Introduction
Back-extrapolation (BE) has been demonstrated to be a suitable procedure for estimating the peak oxygen uptake (˙VO2peak) at the very end of exercise by applying the linear ˙VO2-time relationship to the primary response of the ˙VO2 recovery phase (i.e., fast ˙VO2 off-kinetics) (Léger et al., 1980; Rodríguez et al., 2017; Monteiro et al., 2020). In swimming, BE is a reliable procedure for estimating ˙VO2peak attained in an incremental exercise (Lavoie et al., 1981; Montpetit et al., 1981), and even BE affords a reliable estimate of ˙VO2peak during middle-distance swimming performances (i.e., 200 and 400 m), in which the attainment of the maximal rate of aerobic energy is recognized (Chaverri et al., 2016; Rodríguez et al., 2017). Therefore, the ˙VO2peak estimate from BE is supposed to provide the assessment of maximum ˙VO2 response from submaximal to supramaximal swimming circumstances (Monteiro et al., 2020), and thus BE is also considered a procedure enabling the overcome of contextual constraints imposed by the apparatus for the assessment of ˙VO2 response in the aquatic environment (Chaverri et al., 2016).
However, the linear ˙VO2-time model has been the source of controversial findings on the reliability of BE to estimate ˙VO2peak in swimming (Lavoie et al., 1985; Chaverri et al., 2016). For example, the overestimation of ˙VO2peak assessment of a post 400-m single-trial swimming performance (Lavoie et al., 1981) conflicts with the post incremental step-test values (Montpetit et al., 1981), despite both being swimming circumstances with a recognized maximum ˙VO2 demand (Zacca et al., 2019). Probably, this mismatch in comparing BE estimate vs. incremental test assessment of ˙VO2peak might account for the impairments on physiological response during high-intensity constant work-rate exercise, including either oxidative inertia or the anerobic energy relying on the onset of exercise since both these physiological mechanisms are supposed to modulate ˙VO2 off-kinetics acutely (i.e., slowing or speeding ˙VO2 exponential response post-exercise) (Özyener et al., 2001; Rossiter et al., 2002; Sousa et al., 2015). However, these physiological mechanisms are assumed to impair the attainment of ˙VO2peak during constant-phase exercise, if the reference value for comparison (usually assessed from an incremental exercise protocol) might be considered a reliable ˙VO2peak in swimming (Sousa et al., 2014; Pessôa Filho et al., 2017).
Despite the factors influencing BE reliability to estimate ˙VO2peak, previous reports suggested both the 200- and 400-m performances in swimming as typical middle-distance events, eliciting high aerobic energy release and, therefore, the attainment of ˙VO2peak response, in spite of the differences between each other regarding the aerobic/anerobic energetics balance (Pyne and Sharp, 2014; Almeida et al., 2020; Zacca et al., 2020). In addition, it has been demonstrated that velocities between 95 and 105% of ˙VO2peak in swimming also elicited the ˙VO2peak (Sousa et al., 2014) and showed a similar profile of ˙VO2 response when compared to 200- and 400-m performance (Sousa et al., 2011; Chaverri et al., 2016; Rodríguez et al., 2017). Therefore, the 200- and 400-m trials might be considered suitable for estimating ˙VO2peak by applying BE procedures post all-out performances in swimming (Rodríguez et al., 2017; Zacca et al., 2019).
From these studies, the main lessons are that the BE procedure might overestimate the ˙VO2peak according to the dataset fitting strategies, the exercise intensity during a trial performance (Rodríguez et al., 2017), and exercising conditions previous to the target trial estimating ˙VO2peak (Rodríguez et al., 2017; Zacca et al., 2019). In other words, the mechanisms that affect the reliability of the ˙VO2peak estimate by BE are likely related to the physiological response during exercise that also affects the ˙VO2 kinetic responses in the recovery phase. This is if other sources capable of impairing the accuracy of the BE estimate (e.g., temporal resolution of data sampling, treatment of the dataset, and mathematical curve fitting) are dis-regarded. (for further information on these other sources, see Monteiro et al., 2020; Rodríguez et al., 2017). Such a relationship was theoretically supposed to explain the modification of the constants of the linear function with the increase of the delay for the onset of ˙VO2 recovery, which was in turn, linked to the velocity of ˙VO2 adjustment during exercise (i.e., ˙VO2 on-kinetic) (Rodríguez et al., 2017).
In fact, experimental results have postulated that a high and rapid increase of ˙VO2 during exercise is related to a similar high and rapid reduction in the muscle phosphocreatine (PCr) content, the restoration of which inhibits the rapid decline of oxidative phosphorylation in the initial phase of recovery after exercise (i.e., slow time constant of ˙VO2 off-kinetic—τoff) (Rossiter et al., 2002; Korzeniewski and Zoladz, 2013). Indeed, this assumption might also support the overestimation of ˙VO2peak when applying BE procedures post 400 m rather than post 200 m (Rodríguez et al., 2017). Despite not ever being addressed, the τoff might play an important role for explaining how the reliability of BE to estimate ˙VO2peak is affected by performing exercises in different circumstances, leading to the attainment of the maximal aerobic rate.
Thus, the current study aimed to address the ˙VO2 recovery response and anerobic energy demand post different swimming circumstances in the severe-intensity domain to ascertain whether transients of ˙VO2 off-kinetics account for alterations of the linear adjustments of ˙VO2 response during the initial phase of ˙VO2 off-kinetics. Hence, the gathering of information to analyze the reliability of BE in estimating ˙VO2peak values with correspondence to the maximal ˙VO2 elicited whatever the swimming demand upon anerobic energetics during performances in the severe-intensity domain and correspondence to the maximum ˙VO2 response assessed in incremental exercise. In addition, this study explored whether a 200-m single-trial performance would be a feasible reference for the estimation of ˙VO2peak, adding information to support (or not) that the value estimated by BE is similar to either the ˙VO2peak assessed in an incremental test and/or the maximal ˙VO2 elicited at the end of the trial.
Methods
Subjects
Twenty swimmers (16.7 ± 2.4 years, 173.5 ± 10.2 cm, and 66.4 ± 10.6 kg; men = 12 and women = 8) were voluntarily recruited to participate in the study. The swimmers had at least three annual competitive training seasons and 200-m performances corresponding to 533 ± 83 and 502 ± 75 FINA points in a 25-m swimming pool, respectively, for men and women. The experimental procedures were performed in an indoor 25-m swimming pool, with a water temperature of ∼28°C. The swimmers were evaluated after familiarization with the procedures and devices. They were instructed to refrain from exhaustive training, alcohol, and caffeinated drinks the day before testing and to arrive well-fed and hydrated for the tests. All swimmers (and their legal guardians when they were under 18 years of age) signed a written consent form for their participation. This research was approved by the local ethics committee (CAEE: 54372516.3.0000.5398).
Performance tests and incremental intermittent step test (IIST)
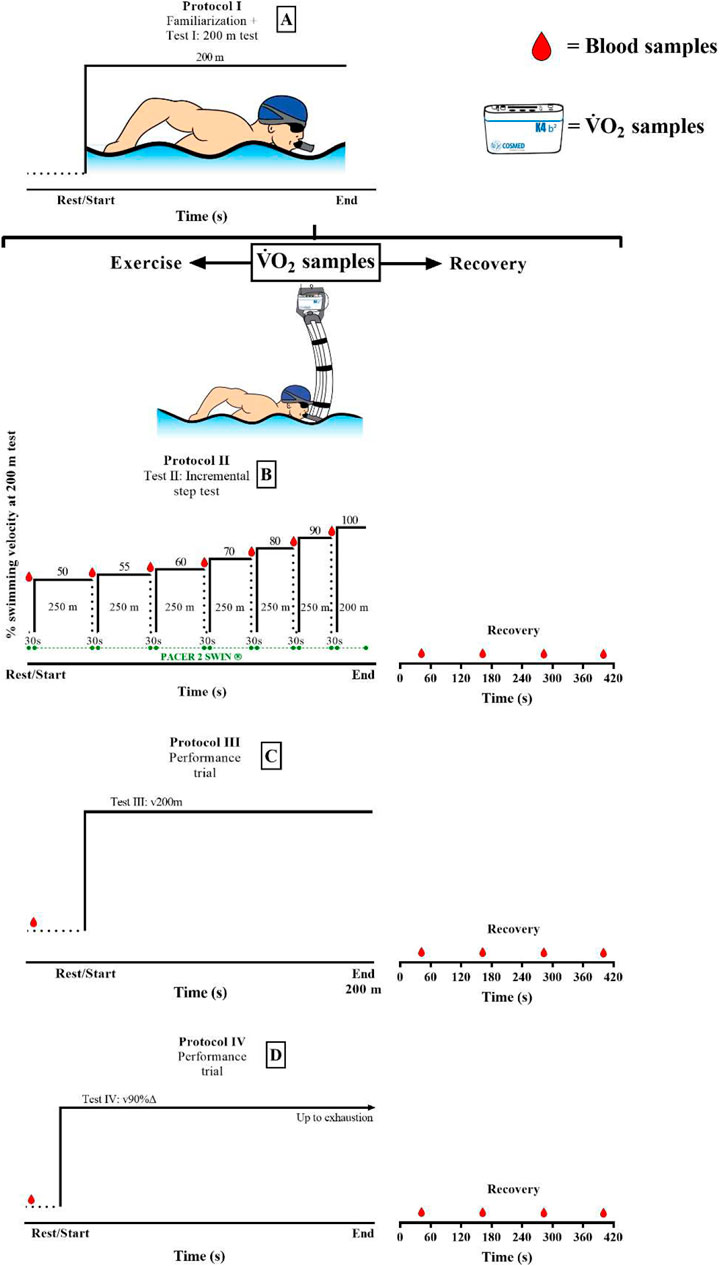
The familiarization phase with the snorkel system took place 24 h before testing procedures, which included all components of a regular training session, emphasizing middle-distance conditioning. All swimmers performed three swimming tests, with the duration between them being at least 48 h (Figure 1), with the second and third tests performed in a randomized order. The tests were 1) an incremental intermittent step-test (IIST) composed of six sets of 250 m in addition to one set of 200 m (IIST_v200m) at 50, 55, 60, 70, 80, 90, and 100% of velocity for 200 m, with 30 s between each step for blood sampling analysis (Almeida et al., 2021). The 200-m test was performed just after familiarization had been accomplished and 24 h before the IIST, following: 1) 1 h of rest from the previous exercise bout and 2) executed maximally with water starting, open turns, and no underwater gliding, as suggested by Massini et al. (2021); 2) a maximal 200-m single-trial performance (v200m); and 3) a transition from rest to the velocity corresponding to 90%Δ (v90%Δ, Eqn. (1)) performed until volitional exhaustion.
v90%∆=vLT+[(v˙VO2max−vLT)×0.9],[1] where vLT is the velocity corresponding to the lactate threshold (LT), defined as the first increase of blood lactate concentration ([la−]) above the resting levels, and determined from log–log bi-segmented plots of [la−] vs. velocity during the IIST (Faude et al., 2009). The swimming speed during all tests was controlled by visual information using an underwater visual pacer placed along the bottom of the pool (Pacer2Swim®, KulzerTEC, Portugal).
Measurements
Breath-by-breath gas exchange was sampled during and after the following experimental conditions: IIST, v200 m, and v90%Δ. For all conditions, the portable CPET unit (K4b2, Cosmed, Italy) was attached to the swimmer by a specific snorkel (new-AquaTrainer®, Cosmed, Italy), which was validated for gas analysis in swimming by Baldari et al. (2013). The CPET unit was calibrated before each test following the manufacturer’s recommendations. Blood samples (25 ul) were obtained from the swimmers’ earlobe at rest and at 1, 3, 5, and 7 min post-exercise, which were diluted in 75 ul 1% NaF solution. The samples were immediately analyzed for [la−] evaluation (YSI, 2300 STAT, Yellow Springs, United States).
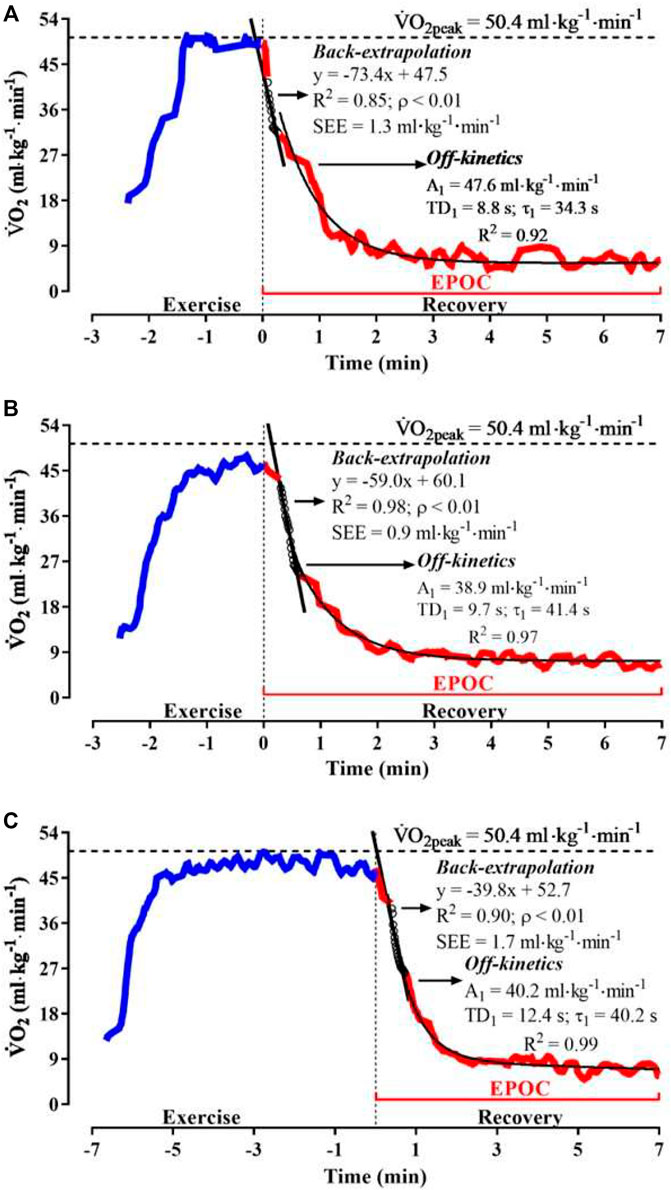
For assessment of ˙VO2peak and peak aerobic velocity (v˙VO2peak) during the IIST, the ˙VO2 data were smoothed (3-data point filter) and time-aligned to the discernibility of exercise and recovery phases. Moving average (30 s) processing was applied to the exercise ˙VO2 raw data, and the highest averaged value was considered the ˙VO2peak (Robergs et al., 2010; Reis et al., 2012). The velocity corresponding to the step of ˙VO2peak occurrence was defined as v˙VO2peak. For modeling of ˙VO2 off-kinetics, the 420-s rough ˙VO2 dataset from each transition at v200 m, v90%Δ, and IIST_v200m was time-aligned, and the noise was excluded and interpolated second-to-second for the analysis of ˙VO2 off-kinetics, as suggested by Özyener et al. (2001), Keir et al. (2014), and Benson et al. (2017). The mathematical modeling of ˙VO2 off-kinetics used a bi-exponential equation, with time delay (TD) (Eqn. (2)), according to the recommendations of Özyener et al. (2001) for the modeling of ˙VO2 off-kinetics in severe exercise:
˙VO2off(t)=EE˙VO2−A1off[1−e−(t−TD1off/τ1off)]−A2off[1−e−(t−TD2off/τ2off)][2] where EE˙VO2 corresponded to the final 30 s averaged ˙VO2 increase during exercise (in ml min−1). A1off and A2off are the net amplitude of ˙VO2 response for each phase of recovery (in ml∙min−1); t is exercise time; τ1off and τ2off are time constants (in seconds, s); and TD1off and TD2off are the time delays (in seconds, s) for ˙VO2 response for each phase of recovery (Özyener et al., 2001). The cardiopulmonary component was excluded by adjusting ˙VO2 response ∼15 s after the onset of exercise recovery (Özyener et al., 2001). The fast-O2debt (i.e., the amount of ˙VO2 response up to a particular time of the initial ˙VO2 recovery phase) was calculated from Eqn. (3), as recommended by Stirling et al. (2005):
Fast−O2debt=A1off·τ1off(1−e(tf−TD1off)τ1off)+A1off×(TD1off−tf)e(tf−TD1off)τ1off,[3] where tf is the time (s) at the end of the recovery sampling protocol. The blood lactate accumulation in equivalents of O2 (O2[la−], in ml∙min−1) was calculated following the recommendations of Prampero and Ferretti (1999) from O2[la−] = β·[la−]net, where β is equivalent to 2.7 ml kg−1 per 1 mmol L−1 of [la−]net, which is the algebraic difference between rest [la−] and peak [la−] post-exercise. The fast-O2debt (in ml·kg−1) and O2[la−] variables indicated the phosphagen and glycolytic components of total anerobic (AnaerTotal) response, respectively, during each swimming performance trial. The mean response time for the fast-O2debt curve was calculated (MRT1off = TD1off + τ1off, s) according to the previous studies in swimming (Almeida et al., 2020; Massini et al., 2021).
The BE method was applied to estimate the ˙VO2peak (BE−˙VO2peak, in ml min−1) and ˙VO2 recovery rate (BE-slope, in ml kg−1) from post-exercise ˙VO2 response (Montpetit et al., 1981) in IIST_v200m, v200 m, and v90%Δ. This procedure adjusted 20 s of the ˙VO2 vs. recovery time dataset by a linear function (f(y) = ax + b) (Léger et al., 1980), in which the delay of ˙VO2 recovery response (i.e., ∼15 s) was excluded before the linear adjustment of the dataset (see details on cardiopulmonary component exclusion for mathematical modeling of ˙VO2 off-kinetics) to the zero-recovery time.
Statistical analysis
The ˙VO2peak, EE˙VO2, and BE−˙VO2peak values (in ml·kg−1 min−1) for each trial were checked for normality with the Shapiro–Wilk test. The one-way ANOVA (Sidak as post hoc) compared ˙VO2peak to BE−˙VO2peak and EE˙VO2 in the IIST_v200m, v200m, and v90%Δ and the values of τ1off, TD1off, MRT1off, A1off, EE˙VO2, fast-O2debt, BE-Slope, and O2[la−] between each of the swimming performance conditions. The coefficient of dispersion (R2) and standard error of estimate (SEE) analyzed the variance between ˙VO2peak and BE−˙VO2peak. Eta squared (η2) was calculated to determine the effect size for ANOVA, considering the threshold values as <0.04 [trivial], 0.04–0.24 [small], 0.25–0.63 [medium], and >0.64 [large] (Fergusson, 2009).
Pearson’s coefficient (r) analyzed the correlation of ˙VO2 off-transients, fast-O2debt, and O2[la−] with EE˙VO2, BE−˙VO2peak, BE-slope, and ˙VO2 off-kinetic components under each swimming condition. The magnitudes of Pearson’s correlation were expressed as weak (0.00–0.29), low (0.30–0.49), moderate (0.50–0.69), strong (0.70–0.89), or very strong (0.90–1.00) (Mukaka, 2012); while R2 was considered <0.04 [trivial], 0.04–0.24 [small], 0.25–0.63 [medium], and >0.64 [strong] (Fergusson, 2009). For all analyses, the significance level was set at ρ ≤ 0.05. Sample power for the observed correlations was calculated considering the sample size (n = 20), correlation coefficient (r) Zα = 1.96 to a security index of α = 0.05, and expected sample power of 80% (β = 0.20). The statistical analysis was performed with SPSS Statistics for Windows (v18.0, IBM®, Chicago, IL, United States), and ˙VO2 data processing and modeling were both performed using OriginPro (OriginLab Corporation®, Northampton, MA, United States).
Results
The ˙VO2peak attained in the IIST was 55.7 ± 7.1 ml·kg−1·min−1, and v˙VO2peak corresponded to 1.26 ± 0.08 m × s−1. The v90%Δ and v200m were performed at 96.3 ± 4.4 and 101.1 ± 5.1% of v˙VO2peak, respectively Figure 2 illustrates the ˙VO2 response profile during exercise and recovery of IIST_v200m, v200 m, and v90%Δ for a male swimmer, which also exemplifies the “off-kinetics” and linear “back-extrapolation” modeling.
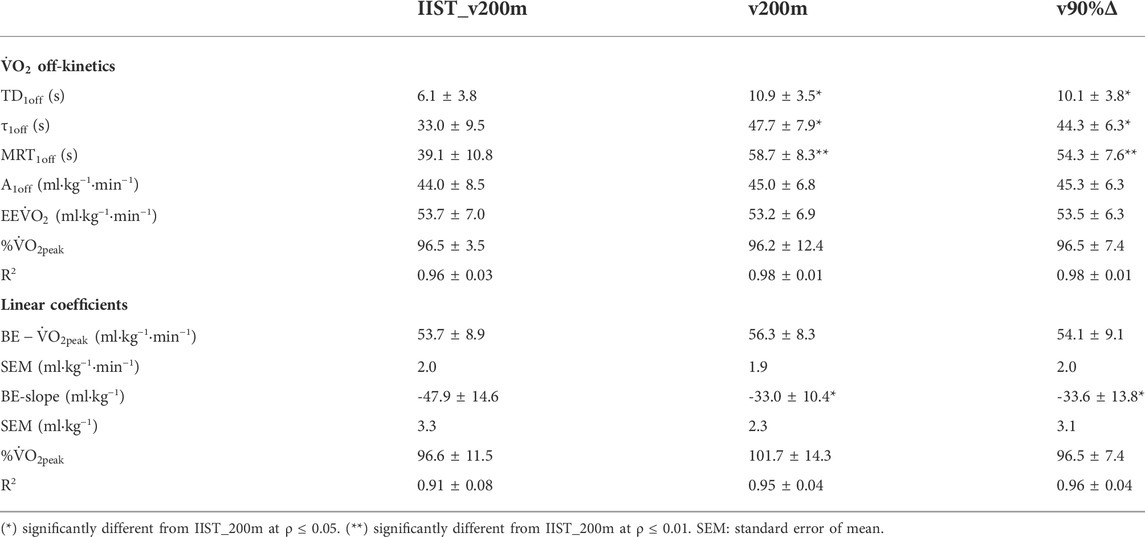
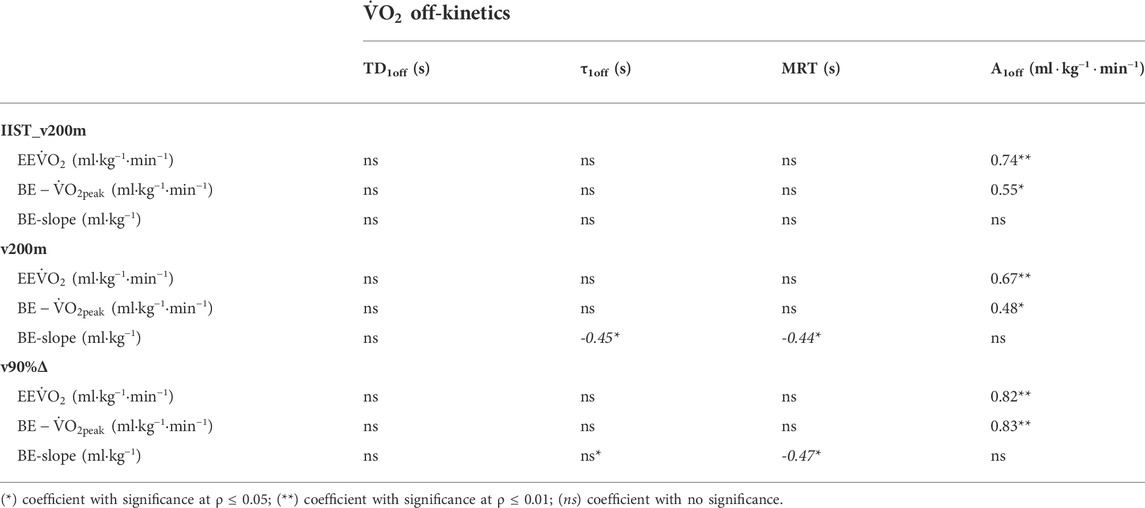
The variables of ˙VO2 off-kinetics and BE are shown in Table 1. Differences were observed for TD1off, τ1off, and MRT1off (p < 0.01, η2= 0.251, 0.397, and 0.479, all considered [medium] effect size), which were lower in IIST_v200m than in v200 m and v90%Δ, but not between v200 m and v90%Δ (ρ = 0.84, 0.45, and 0.35). No differences were observed for A1off (F[2,57] = 0.18, p = 0.83, η2= 0.006 [trivial]) and EE˙VO2 (F[2,57] = 0.04, p = 0.96, η2= 0.001 [trivial]) between trials.
In addition, BE−˙VO2peak values did not differ between trials (p = 0.62), despite BE-slope being higher (p < 0.01, η2= 0.227, considered [small] effect size) in the IIST_v200m than in the v200m and v90%Δ (p < 0.01 for both comparisons), but no difference was observed between v200m and v90%Δ (ρ = 1.00). The values of BE−˙VO2peak assessed for IIST_v200m, v200m, and v90%Δ (Table 1) were not different from those of ˙VO2peak (p = 0.73), neither were differences observed when comparing the EE˙VO2 during each trial for BE−˙VO2peak (p = 0.84) or ˙VO2peak (p = 0.65).
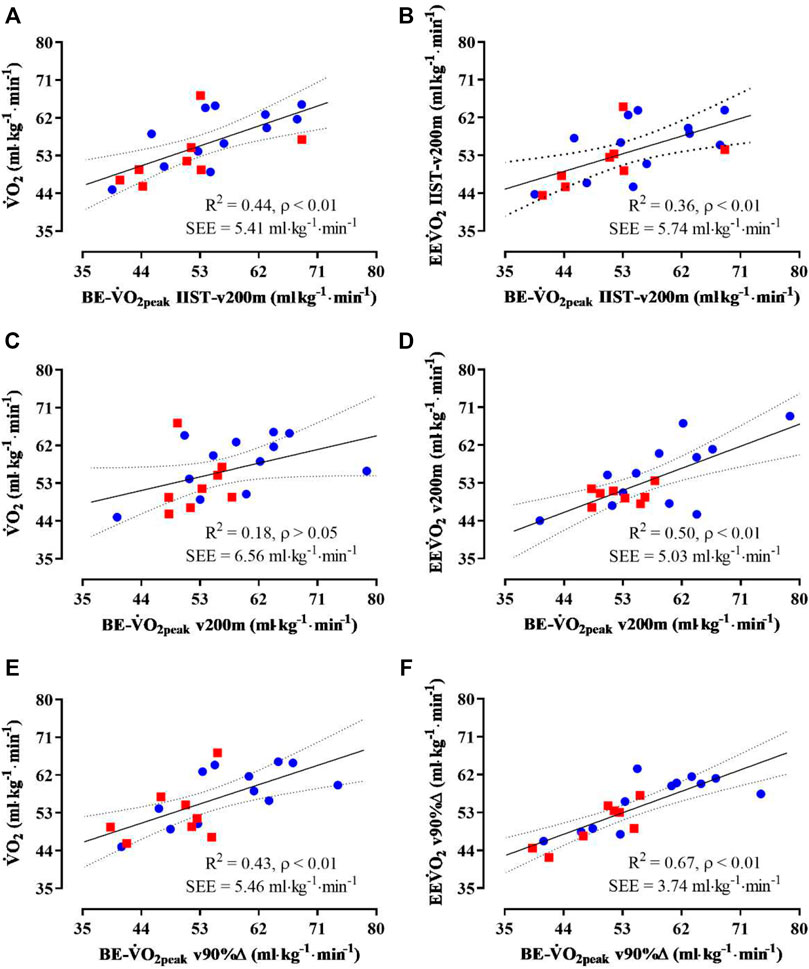
Small-to-medium R2 coefficients were observed between ˙VO2peak and BE−˙VO2peak for IIST_v200m, v200 m, and v90%Δ (Figure 3, panels A, C, and E, respectively), but a non-significant R2 coefficient was observed between ˙VO2peak and BE−˙VO2peak for v200 m. Also, the R2 coefficients were medium to strong between EE˙VO2 and BE−˙VO2peak for IIST_v200m, v200m, and v90%Δ (Figure 3, panels B, D, and F, respectively).
Pearson’s coefficients between parameters of both models (i.e., ˙VO2 off-kinetics vs. BE) attained satisfactory sample power and showed moderate-to-strong correlations between A1off with BE−˙VO2peak and BE-slope for the IIST_v200m and v90%Δ trials, whereas for the v200 m trial, these correlations ranged from low to moderate (Table 2).
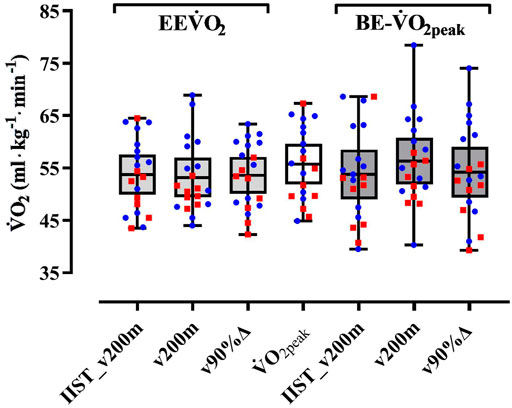
The τ1off correlated, exceptionally, to BE-slope for the v200 m trial, with low level and unsatisfactory sample power, and the MRT1off correlated to BE-slope for both v200 m and v90%Δ trials, but with low level and unsatisfactory sample power. The variability of EE˙VO2 (at IIST_v200m and v90%Δ) values is closer to that observed for ˙VO2peak values when compared to the variability observed for EE˙VO2 at v200 m and BE−˙VO2peak estimates in all trials, with the largest shown in v200 m (Figure 4).
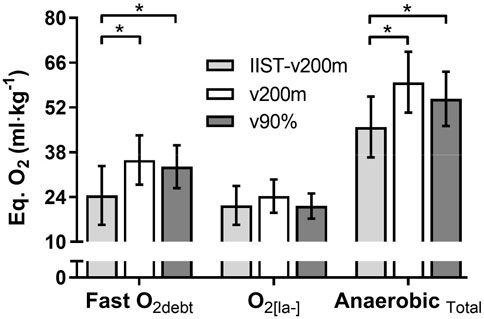
The fast-O2debt, O2[la−] and AnaerTotal demands assessed during the IIST_v200m, v200 m, and v90%Δ trials are shown in Figure 5. The fast-O2debt post IIST_v200m was lower (p < 0.01, η2= 0.281, considered [medium] effect size) than those post v200 m and v90%Δ. However, the values of O2[la−] were not different (p = 0.11) between IIST_v200m, v200 m, and v90%Δ. The AnaerTotal also was lower (p < 0.01, η2= 0.294, considered [medium] effect size) than those post v200m and v90%Δ. No correlations were observed between fast-O2debt and O2[la−] values with the responses of EE˙VO2, BE−˙VO2peak, and BE-slope for IIST_200m, v200 m, and v90%Δ, respectively. However, τ1off and MRT1off were moderately related to AnaerTotal post IIST_v200m (r = 0.64 and r = 0.66; p < 0.01), v200 m (r = 0.61 and r = 0.52; p < 0.01 and p = 0.02), and v90%Δ (r = 0.64 and r = 0.57; p < 0.01).
Discussion
The assumption that maximal ˙VO2 response (i.e., ˙VO2peak) can be elicited, and therefore assessed, during the trials was evidenced from the comparison between mean values of ˙VO2peak, EE˙VO2, and BE−˙VO2peak. In contrast, whether ˙VO2peak can be assessed with reliability by BE procedures applied under different recovery conditions in the severe-intensity domain requires further considerations. For example, the estimated BE−˙VO2peak showed low-to-moderate coefficients for the explained variance of the ˙VO2peak values assessed in the incremental test, with lowest coefficients observed for the 200-m single trial, which means that BE might mismatch actual ˙VO2peak between swimmers irrespective of the trial condition, but mainly in the 200-m trial. Also, when BE−˙VO2peak is estimating EE˙VO2, an improved coefficient of explanation is observed for single-trial conditions, which means that BE provides a satisfactory assessment of ˙VO2 elevation during swimming in the severe-intensity domain. Moreover, the transients of ˙VO2 off-kinetics played an important role on the reliability of BE−˙VO2peak estimate since delayed and slowed time courses of ˙VO2 recovery overshoot the BE values, which seemed to be a direct and positive effect of AnaerTotal release on the transients of ˙VO2 off-kinetics.
First, it is important to note that linear fitting underlying the BE mathematical procedure showed high adjustment coefficients for the 20 s dataset (with fixed TD = 15 s), irrespective of the trial performance in the severe-intensity exercise domain. Hence, the current finding indicating possible mismatching between ˙VO2peak and BE−˙VO2peak should not be addressed to the robustness (i.e., reduced regression power) of the linear procedure applied to the current estimates. The concerns when a fixed delay is considered in the initial phase of ˙VO2 recovery are related to the accuracy of the estimate. Commonly, studies have demonstrated that the accuracy of the BE model is increased when selecting 20 s of data (Chaverri et al., 2016; Rodíguez et al., 2017; Monteiro et al., 2020), applying a linear fit strategy, and considering a short delay (e.g., ∼5–10 s) before dataset fitting, which is, however, not a consensus for BE estimates in different exercise domains (Monteiro et al., 2020) and the exertion level or performance condition at a given exercise domain (Chaverri et al., 2016; Rodíguez et al., 2017). The current finding did not disagree with the aforementioned recommendations for the application of BE procedures but instead suggested that such an arbitrary delay of 15 s shall ensure that the ˙VO2 recovery post-swimming performance in severe-intensity domains has already been initiated, and, indeed, the linear fitting strategy on the 20-s dataset still presents high accuracy for the BE estimate.
Second, there is robust statistical evidence from the comparisons between mean values of ˙VO2peak and BE−˙VO2peak that these measurements are interchangeable, irrespective of the trial in which the BE−˙VO2peak was estimated. Similar evidence was also observed comparing mean values of EE˙VO2 and BE−˙VO2peak. However, dispersion plots of ˙VO2peak vs. BE−˙VO2peak refuted the interchangeable use between each other, showing that the power with which ˙VO2peak was estimated from BE−˙VO2peak in the post IIST_v200m, v90%Δ, and v200 m trials attained, respectively, moderate (44 and 43%) or low (18%) rates, with just the first two rates with satisfactory statistical confidence. Therefore, the BE−˙VO2peak post v200 m seems to be an unreliable assessment of ˙VO2peak, which might be attributed to the tendency (not significant) to overestimate actual values.
However, the ˙VO2 final response during all trials (i.e., EE˙VO2) attained maximal rates, and hence it did not account for the mismatching between ˙VO2peak vs. BE−˙VO2peak either post v200 m or post IIST_v200 m and v90%Δ. Indeed, the assumption that maximal ˙VO2 response is elicited during a 200-m single-trial performance has been well-reported (Almeida et al., 2020; Sousa et al., 2011; Rodríguez et al., 2017) and thus also contributing to recognize no constraints to the attainment of ˙VO2peak in 200 m. Furthermore, the current and previous reports on ˙VO2 response in 200 m also contribute to the typification of the severe-intensity domain in such distance and recognized for swimming conditions ranging from 95 to 105% of v˙VO2max (Sousa et al., 2014), or even for swimming velocity corresponding to 70%Δ (Reis et al., 2012), and just above the respiratory compensation point (Pessoa-Filho et al., 2012).
Third, whether there are no mathematical or physiological concerns about the reliability of BE procedures after all trials, why were the estimates considered poor (and unsatisfactory) for v200 m and moderate (but satisfactory) for IIST_v200m and v90%Δ? The effect of the energetics components during trial performances on the ˙VO2 initial recovery phase might provide new insights into the reliability of BE. Despite the lack of information regarding the effect of aerobic/anerobic energy release on ˙VO2 off-kinetics post-swimming performance in the severe-intensity domain since previous studies just analyzed the ˙VO2 recovery profile in response to exercises at or around maximal aerobic values (i.e., 100% or ranging from 95 to 105% ˙VO2peak, Sousa et al., 2014, 2015) or even at a given distance (i.e., 200 m; Sousa et al., 2011; Almeida et al., 2020), the current findings evidenced that total anerobic energy (i.e., phosphagenic in addition to glycolytic components) released during each trial showed a moderate and positive relationship with the transients τ1off and MRT1off. This means that the trials demanding higher anerobic release might also be associated to slower ˙VO2 recovery, as observed when comparing the slow responses post v90%Δ and v200 m with the fast post IIST_v200m.
In other sports than swimming, longer transients for ˙VO2 off-kinetics were associated with different intramuscular mechanisms such as 1) the rate of phosphocreatine resynthesis (i.e., a higher amount of phosphocreatine to restore requires a longer ˙VO2 decrement phase; Rossiter et al., 2002; Korzeniewski and Zoladz, 2013); 2) lactate clearance (i.e., parallel lactate oxidation and transportation slow the time course of ˙VO2 recovery; (Cunningham et al., 2000;; Özyener et al., 2001); and 3) the pattern of type II fiber recruitment (i.e., the inefficiency of oxidative phosphorylation also accounts to increase the time course of ˙VO2 recovery (Cunningham et al., 2000; Rossiter et al., 2002).
Particularly, in swimming, longer ˙VO2 time course during recovery has also been reported after the trial (200 m) and time-limited performance (Sousa et al., 2011, 2015), which was attributed to both the slower ˙VO2 response until maximal values and to the accumulation of fatigue-related metabolites while performing each swimming condition. Although the current study has no information on the time course of ˙VO2 on-kinetics response, which is therefore a limitation to be more assertive regarding the symmetry between on- and off-transients of ˙VO2 response, the current findings are best aligned with the statement that a longer ˙VO2 decrease is also probably linked to the anerobic reliance during swimming performance in the severe-intensity domain.
Moreover, the EE˙VO2 did not differ between IIST_v200m, v200m, and v90%Δ, and no differences were observed for A1off after each trial. In cycling, the similarity of ˙VO2 values and ˙VO2 on-kinetics between different performances in high-intensity exercise is consistent with the assumption that the attainment of a maximal oxidative response is not affected by the pattern of fast/slow fiber type recruitment, and its particular metabolic profile for each trial, i.e., cost of O2, rate of phosphate utilization, amplitude of slow component, and accumulation of metabolites (Cunninghan et al., 2000; Özyener et al., 2001; Rossiter et al., 2002). Therefore, there are also no physiological arguments to suppose that ˙VO2peak was not attained while performing v200m, IIST_v200m, and v90%Δ, even considering that differences were observed between them regarding total anerobic demand.
However, the aforementioned metabolic statement in cycling also inferred that longer transients of the initial ˙VO2 recovery phase are probably related to the reliance on type II fibers during the performance in the severe-intensity domain, as suggested by higher anerobic release and slow component occurrence, respectively, for higher-intensity short trials (i.e., fast fiber contribution is promptly established) and longer-term trials (i.e., fast fiber contribution is progressively established) (Cunninghan et al., 2000; Özyener et al., 2001; Rossiter et al., 2002). While the current finding on the positive correlation between A1off with BE−˙VO2peak and EE˙VO2 in all trials is aligned with the symmetry between the amplitude of ˙VO2 recovery and its values attained during exercise, the positive correlation in all trials between total anerobic energy and MRT (even if in the moderate level) is also consistent with the muscular bioenergetics (with high reliance on anaerobic energy) having influence on ˙VO2 recovery time course, which therefore accounted for the observation of MRT relationship to BE-slope only in v200 m and v90%Δ.
Finally, the findings suggested that the initial amplitude of ˙VO2 off-kinetics does not account for the possible mismatch between ˙VO2peak and BE−˙VO2peak, unless the attained value of EE˙VO2 is lower than that of ˙VO2peak (i.e., therefore the assumption of maximal ˙VO2 cannot be ensured). Moreover, the anerobic energy released contributes moderately to the longer transients of ˙VO2 off-kinetics, which suggests that the muscular metabolism is one among other variables with effect on BE−˙VO2peak reliability. However, the current results cannot address the reasons underpinning the better matching between ˙VO2peak and BE−˙VO2peak in v90%Δ than in v200 m. Although the aerobic contribution to each trial (i.e., total demand of ˙VO2) was not measured in the current study, it is expected to be higher in v90%Δ than in v200 m as supported when comparing previous reports on the energetics for swimming at velocities surrounding maximal aerobic velocity (Sousa et al., 2014) or at 200 m (Massini et al., 2021).
From the results of these previous studies, the reliance on oxidative metabolism during the performance of v90%Δ is supposed to be higher than that of v200 m, and thus the attainment of a given value of EE˙VO2 not different from EE˙VO2 not different from ˙VO2peak is expected for each swimmer and can be accounted to the low variability of BE−˙VO2peak estimate during v90%Δ. Therefore, the lack of information on aerobic contribution is another limitation of the current study, which should be overcome in future studies aiming to address whether the muscular energetics influence ˙VO2 on-kinetics when comparing distance-limited and time-limited performances in swimming. It can be argued that the poor matching between ˙VO2peak and BE−˙VO2peak in v200 m is a feature of the fixed delay (15 s) applied to the BE procedure. Despite the reliability of the ˙VO2peak estimate being susceptible to different time delays (Rodígues et al., 2017; Monteiro et al., 2020), the initial ˙VO2 recovery seems to differ from 15 s only for IIST_v200m, in which the BE−˙VO2peak estimate was not suspicious.
Although the scope of the current study was not the analysis of the effect of data treatment on the measurements of the transients and amplitudes of ˙VO2 kinetics and BE, an unstudied issue in swimming physiology is whether breathing mechanics (i.e., ventilatory frequency and volume) is disturbed with the AquaTrainer® apparatus by comparing to actual free-swimming condition (e.g., producing larger set of aberrant ˙VO2 data). It is important to investigate whether swimming has an intrinsic characteristic of ventilatory mechanics, which is different from other sports, hence requiring proper ˙VO2 dataset treatment.
When analyzing the practical applications of the current findings, three major comments are discernible: 1) BE is a feasible procedure for the assessment of ˙VO2 response at the end of exercise conditions in the severe-intensity domain (represented by IIST_v200m, v200m, and v90%Δ in the current study), which approached a maximal aerobic value despite the lack of endorsement on its interchangeability with ˙VO2peak; 2) such a maximal aerobic value is, however, meaningful for coaches as it represents the muscular oxidative profile in the severe-intensity domain, and hence enabling the management of aerobic response in middle-distance performance, the adjustments with cardiorespiratory conditioning during training demanding maximal aerobic responses, and the pace reference for training in the severe-intensity domain; and 3) the BE protocol with best reliability to assess the ˙VO2 response that matches ˙VO2peak should allow a proportionally higher reliance on aerobic than anerobic energy contribution, as is probably the case either during longer trials in the severe-intensity domain (e.g., 300–400 m) or shorter distances preceding a similar trial (e.g., 2 × 200 m).
Conclusion
The major contribution of the current study was to determine the effect of anerobic response on the reliability of the estimation of ˙VO2peak by BE, demonstrating that the anerobic demand might also be associated to longer transients of ˙VO2 off-kinetics (i.e., slowed ˙VO2 recovery), which in turn are associated to the alterations of the slope of the regression line (e.g., reducing the inclination), and therefore compromising the reliability of ˙VO2peak estimate, in spite of the strength of these associations observed to be low to moderate. Considering the fact that performance in a single effort with significant contribution of anerobic energy (as observed for v200m and v90%Δ) should probably demand a significant time constant or average response time of ˙VO2 recovery; a useful solution is to ensure faster responses of the transients of ˙VO2 off-kinetics, with the performance of an exercise with the same characteristics of effort intensity as the one where the test is intended to be carried out, as observed in the ISST_v200m situation. In addition, the findings also reinforce that the time delay for ˙VO2 recovery should be considered to apply BE procedures in trials in the severe-intensity domain, being recommendable to encompass a dataset no larger than 15 s. Finally, another important piece of evidence is the response of ˙VO2 at the end of IIST_v200m, v200 m, and v90%Δ corresponding to that typical of the severe-intensity domain, despite the estimation of ˙VO2peak by BE giving no confident value from the v200m test, and hence the estimates from IIST_v200m and v90%Δ are preferable for planning trials, controlling oxidative response, and monitoring the conditioning adjustment needed to perform in the severe-intensity domain.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The studies involving human participants were reviewed and approved by São Paulo State University Ethics Committee (CAEE: 54372516.3.0000.5398). Written informed consent to participate in this study was provided by the participants’ legal guardian/next of kin.
Author contributions
DM, AS, TA, JR, FB, ME, and DPF conceived and designed the study. DM, AS, TA, AM. ME, JR, and DPF conducted experiments and analyzed the data. DM, AS, TA, AM, ME, JR, FB, and DPF wrote the manuscript. All the authors read and approved the manuscript.
Funding
DPF would like to thank São Paulo Research Foundation - FAPESP (PROCESS 2016/04544-3) for the partial financial support and ME to the Foundation for Science and Technology, I.P. Grant/Award Number UIDB/04748/2020. This study was possible thanks to the scholarship granted from the Brazilian Federal Agency for Support and Evaluation of Graduate Education (CAPES), in the scope of the program CAPES-PrInt, process number 88887.310463/2018-00 and Mobility number 88887.580265/2020-00. AM also acknowledges the fellowship from CAPES (Finance Code 001).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Almeida T. A. F., Pessôa Filho D. M., Espada M. A. C., Reis J. F., Sancassani A., Massini D. A., et al. (2021). Physiological responses during high-intensity interval training in young swimmers. Front. Physiol. 12, 662029. doi:10.3389/fphys.2021.662029
PubMed Abstract | CrossRef Full Text | Google Scholar
Almeida T. A. F., Pessôa Filho D. M., Espada M. A. C., Reis J. F., Simionato A. R., Siqueira L. O. C., et al. (2020). VO2 kinetics and energy contribution in simulated maximal performance during short and middle distance-trials in swimming. Eur. J. Appl. Physiol. 120, 1097–1109. doi:10.1007/s00421-020-04348-y
PubMed Abstract | CrossRef Full Text | Google Scholar
Baldari C., Fernandes R., Meucci M., Ribeiro J., Vilas-Boas J. P., Guidetti L. (2013). Is the new AquaTrainer® snorkel valid for VO2 assessment in swimming? Int. J. Sports Med. 34 (4), 336–344. doi:10.1055/s-0032-1321804
PubMed Abstract | CrossRef Full Text | Google Scholar
Benson A. P., Bowen T. S., Ferguson C., Murgatroyd S. R., Rossiter H. B. (2017). Data collection, handling, and fitting strategies to optimize accuracy and precision of oxygen uptake kinetics estimation from breath-by-breath measurements. J. Appl. Physiol. 123 (1), 227–242. doi:10.1152/japplphysiol.00988.2016
PubMed Abstract | CrossRef Full Text | Google Scholar
Chaverri D., Schuller T., Iglesias X., Hoffmann U., Rodríguez F. (2016). A new model for estimating peak oxygen uptake based on post-exercise measurements in swimming. Int. J. Sports Physiol. Perform. 11, 419–424. doi:10.1123/ijspp.2015-0227
PubMed Abstract | CrossRef Full Text | Google Scholar
Cunningham D. A., St Croix C. M., Paterson D. H., Özyener F., Whipp B. J. (2000). The off-transient pulmonary oxygen uptake (VO2) kinetics following attainment of a particular VO2 during heavy-intensity exercise in humans. Exp. Physiol. 85 (3), 339–347. doi:10.1111/j.1469-445X.2000.01919.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Ferguson C. J. (2009). An effect size primer: A guide for clinicians and researchers. Prof. Psychol. Res. Pract. 40, 532–538. doi:10.1037/a0015808
CrossRef Full Text | Google Scholar
Keir D. A., Murias J. M., Paterson D. H., Kowalchuk J. M. (2014). Breath‐by‐breath pulmonary O2 uptake kinetics: Effect of data processing on confidence in estimating model parameters. Exp. Physiol. 99 (11), 1511–1522. doi:10.1113/expphysiol.2014.080812
PubMed Abstract | CrossRef Full Text | Google Scholar
Korzeniewski B., Zoladz J. A. (2013). Slow VO2 off-kinetics in skeletal muscle is associated with fast PCr off-kinetics—And inversely. J. Appl. Physiol. 115, 605–612. doi:10.1152/japplphysiol.00469.2013
PubMed Abstract | CrossRef Full Text | Google Scholar
Lavoie J. M., Léger L. A., Leone M., Provencher P. J. (1985). A maximal multistage swim test to determine the functional and maximal aerobic power of competitive swimmers. J. Swim. Res. 1, 17–22.
Google Scholar
Lavoie J. M., Taylor A. W., Montpetit R. R. (1981). Physiological effects of training in elite swimmers as measured by a free-swimming test. J. Sports Med. Phys. Fit. 21, 38–42.
PubMed Abstract | Google Scholar
Léger L. A., Seliger V., Brassard L. (1980). Backward extrapolation of VO2max values from the O2 recovery curve. Med. Sci. Sports Exerc 12 (1), 24–27. doi:10.1249/00005768-198021000-00006
PubMed Abstract | CrossRef Full Text | Google Scholar
Massini D. A., Almeida T. A. F., Vasconcelos C. M. T., Macedo A. G., Espada M. A. C., Reis J. F., et al. (2021). Are young swimmers short and middle distances energy cost sex-specific? Front. Physiol. 12, 796886. doi:10.3389/fphys.2021.796886
PubMed Abstract | CrossRef Full Text | Google Scholar
Monteiro A. S., Carvalho D. D., Azevedo R., Vilas-Boas J. P., Zacca R., Fernandes R. J. (2020). Post-swim oxygen consumption: Assessment methodologies and kinetics analysis. Physiol. Meas. 41 (10), 105005. doi:10.1088/1361-6579/abb143
PubMed Abstract | CrossRef Full Text | Google Scholar
Montpetit R. R., Léger L. A., Lavoie J. M., Cazorla G. (1981). VO2 peak during free swimming using the backward extrapolation of the O2 recovery curve. Eur. J. Appl. Physiol. Occup. Physiol. 47 (4), 385–391. doi:10.1007/BF02332966
PubMed Abstract | CrossRef Full Text | Google Scholar
Mukaka M. M. (2012). Statistics corner: A guide to appropriate use of correlation coefficient in medical research. Malawi Med. J. 24 (3), 69–71.
PubMed Abstract | Google Scholar
Özyener F., Rossiter H. B., Ward S. A., Whipp B. J. (2001). Influence of exercise intensity on the on- and off-transient kinetics of pulmonary oxygen uptake in humans. Jj. Physiol.Physiol. 533, 891–902. doi:10.1111/j.1469-7793.2001.t01-1-00891.x
CrossRef Full Text | Google Scholar
Pessôa Filho D. M., Siqueira L. O., Simionato A. R., Espada M. A., Pestana D. S., DiMenna F. J. (2017). A rapidly-incremented tethered-swimming test for defining domain-specific training zones. J. Hum. Kinet. 57 (1), 117–128. doi:10.1515/hukin-2017-0053
PubMed Abstract | CrossRef Full Text | Google Scholar
Pessôa-Filho D. M., Alves F. B., Reis J. F., Greco C. C., Denadai B. S. (2012). VO2 kinetics during heavy and severe exercise in swimming. Int. JInt. J. Sports Med. 33, 744–748. doi:10.1055/s-0031-1299753
PubMed Abstract | CrossRef Full Text | Google Scholar
Prampero di, Ferretti G. (1999). The energetics of anaerobic muscle metabolism: A reappraisal of older and recent concept. Respir. Physiol. 118, 103–115. doi:10.1016/s0034-5687(99)00083-3
PubMed Abstract | CrossRef Full Text | Google Scholar
Reis J. F., Alves F. B., Bruno P. M., Vleck V., Millet G. P. (2012). Oxygen uptake kinetics and middle-distance swimming performance. J. Sci. Med. Sport 15, 58–63. doi:10.1016/j.jsams.2011.05.012
PubMed Abstract | CrossRef Full Text | Google Scholar
Robergs R. A., Dwyer D., Astorino T. (2010). Validity of postexercise measurements to estimate peak VO2 in 200-m and 400-m maximal swims. Int. J. Sports Med. 40 (2), 95–111. doi:10.2165/11319670-000000000-00000
PubMed Abstract | CrossRef Full Text | Google Scholar
Rodríguez F. A., Chaverri D., Iglesias X., Schuller T., Hoffmann U. (2017). Validity of postexercise measurements to estimate peak VO2 in 200-m and 400-m maximal swims. Int. J. Sports Med. 38 (6), 426–438. doi:10.1055/s-0042-123707
PubMed Abstract | CrossRef Full Text | Google Scholar
Rossiter H. B., Ward S. A., Kowalchuk J. M., Howe F. A., Griffiths J. R., Whipp B. J. (2002). Dynamic asymmetry of phosphocreatine concentration and O2 uptake between the on- and off-transients of moderate- and high-intensity exercise in humans. J. Physiol. 541, 991–1002. doi:10.1113/jphysiol.2001.012910
PubMed Abstract | CrossRef Full Text | Google Scholar
Sousa A. C., Vilas-Boas J. P., FernandeS R. J. (2014). VO2 kinetics and metabolic contributions whilst swimming at 95, 100, and 105% of the velocity at VO2max. Biomed. Res. Int. 2014, 675363. doi:10.1155/2014/675363
PubMed Abstract | CrossRef Full Text | Google Scholar
Sousa A., Figueiredo P., Keskinen K. L., Rodríguez F. A., Machado L., Vilas-Boas J. P., et al. (2011). VO2 off transient kinetics in extreme intensity swimming. J. Sports Sci. Med. 10 (3), 546–552.
PubMed Abstract | Google Scholar
Sousa A., Figueiredo P., Pendergast D., Kjendlie P. L., Vilas-Boas J. P., Fernandes R. J. (2014). Critical evaluation of oxygen-uptake assessment in swimming. Int. J. Sports Physiol. Perform. 9 (2), 190–202. doi:10.1123/ijspp.2013-0374)
PubMed Abstract | CrossRef Full Text | Google Scholar
Sousa A., Rodríguez F. A., Machado L., Vilas‐Boas J. P., Fernandes R. J. (2015). Exercise modality effect on oxygen uptake off‐transient kinetics at maximal oxygen uptake intensity. Exp. Physiol. 100 (6), 719–729. doi:10.1113/EP085014
PubMed Abstract | CrossRef Full Text | Google Scholar
Stirling J. R., Zakynthinaki M. S., Saltin B. (2005). A model of oxygen uptake kinetics in response to exercise: Including a means of calculating oxygen demand/deficit/debt. Bull. Math. Biol. 67, 989–1015. doi:10.1016/j.bulm.2004.12.005
PubMed Abstract | CrossRef Full Text | Google Scholar
Zacca R., Azevedo R., Peterson Silveira R., Vilas-Boas J. P., Pyne D. B., Castro F. A. S., et al. (2019). Comparison of incremental intermittent and time trial testing in age-group swimmers. J. Strength Cond. Res. 33 (3), 801–810. doi:10.1519/JSC.0000000000002087
PubMed Abstract | CrossRef Full Text | Google Scholar
Zacca R., Azevedo R., Ramos V. R., Abraldes J. A., Vilas-Boas J. P., Castro F. A. S., et al. (2020). Biophysical follow-up of age-group swimmers during a traditional threepeak preparation program. J. Strength Cond. Res. 34 (9), 2585–2595. doi:10.1519/JSC.0000000000002964
PubMed Abstract | CrossRef Full Text | Google Scholar

 Danilo Alexandre Massini1,2
Danilo Alexandre Massini1,2