 Rui Yang
Rui Yang Ranran Wang
Ranran Wang Ajing Xu
Ajing Xu- Department of Clinical Pharmacy, Xinhua Hospital, Shanghai Jiaotong University School of Medicine, Shanghai, China
Neurodegenerative diseases (NDDs) represent a category of serious illnesses characterized by the progressive deterioration of neuronal structure and function. The exploration of natural compounds as potential therapeutic agents has gained increasing attention in recent years owing to their wide range of pharmacological activities and minimal side effects. Baicalin (BAI) and baicalein (BE), polyphenolic flavonoids, derived from the root of Scutellaria baicalensis, evidently show potential in treating NDDs. This review provides an overview of the current understanding of the roles of BAI and BE in alleviating neuroinflammation, a pivotal pathological process implicated in various NDDs. Studies conducted prior to clinical trials have shown that BAI and BE exert protective effects on the nervous system in different animal models of NDDs. Furthermore, mechanistic studies indicate that BAI and BE exert anti-inflammatory effects by inhibiting pro-inflammatory cytokines, suppressing microglial activation, and regulating microglial phenotypes. These effects are mediated through the modulation of inflammatory signaling cascades, including Toll-like receptor 4 (TLR4), mitogen-activated protein kinase (MAPK), amp-activated protein kinase (AMPK), NOD-like receptor thermal protein domain-associated protein 3 (NLRP3) inflammasome, and nuclear factor erythroid 2-related factor 2 (Nrf2)/hemoglobin oxygenase-1 (HO-1). Overall, BAI and BE exhibit promising potential as natural compounds with anti-inflammatory properties and offer innovative therapeutic approaches for managing NDDs.
1 Introduction
Scutellaria baicalensis (S. baicalensis), commonly known as Huangqin or Chinese skullcap, is widely distributed in northern, northwestern, and southwestern China (Shang et al., 2010). It has also been found in the Soviet Union, Mongolia, North Korea, and Japan. Baicalin (BAI; syn.baicalein7-O-β-D-glucuronic acid) and baicalein (BE; 5, 6, 7-trihydroxyflavone), the principal compounds derived from the roots of S. baicalensis, are polyphenolic substances and members of the flavone subclass of flavonoids (Gaire et al., 2014). BAI and its aglycone, BE, have attracted increasing attention from the pharmaceutical industry because of their remarkable biological activities. These two flavonoids share structural similarities and can be converted into each other during metabolism within the body (Liang et al., 2017). BAI, as a glucuronide form of BE, is hydrolyzed by glucuronidases in the intestines, liver, and other tissues, releasing baicalein. This process allows baicalein to exert its biological activities directly. Conversely, BE can be conjugated with glucuronic acid to form BAI through glucuronidation reactions, primarily occurring in the liver. Their pharmacological properties have garnered significant attention in recent years, leading to extensive research on their various therapeutic applications.
BAI is formed by combining BE with glucuronic acid. Due to its glucuronic acid component, BAI exhibits higher water solubility but has relatively poor ability to penetrate the intestinal epithelium, requiring specific transport mechanisms or enzymatic action to enter the bloodstream. In contrast, BE has smaller molecular size and higher lipid solubility, allowing it to more easily penetrate the intestinal epithelium and enter the bloodstream directly. It does not require specific transport mechanisms or enzymatic assistance, making it more efficient in crossing the intestinal barrier (Zhang et al., 2007; Li et al., 2011). BAI is moderately absorbed in the stomach, with limited absorption in the small intestine and colon. Conversely, BE is efficiently absorbed in the stomach and small intestine; however, its absorption in the colon is somewhat restricted. Owing to body dynamics, BE is more completely absorbed and converted back to BAI in the systemic circulation through conjugative reactions. Circulating BAI is expected to return to the gastrointestinal tract via biliary excretion (Taiming and Xuehua, 2006). BAI cannot directly cross the intestine but is hydrolyzed to BE by enzymes and bacteria (Akao et al., 2000; Day et al., 2003). BAI can cross the blood–brain barrier (BBB), quickly spread to the cerebrospinal fluid, and peak at a concentration of 344 μg/L in approximately 30 min after the intravenous administration of 24 mg/kg BAI. BAI has a tendency to build up in the striatum, thalamus, and hippocampus, which helps reinforce its positive impact on the central nervous system (CNS) (Zhang et al., 2006; Huang et al., 2008). Hence, the spread of BAI in the brain reinforces its healing properties on the central nervous system. Upon oral administration of BAI, the presence of both unchanged BAI and BE conjugates of glucuronide and sulfate is reportedly observed in the serum (Lai et al., 2003). BAI and BE are excreted via both the biliary and renal pathways (Akao et al., 2009).
BAI and BE exhibit diverse pharmacological effects, and one of the primary areas of research is their potent antioxidant activity. They can eliminate free radicals and decrease oxidative stress, which helps protect cells from damage and may alleviate various diseases linked to oxidative stress, such as heart problems, brain disorders, and cancer (Waisundara et al., 2009; Ma et al., 2021; Wang et al., 2021; Chen et al., 2022; Nie et al., 2023). Furthermore, BAI and BE exhibit anti-inflammatory effects by modulating various inflammatory mediators and signaling pathways. Their capacity to suppress the generation of pro-inflammatory cytokines enhances their effectiveness in managing inflammatory disorders, such as arthritis, inflammatory bowel disease, and asthma (Chen et al., 2014; Zhang et al., 2014; Dinda et al., 2017; Wang et al., 2021; Wen et al., 2023). Moreover, BAI and BE have been investigated for their neuroprotective properties. They may offer neuroprotection through mechanisms such as reducing neuronal apoptosis, suppressing neuroinflammation, and promoting neuronal regeneration (Yuan et al., 2020; Zhao et al., 2021; Huang et al., 2024; Song et al., 2024). The characteristics of BAI and its aglycone BE make them potential options for treating neurodegenerative diseases (NDDs), such as Alzheimer’s disease (AD), Parkinson’s disease (PD), and stroke.
2 Literature search strategy
To provide an overview of the research articles on BAI and BE in neurodegenerative diseases, we searched the databases PubMed, Web of Science, Embase, and Scopus. The search strategy used in PubMed was as follows: ((“Neurodegenerative Diseases” [MeSH] OR “Alzheimer Disease” [MeSH] OR “Parkinson Disease” [MeSH] OR “Huntington Disease” OR “Amyotrophic Lateral Sclerosis” OR “Multiple Sclerosis”) AND (“Neuroinflammation” [MeSH] OR “Brain Inflammation” OR “Microglia Activation”)) AND (“Baicalin” OR “Baicalein” OR “Flavonoids” OR “Scutellaria baicalensis”). The search strategy used in Web of Science was as follows: ((“Neurodegenerative Diseases” OR “Alzheimer Disease” OR “Parkinson Disease” OR “Huntington Disease” OR “Amyotrophic Lateral Sclerosis” OR “Multiple Sclerosis”) AND (“Neuroinflammation” OR “Brain Inflammation” OR “Microglia Activation”)) AND (“Baicalin” OR “Baicalein” OR “Flavonoids” OR “Scutellaria baicalensis”). The search strategy used in Embase was as follows: ((“Neurodegenerative Diseases” OR “Alzheimer Disease” OR “Parkinson Disease” OR “Huntington Disease” OR “Amyotrophic Lateral Sclerosis” OR “Multiple Sclerosis”) AND (“Neuroinflammation” OR “Brain Inflammation” OR “Microglia Activation”)) AND (“Baicalin” OR ‘Baicalein” OR “Scutellaria baicalensis”). The search strategy used in Scopus was as follows: (TITLE-ABS-KEY (“Neurodegenerative Diseases”) OR TITLE-ABS-KEY (“Alzheimer Disease”) OR TITLE-ABS-KEY (“Parkinson Disease”) OR TITLE-ABS-KEY (“Huntington Disease”) OR TITLE-ABS-KEY (“Amyotrophic Lateral Sclerosis”) OR TITLE-ABS-KEY (“Multiple Sclerosis”)) AND (TITLE-ABS-KEY (“Neuroinflammation”) OR TITLE-ABS-KEY (“Brain Inflammation”) OR TITLE-ABS-KEY (“Microglia Activation”)) AND (TITLE-ABS-KEY (“Baicalin”) OR TITLE-ABS-KEY (“Baicalein”) OR TITLE-ABS-KEY (“Scutellaria baicalensis”)).
3 NDDs and neuroinflammation
NDDs encompass a broad spectrum of neurological disorders characterized by various clinical and pathological hallmarks, affecting specific subsets of neurons within distinct regions of the CNS, mainly including AD, PD, multiple sclerosis (MS), amyotrophic lateral sclerosis (ALS), Huntington’s disease (HD), and multiple system atrophy (MSA). Although the pathogenic mechanisms of these diseases are different, such as different protein aggregates and genetic variations, they all share the common hallmark of chronic neuroinflammation (Mayne et al., 2020). Increasing evidence has demonstrated that neuroinflammation may not merely be a consequence of protein aggregation; rather, it may initiate the accumulation of aggregates at the earliest phase of the disease process (Sosna et al., 2018; Gao et al., 2023).
Microglia, considered macrophages of the CNS, play an important role in neuroinflammation. Resting microglia, also known as M0 microglia, maintain the homeostasis of their adjacent environment during active immune surveillance (Gao et al., 2023). Upon exogenous or endogenous stimuli, microglia are rapidly activated and shift to an activated phenotype, which is typically characterized by two phenotypic states: a classically activated M1 phenotype or an alternatively activated M2 phenotype. Generally, the M1 phenotype is associated with pro-inflammatory and neurotoxic responses, whereas the M2 phenotype predominantly possesses anti-inflammatory and neuroprotective functions (Woodburn et al., 2021). During aging, microglia tend to display a dominant M1-like phenotype associated with neurotoxic responses (Ward et al., 2015). In NDDs, endogenous pathological protein aggregation, neuronal damage, and microglial-related neuroinflammation are interconnected via a positive feedback loop.
In AD, microglia become persistently activated by the aggregation of β-amyloid peptide (Aβ) and subsequently transform into the M1 phenotype. They release a wide variety of pro-inflammatory and toxic productions, amplifying immune responses, leading to neurotoxicity (Meda et al., 2001), and increasing the secretion of Aβ fragments and the aggregation of soluble β-amyloid (Tan and Seshadri, 2010). In PD, excessive aggregation of α-synuclein (α-syn) can be released either directly from neurons or via exosomes, thereby activating microglia. Subsequently, activated microglia exacerbate the disease by releasing cytokines and chemokines that enhance α-syn pathogenicity, induce neuron death, and further enhance microglia activation (Reimer et al., 2018; Pajares et al., 2020). Similar to PD, MSA is also associated with the pathological protein α-syn. Its hallmark pathological characteristic is the presence of glial cytoplasmic inclusions (GCIs), rich in α-syn, within oligodendrocytes. Misfolded α-syn serves as a main trigger for microglial activation, which is believed to accelerate α-syn aggregation and promote the apoptosis of oligodendrocytes (Vieira et al., 2015). In MS, microglia are not the primary triggers but are influenced by a wider immune disturbance (Ransohoff, 2016). They adopt a pro-inflammatory behavior, such as antigen presentation, phagocytosis, and secretion of cytokines and chemokines, which play a central role in the pathogenesis of MS (Li and Barres, 2018; Voet et al., 2019).
Neuroinflammation also plays a vital role in the progression of other NDDs. In ALS, microglia can become overactivated due to the aggregation of TAR DNA-binding protein 43 and Cu/Zn superoxide dismutase 1 (SOD1), producing pro-inflammatory cytokines, which not only directly damage neurons but also exacerbate ALS (Zhao et al., 2015; Calió et al., 2020; Yu et al., 2020). There is a significant increase in activated microglia and pro-inflammatory cytokine levels in the brains of patients with HD and in vivo models (Tai et al., 2007; Sivandzade et al., 2019; Subhramanyam et al., 2019; Saba et al., 2022), which promote neuronal cell death by inducing apoptosis and reactive oxygen species (ROS)/reactive nitrogen species (RNS) production and complement activation, enhancing excitotoxicity and mitochondrial damage (Smith et al., 2012; Saba et al., 2022). In addition to that, chronic cerebral hypoperfusion (CCH) is the common underlying pathophysiological mechanism, which is a major contributor to cognitive decline and degenerative processes (Tian et al., 2022). A continuous decrease in cerebral blood flow causes cell death, and the subsequent release of cell debris will induce the neuroinflammation–immune cascade reaction (Ma et al., 2017). During this process, microglia could turn into an M1 phenotype that releases pro-inflammatory cytokines that further aggravate neuroinflammation and tissue damage, promoting the development of cognitive dysfunction and degeneration (Ma et al., 2017; Tian et al., 2022).
In summary, pathogenic protein aggregation, neuronal damage, and microglial-related neuroinflammation exhibit mutually reinforcing patterns that exacerbate neurodegeneration. Therefore, controlling neuroinflammation is a potential target for NDD therapy. Natural products that modulate neuroinflammation have attracted considerable attention in this field.
4 The effects of BAI and BE on neuroinflammation-related signaling pathways
BAI and BE, essential natural flavonoids extracted from the roots of S. baicalensis, have various biological and pharmacological effects and can cross the BBB (Huang et al., 2008). Extensive research has demonstrated that BAI and BE can inhibit neuroinflammation by targeting various signaling pathways depending on specific NDDs (Figure 1) (Marogianni et al., 2020; Pan et al., 2021; Scheltens et al., 2021; Araujo et al., 2022; Zhang W. et al., 2023).

Figure 1. Plausible molecular mechanisms of BAI and BE in NDDs.
4.1 NF-κB signaling
Nuclear factor-kappa B (NF-κB), implicated in anti-inflammatory processes across various neurodegenerative conditions, typically resides in the cytoplasm bound to the regulatory protein inhibitor of NF-κB (IκB) (Sivandzade et al., 2019; Sun et al., 2022; Xu et al., 2024). When exposed to various stimuli, IκB gets phosphorylated by the IκB kinase complex, made up of IκB kinase (IKK) α, IKKβ, and the scaffolding protein NF-κB essential modulator, resulting in the release of NF-κB dimers. The dimers are subsequently transported into the nucleus and control the transcription of inflammatory cytokines like tumor necrosis factor α (TNF-α), interleukin-1 beta (IL-1β), interleukin (IL)-2, IL-6, IL-18, IL-12, inducible nitric oxide synthase (iNOS), and cyclooxygenase-2 (COX-2) (Guo et al., 2024). BAI or BE mediates anti-inflammatory effects by blocking NF-κB activation through multiple signal transductions such as Toll-like receptor 4 (TLR4), mitogen-activated protein kinase (MAPK), AMP-activated protein kinase (AMPK), and nuclear factor erythroid 2-related factor 2 (Nrf2)/heme oxygenase-1 (HO-1), ultimately exerting neuroprotective effects in NDDs.
4.2 TLR4 signaling
The TLR4 pathway has been implicated in neuroinflammation, which is a common manifestation of various CNS diseases (Adhikarla et al., 2021; Heidari et al., 2022). In vitro studies have shown that BAI inhibits microglial activation and the release of inflammatory factors induced by oxygen–glucose deprivation (OGD) or lipopolysaccharide (LPS). It additionally alters the regulation of associated proteins within the TLR4/MyD88/NF-κB signaling pathway. Interestingly, molecular docking analysis revealed that BAI binds favorably to the active site of TLR4-MD2, indicating a strong and stable interaction with the TLR4 receptor (Hou et al., 2012; Li B. et al., 2022). BAI inhibited neuroinflammation induced by microglia in vivo by blocking the TLR4/NF-κB pathway in amyloid beta precursor protein (APP)/presenilin-1 (PS1) mice (Jin et al., 2019). BAI triggered the TLR4/myeloid differentiation protein 88 (MyD88)/caspase-3 pathway to reduce neurodegeneration in the hippocampal CA3 area, while decreasing the levels of TLR4, NF-κB p65, iNOS, and COX-2 proteins and suppressing the secretion of TNF-α and IL-1β, which resulted in a protective effect on the nervous system (Tu et al., 2011; Yang et al., 2021). BE reduced the protein levels of TLR4, p-IκBα, and p-p65 in activated microglial models, hindering the translocation of NF-κB p65 from the cytoplasm to the nucleus and suppressing the expression of phosphorylated signal transducers and activators of transcription 1 (STAT1), which also contribute to the shift in microglial polarization toward an anti-inflammatory phenotype (Ran et al., 2021). The TLR4/MyD88/NF-κB pathway plays a role in CCH and is implicated in how BE prevents vascular dementia in rats (Song et al., 2024).
4.3 MAPK signaling
The MAPK signaling pathway plays a crucial role in controlling a variety of cellular functions such as cell proliferation, differentiation, and apoptosis. Emerging evidence has suggested a significant association between the dysregulation of the MAPK pathway and CNS disorders (Zheng et al., 2020; Tabaa et al., 2022; Khan et al., 2023). In the LPS-induced BV2 microglia model, Li et al. observed that BAI effectively inhibited LPS-induced phosphorylation of p38, extracellular signal-regulated kinase (ERK), and c-Jun N-terminal kinase (JNK), along with a reduction in the expression of neuroimmune mediators such as NO, prostaglandin E2 (PGE2), and IL-1β, suggesting that MAPK can be targeted by BAI (Li B. et al., 2022). In addition, BE treatment significantly inhibited p38, ERK 1/2, and JNK activation in the brains of PD rats, suggesting that BE can exert an anti-inflammatory effect in PD rats through the MAPK pathway (Zhang et al., 2017).
4.4 NLRP3 inflammasome signaling
The NOD-like receptor thermal protein domain-associated protein 3 (NLRP3) inflammasome, which includes NLRP3, apoptosis-associated speck-like protein containing a caspase recruitment domain (ASC), and caspase-1, breaks down inactive pro-IL-1β and pro-IL-18 to produce mature IL-1β and IL-18, respectively (Juliana et al., 2012; Yin et al., 2018). BAI decreased the activation of NLRP3 and production of IL-1β in the cortex of 3 × Tg-AD mice (Bitto et al., 2017), while BE decreased Aβ plaque accumulation and reduced NLRP3 inflammasome activation and neuronal cell death (Gong et al., 2023). The anti-inflammatory effect of BAI may be associated with its ability to inhibit the activation of the NLRP3 inflammasome, which occurs through the inhibition of NF-κB pathway activation—a preliminary step necessary for NLRP3 inflammasome activation (Zeng et al., 2017). Jin et al. demonstrated that BAI pretreatment significantly inhibited LPS/Aβ-induced elevation of p-IκBα expression and reduction in IκBα protein expression in BV2 microglial cells and simultaneously downregulated NLRP3 protein expression and inhibited caspase-1 activity, thereby reducing IL-1β and IL-18 levels (Jin et al., 2019). In PD, BAI showed a dose-dependent inhibition of the excessive phosphorylation of NF-κB p65 and the upregulation of NLRP3 inflammasomes, along with their resulting caspase-1 and IL-1β (Huang et al., 2024). In contrast, BE alleviated 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-triggered neuroinflammation by inhibiting the NLRP3/caspase-1/gasdermin D (GSDMD) pathway (Rui et al., 2020). BAI treatment also enhanced the neuronal antioxidant capacity (Huang et al., 2021), which is associated with Nrf2/HO-1 activation (Ding et al., 2015; Li X. et al., 2022). The antioxidant effects mediated by Nrf2 are essential for the inhibition of NLRP3 inflammasome signaling by BE (Huang et al., 2024).
4.5 AMPK signaling
In various CNS disorders, the aberrant regulation of AMPK signaling has been implicated in the modulation of neuroinflammation (Chen et al., 2021a; Wang et al., 2022; Steinberg and Hardie, 2023; Wang et al., 2023). AMPK activation reduces the inflammatory response by blocking the NF-κB pathway (Jie et al., 2022). In PD, BE elevates AMPK phosphorylation and inhibits the mammalian target of rapamycin (mTOR) activity, whereas miR-30b-5p overexpression and sirtuin 1 (SIRT1) silencing partially abolish the function of BE in regulating the AMPK/mTOR pathway (Chen et al., 2021b). In addition, BAI attenuates neurological damage following OGD/reperfusion by inhibiting NLRP3 inflammasome activity through enhanced AMPK phosphorylation (Zheng et al., 2021; Li et al., 2017), while the inhibition of the AMPK/Nrf2 pathway may also contribute to BE’s neuroprotective mechanisms (Yuan et al., 2020)
4.6 Other possible mechanisms
The alteration of K (+)-Cl (−) co-transporter and Na-K-2Cl cotransporter-1 expression by BAI affects GABAergic signaling, while the enhancement of heat shock protein expression may also contribute to the neuroprotective mechanisms of BAI (Dai et al., 2013). The regulation of BE on microglial polarization is essential for suppressing neuroinflammation and nerve damage in AD by targeting the heme oxygenase 1/phosphodiesterase 4D axis (Gong et al., 2023).
5 The effects of BE and BAI on neuroinflammation in NDDs
Extensive research has indicated a close association between neuroinflammation and various neurodegenerative pathways (Marogianni et al., 2020; Scheltens et al., 2021; Araujo et al., 2022; Zhang W. et al., 2023). The neuroprotective properties of BAI and BE appear to stem from their anti-inflammatory characteristics (Pan et al., 2021) (Table 1).
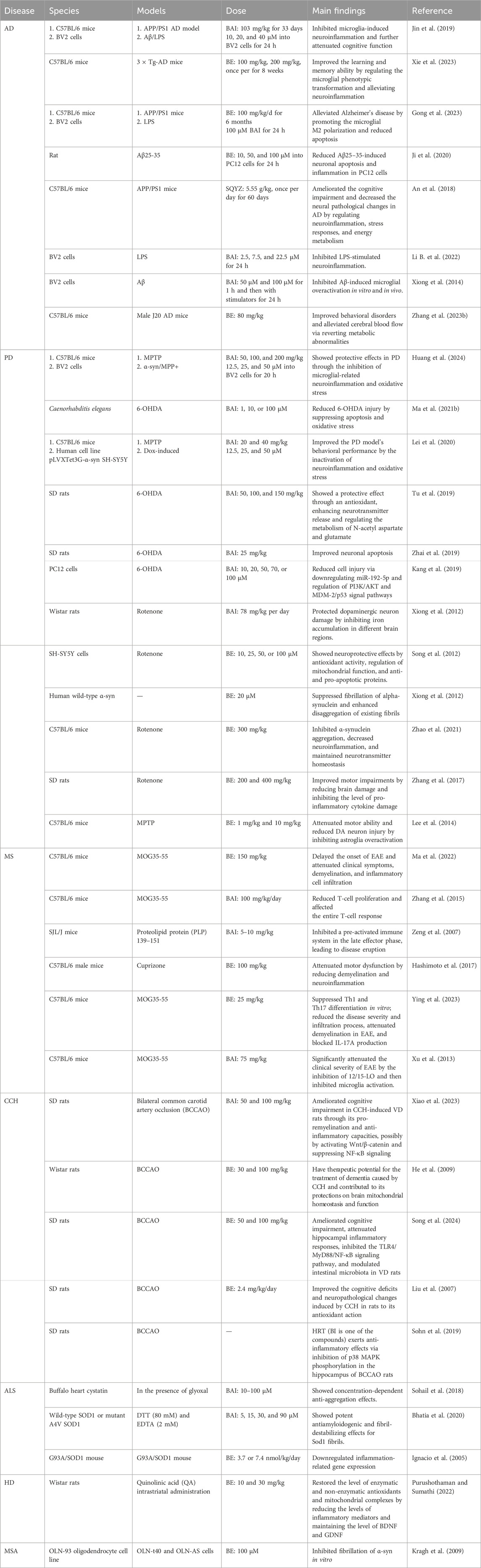
Table 1. BAI and BE show protective effects on NDDs in vivo and in vitro.
5.1 BAI, BE, and AD
AD is the most common neurodegenerative disorder, characterized by significant pathological changes in the brain, including the accumulation of Aβ plaques and neurofibrillary tangles formed hyperphosphorylated tau protein (Scheltens et al., 2021; Thakur et al., 2023). The presence of Aβ plaques and tau tangles leads to chronic neuroinflammation, which exacerbates neuronal damage and cognitive decline (Leng and Edison, 2021). TLR4, NF-κB, MAPK, and the NLRP3 inflammasome are key components in triggering microglial-related neuroinflammation. TLR4 facilitates the recognition of pathological stimuli and then initiates NF-κB and MAPK pathways, which propagate pro-inflammatory signaling (Wu et al., 2022). The NLRP3 inflammasome can be activated by the TLR4/MyD88/NF-κB signaling pathway or directly by oligomers and fiber Aβ, amplifying cytokine release and perpetuating a cycle of neuroinflammation (Yang et al., 2020; Milner et al., 2021). Targeting the inflammation-related signaling pathway could suppress microglial activation, reduce pathogenesis, and improve learning and memory functions, which may be effective therapeutic strategies for AD.
Both BAI and BE exhibit beneficial effects in AD by modulating neuroinflammation. In an Aβ-induced cell model, BAI attenuates Aβ-mediated microglial inflammatory responses and neuroinflammation-associated neuronal apoptosis (Jin et al., 2019; Xiong et al., 2014). Similar effects have been observed with BE, which can alleviate Aβ25–35-stimulated neuronal apoptosis and inflammation (Ji et al., 2020). In addition, BAI significantly alleviates LPS-induced neuroinflammation by suppressing the expression of miR-155, regulating the TLR4/MyD88/NF-κB pathway and MAPK pathway (Li B. et al., 2022).
Furthermore, BAI and BE have demonstrated notable neuroprotective effects in various animal models of AD. BAI treatment attenuated spatial memory dysfunction in APP/PS1 mice by suppressing microglial overactivation (Jin et al., 2019). In a J20 APPSwInd transgenic (Tg) mouse model of AD, BE inhibited hyperactivity and improved spatial learning ability. Metabolic profiling of specific brain regions indicated that BE regulates neuroinflammation, which is associated with the modulation of starch, sucrose, and glycolipid metabolism in the cortex and hippocampus (Zhang L. et al., 2023). Additionally, SQYZ granules, a Chinese herbal preparation containing BAI, ameliorated neural pathological changes in AD, possibly by modulating anti-neuroinflammation, promoting stress recovery, and enhancing energy metabolism in APP/PS1 mice (An et al., 2018).
Moreover, BE and BAI mitigated neuroinflammation by modulating the phenotypic transformation of activated microglia. BE improved cognitive dysfunction in 3 × Tg-AD mice by enhancing M2 microglial polarization (Xie et al., 2023). Furthermore, in both APP/PS1 double-transgenic mice and LPS-stimulated BV2 microglia, BE significantly shifts the microglial phenotype from M1 to M2 (Gong et al., 2023). Moreover, in LPS/interferon γ-induced neuroinflammation, BAI promoted the polarization of microglia from the M1 phenotype to M2 phenotype (He et al., 2022). In conclusion, these studies clearly demonstrated that the protective effects of BAI and BE against neuronal damage in AD are related to the inhibition of neuroinflammation.
5.2 BAI, BE, and PD
PD is a complex neurodegenerative disorder characterized by several pathological manifestations, including the aggregation of α-synuclein, loss of dopaminergic neurons in the nigrostriatal system, and heightened neuroinflammation. The presence of protein aggregates and damaged neurons in PD can initiate an inflammatory response (Gao et al., 2024). Activated microglia release pro-inflammatory cytokines such as IL-1β, IL-6, and TNF-α. These cytokines can further damage neurons and exacerbate the disease process through autocrine and paracrine mechanisms. Additionally, neuroinflammation and oxidative stress can trigger the activation of JNK and p38 MAPK pathways, both of which are involved in exacerbating the pathological progression of PD (Gravandi et al., 2023; Gogna et al., 2024). Currently, dopamine replacement therapy remains the primary treatment approach for PD. However, it is important to note that although these methods are widely used, they may not be universally effective or suitable for all patients.
In a previous investigation using a Caenorhabditis elegans model of PD, it was discovered that BAI enhanced reversal and omega turn behavioral characteristics, along with increasing survival rates. BAI provided protection against 6-hydroxydopamine (6-OHDA) damage by preventing cell death and decreasing oxidative damage (Ma J. et al., 2021). Other in vitro experiments have demonstrated the neuroprotective effects of BAI in PD. BAI inhibited the inflammatory response caused by α-syn/1-methyl-4-phenylpyridinium (MPP+) in BV2 cells (Huang et al., 2024) and reduced neurotoxicity induced by 6-OHDA in PC12 cells (Kang et al., 2019). Additionally, BAI and BE reportedly increase the viability and reduces cell death in dopaminergic SH-SY5Y cells (Song et al., 2012; Lei et al., 2020). Numerous animal models have confirmed the neuroprotective effects of BAI against PD. BAI demonstrated a notable defense mechanism in PD rats induced by 6-OHDA, possibly through antioxidant actions, enhanced neurotransmitter release, and control of N-acetyl aspartate and glutamate metabolism. BAI attenuated substantia nigra neuronal apoptosis in PD rats (Tu et al., 2019; Zhai et al., 2019). Moreover, BAI demonstrates neuroprotective effects in MPTP-induced PD mice by inhibiting pro-inflammatory cytokine expression and reducing oxidative stress. BE also has exhibited a strong protective effect against PD (Lei et al., 2020; Huang et al., 2024). In PD, BE has the potential to modulate pathways mediated by α-syn beyond directly targeting α-syn itself (Li et al., 2023). BE attenuates iron accumulation and iron-induced oxidative stress in the brain of PD rats (Liu et al., 2022). BE has the potential to interfere with both wild-type and E46K/H50Q mutant α-syn fibrils, potentially mitigating PD progression (Yao et al., 2022). Treatment with BE decreases the accumulation of α-syn, prevents inflammation in the brain, balances neurotransmitter levels, and decreases the release of inflammatory molecules like TNF-α and IL-6, all while regulating the activity of astrocytes and microglia in rats with PD (Lee et al., 2014; Zhang et al., 2017; Zhao et al., 2021). These results indicate that BAI and BE have beneficial therapeutic effects in PD, possibly attributed to their anti-inflammatory, antioxidant, and other related actions.
5.3 BAI, BE, and MS
MS is a chronic autoimmune demyelinating disease affecting the central nervous system. Several cytokines play crucial roles in MS, with interferon-gamma (IFN-γ) being produced by activated T cells. IFN-γ binds to its receptor on various central nervous system cells, including astrocytes and microglia, activating the Janus kinase (JAK)–STAT signaling pathway (Kooshki et al., 2023). The associated JAKs are phosphorylated upon receptor activation, which, in turn, phosphorylates STAT proteins. These STAT proteins then dimerize and translocate to the nucleus, regulating the expression of genes related to immune responses and inflammation (Benucci et al., 2023). Additionally, TNF-α plays a significant role by binding to its receptors and activating multiple intracellular pathways, including the NF-κB pathway, which further enhances the expression of pro-inflammatory genes.
Studies of MS have demonstrated that BAI reduces the entry of immune cells into the CNS, suppresses the production of inflammatory substances and chemokines, and hinders the formation of Th1 and Th17 cells (Zhang et al., 2015). Furthermore, BAI can inhibit the development and progression of experimental autoimmune encephalomyelitis (EAE), which is an animal model that has a significant relationship with MS (Zeng et al., 2007). The demyelination in EAE is caused by an autoimmune response that is artificially induced and shares similarities with the unknown trigger of MS demyelination (Manogaran et al., 2018). BE also blocks the activation of microglia/macrophages toward the M1 phenotype in EAE mice by focusing on STAT 1 (Ma et al., 2022) and reduces cuprizone-induced demyelination by suppressing neuroinflammation (Hashimoto et al., 2017). Additional research has indicated that BE can improve EAE by reducing pathogenic C-X-C motif chemokine receptor 6 (CXCR6)+ CD4 cells (Ying et al., 2023) or blocking 12/15-lipoxygenase (Xu et al., 2013). These findings emphasize the importance of exploring the potential roles of BAI and BE in improving therapeutic strategies for MS.
5.4 BAI, BE, and CCH
CCH is characterized by a persistent reduction in cerebral blood flow, which often develops gradually and can significantly impact brain function. This condition activates the MAPK pathway, involving various members of the MAPK family, including ERK, JNK, and p38 MAPK (Zhang et al., 2020). The activation of these kinases is triggered by stimuli associated with the ischemic environment, such as oxidative stress and inflammatory cytokines. In the context of chronic cerebral ischemia, NF-κB is also activated. ROS generated during ischemia, along with other inflammatory mediators, stimulate the activation of the IKK complex (Su et al., 2017).
In addition, numerous studies have demonstrated that CCH is a major contributor to neurodegenerative processes (Daulatzai, 2017). Recent studies have shown that BAI exerts neuroprotective effects against chronic brain ischemia. BAI ameliorates cognitive impairment in CCH-induced VD rats through its pro-remyelination and anti-inflammatory abilities, possibly by activating Wnt/β-catenin and suppressing NF-κB signaling (Xiao et al., 2023). BE alleviates cognitive and motor impairments by decreasing mitochondria reactive oxygen species production (He et al., 2009) and inhibiting the TLR4/MyD88/NF-κB signaling pathway in CCH rats (Song et al., 2024; Liu et al., 2007). Traditional Chinese medicine—Hwangryunhaedok-Tang—containing BAI and BE improves cholinergic dysfunction and inhibits neuroinflammatory responses in CCH rats (Sohn et al., 2019). These results indicate that BAI has a beneficial therapeutic effect against CCH-induced brain injury through its anti-inflammatory properties.
5.5 BAI, BE, and other NDDs
ALS is a severe and progressive neurodegenerative disorder primarily affecting the motor neurons in the brain and spinal cord (Manogaran et al., 2018). HD is an inherited, progressive neurodegenerative disorder that significantly impacts both the physical and mental health of those affected (Gharaba et al., 2024). MSA is a rare and progressive neurodegenerative disorder that impacts multiple systems in the body (Ndayisaba et al., 2024). The MAPK and NF-κB signaling pathways play crucial roles in the pathophysiology of these NDDs.
BAI exhibits concentration-dependent anti-aggregation effects linked to ALS (Sohail et al., 2018). In ALS mouse models, the impact on mutant SOD1 was more pronounced than that on wild-type SOD1, affecting fibril elongation (Bhatia et al., 2020; Ignacio et al., 2005). BE also mitigates the psychological and behavioral changes induced by quinolinic acid in an HD mouse model (Purushothaman and Sumathi, 2022). BE effectively decreases the number of cells exhibiting microtubular retraction and suppresses the aggregation of α-syn in the MSA model (Kragh et al., 2009). In conclusion, BAI and BE demonstrate the potential to ameliorate NDD-associated symptoms.
6 Conclusion and future perspective
NDDs have a profound impact on human health, primarily characterized by the progressive loss of neuronal function and structure, leading to cognitive decline and physical disabilities. Current therapeutic strategies for these disorders are limited, with most treatments focusing on alleviating symptoms rather than addressing the underlying pathology.
Extensive evidence has demonstrated the potent anti-inflammatory and neuroprotective properties of BE and BAI in both in vitro and in vivo models of various NDDs mediated through the activation of multiple signaling pathways. In recent years, two phase-I clinical trials involving BE chewable tablets were completed in healthy Chinese adults, demonstrating that oral BE administration was safe and well-tolerated in healthy subjects (Pang et al., 2016). In a randomized, double-blind, placebo-controlled trial of BAI in patients with coronary heart disease (CAD) and rheumatoid arthritis (RA), it was found that BAI reduces blood lipids and inflammation in patients with both CAD and RA (Hang et al., 2018). BAI affects the innate immunity and apoptosis in leukocytes of children with acute lymphocytic leukemia (Orzechowska et al., 2014). A clinical study on BAI in patients with ulcerative colitis (UC) demonstrated that BAI can balance immune function and alleviate inflammation associated with UC (Yu et al., 2015). Additionally, several clinical trials have investigated BAI or herbal formulations containing BAI in patients with mild–moderate photo-damaged skin, non-surgical periodontal therapy, or after the surgical removal of impacted mandibular third molars (Farris et al., 2014; Isola et al., 2021; Isola et al., 2019). However, there remains a lack of targeted clinical research specifically investigating the effects of BAI and BE on NDDs.
To fully explore the therapeutic potential of BAI and BE, future clinical trials should focus on assessing their efficacy in patients with neurodegenerative disorders. Furthermore, additional research is required to characterize their specific molecular targets or receptors, which is essential for understanding their pharmacological mechanisms, optimizing drug design, and advancing drug development. Given the limited water and lipid solubility of BAI and BE, developing innovative delivery systems—such as nanoparticles or other advanced carriers—will be crucial to enhance bioactivity, improve blood–brain barrier permeability, and increase clinical efficacy (Zhou et al., 2017).
Despite these challenges, accumulating evidence highlights the value of BAI and BE as promising natural compounds. Their therapeutic potential merits continued investigation and exploration, offering hope for new treatment options in the medical and healthcare fields.
Author contributions
RY: conceptualization, funding acquisition, writing–original draft, and writing–review and editing. RW: writing–original draft and writing–review and editing. AX: funding acquisition and writing–review and editing. JZ: writing–review and editing. JM: funding acquisition, writing–original draft, and writing–review and editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This work was funded by the Natural Science Foundation of Shanghai (No. 18ZR1424900), the National Natural Science Foundation of China (No. 82072142), the China Scholarship Council (No. 202206235019), KC Wang Education Foundation (No. 2023-21), Youth Medical Talents–Clinical Pharmacist Program 2022 (No. SHWRS [2022-65]), and the National Key Research and Development Program of China (No. 2020YFC2005502).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Adhikarla, S. V., Jha, N. K., Goswami, V. K., Sharma, A., Bhardwaj, A., Dey, A., et al. (2021). Tlr-mediated signal transduction and neurodegenerative disorders. Brain Sci. 11 (11), 1373. Epub 2021/11/28. doi:10.3390/brainsci11111373
Akao, T., Kawabata, K., Yanagisawa, E., Ishihara, K., Mizuhara, Y., Wakui, Y., et al. (2000). Baicalin, the predominant flavone glucuronide of scutellariae radix, is absorbed from the rat gastrointestinal tract as the aglycone and restored to its original form. J. Pharm. Pharmacol. 52 (12), 1563–1568. doi:10.1211/0022357001777621
Akao, T., Sato, K., and Hanada, M. (2009). Hepatic contribution to a marked increase in the plasma concentration of baicalin after oral administration of its aglycone, baicalein, in multidrug resistance-associated protein 2-deficient rat. Biol. Pharm. Bull. 32 (12), 2079–2082. doi:10.1248/bpb.32.2079
An, H., Wei, D., Qian, Y., Li, N., and Wang, X. (2018). Sqyz granules, a traditional Chinese herbal, attenuate cognitive deficits in Ad transgenic mice by modulating on multiple pathogenesis processes. Am. J. Transl. Res. 10 (11), 3857–3875. Epub 20181115. Available at: https://pmc.ncbi.nlm.nih.gov/articles/PMC6291719/
Araujo, B., Caridade-Silva, R., Soares-Guedes, C., Martins-Macedo, J., Gomes, E. D., Monteiro, S., et al. (2022). Neuroinflammation and Parkinson’s disease-from neurodegeneration to therapeutic opportunities. Cells 11 (18), 2908. Epub 2022/09/24. doi:10.3390/cells11182908
Benucci, M., Bernardini, P., Coccia, C., De Luca, R., Levani, J., Economou, A., et al. (2023). Jak inhibitors and autoimmune rheumatic diseases. Autoimmun. Rev. 22 (4), 103276. Epub 2023/01/18. doi:10.1016/j.autrev.2023.103276
Bhatia, N. K., Modi, P., Sharma, S., and Deep, S. (2020). Quercetin and baicalein act as potent antiamyloidogenic and fibril destabilizing agents for Sod1 fibrils. ACS Chem. Neurosci. 11 (8), 1129–1138. Epub 20200406. doi:10.1021/acschemneuro.9b00677
Bitto, A., Giuliani, D., Pallio, G., Irrera, N., Vandini, E., Canalini, F., et al. (2017). Effects of Cox1-2/5-Lox blockade in Alzheimer transgenic 3xtg-Ad mice. Inflamm. Res. 66 (5), 389–398. Epub 2017/02/27. doi:10.1007/s00011-017-1022-x
Calió, M. L., Henriques, E., Siena, A., Bertoncini, C. R. A., Gil-Mohapel, J., and Rosenstock, T. R. (2020). Mitochondrial dysfunction, neurogenesis, and epigenetics: putative implications for amyotrophic lateral sclerosis neurodegeneration and treatment. Front. Neurosci. 14, 679. Epub 20200715. doi:10.3389/fnins.2020.00679
Chen, M., Huang, N., Liu, J., Huang, J., Shi, J., and Jin, F. (2021a). Ampk: a bridge between diabetes mellitus and Alzheimer’s disease. Behav. Brain Res. 400, 113043. Epub 2020/12/12. doi:10.1016/j.bbr.2020.113043
Chen, M., Peng, L., Gong, P., Zheng, X., Sun, T., Zhang, X., et al. (2021b). Baicalein induces mitochondrial autophagy to prevent Parkinson’s disease in rats via Mir-30b and the Sirt1/Ampk/Mtor pathway. Front. Neurol. 12, 646817. Epub 2022/03/04. doi:10.3389/fneur.2021.646817
Chen, S., Yang, Y., Feng, H., Wang, H., Zhao, R., and Liu, H. (2014). Baicalein inhibits interleukin-1β-induced proliferation of human rheumatoid arthritis fibroblast-like synoviocytes. Inflammation 37 (1), 163–169. Epub 2013/09/06. doi:10.1007/s10753-013-9725-9
Chen, X., Zhou, Y., Wang, S., and Wang, W. (2022). Mechanism of baicalein in brain injury after intracerebral hemorrhage by inhibiting the Ros/Nlrp3 inflammasome pathway. Inflammation 45 (2), 590–602. Epub 2021/10/10. doi:10.1007/s10753-021-01569-x
Dai, J., Chen, L., Qiu, Y. M., Li, S. Q., Xiong, W. H., Yin, Y. H., et al. (2013). Activations of gabaergic signaling, Hsp70 and Mapk cascades are involved in Baicalin's neuroprotection against gerbil global ischemia/reperfusion injury. Brain Res. Bull. 90, 1–9. Epub 2012/10/09. doi:10.1016/j.brainresbull.2012.09.014
Daulatzai, M. A. (2017). Cerebral hypoperfusion and glucose hypometabolism: key pathophysiological modulators promote neurodegeneration, cognitive impairment, and Alzheimer's disease. J. Neurosci. Res. 95 (4), 943–972. Epub 20160627. doi:10.1002/jnr.23777
Day, A. J., Gee, J. M., DuPont, M. S., Johnson, I. T., and Williamson, G. (2003). Absorption of quercetin-3-glucoside and quercetin-4’-glucoside in the rat small intestine: the role of lactase phlorizin hydrolase and the sodium-dependent glucose transporter. Biochem. Pharmacol. 65 (7), 1199–1206. doi:10.1016/s0006-2952(03)00039-x
Dinda, B., Dinda, S., DasSharma, S., Banik, R., Chakraborty, A., and Dinda, M. (2017). Therapeutic potentials of baicalin and its aglycone, baicalein against inflammatory disorders. Eur. J. Med. Chem. 131, 68–80. Epub 20170306. doi:10.1016/j.ejmech.2017.03.004
Ding, H., Wang, H., Zhao, Y., Sun, D., and Zhai, X. (2015). Protective effects of baicalin on Aβ₁₋₄₂-induced learning and memory deficit, oxidative stress, and apoptosis in rat. Cell Mol. Neurobiol. 35 (5), 623–632. Epub 2015/01/19. doi:10.1007/s10571-015-0156-z
Farris, P., Yatskayer, M., Chen, N., Krol, Y., and Oresajo, C. (2014). Evaluation of efficacy and tolerance of a nighttime topical antioxidant containing resveratrol, baicalin, and vitamin E for treatment of mild to moderately photodamaged skin. J. Drugs Dermatol 13 (12), 1467–1472. Epub 2015/01/22. Available at: https://pubmed.ncbi.nlm.nih.gov/25607790/
Gaire, B. P., Moon, S. K., and Kim, H. (2014). Scutellaria baicalensis in stroke management: nature’s blessing in traditional eastern medicine. Chin. J. Integr. Med. 20 (9), 712–720. Epub 20140422. doi:10.1007/s11655-014-1347-9
Gao, C., Jiang, J., Tan, Y., and Chen, S. (2023). Microglia in neurodegenerative diseases: mechanism and potential therapeutic targets. Signal Transduct. Target Ther. 8 (1), 359. Epub 20230922. doi:10.1038/s41392-023-01588-0
Gao, Y., Zhang, J., Tang, T., and Liu, Z. (2024). Hypoxia pathways in Parkinson's disease: from pathogenesis to therapeutic targets. Int. J. Mol. Sci. 25 (19), 10484. Epub 2024/10/16. doi:10.3390/ijms251910484
Gharaba, S., Sprecher, U., Baransi, A., Muchtar, N., and Weil, M. (2024). Characterization of fission and fusion mitochondrial dynamics in hd fibroblasts according to patient's severity status. Neurobiol. Dis. 201, 106667. Epub 2024/09/17. doi:10.1016/j.nbd.2024.106667
Gogna, T., Housden, B. E., and Houldsworth, A. (2024). Exploring the role of reactive oxygen species in the pathogenesis and pathophysiology of Alzheimer's and Parkinson's disease and the efficacy of antioxidant treatment. Antioxidants (Basel) 13 (9), 1138. Epub 2024/09/28 22:43. doi:10.3390/antiox13091138
Gong, Q., Wang, Y., Wang, X., Pan, H., and Yan, C. (2023). Baicalein promotes the microglia M2 polarization and suppresses apoptosis by targeting Hmox1/Pde4d to alleviate Alzheimer's disease. Immunobiology 228 (6), 152761. Epub 20231117. doi:10.1016/j.imbio.2023.152761
Gravandi, M. M., Abdian, S., Tahvilian, M., Iranpanah, A., Moradi, S. Z., Fakhri, S., et al. (2023). Therapeutic targeting of Ras/Raf/Mapk pathway by natural products: a systematic and mechanistic approach for neurodegeneration. Phytomedicine 115, 154821. Epub 2023/04/30. doi:10.1016/j.phymed.2023.154821
Guo, Q., Jin, Y., Chen, X., Ye, X., Shen, X., Lin, M., et al. (2024). NF-κB in biology and targeted therapy: new insights and translational implications. Signal Transduct. Target Ther. 9 (1), 53. Epub 2024/03/04. doi:10.1038/s41392-024-01757-9
Hang, Y., Qin, X., Ren, T., and Cao, J. (2018). Baicalin reduces blood lipids and inflammation in patients with coronary artery disease and rheumatoid arthritis: a randomized, double-blind, placebo-controlled trial. Lipids Health Dis. 17 (1), 146. Epub 2018/06/25. doi:10.1186/s12944-018-0797-2
Hashimoto, M., Yamamoto, S., Iwasa, K., Yamashina, K., Ishikawa, M., Maruyama, K., et al. (2017). The flavonoid baicalein attenuates cuprizone-induced demyelination via suppression of neuroinflammation. Brain Res. Bull. 135, 47–52. Epub 20170915. doi:10.1016/j.brainresbull.2017.09.007
He, C. X., Yu, W. J., Yang, M., Li, Z., Xia, X. F., Li, P., et al. (2022). Baicalin inhibits Lps/Ifn-Γ-induced inflammation via Trem2/Tlr4/Nf-Κb pathway in Bv2 cells. Zhongguo Zhong Yao Za Zhi 47 (6), 1603–1610. doi:10.19540/j.cnki.cjcmm.20211103.401
He, X. L., Wang, Y. H., Gao, M., Li, X. X., Zhang, T. T., and Du, G. H. (2009). Baicalein protects rat brain mitochondria against chronic cerebral hypoperfusion-induced oxidative damage. Brain Res. 1249, 212–221. Epub 20081018. doi:10.1016/j.brainres.2008.10.005
Heidari, A., Yazdanpanah, N., and Rezaei, N. (2022). The role of toll-like receptors and neuroinflammation in Parkinson’s disease. J. Neuroinflammation 19 (1), 135. Epub 2022/06/07. doi:10.1186/s12974-022-02496-w
Hou, J., Wang, J., Zhang, P., Li, D., Zhang, C., Zhao, H., et al. (2012). Baicalin attenuates proinflammatory cytokine production in oxygen-glucose deprived challenged rat microglial cells by inhibiting Tlr4 signaling pathway. Int. Immunopharmacol. 14 (4), 749–757. Epub 2012/11/03. doi:10.1016/j.intimp.2012.10.013
Huang, H., Zhang, Y., Yang, R., and Tang, X. (2008). Determination of baicalin in rat cerebrospinal fluid and blood using microdialysis coupled with ultra-performance liquid chromatography-tandem mass spectrometry. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 874 (1-2), 77–83. Epub 20080910. doi:10.1016/j.jchromb.2008.09.005
Huang, J., Zhang, X., Yang, X., Yv, Q., Ye, F., Chen, S., et al. (2024). Baicalin exerts neuroprotective actions by regulating the Nrf2-Nlrp3 Axis in toxin-induced models of Parkinson’s disease. Chem. Biol. Interact. 387, 110820. Epub 20231126. doi:10.1016/j.cbi.2023.110820
Huang, Z., Guo, L., Huang, L., Shi, Y., Liang, J., and Zhao, L. (2021). Baicalin-loaded macrophage-derived exosomes ameliorate ischemic brain injury via the antioxidative pathway. Mater Sci. Eng. C Mater Biol. Appl. 126, 112123. Epub 2021/06/05. doi:10.1016/j.msec.2021.112123
Ignacio, S., Moore, D. H., Smith, A. P., and Lee, N. M. (2005). Effect of neuroprotective drugs on gene expression in G93a/Sod1 mice. Ann. N. Y. Acad. Sci. 1053, 121–136. doi:10.1196/annals.1344.010
Isola, G., Matarese, M., Ramaglia, L., Iorio-Siciliano, V., Cordasco, G., and Matarese, G. (2019). Efficacy of a drug composed of herbal extracts on postoperative discomfort after surgical removal of impacted mandibular third molar: a randomized, triple-blind, controlled clinical trial. Clin. Oral Investig. 23 (5), 2443–2453. Epub 2018/10/13. doi:10.1007/s00784-018-2690-9
Isola, G., Polizzi, A., Iorio-Siciliano, V., Alibrandi, A., Ramaglia, L., and Leonardi, R. (2021). Effectiveness of a nutraceutical agent in the non-surgical periodontal therapy: a randomized, controlled clinical trial. Clin. Oral Investig. 25 (3), 1035–1045. Epub 2020/06/20. doi:10.1007/s00784-020-03397-z
Ji, Y., Han, J., Lee, N., Yoon, J. H., Youn, K., Ha, H. J., et al. (2020). Neuroprotective effects of baicalein, wogonin, and oroxylin a on amyloid beta-induced toxicity via Nf-Κb/Mapk pathway modulation. Molecules 25 (21), 5087. doi:10.3390/molecules25215087
Jie, F., Yang, X., Yang, B., Liu, Y., Wu, L., and Lu, B. (2022). Stigmasterol attenuates inflammatory response of microglia via NF-κB and NLRP3 signaling by AMPK activation. Biomed. Pharmacother. 153, 113317. Epub 2022/07/01. doi:10.1016/j.biopha.2022.113317
Jin, X., Liu, M. Y., Zhang, D. F., Zhong, X., Du, K., Qian, P., et al. (2019). Baicalin mitigates cognitive impairment and protects neurons from microglia-mediated neuroinflammation via suppressing Nlrp3 inflammasomes and Tlr4/Nf-Κb signaling pathway. CNS Neurosci. Ther. 25 (5), 575–590. Epub 20190124. doi:10.1111/cns.13086
Juliana, C., Fernandes-Alnemri, T., Kang, S., Farias, A., Qin, F., and Alnemri, E. S. (2012). Non-transcriptional priming and deubiquitination regulate Nlrp3 inflammasome activation. J. Biol. Chem. 287 (43), 36617–36622. Epub 2012/09/06. doi:10.1074/jbc.M112.407130
Kang, C., Wang, L., Kang, M., Liu, X., Fu, Y., and Gao, J. (2019). Baicalin alleviates 6-hydroxydopamine-induced neurotoxicity in Pc12 cells by down-regulation of microrna-192-5p. Brain Res. 1708, 84–92. Epub 20181212. doi:10.1016/j.brainres.2018.12.015
Khan, A., Shal, B., Khan, A. U., Bibi, T., Zeeshan, S., Zahra, S. S., et al. (2023). Suppression of Mapk/Nf-Kb and activation of Nrf2 signaling by ajugarin-I in eae model of multiple sclerosis. Phytother. Res. 37 (6), 2326–2343. Epub 2023/02/16. doi:10.1002/ptr.7751
Kooshki, L., Zarneshan, S. N., Fakhri, S., Moradi, S. Z., and Echeverria, J. (2023). The pivotal role of Jak/Stat and Irs/Pi3k signaling pathways in neurodegenerative diseases: mechanistic approaches to polyphenols and alkaloids. Phytomedicine 112, 154686. Epub 2023/02/23. doi:10.1016/j.phymed.2023.154686
Kragh, C. L., Lund, L. B., Febbraro, F., Hansen, H. D., Gai, W. P., El-Agnaf, O., et al. (2009). Alpha-synuclein aggregation and Ser-129 phosphorylation-dependent cell death in oligodendroglial cells. J. Biol. Chem. 284 (15), 10211–10222. Epub 20090209. doi:10.1074/jbc.M809671200
Lai, M. Y., Hsiu, S. L., Chen, C. C., Hou, Y. C., and Chao, P. D. (2003). Urinary pharmacokinetics of baicalein, wogonin and their glycosides after oral administration of scutellariae radix in humans. Biol. Pharm. Bull. 26 (1), 79–83. doi:10.1248/bpb.26.79
Lee, E., Park, H. R., Ji, S. T., Lee, Y., and Lee, J. (2014). Baicalein attenuates astroglial activation in the 1-methyl-4-phenyl-1,2,3,4-tetrahydropyridine-induced Parkinson's disease model by downregulating the activations of nuclear factor-κB, ERK, and JNK. J. Neurosci. Res. 92 (1), 130–139. Epub 20131028. doi:10.1002/jnr.23307
Lei, K. C., Shen, Y. J., He, Y. J., Zhang, L. W., Zhang, J. X., Tong, W. F., et al. (2020). Baicalin represses C/EBPβ via its antioxidative effect in Parkinson's disease. Oxidative Med. Cell. Longev. 2020, 8951907. doi:10.1155/2020/8951907
Leng, F., and Edison, P. (2021). Neuroinflammation and microglial activation in Alzheimer disease: where do we go from here? Nat. Rev. Neurol. 17 (3), 157–172. Epub 2020/12/16. doi:10.1038/s41582-020-00435-y
Li, B., Wang, M., Chen, S., Li, M., Zeng, J., Wu, S., et al. (2022). Baicalin mitigates the neuroinflammation through the Tlr4/Myd88/Nf-Κb and Mapk pathways in Lps-stimulated bv-2 microglia. Biomed. Res. Int. 2022, 3263446. Epub 20221109. doi:10.1155/2022/3263446
Li, C., Lin, G., and Zuo, Z. (2011). Pharmacological effects and pharmacokinetics properties of radix scutellariae and its bioactive flavones. Biopharm. Drug Dispos. 32 (8), 427–445. Epub 20110919. doi:10.1002/bdd.771
Li, Q., and Barres, B. A. (2018). Microglia and macrophages in brain homeostasis and disease. Nat. Rev. Immunol. 18 (4), 225–242. Epub 20171120. doi:10.1038/nri.2017.125
Li, S., Sun, X., Xu, L., Sun, R., Ma, Z., Deng, X., et al. (2017). Baicalin attenuates in vivo and in vitro hyperglycemia-exacerbated ischemia/reperfusion injury by regulating mitochondrial function in a manner dependent on ampk. Eur. J. Pharmacol. 815, 118–126. Epub 2017/07/27. doi:10.1016/j.ejphar.2017.07.041
Li, X., Deng, Q., Kuang, Y., Mao, H., Yao, M., Lin, C., et al. (2023). Identifying NFKB1, STAT3, and CDKN1A as baicalein's potential hub targets in Parkinson's disease-related α-synuclein-mediated pathways by integrated bioinformatics strategies. Curr. Pharm. Des. 29 (30), 2426–2437. doi:10.2174/0113816128259065231011114116
Li, X., Li, S., Ma, C., Li, T., and Yang, L. (2022). Preparation of baicalin-loaded ligand-modified nanoparticles for nose-to-brain delivery for neuroprotection in cerebral ischemia. Drug Deliv. 29 (1), 1282–1298. Epub 2022/04/26. doi:10.1080/10717544.2022.2064564
Liang, W., Huang, X., and Chen, W. (2017). The effects of baicalin and baicalein on cerebral ischemia: a review. Aging Dis. 8 (6), 850–867. Epub 20171201. doi:10.14336/AD.2017.0829
Liu, C., Wu, J., Gu, J., Xiong, Z., Wang, F., Wang, J., et al. (2007). Baicalein improves cognitive deficits induced by chronic cerebral hypoperfusion in rats. Pharmacol. Biochem. Behav. 86 (3), 423–430. Epub 2007/02/10. doi:10.1016/j.pbb.2006.11.005
Liu, R. Z., Zhang, S., Zhang, W., Zhao, X. Y., and Du, G. H. (2022). Baicalein attenuates brain iron accumulation through protecting Aconitase 1 from oxidative stress in rotenone-induced Parkinson's disease in rats. Antioxidants (Basel) 12 (1), 12. Epub 20221221. doi:10.3390/antiox12010012
Ma, J., Wang, R., Chen, T., Jiang, S., and Xu, A. (2021b). Protective effects of baicalin in a Caenorhabditis elegans model of Parkinson’s disease. Toxicol. Res. (Camb) 10 (3), 409–417. Epub 20210426. doi:10.1093/toxres/tfaa107
Ma, L., Wu, F., Shao, Q., Chen, G., Xu, L., and Lu, F. (2021a). Baicalin alleviates oxidative stress and inflammation in diabetic nephropathy via Nrf2 and Mapk signaling pathway. Drug Des. Devel Ther. 15, 3207–3221. Epub 20210721. doi:10.2147/DDDT.S319260
Ma, X., Wang, S., Li, C., Jia, X., Wang, T., Leng, Z., et al. (2022). Baicalein inhibits the polarization of microglia/macrophages to the M1 phenotype by targeting Stat1 in Eae mice. Int. Immunopharmacol. 113 (Pt A), 109373. Epub 20221021. doi:10.1016/j.intimp.2022.109373
Ma, Y., Wang, J., Wang, Y., and Yang, G. Y. (2017). The biphasic function of microglia in ischemic stroke. Prog. Neurobiol. 157, 247–272. Epub 20160202. doi:10.1016/j.pneurobio.2016.01.005
Manogaran, P., Walker-Egger, C., Samardzija, M., Waschkies, C., Grimm, C., Rudin, M., et al. (2018). Exploring experimental autoimmune optic neuritis using multimodal imaging. Neuroimage 175, 327–339. Epub 2018/04/09. doi:10.1016/j.neuroimage.2018.04.004
Marogianni, C., Sokratous, M., Dardiotis, E., Hadjigeorgiou, G. M., Bogdanos, D., and Xiromerisiou, G. (2020). Neurodegeneration and inflammation-an interesting interplay in Parkinson's disease. Int. J. Mol. Sci. 21 (22), 8421. Epub 2020/11/14. doi:10.3390/ijms21228421
Mayne, K., White, J. A., McMurran, C. E., Rivera, F. J., and de la Fuente, A. G. (2020). Aging and neurodegenerative disease: is the adaptive immune system a friend or foe? Front. Aging Neurosci. 12, 572090. Epub 20200923. doi:10.3389/fnagi.2020.572090
Meda, L., Baron, P., and Scarlato, G. (2001). Glial activation in Alzheimer’s disease: the role of abeta and its associated proteins. Neurobiol. Aging 22 (6), 885–893. doi:10.1016/s0197-4580(01)00307-4
Milner, M. T., Maddugoda, M., Gotz, J., Burgener, S. S., and Schroder, K. (2021). The Nlrp3 inflammasome triggers sterile neuroinflammation and Alzheimer’s disease. Curr. Opin. Immunol. 68, 116–124. Epub 2020/11/13. doi:10.1016/j.coi.2020.10.011
Ndayisaba, A., Halliday, G. M., and Khurana, V. (2024). Multiple system atrophy: pathology, pathogenesis, and path forward. Annu. Rev. Pathol., 39405585. Epub 2024/10/15. doi:10.1146/annurev-pathmechdis-051122-104528
Nie, L., Wei, Q., Sun, M., Ding, P., Wang, L., Sun, Y., et al. (2023). Injectable, self-healing, transparent, and antibacterial hydrogels based on chitosan and dextran for wound dressings. Int. J. Biol. Macromol. 233, 123494. Epub 2023/02/04. doi:10.1016/j.ijbiomac.2023.123494
Orzechowska, B., Chaber, R., Wisniewska, A., Pajtasz-Piasecka, E., Jatczak, B., Siemieniec, I., et al. (2014). Baicalin from the extract of Scutellaria baicalensis affects the innate immunity and apoptosis in leukocytes of children with acute lymphocytic leukemia. Int. Immunopharmacol. 23 (2), 558–567. Epub 2014/12/03. doi:10.1016/j.intimp.2014.10.005
Pajares, M., A, I. R., Manda, G., Boscá, L., and Cuadrado, A. (2020). Inflammation in Parkinson’s disease: mechanisms and therapeutic implications. Cells 9 (7), 1687. Epub 20200714. doi:10.3390/cells9071687
Pan, L., Cho, K. S., Yi, I., To, C. H., Chen, D. F., and Do, C. W. (2021). Baicalein, baicalin, and wogonin: protective effects against ischemia-induced neurodegeneration in the brain and retina. Oxid. Med. Cell Longev. 2021, 8377362. Epub 2021/07/27. doi:10.1155/2021/8377362
Pang, H., Xue, W., Shi, A., Li, M., Li, Y., Cao, G., et al. (2016). Multiple-ascending-dose pharmacokinetics and safety evaluation of baicalein chewable tablets in healthy Chinese volunteers. Clin. Drug Investig. 36 (9), 713–724. Epub 2016/06/29. doi:10.1007/s40261-016-0418-7
Purushothaman, B., and Sumathi, T. (2022). 5,6,7 trihydroxy flavone armoured neurodegeneration caused by quinolinic acid induced Huntington’s like disease in rat striatum - reinstating the level of brain neurotrophins with special reference to cognitive-socio behaviour, biochemical and histopathological aspects. Neurosci. Res. 174, 25–35. Epub 20210818. doi:10.1016/j.neures.2021.08.003
Ran, Y., Qie, S., Gao, F., Ding, Z., Yang, S., Tian, G., et al. (2021). Baicalein ameliorates ischemic brain damage through suppressing proinflammatory microglia polarization via inhibiting the TLR4/NF-κB and STAT1 pathway. Brain Res. 1770, 147626. Epub 2021/08/22. doi:10.1016/j.brainres.2021.147626
Ransohoff, R. M. (2016). How neuroinflammation contributes to neurodegeneration. Science 353 (6301), 777–783. doi:10.1126/science.aag2590
Reimer, L., Vesterager, L. B., Betzer, C., Zheng, J., Nielsen, L. D., Kofoed, R. H., et al. (2018). Inflammation kinase PKR phosphorylates α-synuclein and causes α-synuclein-dependent cell death. Neurobiol. Dis. 115, 17–28. Epub 20180301. doi:10.1016/j.nbd.2018.03.001
Rui, W., Li, S., Xiao, H., Xiao, M., and Shi, J. (2020). Baicalein attenuates neuroinflammation by inhibiting Nlrp3/caspase-1/Gsdmd pathway in Mptp induced mice model of Parkinson's disease. Int. J. Neuropsychopharmacol. 23 (11), 762–773. Epub 2020/08/08. doi:10.1093/ijnp/pyaa060
Saba, J., Couselo, F. L., Bruno, J., Carniglia, L., Durand, D., Lasaga, M., et al. (2022). Neuroinflammation in Huntington's disease: a starring role for astrocyte and microglia. Curr. Neuropharmacol. 20 (6), 1116–1143. doi:10.2174/1570159x19666211201094608
Scheltens, P., De Strooper, B., Kivipelto, M., Holstege, H., Chételat, G., Teunissen, C. E., et al. (2021). Alzheimer's disease. Lancet 397 (10284), 1577–1590. Epub 20210302. doi:10.1016/s0140-6736(20)32205-4
Shang, X., He, X., He, X., Li, M., Zhang, R., Fan, P., et al. (2010). The genus Scutellaria an ethnopharmacological and phytochemical review. J. Ethnopharmacol. 128 (2), 279–313. Epub 20100111. doi:10.1016/j.jep.2010.01.006
Sivandzade, F., Prasad, S., Bhalerao, A., and Cucullo, L. (2019). Nrf2 and Nf-Қb interplay in cerebrovascular and neurodegenerative disorders: molecular mechanisms and possible therapeutic approaches. Redox Biol. 21, 101059. Epub 2018/12/24. doi:10.1016/j.redox.2018.11.017
Smith, J. A., Das, A., Ray, S. K., and Banik, N. L. (2012). Role of pro-inflammatory cytokines released from microglia in neurodegenerative diseases. Brain Res. Bull. 87 (1), 10–20. Epub 2011/10/26. doi:10.1016/j.brainresbull.2011.10.004
Sohail, A., Bhat, W. F., Bhat, S. A., Furkan, M., Shah, A., and Bano, B. (2018). Investigating the preventive effects of baicalin and gallocatechin against glyoxal-induced cystatin aggregation. J. Biomol. Struct. Dyn. 36 (14), 3791–3802. Epub 20171130. doi:10.1080/07391102.2017.1400470
Sohn, E., Kim, Y. J., Lim, H. S., Kim, B. Y., and Jeong, S. J. (2019). Hwangryunhaedok-tang exerts neuropreventive effect on memory impairment by reducing cholinergic system dysfunction and inflammatory response in a vascular dementia rat model. Molecules 24 (2), 343. Epub 20190118. doi:10.3390/molecules24020343
Song, J., Li, M., Kang, N., Jin, W., Xiao, Y., Li, Z., et al. (2024). Baicalein ameliorates cognitive impairment of vascular dementia rats via suppressing neuroinflammation and regulating intestinal microbiota. Brain Res. Bull. 208, 110888. Epub 2024/02/01. doi:10.1016/j.brainresbull.2024.110888
Song, J. X., Choi, M. Y., Wong, K. C., Chung, W. W., Sze, S. C., Ng, T. B., et al. (2012). Baicalein antagonizes rotenone-induced apoptosis in dopaminergic Sh-Sy5y cells related to parkinsonism. Chin. Med. 7 (1), 1. Epub 20120121. doi:10.1186/1749-8546-7-1
Sosna, J., Philipp, S., Albay, R., Reyes-Ruiz, J. M., Baglietto-Vargas, D., LaFerla, F. M., et al. (2018). Early long-term administration of the Csf1r inhibitor Plx3397 ablates microglia and reduces accumulation of intraneuronal amyloid, neuritic plaque deposition and pre-fibrillar oligomers in 5xfad mouse model of Alzheimer's disease. Mol. Neurodegener. 13 (1), 11. Epub 20180301. doi:10.1186/s13024-018-0244-x
Steinberg, G. R., and Hardie, D. G. (2023). New insights into activation and function of the ampk. Nat. Rev. Mol. Cell Biol. 24 (4), 255–272. Epub 2022/11/02. doi:10.1038/s41580-022-00547-x
Su, S. H., Wu, Y. F., Lin, Q., and Hai, J. (2017). Cannabinoid receptor agonist WIN55,212-2 and fatty acid amide hydrolase inhibitor URB597 ameliorate neuroinflammatory responses in chronic cerebral hypoperfusion model by blocking NF-κB pathways. Naunyn Schmiedeb. Arch. Pharmacol. 390 (12), 1189–1200. Epub 2017/08/22. doi:10.1007/s00210-017-1417-9
Subhramanyam, C. S., Wang, C., Hu, Q., and Dheen, S. T. (2019). Microglia-mediated neuroinflammation in neurodegenerative diseases. Semin. Cell Dev. Biol. 94, 112–120. Epub 20190511. doi:10.1016/j.semcdb.2019.05.004
Sun, E., Motolani, A., Campos, L., and Lu, T. (2022). The pivotal role of Nf-Kb in the pathogenesis and therapeutics of Alzheimer's disease. Int. J. Mol. Sci. 23 (16), 8972. Epub 2022/08/27. doi:10.3390/ijms23168972
Tabaa, M. M. E., Aboalazm, H. M., Shaalan, M., and Khedr, N. F. (2022). Silymarin constrains diacetyl-Prompted oxidative stress and neuroinflammation in rats: involvements of Dyn/Gdnf and Mapk signaling pathway. Inflammopharmacology 30 (3), 961–980. Epub 2022/04/04. doi:10.1007/s10787-022-00961-9
Tai, Y. F., Pavese, N., Gerhard, A., Tabrizi, S. J., Barker, R. A., Brooks, D. J., et al. (2007). Microglial activation in presymptomatic Huntington’s disease gene carriers. Brain 130 (Pt 7), 1759–1766. Epub 20070330. doi:10.1093/brain/awm044
Taiming, L., and Xuehua, J. (2006). Investigation of the absorption mechanisms of baicalin and baicalein in rats. J. Pharm. Sci. 95 (6), 1326–1333. doi:10.1002/jps.20593
Tan, Z. S., and Seshadri, S. (2010). Inflammation in the Alzheimer's disease cascade: culprit or innocent bystander? Alzheimers Res. Ther. 2 (2), 6. Epub 20100412. doi:10.1186/alzrt29
Thakur, S., Dhapola, R., Sarma, P., Medhi, B., and Reddy, D. H. (2023). Neuroinflammation in Alzheimer’s disease: current progress in molecular signaling and therapeutics. Inflammation 46 (1), 1–17. Epub 20220820. doi:10.1007/s10753-022-01721-1
Tian, Z., Ji, X., and Liu, J. (2022). Neuroinflammation in vascular cognitive impairment and dementia: current evidence, advances, and prospects. Int. J. Mol. Sci. 23 (11), 6224. doi:10.3390/ijms23116224
Tu, L., Wu, Z. Y., Yang, X. L., Zhang, Q., Gu, R., Wang, Q., et al. (2019). Neuroprotective effect and mechanism of baicalin on Parkinson's disease model induced by 6-ohda. Neuropsychiatr. Dis. Treat. 15, 3615–3625. Epub 20191231. doi:10.2147/NDT.S165931
Tu, X. K., Yang, W. Z., Shi, S. S., Chen, Y., Wang, C. H., Chen, C. M., et al. (2011). Baicalin inhibits Tlr2/4 signaling pathway in rat brain following permanent cerebral ischemia. Inflammation 34 (5), 463–470. Epub 2010/09/23. doi:10.1007/s10753-010-9254-8
Vieira, B. D., Radford, R. A., Chung, R. S., Guillemin, G. J., and Pountney, D. L. (2015). Neuroinflammation in multiple system atrophy: response to and cause of α-synuclein aggregation. Front. Cell Neurosci. 9, 437. Epub 2016/01/19. doi:10.3389/fncel.2015.00437
Voet, S., Prinz, M., and van Loo, G. (2019). Microglia in central nervous system inflammation and multiple sclerosis pathology. Trends Mol. Med. 25 (2), 112–123. Epub 20181218. doi:10.1016/j.molmed.2018.11.005
Waisundara, V. Y., Hsu, A., Tan, B. K., and Huang, D. (2009). Baicalin improves antioxidant status of streptozotocin-induced diabetic wistar rats. J. Agric. Food Chem. 57 (10), 4096–4102. Epub 20090413. doi:10.1021/jf8028539
Wang, X., Yu, J. Y., Sun, Y., Wang, H., Shan, H., and Wang, S. (2021). Baicalin protects Lps-induced blood-brain barrier damage and activates Nrf2-mediated antioxidant stress pathway. Int. Immunopharmacol. 96, 107725. Epub 20210528. doi:10.1016/j.intimp.2021.107725
Wang, Y., Tian, M., Tan, J., Pei, X., Lu, C., Xin, Y., et al. (2022). Irisin ameliorates neuroinflammation and neuronal apoptosis through integrin αVβ5/AMPK signaling pathway after intracerebral hemorrhage in mice. J. Neuroinflammation 19 (1), 82. Epub 2022/04/09. doi:10.1186/s12974-022-02438-6
Wang, Z., Cui, J., Li, D., Ran, S., Huang, J., and Chen, G. (2023). Morin exhibits a neuroprotective effect in Mptp-induced Parkinson's disease model via Tfeb/Ampk-mediated mitophagy. Phytomedicine 116, 154866. Epub 2023/05/21. doi:10.1016/j.phymed.2023.154866
Ward, R. J., Dexter, D. T., and Crichton, R. R. (2015). Ageing, neuroinflammation and neurodegeneration. Front. Biosci. Sch. Ed. 7 (1), 189–204. Epub 20150601. doi:10.2741/s433
Wen, Y., Wang, Y., Zhao, C., Zhao, B., and Wang, J. (2023). The pharmacological efficacy of baicalin in inflammatory diseases. Int. J. Mol. Sci. 24 (11), 9317. doi:10.3390/ijms24119317
Woodburn, S. C., Bollinger, J. L., and Wohleb, E. S. (2021). The semantics of microglia activation: neuroinflammation, homeostasis, and stress. J. Neuroinflammation 18 (1), 258. Epub 20211106. doi:10.1186/s12974-021-02309-6
Wu, L., Xian, X., Xu, G., Tan, Z., Dong, F., Zhang, M., et al. (2022). Toll-like receptor 4: a promising therapeutic target for Alzheimer’s disease. Mediat. Inflamm. 2022, 7924199. Epub 2022/09/02. doi:10.1155/2022/7924199
Xiao, Y., Guan, T., Yang, X., Xu, J., Zhang, J., Qi, Q., et al. (2023). Baicalin facilitates remyelination and suppresses neuroinflammation in rats with chronic cerebral hypoperfusion by activating Wnt/β-catenin and inhibiting NF-κB signaling. Behav. Brain Res. 442, 114301. Epub 20230126. doi:10.1016/j.bbr.2023.114301
Xie, X. M., Hao, J. J., Shi, J. Z., Zhou, Y. F., Liu, P. F., Wang, F., et al. (2023). Baicalein ameliorates Alzheimer’s disease via orchestration of Cx3cr1/Nf-Κb pathway in a triple transgenic mouse model. Int. Immunopharmacol. 118, 109994. Epub 20230314. doi:10.1016/j.intimp.2023.109994
Xiong, J., Wang, C., Chen, H., Hu, Y., Tian, L., Pan, J., et al. (2014). Aβ-induced microglial cell activation is inhibited by baicalin through the JAK2/STAT3 signaling pathway. Int. J. Neurosci. 124 (8), 609–620. Epub 20131212. doi:10.3109/00207454.2013.865027
Xiong, P., Chen, X., Guo, C., Zhang, N., and Ma, B. (2012). Baicalin and deferoxamine alleviate iron accumulation in different brain regions of Parkinson’s disease rats. Neural Regen. Res. 7 (27), 2092–2098. doi:10.3969/j.issn.1673-5374.2012.27.002
Xu, G., Dong, F., Su, L., Tan, Z. X., Lei, M., Li, L., et al. (2024). The role and therapeutic potential of nuclear factor κB (NF-κB) in ischemic stroke. Biomed. Pharmacother. 171, 116140. Epub 2024/01/12. doi:10.1016/j.biopha.2024.116140
Xu, J., Zhang, Y., Xiao, Y., Ma, S., Liu, Q., Dang, S., et al. (2013). Inhibition of 12/15-lipoxygenase by baicalein induces microglia PPARβ/δ: a potential therapeutic role for CNS autoimmune disease. Cell Death Dis. 4 (4), e569. Epub 20130404. doi:10.1038/cddis.2013.86
Yang, J., Jia, Z., Xiao, Z., Zhao, J., Lu, Y., Chu, L., et al. (2021). Baicalin rescues cognitive dysfunction, mitigates neurodegeneration, and exerts anti-epileptic effects through activating Tlr4/Myd88/caspase-3 pathway in rats. Drug Des. Devel Ther. 15, 3163–3180. Epub 2021/07/30. doi:10.2147/DDDT.S314076
Yang, J., Wise, L., and Fukuchi, K. I. (2020). Tlr4 cross-talk with Nlrp3 inflammasome and complement signaling pathways in Alzheimer’s disease. Front. Immunol. 11, 724. Epub 2020/05/12. doi:10.3389/fimmu.2020.00724
Yao, Y., Tang, Y., Zhou, Y., Yang, Z., and Wei, G. (2022). Baicalein exhibits differential effects and mechanisms towards disruption of α-synuclein fibrils with different polymorphs. Int. J. Biol. Macromol. 220, 316–325. Epub 20220818. doi:10.1016/j.ijbiomac.2022.08.088
Yin, J., Zhao, F., Chojnacki, J. E., Fulp, J., Klein, W. L., Zhang, S., et al. (2018). Nlrp3 inflammasome inhibitor ameliorates amyloid pathology in a mouse model of Alzheimer’s disease. Mol. Neurobiol. 55 (3), 1977–1987. Epub 2017/03/04. doi:10.1007/s12035-017-0467-9
Ying, S., Yang, H., Gu, Q., Wu, Z., Zou, N., Wang, C. Z., et al. (2023). The small-molecule compound baicalein alleviates experimental autoimmune encephalomyelitis by suppressing pathogenetic Cxcr6(+) Cd4 cells. Int. Immunopharmacol. 114, 109562. Epub 20221209. doi:10.1016/j.intimp.2022.109562
Yu, C. H., Davidson, S., Harapas, C. R., Hilton, J. B., Mlodzianoski, M. J., Laohamonthonkul, P., et al. (2020). Tdp-43 triggers mitochondrial DNA release via Mptp to activate Cgas/sting in als. Cell 183 (3), 636–649. Epub 20201007. doi:10.1016/j.cell.2020.09.020
Yu, F. Y., Huang, S. G., Zhang, H. Y., Chi, H. G., Zou, Y., Lu, R. X., et al. (2015). Effect of baicalin on signal transduction and activating transcription factor expression in ulcerative colitis patients. Zhongguo Zhong Xi Yi Jie He Za Zhi 35 (4), 419–424. Epub 2015/06/06. doi:10.4108/icst.iniscom.2015.258331
Yuan, Y., Men, W., Shan, X., Zhai, H., Qiao, X., Geng, L., et al. (2020). Baicalein exerts neuroprotective effect against ischaemic/reperfusion injury via alteration of Nf-Kb and Lox and Ampk/Nrf2 pathway. Inflammopharmacology 28 (5), 1327–1341. Epub 20200516. doi:10.1007/s10787-020-00714-6
Zeng, J., Chen, Y., Ding, R., Feng, L., Fu, Z., Yang, S., et al. (2017). Isoliquiritigenin alleviates early brain injury after experimental intracerebral hemorrhage via suppressing ROS- and/or NF-κB-mediated NLRP3 inflammasome activation by promoting Nrf2 antioxidant pathway. J. Neuroinflammation 14 (1), 119. Epub 2017/06/15. doi:10.1186/s12974-017-0895-5
Zeng, Y., Song, C., Ding, X., Ji, X., Yi, L., and Zhu, K. (2007). Baicalin reduces the severity of experimental autoimmune encephalomyelitis. Braz J. Med. Biol. Res. 40 (7), 1003–1010. doi:10.1590/s0100-879x2006005000115
Zhai, H., Kang, Z., Zhang, H., Ma, J., and Chen, G. (2019). Baicalin attenuated substantia nigra neuronal apoptosis in Parkinson’s disease rats via the mTOR/AKT/GSK-3β pathway. J. Integr. Neurosci. 18 (4), 423–429. doi:10.31083/j.jin.2019.04.192
Zhang, J., Liu, Y., Zheng, Y., Luo, Y., Du, Y., Zhao, Y., et al. (2020). Trem-2-P38 Mapk signaling regulates neuroinflammation during chronic cerebral hypoperfusion combined with diabetes mellitus. J. Neuroinflammation 17 (1), 2. Epub 2020/01/05. doi:10.1186/s12974-019-1688-9
Zhang, L., Lin, G., Kovacs, B., Jani, M., Krajcsi, P., and Zuo, Z. (2007). Mechanistic study on the intestinal absorption and disposition of baicalein. Eur. J. Pharm. Sci. 31 (3-4), 221–231. Epub 20070409. doi:10.1016/j.ejps.2007.04.001
Zhang, L., Wong, L. R., Wong, P., Shen, W., Yang, S., Huang, L., et al. (2023b). Chronic treatment with baicalein alleviates behavioural disorders and improves cerebral blood flow via reverting metabolic abnormalities in a J20 transgenic mouse model of Alzheimer’s disease. Brain Behav. Immun. Health 28, 100599. Epub 20230131. doi:10.1016/j.bbih.2023.100599
Zhang, L., Xing, D., Wang, W., Wang, R., and Du, L. (2006). Kinetic difference of baicalin in rat blood and cerebral nuclei after intravenous administration of scutellariae radix extract. J. Ethnopharmacol. 103 (1), 120–125. Epub 2005/09/15. doi:10.1016/j.jep.2005.07.013
Zhang, W., Xiao, D., Mao, Q., and Xia, H. (2023a). Role of neuroinflammation in neurodegeneration development. Signal Transduct. Target Ther. 8 (1), 267. Epub 2023/07/12. doi:10.1038/s41392-023-01486-5
Zhang, X., Yang, Y., Du, L., Zhang, W., and Du, G. (2017). Baicalein exerts anti-neuroinflammatory effects to protect against rotenone-induced brain injury in rats. Int. Immunopharmacol. 50, 38–47. Epub 2017/06/18. doi:10.1016/j.intimp.2017.06.007
Zhang, X., Zhu, Y., Chen, X., Zhang, Y., Zhang, Y., Jia, Y., et al. (2014). Baicalein ameliorates inflammatory-related apoptotic and catabolic phenotypes in human chondrocytes. Int. Immunopharmacol. 21 (2), 301–308. Epub 2014/05/27. doi:10.1016/j.intimp.2014.05.006
Zhang, Y., Li, X., Ciric, B., Ma, C. G., Gran, B., Rostami, A., et al. (2015). Therapeutic effect of baicalin on experimental autoimmune encephalomyelitis is mediated by Socs3 regulatory pathway. Sci. Rep. 5, 17407. Epub 20151130. doi:10.1038/srep17407
Zhao, W., Beers, D. R., Bell, S., Wang, J., Wen, S., Baloh, R. H., et al. (2015). Tdp-43 activates microglia through nf-Κb and Nlrp3 inflammasome. Exp. Neurol. 273, 24–35. Epub 20150726. doi:10.1016/j.expneurol.2015.07.019
Zhao, X., Kong, D., Zhou, Q., Wei, G., Song, J., Liang, Y., et al. (2021). Baicalein alleviates depression-like behavior in rotenone- induced Parkinson’s disease model in mice through activating the Bdnf/Trkb/Creb pathway. Biomed. Pharmacother. 140, 111556. Epub 20210601. doi:10.1016/j.biopha.2021.111556
Zheng, W. X., He, W. Q., Zhang, Q. R., Jia, J. X., Zhao, S., Wu, F. J., et al. (2021). Baicalin inhibits Nlrp3 inflammasome activity via the Ampk signaling pathway to alleviate cerebral ischemia-reperfusion injury. Inflammation 44 (5), 2091–2105. Epub 2021/06/04. doi:10.1007/s10753-021-01486-z
Zheng, Y., Han, Z., Zhao, H., and Luo, Y. (2020). Mapk: a key player in the development and progression of stroke. CNS Neurol. Disord. Drug Targets 19 (4), 248–256. Epub 2020/06/14. doi:10.2174/1871527319666200613223018
Zhou, Y., Dong, W., Ye, J., Hao, H., Zhou, J., Wang, R., et al. (2017). A novel matrix dispersion based on phospholipid complex for improving oral bioavailability of baicalein: preparation, in vitro and in vivo evaluations. Drug Deliv. 24 (1), 720–728. Epub 2017/04/25. doi:10.1080/10717544.2017.1311968
Keywords: baicalin, baicalein, neuroinflammation, microglia, neurodegenerative diseases
Citation: Yang R, Wang R, Xu A, Zhang J and Ma J (2024) Mitigating neurodegenerative diseases: the protective influence of baicalin and baicalein through neuroinflammation regulation. Front. Pharmacol. 15:1425731. doi: 10.3389/fphar.2024.1425731
Received: 08 May 2024; Accepted: 07 November 2024;
Published: 02 December 2024.
Edited by:
Cristina Núñez, University of Murcia, SpainReviewed by:
Yan Gao, Capital Medical University, ChinaMashoque Ahmad Rather, University of South Florida, United States
Sanjay, Gachon University, Republic of Korea
Copyright © 2024 Yang, Wang, Xu, Zhang and Ma. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jing Ma, bWFqaW5nQHhpbmh1YW1lZC5jb20uY24=; Jian Zhang, emhhbmdqaWFuQHhpbmh1YW1lZC5jb20uY24=
†These authors have contributed equally to this work and share first authorship
‡These authors have contributed equally to this work