 Nicole Luengo
Nicole Luengo Gary S. Goldfield
Gary S. Goldfield Ana M. Obregón
Ana M. Obregón- 1Escuela de Nutrición y Dietética, Facultad de Ciencias para el cuidado de la Salud, Universidad San Sebastián, Concepción, Chile
- 2Children’s Hospital of Eastern Ontario Research Institute, Ottawa, ON, Canada
Background: A frequent consumption of high sugar/fat foods can affect dopamine signaling in the brain and cause sustained stimulation of the reward system. It has been hypothesized that a hypodopaminergic trait results in an individual overeating in order to increase brain DA. Genetic variants in this route have been connected with addiction and eating behaviors. Most studies focus on a specific SNP, and few studies have used multilocus genetic scores, which quantify genetic risk on a continuum.
Aim: To assess the relationship between multilocus genetic scores based on multiple gene variants in the dopaminergic pathway and measurements of anthropometry, eating behavior, food reinforcement, and food addiction (FA) in Chilean adults.
Methods: We recruited 221 Chilean adults for a cross-sectional study. A standard anthropometric measurement procedure was followed and eating behavior was examined using the Three Factor Eating questionnaire (TFEQ), Food Reinforcement Value Questionnaire (FRVQ), Yale Food Addiction Scale (YFAS) and 24-h diet recall. Multilocus genetic scores were calculated using TaqMan assays (rs1800497-rs1799732-rs6277-rs4680).
Results: No differences were found in the entire sample for anthropometric measurements, by MLGS. We found that participants with a score ≥ 2.0 in the MLGS showed higher food choices on the RVFQ and lower energy intake in protein, lipids, SAFA, MUFA, PUFA, dietary cholesterol, omega-3 and Omega-6 fatty acids in the 24-h recall (p < 0.05). Stratified by nutritional condition, the group with obesity had inferior scores on cognitive restriction, greater scores on uncontrolled eating, emotional eating, and responding to palatable food in the RVFQ. Also, in subjects with obesity, there was more food addiction in the group scoring “MLGS ≥2.0 or low dopamine signaling” (53%), compared to the group scored “MLGS <2.0 or high dopamine signaling” (23%) (p-value; 0.05). Emotional Eating scores correlated positively with MLGS in subjects with obesity.
Conclusion: In adults with obesity, the MLGS of the dopamine pathway, reflecting hypodopaminergic signaling, was associated with greater scores on food addiction and altered eating behavior traits.
Introduction
Obesity is a worldwide problem (1). It could be due to interactions among environmental and genetic factor (2, 3). Considering the 2016 levels of childhood obesity in the USA, simulated growth trajectories suggest 57% of today’s children will be obese by 35 (1). Like many other high-income and developing countries, Chile’s overweight and obesity rate is rapidly increasing, most notably among preschool and school-aged children. The last National Health Survey inform that 74% of the adult population had excess. According to the research, 27.6% of adolescents (15–19 years) are overweight, 12.2% are obese, and 1% are severely obese (4). One of the question that scientists face in this field is related to eating behavior, overeating and food craving with addictive behavior (5, 6). Studies has established that frequent ingestion of high sugar/fat food can produce changes in brain dopamine signaling (7, 8). This can result in abnormally sustained reward system stimulation (9). Food is rewarding, in part, through activation of the mesolimbic dopamine (DA) pathway. A high sugar and fat content in some foods can act like drugs, causing compulsive eating and loss of control (7). There is an emerging literature investigating dopamine genes in relationship to addictive and compulsive appetitive behaviors (10, 11).
A recent 2016 genome-wide association study (GWAS) of 9,314 females of European ancestry who were identified as having food addiction by the modified YFAS (Yale Food Addiction Scale) did not identify a significant association with any single nucleotide polymorphisms (SNPs) or genes implicated in drug addiction (12). Nevertheless, a 2015 study of neurogenetic and neuroimaging evidence for a theoretical model of dopaminergic influences to obesity, found a collection of research involving an association between obesity and genetic variants in DA receptors genes for DA receptors 2, 3, 4 (DRD2, DRD3, and DRD4), dopamine transporter 1 (DAT1) and genes for enzymes implicated with dopamine degradation—catechol-o-methyltransferase (COMT) and monoamine oxidase isomers A and B (13). Although there are no evidences for the involvement of common variants near DRD2 gene from genome-wide association study, there are several association studies that have reported a possible involvement of DRD2 variants in eating behavior traits. In this field, we previously showed that rs1800497 was not associated with food addiction, but in obese female A1 carriers was associated with scores of emotional eating and snacking reinforcement (14). We then looked for the relation between the bilocus genetic profile (rs1799732 + rs1800497) and food addiction in the same sample, showing no association (15). Considering that only two variants account for a minimal percentage of phenotypical variation we decide to explore other methods.
In genetics, to predict an individual’s risk of developing a particular trait or disease based on genetic data, certain tools are often used. Some studies use a Polygenic Risk Score (PRS) which is a numerical representation of the estimated effects of many genetic variants across the genome on an individual’s phenotype, particularly complex traits. A PRS represents an individual’s genetic predisposition to a trait or disease by integrating the cumulative impact of numerous small-effect variants, which are weighted based on their effect sizes in genome-wide association studies. An alternative approach, is the Multilocus Genetic Score (MLGS), which focuses on a specific set of genetic loci believed to contribute to a particular phenotype or disease. This method aggregates the effects of these loci, not necessarily derived from a genome-wide association studies. The MLGS can be seen as a more targeted approach, often reflecting the additive risk from a limited number of genes hypothesized to be involved in a specific biological pathway or trait.
Using the concept of candidate gene clustering, a multilocus genetic score was developed established on polymorphisms in multiple reward markers related to changes in dopamine transmission in the brain. Previously, Nikolova et al. (16) reported that MLGS was associated with higher DA signaling, predicting an increase in reward-related activity in the ventral striatum. According to this finding, a multilocus profiling method could capture the accumulative effect of genetic variants whose single effects might be undetected in small samples. After that, Davis et al. (17) showed that MLGP scores were higher in people with YFAS and that binge eating, cravings, and emotional overeating were positively correlated. In a similar manner, Yokum et al. found that participants with a greater number of alleles associated with DA signaling capacity, showed greater weight gain than those with fewer risk alleles (18).
Given this evidence, the aim if this study was to assess the relationship between multilocus genetic scores based on multiple gene variants in the dopaminergic pathway and measurements of anthropometry, eating behavior, food reinforcement, and food addiction (FA) in Chilean adults. We hypothesized that higher multilocus genetic score would exhibit higher adiposity, scores of unhealthy eating behavior and food addiction compared with lower scores.
Materials and methods
A cross- sectional study was developed between January 2016 and March 2017. The inclusion criteria were (i) subjects aged ≥18 years; without consumption of medications that affected body weight, and without treatment to lose weight. The exclusion criteria were (i) Patients with diseases such as genetic syndromes, pregnant women, individuals with associated diagnoses of cardiovascular, liver, kidney or cancer; and (ii) other pathologies that require dietary restrictions.
A convenience sample was recruited involving 221 adults (74% female, 18–54 years old), 43.8% with obesity, 11.3% overweight, and 44.8% normal weight. Participants were recruited through a variety of sources in the community, including posters, on campus at Universidad San Sebastian, recreational and community centers, as well as online advising on the website.1 Informed consent was achieved from all subjects and laboratory tests occurred at San Sebastian University. The study was approved by the Research and Scientific Ethics Committee of San Sebastian University (#48–2021-20). The protocol was conducted in agreement with the Declaration of Helsinki research ethics guidelines.
Anthropometry
We measured height, weight and waist circumference without shoes, using a weight scale (Seca 700) with a stadiometer included (100 gr. and 0.5 cm sensitivity) (19).
Based on criteria established by the World Health Organization (20), BMI cut-offs were used to determine weight status. Subjects were classified as normal-weight, overweight or obese according to their BMI values (≥ 24.9 Kg/m2, ≥ 25.0–29.9 Kg/m2, or ≥ 30.0 Kg/m2, respectively). After an overnight fast, the body composition was assessed using bioelectrical impedance, based on the manufacturer’s instructions using a Tanita TBF-300MA (Tanita Corporation, Tokyo, Japan).
Eating behavior: four questionnaires validated were used:
(1) Three Factor Eating Behavior Questionnaire: Using this instrument, 18 items are assessed and three components of eating behavior are evaluated. These components are: cognitive restraint (CR), emotional eating (EE), and uncontrolled eating (UE). Using a 4-point Likert scale, subjects rate their level of agreement on each item. Each subscale’s score was calculated by summing individual raw scores and dividing them by the number of items in that subscale (21). A Cronbach-alpha value of 0.60–0.88 was found for all subscales in the present study, suggesting moderate-to-strong internal consistency (22).
(2) Food Reinforcement Value Questionnaire (FRVQ): A 12-item questionnaire assesses the relative reinforcing value of food compared to an alternative reinforcer. Using this task, we assessed subjects’ motivation to work toward obtaining either their most preferred snack food or their highest rated healthy alternative (fruits/vegetables). In this paradigm, work was defined as the number of button presses, with more button presses indicating a higher level of reinforcement. First, a fixed ratio schedule was applied, which required subjects to press the joystick button 20 times to access either snack food or fruit/vegetables. Among the remaining items, the reinforcement schedule for gaining access to preferred snack food increased by 20 button presses to a maximum of 240 button presses for item 12. In contrast, the reinforcement program for preferred fruits/vegetables remained the same. The amount of button presses associated with snack food choices denoted the relative reinforcing value of snack food. Based on the food choices made, it was expressed as a percentage. The validity of this tool has been established against a gold-standard in adults (23), and suggests good predictive validity since they predict weight gain over time (24).
(3) 24-h diet recall: A staff of nutritionists evaluated each participant’s total energy intake, macronutrient, fiber, saturated, monounsaturated, and polyunsaturated fat intake as well as total n-6 and n-3 fatty acids using 24-h dietary recalls on days randomly select. This survey estimate energy and nutrient intake based on exhaustive food descriptions, comprising ingredient names, preparations, portions, and brand (25). Using Food Processor w/PS 10.15, 24-h recalls were analyzed for each patient.
(4) Yale Food Addiction Scale (YFAS) (First version): In accordance with the DSM-IV criteria for substance dependence, a 25-item questionnaire has been developed to assess symptoms of dependence on highly palatable foods (e.g., foods high in fats and/or carbohydrates). The YFAS requires the simultaneous existence of elevated clinically levels of distress for the food addiction diagnosis to be made. Additionally, food addiction symptoms were continuously assessed, with higher scores indicating increased susceptibility. According to Obregón et al., this instrument has been validated in Chilean adults (26), following the original validation (27).
Collection of biologic samples
A registered nurse obtain blood samples in an EDTA-coated tube of 4 mL for molecular analysis, after an overnight fast using a standard vacuum system protocol. We collected blood. After centrifuging the EDTA-coated tube at 3,300 rpm for 10 min at room temperature, plasma was separated from buffy coat and red blood cells. According to the manufacturer’s instructions, DNA was extracted from each blood sample using the QIAGEN QIAamp DNA blood mini kit #51104 (28).
Molecular genotyping
We choose common genetic variants near DRD2 that have been previously associated in several studies with dopamine pathway and eating behavior (16–18).
Genetic variant rs1800497: PCR-RFLP was used previously to assess this variant (14). In order to determine if a given allele was present or absent, the expected sizes of the PCR products were determined: one band of 307 bp was observed for homozygous A1/A1, three bands were observed for heterozygotes of A1/A2, 307 bp, 177 bp, and 127 bp, and two bands were observed for homozygotes of A2/A2 with expected sizes of 177 base pairs and 127 base pairs.
Genetic variants rs1799732, rs4680, rs6277
These variants were identified using a predesigned Taqman assay ID C_33641686_10 (Applied Biosystems) using a QuantStudioTM 3 Real-Time PCR System. For rs1799732 (Homozygous G/G, heterozygous G/Del, and homozygous Del/Del genotype); for rs4680 (Homozygous AA, heterozygous A/G, and homozygous GG) and for rs6277 (Homozygous CC, heterozygous C/T, and homozygous TT) genotype groups were determine.
Multilocus genetic score
We estimated individually Multilocus genetic scores using 4 genetic variants of the dopaminergic system, using a similar approach as other groups (17). There was a score of 1 for genotypes associated with low DA signaling, a score of 0 for genotypes associated with high DA signaling, and a score of 0.5 for intermediate heterozygotes. A score of 1 (“low dopaminergic signaling”) was assigned to TaqIA A1/A1, DRD2-141C Ins/Ins carriers, rs6277 (C957T; T-allele) and rs4680 COMT Met/Met genotypes. A score of 0 (“high dopaminergic signaling”), was assigned to TaqIA A2/A2, DRD2-141C Ins/Del and Del/Del carriers, rs6277 (C957T; C-allele), COMT Val/Val genotypes. Finally, a score of 0.5 (“intermediate dopaminergic signaling”) was given to TaqIA A1/A2 and COMT Met/Val genotypes. The scores were added to build a multilocus genetic score. The global score at each locus will be 0–1, and for the total path a score of 0–4 (Table 1) (29).
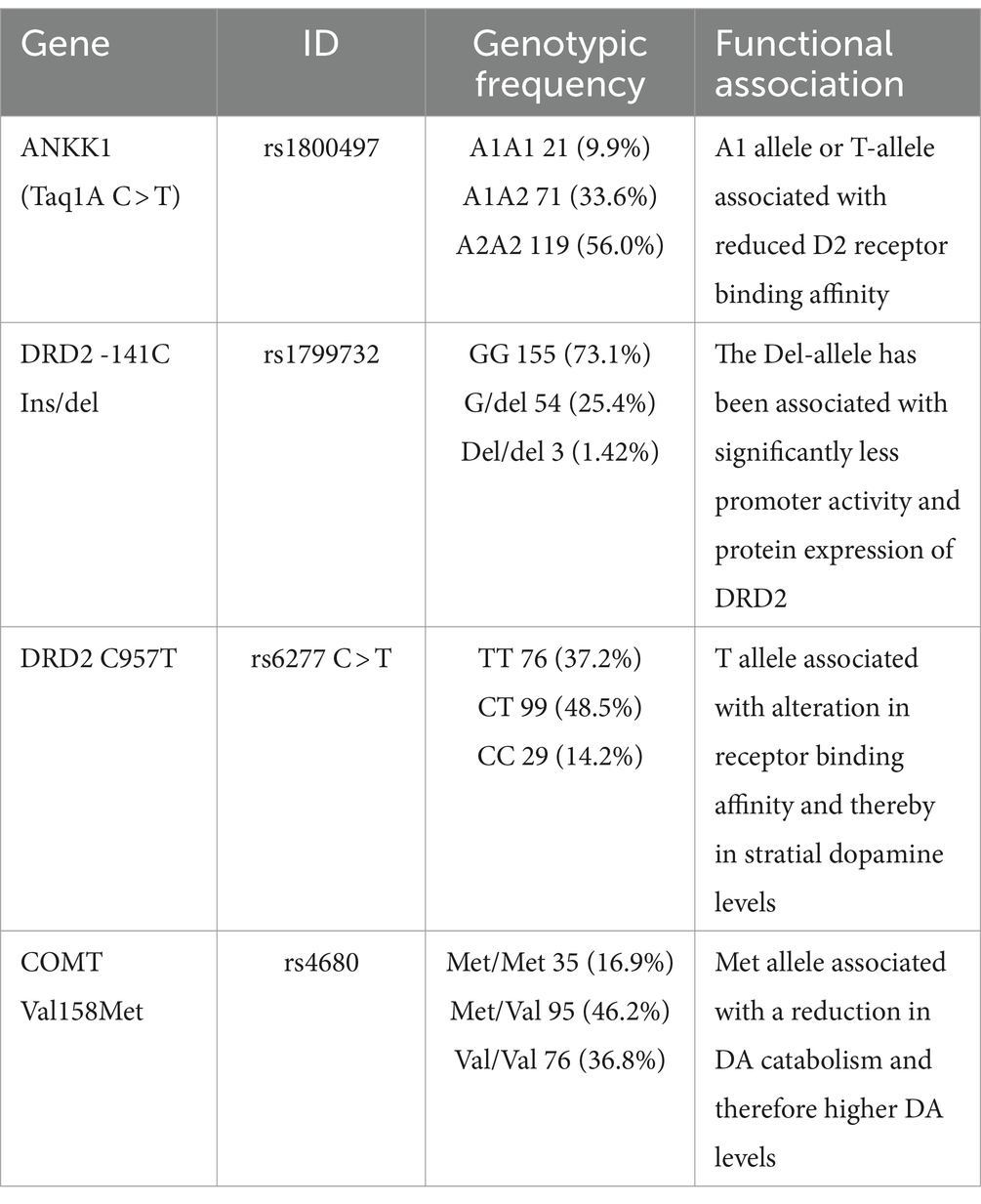
Table 1. Genotypic frequency of the SNPs studied and putatively functional association.
Data analysis
We developed a descriptive analysis of the sample (mean or median and standard deviation). Genotype and allele frequencies were determined. Also the Hardy–Weinberg equilibrium was estimated using the goodness-of-fit X2 test. An examination of differences and associations between groups was conducted using non-parametric statistics (Mann–Whitney and Kruskal-Wallis tests), including a sex-specific analysis. Data were examined with STATA 14.0 software. In order to assess the association between the MLGS and anthropometrics and eating behavior variables, the MLGS was dichotomized into two groups (MLGS <2.0 and MLGS ≥2.0).
Results
Association between MLGS and anthropometric measurements
A total of 204 participants were completely genotypes in the sample. Table 1 presents the genotypic frequencies of genetic variants. All variants meet the Hardy–Weinberg equilibrium. Table 2 presents the frequency of multilocus genetic scores in our sample. The number of participants without risk alleles was only 2/204 (0.98%). 61% of the sample had a score of two or higher. 0.49% of the sample was homozygous for the four polymorphisms examined. To determine if any association existed between the MLGS and anthropometrics and eating behavior variables, the MLGS was dichotomized into two groups, MLGS <2.0 and MLGS ≥2.0. Table 3 shows that there were no differences in the entire sample for anthropometric measurements.

Table 2. Frequency of multilocus genetic score in Chilean university students.
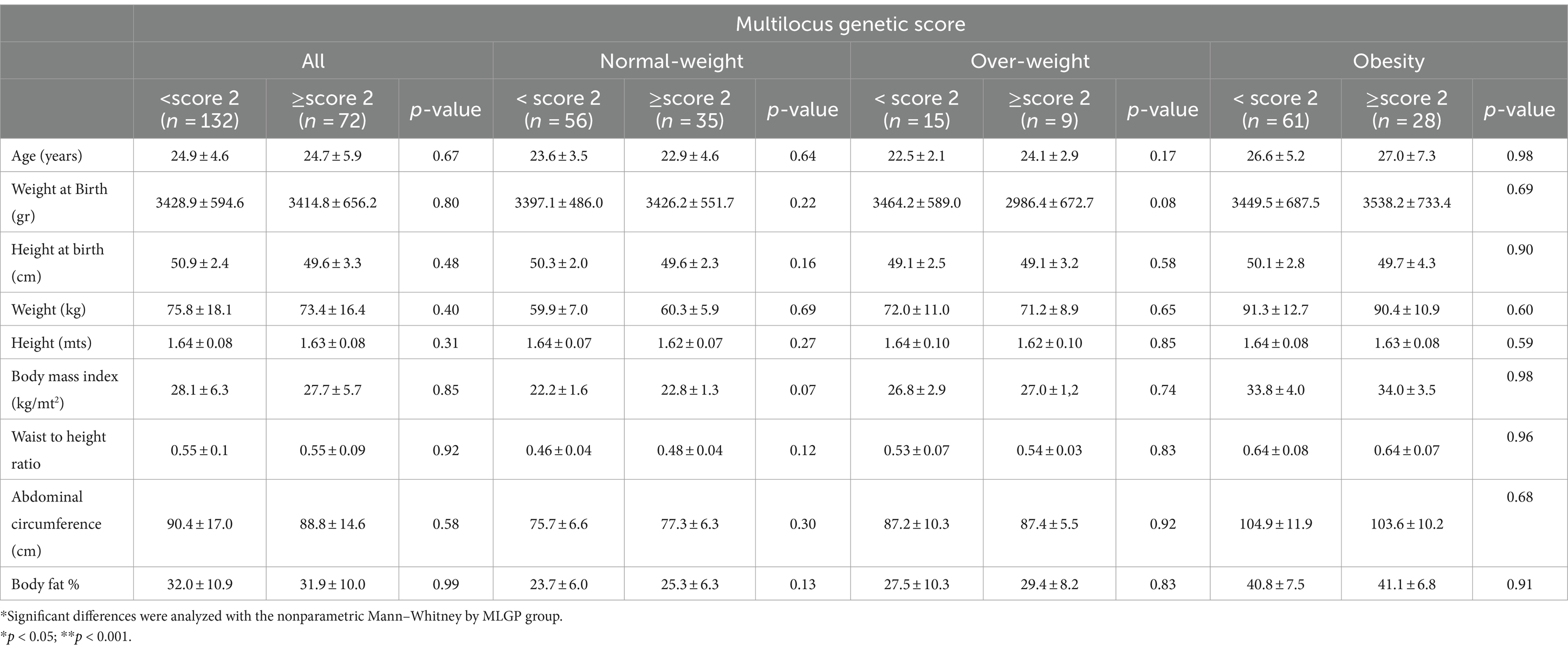
Table 3. Anthropometric measurements by Multilocus genetic score MLGS (rs1799732, rs6277, rs4680, rs1800497).
Association between MLGS and eating behaviors measurements
Table 4 shows the results for eating behavior variables. In the entire sample subjects scoring MLGS ≥2.0 (Low dopamine signaling) showed no difference in the Eating behavior scores. We found higher % of food choice in the RVFQ and lower energy, protein, lipids, SAFA, MUFA, PUFA, dietary cholesterol, omega-3 and Omega-6 fatty acids in the 24-h recall (p < 0.05).
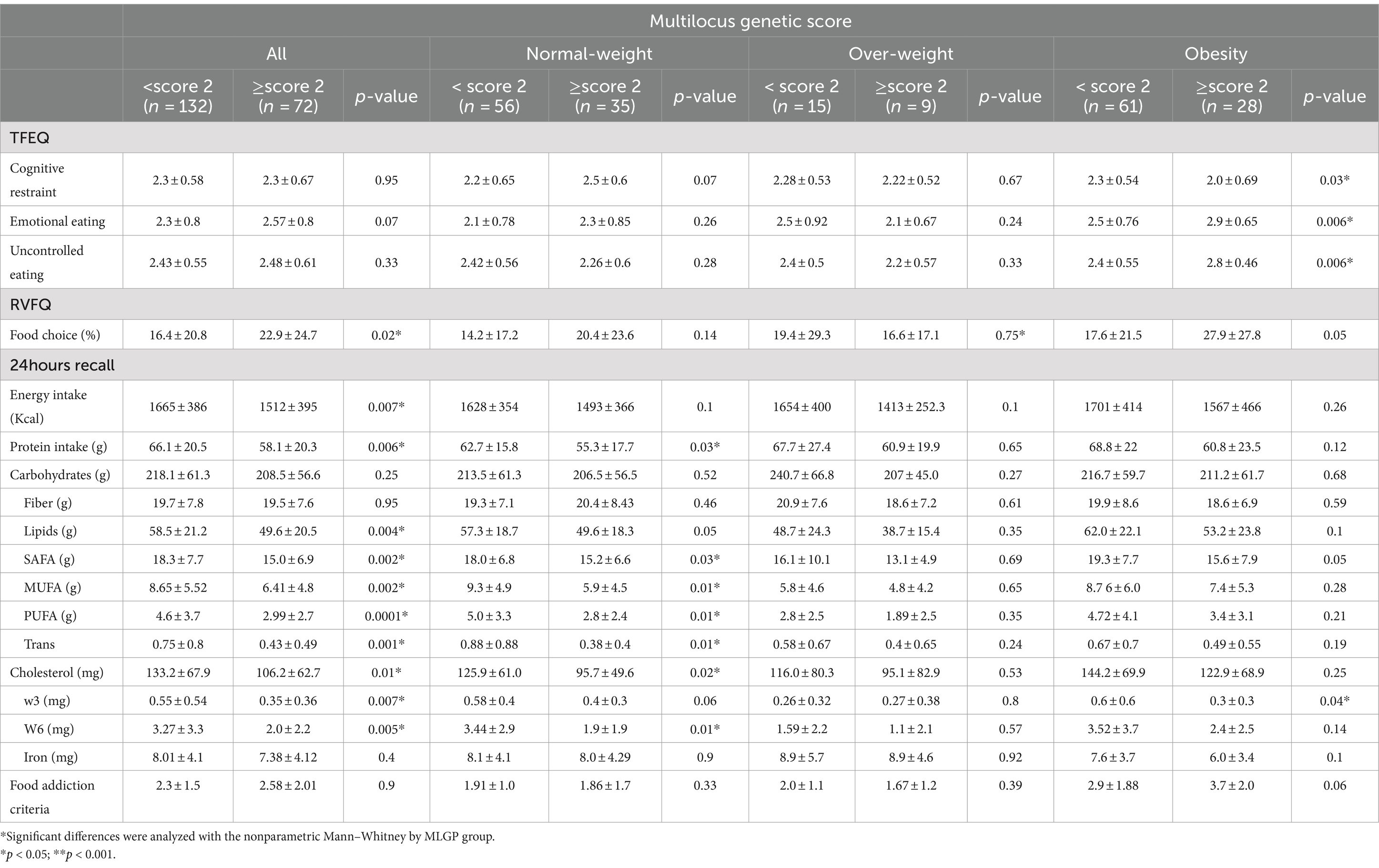
Table 4. Eating behavior by multilocus genetic score MLGS (rs1799732, rs6277, rs4680, rs1800497).
When we categorize by nutritional condition, we saw that in the Normal weight subjects the MLGS ≥2.0 group showed higher scores of cognitive restriction (ns), and lower intake of protein, SAFA, MUFA, PUFA, dietary cholesterol, omega-3 and Omega-6 fatty acids (p < 0.05). No differences were observed in the overweight group. Finally in the subjects with obesity we found lower scores of cognitive restriction and higher scores of Emotional eating, Uncontrolled eating (p < 0.05) and % of food choice in the RVFQ (p = 0.05). In the 24-h recall we found a lower omega-3 fatty acids intake (p < 0.05).
Relation between MLGS score and food addiction
No difference was observed in the frequency of diagnosis of food addiction by categories of MLGS (MLGS <2.0 and MLGS ≥2.0), in the total sample both genders, and by gender. Stratified by nutritional condition it was observed that in participants with obesity a greater % of food addiction was found in the group scored “MLGS ≥2.0 or low dopamine signaling” (53%), compared to the group scored “MLGS <2.0 or high dopamine signaling” (23%) (p-value; 0.05).
In the entire sample we did not find a significant correlation between MLGS, anthropometric and eating behavior variables. When the sample was categorized by nutritional condition, in the normal weight group we observed a positive and significant association between Emotional Eating and Uncontrolled Eating scores. A positive correlation was found between MLGS and Emotional Eating scores in the participants with obesity (r = 0.21; p < 0.05). In females there was a nearly significant positive association between MLGS and % food choice (p = 0.05).
Discussion
The present study evaluate the association between the multilocus genetic score, with anthropometric measurements, eating behavior, food reinforcement and food addiction (FA), in a population of adults from Chile. In our results we observed that 0.98% of the sample did not carrier the risk alleles and 61% had MLGS of two or higher.
According to MLGS, we did not find any differences in anthropometric measurements. These results are in agree with the study of Romer et al., who reported the BMI was not significantly associated with polygenic scores in adults (30), and also with the study conducted in Malaysian university students, which found that three SNPs (rs1800497, rs1079597, rs1800498) in DRD2 are not associated with obesity or adiposity (31). Nevertheless, this is in contrast to the results of Yokum et al. who reported an association with high DA signaling and future weight gain, reflecting that a high DA signaling promt increases in BMI, and with a longitudinal study that showed that the C- allele of the DRD2 rs6277 exhibits protective effects on weight gain (32).
In eating behavior, we observed that participants scoring MLGS ≥2.0 (Low dopamine signaling) had a higher % of food choice in the RVFQ, whereas subjects in the obesity group exhibited lower cognitive restriction scores and higher emotional eating, uncontrolled eating, and percentage of food choice.
In relation to eating behavior and obesity, an important theoretical background have related to the Reward Deficiency Syndrome, which emphasizes the neurofunctional parallels among pathological eating and drug addiction. The theory describes a hyposensitive reward system that motivates an individual to overeat to increase brain DA as a form of “self-medication” due to a hypodopaminergic trait (33, 34). Particularly, contrary to the idea of overfeeding to reestablish low concentrations of brain DA, an alternative thesis led to the Reward Surfeit Model. This indicates that persons with obesity are more reactive to food rewards, resulting in increased sensitivity to rewards (17, 35).
Based on our results, subjects scoring MLGS ≥2.0 (Low dopamine signaling) displayed no differences in their eating behavior scores on the TFEQ in the entire sample, but higher % of snack food choice on the RVFQ, reflecting a higher relative reinforcing value of snack food. Also we observed in subjects with obesity that scored in the MLGS ≥2.0, lower scores of Cognitive Restriction and higher scores of Emotional eating, Uncontrolled eating and % of snack food choice. Also the MLGS correlated positively with Emotional Eating scores (r = 0.21) and % food choice in female. This results are in accordance with Stice et al. who described that individuals with a higher number of these genotypes showed a lower level of activation in reward regions, such as the putamen, caudate, and insula, in response to monetary rewards, suggesting that individuals who have a greater number of variants associated with low DA signaling may perceive food rewards and monetary rewards as more important (29). And also with the results of Diekhof et al., who demonstrated that reward-related activation in the ventral striatum and ventral tegmental area (VTA) was significantly modulated by biologically informed MLGS profiles and sex (36). In relation to the overconsumption of drugs of abuse or palatable food, considering their reinforcing properties it has been described that the 7-repeat (7R) allele of a number of tandem repeats (VNTR) in DRD4, appears as a contributing factor in the neurobiological mechanisms underlying drug abuse, aberrant eating behaviors and related comorbidities (37). Also, the literature have report some longitudinal data. In this sense Fontana et al., developed a prospective cohort study in 359 children recruited at birth. They assessed the relation between genetic variants of dopamine genes such as the DRD4 (exon 3 VNTR) and weight observing that in the first year of life, DRD4.7R variant showed higher BMI Z-scores, and at 3–4 years of life a higher intake of palatable foods and a waist circumference, suggesting that carriers of these alleles can present an increased risk for obesity related to overeating.
In relation to addiction, we found no difference in the frequency of food addiction by categories of MLGS (MLGS <2.0 and MLGS ≥2.0) in the total sample, but in the group with obesity a greater percentage of food addiction was found “MLGS ≥2.0 or low dopamine signaling” (53%) vs. “MLGS <2.0 or high dopamine signaling” (23%). These results are somewhat in line with the results of Steiger et al., who studied the relation between dopamine genetic variations (DRD2 Taq1A, DRD4 7R, and COMT) and the risk of substance abuse in women with binge-purge eating syndromes. It was shown that women who carried high function COMT and low-function DRD4 7R alleles (higher risk) showed more substance abuse (cannabis) (38). Another study in a large cohort of Italian patients with eating disorders has suggested that the specific combination of variants in DRD2 and DRD4 genes are predisposing factors for EDs.
This contrast with previous research, examining the relationship between MLGS and food addiction, supporting that, high dopaminergic signaling genotypes are linked to obesity and higher food addiction scores, through the mechanism of higher responsiveness to eating (17, 30, 39).
Several genotypes isolated are associated with putatively low DA signaling. Individuals with an A1 allele instead of an A2/A2 allele of the TaqIA polymorphism and individuals with an Ins/Ins genotype instead of a Del-allele of the DRD2-141C Ins/Del polymorphism have fewer D2 receptors (40). In this sense, it has been suggest that altered availability of dopamine receptors specifically DA2/3R in extra-striatal and dopamine cell bodies may constitute biological vulnerability traits, for addictions (41).
Stice et al. showed that a lower caudate response predicted body fat gain in adolescents carrying TaqI A1 allele (less dopamine signaling) (42, 43). Cohen et al. showed that TaqIA A1 allele carriers have lower activation of the orbitofrontal cortex (OFC), amygdala, and hippocampal areas to monetary rewards (44) and lower activation in the midbrain, thalamus, and OFC to food rewards (45). Also the single nucleotide exchange in the Catechol-O-methyltransferase gene (COMT Val158Met), have shown fourfold less COMT activity in Met homozygotes compared to Val homozygotes (46) and according to Lachman et al. (47), the former have higher levels of tonic DA and less phasic release in the striatum.
Recently, Arrue et al. explored the relationship of cardiometabolic alterations with single genetic polymorphisms DRD2 in 285 psychiatric patients, they showed that a low dopaminergic activity was related to higher risk of suffering obesity, high diastolic blood pressure (DBP), and hypertriglyceridemia (HTG) (48). Also, Silveira et al. recently showed that variations in a MLGS reflecting DA signaling, was associated with differences in sugar intake in Children that had intrauterine growth restriction, suggesting that DA function is involved in this behavioral feature in these children (49).
This study has several limitations and strength. The fact that the data did not support our hypotheses about food addiction and MLGS could be explained in part by some methodological issues. (i) The small sample size of our study was obtained based on convenience and cannot generalized to all Chilean adults; (ii) Our results could be limited due to the small number of adults who met the criteria for food addiction; (iii) We evaluate dietary intake using 24-h recalls that have document several bias and could be responsible of some inconsistency in our results showing higher food choice, but lower energy and macronutrient intake by MLGS. Additionally, one limitation of our study was that we did not assess anxiety and depression levels in our sample, despite the fact that some studies have shown that anxiety, depression, and emotional eating are closely related (50, 51). This limitation could result in biases and misinterpretations of our results. Nevertheless, this study has the following strengths: (i) This is the first study in Chilean population that considers a multilocus approach, increasing the small contribution of individual polymorphisms to phenotypic variance; (ii) In order to measure eating behavior, we utilized a wide range of tools that were measured face-to-face by highly trained dietitians.
We conclude that, although we did not find any relationship between food addiction and MLGS, these results provide evidence for the involvement of genotypes associated with low dopaminergic signaling in eating behavior, specifically in snack food choice, emotional eating, and uncontrolled eating. These findings strongly encourage further investigations related to genetic susceptibility and the risk of chronic overeating, including the possibility to explore other pathways related to dopamine, such as physical activity. Dopamine is known to regulate physical activity, and in general studies reported in the literature as ours, do not consider this variable, open to the question of whether reduced D2R disrupts energy expenditure and activity. It is frequently suggest that reductions in D2R commonly create a reward deficit and altered appetitive motivation, which induce compulsive eating and obesity. Nevertheless, Beeler et al. developed a D2R knockdown (KD) mouse line and assessed energy expenditure and appetitive motivation under conditions of diet-induced obesity. Interestingly, the KD mice did not gain more weight or showed increased appetitive motivation and in an enriched environment with voluntary exercise opportunities, exhibited dramatically lower activity and became more obese than wild-type mice (52).
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors. Requests can be directed to the corresponding author.
Ethics statement
The studies involving humans were approved by Comite Ético Científico USS. The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
Author contributions
NL: Conceptualization, Investigation, Methodology, Software, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. GG: Conceptualization, Writing – original draft, Writing – review & editing. AO: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This study was supported by Agencia Nacional de Investigación y Desarrollo ANID, Vinculación Internacional, Proyecto FOVI220025.
Acknowledgments
The authors acknowledge the help of the entire team at San Sebastián University, Chile.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abbreviations
DA, Dopamine; GWAS, Genome-wide association study; YFAS, Yale Food Addiction Scale; SNP, Single Nucleotide Polymorphisms; DRD2, Dopamine 2 receptor; DRD3, Dopamine 3 receptor; DRD4, Dopamine 4 receptor; DAT 1, Dopamine transporter 1; COMT, Catechol-o-methyltransferase; MLGS, Multilocus genetic scores; FA, Food addiction; BMI, Body Mass Index; TFEQ, Three Factor Eating Behavior Questionnaire; FRVQ, Food Reinforcement Value Questionnaire; ANKK1, Ankyrin repeat domain containing 1 gene; SAFA, Saturated Fatty Acids; MUFA, Monoinsaturated Fatty Acids; PUFA, Polinsaturated Fatty Acids; PCR-RFLP, Restriction Fragment Length Polymorphism.
Footnotes
References
1. Ward, ZJ, Long, MW, Resch, SC, Giles, CM, Cradock, AL, and Gortmaker, SL. Simulation of growth trajectories of childhood obesity into adulthood. N Engl J Med. (2017) 377:2145–53. doi: 10.1056/NEJMoa1703860
2. Herle, M, Smith, AD, Kininmonth, A, and Llewellyn, C. The role of eating Behaviours in genetic susceptibility to obesity. Curr Obes Rep. (2020) 9:512–21. doi: 10.1007/s13679-020-00402-0
3. Adams, RC, Sedgmond, J, Maizey, L, Chambers, CD, and Lawrence, NS. Food addiction: implications for the diagnosis and treatment of overeating. Nutrients. (2019) 11:2086. doi: 10.3390/nu11092086
4. Available at: http://www.minsal.cl/wp-content/uploads/2017/11/ENS-2016-17_PRIMEROS-RESULTADOS.pdf.
5. Llewellyn, CH, and Fildes, A. Behavioural susceptibility theory: professor Jane Wardle and the role of appetite in genetic risk of obesity. Curr Obes Rep. (2017) 6:38–45. doi: 10.1007/s13679-017-0247-x
6. Wang, GJ. Food addiction a common neurobiological mechanism with drug abuse. Front Biosci. (2018) 23:811–36. doi: 10.2741/4618
7. Volkow, ND, Wang, GJ, and Baler, RD. Reward, dopamine and the control of food intake: implications for obesity. Trends Cogn Sci. (2011) 15:37–46. doi: 10.1016/j.tics.2010.11.001
8. Lennerz, B, and Lennerz, JK. Food addiction, high glycemic index carbohydrates and obesity. Clin Chem. (2018) 64:64–71. doi: 10.1373/clinchem.2017.273532
9. Bello, NT, and Hajnal, A. Dopamine and binge eating behaviors. Pharmacol Biochem Behav. (2010) 97:25–33. doi: 10.1016/j.pbb.2010.04.016
10. Aliasghari, F, Mahdavi, R, Barati, M, Nazm, SA, Yasari, S, Bonyadi, M, et al. Genotypes of ANKK1 and DRD2 genes and risk of metabolic syndrome and its components: a cross-sectional study on Iranian women. Obes Res Clin Pract. (2021) 15:449–54. doi: 10.1016/j.orcp.2021.08.001
11. Cameron, JD, Chaput, JP, Sjödin, AM, and Goldfield, GS. Brain on fire: incentive salience, hedonic hot spots, dopamine, obesity, and other hunger games. Annu Rev Nutr. (2017) 37:183–205. doi: 10.1146/annurev-nutr-071816-064855
12. Cornelis, MC, Flint, A, Field, AE, Kraft, P, Han, J, Rimm, EB, et al. A genome-wide investigation of food addiction: genetics of food addiction. Obesity. (2016) 24:1336–41. doi: 10.1002/oby.21476
13. Stanfill, AG, Conley, Y, Cashion, A, Thompson, C, Homayouni, R, Cowan, P, et al. Neurogenetic and neuroimaging evidence for a conceptual model of dopaminergic contributions to obesity. Biol Res Nurs. (2015) 17:413–21. doi: 10.1177/1099800414565170
14. Obregón, AM, Oyarce, K, García-Robles, MA, Valladares, M, Pettinelli, P, and Goldfield, GS. Association of the dopamine D2 receptor rs1800497 polymorphism with food addiction, food reinforcement, and eating behavior in Chilean adults. Eat Weight Disord-Stud Anorex Bulim Obes. (2022) 27:215–24. doi: 10.1007/s40519-021-01136-1
15. Hidalgo Vira, N, Oyarce, K, Valladares Vega, M, Goldfield, GS, Guzmán-Gutiérrez, E, and Obregón, AM. No association of the dopamine D2 receptor genetic bilocus score (rs1800497/rs1799732) on food addiction and food reinforcement in Chilean adults. Front Behav Neurosci. (2023) 17:1067384. doi: 10.3389/fnbeh.2023.1067384
16. Nikolova, YS, Ferrell, RE, Manuck, SB, and Hariri, AR. Multilocus genetic profile for dopamine signaling predicts ventral striatum reactivity. Neuropsychopharmacology. (2011) 36:1940–7. doi: 10.1038/npp.2011.82
17. Davis, C, Loxton, NJ, Levitan, RD, Kaplan, AS, Carter, JC, and Kennedy, JL. ‘Food addiction’ and its association with a dopaminergic multilocus genetic profile. Physiol Behav. (2013) 118:63–9. doi: 10.1016/j.physbeh.2013.05.014
18. Yokum, S, Marti, CN, Smolen, A, and Stice, E. Relation of the multilocus genetic composite reflecting high dopamine signaling capacity to future increases in BMI. Appetite. (2015) 87:38–45. doi: 10.1016/j.appet.2014.12.202
19. Gordon, C, Chumlea, W, and Roche, A. “Stature, recumbent length and weight”, in Anthropometric standarization reference manual. eds. T. Lohman, A. Roche, and R. Martorell. Abridged Edition. (1991). 3–8
21. Tholin, S, Rasmussen, F, Tynelius, P, and Karlsson, J. Genetic and environmental influences on eating behavior: the Swedish Young male twins study. Am J Clin Nutr. (2005) 81:564–9. doi: 10.1093/ajcn/81.3.564
22. Bland, JM, and Altman, DG. Statistics notes: Cronbach’s alpha. BMJ. (1997) 314:572–2. doi: 10.1136/bmj.314.7080.572
23. Goldfield, GS, Epstein, LH, Davidson, M, and Saad, F. Validation of a questionnaire measure of the relative reinforcing value of food. Eat Behav. (2005) 6:283–92. doi: 10.1016/j.eatbeh.2004.11.004
24. Hill, C, Saxton, J, Webber, L, Blundell, J, and Wardle, J. The relative reinforcing value of food predicts weight gain in a longitudinal study of 7–10-y-old children. Am J Clin Nutr. (2009) 90:276–81. doi: 10.3945/ajcn.2009.27479
25. Vereecken, CA, Covents, M, Sichert-Hellert, W, Alvira, JMF, Le Donne, C, De Henauw, S, et al. Development and evaluation of a self-administered computerized 24-h dietary recall method for adolescents in Europe. Int J Obes. (2008) 32:S26–34. doi: 10.1038/ijo.2008.180
26. Obregón, A, Fuentes, J, and Pettinelli, P. Asociación entre adicción a la comida y estado nutricional en universitarios chilenos. Rev Médica Chile. (2015) 143:589–97. doi: 10.4067/S0034-98872015000500006
27. Gearhardt, AN, Corbin, WR, and Brownell, KD. Preliminary validation of the Yale food addiction scale. Appetite. (2009) 52:430–6. doi: 10.1016/j.appet.2008.12.003
28. QIAGEN QIAamp? DNA Mini and Blood Mini Handbook. 3rd Edn, Germany. (2010) 27–29. Available at: https://www.qiagen.com/us/products/discovery-and-translational-research/dna-rna-purification/dna-purification/genomic-dna/qiaamp-dna-blood-kits?srsltid=AfmBOoqwK4GY_XKNU650DR9_Aa5KWfeNDhy3JqjIVmumd-pKXzgxdm01
29. Stice, E, Yokum, S, Burger, K, Epstein, L, and Smolen, A. Multilocus genetic composite reflecting dopamine signaling capacity predicts reward circuitry responsivity. J Neurosci. (2012) 32:10093–100. doi: 10.1523/JNEUROSCI.1506-12.2012
30. Romer, AL, Su Kang, M, Nikolova, YS, Gearhardt, AN, and Hariri, AR. Dopamine genetic risk is related to food addiction and body mass through reduced reward-related ventral striatum activity. Appetite. (2019) 133:24–31. doi: 10.1016/j.appet.2018.09.010
31. Fy, L, Hh, O, and Yh, S. Association of dopamine receptor D2 gene (DRD2) Taq1 polymorphisms with eating behaviors and obesity among Chinese and Indian Malaysian university students. Asia Pac J Clin Nutr. (2018) 27:707–17. doi: 10.6133/apjcn.092017.09
32. Kvaløy, K, Holmen, J, Hveem, K, and Holmen, TL. Genetic effects on longitudinal changes from healthy to adverse weight and metabolic status–the HUNT study. PLoS One. (2015) 10:e0139632. doi: 10.1371/journal.pone.0139632
33. Leehr, EJ, Schag, K, Brückmann, C, Plewnia, C, Zipfel, S, Nieratschker, V, et al. A putative association of COMT Val(108/158)met with impulsivity in binge eating disorder. Eur Eat Disord Rev. (2016) 24:169–73. doi: 10.1002/erv.2421
34. Blum, K, Braverman, ER, Holder, JM, Lubar, JF, Monastra, VJ, Miller, D, et al. Reward deficiency syndrome: a biogenetic model for the diagnosis and treatment of impulsive, addictive, and compulsive behaviors. J Psychoactive Drugs. (2000) 32:1–112. doi: 10.1080/02791072.2000.10736099
35. Heidinger, BA, Cameron, JD, Vaillancourt, R, de Lisio, M, Ngu, M, Tasca, GA, et al. No association between dopaminergic polymorphisms and response to treatment of binge-eating disorder. Gene. (2021) 781:145538. doi: 10.1016/j.gene.2021.145538
36. Diekhof, EK, Richter, A, Brodmann, K, and Gruber, O. Dopamine multilocus genetic profiles predict sex differences in reactivity of the human reward system. Brain Struct Funct. (2021) 226:1099–114. doi: 10.1007/s00429-021-02227-6
37. Botticelli, L, Micioni Di Bonaventura, E, Del Bello, F, Giorgioni, G, Piergentili, A, Romano, A, et al. Underlying susceptibility to eating disorders and drug abuse: genetic and pharmacological aspects of dopamine D4 receptors. Nutrients. (2020) 12:2288. doi: 10.3390/nu12082288
38. Steiger, H, Thaler, L, Gauvin, L, Joober, R, Labbe, A, Israel, M, et al. Epistatic interactions involving DRD2, DRD4, and COMT polymorphisms and risk of substance abuse in women with binge-purge eating disturbances. J Psychiatr Res. (2016) 77:8–14. doi: 10.1016/j.jpsychires.2016.02.011
39. Loxton, NJ. The role of reward sensitivity and impulsivity in overeating and food addiction. Curr Addict Rep. (2018) 5:212–22. doi: 10.1007/s40429-018-0206-y
40. Jönsson, EG, Nöthen, MM, Grünhage, F, Farde, L, Nakashima, Y, Propping, P, et al. Polymorphisms in the dopamine D2 receptor gene and their relationships to striatal dopamine receptor density of healthy volunteers. Mol Psychiatry. (1999) 4:290–6. doi: 10.1038/sj.mp.4000532
41. Jaworska, N, Cox, SML, Tippler, M, Castellanos-Ryan, N, Benkelfat, C, Parent, S, et al. Extra-striatal D2/3 receptor availability in youth at risk for addiction. Neuropsychopharmacol Off Publ Am Coll Neuropsychopharmacol. (2020) 45:1498–505. doi: 10.1038/s41386-020-0662-7
42. Stice, E, Spoor, S, Bohon, C, and Small, DM. Relation between obesity and blunted striatal response to food is moderated by Taq IA A1 allele. Science. (2008) 322:449–52. doi: 10.1126/science.1161550
43. Stice, E, Burger, KS, and Yokum, S. Reward region responsivity predicts future weight gain and moderating effects of the TaqIA allele. J Neurosci. (2015) 35:10316–24. doi: 10.1523/JNEUROSCI.3607-14.2015
44. Cohen, MX, Young, J, Baek, JM, Kessler, C, and Ranganath, C. Individual differences in extraversion and dopamine genetics predict neural reward responses. Cogn Brain Res. (2005) 25:851–61. doi: 10.1016/j.cogbrainres.2005.09.018
45. Felsted, JA, Ren, X, Chouinard-Decorte, F, and Small, DM. Genetically determined differences in brain response to a primary food reward: figure 1. J Neurosci. (2010) 30:2428–32. doi: 10.1523/JNEUROSCI.5483-09.2010
46. Grossman, MH, Szumlanski, C, Littrell, JB, Weinstein, R, and Weinshilboum, RM. Electrophoretic analysis of low and high activity forms of catechol-O-methyltransferase in human erythrocytes. Life Sci. (1992) 50:473–80. doi: 10.1016/0024-3205(92)90386-4
47. Lachman, HM, Papolos, DF, Saito, T, Yu, YM, Szumlanski, CL, and Weinshilboum, RM. Human catechol-O-methyltransferase pharmacogenetics: description of a functional polymorphism and its potential application to neuropsychiatric disorders. Pharmacogenetics. (1996) 6:243–50. doi: 10.1097/00008571-199606000-00007
48. Arrue, A, Olivas, O, Erkoreka, L, Alvarez, FJ, Arnaiz, A, Varela, N, et al. Multilocus genetic profile reflecting low dopaminergic signaling is directly associated with obesity and Cardiometabolic disorders due to antipsychotic treatment. Pharmaceutics. (2023) 15:2134. doi: 10.3390/pharmaceutics15082134
49. Silveira, PP, Pokhvisneva, I, Gaudreau, H, Atkinson, L, Fleming, AS, Sokolowski, MB, et al. Fetal growth interacts with multilocus genetic score reflecting dopamine signaling capacity to predict spontaneous sugar intake in children. Appetite. (2018) 120:596–601. doi: 10.1016/j.appet.2017.10.021
50. Celik Erden, S, Karakus Yilmaz, B, Kozaci, N, Uygur, AB, Yigit, Y, Karakus, K, et al. The relationship between depression, anxiety, and stress levels and eating behavior in emergency service workers. Cureus. 15:e35504. doi: 10.7759/cureus.35504
51. Papandreou, C, Arija, V, Aretouli, E, Tsilidis, KK, and Bulló, M. Comparing eating behaviours, and symptoms of depression and anxiety between Spain and Greece during the COVID-19 outbreak: cross-sectional analysis of two different confinement strategies. Eur Eat Disord Rev. (2020) 28:836–46. doi: 10.1002/erv.2772
Keywords: food addiction, polymorphism, eating behavior, dopamine, obesity
Citation: Luengo N, Goldfield GS and Obregón AM (2024) Association between dopamine genes, adiposity, food addiction, and eating behavior in Chilean adult. Front. Nutr. 11:1466384. doi: 10.3389/fnut.2024.1466384
Edited by:
Ramón Sotomayor-Zárate, Universidad de Valparaiso, ChileReviewed by:
Pablo R. Moya, Universidad de Valparaiso, ChileJosé Luis Marcos, Viña del Mar University, Chile
Copyright © 2024 Luengo, Goldfield and Obregón. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ana M. Obregón, YW5hLm9icmVnb25AdXNzLmNs; YW5pb2JyZWdvbkBnbWFpbC5jb20=