 Christina Tsigalou1*
Christina Tsigalou1* Avgi Tsolou2
Avgi Tsolou2 Elisavet Stavropoulou1*
Elisavet Stavropoulou1* Theocharis Konstantinidis1
Theocharis Konstantinidis1 Efterpi Zafiriou3
Efterpi Zafiriou3 Efthymios Dardiotis4
Efthymios Dardiotis4 Alexandra Tsirogianni5
Alexandra Tsirogianni5 Dimitrios Bogdanos6
Dimitrios Bogdanos6- 1Laboratory of Hygiene and Environmental Protection, Medical School, Democritus University of Thrace, University Hospital, Alexandroupolis, Greece
- 2Laboratory of Molecular Cell Biology, Cell Cycle and Proteomics, Department of Molecular Biology and Genetics, Democritus University of Thrace, Alexandroupolis, Greece
- 3Department of Dermatology, Faculty of Medicine, School of Health Sciences, University of Thessaly, Larissa, Greece
- 4Department of Neurology, Faculty of Medicine, School of Health Sciences, University of Thessaly, Larissa, Greece
- 5Department of Immunology-Histocompatibility, Evangelismos General Hospital, Athens, Greece
- 6Department of Rheumatology and Clinical Immunology, Faculty of Medicine, School of Health Sciences, University of Thessaly, Larissa, Greece
The nutritional habits regulate the gut microbiota and increase risk of an autoimmune disease. Western diet is rich in sugars, meat, and poly-unsaturated fatty acids, which lead to dysbiosis of intestinal microbiota, disruption of gut epithelial barrier and chronic mucosal inflammation. In contrast, the Mediterranean Diet (MedDiet) is abundant in ω3 fatty acids, fruits, and vegetables, possessing anti-inflammatory properties that contribute to the restoration of gut eubiosis. Numerous studies have extensively examined the impact of MedDiet and its components on both health and various disease states. Additionally, specific investigations have explored the correlation between MedDiet, microbiota, and the risk of autoimmune diseases. Furthermore, the MedDiet has been linked to a reduced risk of cardiovascular diseases, playing a pivotal role in lowering mortality rates among individuals with autoimmune diseases and comorbidities. The aim of the present review is to specifically highlight current knowledge regarding possible interactions of MedDiet with the patterns of intestinal microbiota focusing on autoimmunity and a blueprint through dietary modulations for the prevention and management of disease’s activity and progression.
Introduction
There is a widely held belief that the Mediterranean Sea provides an ideal temperate environment, characterized by favorable conditions in temperature, humidity, and sunlight, for the countries and nations in its vicinity. The prevailing dietary habits among the inhabitants of the Mediterranean Sea basin are believed by scientists to contribute to a healthy way of life. From the early 1960s onwards, it became evident that individuals in the Mediterranean region adhered to a consistent eating pattern with subtle variations, commonly referred to as the “Mediterranean Diet” (MedDiet). However, it wasn’t until 1993 that the Harvard School of Public Health, Ordway’s Preservation and Exchange Trust and the European Office of World Health Organization, together with Greek researchers introduced the MedDiet Pyramid, based mainly on the eating patterns of Crete Island and Southern Italy (1, 2). The Mediterranean Diet (MedDiet) emphasizes a plant-based approach, with significant consumption of vegetables, cereals, nuts, and fruits, providing ample fiber. It includes smaller quantities of animal products, with a preference for seafood and fish.
Over the last 15 years, research has yielded a wealth of knowledge regarding human microbiome. The inconceivable diversity and abundance of all living organisms (microbes, fungi, viruses, parasites), which co-evolved with humans for thousands of years and inhabit the human body was revealed by next generation technologies. Observations in both experimental models and in humans struggle to define the precise relations and interactions between microbiota and health, as well as their correlation to different diseases. Symbionts and pathobionts are in a constant battle within gut microbiota contributing to a final state of symbiosis or dysbiosis. Different external and internal factors affect the composition of gut microbiome and diet appears to be one of the most important. Dietary habits have figurative results in intestinal microbiota and MedDiet seems to favor specific phyla while suppressing others (3).
It is widely-recognized that dietary shifts largely impact on microbial populations, which can, in turn, modulate innate and adaptive immunity (4). Modern techniques, as Next Generation Sequencing (NGS) managed to provide evidence for different stages of dysbiosis related to various immune-mediated and autoimmune diseases. However, our comprehension of the cause-and-effect model and the exact outcomes and implications of these interconnections remains limited (5, 6).
The literature research process involved searching the PubMed Database using Boolean operators (AND, OR, NOT) and combinations of keywords related to Autoimmune Disease, Autoimmunity, Dysbiosis, Mediterranean diet, Microbiome, and Intestinal Microbiota. Articles were included based on their relevance to how the Mediterranean diet influences intestinal microbiota patterns and their implications for autoimmunity. We refined our investigation to encompass studies published in English, exclusively concentrating on those conducted on human subjects. Our focus was specifically on articles discussing the Mediterranean diet as a whole, while excluding studies centered on specific foods within this dietary pattern.
The objective is to offer a comprehensive overview exploring the interplay between the Mediterranean Diet and Gut Microbiota in Autoimmune Resilience. The review aims to synthesize information in these areas, focusing on how the Mediterranean Diet influences intestinal microbiota patterns and their implications for autoimmunity. Additionally, it seeks to outline dietary modulations for preventing and managing autoimmune diseases.
Discussion covers the pathophysiology of autoimmune diseases, modulation of microbial dysbiosis, and attenuation of autoimmune-related inflammation. It’s evident that diet impacts various aspects of innate and adaptive immunity through diverse mechanisms, microbiota modulation to enhance the effectiveness of dietary interventions in managing autoimmune diseases. Articles were excluded if they lacked measurement methods and outcomes or focused exclusively on children and adolescents.
On a global scale, we seek to encapsulate the probable connections between adhering to the Mediterranean Diet (MedDiet) and the intestinal microbiota, particularly in the context of autoimmunity. Additionally, we explore potential dietary adjustments for the early prevention and effective management of autoimmune diseases.
Special features of the Mediterranean diet and links to disease prevention or improvement
Mediterranean coastal countries have incorporated in their traditional food habits for years a healthy style of cooking and eating. As previously stated, the key components of MedDiet emphasize firstly, on the consumption of fruits and vegetables, nuts and whole grains, accompanied by healthy fats, mainly olive oil, spices and herbs, and secondly, on the small quantities of red meat and larger portions of fish and poultry. Additionally, the moderate drinking of red wine and physical activity/exercise is of great importance. In 2010, UNESCO introduced an expanded definition of the Mediterranean Diet (MedDiet), encompassing “a set of skills, knowledge, practices and traditions ranging from the landscape to the table, including the crops, harvesting, fishing, conservation, processing, preparation and, particularly, consumption of food” (7).
Large-scale projects demonstrated that the advantageous effects on the well-being of the participants seem to be attributed mainly to healthy mono-unsaturated fatty acids (MUFAs) in olive oil and flavonoids in red wine, nuts, spices and more (8–11). These effects are accomplished through the anti-inflammatory and anti-oxidative actions of these components, leading to the decrease of inflammation and oxidative stress (8). Yet, it’s important to also highlight the significant contribution of fiber from fruits and vegetables (1). Fiber plays a crucial role in digestive health and disease prevention (3). Overall, the Mediterranean Diet’s holistic approach, including a variety of nutrient-dense foods, has been linked to numerous health benefits, underscoring its importance for overall well-being and disease prevention.
Research conducted over the last two decades has consistently shown that adherence to MedDiet is beneficial for preventing or alleviating various inflammatory diseases. The lion’s share of the studies underlines the significant role of the Mediterranean dietary pattern in preventing cardiovascular diseases (9), type II Diabetes (10), obesity and metabolic syndrome (11, 12). Moreover, increased interest in the diet’s results on human morbidities underscores the effect of prolonged survival of the Elderly (13), better sleep and academic performance in teens (14), increased vitamin D levels, improvement of neck bone mineral density in adults who already had osteoporosis (15), longer length of telomeres in women (16), decreased depression in the Elderly (17), lower risk of fatty liver (18), lower risk of aggressive prostate cancer (19), prevention of colon cancer (20), reduced incidence of gestational diabetes and premature births (21), longevity (22), and improvement of arthritis symptoms (7). Studies suggest that the essence of the Mediterranean Diet’s benefits lies in the combined consumption of multiple health-promoting foods rather than strict adherence to a specific eating pattern. The protective effects of specific foods or nutrients in the diet appear to be less significant in combating diseases compared to embracing the entire dietary plan (23).
Effects of diet on gut microbial communities
According to Hippocrates, the father of modern medicine, ‘all diseases begin in the gut’. Centuries later, the truth of this wisdom has been unraveled through numerous studies, intricate reflections, and ambitious projects. Culture-independent techniques, as NGS in collaboration with bioinformatics, successfully accomplished the characterization of the inhabitants of our body from various anatomical sites concluding that the gut is, by far, the most populated site of the human body (24). Nowadays, it is well recognized that nutritional habits regulate the gut microbiota, and may through its effects on microbiome, have an impact on homeostasis and immune response and autoimmunity (Figure 1).
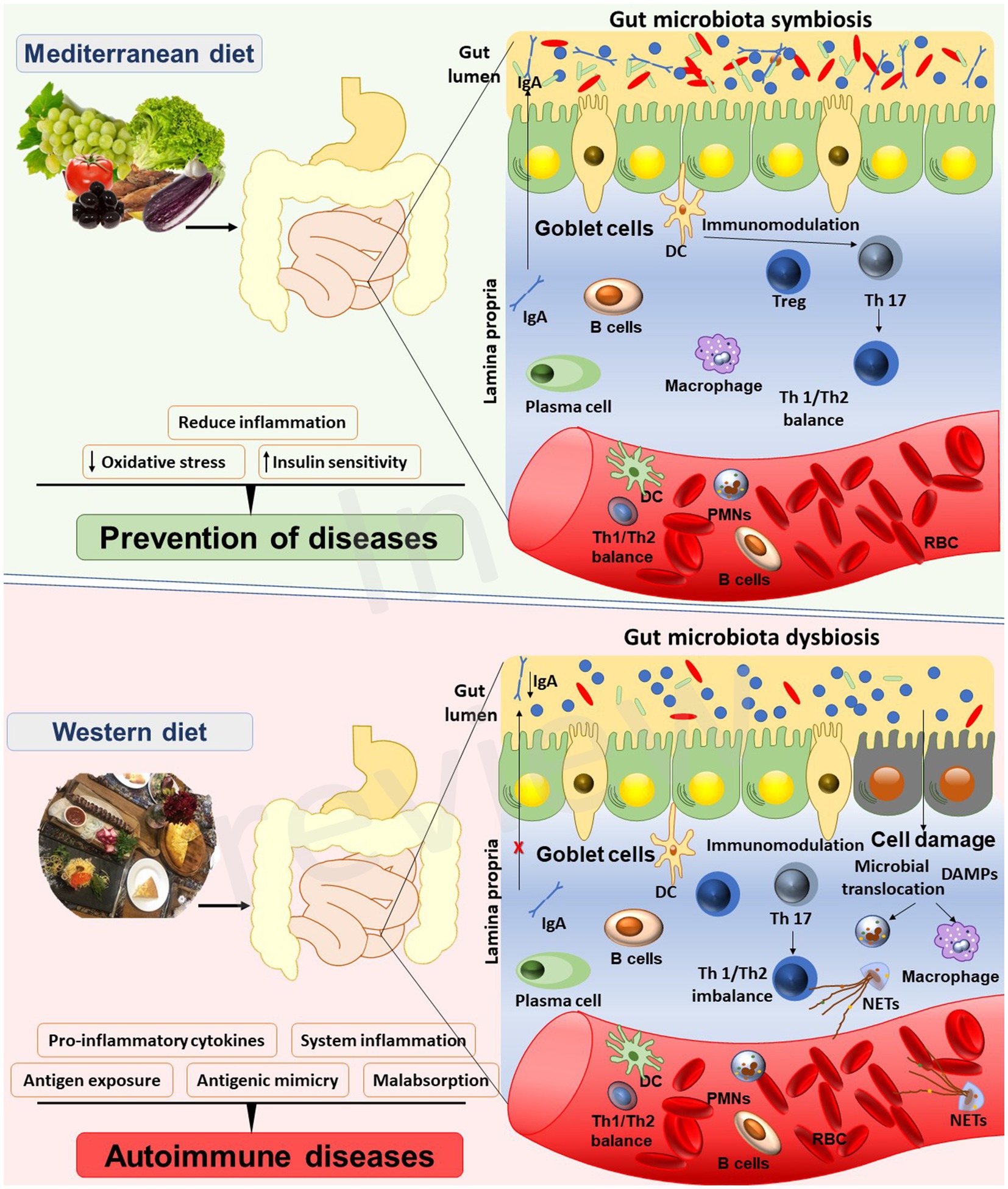
Figure 1. Interplay between the Mediterranean diet, gut microbiota, and autoimmune diseases.
Advancements in technology and research have led to new insights, particularly in distinguishing between the microbiome and microbiota, two terms still under debate (25). Recently, there has been a resurgence in the original definitions proposed by Whipps et al. in 1988 (26). The microbiota, as defined, represents a characteristic microbial community within a well-defined habitat, characterized by specific physiochemical properties. It encompasses not only the microorganisms themselves but also their activities, forming distinct ecological niches. This dynamic and interactive micro-ecosystem undergoes temporal and spatial changes and is intricately linked with macro-ecosystems, including eukaryotic hosts, playing a crucial role in their functionality and health.
On the other hand, the microbiome consists of a collection of microorganisms from various kingdoms (Prokaryotes such as Bacteria and Archaea, as well as Eukaryotes like Protozoa, Fungi, and Algae). Its scope extends beyond the organisms themselves to include their activities, encompassing microbial structures, metabolites, mobile genetic elements (such as transposons, phages, and viruses), and relic DNA, all of which are influenced by the environmental conditions of the habitat.
Large-scale studies have dictated that different eating patterns promote a variety of gut microbiota composition and diversity and highlight diet as one of the most significant influencing factors (3, 6, 27, 28).
Different dietary practices, alterations in dietary components -such as fats, proteins and carbohydrates-, food complements as probiotics and prebiotics, even salt composition, can all modulate the gut microbiota (29). However, such changes can exert an effect not just at microbial species level but also at phyla differences (30).
The majority of study findings are based on animal experiments, particularly the reshaping of the gut microbiome due to a Western diet, resulting in a significant expansion of the Mollicute lineage within the Firmicutes, alongside a prevailing reduction in Bacteroidetes in the microbial community. Furthermore, research in both animals and humans indicates that increased fructose consumption contributes to small intestine bacterial overgrowth and elevated intestinal permeability, leading to high levels of endotoxins (31–35). Nevertheless, the supplementation of prebiotics has the potential to reverse microbial population shifts induced by a high-fat diet in obese mice (36). Furthermore, mice with an elevated protein intake exhibited notable increases of Lactobacillaceae/Lactobacillus and decreased Clostridiaceae/Clostridium within their gut microbiota, with the extent of these changes being dose-dependent (37). In humans, gut microbiota varies at the species level, depending on fat and fiber or carbohydrate consumption, between omnivores and vegetarians, or even among vegetarian subgroups (3). On the whole, dietary patterns play a role in shaping the diversity of gut microbiota. This diversity lies on higher taxonomy levels, as phylum, family, and genus, rather than at the species level. Changes in individual dietary components, such as gluten, may also change composition and function of the gut microbiome and host physiology of healthy individuals (38). Investigating geographical and socio-economic differences in various regions, numerous studies shed light on how diet influences gut microbiota, comparing a spectrum of dietary habits (39–41).
Interestingly, a large-cohort study on US immigrants from non- Western countries led to a relatively fast and enduring loss of gut microbiome diversity, accompanied by the decline of bacterial enzymes related to plant fiber degradation and the displacement of native strains and functions within the first 9 months of immigration and, as a result, predisposition of individuals to obesity and metabolic diseases. Substantial disruptions in the gut microbiome can be partially explained by dietary variations (42).
Gut microbiome’s link to autoimmunity
It seems that apart from the identification of all taxa of microbes, the ultimate challenge is to define the benefits and drawbacks of the ‘healthy gut’. The human microbiota, and the gut in particular, has been deemed an “essential organ,” containing approximately 1,000 different microbial species, and counting for over 150 times more genes than those of the whole human genome (43, 44).
The pivotal role of human microbiota in health and disease involves a diverse array of functions. The microbiota assists energy extraction from food, supplies unique enzymes and biochemical pathways to the human body, and acts as a physical barrier against foreign pathogens through antimicrobial substances or space competition (45–47). Last but not least, the gut microbiota is essential for the normal development of both the intestinal mucosa and the host humoral and cellular immune system, as signals and metabolites from the microorganisms are sensed by cells of the innate immune system and are recognized as physiological responses (48–50). The term “autoimmunity” refers to activation of adaptive immune responses, with the involvement of T and B lymphocytes against self-antigens, i.e., against its own healthy cells and tissues. The actual causes of autoimmunity are not fully understood, but various environmental factors, including diet, lifestyle and infections, along with the genetic background of the host, appear to play a crucial role (51, 52). The initiation of adaptive immune responses is rooted in the connection between the innate and adaptive immune systems. It is the innate immune system that can discriminate between self and non-self-antigens, whereas the adaptive immune system recognizes either native antigens or peptides presented in the context of major histocompatibility complex molecule (MHC) (53). Therefore, when a microbial pathogen or parasite invades the host, both the adaptive immune system and the innate system respond accordingly. Studies in both humans and animal models designate the involvement of commensal microbiota in autoimmunity (54, 55).
The impact of the microbiota on autoimmune diseases, which rely on the innate-adaptive immune system connection, can range from being neutral to being essential for the initiation of autoimmunity. The loss of immune tolerance to self-antigens can occur due to changes in microbial composition. Consequently, the human microbiota becomes a crucial participant in the initiation and perpetuation of autoreactive immune responses, ultimately resulting in self-tissue destruction and the overt of autoimmune diseases (54, 55). Several factors contribute to the loss of immune tolerance and induction of autoimmunity by microorganisms, including molecular mimicry, bystander activation, and viral persistence with or without epitope spreading (56–60) focused, dominant epitope-specific immune response. This expansion targets self or foreign proteins, encompassing subdominant and/or cryptic epitopes on the same protein (intramolecular spreading) or other proteins (intermolecular spreading) (61).
Self-antigens can be a result of slightly changed antigens, even at an amino acid residue. As a result, the immune reaction will affect both the “wild” protein or the altered one (61, 62).
Microorganisms could trigger autoimmune responses through the expression of heat- shock proteins in their cells when under stress, such as high temperatures. These proteins can subsequently become targets of the immune system’s response. Heat shock proteins produced by microbes have the potential to trigger self-reactivity toward the host’s own heat shock proteins, potentially leading to autoimmune diseases. This self-reactivity toward heat shock proteins serves to protect the host against disease by regulating the induction and release of pro-inflammatory cytokines. Nevertheless, antibodies targeting self heat shock proteins have been implicated in the pathogenesis of autoimmune diseases such as arthritis and atherosclerosis (63).
In ‘molecular mimicry’, a shared immunologic epitope between a microbe and the host is the prerequisite for the initiation of cross-reactive immune responses. A notable illustration of molecular mimicry as a mechanism leading to the onset of autoimmune disease is evident in individuals with rheumatic fever after being infected with group A beta-hemolytic Streptococci. Analysis of infected hosts’ sera demonstrated the presence of antibodies reactive with heart, joints, brain, and skin. Moreover, patients’ antibodies are found to cross-react with streptococcal antigens, like the group A carbohydrate antigen, the M protein (a Streptococcus- related virulence factor) and to cross-react with myosin. Cross-reactive peptides from M protein and cardiac myosin may provoke the onset of autoimmune disease in mice with rheumatic heart disease (62). The mechanism of molecular mimicry can also operate at the T-cell level involving antigenic epitopes of human and foreign origin which serve as targets of CD4 and CD8 T-cell responses. Previous studies of our group have meticulously addressed the role of molecular mimicry in the induction of autoimmune diseases, primarily affecting the liver and the gastrointestinal tract (62–82).
‘Bystander activation/killing’ is another mechanism resulting in autoimmune diseases. Viral infections can activate Antigen-Presenting Cells (APCs) which, in turn, activate primed autoreactive T cells for the onset of autoimmune disease. Furthermore, initiation of bystander activation can be a result of virus-specific T cells. Bystander effect may lead to killing of the uninfected neighboring cells, that increases the immunopathology at the infected area (82). Finally, persistent viral infections may cause immunopathology, as a result of the permanent presence of viral antigens challenging the immune system (83).
Adherence in MedDiet and its effects on microbiome related to autoimmune diseases: the case of rheumatoid arthritis
The Mediterranean diet is rich in fiber, antioxidants and vitamins and encompasses anti- inflammatory properties (84). Various data have demonstrated that consumption of cereals, fruits and vegetables, nuts and legumes, omega-3 polyunsaturated fatty acids in olive oil and moderate consumption of red wine flavonoids leads to the reduction of pro-inflammatory cytokines, the increase of anti-inflammatory cytokines and the decrease of oxidative stress (83, 84). It has been shown, that adherence to a Mediterranean pattern diet leads to the reduction of CRP and TNF-α levels (85, 86).
Several studies investigated the role of MedDiet in Rheumatoid Arthritis (RA), a common autoimmune rheumatic disease. Highlighting the immunopathological characteristics of rheumatoid arthritis (RA), underscore the distinctive cytokine profiles associated with the condition, notably the heightened levels of Th1/Th17 cytokines and the compromised function of Tregs (87). Immune dysregulation, and imbalance between the function, differentiation, and regulation of Th17 and Treg cell plays a highlighting role in disease onset (87) disorders, rheumatoid arthritis is associated with oxidative stress. This refers to a state being a quintessential example of chronic inflammatory autoimmune where the level of reactive oxygen species gradually rises, either due to increased production, diminished antioxidant defenses, or both, ultimately leading to disruptions in redox signaling (88). Data suggest that adopting a Mediterranean Diet model reduces the inflammatory activity of the disease, enhances functionality, and improves the quality of life for patients with RA (89). Another study claims that adherence to MedDiet is associated with decreased disease activity, improved physical function, and heightened vitality individuals with RA (87). Subsequent studies have yielded inconsistent findings (90–98). A recent systematic review analyzing the data so far provided from prospective human studies, concluded that MedDiet has beneficial effects in reducing pain and increasing physical function in people with RA but underline that there is insufficient evidence to support the widespread recommendation of the Mediterranean Diet for the prevention and management of RA (7).
Data from studies on short term MedDiet and fasting revealed no significant impact of these diets on the gut microbial profile in individuals with RA or fibromyalgia (98).
Recent studies have underscored the importance of long-term adherence, lasting more than 3 months to MedDiet is required to produce significant diversity in the gut microbiome of overweight omnivores (99). The CARDIVEG study reported that MedDiet significantly alters the abundance of Lachnoclostridium, Enterorhabdus and Parabacteroides, while vegetarian diet significantly disturbs the abundance of Streptococcus, Anaerostipes, Clostridium sensustricto, and Odoribacter (99).
A recent study suggests the potential role of P. copri in the preclinical evolution and pathogenesis of synovitis in RA, based on the analysis of anti-P. copri antibody responses in different RA cohorts (100).
RA drugs include disease-modifying antirheumatic drugs (DMARDs), target cytokines and immune responses. Patients usually initiate treatment with conventional DMARDs such as methotrexate. However, exploring alternative approaches like probiotics, prebiotics, herbal remedies, and dietary interventions is crucial for discovering new avenues for treating the disease.
Modulating gut microbiota holds promise as a potential approach for treating rheumatoid arthritis (RA) microbiota transplantation (FMT) is being investigated for its ability to rebalance the gut microbiota and improve RA symptoms (101).
In rheumatoid arthritis (RA), the presence of Prevotella spp., particularly P. copri, exacerbates the disease by promoting proinflammatory metabolite production. Colonization with P. copri and a fiber-rich diet leads to dysbiosis, characterized by metabolites like succinate and fumarate, worsens RA symptoms (102). Conversely, a Mediterranean diet’s effectiveness in RA treatment may vary based on gut microbiome personalized treatment approaches and may aid in identifying predictive biomarkers microbiota in mediating dietary effects on RA could help elucidate why certain dietary enhancing the efficacy of dietary interventions in RA management, ameliorate and exacerbate colitis in animal models. While they have the potential to improve colitis symptoms, caution is advised as certain prebiotic fibers may contribute to gut dysbiosis and lead to excessive production of colonic butyrate, potentially worsening inflammatory bowel disease (IBD) (103). The fermentation products generated by consuming high-fiber diets (and possibly Mediterranean diets) can have adverse effects, particularly in the presence of intestinal inflammation. For instance, the production of butyrate after diet fermentation may exacerbate inflammation by promoting NLRP3 activation.
Meddiet lead to changes of intestinal short-chain fatty acids (SCFA)
Such studies on gut microbiome changes in patients with RA adherent to MedDiet have not been performed so far, but several data from research conducted on the gut microbiome and its relation to RA may be relevant. Several studies have assessed changes of the microbiota in relation to MedDiet in patients with RA. An early study suggested that changes of intestinal Short-Chain Fatty Acids (SCFA) from the microbiota are not necessarily correlated with clinical improvements and disease activity in RA (104).
An association between MD and increased SCFAs production is well known (105). Rich fiber foods like fruits, vegetables, and legumes, commonly consumed by those following the Mediterranean Diet, are broken down by Firmicutes and Bacteroidetes bacteria. This process results in the production of elevated levels of fecal Short-Chain Fatty Acids (SCFA) (106, 107).
Among these SCFAs, butyrate, extensively studied for its functional role, is produced during the fermentation of dietary fiber by the anaerobic intestinal microbiota. Butyrate has beneficial effects on intestinal barrier integrity, as it enhances the expression of tight junction proteins. Moreover, it helps prevent deleterious intestinal permeability and bacterial translocation, thereby exerting a protective influence on the initiation of pro- inflammatory responses (106, 108, 109). To relevance in collagen-induced arthritis, an animal model of RA, butyrate is able to suppress RA features and this is achieved via a butyrate- mediated increase of IL-10 producing Tregs and a decrease of Th17 (110). However, not all Mediterranean type of diets increase SCFAs, and butyrate in particular at the same extent, and this may have an impact in their ability to suppress anti- inflammatory immune responses or to influence intestinal microbiota-related influence of the immune system.
Modified Mediterranean type enriched for SCFA production are increasingly popular but their effect in RA has not been assessed. Omnivores who consume a MedDiet-pattern diet rich in fruit, legumes and vegetables not only have increased SCFAs (106), but also decreased trimethylamine N-oxide (TMAO), a microbial metabolite the precursors of which are carnitine and choline which are primarily found in foods of animal origin (106, 111).
Several microbial genera, like L-Ruminococcus, have been linked to the intake of animal proteins such as a diet plenty in red meat consumption and increased TMAO levels. This is very interesting, in view of recent data demonstrating a twofold to threefold increased abundance of Ruminococcus gnavus in patients with spondylarthritis and to a lesser extent in RA patients compared to healthy controls (112).
However, the most notable association over the last few years is that linking P. copri the TMAO-producing anaerobic, Gram-negative Bacteriodetes, with the development of RA, an association thoroughly reviewed elsewhere (113, 114). Prevotella spp. are abundant in the periodontium, the intestine, and the respiratory system and its heightened presence is deemed a risk factor for RA and features associated with RA, such as cardiovascular risk- events (113, 115–118). Furthermore, it may impact the metabolism of the microbiota to reduce the effectiveness of the common disease-modifying anti-rheumatic drug (DMARD) methotrexate (119). The question arises as to whether adherence to MedDiet alone or in combination with other diet supplementation can alter gut dysbiosis to a state that Prevotella spp. are not dominant (62, 120, 121). This event would stop the vicious circle of immunological events that take place, Prevotella being in the center of it and could prevent from RA.
A recent elegant study has shown that the microbiota of individuals in pre-clinical early RA stages had significantly altered fecal microbiota composition compared with their first- degree relatives (FDRs) (116). In these pre-clinical RA individuals, who had either developed anticitrullinated peptide antibodies or rheumatoid factor positivity, and/or exhibited symptomatology and features associated with possible RA in the ‘pre-clinical stages,’ their feces were significantly enriched in the bacterial family Prevotellaceae, particularly Prevotella spp., compared to their first-degree relatives (FDRs) (116). These data clearly demonstrate that Prevotella spp. enrichment in early RA and very early RA may indeed be a characteristic feature of these subclinical phenotype raising the expectation that Prevotella spp. are pathogenically relevant to the development of the disease, from early stages, rather than consequence of established disease state.
We recently reviewed the existent literature tackling this topic and thoroughly discussed mechanistic scenarios, which could implicate Prevotella species in the establishment of RA (122). We underlined the decisive role of Prevotella species in the potential induction of either ACPA-positive or ACPA-negative RA. This contrasts that of the well-known association of P. gingivalis and ACPA-positive RA and the inability of this oral commensal to explain the induction of ACPA-negative RA. Though several components of the Prevotella-host interactions are still puzzling and improperly explored, some of the features linking P. copri, as well as oral Prevotella species, to RA are striking. One of the most fascinating has recently been obtained by Pianta et al. (123). These authors using liquid chromatography–tandem mass spectrometry identified two novel RA autoantigens, targeted by half of the ACPA-pos and ACPA-negative patients with RA. The two autoantigens were the N-acetyloglucosamine- 6- sulfatase (GNS) and filamin A (FLNA). Of interest, both GNS and FLNA were expressed in synovia, a finding that supports the notion that they could be relevant to immune-mediated tissue destruction. The former also appeared to be citrullinated, which makes it likely target of ACPA antibodies. Data indicating that antibody concentrations of these autoantibodies are correlated further supports the likely association of anti-GNS antibodies with ACPA (123). These autoantigens were recognized by more than half of the RA patients, and were also present in ACPA-negative RA. Of immunological relevance, Pianta et al. (123) found that the epitopic regions of GNS and FLNA not only are highly homologous to Prevotella copri but are also targeted by B and T-cells responses, also cross-recognizing the Prevotella homologs. Thus, there is evidence that a molecular mimicry mechanism is in operation, which could account for the induction of those autoantibodies (123). Of relevance, molecular mimicry involving an oral Prevotella sp. and collagen I, have been previously reported and has been considered a likely trigger for chronic periodontitis and possibly inflammatory (124).
The GNS peptidyl sequence was highly homologous to a sequence from sulfatase proteins of the Prevotella sp. and Parabacteroides sp. Finding marked homologies between human and microbial highly conserved proteins is extremely common and the homology reported by Pianta et al. belongs to this category (123). In a similar vein, homologies between human and microbial heat shock proteins and human and microbial 2-oxo-acid dehydrogenase complexes have been identified and suggested as triggers of various organ and non-organ specific immune-mediated and autoimmune diseases. Because they are extremely common, several investigators, including authorities in the field, suggested that molecular mimicry involving such homologs must not be regarded as a perpetuator of autoimmunity.
Similarities between human GNS and Prevotella were shown by Pianta et al. (123). The same human GNS sequence had also marked amino acid similarity with the gut commensal Parabacteroides sp. Using a BLAST program, they investigated for additional similarities that could be potentially homologous to the human GNS epitope. Microbial mimics that have similarities to the core epitope region of the human GNS epitope were also presented.
Particularly, the pentameric -FFMMI- 224-228 aa of human GNS is contained in the transmembrane protein of Streptococcus gordonii (aa 43–47), transcriptional regulator of Lactobacillus casei (aa 464-468), acetyl-CoA carboxylase, carboxyl transferase subunit beta of Clostridium cellulolyticum H10 (aa 133-137), hypothetical protein of Vibrio phage KVP40 (aa 23-27) and several other foreign proteins, suggesting that many other triggering factors may really exacerbate GNS-specific autoreactivity in RA by molecular mimicry. Thus, Pianta’s homologs (123) cannot be disregarded as the relevant mimics are targets of cross-reactive responses and the humoral responses against the microbial peptides are correlated with disease-specific autoantibodies, as ACPAs. Moreover, gut dysbiosis leads to immunologic alterations, which are pivotal for the loss of immunological tolerance to RA-specific autoantigens. This was achieved in a stepwise manner and that several immunological mechanisms are involved, molecular mimicry being just one of those. Firstly, an external parameter leads to changes in the gut microbiome, as well as changes on the microbiome of the oral cavity and an establishment of gut dysbiosis (123). This parameter or combination of parameters could be drugs, infections, changes of diet habits from Mediterranean diet to Western diet and more. Gut dysbiosis, in turn, leads to the establishment of an immunological environment, which alter the composition of regulatory T and B cells and diminishes their capacity to suppress autoreactive immune responses and augments pro- inflammatory Th17 responses. The enrichment of specific species, such as Prevotella, has additional consequences, notably the activation of the immune system against a gut microbe. This can initiate anti-Prevotella responses, which, through mechanisms like molecular mimicry and others, in conjunction with various factors, may lead to the induction of autoreactive responses. This initiation can lead to the development of autoimmune disease only in susceptible individuals (125).
Of indirect relevance to the topic, among the American indigenous populations, the Canadian Inuit population has the lowest age-adjusted prevalence of RA (at 0.65%, with an incidence of 48.2 per 100,000 per year) (126). This is of interest because a microbiome study has found that Prevotella spp., were enriched among the Inuit consuming a Western diet.
However, the gut microbiome of Inuit consuming a traditional high-fiber diet (127) had significantly less genetic diversity within the Prevotella genus, compared to the Inuit consuming a Western diet further, indicating that a low-fiber diet might not only select against Prevotella but also decrease its diversity, a factor which could be relevant to the induction of autoreactive responses implicating Prevotella in inflammatory arthritis.
A recent study has shown that berberine modulates gut microflora and exerts an anti- inflammatory effect on collagen-induced arthritis (128). This is achieved because the abundance of Prevotella is diminished and the abundance of butyrate-producing bacteria in CIA rats is increased (128).
Nonetheless, a study examining changes in gut microbiota associated with the MedDiet discovered a greater presence of Bacteroidetes and a lower Firmicutes–Bacteroidetes ratio in those with a higher Mediterranean Diet score (129). However, the study did not report significant differences in levels of Bacteroides and Prevotella, genera included in the Bacteroidetes phylum (129). Again, it is not clear whether long-term adherence to MedDiet may indeed exert an influence on the levels of Prevotella, which could in turn utilize beneficial effects in preventing from RA or ameliorating disease’s clinical features. Another study conducted an integrative analysis of distal gut microbiota composition and functions, as well as intestinal metabolites in Egyptian teenagers consuming Mediterranean Diet- related products. The study compared this cohort with a group of USA teenagers consuming a Western-type diet enriched in animal proteins, fats, and processed carbohydrates (130). The Prevotella enterotype predominated in gut microbial communities of the Egyptian teenagers while the Bacteroides enterotype dominated the USA cohort. As it was expected the intestinal environment of Egyptian teenagers was characterized by higher levels of SCFA, an increased prevalence of microbial polysaccharide degradation-encoding genes, and a higher proportion of several polysaccharide-degrading genera while the gut environment of the American children was enriched in proteolytic microbes and end products of protein and fat metabolism (130).
Adherence in MedDiet and its effects on microbiome related to autoimmune diseases: the case of autoimmune disease of the central neuros system (CNS)
Multiple sclerosis (MS) and Neuromyelitis optica (NMO) is an autoimmune disease of the CNS, with some overlapping symptoms. A few studies have shown that both genetic and environmental factors may affect the etiology of the MS, including dietary habits. Moreover, the onset of the disease post microbial infections via molecular mimicry and bystander activation mechanisms is also a scenario (131, 132). The multifaced roles of gut microbiome on MS pathogenesis has been shown in a reciprocal way; germ free or gnotobiotic mice demonstrated considerably reduced susceptibility to experimental autoimmune encephalomyelitis (EAE), a MS model, while CNS disease may affect gut homeostasis. The gut-brain connection is largely supported by evidence on individuals with MS (133, 134).
Th17 cells are normally required for host defense against invaders, but they may have vicious effects in terms of autoimmunity, also suggested by their increased numbers in MS. The role of gut microbiota in Th17 cells and their IL-17 product, and their increased numbers of the latter ones in MS have been reviewed in detail (135, 136). On top of that, the main role of Tregs is the suppression of autoreactive T cells, thus maintaining peripheral tolerance. Although abundant, demonstrate reduced function in MS patients. Taken together, along with the outcome of several studies regarding gut microbiota composition in CNS patients, the gut of MS individuals is characterized by microbial dysbiosis, i.e. impaired intestinal microbiota (131). It is widely accepted that dietary habits affect the composition of human microbiota. High-fiber foods enhance gut populations of the Firmicutes and Bacteroidetes phyla, which produce short-chain fatty acids (SCFAs), which, in turn, suppress inflammation via Treg induction (137).
In 2016, the findings of a small, randomized control trial (RCT) of a Mediterranean-style dietary intervention for MS were published (138). Thirty-three Relapsing–Remitting MS (RRMS) patients were randomized into three groups: group 1 undertook a dietary intervention with vitamin D supplementation, group 2 started vitamin D without a dietary intervention, and group 3 underwent a dietary intervention with vitamin D and other nutritional supplements, including a multivitamin and fish oil supplement (138). The study failed to find a significant effect of the intervention on the Expanded Disability Status Score (EDSS) or the Fatigue Severity Scale (FSS) (138). Published data evaluating adherence to the Mediterranean Diet pattern, assessed using alternate MedDiet score, and the risk of an initial clinical diagnosis of the precursor of multiple sclerosis (MS), specifically central nervous demyelination (139). The researchers found that a Mediterranean diet, included unprocessed red meat, was associated with a reduced risk of demyelination in the Australian adult population. Their conclusion suggested that integrating unprocessed red meat into a Mediterranean diet might bring about beneficial effects for individuals at a high risk of multiple sclerosis (MS).
Adherence in MedDiet and its effects on microbiome related to autoimmune diseases: the case of systemic lupus erythematosus (SLE)
Apart from MS and RA, there are other autoimmune diseases related with the gut microbiota, as Systemic Lupus Erythematosus (SLE), and the Inflammatory Bowel Disease (140–142). Concerning Systemic Lupus Erythematosus, there are conflicting results from a limited number of studies indicating a smaller Firmicutes-to-Bacteroidetes (F:B) ratio compared to healthy individuals. It appears that this ratio is not well-established as a clear cause or consequence, as it could be both simultaneously (143). Howbeit, Enterococcus gallinarum, member of gut commensals of the Firmicutes, appears to promote a lupus-like disease (140) In Inflammatory Bowel Disease (IBD), specifically in ulcerative colitis (UC) patients, Faecalibacterium prausnitzii is reduced in feces. In Crohn’s disease biopsies, Faecalibacterium prausnitzii (F. prausnitzii) is also detected (143).
In conclusion, upcoming research focusing on the influence of specific components or the entirety of the Mediterranean diet in ameliorating microbial dysbiosis and mitigating autoimmune-related inflammatory reactions holds significant importance. The diet seems to influence components of both innate and adaptive immunity through a myriad of mechanisms, either independently or collaboratively.
Restoring microbiota through ‘nutraceuticals’
Nutraceuticals, a term coined by their “Godfather” Dr. Stephen De Felice in 1989, embody a fusion of nutritional and pharmaceutical concepts. They encompass products that are isolated or purified from foods. Established nutraceuticals include probiotics, prebiotics, omega-3 and -6 fatty acids, and others like polyphenols, phytoestrogens, flavonoids and antioxidants, with already recognized favorable effects under specific conditions (143–148). Nutraceuticals are “related” to the human microbiota that includes 6 taxonomic bacterial phyla with Firmicutes and Bacteriodetes occupying the 90% of the host’s colonized areas (149). Nutraceuticals include probiotics, which WHO defines as “live micro-organisms” which, when administered in adequate amounts, confer a health benefit on the host.
Probiotics produce short chain fatty acids (SCFAs), which are able to restore both population numbers and diversity of microbiota. Lactobacillus species may decrease or even prevent the symptoms of antibiotic associated diarrhea (AAD) (150), while a meta-analysis study showed positive effect of probiotics on AAD (151). Furthermore, Lactobacillus, Bifidobacterium or Escherichia coli species have positive impact on host against metabolic diseases or gastrointestinal disorders (149).
Prebiotics, on the other hand, are “dietary carbohydrates stimulating the development of gut bacteria or probiotics post external administration, having advantageous results on the host” (152). Breakdown of carbohydrates supply the body with SCFAs, − acetate, propionate and butyrate- which have a beneficial role on the composition and diversity of human microbiota (153). Short-chain fatty acids (SCFAs) have immunomodulatory effects, but the underlying mechanisms are not well understood. SCFAs promote the differentiation of Tregs and protect the integrity of the gut barrier function (153). Also, these fatty acids may regulate immune procedures through G-protein coupled receptors (GPCRs). Gut homeostasis is critically correlated to these specific metabolites as acetate, butyrate and propionate and its disruption could lead to Inflammation Bowel Disease (IBD). SCFAs are very promising interventions for IBD therapy as they reinforce the gut barrier (153). Another important epigenetic factor is the family of histone deacetylase (HDACs) and particularly ADACs inhibitors coming from diet components for example chrysin found in fruits, vegetables, olive oil and red wine (154, 155). Experimental studies present models with HDAC inhibitors for treating T-cell mediated autoimmune diseases (156). Yet, in IBD again particular diet compounds from MedDiet may regulate epigenetic changes to diminish inflammation and the cancer risk (157).
Moreover, the advantageous impact of prebiotics on host health comprises alterations in gut microbiota composition, immune host capacity, energy production, enhanced mineral absorption and better functions of the intestinal barrier (158). Administration of inulin or Fructo-oligosaccharides (FOS) on cancer patients demonstrated a positive impact of these components on gut populations of Lactobacillus and Bifidobacteria (159). Ingredients in onions and garlic may have beneficial effect on particular gut microbial species populations, and, on the other hand, lethal effects on pathogens like E. coli and S. aureus (160).
Phytoestrogens include flavonoids, which regulate the intestinal barrier and own antimicrobial effect against pathogens, being characterized as alternative antibiotics (161). Anthocyanins and flavonoids, found in fruits like grapes and apples, may prevent a wide range of diseases (162). Polyphenols, as quercetin, found in apples, grapes, onions, tomatoes, nuts and seeds, alters the gut microbiota in overweight mice fed with high-fat sucrose diet (163). Resveratrol, another polyphenol taken as a food supplement regulates gut microbiota dysbiosis caused by high-fat diet, by enhancing growth of Lactobacillus and Bifidobacterium, raising the ratio of Bacteroidetes/Firmicutes and by hampering growth of E. faecalis (164). Carvacrol and thymol, phenols in the aromatic plant Oregano vulgare, have antibacterial properties and ability to affect the gut microbiota and the immune status in animal models (165–167). Use of omega-3 and -6 polyunsaturated fatty acids, as supplements, alter gut microbiota composition, by increasing Bacteroidetes/Firmicutes ratio, restrains growth of pathogenic bacteria like Helicobacter, Firmicutes, Pseudomonas sp., thus, avoiding immunological disturbances (165).
Conclusion
Taking together, there appears to be a strong correlation between dietary habits, whether in the form of foods or dietary supplements like nutraceuticals, the modulation of gut microbiota, and the critical role of human commensals in disease prevention and regulation. This extends to the onset and development of various immunological disturbances, including autoimmune and metabolic diseases. The Mediterranean diet is based on fruits, vegetables, seeds, nuts, fish, whose composition is rich in prebiotics, phytoestrogens as flavonoids, polyphenols and omega −3 and − 6 polyunsaturated fats. Several studies have established a correlation between Mediterranean dietary habits and favorable effects on gut microbiota composition. This association suggests a potential positive impact on the risk and progression of inflammatory diseases. Consequently, it could be critically important to advocate for the adoption or reinforcement of Mediterranean dietary habits. Such habits may contribute to the modulation of microbial homeostasis, mitigate the effects of pathogen invasions, and influence the inflammatory pathway, ultimately benefiting health. However, further studies, on its individual components and/or total/overall compounds of the Mediterranean diet “philosophy” on health’s benefit via microbiota homeostasis should be conducted, to address the effect of the diet and its constituents in the prevention and clinical management of patients affect with specific autoimmune disorders.
Author contributions
CT: Conceptualization, Data curation, Writing – original draft. AvT: Data curation, Resources, Writing – original draft. ES: Writing – review & editing. TK: Data curation, Resources, Software, Writing – review & editing. EZ: Investigation, Resources, Software, Writing – review & editing. ED: Formal analysis, Investigation, Project administration, Writing – review & editing. AlT: Formal analysis, Methodology, Validation, Writing – review & editing. DB: Supervision, Validation, Writing – original draft.
Funding
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abbreviations
ACPA, Anti-citrullinated protein antibody; APCs, Antigen-presenting cells; CRP, C-reactive protein; DMARD, Disease-modifying anti-Rheumatic Drug; EDSS, Expanded Disability Status Score; GNS, Glucosamine (N-acetyl)-6-Sulfatase; MedDiet, Mediterranean Diet; MS, Multiple sclerosis; NGS, Next Generation Sequencing; NMO, Neuromyelitis optica; RA, Rheumatoid arthritis; TNF, Tumor necrosis factor; TNF-α, Tumor necrosis factor alpha; SCFA, Short-Chain Fatty Acids.
References
1. Willett, WC, Sacks, F, Trichopoulou, A, Drescher, G, Ferro-Luzzi, A, Helsing, E, et al. Mediterranean diet pyramid: a cultural model for healthy eating. Am J Clin Nutr. (1995) 61:1402S–6S. doi: 10.1093/ajcn/61.6.1402S
2. Gifford, KD . Dietary fats, eating guides, and public policy: history, critique, and recommendations. Am J Med. (2002) 113:89S–106S. doi: 10.1016/s0002-9343(01)00996-2
3. Senghor, B, Sokhna, C, Ruimy, R, and Lagier, J-C. Gut microbiota diversity according to dietary habits and geographical provenance. Hum Microbiome J. (2018) 7-8:1–9. doi: 10.1016/j.humic.2018.01.001
4. Kau, AL, Ahern, PP, Griffin, NW, Goodman, AL, and Gordon, JI. Human nutrition, the gut microbiome and the immune system. Nature. (2011) 474:327–36. doi: 10.1038/nature10213
5. Li, B, Selmi, C, Tang, R, Gershwin, ME, and Ma, X. The microbiome and autoimmunity: a paradigm from the gut-liver axis. Cell Mol Immunol. (2018) 15:595–609. doi: 10.1038/cmi.2018.7
6. Vieira, SM, Pagovich, OE, and Kriegel, MA. Diet, microbiota and autoimmune diseases. Lupus. (2014) 23:518–26. doi: 10.1177/0961203313501401
7. Forsyth, C, Kouvari, M, D’Cunha, NM, Georgousopoulou, EN, Panagiotakos, DB, Mellor, DD, et al. The effects of the Mediterranean diet on rheumatoid arthritis prevention and treatment: a systematic review of human prospective studies. Rheumatol Int. (2018) 38:737–47. doi: 10.1007/s00296-017-3912-1
8. Stamostergiou, J, Theodoridis, X, Ganochoriti, V, Bogdanos, DP, and Sakkas, LI. The role of the Mediterranean diet in hyperuricemia and gout. MJR. (2018) 29:21–5. doi: 10.31138/mjr.29.1.21
9. Estruch, R, Ros, E, Salas-Salvadó, J, Covas, M-I, Corella, D, Arós, F, et al. Primary prevention of cardiovascular disease with a Mediterranean diet supplemented with extra-virgin olive oil or nuts. N Engl J Med. (2018) 378:389. doi: 10.1056/NEJMoa1800389
10. Martínez-González, MA, de la Fuente-Arrillaga, C, Nunez-Cordoba, JM, Basterra- Gortari, FJ, Beunza, JJ, Vazquez, Z, et al. Adherence to Mediterranean diet and risk of developing diabetes: prospective cohort study. BMJ. (2008) 336:1348–51. doi: 10.1136/bmj.39561.501007.BE
11. Echeverría, G, McGee, EE, Urquiaga, I, Jiménez, P, D’Acuña, S, Villarroel, L, et al. Inverse associations between a locally validated Mediterranean diet index, overweight/obesity, and metabolic syndrome in Chilean adults. Nutrients. (2017) 9:862. doi: 10.3390/nu9080862
12. Kastorini, C-M, Milionis, HJ, Esposito, K, Giugliano, D, Goudevenos, JA, and Panagiotakos, DB. The effect of Mediterranean diet on metabolic syndrome and its components: a meta-analysis of 50 studies and 534,906 individuals. J Am Coll Cardiol. (2011) 57:1299–313. doi: 10.1016/j.jacc.2010.09.073
13. Bonaccio, M, Di Castelnuovo, A, Costanzo, S, Gialluisi, A, Persichillo, M, Cerletti, C, et al. Mediterranean diet and mortality in the elderly: a prospective cohort study and a meta-analysis. Br J Nutr. (2018) 120:841–54. doi: 10.1017/S0007114518002179
14. Godos, J, Ferri, R, Caraci, F, Cosentino, FII, Castellano, S, Shivappa, N, et al. Dietary inflammatory index and sleep quality in southern Italian adults. Nutrients. (2019) 11:1324. doi: 10.3390/nu11061324
15. Jennings, A, Cashman, KD, Gillings, R, Cassidy, A, Tang, J, Fraser, W, et al. A Mediterranean-like dietary pattern with vitamin D3 (10 μg/d) supplements reduced the rate of bone loss in older Europeans with osteoporosis at baseline: results of a 1-y randomized controlled trial. Am J Clin Nutr. (2018) 108:633–40. doi: 10.1093/ajcn/nqy122
16. Leung, CW, Fung, TT, McEvoy, CT, Lin, J, and Epel, ES. Diet quality indices and leukocyte telomere length among healthy US adults: data from the National Health and nutrition examination survey, 1999-2002. Am J Epidemiol. (2018) 187:2192–201. doi: 10.1093/aje/kwy124
17. Pagliai, G, Sofi, F, Vannetti, F, Caiani, S, Pasquini, G, Molino Lova, R, et al. Mediterranean diet, food consumption and risk of late-life depression: the Mugello study. J Nutr Health Aging. (2018) 22:569–74. doi: 10.1007/s12603-018-1019-3
18. Ma, J, Hennein, R, Liu, C, Long, MT, Hoffmann, U, Jacques, PF, et al. Improved diet quality associates with reduction in liver fat, particularly in individuals with high genetic risk scores for nonalcoholic fatty liver disease. Gastroenterology. (2018) 155:107–17. doi: 10.1053/j.gastro.2018.03.038
19. Castelló, A, Boldo, E, Amiano, P, Castaño-Vinyals, G, Aragonés, N, Gómez-Acebo, I, et al. Mediterranean dietary pattern is associated with low risk of aggressive prostate Cancer: MCC-Spain study. J Urol. (2018) 199:430–7. doi: 10.1016/j.juro.2017.08.087
20. Haslam, A, Robb, SW, Hébert, JR, Huang, H, and Ebell, MH. Greater adherence to a Mediterranean diet is associated with lower prevalence of colorectal adenomas in men of all races. Nutr Res N Y N. (2017) 48:76–84. doi: 10.1016/j.nutres.2017.10.003
21. Assaf-Balut, C, García de la Torre, N, Durán, A, Fuentes, M, Bordiú, E, Del Valle, L, et al. A Mediterranean diet with additional extra virgin olive oil and pistachios reduces the incidence of gestational diabetes mellitus (GDM): a randomized controlled trial: the St. Carlos GDM prevention study. PloS One. (2017) 12:185873. doi: 10.1371/journal.pone.0185873
22. Sotos-Prieto, M, Bhupathiraju, SN, Mattei, J, Fung, TT, Li, Y, Pan, A, et al. Association of Changes in diet quality with Total and cause- specific mortality. N Engl J Med. (2017) 377:143–53. doi: 10.1056/NEJMoa1613502
23. Crous-Bou, M, Fung, TT, Prescott, J, Julin, B, Du, M, Sun, Q, et al. Mediterranean diet and telomere length in nurses’ health study: population based cohort study. BMJ. (2014) 349:g6674. doi: 10.1136/bmj.g6674
24. Relman, DA . Microbiology: learning about who we are. Nature. (2012) 486:194–5. doi: 10.1038/486194a
25. Berg, G, Rybakova, D, Fischer, D, Cernava, T, Vergès, M, Charles, T, et al. Microbiome definition re-visited: old concepts and new challenges. Microbiome. (2020) 8:1–22. doi: 10.1186/s40168-020-00875-0
26. Whipps, JM, Lewis, K, and Cooke, RC. Mycoparasitism and plant disease control 161–187 In: NM Burge , editor. Fungi in biological control systems. Manchester: Manchester University Press (1988). 176.
27. Muegge, BD, Kuczynski, J, Knights, D, Clemente, JC, González, A, Fontana, L, et al. Diet drives convergence in gut microbiome functions across mammalian phylogeny and within humans. Science. (2011) 332:970–4. doi: 10.1126/science.1198719
28. Ley, RE, Hamady, M, Lozupone, C, Turnbaugh, PJ, Ramey, RR, Bircher, JS, et al. Evolution of mammals and their gut microbes. Science. (2008) 320:1647–51. doi: 10.1126/science.1155725
29. Kashtanova, DA, Popenko, AS, Tkacheva, ON, Tyakht, AB, Alexeev, DG, and Boytsov, SA. Association between the gut microbiota and diet: fetal life, early childhood, and further life. Nutr Burbank Los Angel Cty Calif. (2016) 32:620–7. doi: 10.1016/j.nut.2015.12.037
30. Houghton, D, Stewart, CJ, Day, CP, and Trenell, M. Gut microbiota and lifestyle interventions in NAFLD. Int J Mol Sci. (2016) 17:447. doi: 10.3390/ijms17040447
31. Murphy, EF, Cotter, PD, Healy, S, Marques, TM, O’Sullivan, O, Fouhy, F, et al. Composition and energy harvesting capacity of the gut microbiota: relationship to diet, obesity and time in mouse models. Gut. (2010) 59:1635–42. doi: 10.1136/gut.2010.215665
32. Turnbaugh, PJ, Bäckhed, F, Fulton, L, and Gordon, JI. Diet-induced obesity is linked to marked but reversible alterations in the mouse distal gut microbiome. Cell Host Microbe. (2008) 3:213–23. doi: 10.1016/j.chom.2008.02.015
33. Hildebrandt, MA, Hoffmann, C, Sherrill-Mix, SA, Keilbaugh, SA, Hamady, M, Chen, Y-Y, et al. High-fat diet determines the composition of the murine gut microbiome independently of obesity. Gastroenterology. (2009) 137:e1–2. doi: 10.1053/j.gastro.2009.08.042
34. Daniel, H, Gholami, AM, Berry, D, Desmarchelier, C, Hahne, H, Loh, G, et al. High-fat diet alters gut microbiota physiology in mice. ISME J. (2014) 8:295–308. doi: 10.1038/ismej.2013.155
35. de Wit, NJW, Afman, LA, Mensink, M, and Müller, M. Phenotyping the effect of diet on non-alcoholic fatty liver disease. J Hepatol. (2012) 57:1370–3. doi: 10.1016/j.jhep.2012.07.003
36. Mujico, JR, Baccan, GC, Gheorghe, A, Díaz, LE, and Marcos, A. Changes in gut microbiota due to supplemented fatty acids in diet-induced obese mice. Br J Nutr. (2013) 110:711–20. doi: 10.1017/S0007114512005612
37. McAllan, L, Skuse, P, Cotter, PD, O’Connor, P, Cryan, JF, Ross, RP, et al. Protein quality and the protein to carbohydrate ratio within a high fat diet influences energy balance and the gut microbiota in C57BL/6J mice. PLoS One. (2014) 9:e88904. doi: 10.1371/journal.pone.0088904
38. Hansen, LBS, Roager, HM, Søndertoft, NB, Gøbel, RJ, Kristensen, M, Vallès- Colomer, M, et al. A low-gluten diet induces changes in the intestinal microbiome of healthy Danish adults. Nat Commun. (2018) 9:4630. doi: 10.1038/s41467-018-07019-x
39. Ferrocino, I, Di Cagno, R, De Angelis, M, Turroni, S, Vannini, L, Bancalari, E, et al. Fecal microbiota in healthy subjects following omnivore, vegetarian and vegan diets: Culturable populations and rRNA DGGE profiling. PLoS One. (2015) 10:e0128669. doi: 10.1371/journal.pone.0128669
40. Rothschild, D, Weissbrod, O, Barkan, E, Kurilshikov, A, Korem, T, Zeevi, D, et al. Environment dominates over host genetics in shaping human gut microbiota. Nature. (2018) 555:210–5. doi: 10.1038/nature25973
41. Obregon-Tito, AJ, Tito, RY, Metcalf, J, Sankaranarayanan, K, Clemente, JC, Ursell, LK, et al. Subsistence strategies in traditional societies distinguish gut microbiomes. Nat Commun. (2015) 6:6505. doi: 10.1038/ncomms7505
42. Vangay, P, Johnson, AJ, Ward, TL, Al-Ghalith, GA, Shields-Cutler, RR, Hillmann, BM, et al. US immigration westernizes the human gut microbiome. Cell. (2018) 175:962–972.e10. doi: 10.1016/j.cell.2018.10.029
43. O’Hara, AM, and Shanahan, F. The gut flora as a forgotten organ. EMBO Rep. (2006) 7:688–93. doi: 10.1038/sj.embor.7400731
44. Ursell, LK, Haiser, HJ, Van Treuren, W, Garg, N, Reddivari, L, Vanamala, J, et al. The intestinal metabolome: an intersection between microbiota and host. Gastroenterology. (2014) 146:1470–6. doi: 10.1053/j.gastro.2014.03.001
45. Cash, HL, Whitham, CV, Behrendt, CL, and Hooper, LV. Symbiotic bacteria direct expression of an intestinal bactericidal lectin. Science. (2006) 313:1126–30. doi: 10.1126/science.1127119
46. Gill, SR, Pop, M, Deboy, RT, Eckburg, PB, Turnbaugh, PJ, Samuel, BS, et al. Metagenomic analysis of the human distal gut microbiome. Science. (2006) 312:1355–9. doi: 10.1126/science.1124234
47. Hooper, LV, Stappenbeck, TS, Hong, CV, and Gordon, JI. Angiogenins: a new class of microbicidal proteins involved in innate immunity. Nat Immunol. (2003) 4:269–73. doi: 10.1038/ni888
48. Rakoff-Nahoum, S, and Medzhitov, R. Innate immune recognition of the indigenous microbial flora. Mucosal Immunol. (2008) 1:S10–4. doi: 10.1038/mi.2008.49
49. Macpherson, AJ, and Harris, NL. Interactions between commensal intestinal bacteria and the immune system. Nat Rev Immunol. (2004) 4:478–85. doi: 10.1038/nri1373
50. Thaiss, CA, Zmora, N, Levy, M, and Elinav, E. The microbiome and innate immunity. Nature. (2016) 535:65–74. doi: 10.1038/nature18847
51. Miller, FW, Pollard, KM, Parks, CG, Germolec, DR, Leung, PSC, Selmi, C, et al. Criteria for environmentally associated autoimmune diseases. J Autoimmun. (2012) 39:253–8. doi: 10.1016/j.jaut.2012.05.001
52. Ramos-Casals, M, Brito-Zerón, P, Kostov, B, Sisó-Almirall, A, Bosch, X, Buss, D, et al. Google-driven search for big data in autoimmune geoepidemiology: analysis of 394,827 patients with systemic autoimmune diseases. Autoimmun Rev. (2015) 14:670–9. doi: 10.1016/j.autrev.2015.03.008
53. Chervonsky, AV . Microbiota and autoimmunity. Cold Spring Harb Perspect Biol. (2013) 5:a007294. doi: 10.1101/cshperspect.a007294
54. Belkaid, Y, and Hand, TW. Role of the microbiota in immunity and inflammation. Cell. (2014) 157:121–41. doi: 10.1016/j.cell.2014.03.011
55. Shamriz, O, Mizrahi, H, Werbner, M, Shoenfeld, Y, Avni, O, and Koren, O. Microbiota at the crossroads of autoimmunity. Autoimmun Rev. (2016) 15:859–69. doi: 10.1016/j.autrev.2016.07.012
56. De Luca, F, and Shoenfeld, Y. The microbiome in autoimmune diseases. Clin Exp Immunol. (2019) 195:74–85. doi: 10.1111/cei.13158
57. Agmon-Levin, N, Ram, M, Barzilai, O, Porat-Katz, BS, Parikman, R, Selmi, C, et al. Prevalence of hepatitis C serum antibody in autoimmune diseases. J Autoimmun. (2009) 32:261–6. doi: 10.1016/j.jaut.2009.02.017
58. Getts, DR, Chastain, EML, Terry, RL, and Miller, SD. Virus infection, antiviral immunity, and autoimmunity. Immunol Rev. (2013) 255:197–209. doi: 10.1111/imr.12091
59. Guilherme, L, Kalil, J, and Cunningham, M. Molecular mimicry in the autoimmune pathogenesis of rheumatic heart disease. Autoimmunity. (2006) 39:31–9. doi: 10.1080/08916930500484674
60. Vojdani, A . A potential link between environmental triggers and autoimmunity. Autoimmune Dis. (2014) 2014:437231:1–18. doi: 10.1155/2014/437231
61. Vanderlugt, CJ, and Miller, SD. Epitope spreading. Curr Opin Immunol. (1996) 8:831–6. doi: 10.1016/S0952-7915(96)80012-4
62. Vanderlugt, C, and Miller, S. Epitope spreading in immune-mediated diseases: implications for immunotherapy. Nat Rev Immunol. (2002) 2:85–95. doi: 10.1038/nri724
63. Kaul, G, and Thippeswamy, H. Role of heat shock proteins in diseases and their therapeutic potential. Indian J Microbiol. (2011) 51:124–31. doi: 10.1007/s12088-011-0147-9
64. Cunningham, MW . Pathogenesis of group a streptococcal infections. Clin Microbiol Rev. (2000) 13:470–511. doi: 10.1128/CMR.13.3.470
65. Bogdanos, DP, and Sakkas, LI. From microbiome to infectome in autoimmunity. Curr Opin Rheumatol. (2017) 29:369–73. doi: 10.1097/BOR.0000000000000394
66. Polymeros, D, Tsiamoulos, ZP, Koutsoumpas, AL, Smyk, DS, Mytilinaiou, MG, Triantafyllou, K, et al. Bioinformatic and immunological analysis reveals lack of support for measles virus related mimicry in Crohn’s disease. BMC Med. (2014) 12:139. doi: 10.1186/s12916-014-0139-9
67. Ehser, J, Holdener, M, Christen, S, Bayer, M, Pfeilschifter, JM, Hintermann, E, et al. Molecular mimicry rather than identity breaks T-cell tolerance in the CYP2D6 mouse model for human autoimmune hepatitis. J Autoimmun. (2013) 42:39–49. doi: 10.1016/j.jaut.2012.11.001
68. Koutsoumpas, A, Polymeros, D, Tsiamoulos, Z, Smyk, D, Karamanolis, G, Triantafyllou, K, et al. Peculiar antibody reactivity to human connexin 37 and its microbial mimics in patients with Crohn’s disease. J Crohns Colitis. (2011) 5:101–9. doi: 10.1016/j.crohns.2010.10.009
69. Koutsoumpas, A, Mytilinaiou, M, Polymeros, D, Dalekos, GN, and Bogdanos, DP. Anti- Helicobacter pylori antibody responses specific for VacA do not trigger primary biliary cirrhosis-specific antimitochondrial antibodies. Eur J Gastroenterol Hepatol. (2009) 21:1220. doi: 10.1097/MEG.0b013e32831a4807
70. Bogdanos, D-P, and Rigopoulou, EI. Viral/self-mimicry and immunological cross- reactivity as a trigger of hepatic C virus associated autoimmune diabetes. Diabetes Res Clin Pract. (2007) 77:155–6. doi: 10.1016/j.diabres.2006.10.012
71. Bogdanos, D-P, Koutsoumpas, A, Baum, H, and Vergani, D. Borrelia Burgdorferi: a new self-mimicking trigger in primary biliary cirrhosis. Dig Liver Dis. (2006) 38:781–2; author reply 782-783. doi: 10.1016/j.dld.2006.05.010
72. Polymeros, D, Bogdanos, DP, Day, R, Arioli, D, Vergani, D, and Forbes, A. Does cross- reactivity between mycobacterium avium paratuberculosis and human intestinal antigens characterize Crohn’s disease? Gastroenterology. (2006) 131:85–96. doi: 10.1053/j.gastro.2006.04.021
73. Bogdanos, D-P, Smith, H, Ma, Y, Baum, H, Mieli-Vergani, G, and Vergani, D. A study of molecular mimicry and immunological cross-reactivity between hepatitis B surface antigen and myelin mimics. Clin Dev Immunol. (2005) 12:217–24. doi: 10.1080/17402520500285247
74. Muratori, L, Bogdanos, DP, Muratori, P, Lenzi, M, Granito, A, Ma, Y, et al. Susceptibility to thyroid disorders in hepatitis C. Clin Gastroenterol Hepatol. (2005) 3:595–603. doi: 10.1016/s1542-3565(05)00018-2
75. Bogdanos, D-P, Baum, H, Gunsar, F, Arioli, D, Polymeros, D, Ma, Y, et al. Extensive homology between the major immunodominant mitochondrial antigen in primary biliary cirrhosis and Helicobacter pylori does not lead to immunological cross-reactivity. Scand J Gastroenterol. (2004) 39:981–7. doi: 10.1080/00365520410003236
76. Bogdanos, D-P, Pares, A, Baum, H, Caballeria, L, Rigopoulou, EI, Ma, Y, et al. Disease-specific cross-reactivity between mimicking peptides of heat shock protein of Mycobacterium gordonae and dominant epitope of E2 subunit of pyruvate dehydrogenase is common in Spanish but not British patients with primary biliary cirrhosis. J Autoimmun. (2004) 22:353–62. doi: 10.1016/j.jaut.2004.03.002
77. Bogdanos, D-P, Lenzi, M, Okamoto, M, Rigopoulou, EI, Muratori, P, Ma, Y, et al. Multiple viral/self immunological cross-reactivity in liver kidney microsomal antibody positive hepatitis C virus infected patients is associated with the possession of HLA B51. Int J Immunopathol Pharmacol. (2004) 17:83–92. doi: 10.1177/039463200401700112
78. Bogdanos, D-P, Baum, H, Grasso, A, Okamoto, M, Butler, P, Ma, Y, et al. Microbial mimics are major targets of crossreactivity with human pyruvate dehydrogenase in primary biliary cirrhosis. J Hepatol. (2004) 40:31–9. doi: 10.1016/s0168-8278(03)00501-4
79. Gregorio, GV, Choudhuri, K, Ma, Y, Pensati, P, Iorio, R, Grant, P, et al. Mimicry between the hepatitis C virus polyprotein and antigenic targets of nuclear and smooth muscle antibodies in chronic hepatitis C virus infection. Clin Exp Immunol. (2003) 133:404–13. doi: 10.1046/j.1365-2249.2003.02229.x
80. Bogdanos, D-P, Baum, H, Sharma, UC, Grasso, A, Ma, Y, Burroughs, AK, et al. Antibodies against homologous microbial caseinolytic proteases P characterise primary biliary cirrhosis. J Hepatol. (2002) 36:14–21. doi: 10.1016/s0168-8278(01)00252-5
81. Bogdanos, DP, Choudhuri, K, and Vergani, D. Molecular mimicry and autoimmune liver disease: virtuous intentions, malign consequences. Liver. (2001) 21:225–32. doi: 10.1034/j.1600-0676.2001.021004225.x
82. Baum, H, Bogdanos, DP, and Vergani, D. Antibodies to Clp protease in primary biliary cirrhosis: possible role of a mimicking T-cell epitope. J Hepatol. (2001) 34:785–7. doi: 10.1016/s0168-8278(01)00059-9
83. Bogdanos, DP, Mieli-Vergani, G, and Vergani, D. Virus, liver and autoimmunity. Dig Liver Dis. (2000) 32:440–6. doi: 10.1016/s1590-8658(00)80266-2
84. Fujinami, RS, von Herrath, MG, Christen, U, and Whitton, JL. Molecular mimicry, bystander activation, or viral persistence: infections and autoimmune disease. Clin Microbiol Rev. (2006) 19:80–94. doi: 10.1128/CMR.19.1.80-94.2006
85. Kopf, JC, Suhr, MJ, Clarke, J, Eyun, S-I, Riethoven, J-JM, Ramer-Tait, AE, et al. Role of whole grains versus fruits and vegetables in reducing subclinical inflammation and promoting gastrointestinal health in individuals affected by overweight and obesity: a randomized controlled trial. Nutr J. (2018) 17:72. doi: 10.1186/s12937-018-0381-7
86. Koloverou, E, Panagiotakos, DB, Pitsavos, C, Chrysohoou, C, Georgousopoulou, EN, Grekas, A, et al. Adherence to Mediterranean diet and 10-year incidence (2002-2012) of diabetes: correlations with inflammatory and oxidative stress biomarkers in the ATTICA cohort study. Diabetes Metab Res Rev. (2016) 32:73–81. doi: 10.1002/dmrr.2672
87. Yan, S, Kotschenreuther, K, Deng, S, and Kofler, DM. Regulatory T cells in rheumatoid arthritis: functions, development, regulation, and therapeutic potential. Cell Mol Life Sci. (2022) 79:533. doi: 10.1007/s00018-022-04563-0
88. da Fonseca, LJS, Nunes-Souza, V, Goulart, MOF, and Rabelo, LA. Oxidative stress in rheumatoid arthritis: what the future might hold regarding novel biomarkers and add-on therapies. Oxidative Med Cell Longev. (2019) 2019:7536805–16. doi: 10.1155/2019/7536805
89. Neale, EP, Batterham, MJ, and Tapsell, LC. Consumption of a healthy dietary pattern results in significant reductions in C-reactive protein levels in adults: a meta-analysis. Nut Res N Y N. (2016) 36:391–401. doi: 10.1016/j.nutres.2016.02.009
90. Sköldstam, L, Hagfors, L, and Johansson, G. An experimental study of a Mediterranean diet intervention for patients with rheumatoid arthritis. Ann Rheum Dis. (2003) 62:208–14. doi: 10.1136/ard.62.3.208
91. Porras, M, Rada, G, and Durán, J. Effects of Mediterranean diet on the treatment of rheumatoid arthritis. Medwave. (2019) 19:e7640. doi: 10.5867/medwave.2019.05.7639
92. García-Morales, JM, Lozada-Mellado, M, Hinojosa-Azaola, A, Llorente, L, Ogata- Medel, M, Pineda-Juárez, JA, et al. Effect of a dynamic exercise program in combination with Mediterranean diet on quality ofLife in women with rheumatoid arthritis. J Clin Rheumatol Pract Rep Rheum Musculoskelet Dis. (2019) 26:S116–22. doi: 10.1097/RHU.0000000000001064
93. Johansson, K, Askling, J, Alfredsson, L, and Di Giuseppe, D. EIRA study group Mediterranean diet and risk of rheumatoid arthritis: a population-based case-control study. Arthritis Res Ther. (2018) 20:175. doi: 10.1186/s13075-018-1680-2
94. Hu, Y, Costenbader, KH, Gao, X, Hu, FB, Karlson, EW, and Lu, B. Mediterranean diet and incidence of rheumatoid arthritis in women. Arthritis Care Res. (2015) 67:597–606. doi: 10.1002/acr.22481
95. McKellar, G, Morrison, E, McEntegart, A, Hampson, R, Tierney, A, Mackle, G, et al. A pilot study of a Mediterranean-type diet intervention in female patients with rheumatoid arthritis living in areas of social deprivation in Glasgow. Ann Rheum Dis. (2007) 66:1239–43. doi: 10.1136/ard.2006.065151
96. Hagfors, L, Leanderson, P, Sköldstam, L, Andersson, J, and Johansson, G. Antioxidant intake, plasma antioxidants and oxidative stress in a randomized, controlled, parallel, Mediterranean dietary intervention study on patients with rheumatoid arthritis. Nutr J. (2003) 2:5. doi: 10.1186/1475-2891-2-5
97. Hagfors, L, Nilsson, I, Sköldstam, L, and Johansson, G. Fat intake and composition of fatty acids in serum phospholipids in a randomized, controlled, Mediterranean dietary intervention study on patients with rheumatoid arthritis. Nutr Metab. (2005) 2:26. doi: 10.1186/1743-7075-2-26
98. Michalsen, A, Riegert, M, Lüdtke, R, Bäcker, M, Langhorst, J, Schwickert, M, et al. Mediterranean diet or extended fasting’s influence on changing the intestinal microflora, immunoglobulin a secretion and clinical outcome in patients with rheumatoid arthritis and fibromyalgia: an observational study. BMC Complement Altern Med. (2005) 5:22. doi: 10.1186/1472-6882-5-22
99. Pagliai, G, Russo, E, Niccolai, E, Dinu, M, Di Pilato, V, Magrini, A, et al. Influence of a 3-month low-calorie Mediterranean diet compared to the vegetarian diet on human gut microbiota and SCFA: the CARDIVEG study. Eur J Nutr. (2019) 59:2011–24. doi: 10.1007/s00394-019-02050-0
100. Deane, KD, Norris, JM, Steere, AC, and Holers, VM. Association of Antibodies to Prevotella copri in anti-cyclic Citrullinated peptide-positive individuals at risk of developing rheumatoid arthritis and in patients with early or established rheumatoid arthritis. Arthritis Rheumatol. (2023) 75:507–16. doi: 10.1002/art.42370
101. Zhao, T, Wei, Y, Zhu, Y, Xie, Z, Hai, Q, Li, Z, et al. Gut microbiota and rheumatoid arthritis: from pathogenesis to novel therapeutic opportunities. Front Immunol. (2022) 13:1007165. doi: 10.3389/fimmu.2022.1007165
102. Jiang, L, Shang, M, Yu, S, Liu, Y, Zhang, H, Zhou, Y, et al. A high-fiber diet synergizes with Prevotella copri and exacerbates rheumatoid arthritis. Cell Mol Immunol. (2022) 19:1414–24. doi: 10.1038/s41423-022-00934-6
103. Singh, V, Yeoh, BS, Walker, RE, Xiao, X, Saha, P, Golonka, RM, et al. Microbiota fermentation-NLRP3 axis shapes the impact of dietary fibres on intestinal inflammation. Gut. (2019) 68:1801–12. doi: 10.1136/gutjnl-2018-316250
104. Abendroth, A, Michalsen, A, Lüdtke, R, Rüffer, A, Musial, F, Dobos, GJ, et al. Changes of intestinal microflora in patients with rheumatoid arthritis during fasting or a Mediterranean diet. Forsch Komplementarmedizin. (2006) 17:307–13. doi: 10.1159/000322313
105. Gutiérrez-Díaz, I, Fernández-Navarro, T, Sánchez, B, Margolles, A, and González, S. Mediterranean diet and faecal microbiota: a transversal study. Food Funct. (2016) 7:2347–56. doi: 10.1039/c6fo00105j
106. de Filippis, F, Pellegrini, N, Vannini, L, Jeffery, IB, La Storia, A, Laghi, L, et al. High-level adherence to a Mediterranean diet beneficially impacts the gut microbiota and associated metabolome. Gut. (2016) 65:1812–21. doi: 10.1136/gutjnl-2015-309957
107. Mitsou, EK, Kakali, A, Antonopoulou, S, Mountzouris, KC, Yannakoulia, M, Panagiotakos, DB, et al. Adherence to the Mediterranean diet is associated with the gut microbiota pattern and gastrointestinal characteristics in an adult population. Br J Nutr. (2017) 117:1645–55. doi: 10.1017/S0007114517001593
108. De Santis, S, Cavalcanti, E, Mastronardi, M, Jirillo, E, and Chieppa, M. Nutritional keys for intestinal barrier modulation. Front Immunol. (2015) 6:612. doi: 10.3389/fimmu.2015.00612
109. Lerner, A, Patricia, J, and Matthias, T. Nutrients, bugs and us: the short-chain fatty acids story in celiac disease. Int J Celiac Dis. (2016) 4:92–4. doi: 10.12691/ijcd-4-3-12
110. Hui, W, Yu, D, Cao, Z, and Zhao, X. Butyrate inhibit collagen-induced arthritis via Treg/IL-10/Th17 axis. Int Immunopharmacol. (2019) 68:226–33. doi: 10.1016/j.intimp.2019.01.018
111. Obeid, R, Awwad, HM, Keller, M, and Geisel, J. Trimethylamine-N-oxide and its biological variations in vegetarians. Eur J Nutr. (2017) 56:2599–609. doi: 10.1007/s00394-016-1295-9
112. Breban, M, Tap, J, Leboime, A, Said-Nahal, R, Langella, P, Chiocchia, G, et al. Faecal microbiota study reveals specific dysbiosis in spondyloarthritis. Ann Rheum Dis. (2017) 76:1614–22. doi: 10.1136/annrheumdis-2016-211064
113. Scher, JU, Sczesnak, A, Longman, RS, Segata, N, Ubeda, C, Bielski, C, et al. Expansion of intestinal Prevotella copri correlates with enhanced susceptibility to arthritis. eLife. (2013) 2:e01202. doi: 10.7554/eLife.01202
114. Pianta, A, Arvikar, S, Strle, K, Drouin, EE, Wang, Q, Costello, CE, et al. Evidence of the immune relevance of Prevotella copri, a gut microbe, in patients with rheumatoid arthritis. Arthritis Rheumatol. (2017) 69:964–75. doi: 10.1002/art.40003
115. Vaahtovuo, J, Munukka, E, Korkeamäki, M, Luukkainen, R, and Toivanen, P. Fecal microbiota in early rheumatoid arthritis. J Rheumatol. (2008) 35:1500–5.
116. Alpizar-Rodriguez, D, Lesker, TR, Gronow, A, Gilbert, B, Raemy, E, Lamacchia, C, et al. Prevotella copri in individuals at risk for rheumatoid arthritis. Ann Rheum Dis. (2019) 78:590–3. doi: 10.1136/annrheumdis-2018-214514
117. Wells, PM, Williams, FMK, Matey-Hernandez, ML, Menni, C, and Steves, CJ. RA and the microbiome: do host genetic factors provide the link? J Autoimmun. (2019) 99:104–15. doi: 10.1016/j.jaut.2019.02.004
118. Schwenzer, A, Quirke, A-M, Marzeda, AM, Wong, A, Montgomery, AB, Sayles, HR, et al. Association of Distinct Fine Specificities of anti-Citrullinated peptide antibodies with elevated immune responses to Prevotella intermedia in a subgroup of patients with rheumatoid arthritis and periodontitis. Arthritis Rheumatol. (2017) 69:2303–13. doi: 10.1002/art.40227
119. Chung, YL, Rider, LG, Bell, JD, Summers, RM, Zemel, LS, Rennebohm, RM, et al. Muscle metabolites, detected in urine by proton spectroscopy, correlate with disease damage in juvenile idiopathic inflammatory myopathies. Arthritis Rheumatol. (2005) 53:565–70. doi: 10.1002/art.21331
120. Asteriou, E, Gkoutzourelas, A, Mavropoulos, A, Katsiari, C, Sakkas, LI, and Bogdanos, DP. Curcumin for the Management of Periodontitis and Early ACPA-positive rheumatoid arthritis: killing two birds with one Stone. Nutrients. (2018) 10:908. doi: 10.3390/nu10070908
121. Kelepouri, D, Mavropoulos, A, Bogdanos, DP, and Sakkas, LI. The role of flavonoids in inhibiting Th17 responses in inflammatory arthritis. J Immunol Res. (2018) 2018:9324357–11. doi: 10.1155/2018/9324357
122. Sakkas, LI, and Bogdanos, DP. Multiple hit infection and autoimmunity: the dysbiotic microbiota-ACPA connection in rheumatoid arthritis. Curr Opin Rheumatol. (2018) 30:403–9. doi: 10.1097/BOR.0000000000000503
123. Pianta, A, Arvikar, SL, Strle, K, Drouin, EE, Wang, Q, Costello, CE, et al. Two rheumatoid arthritis-specific autoantigens correlate microbial immunity with autoimmune responses in joints. J Clin Invest. (2017) 127:2946–56. doi: 10.1172/JCI93450
124. Obando-Pereda, GA . GAKG-RGEKG an epitope that provokes immune cross- reactivity between Prevotella sp. and human collagen: evidence of molecular mimicry in chronic periodontitis. Autoimmune Dis. (2016) 2016:5472320. doi: 10.1155/2016/5472320
125. McInnes, IB, and Schett, G. The pathogenesis of rheumatoid arthritis. N Engl J Med. (2011) 365:2205–19. doi: 10.1056/NEJMra1004965
126. Oen, K, Postl, B, Chalmers, IM, Ling, N, Schroeder, ML, Baragar, FD, et al. Rheumatic diseases in an Inuit population. Arthritis Rheum. (1986) 29:65–74. doi: 10.1002/art.1780290109
127. Mulvad, G, Pedersen, HS, Hansen, JC, Dewailly, E, Jul, E, Pedersen, M, et al. The Inuit diet. Fatty acids and antioxidants, their role in ischemic heart disease, and exposure to organochlorines and heavy metals. An international study Arctic Med Res. (1996) 55:20–4.
128. Yue, M, Tao, Y, Fang, Y, Lian, X, Zhang, Q, Xia, Y, et al. The gut microbiota modulator berberine ameliorates collagen-induced arthritis in rats by facilitating the generation of butyrate and adjusting the intestinal hypoxia and nitrate supply. FASEB J. (2019) 33:12311–23. doi: 10.1096/fj.201900425RR
129. Garcia-Mantrana, I, Selma-Royo, M, Alcantara, C, and Collado, MC. Shifts on gut microbiota associated to Mediterranean diet adherence and specific dietary intakes on general adult population. Front Microbiol. (2018) 9:890. doi: 10.3389/fmicb.2018.00890
130. Shankar, V, Gouda, M, Moncivaiz, J, Gordon, A, Reo, NV, Hussein, L, et al. Differences in gut metabolites and microbial composition and functions between Egyptian and U.S. Children Are Consistent with Their Diets. mSystems. (2017) 2:169. doi: 10.1128/mSystems.00169-16
131. Colpitts, SL, and Kasper, LH. Influence of the gut microbiome on autoimmunity in the central nervous system. J Immunol. (1950) 198:596–604. doi: 10.4049/jimmunol.1601438
132. Hirschberg, S, Gisevius, B, Duscha, A, and Haghikia, A. Implications of diet and the gut microbiome in Neuroinflammatory and neurodegenerative diseases. Int J Mol Sci. (2019) 20:3109. doi: 10.3390/ijms20123109
133. Berer, K, Mues, M, Koutrolos, M, Rasbi, ZA, Boziki, M, Johner, C, et al. Commensal microbiota and myelin autoantigen cooperate to trigger autoimmune demyelination. Nature. (2011) 479:538–41. doi: 10.1038/nature10554
134. Lee, YK, Menezes, JS, Umesaki, Y, and Mazmanian, SK. Proinflammatory T-cell responses to gut microbiota promote experimental autoimmune encephalomyelitis. Proc Natl Acad Sci USA. (2011) 108:4615–22. doi: 10.1073/pnas.1000082107
135. Dos Passos, GR, Sato, DK, Becker, J, and Fujihara, K. Th17 cells pathways in multiple sclerosis and Neuromyelitis Optica Spectrum disorders: pathophysiological and therapeutic implications. Mediat Inflamm. (2016) 2016:5314541–11. doi: 10.1155/2016/5314541
136. Chewning, JH, and Weaver, CT. Development and survival of Th17 cells within the intestines: the influence of microbiome- and diet-derived signals. J Immunol. (1950) 193:4769–77. doi: 10.4049/jimmunol.1401835
137. Smith, PM, Howitt, MR, Panikov, N, Michaud, M, Gallini, CA, Bohlooly-Y, M, et al. The microbial metabolites, short-chain fatty acids, regulate colonic Treg cell homeostasis. Science. (2013) 341:569–73. doi: 10.1126/science.1241165
138. Riccio, P, Rossano, R, Larocca, M, Trotta, V, Mennella, I, Vitaglione, P, et al. Anti-inflammatory nutritional intervention in patients with relapsing-remitting and primary-progressive multiple sclerosis: a pilot study. Exp Biol Med Maywood NJ. (2016) 241:620–35. doi: 10.1177/1535370215618462
139. Black, LJ, Baker, K, Ponsonby, A-L, van der Mei, I, Lucas, RM, and Pereira, G. Ausimmune Investigator Group a higher Mediterranean diet score, including unprocessed red meat, is associated with reduced risk of central nervous system demyelination in a case-control study of Australian adults. J Nutr. (2019) 149:1385–92. doi: 10.1093/jn/nxz089
140. Ruiz, L, López, P, Suárez, A, Sánchez, B, and Margolles, A. The role of gut microbiota in lupus: what we know in 2018? Expert Rev Clin Immunol. (2018) 14:787–92. doi: 10.1080/1744666X.2018.1519395
141. Luo, XM, Edwards, MR, Mu, Q, Yu, Y, Vieson, MD, Reilly, CM, et al. Gut microbiota in human systemic lupus erythematosus and a mouse model of lupus. Appl Environ Microbiol. (2018) 84:2288. doi: 10.1128/AEM.02288-17
142. Zuo, T, and Ng, SC. The gut microbiota in the pathogenesis and therapeutics of inflammatory bowel disease. Front Microbiol. (2018) 9:2247. doi: 10.3389/fmicb.2018.02247
143. Yamamoto, EA, and Jørgensen, TN. Relationships between vitamin D, gut microbiome, and systemic autoimmunity. Front Immunol. (2020) 10:3141. doi: 10.3389/fimmu.2019.03141
144. Vieira, SM, Hiltensperger, M, Kumar, V, Zegarra-Ruiz, D, Dehner, C, Khan, N, et al. Translocation of a gut pathobiont drives autoimmunity in mice and humans. Science. (2018) 359:1156–61. doi: 10.1126/science.aar7201
145. Chauhan, B, Kumar, G, Kalam, N, and Ansari, SH. Current concepts and prospects of herbal nutraceutical: a review. J Adv Pharm Technol Res. (2013) 4:4–8. doi: 10.4103/2231-4040.107494
147. Woting, A, and Blaut, M. The intestinal microbiota in metabolic disease. Nutrients. (2016) 8:202. doi: 10.3390/nu8040202
148. Park, MS, Kwon, B, Ku, S, and Ji, GE. The efficacy of Bifidobacterium longum BORI and Lactobacillus acidophilus AD031 probiotic treatment in infants with rotavirus infection. Nutrients. (2017) 9:887. doi: 10.3390/nu9080887
149. Johnston, BC, Goldenberg, JZ, Vandvik, PO, Sun, X, and Guyatt, GH. Probiotics for the prevention of pediatric antibiotic-associated diarrhea. Cochrane Database Syst Rev. (2011):CD004827. doi: 10.1002/14651858.CD004827.pub3
150. Bindels, LB, Neyrinck, AM, Salazar, N, Taminiau, B, Druart, C, Muccioli, GG, et al. Non digestible oligosaccharides modulate the gut microbiota to control the development of leukemia and associated Cachexia in mice. PLoS One. (2015) 10:e0131009. doi: 10.1371/journal.pone.0131009
151. Shashkova, T, Popenko, A, Tyakht, A, Peskov, K, Kosinsky, Y, Bogolubsky, L, et al. Agent based modeling of human gut microbiome interactions and perturbations. PLoS One. (2016) 11:e0148386. doi: 10.1371/journal.pone.0148386
152. Luu, M, Pautz, S, Kohl, V, Singh, R, Romero, R, Lucas, S, et al. The short-chain fatty acid pentanoate suppresses autoimmunity by modulating the metabolic-epigenetic crosstalk in lymphocytes. Nat Commun. (2019) 10:760. doi: 10.1038/s41467-019-08711-2
153. Parada Venegas, D, De la Fuente, MK, Landskron, G, González, MJ, Quera, R, Dijkstra, G, et al. Short chain fatty acids (SCFAs)- mediated gut epithelial and immune regulation and its relevance for inflammatory bowel diseases. Front Immunol. (2019) 10:277. doi: 10.3389/fimmu.2019.00277
154. Bassett, SA, and Barnett, MPG. The role of dietary histone deacetylases (HDACs) inhibitors in health and disease. Nutrients. (2014) 6:4273–301. doi: 10.3390/nu6104273
155. Pal-Bhadra, M, Ramaiah, MJ, Reddy, TL, Krishnan, A, Pushpavalli, S, Babu, KS, et al. Plant HDAC inhibitor chrysin arrest cell growth and induce p21WAF1 by altering chromatin of STAT response element in A375 cells. BMC Cancer. (2012) 12:180. doi: 10.1186/1471-2407-12-180
156. Ellmeier, W, and Seiser, C. Histone deacetylase function in CD4+ T cells. Nat Rev Immunol. (2018) 18:617–34. doi: 10.1038/s41577-018-0037-z
157. Ferguson, LR . Nutrigenomics and Nutrigenetics in functional foods and personalized nutrition. Boca Raton, FL: CRC Press (2016). 5369 p.
158. Bron, PA, Kleerebezem, M, Brummer, R-J, Cani, PD, Mercenier, A, MacDonald, TT, et al. Can probiotics modulate human disease by impacting intestinal barrier function? Br J Nutr. (2017) 117:93–107. doi: 10.1017/S0007114516004037
159. García-Peris, P, Velasco, C, Lozano, MA, Moreno, Y, Paron, L, de la Cuerda, C, et al. Effect of a mixture of inulin and fructo-oligosaccharide on Lactobacillus and Bifidobacterium intestinal microbiota of patients receiving radiotherapy: a randomised, double-blind, placebo-controlled trial. Nutr Hosp. (2012) 27:1908–15. doi: 10.3305/nh.2012.27.6.5992
160. Catinean, A, Neag, MA, Muntean, DM, Bocsan, IC, and Buzoianu, AD. An overview on the interplay between nutraceuticals and gut microbiota. PeerJ. (2018) 6:e4465. doi: 10.7717/peerj.4465
161. Iranshahi, M, Rezaee, R, Parhiz, H, Roohbakhsh, A, and Soltani, F. Protective effects of flavonoids against microbes and toxins: the cases of hesperidin and hesperetin. Life Sci. (2015) 137:125–32. doi: 10.1016/j.lfs.2015.07.014
162. Chaiyasut, C, Pengkumsri, N, Sirilun, S, Peerajan, S, Khongtan, S, and Sivamaruthi, BS. Assessment of changes in the content of anthocyanins, phenolic acids, and antioxidant property of Saccharomyces cerevisiae mediated fermented black rice bran. AMB Express. (2017) 7:114. doi: 10.1186/s13568-017-0411-4
163. Li, Y, Yao, J, Han, C, Yang, J, Chaudhry, MT, Wang, S, et al. Quercetin, inflammation and immunity. Nutrients. (2016) 8:167. doi: 10.3390/nu8030167
164. Larrosa, M, Yañéz-Gascón, MJ, Selma, MV, González-Sarrías, A, Toti, S, Cerón, JJ, et al. Effect of a low dose of dietary resveratrol on colon microbiota, inflammation and tissue damage in a DSS-induced colitis rat model. J Agric Food Chem. (2009) 57:2211–20. doi: 10.1021/jf803638d
165. Fournomiti, M, Kimbaris, A, Mantzourani, I, Plessas, S, Theodoridou, I, Papaemmanouil, V, et al. Antimicrobial activity of essential oils of cultivated oregano (Origanum vulgare), sage (Salvia officinalis), and thyme (Thymus vulgaris) against clinical isolates of Escherichia coli, Klebsiella oxytoca, and Klebsiella pneumoniae. Microb Ecol Health Dis. (2015) 26:23289. doi: 10.3402/mehd.v26.23289
166. Widmer, RJ, Flammer, AJ, Lerman, LO, and Lerman, A. The Mediterranean diet, its components, and cardiovascular disease. Am J Med. (2015) 128:229–38. doi: 10.1016/j.amjmed.2014.10.014
Keywords: autoimmune diseases, autoimmunity, dysbiosis, Mediterranean diet, microbiome
Citation: Tsigalou C, Tsolou A, Stavropoulou E, Konstantinidis T, Zafiriou E, Dardiotis E, Tsirogianni A and Bogdanos D (2024) Unraveling the intricate dance of the Mediterranean diet and gut microbiota in autoimmune resilience. Front. Nutr. 11:1383040. doi: 10.3389/fnut.2024.1383040
Edited by:
Bisheng Zhou, University of Illinois Chicago, United StatesReviewed by:
Zyanya Reyes-Castillo, University of Guadalajara, MexicoAli Osman Gürol, Istanbul University, Türkiye
Copyright © 2024 Tsigalou, Tsolou, Stavropoulou, Konstantinidis, Zafiriou, Dardiotis, Tsirogianni and Bogdanos. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Christina Tsigalou, Y3RzaWdhbG9AbWVkLmR1dGguZ3I=; Elisavet Stavropoulou, ZWxpc2FiZXRoLnN0YXZyb3BvdWxvdUBnbWFpbC5jb20=