
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Pharmacol. , 19 March 2025
Sec. Neuropharmacology
Volume 15 - 2024 | https://doi.org/10.3389/fphar.2024.1505867
This article is part of the Research Topic Targeting Mitochondrial Dysfunction for the Discovery and Development of Novel CNS Therapies in Rare and Neurodegenerative Diseases View all 8 articles
 Delphine Ji1,2*
Delphine Ji1,2* Shanthini Mylvaganam1Prathyusha Ravi Chander1Mark Tarnopolsky3Keiran Murphy1Peter Carlen1,2,4*
Shanthini Mylvaganam1Prathyusha Ravi Chander1Mark Tarnopolsky3Keiran Murphy1Peter Carlen1,2,4*Epilepsy, affecting approximately 50 million individuals worldwide, is a neurological disorder characterized by recurrent seizures. Mitochondrial dysfunction and oxidative stress are critical factors in its pathophysiology, leading to neuronal hyperexcitability and cell death. Because of the multiple mitochondrial pathways that can be involved in epilepsy and mitochondrial dysfunction, it is optimal to treat epilepsy with multiple antioxidants in combination. Recent advancements highlight the potential of antioxidant therapy as a novel treatment strategy. This approach involves tailoring antioxidant interventions—such as melatonin, idebenone, and plant-derived compounds—based on individual mitochondrial health, including mitochondrial DNA mutations and haplogroups that influence oxidative stress susceptibility and treatment response. By combining antioxidants that target multiple pathways, reducing oxidative stress, modulating neurotransmitter systems, and attenuating neuroinflammation, synergistic effects can be achieved, enhancing therapeutic efficacy beyond that of a single antioxidant on its own. Future directions include conducting clinical trials to evaluate these combination therapies, and to translate preclinical successes into effective clinical interventions. Targeting oxidative stress and mitochondrial dysfunction through combination antioxidant therapy represents a promising adjunctive strategy to modify disease progression and improve outcomes for individuals living with epilepsy.
Epilepsy is a common and complex neurological disorder characterized by chronic, unprovoked seizures and affects approximately 50 million individuals worldwide (World Health, 2019). It is defined by the International League Against Epilepsy (ILAE) as transient occurrences of signs or symptoms due to abnormal excessive or synchronous neuronal activity in the brain (Scheffer et al., 2017). The hallmark characteristic is spontaneous recurrent seizures (SRS), corresponding with increased neuronal synchrony and excitability. The etiology ranges from genetic mutations, structural abnormalities, and, importantly, metabolic and/or mitochondrial dysfunction. Since the brain already has high aerobic metabolic demands and polyunsaturated fatty acids, it becomes particularly susceptible to insult and functional disturbances (Patel, 2002; Rho and Boison, 2022) (Supplementary Table S1; Table 1).
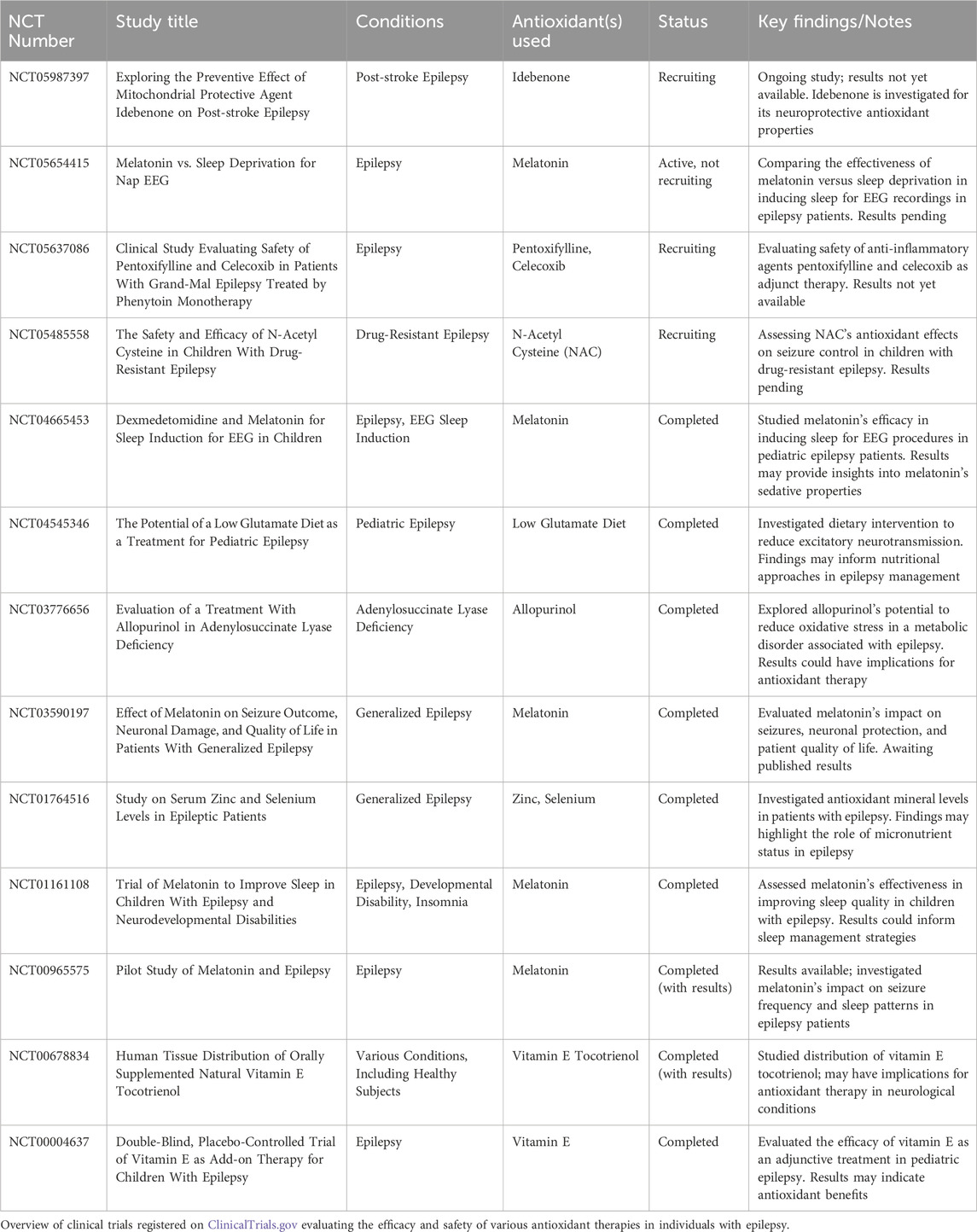
Table 1. Summary of clinical trials investigating antioxidant therapies in epilepsy.
Although the exact mechanisms of epileptogenesis have not been fully elucidated, there is increasing evidence for the involvement of mitochondria as crucial organelles in cellular energy production. In neurons, mitochondria are essential for maintaining membrane potential, calcium regulation (Bierhansl et al., 2024), and bioenergetics that support the high energetic demands for synaptic transmission. Dysfunction of mitochondrial processes, such as in ATP generation, calcium buffering, and the regulation of apoptosis, are increasingly recognized as contributors to the development and progression of epilepsy, particularly in mitochondrial epilepsies (Lopriore et al., 2022). In primary mitochondrial diseases such as MELAS or MERRF syndromes, defects in oxidative phosphorylation (OXPHOS) can lead to ATP depletion, impairing neuronal hyperpolarization and contributing to excessive excitatory activity. This disruption of energy homeostasis not only changes sodium-potassium pump (Na+/K+ ATPase) activity but also leads to the death of inhibitory interneurons, which are particularly vulnerable to OXPHOS deficiencies (Reinecke et al., 2009). This results in reduced GABAergic inhibition, increased glutamate release from astrocytes, and an overall hyperexcitable network, lowering the seizure threshold.
Oxidative stress plays a complex role in the pathophysiology of epilepsy, particularly in patients with mitochondrial dysfunction (Patel, 2002). Under normal physiological conditions, reactive oxygen species (ROS) and reactive nitrogen species (RNS) are produced as byproducts of cellular metabolism, especially in the mitochondrial respiratory chain, which accounts for many of the free radicals produced in the body (Juan et al., 2021; Kowalczyk et al., 2021). These species, including superoxide (O2−⋅), hydrogen peroxide (H2O2), and the hydroxyl radical (HO⋅), are involved in cellular signalling but can become harmful when produced in excess (Pizzino et al., 2017).
Specifically in mitochondria, superoxide radicals are generated through the reduction of molecular oxygen during electron transport (Turrens, 2003). While hydrogen peroxide is a by-product of this reaction and is not a free radical, it can be converted into hydroxyl radicals, some of the most damaging forms of ROS. The reaction between superoxide and nitric oxide (NO) forms peroxynitrite (ONOO−), a highly reactive molecule capable of initiating lipid peroxidation, protein nitration, and DNA damage (Castro and Freeman, 2001; Rowley et al., 2015). NADPH oxidase (Nox2) and cyclooxygenase-2 (COX-2) are additional enzymes that contribute to the oxidative stress burden in epilepsy (Rawat et al., 2019; Almeida et al., 2022). COX-2 is expressed in astrocytes, which are responsible for releasing proinflammatory cytokines, further exacerbating neuronal damage (Rawat et al., 2019). Oxidative stress in epilepsy results from an imbalance between pro-oxidant species and the potential of antioxidant defences that normally neutralize them. When this balance is disturbed, ROS and RNS can cause oxidative damage to DNA (Di Meo et al., 2016), proteins, and lipids, leading to cellular damage and apoptosis (Schieber and Chandel, 2014). Oxidative stress-induced damage has been implicated in numerous diseases, including epilepsy, atherosclerosis (Batty et al., 2022), diabetes complications (Volpe et al., 2018), and cancer (Glorieux et al., 2024). In the literature, the role of free radicals in causing malondialdehyde (MDA) elevation is particularly significant in epilepsy, as MDA is a marker of lipid peroxidation and higher levels have been observed in patients with recurrent seizures (Yilgor and Demir, 2024).
More specifically in epilepsy, oxidative stress can further impair mitochondrial function and set up a “vicious cycle”, leading to metabolic disturbances in neurons. It is generally understood that low levels of antioxidant enzymes such as superoxide dismutase (SOD) and catalase (CAT), combined with high levels of MDA, are indicative that the antioxidant defence system is overwhelmed by ROS production (Yilgor and Demir, 2024). Non-monogenic epilepsies are well-documented to be associated with mitochondrial dysfunction. There is evidence from both preclinical (Supplementary Table S1) and clinical studies (Table 1) that oxidative stress plays a key role in the initiation and progression of epilepsy. This is primarily founded on animal models using kainic acid or pilocarpine models of status epilepticus (Dal-Pizzol et al., 2000).
Mitochondrial disorders are frequently associated with epilepsy, and, conversely, seizures are associated with causing mitochondrial dysfunction with oxidative stress (Kunz, 2002; Patel, 2002). It has been hypothesized that impaired mitochondrial energy production could be the basis of pharmacoresistance in epilepsy (Yuen and Sander, 2011). The standard method of treating epilepsy patients consists of successive trials of many antiepileptic drugs, most of which raise the seizure threshold without addressing other aspects of their disorder, such as pervasive mitochondrial dysfunction. Status epilepticus (SE), sometimes fatal, is associated with oxidative stress, bioenergetic failure, and impaired mitochondrial dynamics in both mature and immature brains, all ameliorated by antioxidant treatments (Folbergrová and Kunz, 2012). Remarkably, antioxidant therapy is not used in SE, a treatment which could be quite safe and easily translatable. As argued below, it is becoming clear that the approach of using multiple rather than one or two antioxidants appears to be an improved approach for treating the many facets of mitochondrial dysfunction in epilepsy, as well as other neurological/neurodegenerative disorders. Neuroprotectants, particularly antioxidants, may have therapeutic value in epilepsy by reducing oxidative stress and its damaging effects on neurons. Increased levels of antioxidant enzymes such as SOD, CAT, and glutathione (GSH) have been proposed as potential therapeutic targets in refractory epilepsy (Cardenas-Rodriguez et al., 2013).
Predictive, preventive, and personalized medicine (PPPM/3PM) is a shift from the traditional “one-size-fits-all” approach in diseases that involve significant metabolic dysfunctions like mitochondrial diseases (MDs). Mitochondrial genetics significantly affect individual responses to therapies, including antioxidant treatments, which are increasingly being explored for their potential to counteract mitochondrial oxidative stress. One example includes mitochondria being a critical target in the context of hypoxic and ischemic injury, being central to therapeutic strategies aimed at improving outcomes in stroke patients (Ham III and Raju, 2017). Mitochondrial health and quality control are pivotal not only for assessing the risk of ischemic stroke but also for protecting neural tissue, supporting survival, and enhancing recovery outcomes on a personalized basis (Anzell et al., 2018; He et al., 2020). Furthermore, damage to the blood-brain barrier in the peri-infarct area, which often results in secondary injury, is strongly associated with limited recovery and significant disruptions in mitochondrial function (Nahirney et al., 2016). Another example is shown in tuberculosis (TB) and the body’s redox response to infection. The non-protein thiol glutathione (GSH) protects against Mycobacterium tuberculosis (MTB) infection. GSH, in conjunction with the transcription factor Nrf2 (nuclear factor erythroid 2-related factor 2), are crucial in counteracting the redox imbalance induced by MTB. Nrf2 mediates the expression of numerous antioxidant genes, and its antioxidant response element (ARE) signalling pathway is increasingly recognized as central to the pathogenesis of TB (Palanisamy et al., 2011). Personalized modulation of Nrf2-target genes highlights the potential of antioxidant therapies to enhance the efficacy of TB treatments (Petrillo et al., 2022).
Mitochondrial DNA (mtDNA) mutations have been increasingly associated with many neurological disorders, including epilepsy. While most genetic neurological conditions are linked to nuclear DNA mutations, defects in mtDNA significantly contribute to diseases such as Autism Spectrum Disorder (ASD) (Varga et al., 2018; Wang et al., 2022), Huntington’s disease (Ayala-Peña, 2013; Neueder et al., 2024), bipolar disorder (Munakata et al., 2004), and Leigh Syndrome (Ball et al., 1993). The mtDNA is a circular, double-stranded molecule comprising 16,659 nucleotides. It encodes 13 protein-coding genes essential for the electron transport chain (ETC.) complexes I, III, IV, and V, crucial for OXPHOS and ATP production (Anderson et al., 1981). Mutations in these genes can impair mitochondrial function, leading to decreased ATP production and increased ROS generation, contributing to neuronal hyperexcitability and seizures (Larsson et al., 1998). Specific mtDNA mutations are linked to mitochondrial encephalomyopathies associated with epilepsy, notably Mitochondrial Encephalopathy, Lactic Acidosis, Stroke-like episodes (MELAS) and Myoclonic Epilepsy with Ragged Red Fibers (MERRF) syndromes (Zeviani et al., 1993). In MELAS, the m.3243A > G mutation in the tRNA^Leu(UUR) gene affects mitochondrial protein synthesis, leading to defective OXPHOS and increased oxidative stress (Pia and Lui, 2024). MERRF is commonly associated with the m.8344A > G mutation in the tRNA^Lys gene, resulting in similar mitochondrial dysfunction (Hameed and Tadi, 2024). Variations in mtDNA can also influence mitochondrial biogenesis, apoptosis, calcium ion regulation, and other essential cellular processes (Osellame et al., 2012). Heteroplasmy, the coexistence of mutant and wild-type mtDNA within cells, contributes to variability in clinical presentations and disease severity (Stewart and Chinnery, 2015). In rat models, heteroplasmy has been shown to alter metabolic function, causing behavioural and cognitive deficits (Stewart and Chinnery, 2015).
Mitochondrial haplogroups, defined by specific mtDNA polymorphisms inherited maternally, may influence susceptibility to oxidative stress and response to antioxidant therapies (Gómez-Durán et al., 2010). Different haplogroups can affect mitochondrial efficiency and ROS production, potentially altering an individual’s vulnerability to mitochondrial dysfunction-related epilepsy (Amo et al., 2008). Certain haplogroups may be associated with higher baseline ROS production due to less efficient electron transport, increasing oxidative stress and seizure susceptibility; however, in cases of neurodegeneration, the potential as a risk factor remains controversial (Mancuso et al., 2008; Ingram et al., 2012). Understanding the role of mitochondrial haplogroups in epilepsy could aid in predicting disease risk and tailoring antioxidant therapies (Rea et al., 2013). Mitochondrial dysfunction resulting from genetic mutations leads to increased ROS production, contributing to neuronal damage and epileptogenesis (Soini et al., 2013). Antioxidant therapies have been explored to mitigate oxidative stress in epilepsy, but responses vary based on mitochondrial genetic background (Vergani et al., 2004). Studies have demonstrated that patients with mitochondrial disorders respond differently to antioxidants like Coenzyme Q10 (CoQ10) (Quinzii and Hirano, 2010). In cases of primary CoQ10 deficiency, resulting from mutations in nuclear genes involved in CoQ10 biosynthesis (e.g., PDSS2, COQ9, ADCK3), supplementation with CoQ10 has led to improvements in animal experiments, including reduced seizure duration (Sattarinezhad et al., 2014; Simani et al., 2020) and has been measured in various mitochondrial disorders in an RCT (Glover et al., 2010). In mitochondrial disorders caused by mtDNA mutations, such as MELAS, the efficacy of antioxidant therapies shows some evidence (Rodriguez et al., 2007). The variable responses may be due to differences in how specific mutations affect mitochondrial function and the resulting oxidative stress levels. Given the heterogeneity of mitochondrial genetic defects, personalized antioxidant therapies hold promise for improving treatment outcomes. By identifying specific mitochondrial mutations or haplogroups present in a patient, clinicians can tailor antioxidant strategies to target the underlying mitochondrial dysfunction more effectively (Meng et al., 2021). For instance, patients with mutations leading to deficiencies in, ETC complexes might benefit from antioxidants that support electron transport and reduce ROS production. Mitochondrial-targeted antioxidants, such as MitoQ or SkQ1, are designed to accumulate within mitochondria and directly neutralize ROS at the source (Murphy and Smith, 2007; Skulachev et al., 2023). These targeted therapies could enhance treatment efficacy and reduce potential toxicity associated with higher doses of non-specific antioxidants (Wang et al., 2011).
The potential for the use of multi-ingredient supplements to target the multiple final common pathways of neuronal dysfunction was first proposed in 2001 by Tarnopolsky and Beal (2001). Others have supported this contention, suggesting that synergism occurs when the combined effect of multiple antioxidants is greater than the sum of their individual effects (Thoo et al., 2013). This phenomenon can arise from various mechanisms, including antioxidant regeneration, differences in cellular localization, and complementary actions on oxidative pathways (Wang et al., 2011). For instance, one antioxidant may regenerate another by donating electrons to restore its active form, thereby extending its antioxidant activity. Several mechanisms contribute to this improved synergism such as redox cycling, antioxidant partitioning, and various formulations and combinations. In redox cycling, one antioxidant regenerates another by donating electrons, restoring its active form. For example, ascorbic acid (vitamin C) can regenerate α-tocopherol (vitamin E) by reducing the α-tocopheroxyl radical back to α-tocopherol (Niki, 1987). This process maintains antioxidant activity and prolongs protection against oxidative damage. Antioxidants with varying solubilities will also localize differently within biological systems, targeting oxidative stress in multiple compartments. Lipid-soluble antioxidants protect cell membranes, while water-soluble antioxidants defend the cytosol (Sharifi-Rad et al., 2020). Their distinct localization can enhance overall antioxidant efficacy. Together, antioxidants that scavenge free radicals and those that chelate pro-oxidant metal ions can more effectively reduce oxidative stress by addressing multiple pathways simultaneously (Halliwell, 1987). The concept of the use of multi-ingredient supplements for genetic mitochondrial disease was first studied in a randomized, double-blind study showing that the use of a multi-ingredient supplement that provided an alternative energy source (creatine monohydrate) + a membrane anti-oxidant (vitamin E) and two mitochondrial localized anti-oxidants/redox couple (COQ10 + alpha lipoic acid) lowered ROS markers and lactate (improved mitochondrial function) (Rodriguez et al., 2007; Tarnopolsky and Beal, 2001). Support for the superiority of the multi-ingredient supplement approach vs. a single agent targeting one pathway (ROS) was reflected in the fact that very high doses of CoQ10 (600 mg bid) neither lowered oxidative stress nor lactate in a similar cohort genetic mitochondrial disease patients (Glover et al., 2010).
For example, the combination of ascorbic acid and α-tocopherol has shown synergistic antioxidant effects in protecting phospholipid bilayers (Liebler et al., 1986). Ascorbic acid regenerates α-tocopherol from its radical form, sustaining membrane protection against lipid peroxidation. Flavonoids like quercetin and myricetin, which have lower redox potentials than α-tocopherol, can regenerate α-tocopherol and enhance its antioxidant activity (Marinova et al., 2008). Studies have reported synergistic interactions between α-tocopherol and flavonoids in inhibiting lipid oxidation (Bayram and Decker, 2023). Combining mitochondrial-targeted antioxidants with agents that modulate gene expression can restore mitochondrial function more effectively. Activation of Nrf2, a transcription factor that upregulates antioxidant defences, has shown promise in enhancing cellular resilience to oxidative stress (Li and Kong, 2009). Epigenetic therapies influence gene expression without altering the DNA sequence, affecting pathways involved in oxidative stress and mitochondrial health (Shaughnessy Daniel et al., 2014). By combining antioxidants with epigenetic modulators, it is possible to target genetic pathways that restore mitochondrial function and reduce seizure susceptibility. Nrf2 controls the expression of antioxidant enzymes and cytoprotective proteins by activating an ARE (Cardenas-Rodriguez et al., 2013). Activating Nrf2 enhances the cell’s endogenous antioxidant capacity. Compounds like RTA 408, an Nrf2 activator, have shown neuroprotective effects in preclinical models (Shekh-Ahmad et al., 2019). NOX enzymes are significant sources of ROS in the central nervous system. Inhibiting NOX reduces ROS generation during seizures. Agents like AEBSF, a NOX inhibitor, can decrease oxidative damage when combined with antioxidants (Shekh-Ahmad et al., 2019). This combination prevented seizure-induced mitochondrial depolarization, ROS generation, and neuronal cell death more effectively than either agent alone. In vivo, the combination therapy increased antioxidant capacity following kainic acid (KA)-induced SE, prevented the development of epilepsy, and reduced seizure frequency in established epilepsy models (Shekh-Ahmad et al., 2019).
The use of multiple antioxidants offers a promising strategy to address the complex pathophysiology of epilepsy, particularly in cases where personalized medicine is not feasible. Mitochondrial dysfunction plays a central role in epilepsy, even in monogenic forms of the disease, with downstream effects including excitotoxicity, calcium dysregulation, excessive reactive oxygen species (ROS) production, and neuroinflammation. Combining antioxidants that target diverse pathways associated with these dysfunctions can enhance therapeutic efficacy. Evidence from animal models and clinical studies highlights the potential of antioxidants such as vitamin E, melatonin, coenzyme Q10, and polyphenols to reduce seizure frequency and severity. However, when used as monotherapies, antioxidants may exhibit pro-oxidant effects under certain conditions, as demonstrated by Tarnopolsky (Tarnopolsky, 2008), making combination therapies a safer and more effective approach. Like “mitochondrial cocktails,” multi-antioxidant regimens provide broad-spectrum coverage, mitigating oxidative stress while modulating neuroinflammatory and neurotransmitter pathways. This strategy has shown promise in epilepsy-related dietary interventions like the ketogenic diet, which enhances mitochondrial health (Miller et al., 2020). Additionally, a multi-antioxidant approach can address comorbidities frequently associated with mitochondrial dysfunction, such as cognitive decline and mood disorders (Fattal et al., 2007). Further, by combining multiple antioxidants to target various regions in the oxidative and inflammatory cascades, antioxidant therapies can synergistically counteract the multifactorial nature of epilepsy pathogenesis, offering a robust alternative when personalized treatments are unattainable. This has been introduced as a preventative method from DNA injury in diagnostic radiation exposure (Merlin et al., 2022; Xhuti et al., 2023).
A substantial body of preclinical research has explored the therapeutic potential of antioxidant compounds in managing epilepsy. These studies have employed animal models to investigate how antioxidants can potentially mitigate seizure activity, prevent neuronal damage, and modulate oxidative stress and neuroinflammatory pathways associated with epileptogenesis.
Natural antioxidants derived from plants and other sources have been featured in many of these articles. For instance, royal jelly (RJ) demonstrated significant neuroprotective effects in kainic acid-induced TLE in rats by reducing seizure severity and oxidative stress markers, while enhancing total antioxidant capacity and preventing hippocampal neuronal damage (Hashemi et al., 2023). Proanthocyanidins (PACs) exhibited anticonvulsant effects in pentylenetetrazole (PTZ)-induced epilepsy in mice through activation of the Nrf2 pathway, leading to decreased oxidative stress, inflammation, and neuronal apoptosis (Alyami et al., 2022). Sulforaphane (SFN), another Nrf2 activator, reduced ROS production, restored glutathione levels, and attenuated neuronal death in kainic acid-induced SE in rats (Sandouka and Shekh-Ahmad, 2021). Other plant-derived compounds, such as curcumin derivatives, lycopene, and extracts from Melissa officinalis, Echinops spinosus, and Syzygium cumini, have also shown significant anticonvulsant and neuroprotective effects. These effects are primarily mediated through antioxidant mechanisms, modulation of neurotransmitter systems (e.g., GABA), and attenuation of neuroinflammation (Mahmoudi et al., 2020; Taskiran and Tastemur, 2021; Abd Allah et al., 2022; Kandeda et al., 2022; Alkhudhayri et al., 2023). Synthetic antioxidants and pharmaceuticals have been evaluated for their efficacy in epilepsy models as well. Tempol, a membrane-permeable radical scavenger, could attenuate PTZ-induced seizures in mice by reducing oxidative and nitrosative stress, enhancing GABAergic neurotransmission, and inhibiting pro-inflammatory cytokines (Zhang et al., 2018). Lacosamide, an antiepileptic drug (AED), not only decreased seizure activity in pilocarpine-induced SE in rats but also exerted antioxidant effects by restoring superoxide dismutase (SOD) activity and glutathione (GSH) levels (Shishmanova-Doseva et al., 2021). Mitochondrial dysfunction has emerged as a critical factor in epileptogenesis. Succinate accumulation contributed to increased oxidative stress and mitochondrial ROS levels, leading to neuronal degeneration and SE in kainic acid-induced models. Inhibiting succinate dehydrogenase (SDH) and related metabolic pathways reduced seizure severity and oxidative damage (Zhang et al., 2020). Interventions targeting mitochondrial bioenergetics, such as treatment with ascorbic acid, alpha-tocopherol, and sodium pyruvate (AATP), improved mitochondrial function, reduced seizure burden, and enhanced synaptic activity in temporal lobe epilepsy models (Simeone et al., 2014). Combination antioxidant therapies targeting multiple pathways have shown promise in providing enhanced neuroprotection. Preservation of ion channel function and enzyme activities has also been a focus. Agents like lipoic acid (LA) and idebenone prevented seizures and restored the activities of critical enzymes such as Na⁺/K⁺-ATPase and δ-aminolevulinic acid dehydratase (δ-ALA-D), which are essential for maintaining neuronal excitability and metabolic homeostasis (de Sales Santos et al., 2010; Ahmed, 2014).
Despite the promising results from preclinical studies demonstrating the neuroprotective and anticonvulsant effects of antioxidant therapies in epilepsy, several challenges hinder the translation of these findings into clinical practice. One significant challenge lies in the selective uptake limitations of mitochondria-targeted antioxidants. Damaged mitochondria, which typically exhibit lower membrane potential, may uptake these antioxidants less efficiently than their healthy counterparts, thereby reducing the efficacy of treatments aimed at mitigating oxidative stress within the very mitochondria that require intervention (Plotnikov and Zorov, 2019). Additionally, there is a risk of reductive stress, where excessive antioxidant supplementation disrupts the delicate balance of reactive oxygen species (ROS) necessary for normal cellular signaling and physiological functions. Over-suppression of ROS can impair essential processes such as cell differentiation, apoptosis, and immune responses, potentially leading to adverse cellular outcomes. There is also a potential for prooxidant activity under certain conditions, such as high concentrations or the presence of transition metals, which can paradoxically exacerbate oxidative stress rather than mitigate it (Podmore et al., 1998).
Determining the optimal dosage and administration regimen is complex, as factors such as bioavailability, pharmacokinetics, and individual patient variability influence therapeutic outcomes. Additionally, the lack of standardized methods for evaluating the efficacy and safety of these antioxidants poses significant regulatory challenges, making it difficult to establish universally accepted guidelines for their use. Addressing these disadvantages requires a multifaceted approach. Future research should focus on conducting clinical trials to evaluate the efficacy and safety of antioxidant compounds in patients with epilepsy. Personalized medicine holds great potential in optimizing antioxidant therapies, considering the variability in mitochondrial genetics among individuals. Exploring the role of mitochondrial DNA mutations and haplogroups in influencing the response to antioxidant treatments could enable the tailoring of therapies to individual patient profiles. Identifying oxidative stress and mitochondrial dysfunction biomarkers may further aid in customizing antioxidant interventions, enhancing therapeutic outcomes.
Moreover, combining antioxidants with anti-inflammatory agents or traditional antiepileptic drugs may provide synergistic effects, as suggested by preclinical studies (Pauletti et al., 2019; Shekh-Ahmad et al., 2019). The development of novel mitochondria-targeted antioxidants, such as MitoQ and SkQ1, offers the potential for a more effective reduction of oxidative stress at its primary source within neurons (Snow et al., 2010). Additional research is needed to understand the precise mechanisms by which antioxidants exert their anticonvulsant effects. Investigations into the role of the Nrf2 pathway, mitochondrial bioenergetics, and ion channel modulation in the context of antioxidant treatment could provide deeper insights (Waldbaum and Patel, 2010). Furthermore, the advantage of a multi-ingredient approach is notable, as it is likely to address a broader range of disorders compared to single-agent therapies. Given that mitochondrial genetic disorders, epilepsy, and most other neurological disorders converge on common pathways such as mitochondrial dysfunction, excitotoxicity, apoptosis, calcium dysregulation, ROS excess, and inflammation (Madireddy and Madireddy, 2023).
Oxidative stress and mitochondrial dysfunction play critical roles in the pathophysiology of epilepsy, contributing to neuronal hyperexcitability and cell death. Preclinical studies have provided substantial evidence that antioxidant therapies can mitigate these pathological processes, reduce seizure activity, and protect neuronal integrity. Compounds such as melatonin, sulforaphane, and various plant extracts have demonstrated significant anticonvulsant and neuroprotective effects in animal models. While clinical trials investigating antioxidant therapies in epilepsy are limited, preliminary findings suggest potential benefits. However, more extensive clinical research is necessary to confirm these effects and to establish optimal dosing regimens, safety profiles, and patient selection criteria. Considering individual genetic and metabolic differences may enhance the efficacy of antioxidant treatments. Antioxidant therapies represent a promising adjunctive strategy in the management of epilepsy. By targeting oxidative stress and mitochondrial dysfunction, these agents have the potential to modify disease progression and improve patient outcomes. Continued research efforts are essential to translate preclinical successes into effective clinical interventions for individuals living with epilepsy.
DJ: Writing–original draft, Writing–review and editing. SM: Writing–review and editing. PR: Writing–review and editing. MT: Conceptualization, Writing–review and editing. KM: Writing–review and editing. PC: Writing–original draft, Writing–review and editing.
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphar.2024.1505867/full#supplementary-material
Abd Allah, H. N., Abdul-Hamid, M., Mahmoud, A. M., and Abdel-Reheim, E. S. (2022). Melissa officinalis L. ameliorates oxidative stress and inflammation and upregulates Nrf2/HO-1 signaling in the hippocampus of pilocarpine-induced rats. Environ. Sci. Pollut. Res. 29, 2214–2226. doi:10.1007/s11356-021-15825-y
Ahmed, M. a.E. (2014). Neuroprotective effects of idebenone against pilocarpine-induced seizures: modulation of antioxidant status, DNA damage and Na+, K+-ATPase activity in rat Hippocampus. Neurochem. Res. 39, 394–402. doi:10.1007/s11064-014-1236-z
Alkhudhayri, A., Abdel Moneim, A. E., Rizk, S., Bauomy, A. A., and Dkhil, M. A. (2023). The neuroprotective effect associated with Echinops spinosus in an acute seizure model induced by pentylenetetrazole. Neurochem. Res. 48, 273–283. doi:10.1007/s11064-022-03738-2
Almeida, C., Pongilio, R. P., Móvio, M. I., Higa, G. S. V., Resende, R. R., Jiang, J., et al. (2022). Distinct cell-specific roles of NOX2 and MyD88 in epileptogenesis. Front. Cell Dev. Biol. 10, 926776. doi:10.3389/fcell.2022.926776
Alshabi, A. M., Shaikh, I. A., and Asdaq, S. M. B. (2022). The antiepileptic potential of Vateria indica Linn in experimental animal models: effect on brain GABA levels and molecular mechanisms. Saudi J. Biol. Sci. 29, 3600–3609. doi:10.1016/j.sjbs.2022.02.059
Alyami, N. M., Abdi, S., Alyami, H. M., and Almeer, R. (2022). Proanthocyanidins alleviate pentylenetetrazole-induced epileptic seizures in mice via the antioxidant activity. Neurochem. Res. 47, 3012–3023. doi:10.1007/s11064-022-03647-4
Amin, F., Tabassum, S., Sarwar, S., Qureshi, R., Sohaib Khalid, M., Riaz, N., et al. (2022). Neuroprotective effect of otostegia limbata against PTZ-induced mice model of epilepsy by attenuated expression of p-nfκb and TNF-α. Front. Neurosci. 16 16. doi:10.3389/fnins.2022.779681
Amini, S., Hooshmandi, Z., Setorki, M., and Teymourivand, Y. (2020). Protective effects of Biarum carduchrum ethyl acetate extract on seizure severity, depression, memory, and learning in pentylenetetrazole-induced kindling rats. IranRedCrescentMedJ. 22 (8), e76. doi:10.32592/ircmj.2020.22.8.76
Amo, T., Yadava, N., Oh, R., Nicholls, D. G., and Brand, M. D. (2008). Experimental assessment of bioenergetic differences caused by the common European mitochondrial DNA haplogroups H and T. Gene 411, 69–76. doi:10.1016/j.gene.2008.01.007
Anderson, S., Bankier, A. T., Barrell, B. G., De Bruijn, M. H., Coulson, A. R., Drouin, J., et al. (1981). Sequence and organization of the human mitochondrial genome. Nature 290, 457–465. doi:10.1038/290457a0
Anzell, A. R., Maizy, R., Przyklenk, K., and Sanderson, T. H. (2018). Mitochondrial quality control and disease: insights into ischemia-reperfusion injury. Mol. Neurobiol. 55, 2547–2564. doi:10.1007/s12035-017-0503-9
Ata Yaseen Abdulqader, Y., Abdel Kawy, H. S., Mohammed Alkreathy, H., and Abdullah Rajeh, N. (2021). The potential antiepileptic activity of astaxanthin in epileptic rats treated with valproic acid. Saudi Pharm. J. 29, 418–426. doi:10.1016/j.jsps.2021.04.002
Ayala-Peña, S. (2013). Role of oxidative DNA damage in mitochondrial dysfunction and Huntington’s disease pathogenesis. Free Radic. Biol. Med. 62, 102–110. doi:10.1016/j.freeradbiomed.2013.04.017
Azizi, V., Allahyari, F., and Hosseini, A. (2018). The antiepileptic and neuroprotective effect of the Buxus hyrcana Pojark hydroethanolic extract against the pentylentetrazol induced model of the seizures in the male rats. Int. J. Neurosci. 128, 913–919. doi:10.1080/00207454.2018.1443925
Ball, M., Thorburn, D. R., and Rahman, S. (1993). “Mitochondrial DNA-associated Leigh syndrome spectrum,” in GeneReviews(®). Editors M. P. Adam, J. Feldman, G. M. Mirzaa, R. A. Pagon, S. E. Wallace, L. J. H. Beanet al. (Seattle, WA: University of Washington). GeneReviews is a registered trademark of the University of Washington, Seattle.
Batty, M., Bennett, M. R., and Yu, E. (2022). The role of oxidative stress in atherosclerosis. Cells 11, 3843. doi:10.3390/cells11233843
Bayram, I., and Decker, E. A. (2023). Underlying mechanisms of synergistic antioxidant interactions during lipid oxidation. Trends Food Sci. and Technol. 133, 219–230. doi:10.1016/j.tifs.2023.02.003
Bierhansl, L., Gola, L., Narayanan, V., Dik, A., Meuth, S. G., Wiendl, H., et al. (2024). Neuronal mitochondrial calcium uniporter (MCU) deficiency is neuroprotective in hyperexcitability by modulation of metabolic pathways and ROS balance. Mol. Neurobiol. 61, 9529–9538. doi:10.1007/s12035-024-04148-x
Bortolatto, C. F., Jesse, C. R., Wilhelm, E. A., Ribeiro, L. R., Rambo, L. M., Royes, L. F. F., et al. (2011). Protective effect of 2,2′-dithienyl diselenide on kainic acid-induced neurotoxicity in rat hippocampus. Neuroscience 193, 300–309. doi:10.1016/j.neuroscience.2011.07.038
Cao, J., Tang, C., Gao, M., Rui, Y., Zhang, J., Wang, L., et al. (2020). Hyperoside alleviates epilepsy-induced neuronal damage by enhancing antioxidant levels and reducing autophagy. J. Ethnopharmacol. 257, 112884. doi:10.1016/j.jep.2020.112884
Cardenas-Rodriguez, N., Huerta-Gertrudis, B., Rivera-Espinosa, L., Montesinos-Correa, H., Bandala, C., Carmona-Aparicio, L., et al. (2013). Role of oxidative stress in refractory epilepsy: evidence in patients and experimental models. Int. J. Mol. Sci. 14, 1455–1476. doi:10.3390/ijms14011455
Castro, L., and Freeman, B. A. (2001). Reactive oxygen species in human health and disease. Nutrition 17, 163–165. doi:10.1016/s0899-9007(00)00570-0
Dal-Pizzol, F., Klamt, F., Vianna, M. M. R., Schröder, N., Quevedo, J., Benfato, M. S., et al. (2000). Lipid peroxidation in hippocampus early and late after status epilepticus induced by pilocarpine or kainic acid in Wistar rats. Neurosci. Lett. 291, 179–182. doi:10.1016/s0304-3940(00)01409-9
Della-Pace, I. D., Rambo, L. M., Ribeiro, L. R., Saraiva, A. L. L., De Oliveira, S. M., Silva, C. R., et al. (2013). Triterpene 3β, 6β, 16β trihidroxilup-20(29)-ene protects against excitability and oxidative damage induced by pentylenetetrazol: the role of Na(+),K(+)-ATPase activity. Neuropharmacology 67, 455–464. doi:10.1016/j.neuropharm.2012.10.022
de Sales Santos, Í. M., Da Rocha Tomé, A., Feitosa, C. M., De Souza, G. F., Feng, D., De Freitas, R. M., et al. (2010). Lipoic acid blocks seizures induced by pilocarpine via increases in δ-aminolevulinic dehydratase and Na+, K+-ATPase activity in rat brain. Pharmacol. Biochem. Behav. 95, 88–91. doi:10.1016/j.pbb.2009.12.011
Di Meo, S., Reed, T. T., Venditti, P., and Victor, V. M. (2016). Role of ROS and RNS sources in physiological and pathological conditions. Oxidative Med. Cell. Longev. 2016, 1245049. doi:10.1155/2016/1245049
El Nashar, E. M., Obydah, W., Alghamdi, M. A., Saad, S., Yehia, A., Maryoud, A., et al. (2022). Effects of Stevia rebaudiana Bertoni extracts in the rat model of epilepsy induced by pentylenetetrazol: sirt-1, at the crossroads between inflammation and apoptosis. J. Integr. Neurosci. 21, 21. doi:10.31083/j.jin2101021
Essawy, A. E., El-Sayed, S. A., Tousson, E., Abd El-Gawad, H. S., Alhasani, R. H., and Abd Elkader, H.-T. a.E. (2022). Anti-kindling effect of Ginkgo biloba leaf extract and L-carnitine in the pentylenetetrazol model of epilepsy. Environ. Sci. Pollut. Res. 29, 48573–48587. doi:10.1007/s11356-022-19251-6
Fattal, O., Link, J., Quinn, K., Cohen, B. H., and Franco, K. (2007). Psychiatric comorbidity in 36 adults with mitochondrial cytopathies. CNS Spectrums 12, 429–438. doi:10.1017/s1092852900015303
Fighera, M. R., Royes, L. F., Furian, A. F., Oliveira, M. S., Fiorenza, N. G., Frussa-Filho, R., et al. (2006). GM1 ganglioside prevents seizures, Na+,K+-ATPase activity inhibition and oxidative stress induced by glutaric acid and pentylenetetrazole. Neurobiol. Dis. 22, 611–623. doi:10.1016/j.nbd.2006.01.002
Folbergrová, J., and Kunz, W. S. (2012). Mitochondrial dysfunction in epilepsy. Mitochondrion 12, 35–40. doi:10.1016/j.mito.2011.04.004
Fuchs, M., Viel, C., Lehto, A., Lau, H., and Klein, J. (2023). Oxidative stress in rat brain during experimental status epilepticus: effect of antioxidants. Front. Pharmacol. 14, 1233184. doi:10.3389/fphar.2023.1233184
Glorieux, C., Liu, S., Trachootham, D., and Huang, P. (2024). Targeting ROS in cancer: rationale and strategies. Nat. Rev. Drug Discov. 23, 583–606. doi:10.1038/s41573-024-00979-4
Glover, E. I., Martin, J., Maher, A., Thornhill, R. E., Moran, G. R., and Tarnopolsky, M. A. (2010). A randomized trial of coenzyme Q10 in mitochondrial disorders. Muscle Nerve 42, 739–748. doi:10.1002/mus.21758
Gómez-Durán, A., Pacheu-Grau, D., López-Gallardo, E., Díez-Sánchez, C., Montoya, J., López-Pérez, M. J., et al. (2010). Unmasking the causes of multifactorial disorders: OXPHOS differences between mitochondrial haplogroups. Hum. Mol. Genet. 19, 3343–3353. doi:10.1093/hmg/ddq246
Halliwell, B. (1987). Free radicals and metal ions in health and disease. Proc. Nutr. Soc. 46, 13–26. doi:10.1079/pns19870004
Hameed, S., and Tadi, P. (2024). Myoclonic epilepsy and ragged red Fibers. Treasure Island, FL: StatPearls Publishing.
Ham Iii, P. B., and Raju, R. (2017). Mitochondrial function in hypoxic ischemic injury and influence of aging. Prog. Neurobiol. 157, 92–116. doi:10.1016/j.pneurobio.2016.06.006
Hashemi, P., and Ahmadi, S. (2023). Alpha-pinene exerts antiseizure effects by preventing oxidative stress and apoptosis in the Hippocampus in a rat model of temporal lobe epilepsy induced by kainate. Mol. Neurobiol. 60, 3227–3238. doi:10.1007/s12035-023-03274-2
Hashemi, P., Moloudi, M. R., Vahabzadeh, Z., and Izadpanah, E. (2023). Anticonvulsant effects of royal jelly in kainic acid-induced animal model of temporal lobe epilepsy through antioxidant activity. Neurochem. Res. 48, 2187–2195. doi:10.1007/s11064-023-03897-w
He, Z., Ning, N., Zhou, Q., Khoshnam, S. E., and Farzaneh, M. (2020). Mitochondria as a therapeutic target for ischemic stroke. Free Radic. Biol. Med. 146, 45–58. doi:10.1016/j.freeradbiomed.2019.11.005
Ingram, C. J., Weale, M. E., Plaster, C. A., Morrison, K. E., Goodall, E. F., Pall, H. S., et al. (2012). Analysis of European case-control studies suggests that common inherited variation in mitochondrial DNA is not involved in susceptibility to amyotrophic lateral sclerosis. Amyotroph. Lateral Scler. 13, 341–346. doi:10.3109/17482968.2012.654394
Ishola, I. O., Akinleye, M. O., Afolayan, O. O., Okonkwo, H. E., Animashaun, O. T., and Agbaje, E. O. (2022). Anticonvulsant activity of Nymphaea lotus Linn. extract in mice: the role of GABAergic-glutamatergic neurotransmission and antioxidant defence mechanisms. Epilepsy Res. 181, 106871. doi:10.1016/j.eplepsyres.2022.106871
Jaiswal, G., and Kumar, P. (2022). Neuroprotective role of apocynin against pentylenetetrazole kindling epilepsy and associated comorbidities in mice by suppression of ROS/RNS. Behav. Brain Res. 419, 113699. doi:10.1016/j.bbr.2021.113699
Juan, C. A., Pérez De La Lastra, J. M., Plou, F. J., and Pérez-Lebeña, E. (2021). The chemistry of reactive oxygen species (ROS) revisited: outlining their role in biological macromolecules (DNA, lipids and proteins) and induced pathologies. Int. J. Mol. Sci. 22 22, 4642. doi:10.3390/ijms22094642
Kandeda, A. K., Nodeina, S., and Mabou, S. T. (2022). An aqueous extract of Syzygium cumini protects against kainate-induced status epilepticus and amnesia: evidence for antioxidant and anti-inflammatory intervention. Metab. Brain Dis. 37, 2581–2602. doi:10.1007/s11011-022-01052-y
Kandhare, A. D., Mukherjee, A. A., and Bodhankar, S. L. (2018). Anti-epileptic effect of morin against experimental pentylenetetrazol-induced seizures via modulating brain monoamines and oxidative stress. Asian Pac. J. Trop. Biomed. 8 8, 352. doi:10.4103/2221-1691.237078
Kaproń, B., Czarnomysy, R., Wysokiński, M., Andrys, R., Musilek, K., Angeli, A., et al. (2020). 1,2,4-Triazole-based anticonvulsant agents with additional ROS scavenging activity are effective in a model of pharmacoresistant epilepsy. J. Enzyme Inhib. Med. Chem. 35, 993–1002. doi:10.1080/14756366.2020.1748026
Karabulut, S., and Taskiran, A. S. (2021). Effect of pre-treatment with acetaminophen on hippocampal oxidative, inflammatory, and apoptotic parameters in PTZ-induced acute seizure mice model. Neurochem. J. 15, 79–85. doi:10.1134/s1819712421010050
Kazmi, Z., Zeeshan, S., Khan, A., Malik, S., Shehzad, A., Seo, E. K., et al. (2020). Anti-epileptic activity of daidzin in PTZ-induced mice model by targeting oxidative stress and BDNF/VEGF signaling. NeuroToxicology 79, 150–163. doi:10.1016/j.neuro.2020.05.005
Khamse, S., Haftcheshmeh, S. M., Sadr, S. S., Roghani, M., Kamalinejad, M., Moghaddam, P. M., et al. (2021). The potential neuroprotective roles of olive leaf extract in an epilepsy rat model induced by kainic acid. Res. Pharm. Sci. 16, 48–57. doi:10.4103/1735-5362.305188
Kowalczyk, P., Sulejczak, D., Kleczkowska, P., Bukowska-Ośko, I., Kucia, M., Popiel, M., et al. (2021). Mitochondrial oxidative stress-A causative factor and therapeutic target in many diseases. Int. J. Mol. Sci. 22 22, 13384. doi:10.3390/ijms222413384
Kunz, W. S. (2002). The role of mitochondria in epileptogenesis. Curr. Opin. neurology 15, 179–184. doi:10.1097/00019052-200204000-00009
Larsson, N.-G., Wang, J., Wilhelmsson, H., Oldfors, A., Rustin, P., Lewandoski, M., et al. (1998). Mitochondrial transcription factor A is necessary for mtDNA maintenance and embryogenesis in mice. Nat. Genet. 18, 231–236. doi:10.1038/ng0398-231
Li, W., and Kong, A. N. (2009). Molecular mechanisms of Nrf2-mediated antioxidant response. Mol. Carcinog. 48, 91–104. doi:10.1002/mc.20465
Liebler, D. C., Kling, D. S., and Reed, D. J. (1986). Antioxidant protection of phospholipid bilayers by alpha-tocopherol. Control of alpha-tocopherol status and lipid peroxidation by ascorbic acid and glutathione. J. Biol. Chem. 261, 12114–12119. doi:10.1016/s0021-9258(18)67210-2
Lopriore, P., Gomes, F., Montano, V., Siciliano, G., and Mancuso, M. (2022). Mitochondrial epilepsy, a challenge for neurologists. Int. J. Mol. Sci. 23, 13216. doi:10.3390/ijms232113216
Loshali, A., Joshi, B. C., Sundriyal, A., and Uniyal, S. (2021). Antiepileptic effects of antioxidant potent extract from Urtica dioica Linn. root on pentylenetetrazole and maximal electroshock induced seizure models. Heliyon 7, e06195. doi:10.1016/j.heliyon.2021.e06195
Lu, W., Wu, Z., Zhang, C., Gao, T., Ling, X., Xu, M., et al. (2022). Jujuboside A exhibits an antiepileptogenic effect in the rat model via protection against traumatic epilepsy-induced oxidative stress and inflammatory responses. Evid. Based Complement. Altern. Med. 2022, 7792791. doi:10.1155/2022/7792791
Madireddy, S., and Madireddy, S. (2023). Therapeutic strategies to ameliorate neuronal damage in epilepsy by regulating oxidative stress, mitochondrial dysfunction, and neuroinflammation. Brain Sci. 13, 784. doi:10.3390/brainsci13050784
Mahmoudi, T., Lorigooini, Z., Rafieian-Kopaei, M., Arabi, M., Rabiei, Z., Bijad, E., et al. (2020). Effect of Curcuma zedoaria hydro-alcoholic extract on learning, memory deficits and oxidative damage of brain tissue following seizures induced by pentylenetetrazole in rat. Behav. Brain Funct. 16, 7. doi:10.1186/s12993-020-00169-3
Mancuso, M., Filosto, M., Orsucci, D., and Siciliano, G. (2008). Mitochondrial DNA sequence variation and neurodegeneration. Hum. Genomics 3, 71–78. doi:10.1186/1479-7364-3-1-71
Mansouri, S., Hosseini, M., Beheshti, F., Sobhanifar, M. A., Rakhshandeh, H., and Anaeigoudari, A. (2021). Neuroprotective effects of Pinus eldarica in a mouse model of pentylenetetrazole-induced seizures. Avicenna J. Phytomed 11, 610–621. doi:10.22038/AJP.2021.18562
Marinova, E., Toneva, A., and Yanishlieva, N. (2008). Synergistic antioxidant effect of α-tocopherol and myricetin on the autoxidation of triacylglycerols of sunflower oil. Food Chem. 106, 628–633. doi:10.1016/j.foodchem.2007.06.022
Mehmet, K., Alper, E. M., and Oytun, E. (2022). Anti-seizure effect of zinc on PTZ-induced epilepsy in rat model. Bratisl. Med. Journal/Bratislavské Lekárske Listy 123 123, 648–652. doi:10.4149/BLL_2022_104
Meng, J., Lv, Z., Zhang, Y., Wang, Y., Qiao, X., Sun, C., et al. (2021). Precision redox: the key for antioxidant pharmacology. Antioxid. Redox Signal 34, 1069–1082. doi:10.1089/ars.2020.8212
Merlin, J. P. J., Mathavarajah, S., Dellaire, G., Murphy, K. P. J., and Rupasinghe, H. P. V. (2022). A dietary antioxidant formulation ameliorates DNA damage caused by γ-irradiation in normal human bronchial epithelial cells in vitro. Antioxidants 11, 1407. doi:10.3390/antiox11071407
Miller, V. J., Lafountain, R. A., Barnhart, E., Sapper, T. S., Short, J., Arnold, W. D., et al. (2020). A ketogenic diet combined with exercise alters mitochondrial function in human skeletal muscle while improving metabolic health. Am. J. Physiology-Endocrinology Metabolism 319, E995-E1007–E1007. doi:10.1152/ajpendo.00305.2020
Mishra, P., Mittal, A. K., Rajput, S. K., and Sinha, J. K. (2021). Cognition and memory impairment attenuation via reduction of oxidative stress in acute and chronic mice models of epilepsy using antiepileptogenic Nux vomica. J. Ethnopharmacol. 267, 113509. doi:10.1016/j.jep.2020.113509
Munakata, K., Tanaka, M., Mori, K., Washizuka, S., Yoneda, M., Tajima, O., et al. (2004). Mitochondrial DNA 3644T-->C mutation associated with bipolar disorder. Genomics 84, 1041–1050. doi:10.1016/j.ygeno.2004.08.015
Murphy, M. P., and Smith, R. A. (2007). Targeting antioxidants to mitochondria by conjugation to lipophilic cations. Annu. Rev. Pharmacol. Toxicol. 47, 629–656. doi:10.1146/annurev.pharmtox.47.120505.105110
Nahirney, P. C., Reeson, P., and Brown, C. E. (2016). Ultrastructural analysis of blood–brain barrier breakdown in the peri-infarct zone in young adult and aged mice. J. Cereb. Blood Flow and Metabolism 36, 413–425. doi:10.1177/0271678X15608396
Neueder, A., Kojer, K., Gu, Z., Wang, Y., Hering, T., Tabrizi, S., et al. (2024). Huntington’s disease affects mitochondrial network dynamics predisposing to pathogenic mitochondrial DNA mutations. Brain 147, 2009–2022. doi:10.1093/brain/awae007
Nguezeye, Y., Yadang, F. S. A., Pale, S., Jugha, V. T., Mambou, H. M. a.Y., Bila, R. B., et al. (2023). Anticonvulsant effects of chrysanthellum americanum L. (Vatke) aqueous extract in mice pilocarpine model of epilepsy and associated memory impairment: role of antioxidant defense system and cholinergic transmission. J. Biosci. Med. 11, 81–102. doi:10.4236/jbm.2023.116006
Niki, E. (1987). Interaction of ascorbate and alpha-tocopherol. Ann. N. Y. Acad. Sci. 498, 186–199. doi:10.1111/j.1749-6632.1987.tb23761.x
Nkwingwa, B. K., Wado, E. K., Foyet, H. S., Bouvourne, P., Jugha, V. T., Mambou, A., et al. (2023). Ameliorative effects of Albizia adianthifolia aqueous extract against pentylenetetrazole-induced epilepsy and associated memory loss in mice: role of GABAergic, antioxidant defense and anti-inflammatory systems. Biomed. Pharmacother. 165, 115093. doi:10.1016/j.biopha.2023.115093
Onishi, K., Kamida, T., Fujiki, M., Momii, Y., and Sugita, K. (2023). Anticonvulsant and antioxidant effects of lamotrigine on pilocarpine-induced status epilepticus in mice. NeuroReport 34 34, 61–66. doi:10.1097/WNR.0000000000001859
Osellame, L. D., Blacker, T. S., and Duchen, M. R. (2012). Cellular and molecular mechanisms of mitochondrial function. Best. Pract. Res. Clin. Endocrinol. Metab. 26, 711–723. doi:10.1016/j.beem.2012.05.003
Palanisamy, G. S., Kirk, N. M., Ackart, D. F., Shanley, C. A., Orme, I. M., and Basaraba, R. J. (2011). Evidence for oxidative stress and defective antioxidant response in Guinea pigs with tuberculosis. PloS one 6, e26254. doi:10.1371/journal.pone.0026254
Patel, M. N. (2002). Oxidative stress, mitochondrial dysfunction, and epilepsy. Free Radic. Res. 36, 1139–1146. doi:10.1080/1071576021000016391
Pauletti, A., Terrone, G., Shekh-Ahmad, T., Salamone, A., Ravizza, T., Rizzi, M., et al. (2019). Targeting oxidative stress improves disease outcomes in a rat model of acquired epilepsy. Brain 142, e39. doi:10.1093/brain/awz130
Peng, Y., Chen, L., Qu, Y., Wang, D., Zhu, Y., and Zhu, Y. (2021). Rosiglitazone prevents autophagy by regulating Nrf2-antioxidant response element in a rat model of lithium-pilocarpine-induced status epilepticus. Neuroscience 455, 212–222. doi:10.1016/j.neuroscience.2020.10.026
Petrillo, S., Gallo, M. G., Santoro, A., Brugaletta, R., Nijhawan, P., Russo, C., et al. (2022). Personalized profiles of antioxidant signaling pathway in patients with tuberculosis. J. Microbiol. Immunol. Infect. 55, 405–412. doi:10.1016/j.jmii.2021.07.004
Pires, L. F., Costa, L. M., De Almeida, A. a.C., Silva, O. A., Cerqueira, G. S., De Sousa, D. P., et al. (2015). Neuropharmacological effects of carvacryl acetate on δ-aminolevulinic dehydratase, Na+, K+-ATPase activities and amino acids levels in mice hippocampus after seizures. Chemico-Biological Interact. 226, 49–57. doi:10.1016/j.cbi.2014.12.001
Pizzino, G., Irrera, N., Cucinotta, M., Pallio, G., Mannino, F., Arcoraci, V., et al. (2017). Oxidative stress: harms and benefits for human health. Oxid. Med. Cell Longev. 2017, 8416763. doi:10.1155/2017/8416763
Plotnikov, E. Y., and Zorov, D. B. (2019). Pros and cons of use of mitochondria-targeted antioxidants. Antioxidants (Basel) 8 8, 316. doi:10.3390/antiox8080316
Podmore, I. D., Griffiths, H. R., Herbert, K. E., Mistry, N., Mistry, P., and Lunec, J. (1998). Vitamin C exhibits pro-oxidant properties. Nature 392, 559. doi:10.1038/33308
Quinzii, C. M., and Hirano, M. (2010). Coenzyme Q and mitochondrial disease. Dev. Disabil. Res. Rev. 16, 183–188. doi:10.1002/ddrr.108
Ramazi, S., Fahanik-Babaei, J., Mohamadi-Zarch, S.-M., Tashakori-Miyanroudi, M., Nourabadi, D., Nazari-Serenjeh, M., et al. (2020). Neuroprotective and anticonvulsant effects of sinomenine in kainate rat model of temporal lobe epilepsy: involvement of oxidative stress, inflammation and pyroptosis. J. Chem. Neuroanat. 108, 101800. doi:10.1016/j.jchemneu.2020.101800
Rambo, L. M., Ribeiro, L. R., Della-Pace, I. D., Stamm, D. N., Da Rosa Gerbatin, R., Prigol, M., et al. (2013). Acute creatine administration improves mitochondrial membrane potential and protects against pentylenetetrazol-induced seizures. Amino Acids 44, 857–868. doi:10.1007/s00726-012-1408-6
Rawat, C., Kukal, S., Dahiya, U. R., and Kukreti, R. (2019). Cyclooxygenase-2 (COX-2) inhibitors: future therapeutic strategies for epilepsy management. J. Neuroinflammation 16, 197. doi:10.1186/s12974-019-1592-3
Rea, I. M., Mcnerlan, S. E., Archbold, G. P., Middleton, D., Curran, M. D., Young, I. S., et al. (2013). Mitochondrial J haplogroup is associated with lower blood pressure and anti-oxidant status: findings in octo/nonagenarians from the BELFAST Study. Age (Dordr) 35, 1445–1456. doi:10.1007/s11357-012-9444-4
Reinecke, F., Smeitink, J. a.M., and Van Der Westhuizen, F. H. (2009). OXPHOS gene expression and control in mitochondrial disorders. Biochimica Biophysica Acta (BBA) - Mol. Basis Dis. 1792, 1113–1121. doi:10.1016/j.bbadis.2009.04.003
Rho, J. M., and Boison, D. (2022). The metabolic basis of epilepsy. Nat. Rev. Neurol. 18, 333–347. doi:10.1038/s41582-022-00651-8
Rodriguez, M. C., Macdonald, J. R., Mahoney, D. J., Parise, G., Beal, M. F., and Tarnopolsky, M. A. (2007). Beneficial effects of creatine, CoQ10, and lipoic acid in mitochondrial disorders. Muscle Nerve 35, 235–242. doi:10.1002/mus.20688
Rostamian, S., Keshavarz Hedayati, S., Khosraviani, S., Aali, E., and Naderi, Y. (2021). Anticonvulsive and antioxidant effects of pioglitazone on pilocarpine-induced seizures in mice. Iran. J. Toxicol. 15, 271–278. doi:10.32598/ijt.15.4.833.1
Rowley, S., Liang, L.-P., Fulton, R., Shimizu, T., Day, B., and Patel, M. (2015). Mitochondrial respiration deficits driven by reactive oxygen species in experimental temporal lobe epilepsy. Neurobiol. Dis. 75, 151–158. doi:10.1016/j.nbd.2014.12.025
Saint Martin, E. M., Sosa, A. M., Martinez, C. S., Prieto, J., Marrassini, C., Dobrecky, C. B., et al. (2024). In vitro antioxidant activity and anticonvulsant properties on zebrafish PTZ-induced seizure model of a Tilia viridis aqueous extract. J. Pharmacopuncture 27, 211–222. doi:10.3831/KPI.2024.27.3.211
Sandouka, S., and Shekh-Ahmad, T. (2021). Induction of the Nrf2 pathway by sulforaphane is neuroprotective in a rat temporal lobe epilepsy model. Antioxidants 10, 1702. doi:10.3390/antiox10111702
Sattarinezhad, E., Shafaroodi, H., Sheikhnouri, K., Mousavi, Z., and Moezi, L. (2014). The effects of coenzyme Q10 on seizures in mice: the involvement of nitric oxide. Epilepsy Behav. 37, 36–42. doi:10.1016/j.yebeh.2014.05.024
Scheffer, I. E., Berkovic, S., Capovilla, G., Connolly, M. B., French, J., Guilhoto, L., et al. (2017). ILAE classification of the epilepsies: position paper of the ILAE commission for classification and terminology. Epilepsia 58, 512–521. doi:10.1111/epi.13709
Schieber, M., and Chandel, N. S. (2014). ROS function in redox signaling and oxidative stress. Curr. Biol. 24, R453–R462. doi:10.1016/j.cub.2014.03.034
Schneider Oliveira, M., Flávia Furian, A., Freire Royes, L. F., Rechia Fighera, M., De Carvalho Myskiw, J., Gindri Fiorenza, N., et al. (2004). Ascorbate modulates pentylenetetrazol-induced convulsions biphasically. Neuroscience 128, 721–728. doi:10.1016/j.neuroscience.2004.07.012
Schwarz, A. P., Dyomina, A. V., Zakharova, M. V., Kovalenko, A. A., Gryaznova, M. O., Ischenko, A. M., et al. (2022). The reference gene validation in the brain of rats during antioxidant and anti-inflammatory treatment in the lithium-pilocarpine model of temporal epilepsy. J. Evol. Biochem. Physiology 58, 930–940. doi:10.1134/s0022093022030279
Sharifi-Rad, M., Anil Kumar, N. V., Zucca, P., Varoni, E. M., Dini, L., Panzarini, E., et al. (2020). Lifestyle, oxidative stress, and antioxidants: back and forth in the pathophysiology of chronic diseases. Front. Physiol. 11, 694. doi:10.3389/fphys.2020.00694
Shaughnessy Daniel, T., Mcallister, K., Worth, L., Haugen Astrid, C., Meyer Joel, N., Domann Frederick, E., et al. (2014). Mitochondria, energetics, epigenetics, and cellular responses to stress. Environ. Health Perspect. 122, 1271–1278. doi:10.1289/ehp.1408418
Shekh-Ahmad, T., Lieb, A., Kovac, S., Gola, L., Wigley, W. C., Abramov, A. Y., et al. (2019). Combination antioxidant therapy prevents epileptogenesis and modifies chronic epilepsy. Redox Biol. 26, 101278. doi:10.1016/j.redox.2019.101278
Shishmanova-Doseva, M., Peychev, L., Yoanidu, L., Uzunova, Y., Atanasova, M., Georgieva, K., et al. (2021). Anticonvulsant effects of topiramate and lacosamide on pilocarpine-induced status epilepticus in rats: a role of reactive oxygen species and inflammation. Int. J. Mol. Sci. [Online] 22, 2264. doi:10.3390/ijms22052264
Simani, L., Rezaei, O., Ryan, F., Sadeghi, M., Hooshmandi, E., Ramezani, M., et al. (2020). Coenzyme Q10 insufficiency contributes to the duration and frequency of seizures in epileptic patients. Basic Clin. Neurosci. 11, 765–771. doi:10.32598/bcn.11.6.1100.1
Simeone, K. A., Matthews, S. A., Samson, K. K., and Simeone, T. A. (2014). Targeting deficiencies in mitochondrial respiratory complex I and functional uncoupling exerts anti-seizure effects in a genetic model of temporal lobe epilepsy and in a model of acute temporal lobe seizures. Exp. Neurol. 251, 84–90. doi:10.1016/j.expneurol.2013.11.005
Şirinyıldız, F., and Ek, R. O. (2021). Effects of melatonin at different doses on experimental epilepsy model induced by pentylenetetrazole. Meandros Med. Dent. J. 22, 177–186. doi:10.4274/meandros.galenos.2021.49358
Skulachev, V., Vyssokikh, M. Y., Chernyak, B., Averina, O., Andreev-Andrievskiy, A., Zinovkin, R., et al. (2023). Mitochondrion-targeted antioxidant SkQ1 prevents rapid animal death caused by highly diverse shocks. Sci. Rep. 13, 4326. doi:10.1038/s41598-023-31281-9
Smilin Bell Aseervatham, G., Abbirami, E., Sivasudha, T., and Ruckmani, K. (2020). Passiflora caerulea L. fruit extract and its metabolites ameliorate epileptic seizure, cognitive deficit and oxidative stress in pilocarpine-induced epileptic mice. Metab. Brain Dis. 35, 159–173. doi:10.1007/s11011-019-00501-5
Snow, B. J., Rolfe, F. L., Lockhart, M. M., Frampton, C. M., O'sullivan, J. D., Fung, V., et al. (2010). A double-blind, placebo-controlled study to assess the mitochondria-targeted antioxidant MitoQ as a disease-modifying therapy in Parkinson's disease. Mov. Disord. 25, 1670–1674. doi:10.1002/mds.23148
Soini, H. K., Moilanen, J. S., Vilmi-Kerälä, T., Finnilä, S., and Majamaa, K. (2013). Mitochondrial DNA variant m.15218A > G in Finnish epilepsy patients who have maternal relatives with epilepsy, sensorineural hearing impairment or diabetes mellitus. BMC Med. Genet. 14, 73. doi:10.1186/1471-2350-14-73
Souza, M. A., Mota, B. C., Gerbatin, R. R., Rodrigues, F. S., Castro, M., Fighera, M. R., et al. (2013). Antioxidant activity elicited by low dose of caffeine attenuates pentylenetetrazol-induced seizures and oxidative damage in rats. Neurochem. Int. 62, 821–830. doi:10.1016/j.neuint.2013.02.021
Stewart, J. B., and Chinnery, P. F. (2015). The dynamics of mitochondrial DNA heteroplasmy: implications for human health and disease. Nat. Rev. Genet. 16, 530–542. doi:10.1038/nrg3966
Taiwe, G. S., Ndieudieu Kouamou, A. L., Dabole, B., Ambassa, A. R. M., Mambou, H., Bila, R. B., et al. (2021). Protective effects of anthocleista djalonensis extracts against pentylenetetrazole-induced epileptic seizures and neuronal cell loss: role of antioxidant defense system. Evid. Based Complement. Altern. Med. 2021, 5523705. doi:10.1155/2021/5523705
Tao, Z., Chun-Yan, H., Hua, P., Bin-Bin, Y., and Xiaoping, T. (2020). Phyllathin from phyllanthus amarus ameliorates epileptic convulsion and kindling associated post-Ictal depression in mice via inhibition of NF-κB/TLR-4 pathway. Dose-Response 18, 1559325820946914. doi:10.1177/1559325820946914
Tarnopolsky, M. A. (2008). The mitochondrial cocktail: rationale for combined nutraceutical therapy in mitochondrial cytopathies. Adv. Drug Deliv. Rev. 60, 1561–1567. doi:10.1016/j.addr.2008.05.001
Tarnopolsky, M. A., and Beal, M. F. (2001). Potential for creatine and other therapies targeting cellular energy dysfunction in neurological disorders. Ann. Neurol. 49, 561–574. doi:10.1002/ana.1028.abs
Taskiran, A. S., and Tastemur, Y. (2021). The role of nitric oxide in anticonvulsant effects of lycopene supplementation on pentylenetetrazole-induced epileptic seizures in rats. Exp. Brain Res. 239, 591–599. doi:10.1007/s00221-020-06012-5
Tavakoli, Z., Tahmasebi Dehkordi, H., Lorigooini, Z., Rahimi-Madiseh, M., Korani, M. S., and Amini-Khoei, H. (2023). Anticonvulsant effect of quercetin in pentylenetetrazole (PTZ)-induced seizures in male mice: the role of anti-neuroinflammatory and anti-oxidative stress. Int. Immunopharmacol. 116, 109772. doi:10.1016/j.intimp.2023.109772
Thoo, Y. Y., Abas, F., Lai, O. M., Ho, C. W., Yin, J., Hedegaard, R. V., et al. (2013). Antioxidant synergism between ethanolic Centella asiatica extracts and α-tocopherol in model systems. Food Chem. 138, 1215–1219. doi:10.1016/j.foodchem.2012.11.013
Turrens, J. F. (2003). Mitochondrial formation of reactive oxygen species. J. Physiol. 552, 335–344. doi:10.1113/jphysiol.2003.049478
Varga, N., Pentelényi, K., Balicza, P., Gézsi, A., Reményi, V., Hársfalvi, V., et al. (2018). Mitochondrial dysfunction and autism: comprehensive genetic analyses of children with autism and mtDNA deletion. Behav. Brain Funct. 14, 4. doi:10.1186/s12993-018-0135-x
Vergani, L., Floreani, M., Russell, A., Ceccon, M., Napoli, E., Cabrelle, A., et al. (2004). Antioxidant defences and homeostasis of reactive oxygen species in different human mitochondrial DNA-depleted cell lines. Eur. J. Biochem. 271, 3646–3656. doi:10.1111/j.1432-1033.2004.04298.x
Volpe, C. M. O., Villar-Delfino, P. H., Dos Anjos, P. M. F., and Nogueira-Machado, J. A. (2018). Cellular death, reactive oxygen species (ROS) and diabetic complications. Cell Death Dis. 9, 119. doi:10.1038/s41419-017-0135-z
Waldbaum, S., and Patel, M. (2010). Mitochondria, oxidative stress, and temporal lobe epilepsy. Epilepsy Res. 88, 23–45. doi:10.1016/j.eplepsyres.2009.09.020
Wang, K., Liu, Y., Shi, Y., Yan, M., Rengarajan, T., and Feng, X. (2021). Amomum tsaoko fruit extract exerts anticonvulsant effects through suppression of oxidative stress and neuroinflammation in a pentylenetetrazol kindling model of epilepsy in mice. Saudi J. Biol. Sci. 28, 4247–4254. doi:10.1016/j.sjbs.2021.06.007
Wang, S., Meckling, K. A., Marcone, M. F., Kakuda, Y., and Tsao, R. (2011). Synergistic, additive, and antagonistic effects of food mixtures on total antioxidant capacities. J. Agric. Food Chem. 59, 960–968. doi:10.1021/jf1040977
Wang, Y., Guo, X., Hong, X., Wang, G., Pearson, C., Zuckerman, B., et al. (2022). Association of mitochondrial DNA content, heteroplasmies and inter-generational transmission with autism. Nat. Commun. 13, 3790. doi:10.1038/s41467-022-30805-7
Xhuti, D., Rebalka, I. A., Minhas, M., May, L., Murphy, K., Nederveen, J. P., et al. (2023). The acute effect of multi-ingredient antioxidant supplementation following ionizing radiation. Nutrients 15, 207. doi:10.3390/nu15010207
Yamamoto, H.-A., and Mohanan, P. V. (2003). Ganglioside GT1B and melatonin inhibit brain mitochondrial DNA damage and seizures induced by kainic acid in mice. Brain Res. 964, 100–106. doi:10.1016/s0006-8993(02)04083-0
Yilgor, A., and Demir, C. (2024). Determination of oxidative stress level and some antioxidant activities in refractory epilepsy patients. Sci. Rep. 14, 6688. doi:10.1038/s41598-024-57224-6
Yuen, A. W. C., and Sander, J. W. (2011). Impaired mitochondrial energy production: the basis of pharmacoresistance in epilepsy. Med. Hypotheses 77, 536–540. doi:10.1016/j.mehy.2011.06.025
Yunusoğlu, O., Dzhafar, S., Dalar, A., Mükemre, M., Ekin, S., and Yıldız, D. (2020). Phytochemical profile and in vitro and in vivo anticonvulsant and antioxidant activities of epilobium hirsutum. Int. J. Second. Metabolite 7, 63–76. doi:10.21448/ijsm.669451
Zeviani, M., Muntoni, F., Savarese, N., Serra, G., Tiranti, V., Carrara, F., et al. (1993). A MERRF/MELAS overlap syndrome associated with a new point mutation in the mitochondrial DNA tRNALys gene. Eur. J. Hum. Genet. 1, 80–87. doi:10.1159/000472390
Zhang, L., Wu, T., Kandhare, A., Mukherjee, A., Guo, G., and Bodhankar, S. L. (2018). Elucidation of the molecular mechanism of tempol in pentylenetetrazol-induced epilepsy in mice: role of gamma-aminobutyric acid, tumor necrosis factor-alpha, interleukin-1β and c-Fos. Pharmacogn. Mag. 14 14, 520. doi:10.4103/pm.pm_263_18
Zhang, X., Wu, S., Guo, C., Guo, K., Hu, Z., Peng, J., et al. (2022). Vitamin E exerts neuroprotective effects in pentylenetetrazole kindling epilepsy via suppression of ferroptosis. Neurochem. Res. 47, 739–747. doi:10.1007/s11064-021-03483-y
Keywords: epilepsy, antioxidant therapy, oxidative stress, mitochondrial dysfunction, reactive oxygen species
Citation: Ji D, Mylvaganam S, Ravi Chander P, Tarnopolsky M, Murphy K and Carlen P (2025) Mitochondria and oxidative stress in epilepsy: advances in antioxidant therapy. Front. Pharmacol. 15:1505867. doi: 10.3389/fphar.2024.1505867
Received: 03 October 2024; Accepted: 26 December 2024;
Published: 19 March 2025.
Edited by:
Eva Zerovnik, Institut Jožef Stefan (IJS), SloveniaReviewed by:
Wieslawa Agnieszka Fogel, Polish Academy of Sciences, PolandCopyright © 2025 Ji, Mylvaganam, Ravi Chander, Tarnopolsky, Murphy and Carlen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Peter Carlen, cGV0ZXIuY2FybGVuQHVobi5jYQ==; Delphine Ji, ZGVscGhpbmUuamlAbWFpbC51dG9yb250by5jYQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.