 Konstancja Grabowska1,2†
Konstancja Grabowska1,2† Mateusz Grabowski1,2†
Mateusz Grabowski1,2† Marta Przybyła1,2
Marta Przybyła1,2 Natalia Pondel1
Natalia Pondel1 Jarosław J. Barski2,3
Jarosław J. Barski2,3 Marta Nowacka-Chmielewska1‡
Marta Nowacka-Chmielewska1‡ Daniela Liśkiewicz1,4,5*‡
Daniela Liśkiewicz1,4,5*‡- 1Laboratory of Molecular Biology, Institute of Physiotherapy and Health Sciences, Academy of Physical Education, Katowice, Poland
- 2Department for Experimental Medicine, Faculty of Medical Sciences in Katowice, Medical University of Silesia, Katowice, Poland
- 3Department of Physiology, Faculty of Medical Sciences in Katowice, Medical University of Silesia, Katowice, Poland
- 4Institute of Diabetes and Obesity, Helmholtz Center Munich, Neuherberg, Germany
- 5German Center for Diabetes Research (DZD), Neuherberg, Germany
As a journal page for full details. The ketogenic diet (KD) has been established as a treatment for epilepsy, but more recently it has been explored as an alternative or add-on therapy for many other diseases ranging from weight loss to neurological disorders. Animal models are widely used in studies investigating the therapeutic effects of the KD as well as underlying mechanisms. Especially in the context of neurological, psychiatric, and neurodevelopmental disorders essential endpoints are assessed by behavioral and motor tests. Here we summarized research evaluating the influence of the KD on cognition, depressive and anxiety-related behaviors, and social and nutritional behaviors of laboratory rodents. Each section contains a brief description of commonly used behavioral tests highlighting their limitations. Ninety original research articles, written in English, performed on mice or rats, providing measurement of blood beta-hydroxybutyrate (BHB) levels and behavioral evaluation were selected for the review. The majority of research performed in various disease models shows that the KD positively impacts cognition. Almost an equal number of studies report a reduction or no effect of the KD on depressive-related behaviors. For anxiety-related behaviors, the majority of studies show no effect. Despite the increasing use of the KD in weight loss and its appetite-reducing properties the behavioral evaluation of appetite regulation has not been addressed in preclinical studies. This review provides an overview of the behavioral effects of nutritional ketosis addressed to a broad audience of scientists interested in the KD field but not necessarily specializing in behavioral tests.
1 Introduction
The ketogenic diet (KD) is a very low-carbohydrate, high-fat, and adequate protein nutritional approach that induces a metabolic shift to the use of ketone bodies as an additional energy source (1, 2). In the 1920s, physicians introduced the KD as a treatment for epilepsy, especially in patients poorly responding to pharmacotherapy. By the end of the XX century, the KD resurfaced, gaining popularity with the general public mainly due to its efficiency in treating obesity (3). Scientific interest in the KD also significantly increased, as illustrated by the fact that a PubMed search for ‘ketogenic diet’ shows 254 results until 2000 and 4,682 hits in the years 2000–2023. As a result of extensive research, it is now well established that, besides the well-known metabolic effects including ketosis and decreased blood glucose levels, the KD influences inflammatory processes, oxidative stress, gut microbiota, and intracellular signaling pathways (4–6). The KD also has a pleiotropic impact on brain functioning, including gene expression (7–9), neurotransmission (10), the level of neurotrophic factors (11, 12), protein phosphorylation (13), and the metabolism of amino acids (14). Due to this multifaceted effect on physiology, the KD has been increasingly investigated as an alternative or add-on therapy for many diseases (15, 16). Findings from large-scale clinical trials remain limited, and animal models are widely utilized in studies investigating the therapeutic effects of the KD as well as underlying mechanisms. Especially in the context of neurological, psychiatric, and neurodevelopmental disorders, essential endpoints are evaluated by behavioral tests, including the assessment of cognitive functions, and behaviors related to anxiety and depression. The examination of social behavior is important in studies related to Autism Spectrum Disorders (ASD). The reduction of appetite is considered crucial for the effectiveness of the KD in treating obesity in humans (17, 18). Therefore, the influence of the ketogenic diet on nutritional behavior is another interesting aspect that can be explored using animal models.
While there are several important considerations when designing behavioral experiments in animal models, one particularly crucial aspect in the KD field is the composition of the chow. Numerous variations of ketogenic chows are employed in animal research, with the most significant distinctions revolving around the macronutrient ratio, source of fat as well as macronutrient and vitamin content (19, 20). Appropriately chosen macronutrient ratio, not only carbohydrate restriction but also adequate protein content, determines the level of ketosis (21). In addition, particular fat content like medium chain triglycerides (MCT) can enhance ketone production (22–24). Since possible nutrient deficiencies resulting from a very restrictive diet are a common adverse effect of KD in humans (25), currently, a lot of attention is given to the composition of a dietary plan. The same should apply to animal models, where proper micronutrient supplementation of the ketogenic chow is critical to avoid adverse effects such as weakness and growth inhibition (26, 27).
Here, we review studies examining the influence of KD on the behavior of laboratory rodents. Included in the review were only studies that reported the level of ketosis, ensuring a minimal requirement in terms of diet composition for achieving nutritional ketosis. The article is organized into sections dedicated to cognition, depressive and anxiety-related behaviors, as well as social and nutritional behavior. Each section begins with a concise explanation of the rationale for assessing specific behaviors within the context of nutritional ketosis. Evaluation of animal behavior, interpretation of results, and translation of the findings to the clinically relevant situation require an understanding of the behavioral tests employed and inherent their limitations. Consequently, each section offers a brief overview of the available methods for evaluating the behavior in question, highlighting both limitations and potential caveats in result interpretation.
2 Search strategy and study selection
The term “ketogenic diet” was searched in PubMed without a year-of-publication restriction identifying 4,918. Only articles written in English were included. Articles were divided among all authors for screening of titles and abstracts in order to select full-text original papers conducted on mice or rats. In this first step of screening 880 publications were included and full texts were independently reviewed by two investigators in order to select studies showing behavioral or functional tests investigating: depressive-, and anxiety-related behaviors, cognition, and social behavior. One hundred and forty-five studies were selected for evaluation of eligibility. Separate screening was performed to identify studies reporting food intake which was discussed as an approximate of nutritional behavior.
This screening resulted in the identification of 65 articles. Only studies reporting the level of ketosis, as a measurement of blood beta-hydroxybutyrate (BHB) level, were included. Finally, 90 articles were included in the review. The last search was performed on September 26th, 2023. The flow chart illustrating the article selection process is shown in Figure 1.

Figure 1. Flow chart showing the procedure of studies selection included in review.
3 Behavioral effects of KD treatment
3.1 Cognition
Nutritional ketosis reduces neuroinflammation, and oxidative stress, and improves mitochondrial function (28, 29). All these processes have a profound impact on brain health and in turn on neurocognitive functions. The neuroprotective properties of KDs were demonstrated in animal models of epilepsy, aging, dementia, and neurodegenerative diseases (30–32).
Cognition is a very complex construct that encompasses several aspects of intellectual functioning. Therefore, results arising from animal studies can only capture some of the meanings of this term.
Despite the availability of a variety of tests that measure slightly different, but also overlapping and interacting aspects of cognition, obtaining reliable and disease-relevant results requires careful choice of methods and usually using a combination of different paradigms.
The most commonly used behavioral test to measure impairment of cognitive functions is the Morris water maze (MWM). The MWM paradigm is designed to assess spatial learning and long-term memory by observing and recording escape latency, thigmotaxis duration, distance moved, and velocity during the time spent in the circular water tank with a hidden platform, where rodents are required to find the escape route to the platform by remembering visual cues (33). Another common test to evaluate learning and memory, particularly recognition memory, is the novel object recognition test (NOR). NOR test uses innate preference to interact with unknown objects in relation to known objects (e.g., blocks, balls) (34). Other behavioral tests used in animal studies involve modifications of maze tasks, for example, Y-maze, T-maze, V-maze, Hebb Williams Maze, or Barnes Maze. Above-described tests can be used to assess the rodents’ short- and long-term memory, learning, and spatial learning (35). It is important to note that the tests evaluating cognitive functions depend on the exploratory behavior of animals and their interaction with the environment. Other parameters like locomotor activity or levels of anxiety can significantly impact the outcomes of cognitive performance during tests, in addition to cognitive abilities themselves. In some paradigms, palatable food is used as a reward for conditional learning. Using these tests when comparing animals fed with KD or standard diet requires careful consideration since the animals fed with different diets may have various levels of motivation towards obtaining food. Moreover, an inappropriately selected snack may influence the level of ketosis.
The impact of KD on cognitive functions is most commonly described in epilepsy (37–44), traumatic brain injury (TBI) (45–49), Alzheimer’s disease (AD) (50–54), and also in healthy animals (55–62) especially in the context of aging (63–72).
In the animal models of epilepsy, it was reported in nine research articles (37–44). Despite the indisputable anti-seizure effects of KD, these studies do not bring unequivocal results regarding cognitive function. In different animal models of epilepsy: electrically elicited (kindled) seizures (36), spontaneously epileptic Kcna1-null mice (37), the pilocarpine-induced status epilepticus (38), and the pentylenetetrazol (PTZ)-kindled model (39) KD treatment led to improved memory functions. In the PTZ-kindled model of epilepsy KD improved spatial memory in the novel placement recognition test in rats, without changes in memory acquisition based on the MWM test results (39). Su et al. (40) drew attention to the importance of the timing of KD initiation in the pharmacological model of epilepsy, showing that early KD initiation (2 days after status epilepticus) resulted in weaker spatial learning in the MWM than observed in rats on a control diet or rats that started the KD 2 weeks after status epilepticus. In the context of epilepsy seizure activity is a primary cause of cognitive deficits (73, 74). Therefore, it is possible that the cognition-enhancing effect of the KD observed in the models of epilepsy is an indirect result of seizure mitigation. However, despite effective seizure mitigation, no improvements in cognitive performance after KD treatment were observed in other studies in a genetic model for idiopathic epilepsy (42), kindling model of epilepsy (43), and Dravet syndrome model (44). One study conducted in the lithium pilocarpine model of epilepsy showed substantially worsened performances in the MWM of young rats treated with KD. The severe impairment in visual–spatial memory was accompanied by decreased brain growth (41). Interestingly, when applying the ketogenic chow of the same composition to rats just after weaning, we observed adverse effects such as weakness, growth inhibition, and brain undergrowth. These adverse effects were mitigated when we supplemented the diet with wheat bran (27). Similarly, other authors achieved a significant reduction in adverse effects of the same ketogenic chow through supplementation with choline or methionine (26). It is important to mention that the same ketogenic chow (Bio-Serv F3666) used in mice, for example, in spontaneously epileptic Kcna1-null mice (37), in a TBI model (47) or in naive mice, led to improved cognitive measures. These studies are discussed in the further part of this section.
In the context of aging, healthspan, and lifespan numerous studies demonstrated improved cognitive function in KD-fed animals (63–72). KD treatment introduced at both old (20 months) and young (4 months) ages, enhanced cognition across the lifespan, regardless of sex. It resulted in enhanced performance on both the elevated figure-8 maze alternation task and a cognitive dual task that involved working memory. Also, the authors noted that the observed differences in protein expression related to metabolism and vesicular transport in the prefrontal cortex and hippocampus could contribute to the development of further therapies for age-related cognitive decline (68). Zhou et al. (63) showed that KD introduced at 18 months of age improved spatial learning and memory at 26 months of age. Roberts et al. (65) showed that aged male mice fed with KD show memory improvement in the NOR test as compared to the control but also to low-carbohydrate diet-fed animals. Authors suggested that nutritional ketosis, rather than the low glycemic index, affects index lifespan and slows down age-related cognitive impairment in old mice. In aged female mice after 2 months of KD, an improvement in spatial learning but no recognition memory or short-term working memory was observed (69). Newman et al. (64) reported that long-term exposure to the cyclic KD (given every other week) reduces mid-life mortality and preserves memory in aging males. Similarly, Hernandez et al. (67) found that time-restricted access to the KD in middle-aged mice positively affected cognitive functions compared to animals fed ad libitum with standard chow. Additionally, there were significant differences in gut microbiome diversity and composition in both diets. Authors suggested that improved cognition was associated with an altered gut microbiome, especially lowered Allobaculum abundance. Observed improvement may result from time-restricted feeding paradigm rather than macronutrient composition of KD and standard chow (67). These results provide evidence that the KD may beneficially affect cognitive function in female and male rodents, especially in middle-aged and old animals, not only after long-term exposure to diet (64, 65, 67–69), but also after a few weeks of treatment (66). On the other hand, exposure to KD from postnatal day (P) 20 - P32 for at least 5 weeks may adversely impact cognition later in life in laboratory rodents (40, 41, 71). Recently, Miles and Skelton (72) reported that early-life (from P21 through young adulthood ~P90) exposure to nutritional ketosis could impair learning and memory abilities. These data may suggest that the nutritional content of a KD is not sufficient to ensure proper neurodevelopment in young animals, while other studies argue that adequate composition of the diet (in terms of micronutrients and vitamins) is crucial for the health and development of young rodents fed with KD (26, 27, 75). We have previously demonstrated that modification of commonly used ketogenic chow allows for its application in developing rats, without causing detrimental side effects (27). Importantly, no adverse effects on neurodevelopment were observed in children using a KD to treat epilepsy (41). Hence, the observed underdevelopment and reduced cognitive abilities of young rodents fed with KD seem to be associated with inadequate composition of the diet or to be species-specific.
In animal models of TBI, the beneficial effect of the KD on cognition was demonstrated in four studies (45–48) while one study reported no effect (49). The KD-fed male adolescent (45–47) or young-adult rodents (47, 48) showed significantly improved recovery after injury and spatial memory in a variety of behavioral tests (MWM, NOR, or Y-maze) compared to the injured animals fed the control diet. Besides cognitive improvement, post-TBI KD administration resulted in better neurological outcomes including decreased degeneration of neurons in the dentate gyrus (45), attenuated neuroinflammation (46, 48), white matter damage, microgliosis (46), astrogliosis (45, 46), and oligodendrocyte loss (48), and improved sensorimotor functions (46, 48). Interestingly, Appelberg et al. (47) showed that KD introduced for 1 week immediately after TBI significantly improves cognitive recovery in adolescent rats but not in adult rats, suggesting that the effectiveness of ketones as an alternative fuel after TBI may be age-dependent. However, other authors demonstrate that applying an alternative KD formulation, with a fat-to-carbohydrate plus protein ratio of 2:1, containing MCT, docosahexaenoic acid, low glycemic index carbohydrates, fibers, and leucine, extends its neuroprotective potential in TBI to adult mice (48), again pointing out the importance of diet composition. One study reported that both pre-mild TBI and post-mild TBI exposure to the KD did not affect performance in novel context mismatch test in adolescent rats of both sexes. However, other parameters like balance and motor impairments, exploratory behavior, and telomere length were improved (49).
The observation that defective insulin signaling leading to decreased glucose metabolism may contribute to the progression of AD. It may be speculated that dietary interventions improving glucose and insulin metabolism might serve as a novel therapeutic approach to AD (76). Pre-clinical studies showed that the KD can mitigate some of the molecular and cellular changes associated with AD pathophysiology, resulting from enhancement in mitochondrial function, neuroprotection, reduction in neuroinflammatory response, and the expression of apoptotic mediators (21, 31). Moreover, the KD can help to eliminate brain amyloid-beta (Aβ) plaques by increasing the concentration of low density lipoprotein receptor-related protein 1 (LRP1), glycoprotein P (P-gp), and phosphatidylinositol binding clathrin assembly protein (PICALM) (77, 78). For instance, feeding with a KD decreased astroglial response to Aβ-plaques and lowered expression of the proinflammatory cytokines in the model of familial AD (79).
KD improved cognitive deficiency in female mice in a model of sleep deprivation-induced AD and a study with APP/PS1 mice, where KD was administered for 1 or 12 months (50, 51). While in female mice carrying the “London” APP mutation (APP/V717I) feeding with KD was not able to improve cognitive measures although it reduced Aβ40 and 42 levels by 25% (52). In other studies using genetic (APP/PS1, Tg4510) or pharmacological AD models (Aβ infusions) nutritional ketosis did not rescue memory deficits in a variety of experimental paradigms, varying in the time of KD exposure or diet composition (53, 54).
In addition to the aforementioned studies, there is further evidence indicating that nutritional ketosis can affect cognitive functions in animal models of neural disorders, stress, and obesity. KD administration improved cognition accompanied by histone modification in the model of neural disorders resulting from hypoxia injury (80), and Kmt2d+/βGeo mice (model of Kabuki syndrome) (81). A similar, positive effect of KD on cognition, related to peripheral metabolism (82) and biochemical changes in the hippocampus was shown in the rat model of chronic variable stress (83). We have previously shown that obesity-induced impairment in cognitive performance was ameliorated after weight loss achieved by either calorie restriction or KD. However, rats fed with a calorie-restricted KD performed better in MWM than those fed with a calorie-restricted standard diet (84). Finally, Fukushima et al. (55) concluded that the improvement in Y-maze performance may result from KD-induced increased hippocampal expression of the AMPA receptor subunit, GluR1 of naive adult rats. Also, the administration of KD with a ketogenic ratio of 6.6:0 has been shown to improve the Y-maze performance of naive adult rats in comparison to those fed with a standard diet. While no changes were observed with a ketogenic ratio of 3.0:0 (56). No effects of KD on cognition were reported in a few studies regarding synaptic functions (57–59), social behavior (60), evaluation of hippocampal involvement in spatial-cognitive behavior (61), or behavioral profiling (62) in wild type mice and rats.
Many studies suggest that a KD has a positive impact on cognition, especially in animal models of epilepsy, TBI, and aging. These studies generally report improvements, although sometimes there is no noticeable effect. The mitigation of cognitive impairment often goes hand in hand and may be secondary to other improvements in neurological and health outcomes such as reduced seizures in epilepsy models or reduced midlife mortality and improved health in old mice (64). However, in models of AD, the effects on cognition are usually moderate or even absent, despite reductions in amyloid deposition and other processes contributing to disease progression. In naive adult rodents, six articles report no effect (57–62), while two note cognition enhancement (55, 56). Four publications show that the application of a KD directly after weaning results in cognitive impairments later in life (40, 41, 71, 72).
3.2 Depressive-like behavior
The potential use of a KD in the treatment of depression is explored in the literature. Mechanisms through which a KD may potentially positively influence depression symptoms involve the modulation of the glutamate-glutamine cycle, gamma-aminobutyric acid (GABA) neurotransmission and, monoamine levels (85). Additionally, the diet provides nutrients such as ω-3 fatty acids, which may contribute to improvements in depression, and it may also influence the composition of the gut microbiota (86). It was proposed that KD through modulation of gut bacteria and its metabolites improves gut dysbiosis, decreases cytokine production, and lowers overall inflammation observed in depression (87). Another important finding underlying the therapeutic potential of nutritional ketosis comes from metabolic and behavioral analysis of Dravet mice fed with KD (44). KD reduced preference for saccharin in the sucrose preference test (SPT) in wild-type and Dravet mice, however, hippocampal levels of glutamate precursor ⍺-ketoglutarate and ⍺-D-glucose-1-phosphate correlated positively with saccharin preference in Dravet but not in wild-type mice (44). The SPT bases on natural rodents’ preference to selectively drink sweet solution when given a two-bottle free-choice regimen with access to both sucrose solution and water. A reduction in the sucrose preference ratio is indicative of anhedonia used for detection of depressive-like behavior in rodents (88).
The influence of nutritional ketosis on depressive-like behaviors and stress response was investigated in a few experimental studies (49, 57, 71, 89–96). Four studies have reported that nutritional ketosis may positively impact depressive-like behaviors (89–91, 93), as measured by the forced swim test (FST) and tail suspension test (TST), two classical behavior paradigms designed to measure depression levels, including changes observed in response to acute stress (97). The prolonged immobility in TST and FST are used for estimating depression-related behavior. Both of these models work similarly in assessing behavioral symptoms of feeling despair but not anhedonia (98).
One of the studies revealed that KD-fed rats exhibited less immobility duration in the FST when compared to those fed the standard diet (89). Additionally, the other study found that young adult CD-1 mice-offspring of mothers fed with a KD during pregnancy, exhibited reduced susceptibility to anxiety and depression (93). These observations were confirmed by Arqoub et al. (90) who observed that gestational exposure to KD reduced the expression of depressive-like behaviors in the FST. The latest study by Guan et al. (91), revealed that KD treatment decreased immobility duration in the TST and FST, and increased sucrose preference in the anhedonia-based SPT in repeated social defeat stress (R-SDS) and lipopolysaccharide (LPS) depression models. In the most recent paper by Gumus et al. (94), a combination of regular voluntary exercise with a KD decreased depressive-like behaviors in adult male mice, which was correlated with a decline of insulin and glucose or low/high-density lipoprotein (LDL/HDL) ratio and an increase of BHB levels. To the best of our knowledge, no studies are reporting the worsening of depressive-like behaviors in nutritional ketosis. No influence of KD feeding on depressive-like behaviors was reported in naive animals (57, 95), and the genetic model of Fragile X Syndrome (96).
The results from experimental studies suggest that nutritional ketosis may exhibit a beneficial influence on depressive-like behaviors. Considering the applied experimental paradigms – application of diet before acute stress or in utero - the effect may have a preventive character. Moreover, the differences observed in the experimental designs (e.g., the type of diet used, and time of administration), further emphasize the pressing need to investigate the underlying biological mechanisms of the anti-depression effects of the KD. Considering the positive impact of KD on depressive-like behavior, it might be postulated that nutritional ketosis might be used in depression treatment.
3.3 Anxiety-like behavior
The rationale behind the potential use of KD in anxiety disorders comes from its ability to counteract pathological changes in neurotransmission that are strongly linked to anxiety. These include GABA deficiency (99, 100) and increased neuronal excitability (15). The usefulness of the KD in anxiety disorders may also arise from its impact on gut microbiota, improvement of intestinal barrier function (101), its anti-inflammatory effects (102), and reduced production of reactive oxygen species (ROS) (103). The mechanisms substantiating the potential applicability of KDs in anxiety disorders were comprehensively discussed by Zhu et al. (15) and Wlodarczyk et al. (104) while the available clinical evidence was recently systematically reviewed by Dietch et al. (105).
Numerous behavioral tests have been developed to measure anxiety in rodents (106–108). In tests like open field, dark/light compartment tests, or elevated plus maze (EPM) the assessment of anxiety relies on the fact that laboratory rodents prefer closed and dark over open and light spaces (109). Other popular type of tests, used mostly for anxiolytic screening, are “conflict” tests like the Geller-Seifter or Vogel test, in which a hungry or thirsty animal is given an option to obtain, respectively, food or water by pressing a lever that can also elicit electric shock (110, 111). Due to the nature of anxiety tests separating anxiety, exploratory, activity, and learning responses is often not possible. Therefore, for the interpretation of results and understanding of their translational potential, it is crucial to recognize that a multitude of factors influences animal behavior in those tests. The overview of most common animal tests of anxiety alongside the consideration of conceptual issues regarding methodological details, interpretation of results, and intraspecies translation is comprehensively discussed in excellent reviews that focus on anxiety evaluation in preclinical settings (106–108).
Twenty articles evaluating the influence of nutritional ketosis on anxiety-related behaviors met the eligibility criteria for this review. Most data indicate that the KD does not influence anxiety-related behaviors (39, 55, 57, 60, 64, 69, 92, 95, 96, 112–115).
Among studies performed on rats, two reported positive effects of the KD on anxiety. Both young and aged naive rats fed with a KD showed resilience against the anxiogenic open arm in the EPM test (68). KD treatment showed protective properties by reducing anxiety levels in a model of TBI. Rats exposed to the KD post-injury showed reduced anxiety- and depressive-like behaviors acutely post-TBI. While pre-injury exposure to the KD resulted in even more pronounced improvement of outcomes like reduced balance and motor impairments (49). Interestingly, one study reported increased anxiety levels and decreased locomotor activity on a KD that were reversed by environmental enrichment (71). Other studies performed on rats report no effect of a KD on anxiety-related behaviors (39, 60, 95, 112, 113).
In naive mice, one study reported that a combination of a KD and regular voluntary exercise ameliorated anxiety and depression-like behaviors (Balb/c mice) (94). However, other studies performed on naive mice did not show changes in anxiety-related behaviors (55), also in the context of aging, in male and female mice, despite improvements in other neurocognitive functions (54, 69). Gestational exposure to a KD resulted in reduced susceptibility to anxiety and depression in adulthood, alongside many neuro-anatomical differences (93). Reduced anxiety under nutritional ketosis was also reported in a study performed on a model of ASD in BTBR mice (116). Moreover, a general improvement of autism symptoms after KD treatment was observed in numerous preclinical models of ASD (10, 116–119). Other research showed that a KD supplemented with ketone monoester reduced handling-induced convulsions and anxiety-like behaviors in early alcohol withdrawal (120). The tendency toward lower anxiety-like behaviors was reported in other studies performed on mice including the model of MPC1 deficiency in adult glutamatergic neurons (114) or Fragile X Syndrome (96), but none of the studies reported anxiogenic effects of nutritional ketosis. Interestingly, a study evaluating the effects of chronic or subchronic (7 days) administration of exogenous ketones alongside a standard diet reported a reduction of anxiety assessed with EPM test in all treatment conditions (121). Summing up, studies evaluating anxiety-related behaviors in nutritional ketosis report either no effect or reduced anxiety in the majority. The latter seems to be an indirect effect originating from the mitigation of pathophysiological changes specific to the examined disease model.
3.4 Social behavior
Given the beneficial effect of nutritional ketosis on epilepsy, mitochondrial function, carbohydrate metabolism, and inflammation, it has been proposed that treatment with a KD has the potential to reduce some of the ASD-associated symptoms, including impaired social interactions (122). The social behavior of laboratory rodents is most commonly evaluated with a 3-chamber test which allows for assessing sociability (time spent in the chamber with mouse vs. chamber with object) and preference for social novelty (time spent with unknown vs. known mouse). Other commonly used tests include: social transmission of food preference or analysis of the social activity in a home cage where behaviors like sniffing and following are analyzed, or in the case of juvenile rodents, also play responses like evasion or rotation (123, 124). The influence of the KD on social behavior has been tested in nine experimental studies (10, 57, 60, 90, 115–119). Studies conducted in rodent models of ASD, i.e., the BTBR model (116), the prenatal valproic acid (VPA) model (117), Shank3+/ΔC mice (115), Engrailed 2 null mice (10, 118), and the maternal immune activation model of ASD (119) reported improvement of social deficits. An increase in social activity has also been reported in wild-type rats fed with a KD (60, 117) and in offsprings of dams fed a KD during gestation (90). Only one study reported that feeding with KD has not affected sociability in naive mice (57). Despite differences in the used models, age of the animals, time of the treatment, and employed behavioral tests, most of all these studies coherently show the increased social activity of rodents fed with the KD. This suggests that the mechanism by which the KD increases social activity is independent of the alterations underlying social impairment. Another line of evidence supporting this conclusion will be the observation that KD-induced reduction in social impairment in BTBR mice is not secondary to the well-known antiepileptic properties of this diet (116). It can be hypothesized that increased social activity results from other behavioral changes like increased arousal which translates to greater locomotor activity or reduced anxiety which leads to enhanced interest in the environment in general. However, neither reduced anxiety, increased locomotor activity nor changes in memory were reported in the studies mentioned above (10, 60). The effect is not persistent since the level of social activity is restored to control levels after the cessation of the diet (10, 60). In contrast, KD ameliorated autism-like social deficits observed in Shank3+/ΔC mice, and this positive impact endured for up to 6 weeks after discontinuation of the diet (115). Most of the above-mentioned studies were conducted on male rodents, however, it has been also demonstrated that in Engrailed 2 null mice KD improved multiple measures of sociability in females, with limited effects in males (118). Although mechanisms underlying changes in social behavior in nutritional ketosis have not been deeply investigated, some insight was provided by Verpeut et al. (10) in the study performed on Engrailed 2 null mice, where immunohistochemical analysis demonstrated that groups exposed to the KD, regardless of genotype, showed increased neuronal activation in response to novel animal exposure. The KD-fed animals had more c-Fos positive cells in brain regions associated with social behaviors including the cingulate cortex, lateral septal nuclei, and anterior bed nucleus of the stria terminalis (10). This supports the idea that an increase in various aspects of sociability observed in the abovementioned studies arises from the impact of the KD on neuronal circuits controlling social behavior, independently of particular pathology underlying social impairment in used disease models.
3.5 Nutritional behavior
The KD is increasingly used for the treatment of obesity and as an add-on therapy in the management of type 2 diabetes (T2DM) (125). Meta-analyses comparing the effectivity of KDs to low-fat diets consistently show slightly greater weight loss, improved HDL-cholesterol, triacylglycerol (TAG), and other cardiometabolic markers but increased LDL-cholesterol (126–128). Although there is no consensus on the precise mechanism that determines the efficiency of weight loss under nutritional ketosis suppression of appetite is considered the play a leading role (17, 129, 130). Increased feeling of hunger is a common side effect of diet-induced weight loss that in the long term leads to reduced patient adherence compromising the results of the therapy and finally promoting weight regain (131, 132). Therefore understanding the mechanism of appetite suppression on a KD can significantly contribute to the improvement of weight loss therapies both in terms of lifestyle intervention as well as the development of new drugs. Studying the appetite-controlling neuroendocrine network on molecular and cellular levels is almost exclusively possible with the use of animal models. However, a question arises if animal models are suitable for studying appetite regulation under nutritional ketosis, i.e., if reduction of appetite occurs in laboratory rodents fed with the KD and how it can be measured, and finally what are the best experimental conditions to reflect the human situation. In the field of nutrition obesity on the behavior of rodents is most commonly evaluated by measuring food intake expressed in grams or in calories if different diets are compared. The most common methods for quantifying food consumption are manual weighing of the chow, but also automated chow counters, pellet dispensers, or video monitoring (133). The important factor compromising the accuracy of these methods is the fact that not the whole amount of chow leaving the food containers is consumed by animals. Especially in the case of high-fat diets (HFD) due to their consistency the chow crumbles (or is shredded by the animals) and falls into the bedding. This impacts the reliability of comparison between chows having various consistencies like standard chow and ketogenic chow. Another aspect important for methodological considerations is performing food deprivation studies or using food as a reward or reinforcement. Inappropriately chosen snack may lower the level of nutritional ketosis. Moreover, animals fed with different diets (standard and KD) may vary in their interaction with food, e.g., the motivation to obtain food, or may have different fasting tolerance. In the first couple of days after switching to a KD rodents usually reduce food/calorie intake (134–136). This reflects the need to acclimate to the type and consistency of new chow rather than a reduction of appetite since the calorie intake quickly goes back to baseline levels (134–136). After the habituation period, rodents fed with a KD usually consume less food expressed in grams but equal calories as the animals fed with standard chow. This trend was observed in wild-type animals fed with KD for up to 2 months (26, 55, 57, 83, 137–141) or longer (58, 142–145) as well as in different disease models like T2DM model (146), AD model (54), in the stress model (92), glaucoma model in both females and males (147), models of hepatic enzyme disturbances (148, 149), and in acute alcohol withdrawal symptoms (120). However, despite the equivalent caloric intake, feeding with KD often results in improved body mass (26, 83, 138, 140, 144) and metabolic health in long-term treatment (65). In some studies, rodents fed with KD consumed more calories than chow-fed controls, which was probably associated with lower energy assimilation because weight gain was not increased (44, 60, 95, 135, 150–158). One exception is a study in the model of Dravet syndrome where increased calorie intake was accompanied by increased body weight probably resulting from improvement of other disease symptoms (44). Decreased calorie intake, accompanied by decreased body weight during nutritional ketosis was reported in a couple of studies (53, 159–163). Taken together the majority of studies show that a KD, in comparison to standard chow, does not induce a spontaneous reduction in food intake. This suggests that the reduction in appetite observed in humans following a KD either does not occur in laboratory rodents, at least when compared to a standard chow, or is not detected through routine monitoring of food intake.
To answer the question of whether rodent models can be used to study mechanisms of appetite regulation under nutritional ketosis assessment of appetite could be performed in conditions where animals significantly overeat. This can be achieved by testing the changes in the appetite of diet induced obesity (DIO) animals after switching them to a standard or KD or by offering palatable snacks to animals fed with either KD or standard diet. It is a matter of ongoing discussion whether the disruption of homeostatic hunger regulation or the hedonic reward system governs overeating in rodents exposed to HFDs (164).
Therefore, experimental conditions need to be planned carefully with consideration of the underlying neuroendocrine status, choice of food/snacks, and test methods. Not only monitoring food intake but also applying behavioral tests like the food/risk competition test or food preference test would be informative (165). According to our best knowledge, such studies have not been performed so far. However, one study showed that animals previously fed with KD or HFD preferred to obtain morecalories from HFD, than animals fed with standard chow (55).
Laboratory rodents are widely used in research exploring mechanisms of KD action. However, the aspect of appetite regulation was not addressed in those studies so far and it remains an open question whether animal models are adequate research tools in this regard. Multiple factors including the differences in feeding patterns of humans and laboratory rodents or mechanisms that govern overeating need to be taken into account. Although study design seems to be challenging, the potential benefits of understanding KD-induced changes in the neuroendocrine network controlling appetite may have implications for the treatment of obesity that go beyond the use of the KD itself.
4 Summary
The majority of research performed in various disease models shows that the KD positively impacts cognition, especially in the models of epilepsy, TBI, and aging but also in naive animals (Supplementary Table S1). Almost an equal number of studies reports a reduction or no effect of the KD on depressive-related behaviors.
For anxiety-related behaviors, the majority of studies show no effect of the KD treatment. The adverse influence of the KD on cognition, anxiety-, and depressive-related behaviors was rarely reported (Table 1). Beneficial effects observed in behavioral measures seem to arise from the neuroprotective properties of the KD. Since the behavioral changes are accompanied by improvements in other outcomes specific to a disease model. In contrast, the increase in social activity seems to be unspecific and independent of the underlying cause of social impairment.
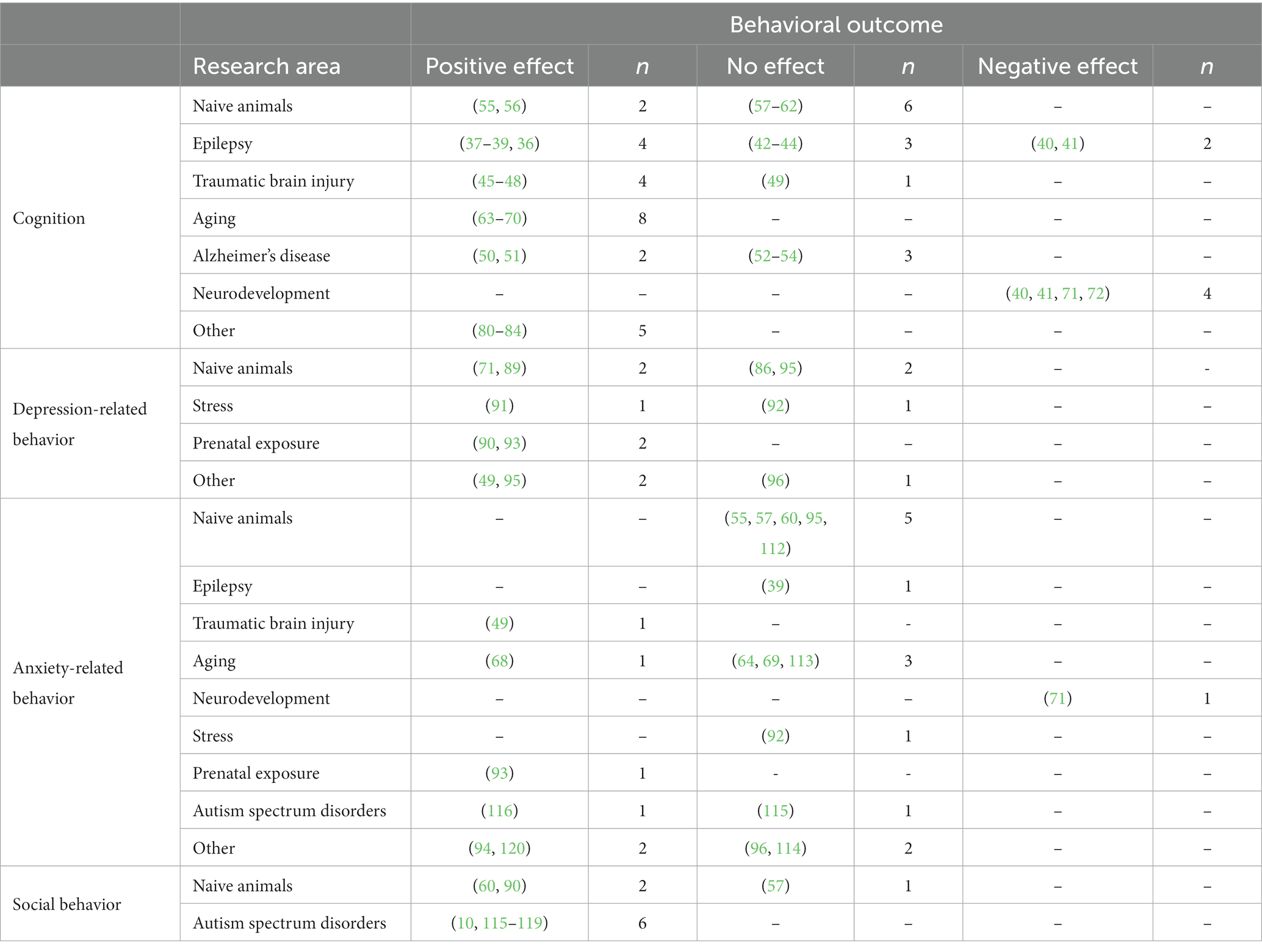
Table 1. The behavioral effects of ketogenic diet treatment.
Despite the growing use of the KD in the treatment of obesity, accompanied by scientifically proven efficiency and appetite-reducing properties of the KD, the aspect of nutritional behavior of KD-fed animals has not been addressed so far. It remains an open question whether animal models are adequate research tools to study appetite regulation under nutritional ketosis.
Using animal models to evaluate the influence of the KD on behavioral measures requires paying special attention to the composition of the diet. While the proper macronutrient ratio is reflected by elevated blood BHB levels, an adequate micronutrient supply must be provided to avoid malnutrition that will affect behavioral measures (26, 27, 75). A couple of studies point out that additional supplementation of the KD with, e.g., MCT, ketogenic amino acids, or exogenous ketones may impact not only physiological but also behavioral effects of the diet in laboratory animals (48, 166) but also in humans (167). Although currently, many scientists acknowledge the significance of the KD’s composition in determining its properties, most studies primarily investigate its effects in comparison to standard rodent chow. The differences between KDs of various compositions are rarely tested. Therefore, drawing direct conclusions about the role of diet composition or the presence of a particular ingredient that would be backed by strong scientific evidence is usually not possible.
Author contributions
KG: Investigation, Visualization, Writing – original draft. MG: Investigation, Writing – original draft. MP: Investigation, Writing – original draft. NP: Investigation, Writing – original draft. JJB: Conceptualization, Supervision, Writing – review & editing. MN-Ch: Investigation, Methodology, Project administration, Resources, Supervision, Writing – original draft, Writing – review & editing. DL: Conceptualization, Methodology, Project administration, Supervision, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by the statutory grants: AWF/NF/ZB1/2023/1/3 from the Academy of Physical Education, Katowice, Poland and PCN-2-057/K/2/I, PCN-2-023/N/1/O from the Medical University of Silesia, Katowice, Poland.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnut.2024.1322509/full#supplementary-material
References
1. Masood, W, Annamaraju, P, Khan Suheb, MZ, and Uppaluri, KR. Ketogenic Diet. Treasure Island: StatPearls Publishing (2022).
2. Barzegar, M, Afghan, M, Tarmahi, V, Behtari, M, Rahimi Khamaneh, S, and Raeisi, S. Ketogenic diet: overview, types, and possible anti-seizure mechanisms. Nutr Neurosci. (2021) 24:307–16. doi: 10.1080/1028415X.2019.1627769
3. Wheless, JW. History of the ketogenic diet. Epilepsia. (2008) 49:3–5. doi: 10.1111/j.1528-1167.2008.01821.x
4. Greenhill, C. Ketogenic diet affects immune cells in mice. Nat Rev Endocrinol. (2020) 16:196–7. doi: 10.1038/s41574-020-0328-x
5. Rojas-Morales, P, León-Contreras, JC, Sánchez-Tapia, M, Silva-Palacios, A, Cano-Martínez, A, González-Reyes, S, et al. A ketogenic diet attenuates acute and chronic ischemic kidney injury and reduces markers of oxidative stress and inflammation. Life Sci. (2022) 289:120227. doi: 10.1016/j.lfs.2021.120227
6. Gentile, CL, and Weir, TL. The gut microbiota at the intersection of diet and human health. Science. (2018) 362:776–80. doi: 10.1126/science.aau5812
7. Cheng, CM, Kelley, B, Wang, J, Strauss, D, Eagles, DA, and Bondy, CA. A ketogenic diet increases brain insulin-like growth factor receptor and glucose transporter gene expression. Endocrinology. (2003) 144:2676–82. doi: 10.1210/en.2002-0057
8. Noh, HS, Kim, DW, Kang, SS, Kim, YH, Cho, GJ, and Choi, WS. Ketogenic diet decreases the level of proenkephalin mRNA induced by kainic acid in the mouse hippocampus. Neurosci Lett. (2006) 395:87–92. doi: 10.1016/j.neulet.2005.10.073
9. Stafford, P, Abdelwahab, MG, Kim, DY, Preul, MC, Rho, JM, and Scheck, AC. The ketogenic diet reverses gene expression patterns and reduces reactive oxygen species levels when used as an adjuvant therapy for glioma. Nutr Metab (Lond). (2010) 7:74. doi: 10.1186/1743-7075-7-74
10. Verpeut, JL, DiCicco-Bloom, E, and Bello, NT. Ketogenic diet exposure during the juvenile period increases social behaviors and forebrain neural activation in adult engrailed 2 null mice. Physiol Behav. (2016) 161:90–8. doi: 10.1016/j.physbeh.2016.04.001
11. Vizuete, AF, de Souza, DF, Guerra, MC, Batassini, C, Dutra, MF, Bernardi, C, et al. Brain changes in BDNF and S100B induced by ketogenic diets in Wistar rats. Life Sci. (2013) 92:923–8. doi: 10.1016/j.lfs.2013.03.004
12. Mangiarotti Marchi, MA, Marchi, AG, Fasola, V, and Gregoretti, S. Studio di attività enzimatiche in linfociti fito-stimolati nelle malattie congenite del metabolismo [Enzymatic lysosomal activity in phyto-stimulated lymphocytes in congenital metabolic diseases]. Minerva Pediatr. (1971) 23:1847–50.
13. Ziegler, DR, Araújo, E, Rotta, LN, Perry, ML, and Gonçalves, CA. A ketogenic diet increases protein phosphorylation in brain slices of rats. J Nutr. (2002) 132:483–7. doi: 10.1093/jn/132.3.483
14. Yudkoff, M, Daikhin, Y, Melø, TM, Nissim, I, Sonnewald, U, and Nissim, I. The ketogenic diet and brain metabolism of amino acids: relationship to the anticonvulsant effect. Annu Rev Nutr. (2007) 27:415–30. doi: 10.1146/annurev.nutr.27.061406.093722
15. Zhu, H, Bi, D, Zhang, Y, Kong, C, Du, J, Wu, X, et al. Ketogenic diet for human diseases: the underlying mechanisms and potential for clinical implementations. Signal Transduct Target Ther. (2022) 7:11. doi: 10.1038/s41392-021-00831-w
16. Batch, JT, Lamsal, SP, Adkins, M, Sultan, S, and Ramirez, MN. Advantages and disadvantages of the ketogenic diet: a review article. Cureus. (2020) 12:e9639. doi: 10.7759/cureus.9639
17. Gibson, AA, Seimon, RV, Lee, CM, Ayre, J, Franklin, J, Markovic, TP, et al. Do ketogenic diets really suppress appetite? A systematic review and meta-analysis. Obes Rev. (2015) 16:64–76. doi: 10.1111/obr.12230
18. Nymo, S, Coutinho, SR, Jørgensen, J, Rehfeld, JF, Truby, H, Kulseng, B, et al. Timeline of changes in appetite during weight loss with a ketogenic diet. Int J Obes. (2017) 41:1224–31. doi: 10.1038/ijo.2017.96
19. Titcomb, TJ, Liu, B, Wahls, TL, Snetselaar, LG, Shadyab, AH, Tabung, FK, et al. Comparison of the ketogenic ratio of macronutrients with the low-carbohydrate diet score and their association with risk of type 2 diabetes in postmenopausal women: a secondary analysis of the Women’s Health Initiative. J Acad Nutr Diet. (2023) 123:1152–1161.e4. doi: 10.1016/j.jand.2022.12.004
20. Dunn, E, Zhang, B, Sahota, VK, and Augustin, H. Potential benefits of medium chain fatty acids in aging and neurodegenerative disease. Front Aging Neurosci. (2023) 15:1230467. doi: 10.3389/fnagi.2023.1230467
21. Xu, Y, Jiang, C, Wu, J, Liu, P, Deng, X, Zhang, Y, et al. Ketogenic diet ameliorates cognitive impairment and neuroinflammation in a mouse model of Alzheimer’s disease. CNS Neurosci Ther. (2022) 28:580–92. doi: 10.1111/cns.13779
22. Takeishi, J, Tatewaki, Y, Nakase, T, Takano, Y, Tomita, N, Yamamoto, S, et al. Alzheimer’s disease and type 2 diabetes mellitus: the use of MCT oil and a ketogenic diet. Int J Mol Sci. (2021) 22:12310. doi: 10.3390/ijms222212310
23. Khodabakhshi, A, Akbari, ME, Mirzaei, HR, Mehrad-Majd, H, Kalamian, M, and Davoodi, SH. Feasibility, safety, and beneficial effects of MCT-based ketogenic diet for breast cancer treatment: a randomized controlled trial study. Nutr Cancer. (2020) 72:627–34. doi: 10.1080/01635581.2019.1650942
24. Lee, RWY, Corley, MJ, Pang, A, Arakaki, G, Abbott, L, Nishimoto, M, et al. A modified ketogenic gluten-free diet with MCT improves behavior in children with autism spectrum disorder. Physiol Behav. (2018) 188:205–11. doi: 10.1016/j.physbeh.2018.02.006
25. O’Neill, B, and Raggi, P. The ketogenic diet: pros and cons. Atherosclerosis. (2020) 292:119–26. doi: 10.1016/j.atherosclerosis.2019.11.021
26. Pissios, P, Hong, S, Kennedy, AR, Prasad, D, Liu, FF, and Maratos-Flier, E. Methionine and choline regulate the metabolic phenotype of a ketogenic diet. Mol Metab. (2013) 2:306–13. doi: 10.1016/j.molmet.2013.07.003
27. Liśkiewicz, AD, Kasprowska-Liśkiewicz, D, Sługocka, A, Nowacka-Chmielewska, MM, Wiaderkiewicz, J, Jędrzejowska-Szypułka, H, et al. The modification of the ketogenic diet mitigates its stunting effects in rodents. Appl Physiol Nutr Metab. (2018) 43:203–10. doi: 10.1139/apnm-2017-0374
28. Chinna-Meyyappan, A, Gomes, FA, Koning, E, Fabe, J, Breda, V, and Brietzke, E. Effects of the ketogenic diet on cognition: a systematic review. Nutr Neurosci. (2022) 26:1258–78. doi: 10.1080/1028415X.2022.2143609
29. Hallböök, T, Ji, S, Maudsley, S, and Martin, B. The effects of the ketogenic diet on behavior and cognition. Epilepsy Res. (2012) 100:304–9. doi: 10.1016/j.eplepsyres.2011.04.017
30. Davis, JJ, Fournakis, N, and Ellison, J. Ketogenic diet for the treatment and prevention of dementia: a review. J Geriatr Psychiatry Neurol. (2021) 34:3–10. doi: 10.1177/0891988720901785
31. Rusek, M, Pluta, R, Ułamek-Kozioł, M, and Czuczwar, SJ. Ketogenic diet in Alzheimer’s disease. Int J Mol Sci. (2019) 20:3892. doi: 10.3390/ijms20163892
32. Hersant, H, and Grossberg, G. The ketogenic diet and Alzheimer’s disease. J Nutr Health Aging. (2022) 26:606–14. doi: 10.1007/s12603-022-1807-7
33. Tian, H, Ding, N, Guo, M, Wang, S, Wang, Z, Liu, H, et al. Analysis of learning and memory ability in an Alzheimer’s disease mouse model using the Morris water maze. J Vis Exp. (2019):152. doi: 10.3791/60055
34. Lueptow, LM. Novel object recognition test for the investigation of learning and memory in mice. J Vis Exp. (2017) 126:55718. doi: 10.3791/55718
35. Hölter, SM, Garrett, L, Einicke, J, Sperling, B, Dirscherl, P, Zimprich, A, et al. Assessing cognition in mice. Curr Protoc Mouse Biol. (2015) 5:331–58. doi: 10.1002/9780470942390.mo150068
36. Gom, RC, Bhatt, D, Villa, BR, George, AG, Lohman, AW, Mychasiuk, R, et al. The ketogenic diet raises brain oxygen levels, attenuates postictal hypoxia, and protects against learning impairments. Neurobiol Dis. (2021) 154:105335. doi: 10.1016/j.nbd.2021.105335
37. Kim, DY, Simeone, KA, Simeone, TA, Pandya, JD, Wilke, JC, Ahn, Y, et al. Ketone bodies mediate antiseizure effects through mitochondrial permeability transition. Ann Neurol. (2015) 78:77–87. doi: 10.1002/ana.24424
38. Qiao, Q, Qu, Z, Tian, S, Cao, H, Zhang, Y, Sun, C, et al. Ketogenic diet alleviates hippocampal neurodegeneration possibly via ASIC1a and the mitochondria-mediated apoptotic pathway in a rat model of temporal lobe epilepsy. Neuropsychiatr Dis Treat. (2022) 18:2181–98. doi: 10.2147/NDT.S376979
39. Jiang, Y, Lu, Y, Jia, M, Wang, X, Zhang, Z, Hou, Q, et al. Ketogenic diet attenuates spatial and item memory impairment in pentylenetetrazol-kindled rats. Brain Res. (2016) 1646:451–8. doi: 10.1016/j.brainres.2016.06.029
40. Su, SW, Cilio, MR, Sogawa, Y, Silveira, DC, Holmes, GL, and Stafstrom, CE. Timing of ketogenic diet initiation in an experimental epilepsy model. Brain Res Dev Brain Res. (2000) 125:131–8. doi: 10.1016/s0165-3806(00)00130-9
41. Zhao, Q, Stafstrom, CE, Fu, DD, Hu, Y, and Holmes, GL. Detrimental effects of the ketogenic diet on cognitive function in rats. Pediatr Res. (2004) 55:498–506. doi: 10.1203/01.PDR.0000112032.47575.D1
42. Todorova, MT, Tandon, P, Madore, RA, Stafstrom, CE, and Seyfried, TN. The ketogenic diet inhibits epileptogenesis in EL mice: a genetic model for idiopathic epilepsy. Epilepsia. (2000) 41:933–40. doi: 10.1111/j.1528-1157.2000.tb00275.x
43. Hori, A, Tandon, P, Holmes, GL, and Stafstrom, CE. Ketogenic diet: effects on expression of kindled seizures and behavior in adult rats. Epilepsia. (1997) 38:750–8. doi: 10.1111/j.1528-1157.1997.tb01461.x
44. Miljanovic, N, van Dijk, RM, Buchecker, V, and Potschka, H. Metabolomic signature of the Dravet syndrome: a genetic mouse model study. Epilepsia. (2021) 62:2000–14. doi: 10.1111/epi.16976
45. Har-Even, M, Rubovitch, V, Ratliff, WA, Richmond-Hacham, B, Citron, BA, and Pick, CG. Ketogenic diet as a potential treatment for traumatic brain injury in mice. Sci Rep. (2021) 11:23559. doi: 10.1038/s41598-021-02849-0
46. Dilimulati, D, Zhang, F, Shao, S, Lv, T, Lu, Q, Cao, M, et al. Ketogenic diet modulates Neuroinflammation via metabolites from Lactobacillus reuteri after repetitive mild traumatic brain injury in adolescent mice. Cell Mol Neurobiol. (2023) 43:907–23. doi: 10.1007/s10571-022-01226-3
47. Appelberg, KS, Hovda, DA, and Prins, ML. The effects of a ketogenic diet on behavioral outcome after controlled cortical impact injury in the juvenile and adult rat. J Neurotrauma. (2009) 26:497–506. doi: 10.1089/neu.2008.0664
48. Thau-Zuchman, O, Svendsen, L, Dyall, SC, Paredes-Esquivel, U, Rhodes, M, Priestley, JV, et al. A new ketogenic formulation improves functional outcome and reduces tissue loss following traumatic brain injury in adult mice. Theranostics. (2021) 11:346–60. doi: 10.7150/thno.48995
49. Salberg, S, Weerwardhena, H, Collins, R, Reimer, RA, and Mychasiuk, R. The behavioural and pathophysiological effects of the ketogenic diet on mild traumatic brain injury in adolescent rats. Behav Brain Res. (2019) 376:112225. doi: 10.1016/j.bbr.2019.112225
50. Yang, Y, Wang, X, Xiao, A, Han, J, Wang, Z, and Wen, M. Ketogenic diet prevents chronic sleep deprivation-induced Alzheimer’s disease by inhibiting iron dyshomeostasis and promoting repair via Sirt1/Nrf2 pathway. Front Aging Neurosci. (2022) 14:998292. doi: 10.3389/fnagi.2022.998292
51. Qin, Y, Bai, D, Tang, M, Zhang, M, Zhao, L, Li, J, et al. Ketogenic diet alleviates brain iron deposition and cognitive dysfunction via Nrf2-mediated ferroptosis pathway in APP/PS1 mouse. Brain Res. (2023) 1812:148404. doi: 10.1016/j.brainres.2023.148404
52. Van der Auwera, I, Wera, S, Van Leuven, F, and Henderson, ST. A ketogenic diet reduces amyloid beta 40 and 42 in a mouse model of Alzheimer’s disease. Nutr Metab (Lond). (2005) 2:28. doi: 10.1186/1743-7075-2-28
53. Park, S, Zhang, T, Wu, X, and Yi, QJ. Ketone production by ketogenic diet and by intermittent fasting has different effects on the gut microbiota and disease progression in an Alzheimer’s disease rat model. J Clin Biochem Nutr. (2020) 67:188–98. doi: 10.3164/jcbn.19-87
54. Brownlow, ML, Benner, L, D’Agostino, D, Gordon, MN, and Morgan, D. Ketogenic diet improves motor performance but not cognition in two mouse models of Alzheimer’s pathology. PLoS One. (2013) 8:75713. doi: 10.1371/journal.pone.0075713
55. Fukushima, A, Ogura, Y, Furuta, M, Kakehashi, C, Funabashi, T, and Akema, T. Ketogenic diet does not impair spatial ability controlled by the hippocampus in male rats. Brain Res. (2015) 1622:36–42. doi: 10.1016/j.brainres.2015.06.016
56. Ruskin, DN, Suter, TA, Ross, JL, and Masino, SA. Ketogenic diets and thermal pain: dissociation of hypoalgesia, elevated ketones, and lowered glucose in rats. J Pain. (2013) 14:467–74. doi: 10.1016/j.jpain.2012.12.015
57. Huang, J, Li, YQ, Wu, CH, Zhang, YL, Zhao, ST, Chen, YJ, et al. The effect of ketogenic diet on behaviors and synaptic functions of naive mice. Brain Behav. (2019) 9:e01246. doi: 10.1002/brb3.1246
58. Hargrave, SL, Davidson, TL, Lee, TJ, and Kinzig, KP. Brain and behavioral perturbations in rats following Western diet access. Appetite. (2015) 93:35–43. doi: 10.1016/j.appet.2015.03.037
59. Thio, LL, Rensing, N, Maloney, S, Wozniak, DF, Xiong, C, and Yamada, KA. A ketogenic diet does not impair rat behavior or long-term potentiation. Epilepsia. (2010) 51:1619–23. doi: 10.1111/j.1528-1167.2009.02515.x
60. Kasprowska-Liśkiewicz, D, Liśkiewicz, AD, Nowacka-Chmielewska, MM, Nowicka, J, Małecki, A, and Barski, JJ. The ketogenic diet affects the social behavior of young male rats. Physiol Behav. (2017) 179:168–77. doi: 10.1016/j.physbeh.2017.06.007
61. Silva, MC, Rocha, J, Pires, CS, Ribeiro, LC, Brolese, G, Leite, MC, et al. Transitory gliosis in the CA3 hippocampal region in rats fed on a ketogenic diet. Nutr Neurosci. (2005) 8:259–64. doi: 10.1080/10284150500475032
62. Ródenas-González, F, Blanco-Gandía, MC, Miñarro, J, and Rodríguez-Arias, M. Cognitive profile of male mice exposed to a ketogenic diet. Physiol Behav. (2022) 254:113883. doi: 10.1016/j.physbeh.2022.113883
63. Zhou, Z, Kim, K, Ramsey, JJ, and Rutkowsky, JM. Ketogenic diets initiated in late mid-life improved measures of spatial memory in male mice. Geroscience. (2023) 45:2481–94. doi: 10.1007/s11357-023-00769-7
64. Newman, JC, Covarrubias, AJ, Zhao, M, Yu, X, Gut, P, Ng, CP, et al. Ketogenic diet reduces midlife mortality and improves memory in aging mice. Cell Metab. (2017) 26:547–557.e8. doi: 10.1016/j.cmet.2017.08.004
65. Roberts, MN, Wallace, MA, Tomilov, AA, Zhou, Z, Marcotte, GR, Tran, D, et al. A ketogenic diet extends longevity and Healthspan in adult mice. Cell Metab. (2017) 26:539–546.e5. doi: 10.1016/j.cmet.2017.08.005
66. Xu, K, Sun, X, Eroku, BO, Tsipis, CP, Puchowicz, MA, and LaManna, JC. Diet-induced ketosis improves cognitive performance in aged rats. Adv Exp Med Biol. (2010) 662:71–5. doi: 10.1007/978-1-4419-1241-1_9
67. Hernandez, AR, Watson, C, Federico, QP, Fletcher, R, Brotgandel, A, Buford, TW, et al. Twelve months of time-restricted feeding improves cognition and alters microbiome composition independent of macronutrient composition. Nutrients. (2022) 14:3977. doi: 10.3390/nu14193977
68. Hernandez, AR, Hernandez, CM, Campos, K, Truckenbrod, L, Federico, Q, Moon, B, et al. A ketogenic diet improves cognition and has biochemical effects in prefrontal cortex that are dissociable from hippocampus. Front Aging Neurosci. (2018) 10:391. doi: 10.3389/fnagi.2018.00391
69. Pathak, SJ, Zhou, Z, Steffen, D, Tran, T, Ad, Y, Ramsey, JJ, et al. 2-month ketogenic diet preferentially alters skeletal muscle and augments cognitive function in middle aged female mice. Aging Cell. (2022) 21:e13706. doi: 10.1111/acel.13706
70. Saito, ER, Warren, CE, Hanegan, CM, Larsen, JG, du Randt, JD, Cannon, M, et al. A novel ketone-supplemented diet improves recognition memory and hippocampal mitochondrial efficiency in healthy adult mice. Meta. (2022) 12:1019. doi: 10.3390/metabo12111019
71. Scichilone, JM, Yarraguntla, K, Charalambides, A, Harney, JP, and Butler, D. Environmental enrichment mitigates detrimental cognitive effects of ketogenic diet in weanling rats. J Mol Neurosci. (2016) 60:1–9. doi: 10.1007/s12031-016-0753-4
72. Miles, KN, and Skelton, MR. Male mice placed on a ketogenic diet from postnatal day (P) 21 through adulthood have reduced growth, are hypoactive, show increased freezing in a conditioned fear paradigm, and have spatial learning deficits. Brain Res. (2020) 1734:146697. doi: 10.1016/j.brainres.2020.146697
73. Singh, T, Mishra, A, and Goel, RK. PTZ kindling model for epileptogenesis, refractory epilepsy, and associated comorbidities: relevance and reliability. Metab Brain Dis. (2021) 36:1573–90. doi: 10.1007/s11011-021-00823-3
74. Novak, A, Vizjak, K, and Rakusa, M. Cognitive impairment in people with epilepsy. J Clin Med. (2022) 11:267. doi: 10.3390/jcm11010267
75. Schugar, RC, Huang, X, Moll, AR, Brunt, EM, and Crawford, PA. Role of choline deficiency in the fatty liver phenotype of mice fed a low protein, very low carbohydrate ketogenic diet. PLoS One. (2013) 8:74806. doi: 10.1371/journal.pone.0074806
76. Arvanitakis, Z, Wilson, RS, Bienias, JL, Evans, DA, and Bennett, DA. Diabetes mellitus and risk of Alzheimer disease and decline in cognitive function. Arch Neurol. (2004) 61:661–16. doi: 10.1001/archneur.61.5.661
77. Broom, GM, Shaw, IC, and Rucklidge, JJ. The ketogenic diet as a potential treatment and prevention strategy for Alzheimer’s disease. Nutrition. (2019) 60:118–21. doi: 10.1016/j.nut.2018.10.003
78. Versele, R, Corsi, M, Fuso, A, Sevin, E, Businaro, R, Gosselet, F, et al. Ketone bodies promote amyloid-β1-40 clearance in a human in vitro blood-brain barrier model. Int J Mol Sci. (2020) 21:934. doi: 10.3390/ijms21030934
79. Aso, E, Semakova, J, Joda, L, Semak, V, Halbaut, L, Calpena, A, et al. Triheptanoin supplementation to ketogenic diet curbs cognitive impairment in APP/PS1 mice used as a model of familial Alzheimer’s disease. Curr Alzheimer Res. (2013) 10:290–7. doi: 10.2174/15672050112099990128
80. Zhao, M, Huang, X, Cheng, X, Lin, X, Zhao, T, Wu, L, et al. Ketogenic diet improves the spatial memory impairment caused by exposure to hypobaric hypoxia through increased acetylation of histones in rats. PLoS One. (2017) 12:e0174477. doi: 10.1371/journal.pone.0174477
81. Benjamin, JS, Pilarowski, GO, Carosso, GA, Zhang, L, Huso, DL, Goff, LA, et al. A ketogenic diet rescues hippocampal memory defects in a mouse model of kabuki syndrome. Proc Natl Acad Sci USA. (2017) 114:125–30. doi: 10.1073/pnas.1611431114
82. Davidson, TL, Hargrave, SL, Swithers, SE, Sample, CH, Fu, X, Kinzig, KP, et al. Inter-relationships among diet, obesity and hippocampal-dependent cognitive function. Neuroscience. (2013) 253:110–22. doi: 10.1016/j.neuroscience.2013.08.044
83. Brownlow, ML, Jung, SH, Moore, RJ, Bechmann, N, and Jankord, R. Nutritional ketosis affects metabolism and behavior in Sprague-Dawley rats in both control and chronic stress environments. Front Mol Neurosci. (2017) 10:129. doi: 10.3389/fnmol.2017.00129
84. Liśkiewicz, AD, Liśkiewicz, D, Marczak, Ł, Przybyła, M, Grabowska, K, Student, S, et al. Obesity-associated deterioration of the hippocampus is partially restored after weight loss. Brain Behav Immun. (2021) 96:212–26. doi: 10.1016/j.bbi.2021.05.030
85. Brietzke, E, Mansur, RB, Subramaniapillai, M, Balanzá-Martínez, V, Vinberg, M, González-Pinto, A, et al. Ketogenic diet as a metabolic therapy for mood disorders: evidence and developments. Neurosci Biobehav Rev. (2018) 94:11–6. doi: 10.1016/j.neubiorev.2018.07.020
86. Włodarczyk, A, Cubała, WJ, and Stawicki, M. Ketogenic diet for depression: a potential dietary regimen to maintain euthymia? Prog Neuro-Psychopharmacol Biol Psychiatry. (2021) 109:110257. doi: 10.1016/j.pnpbp.2021.110257
87. Rawat, K, Singh, N, Kumari, P, and Saha, L. A review on preventive role of ketogenic diet (KD) in CNS disorders from the gut microbiota perspective. Rev Neurosci. (2020) 32:143–57. doi: 10.1515/revneuro-2020-0078
88. Primo, MJ, Fonseca-Rodrigues, D, Almeida, A, Teixeira, PM, and Pinto-Ribeiro, F. Sucrose preference test: a systematic review of protocols for the assessment of anhedonia in rodents. Eur Neuropsychopharmacol. (2023) 77:80–92. doi: 10.1016/j.euroneuro.2023.08.496
89. Murphy, P, Likhodii, S, Nylen, K, and Burnham, WM. The antidepressant properties of the ketogenic diet. Biol Psychiatry. (2004) 56:981–3. doi: 10.1016/j.biopsych.2004.09.019
90. Arqoub, AMS, Flynn, KG, and Martinez, LA. Gestational exposure to a ketogenic diet increases sociability in CD-1 mice. Behav Neurosci. (2020) 134:358–68. doi: 10.1037/bne0000368
91. Guan, YF, Huang, GB, Xu, MD, Gao, F, Lin, S, Huang, J, et al. Anti-depression effects of ketogenic diet are mediated via the restoration of microglial activation and neuronal excitability in the lateral habenula. Brain Behav Immun. (2020) 88:748–62. doi: 10.1016/j.bbi.2020.05.032
92. Sahagun, E, Ward, LM, and Kinzig, KP. Attenuation of stress-induced weight loss with a ketogenic diet. Physiol Behav. (2019) 212:112654. doi: 10.1016/j.physbeh.2019.112654
93. Sussman, D, Germann, J, and Henkelman, M. Gestational ketogenic diet programs brain structure and susceptibility to depression & anxiety in the adult mouse offspring. Brain Behav. (2015) 5:e00300. doi: 10.1002/brb3.300
94. Gumus, H, Ilgin, R, Koc, B, Yuksel, O, Kizildag, S, Guvendi, G, et al. A combination of ketogenic diet and voluntary exercise ameliorates anxiety and depression-like behaviors in Balb/c mice. Neurosci Lett. (2022) 770:136443. doi: 10.1016/j.neulet.2021.136443
95. Ryan, KK, Packard, AEB, Larson, KR, Stout, J, Fourman, SM, Thompson, AMK, et al. Dietary manipulations that induce ketosis activate the HPA Axis in male rats and mice: a potential role for fibroblast growth Factor-21. Endocrinology. (2018) 159:400–13. doi: 10.1210/en.2017-00486
96. Westmark, PR, Gutierrez, A, Gholston, AK, Wilmer, TM, and Westmark, CJ. Preclinical testing of the ketogenic diet in fragile X mice. Neurochem Int. (2020) 134:104687. doi: 10.1016/j.neuint.2020.104687
97. Acikgoz, B, Dalkiran, B, and Dayi, A. An overview of the currency and usefulness of behavioral tests used from past to present to assess anxiety, social behavior and depression in rats and mice. Behav Process. (2022) 200:104670. doi: 10.1016/j.beproc.2022.104670
98. Cryan, JF, Markou, A, and Lucki, I. Assessing antidepressant activity in rodents: recent developments and future needs. Trends Pharmacol Sci. (2002) 23:238–45. doi: 10.1016/s0165-6147(02)02017-5
99. Möhler, H. The GABA system in anxiety and depression and its therapeutic potential. Neuropharmacology. (2012) 62:42–53. doi: 10.1016/j.neuropharm.2011.08.040
100. Calderón, N, Betancourt, L, Hernández, L, and Rada, P. A ketogenic diet modifies glutamate, gamma-aminobutyric acid and agmatine levels in the hippocampus of rats: a microdialysis study. Neurosci Lett. (2017) 642:158–62. doi: 10.1016/j.neulet.2017.02.014
101. Attaye, I, van Oppenraaij, S, Warmbrunn, MV, and Nieuwdorp, M. The role of the Gut microbiota on the beneficial effects of ketogenic diets. Nutrients. (2021) 14:191. doi: 10.3390/nu14010191
102. Koh, S, Dupuis, N, and Auvin, S. Ketogenic diet and Neuroinflammation. Epilepsy Res. (2020) 167:106454. doi: 10.1016/j.eplepsyres.2020.106454
103. Tieu, K, Perier, C, Caspersen, C, Teismann, P, Wu, DC, Yan, SD, et al. D-beta-hydroxybutyrate rescues mitochondrial respiration and mitigates features of Parkinson disease. J Clin Invest. (2003) 112:892–901. doi: 10.1172/JCI18797
104. Włodarczyk, A, Cubała, WJ, and Wielewicka, A. Ketogenic diet: a dietary modification as an anxiolytic approach? Nutrients. (2020) 12:3822. doi: 10.3390/nu12123822
105. Dietch, DM, Kerr-Gaffney, J, Hockey, M, Marx, W, Ruusunen, A, Young, AH, et al. Efficacy of low carbohydrate and ketogenic diets in treating mood and anxiety disorders: systematic review and implications for clinical practice. BJPsych Open. (2023) 9:e70. doi: 10.1192/bjo.2023.36
106. Cryan, JF, and Sweeney, FF. The age of anxiety: role of animal models of anxiolytic action in drug discovery. Br J Pharmacol. (2011) 164:1129–61. doi: 10.1111/j.1476-5381.2011.01362.x
107. Harro, J. Animals, anxiety, and anxiety disorders: how to measure anxiety in rodents and why. Behav Brain Res. (2018) 352:81–93. doi: 10.1016/j.bbr.2017.10.016
108. Hoffman, KL. New dimensions in the use of rodent behavioral tests for novel drug discovery and development. Expert Opin Drug Discov. (2016) 11:343–53. doi: 10.1517/17460441.2016.1153624
109. Ennaceur, A. Tests of unconditioned anxiety - pitfalls and disappointments. Physiol Behav. (2014) 135:55–71. doi: 10.1016/j.physbeh.2014.05.032
110. Vogel, JR, Beer, B, and Clody, DE. A simple and reliable conflict procedure for testing anti-anxiety agents. Psychopharmacologia. (1971) 21:1–7. doi: 10.1007/BF00403989
111. Geller, I, Kulak, JT Jr, and Seifter, J. The effects of chlordiazepoxide and chlorpromazine on a punishment discrimination. Psychopharmacologia. (1962) 3:374–85. doi: 10.1007/BF00408322
112. Ziegler, DR, Gamaro, GD, Araújo, E, Bassani, MG, Perry, ML, Dalmaz, C, et al. Nociception and locomotor activity are increased in ketogenic diet fed rats. Physiol Behav. (2005) 84:421–7. doi: 10.1016/j.physbeh.2005.01.003
113. Wang, D, and Mitchell, ES. Cognition and synaptic-plasticity related changes in aged rats supplemented with 8- and 10-carbon medium chain triglycerides. PLoS One. (2016) 11:e0160159. doi: 10.1371/journal.pone.0160159
114. De La Rossa, A, Laporte, MH, Astori, S, Marissal, T, Montessuit, S, Sheshadri, P, et al. Paradoxical neuronal hyperexcitability in a mouse model of mitochondrial pyruvate import deficiency. elife. (2022) 11:72595. doi: 10.7554/eLife.72595
115. Qin, L, Ma, K, and Yan, Z. Rescue of histone hypoacetylation and social deficits by ketogenic diet in a Shank3 mouse model of autism. Neuropsychopharmacology. (2022) 47:1271–9. doi: 10.1038/s41386-021-01212-1
116. Ruskin, DN, Svedova, J, Cote, JL, Sandau, U, Rho, JM, Kawamura, M Jr, et al. Ketogenic diet improves core symptoms of autism in BTBR mice. PLoS One. (2013) 8:65021. doi: 10.1371/journal.pone.0065021
117. Ahn, Y, Narous, M, Tobias, R, Rho, JM, and Mychasiuk, R. The ketogenic diet modifies social and metabolic alterations identified in the prenatal valproic acid model of autism spectrum disorder. Dev Neurosci. (2014) 36:371–80. doi: 10.1159/000362645
118. Ruskin, DN, Fortin, JA, Bisnauth, SN, and Masino, SA. Ketogenic diets improve behaviors associated with autism spectrum disorder in a sex-specific manner in the EL mouse. Physiol Behav. (2017) 168:138–45. doi: 10.1016/j.physbeh.2016.10.023
119. Ruskin, DN, Murphy, MI, Slade, SL, and Masino, SA. Ketogenic diet improves behaviors in a maternal immune activation model of autism spectrum disorder. PLoS One. (2017) 12:e0171643. doi: 10.1371/journal.pone.0171643
120. Bornebusch, AB, Mason, GF, Tonetto, S, Damsgaard, J, Gjedde, A, Fink-Jensen, A, et al. Effects of ketogenic diet and ketone monoester supplement on acute alcohol withdrawal symptoms in male mice. Psychopharmacology. (2021) 238:833–44. doi: 10.1007/s00213-020-05735-1
121. Ari, C, Kovács, Z, Juhasz, G, Murdun, C, Goldhagen, CR, Koutnik, AP, et al. Exogenous ketone supplements reduce anxiety-related behavior in Sprague-Dawley and Wistar albino Glaxo/Rijswijk rats. Front Mol Neurosci. (2016) 9:137. doi: 10.3389/fnmol.2016.00137
122. Napoli, E, Dueñas, N, and Giulivi, C. Potential therapeutic use of the ketogenic diet in autism spectrum disorders. Front Pediatr. (2014) 2:69. doi: 10.3389/fped.2014.00069
123. Dougnon, G, and Matsui, H. Modelling autism Spectrum disorder (ASD) and attention-deficit/hyperactivity disorder (ADHD) using mice and zebrafish. Int J Mol Sci. (2022) 23:7550. doi: 10.3390/ijms23147550
124. Kim, DG, Gonzales, EL, Kim, S, Kim, Y, Adil, KJ, Jeon, SJ, et al. Social interaction test in home cage as a novel and ethological measure of social behavior in mice. Exp Neurobiol. (2019) 28:247–60. doi: 10.5607/en.2019.28.2.247
125. Tinguely, D, Gross, J, and Kosinski, C. Efficacy of ketogenic diets on type 2 diabetes: a systematic review. Curr Diab Rep. (2021) 21:32. doi: 10.1007/s11892-021-01399-z
126. Bueno, NB, de Melo, IS, de Oliveira, SL, and da Rocha, AT. Very-low-carbohydrate ketogenic diet v. low-fat diet for long-term weight loss: a meta-analysis of randomised controlled trials. Br J Nutr. (2013) 110:1178–87. doi: 10.1017/S0007114513000548
127. Chawla, S, Tessarolo Silva, F, Amaral Medeiros, S, Mekary, RA, and Radenkovic, D. The effect of low-fat and low-carbohydrate diets on weight loss and lipid levels: a systematic review and meta-analysis. Nutrients. (2020) 12:3774. doi: 10.3390/nu12123774
128. Mansoor, N, Vinknes, KJ, Veierød, MB, and Retterstøl, K. Effects of low-carbohydrate diets v. low-fat diets on body weight and cardiovascular risk factors: a meta-analysis of randomised controlled trials. Br J Nutr. (2016) 115:466–79. doi: 10.1017/S0007114515004699
129. Paoli, A, Bosco, G, Camporesi, EM, and Mangar, D. Ketosis, ketogenic diet and food intake control: a complex relationship. Front Psychol. (2015) 6:27. doi: 10.3389/fpsyg.2015.00027
130. Roekenes, J, and Martins, C. Ketogenic diets and appetite regulation. Curr Opin Clin Nutr Metab Care. (2021) 24:359–63. doi: 10.1097/MCO.0000000000000760
131. Sumithran, P, Prendergast, LA, Delbridge, E, Purcell, K, Shulkes, A, Kriketos, A, et al. Long-term persistence of hormonal adaptations to weight loss. N Engl J Med. (2011) 365:1597–604. doi: 10.1056/NEJMoa1105816
132. Polidori, D, Sanghvi, A, Seeley, RJ, and Hall, KD. How strongly does appetite counter weight loss? Quantification of the feedback control of human energy intake. Obesity (Silver Spring). (2016) 24:2289–95. doi: 10.1002/oby.21653
133. Ali, MA, and Kravitz, AV. Challenges in quantifying food intake in rodents. Brain Res. (2018) 1693:188–91. doi: 10.1016/j.brainres.2018.02.040
134. Streijger, F, Plunet, WT, Lee, JH, Liu, J, Lam, CK, Park, S, et al. Ketogenic diet improves forelimb motor function after spinal cord injury in rodents. PLoS One. (2013) 8:78765. doi: 10.1371/journal.pone.0078765
135. Liśkiewicz, D, Liśkiewicz, A, Grabowski, M, Nowacka-Chmielewska, MM, Jabłońska, K, Wojakowska, A, et al. Upregulation of hepatic autophagy under nutritional ketosis. J Nutr Biochem. (2021) 93:108620. doi: 10.1016/j.jnutbio.2021.108620
136. Kinzig, KP, and Taylor, RJ. Maintenance on a ketogenic diet: voluntary exercise, adiposity and neuroendocrine effects. Int J Obes. (2009) 33:824–30. doi: 10.1038/ijo.2009.109
137. Hsu, YJ, Huang, CC, and Lin, CI. The effect of a low carbohydrate ketogenic diet with or without exercise on postpartum weight retention, metabolic profile and physical activity performance in postpartum mice. J Nutr Biochem. (2022) 102:108941. doi: 10.1016/j.jnutbio.2022.108941
138. Srivastava, S, Baxa, U, Niu, G, Chen, X, and Veech, RL. A ketogenic diet increases brown adipose tissue mitochondrial proteins and UCP1 levels in mice. IUBMB Life. (2013) 65:58–66. doi: 10.1002/iub.1102
139. Guo, M, Wang, X, Zhao, Y, Yang, Q, Ding, H, Dong, Q, et al. Ketogenic diet improves brain ischemic tolerance and inhibits NLRP3 Inflammasome activation by preventing Drp1-mediated mitochondrial fission and endoplasmic reticulum stress. Front Mol Neurosci. (2018) 11:86. doi: 10.3389/fnmol.2018.00086
140. Jornayvaz, FR, Jurczak, MJ, Lee, HY, Birkenfeld, AL, Frederick, DW, Zhang, D, et al. A high-fat, ketogenic diet causes hepatic insulin resistance in mice, despite increasing energy expenditure and preventing weight gain. Am J Physiol Endocrinol Metab. (2010) 299:E808–15. doi: 10.1152/ajpendo.00361.2010
141. Morrow, NM, Locatelli, CAA, Trzaskalski, NA, Klein, CT, Hanson, AA, Alhadi, H, et al. Adaptation to short-term extreme fat consumption alters intestinal lipid handling in male and female mice. Biochim Biophys Acta Mol Cell Biol Lipids. (2022) 1867:159208. doi: 10.1016/j.bbalip.2022.159208
142. Likhodii, SS, Musa, K, Mendonca, A, Dell, C, Burnham, WM, and Cunnane, SC. Dietary fat, ketosis, and seizure resistance in rats on the ketogenic diet. Epilepsia. (2000) 41:1400–10. doi: 10.1111/j.1528-1157.2000.tb00115.x
143. Ma, D, Wang, AC, Parikh, I, Green, SJ, Hoffman, JD, Chlipala, G, et al. Ketogenic diet enhances neurovascular function with altered gut microbiome in young healthy mice. Sci Rep. (2018) 8:6670. doi: 10.1038/s41598-018-25190-5
144. Kennedy, AR, Pissios, P, Otu, H, Roberson, R, Xue, B, Asakura, K, et al. A high-fat, ketogenic diet induces a unique metabolic state in mice. Am J Physiol Endocrinol Metab. (2007) 292:E1724–39. doi: 10.1152/ajpendo.00717.2006
145. Holcomb, LE, O’Neill, CC, DeWitt, EA, and Kolwicz, SC Jr. The effects of fasting or ketogenic diet on endurance exercise performance and metabolism in female mice. Meta. (2021) 11:397. doi: 10.3390/metabo11060397
146. Zhou, J, Lu, Y, Jia, Y, Lu, J, Jiang, Z, and Chen, K. Ketogenic diet ameliorates lipid dysregulation in type 2 diabetic mice by downregulating hepatic pescadillo 1. Mol Med. (2022) 28:1. doi: 10.1186/s10020-021-00429-6
147. Harun-Or-Rashid, M, Pappenhagen, N, Palmer, PG, Smith, MA, Gevorgyan, V, Wilson, GN, et al. Structural and functional Rescue of Chronic Metabolically Stressed Optic Nerves through respiration. J Neurosci. (2018) 38:5122–39. doi: 10.1523/JNEUROSCI.3652-17.2018
148. Lee, J, Choi, J, Scafidi, S, and Wolfgang, MJ. Hepatic fatty acid oxidation restrains systemic catabolism during starvation. Cell Rep. (2016) 16:201–12. doi: 10.1016/j.celrep.2016.05.062
149. Guo, Y, Liu, X, Li, T, Zhao, J, Yang, Y, Yao, Y, et al. Alternate-day ketogenic diet feeding protects against heart failure through preservation of Ketogenesis in the liver. Oxidative Med Cell Longev. (2022) 2022:4253651–13. doi: 10.1155/2022/4253651
150. Moore, MP, Cunningham, RP, Kelty, TJ, Boccardi, LR, Nguyen, NY, Booth, FW, et al. Ketogenic diet in combination with voluntary exercise impacts markers of hepatic metabolism and oxidative stress in male and female Wistar rats. Appl Physiol Nutr Metab. (2020) 45:35–44. doi: 10.1139/apnm-2019-0042
151. da Silva, IV, Gullette, S, Florindo, C, Huang, NK, Neuberger, T, Ross, AC, et al. The effect of nutritional ketosis on aquaporin expression in apolipoprotein E-deficient mice: potential implications for energy homeostasis. Biomedicines. (2022) 10:1159. doi: 10.3390/biomedicines10051159
152. Ma, S, Huang, Q, Yada, K, Liu, C, and Suzuki, K. An 8-week ketogenic low carbohydrate, high fat diet enhanced exhaustive exercise capacity in mice. Nutrients. (2018) 10:673. doi: 10.3390/nu10060673
153. Irfannuddin, I, Sarahdeaz, SFP, Murti, K, Santoso, B, and Koibuchi, N. The effect of ketogenic diets on neurogenesis and apoptosis in the dentate gyrus of the male rat hippocampus. J Physiol Sci. (2021) 71:3. doi: 10.1186/s12576-020-00786-7
154. Castro, R, Whalen, CA, Gullette, S, Mattie, FJ, Florindo, C, Heil, SG, et al. A Hypomethylating ketogenic diet in apolipoprotein E-deficient mice: a pilot study on vascular effects and specific epigenetic changes. Nutrients. (2021) 13:3576. doi: 10.3390/nu13103576
155. Huang, Q, Ma, S, Tominaga, T, Suzuki, K, and Liu, C. An 8-week, low carbohydrate, high fat, ketogenic diet enhanced exhaustive exercise capacity in mice part 2: effect on fatigue recovery, post-exercise biomarkers and anti-oxidation capacity. Nutrients. (2018) 10:1339. doi: 10.3390/nu10101339
156. Kosiek, W, Rauk, Z, Szulc, P, Cichy, A, Rugieł, M, Chwiej, J, et al. Ketogenic diet impairs neurological development of neonatal rats and affects biochemical composition of maternal brains: evidence of functional recovery in pups. Brain Struct Funct. (2022) 227:1099–113. doi: 10.1007/s00429-021-02450-1
157. Cortez, NE, Pathak, S, Rodriguez Lanzi, C, Hong, BV, Crone, R, Sule, R, et al. A ketogenic diet in combination with gemcitabine mitigates pancreatic cancer-associated cachexia in male and female KPC mice. Int J Mol Sci. (2023) 24:10753. doi: 10.3390/ijms241310753
158. Lin, J, Huang, Z, Liu, J, Huang, Z, Liu, Y, Liu, Q, et al. Neuroprotective effect of ketone metabolism on inhibiting inflammatory response by regulating macrophage polarization after acute cervical spinal cord injury in rats. Front Neurosci. (2020) 14:583611. doi: 10.3389/fnins.2020.583611
159. Takeuchi, F, Nishikata, N, Nishimura, M, Nagao, K, and Kawamura, M Jr. Leucine-enriched essential amino acids enhance the Antiseizure effects of the ketogenic diet in rats. Front Neurosci. (2021) 15:637288. doi: 10.3389/fnins.2021.637288
160. Nakao, R, Abe, T, Yamamoto, S, and Oishi, K. Ketogenic diet induces skeletal muscle atrophy via reducing muscle protein synthesis and possibly activating proteolysis in mice. Sci Rep. (2019) 9:19652. doi: 10.1038/s41598-019-56166-8
161. Kalafut, KC, Mitchell, SJ, MacArthur, MR, and Mitchell, JR. Short-term ketogenic diet induces a molecular response that is distinct from dietary protein restriction. Front Nutr. (2022) 9:839341. doi: 10.3389/fnut.2022.839341
162. Kraeuter, AK, van den Buuse, M, and Sarnyai, Z. Ketogenic diet prevents impaired prepulse inhibition of startle in an acute NMDA receptor hypofunction model of schizophrenia. Schizophr Res. (2019) 206:244–50. doi: 10.1016/j.schres.2018.11.011
163. Tozzi, R, Campolo, F, Baldini, E, Venneri, MA, Lubrano, C, Ulisse, S, et al. Ketogenic diet increases serum and white adipose tissue SIRT1 expression in mice. Int J Mol Sci. (2022) 23:15860. doi: 10.3390/ijms232415860
164. Licholai, JA, Nguyen, KP, Fobbs, WC, Schuster, CJ, Ali, MA, and Kravitz, AV. Why do mice overeat high-fat diets? How high-fat diet alters the regulation of daily caloric intake in mice. Obesity (Silver Spring). (2018) 26:1026–33. doi: 10.1002/oby.22195
165. de Wouters, d’O A, Rastelli, M, Van Hul, M, Delzenne, NM, Cani, PD, and Everard, A. Gut microbes participate in food preference alterations during obesity. Gut Microbes. (2021) 13:1959242. doi: 10.1080/19490976.2021.1959242
166. da Rocha, LS, Mendes, CB, Silva, JS, Alcides, RLGF, Mendonça, IP, Andrade-da-Costa, BLS, et al. Triheptanoin, an odd-medium-chain triglyceride, impacts brain cognitive function in young and aged mice. Nutr Neurosci. (2023) 21:1–11. doi: 10.1080/1028415X.2023.2178096
167. Kackley, ML, Brownlow, ML, Buga, A, Crabtree, CD, Sapper, TN, O’Connor, A, et al. The effects of a 6-week controlled, hypocaloric ketogenic diet, with and without exogenous ketone salts, on cognitive performance and mood states in overweight and obese adults. Front Neurosci. (2022) 16:971144. doi: 10.3389/fnins.2022.971144
Glossary
Keywords: ketogenic diet, cognition, depressive-like behavior, anxiety-like behavior, social behavior, nutritional behavior, nutritional ketosis, animal models
Citation: Grabowska K, Grabowski M, Przybyła M, Pondel N, Barski JJ, Nowacka-Chmielewska M and Liśkiewicz D (2024) Ketogenic diet and behavior: insights from experimental studies. Front. Nutr. 11:1322509. doi: 10.3389/fnut.2024.1322509
Edited by:
Beth Ann Zupec-Kania, Ketogenic Therapies, LLC, United StatesReviewed by:
Heather M. Wilkins, University of Kansas Medical Center Research Institute, United StatesMadison Lee Kackley, The Ohio State University, United States
Abbi R. Hernandez, University of Alabama at Birmingham, United States
Aycan Ünalp, University of Health Sciences, Türkiye
Copyright © 2024 Grabowska, Grabowski, Przybyła, Pondel, Barski, Nowacka-Chmielewska and Liśkiewicz. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Daniela Liśkiewicz, ZC5saXNraWV3aWN6QGF3Zi5rYXRvd2ljZS5wbA==
†These authors have contributed equally to this work and share first authorship
‡These authors have contributed equally to this work