.404
Olshansky B., Sabbah H. N., Hauptman P. J., Colucci W. S. (2008). Parasympathetic nervous system and heart failure: pathophysiology and potential implications for therapy. Circulation 118 (8), 863–871. doi:10.1161/CIRCULATIONAHA.107.760405
Olson E. B., Vidruk E. H., Dempsey J. A. (1988). Carotid body excision significantly changes ventilatory control in awake rats. J. Appl. physiology (Bethesda, Md. 1985) 64 (2), 666–671. doi:10.1152/jappl.1988.64.2.666
Pamenter M. E., Carr J. A., Go A., Fu Z., Reid S. G., Powell F. L. (2014). Glutamate receptors in the nucleus tractus solitarius contribute to ventilatory acclimatization to hypoxia in rat. J. Physiol. 592 (8), 1839–1856. doi:10.1113/jphysiol.2013.268706
Perini R., Milesi S., Biancardi L., Veicsteinas A. (1996). Effects of high altitude acclimatization on heart rate variability in resting humans. Eur. J. Appl. Physiol. Occup. Physiol. 73 (6), 521–528. doi:10.1007/BF00357674
Persson P., Ehmke H., Kirchheim H., Seller H. (1988). Effect of sino-aortic denervation in comparison to cardiopulmonary deafferentiation on long-term blood pressure in conscious dogs. Pflugers Arch. 411 (2), 160–166. doi:10.1007/BF00582309
Pijacka W., Katayama P. L., Salgado H. C., Lincevicius G. S., Campos R. R., McBryde F. D., et al. (2018). Variable role of carotid bodies in cardiovascular responses to exercise, hypoxia and hypercapnia in spontaneously hypertensive rats. J. Physiol. 596 (15), 3201–3216. doi:10.1113/JP275487
Ponikowski P., Chua T. P., Anker S. D., Francis D. P., Doehner W., Banasiak W., et al. (2001). Peripheral chemoreceptor hypersensitivity: an ominous sign in patients with chronic heart failure. Circulation 104 (5), 544–549. doi:10.1161/hc3101.093699
Prabhakar N. R. (2006). O2 sensing at the mammalian carotid body: why multiple O2 sensors and multiple transmitters? O2sensing at carotid body. Exp. Physiol. 91 (1), 17–23. doi:10.1113/expphysiol.2005.031922
Prabhakar N. R., Kumar G. K., Peng Y. J. (2012). Sympatho-adrenal activation by chronic intermittent hypoxia. J. Appl. Physiol. (1985) 113 (8), 1304–1310. doi:10.1152/japplphysiol.00444.2012
Pringle J. S., Jones A. M. (2002). Maximal lactate steady state, critical power and EMG during cycling. Eur. J. Appl. Physiol. 88 (3), 214–226. doi:10.1007/s00421-002-0703-4
Rosin D. L., Chang D. A., Guyenet P. G. (2006). Afferent and efferent connections of the rat retrotrapezoid nucleus. J. Comp. Neurol. 499 (1), 64–89. doi:10.1002/cne.21105
Sampson K. J., Kass R. S. (2010). Molecular mechanisms of adrenergic stimulation in the heart. Heart rhythm. 7 (8), 1151–1153. doi:10.1016/j.hrthm.2010.02.013
Schmitt L., Fouillot J. P., Millet G. P., Robach P., Nicolet G., Brugniaux J., et al. (2008). Altitude, heart rate variability and aerobic capacities. Int. J. Sports Med. 29 (4), 300–306. doi:10.1055/s-2007-965355
Schultz H. D., Marcus N. J., Del Rio R. (2015). Mechanisms of carotid body chemoreflex dysfunction during heart failure. Exp. Physiol. 100 (2), 124–129. doi:10.1113/expphysiol.2014.079517
Schultz H. D., Sun S. Y. (2000). Chemoreflex function in heart failure. Heart Fail Rev. 5 (1), 45–56. doi:10.1023/A:1009846123893
Seals D. R., Johnson D. G., Fregosi R. F. (1991). Hyperoxia lowers sympathetic activity at rest but not during exercise in humans. Am. J. physiology 260 (5 Pt 2), R873–R878. doi:10.1152/ajpregu.1991.260.5.R873
Sevre K., Bendz B., Hankø E., Nakstad A. R., Hauge A., Kåsin J. I., et al. (2001). Reduced autonomic activity during stepwise exposure to high altitude. Acta Physiol. Scand. 173 (4), 409–417. doi:10.1046/j.1365-201X.2001.00925.x
Simonson T. S., Huff C. D., Witherspoon D. J., Prchal J. T., Jorde L. B. (2015). Adaptive genetic changes related to haemoglobin concentration in native high-altitude Tibetans. Exp. Physiol. 100 (11), 1263–1268. doi:10.1113/EP085035
Smith C. A., Forster H. V., Blain G. M., Dempsey J. A. (2010). An interdependent model of central/peripheral chemoreception: evidence and implications for ventilatory control. Respir. Physiol. Neurobiol. 173 (3), 288–297. doi:10.1016/j.resp.2010.02.015
Somers V. K., Mark A. L., Abboud F. M. (1991). Interaction of baroreceptor and chemoreceptor reflex control of sympathetic nerve activity in normal humans. J. Clin. Invest. 87 (6), 1953–1957. doi:10.1172/JCI115221
Spiller P. F., da Silva M. P., Moraes D. J. A. (2021). Lactate does not activate the carotid body of Wistar rat. Respir. physiology Neurobiol. 285 (103593), 103593. doi:10.1016/j.resp.2020.103593
Stickland M. K., Fuhr D. P., Haykowsky M. J., Jones K. E., Paterson D. I., Ezekowitz J. A., et al. (2011). Carotid chemoreceptor modulation of blood flow during exercise in healthy humans. J. Physiol. 589 (Pt 24), 6219–6230. doi:10.1113/jphysiol.2011.218099
Stickland M. K., Miller J. D., Smith C. A., Dempsey J. A. (2007). Carotid chemoreceptor modulation of regional blood flow distribution during exercise in health and chronic heart failure. Circ. Res. 100 (9), 1371–1378. doi:10.1161/01.RES.0000266974.84590.d2
Subudhi A. W., Fan J. L., Evero O., Bourdillon N., Kayser B., Julian C. G., et al. (2014). AltitudeOmics: effect of ascent and acclimatization to 5260 m on regional cerebral oxygen delivery. Exp. Physiol. 99 (5), 772–781. doi:10.1113/expphysiol.2013.075184
Swynghedauw B. (1999). Molecular mechanisms of myocardial remodeling. Physiol. Rev. 79 (1), 215–262. doi:10.1152/physrev.1999.79.1.215
Teppema L. J., Dahan A. (2010). The ventilatory response to hypoxia in mammals: mechanisms, measurement, and analysis. Physiol. Rev. 90 (2), 675–754. doi:10.1152/physrev.00012.2009
Thrasher T. N. (2002). Unloading arterial baroreceptors causes neurogenic hypertension. Am. J. Physiol. Regul. Integr. Comp. Physiol. 282 (4), R1044–R1053. doi:10.1152/ajpregu.00431.2001
Tocco F., Crisafulli A., Milia R., Marongiu E., Mura R., Roberto S., et al. (2015). Nervous facilitation in cardiodynamic response of exercising athletes to superimposed mental tasks: implications in depressive disorder. Clin. Pract. Epidemiol. Ment. Health 11, 166–173. doi:10.2174/1745017901511010166
Torres-Torrelo H., Ortega-Saenz P., Gao L., Lopez-Barneo J. (2021). Lactate sensing mechanisms in arterial chemoreceptor cells. Nat. Commun. 12 (1), 4166. doi:10.1038/s41467-021-24444-7
Vizek M., Pickett C. K., Weil J. V. (1987). Increased carotid body hypoxic sensitivity during acclimatization to hypobaric hypoxia. J. Appl. Physiol. (1985) 63, 2403–2410. doi:10.1152/jappl.1987.63.6.2403
Vogt M., Hoppeler H. (2010). Is hypoxia training good for muscles and exercise performance? Prog. Cardiovasc. Dis. 52 (6), 525–533. doi:10.1016/j.pcad.2010.02.013
Wagner P. D. (2010). The physiological basis of reduced VO2max in Operation Everest II. High. Alt. Med. Biol. 11 (3), 209–215. doi:10.1089/ham.2009.1058
Wan H. Y., Bunsawat K., Amann M. (2023). Autonomic cardiovascular control during exercise. Am. J. Physiol. Heart Circ. Physiol. 325 (4), H675–H686. doi:10.1152/ajpheart.00303.2023
Wang Z. Y., Olson E. B., Bjorling D. E., Mitchell G. S., Bisgard G. E. (2008). Sustained hypoxia-induced proliferation of carotid body type I cells in rats. J. Appl. Physiol. (1985) 104 (3), 803–808. doi:10.1152/japplphysiol.00393.2007
Wehrlin J. P., Hallen J. (2006). Linear decrease in.VO2max and performance with increasing altitude in endurance athletes. Eur. J. Appl. Physiol. 96 (4), 404–412. doi:10.1007/s00421-005-0081-9
Weil J. V., Byrne-Quinn E., Sodal I. E., Kline J. S., McCullough R. E., Filley G. F. (1972). Augmentation of chemosensitivity during mild exercise in normal man. J. Appl. Physiol. 33 (6), 813–819. doi:10.1152/jappl.1972.33.6.813
Whipp B. J., Ward S. A., Rossiter H. B. (2005). Pulmonary O2 uptake during exercise: conflating muscular and cardiovascular responses. Med. Sci. Sports Exerc 37 (9), 1574–1585. doi:10.1249/01.mss.0000177476.63356.22
Windsor J. S., Rodway G. W. (2007). Heights and haematology: the story of haemoglobin at altitude. Postgrad. Med. J. 83 (977), 148–151. doi:10.1136/pgmj.2006.049734
Young A. J., Sawka M. N., Muza S. R., Boushel R., Lyons T., Rock P. B., et al. (1996). Effects of erythrocyte infusion on VO2max at high altitude. J. Appl. physiology (Bethesda, Md. 1985) 81 (1), 252–259. doi:10.1152/jappl.1996.81.1.252
Zhang W., Carreño F. R., Cunningham J. T., Mifflin S. W. (2009). Chronic sustained hypoxia enhances both evoked EPSCs and norepinephrine inhibition of glutamatergic afferent inputs in the nucleus of the solitary tract. he J. Neurosci. official J. Soc. Neurosci. 29 (10), 3093–3102. doi:10.1523/JNEUROSCI.2648-08.2009
Zoccal D. B., Bonagamba L. G. H., Paton J. F. R., Machado B. H. (2009). Sympathetic-mediated hypertension of awake juvenile rats submitted to chronic intermittent hypoxia is not linked to baroreflex dysfunction: hypertension and chronic intermittent hypoxia. Exp. Physiol. 94 (9), 972–983. doi:10.1113/expphysiol.2009.048306
Keywords: chemoreflex, carotid body, baroreflex, baroreceptors, high altitude exposure, breathing, ventilation, arterial pressure
Citation: Alvarez-Araos P, Jiménez S, Salazar-Ardiles C, Núñez-Espinosa C, Paez V, Rodriguez-Fernandez M, Raberin A, Millet GP, Iturriaga R and Andrade DC (2024) Baroreflex and chemoreflex interaction in high-altitude exposure: possible role on exercise performance. Front. Physiol. 15:1422927. doi: 10.3389/fphys.2024.1422927
Received: 24 April 2024; Accepted: 15 May 2024;
Published: 04 June 2024.
Edited by:
Steffen Schulz, Charité University Medicine Berlin, GermanyReviewed by:
Nicholas Burgraff, Seattle Children’s Research Institute, United StatesCopyright © 2024 Alvarez-Araos, Jiménez, Salazar-Ardiles, Núñez-Espinosa, Paez, Rodriguez-Fernandez, Raberin, Millet, Iturriaga and Andrade. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: David C. Andrade, ZGF2aWQuYW5kcmFkZUB1YW50b2YuY2w=, ZGNhbmRyYWRlQHVjLmNs
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.