 Janaina L. S. Donadio
Janaina L. S. Donadio João Paulo Fabi
João Paulo Fabi Marcelo B. Sztein
Marcelo B. Sztein Rosângela Salerno-Gonçalves
Rosângela Salerno-Gonçalves
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Nutr. , 12 January 2024
Sec. Nutrition and Microbes
Volume 10 - 2023 | https://doi.org/10.3389/fnut.2023.1286138
This article is part of the Research Topic Exploring Dietary Polysaccharides: Molecular Insights into Gastrointestinal Health and the Brain-Gut Axis View all 6 articles
Pectins, a class of dietary fibers abundant in vegetables and fruits, have drawn considerable interest due to their potential anti-inflammatory properties. Numerous studies have indicated that incorporating pectins into infant formula could be a safe strategy for alleviating infant regurgitation and diarrhea. Moreover, pectins have been shown to modulate cytokine production, macrophage activity, and NF-kB expression, all contributing to their anti-inflammatory effects. Despite this promising evidence, the exact mechanisms through which pectins exert these functions and how their structural characteristics influence these processes remain largely unexplored. This knowledge is particularly significant in the context of gut inflammation in developing preterm babies, a critical aspect of necrotizing enterocolitis (NEC), and in children and adults dealing with inflammatory bowel disease (IBD). Our mini review aims to provide an up-to-date compilation of relevant research on the effects of pectin on gut immune responses, specifically focusing on preterms and newborns. By shedding light on the underlying mechanisms and implications of pectin-mediated anti-inflammatory properties, this review seeks to advance our knowledge in this area and pave the way for future research and potential therapeutic interventions.
Pectins, a class of dietary fibers naturally found in fruits and vegetables (1), exhibit highly viscous and water-soluble properties and are susceptible to fermentation by the intestinal microbiota (2). Multiple studies have suggested that incorporating pectins into infant formula is safe and could alleviate infant regurgitation and diarrhea, while also promoting overall physical development (3–10). Several clinical trials involving regurgitating infants fed with a pectin-thickened formula have reported reduced regurgitation episodes (3–7). Furthermore, due to its remarkable water-retention properties (11), pectin may also influence infant stool frequency, consistency, and potentially reduce the incidence of diarrhea (3, 8, 9, 12). Pectins, being extracted from plant material without undergoing chemical modification, primarily consist of polysaccharides with a small proportion of oligosaccharides (13). While there are structural differences between pectin oligosaccharides (POS) and human milk oligosaccharides (HMOs) (14), the ingestion of POS has been shown to influence the composition of the infant’s fecal microbiota and provide benefits akin to HMOs, including a decreased risk of infection by pathogenic bacteria and virus (15, 16). It is worth noting that HMOs represent the most significant solid component in breast milk, surpassing carbohydrates and fat (17). HMO concentration is highest in colostrum (20 g/L) and then decreases by about 20% at day 30 of lactation (18).
The molecular structure of pectins is notably complex, consisting of a homogalacturonan (HG) backbone core region comprising 1,4-α-D-galacturonic acid (GalA) units that can undergo methylation or acetylation, along with branched regions primarily composed of rhamnogalacturonan type I (RGI) (19). The HG backbone incorporates monosaccharides with β-(1,3)-D-xylose residues (xylogalacturonan) or branched structures with alternating α-(1,4)-GalA and α-(1,2)-rhamnose (Rha), the latter of which can be linked to RGI (20). RGI possesses intricate side chains of neutral sugars attached to the rhamnose residues, while the HG backbone may also include complex branches like aceric acid and apiose with distinct side chains, referred to as rhamnogalacturonan type II (RGII) (21). After HG, RGI is plants’ second major pectic fraction (21) (Figure 1). While pectins share the same fundamental repeating elements, their quantities and chemical structures can vary depending on their source, location within the plant, and the extraction methods employed (15). Pectins demonstrated varying degrees of esterification and a wide range of molecular weights, ranging from 60 to 900 kDa, precluding their intestinal absorption (19, 20, 22, 23). Notably, the structural characteristics of pectin directly influence the development of gut microbial communities (15).
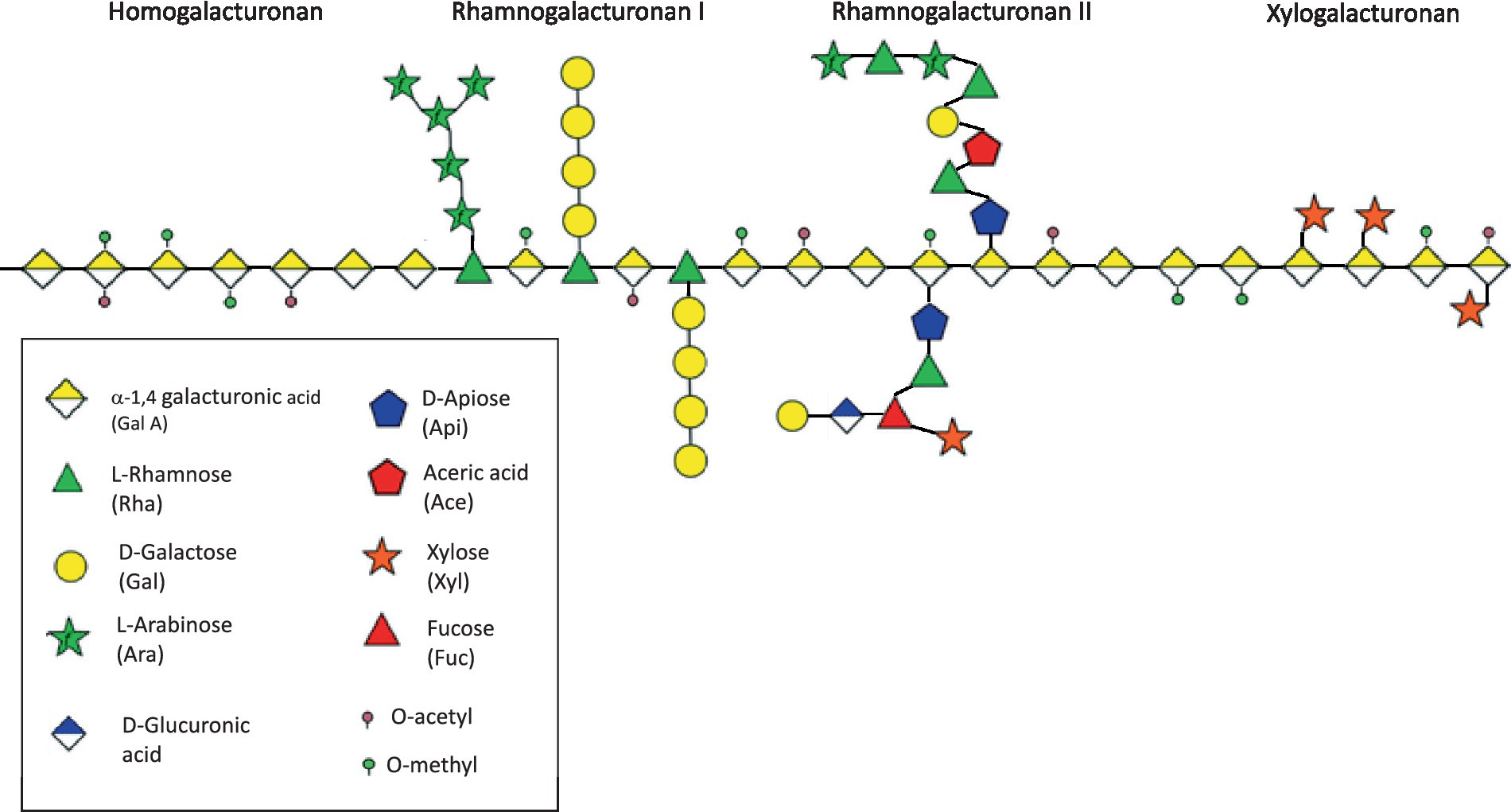
Figure 1. Schematic structure of pectins.
In addition, pectins can have dual immunomodulatory effects: (a) direct interactions with the intestinal barrier and engagement with immune receptors, such as Toll-like receptors (TLRs), resulting in reduced inflammation, and (b) indirect effects via modulation of the gut microbiota through fermentation and production of SCFA (24–26). These properties are particularly relevant in intestinal inflammatory diseases, such as necrotizing enterocolitis (NEC), which remains a leading cause of mortality in premature neonates (27). In this review, we aim to provide a comprehensive overview of the immunomodulatory effects of pectins in the context of preterms and newborns and their potential role in modulating the neonatal gut epithelial barrier and microbiota. Understanding the impact of pectins on the epithelium and microbiota can offer valuable insights into their potential therapeutic applications for mitigating intestinal inflammation and promoting neonatal health.
Preterm neonates exhibit a distinct gut microbiota composition compared to full-term neonates (28). Unlike vaginally born infants, preterm neonates delivered by cesarean section (C-section) tend to be colonized by maternal skin bacteria, leading to an abundance of Staphylococcus, Corynebacterium, Propionibacterium spp., and a deficiency of Lactobacillus, Bifidobacterium, and Bacteroides (28). This difference in microbiota colonization can significantly affect the neonate’s immune system (28, 29). Notably, C-section delivery has been associated with an increased risk of celiac disease, asthma, obesity, and type 1 diabetes in newborns (29). During vaginal delivery, Escherichia coli, Staphylococcus, and Streptococcus, play a crucial role in creating an anaerobic environment, which allows the colonization of strict commensal anaerobes such as Bacteroides, Clostridium, and Bifidobacterium spp. (29, 30). The diversity of the infant gut microbiota continues to increase over time with a significant shift at weaning (31, 32). This process is delayed in preterm neonates delivered by C-section, potentially affecting the growth of commensal anaerobic bacteria (29).
Apart from the mode of delivery, other factors can disrupt the neonate’s microbiota, including early antibiotic use and feeding practices. Early antibiotic administration reduces microbiome diversity in the neonate’s stool and eliminates the growth of commensal bacteria impairing the innate immune defense (29, 33, 34). Microbial colonization triggers and accompanies rapid morphological and functional changes in the gut (31, 32). It has been proposed that impaired colonization in preterm infants leads to dysmotility of the intestinal tract, and uncontrolled inflammation, triggering disease states including neonatal NEC (27, 35). The host’s central strategy to maintain its homeostatic relationship with the microbiota is to minimize contact between the microbiota and the epithelial cell surface, thereby limiting tissue inflammation (36). This segregation is partially accomplished by secretory-IgA (SIgA) (37). However, the presence of SIgA, a predominant immunoglobulin in the human gut, relies on the gastrointestinal tract’s colonization by microbiota. SIgA is critical in attenuating inflammatory reactions in the intestine, particularly in preterm infants (37–40). For example, germ-free mice, lacking microbiota, demonstrate a marked reduction of SIgA in the gut (41–43). Moreover, previous studies have shown that pectin-fed animals have significantly higher levels of SIgA and IgA than controls (e.g., cellulose-fed animals) (44–47).
The feeding modality can also shape the infant’s gut microbiota. Formula feeding, for instance, can impact the microbiota by increasing the prevalence of Clostridium difficile, Bacteroides fragilis, and Escherichia coli, while reducing the prevalence of beneficial commensal bifidobacteria (29). Lack of breastfeeding prevents neonates from acquiring prebiotics from human breast milk, resulting in lower microbial diversity and unusual gut colonization with pathogenic proteobacteria, such as Enterobacter, Escherichia, and Klebsiella in preterm newborns (48–50). The immature intestinal mucosal barrier in preterms also allows the passage of pathogenic bacteria and bacterial toxins through the epithelial cells (49). Finally, infants can inherit bacteria associated with antibiotic resistance from their mothers through breastfeeding (51).
Since, pectin and POS are fermented in the intestine by Bacteroides, Bifidobacteria, Lactobacilli, Enterococcus, and Clostridium (13), it is anticipated that the microbial community in preterm neonates will differ in its ability to metabolize pectin substrates and, consequently, produce short-chain fatty acids (SCFA) compared to full-term babies. Unlike full-term babies, preterm infants with a deficiency of Lactobacillus, Bifidobacterium, and Bacteroides are expected to exhibit a reduced capacity for pectin degradation. Bacteroides, known for their pivotal role in breaking down various plant polymers, possess many carbohydrate-active enzyme (CAZymes) genes (52). Furthermore, pectin substrates may also foster the growth of beneficial bacteria. For instance, RGI-enriched citrus pectin has been shown to selectively promote the growth of Bifidobacterium, Lactobacillus, and Faecalibaculum spp. (53). Larsen and colleagues have also demonstrated that the abundance of beneficial bacteria such as Bifidobacterium, Christensenellaceae, Prevotella copri, and Bacteroides spp. can either increase or decrease depending on the specific pectin substrate, suggesting that the microbial community in preterm infants can be modulated using structurally different pectins to promote the growth of more beneficial bacteria (54).
NEC is a severe inflammatory disease of premature neonates’ gastrointestinal tract, characterized by intense intestinal necrosis (55, 56). NEC’s mortality rate can reach 30%, leaving the survivors with severe neurodevelopmental delays (56, 57). While the exact mechanisms responsible for NEC development are still debated, they may involve factors such as the prematurity of the intestine, intense production of inflammatory cytokines, defective mucus production, and low expression of tight junctions (TJ) proteins, leading to increased intestinal permeability and penetration of pathogenic bacteria and toxin, causing tissue injury and intestinal necrosis (56, 58). One potential mechanism by which pectins can contribute to preventing intestinal inflammatory diseases is by preserving the integrity of the intestinal layer and enhancing mucosal immunity (24). The intestinal layer is safeguarded by several physical barriers, including gastric acid, the mucus layer, and a tight monolayer of intestinal epithelial cells (IECs) held together by TJ to prevent the transfer of pathogens and toxins from the lumen into the circulation (59). These TJ play a crucial role in maintaining the gastrointestinal barrier’s integrity by regulating the permeability of the intestinal cell layer (58).
The mucus layer is a protective barrier separating the IECs from the luminal content and the microbiota. It mainly consists of mucins, glycoproteins produced by goblet cells, with MUC2 being the predominant mucin in the small and large intestines (59). Pectins can stimulate the production of MUC2 (60). Additionally, the mucus layer contains defensins, antimicrobial components derived from Paneth cells, and SIgA that protects against pathogen invasion (61–63). The mucus coating is composed of two layers: an outer layer, housing commensal bacteria, and an inner layer, which acts as a barrier against bacterial penetration (64). Neonates with NEC may have fewer goblet cells producing mucus, resulting in impaired mucus production after infection (65, 66). While not yet confirmed in humans, a neonatal rat NEC model demonstrates that HMOs provide protection against NEC (67). Survival rates and pathology scores show significant improvement when HMOs are introduced into orally administered formula. These beneficial effects are hypothesized to be mediated through specific receptors that mimic pathogen lectins, preventing interactions with host glycans. It is worth noting that pectin and POS also exhibit antiadhesive and antimicrobial properties, similar to HMOs (14). Additionally, pectins possess mucoadhesive properties by adhering to densely mucin-grafted glycans (68) and preventing pathogen colonization.
NEC patients often exhibit higher production of pro-inflammatory cytokines, such as TNF-α and IL-1β, which increase intestinal TJ permeability, causing bacterial translocation and boosting the inflammatory state in the gut (58). Pectins can also directly interact with TLR signaling pathways, thereby reducing inflammation (69, 70). For instance, pectins have been found to inhibit IL-6 secretion induced by TLR2-1 (70, 71). Additionally, TLR4 can serve as a receptor for non-canonical ligands, including carbohydrates present in pectins (72). It is believed that the increased expression of TLR4 on IECs might explain the excessive inflammatory response with high production of pro-inflammatory cytokines including IL-6, IL-8, and TNF-α found in NEC patients (73). TLR4 is responsible for detecting lipopolysaccharide (LPS), a critical outer membrane component of gram-negative bacteria, which stimulates the NF-κb pathway to produce pro-inflammatory cytokines, such as IL-8 and TNF-α (74). TLR4 also increases intestinal stem cell apoptosis and decreases IEC proliferation and migration, impairing intestinal mucosal healing and regeneration, thereby favoring NEC development (56).
Several studies have observed that pectins with different structures have distinct modulatory effects on the immune system. Most of the studies were conducted using pectins from citrus (e.g., lemon) (75, 76), but also with pectins from apple (77), cacao (78), and papaya (69). Pectins have a backbone composed of GalA that varies in the degree of methyl-esterification (DM), and the immunomodulatory effects of pectins are dependent on the DM. In vitro and in vivo studies (Tables 1, 2, respectively) showed that pectins, with different chemical structures and degrees of DM, can strengthen the mucus layer by directly stimulating mucin production by goblet cells.
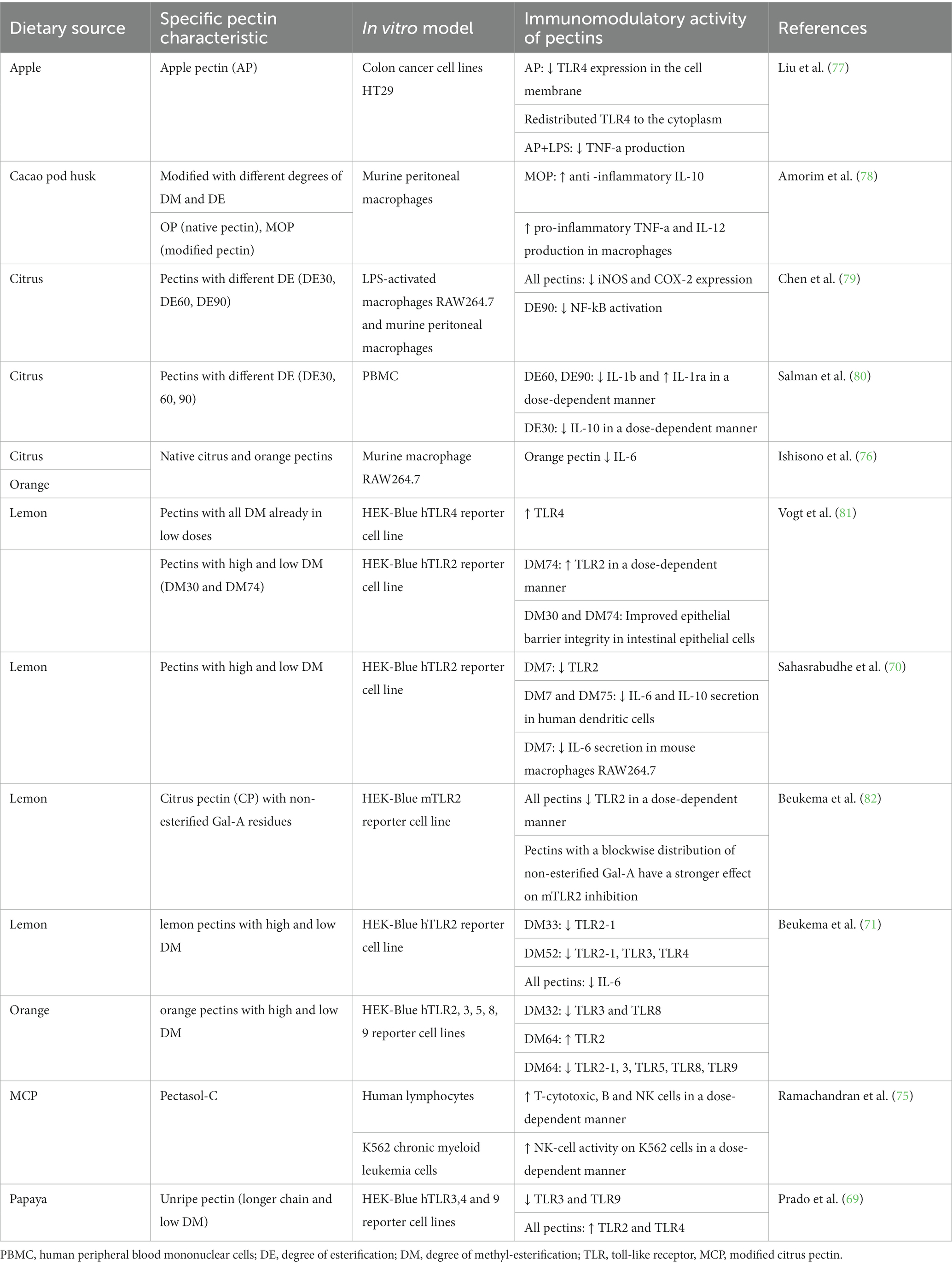
Table 1. In vitro studies on the immunoregulatory activities of pectins.
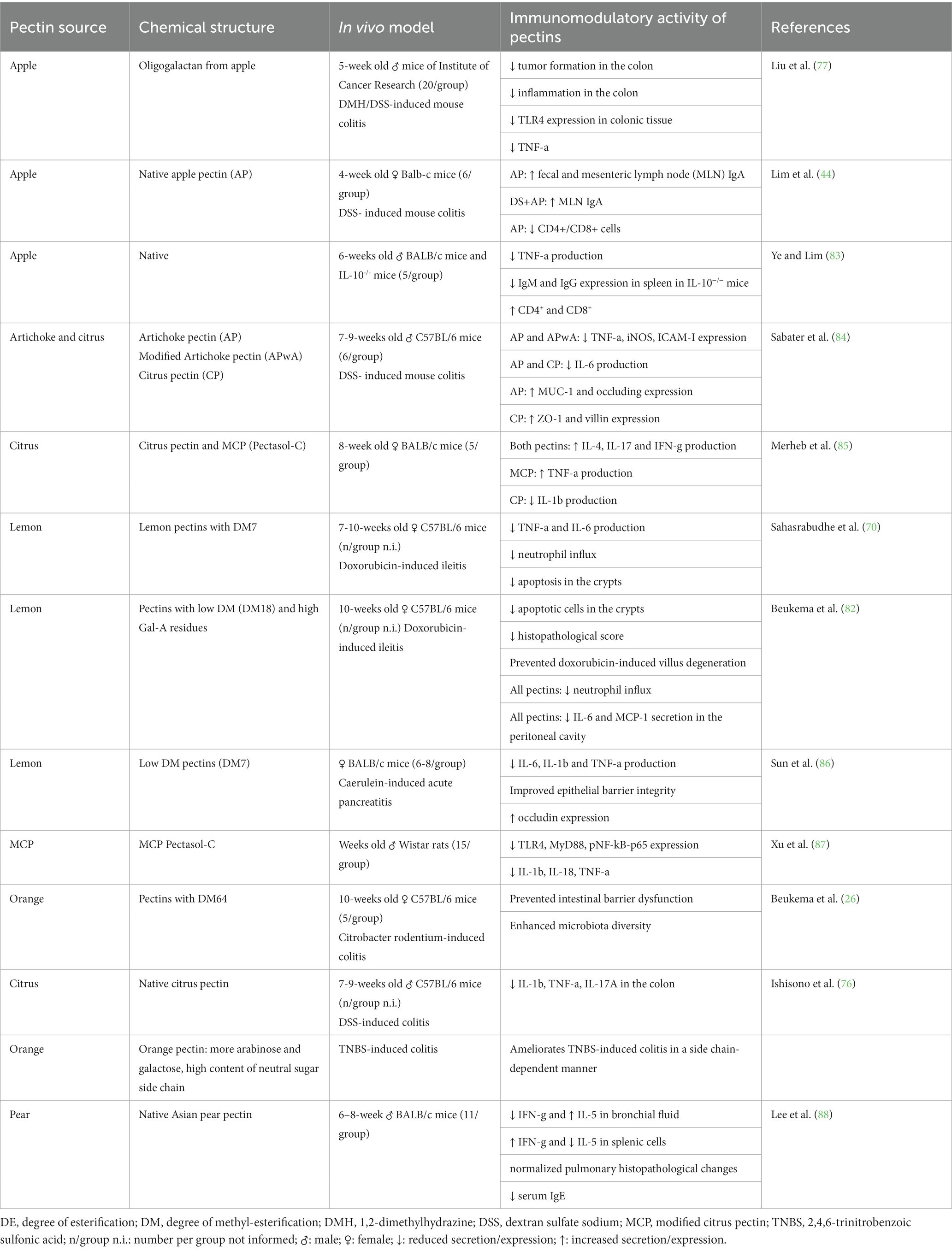
Table 2. In vivo studies on the immunoregulatory effects of pectins.
In vitro studies have shown that pectins from citrus, lemon, and orange with higher DM (DM52 to 90) were able to reduce expression of NF-κB, IL-1β, IL-6, and IL-10 in a dose-dependent manner, reduce the activation of TLR2-1, TLR3, and TLR4, and increase the activation of TLR2 (70, 71, 79–81). In contrast, pectins with low DM (DM7 to 30) from citrus, lemon, orange, and papaya improved epithelial barrier integrity, reduced secretion of IL-10, and IL-6 in a dose-dependent manner, and reduced the activation of TLR2, TLR2-1, TLR3, TLR8, TLR9 [69; 70; 71; 80; 81]. Interestingly, the reduction of IL-6, iNOS, and COX-2 expression, the activation of TLR2 and TLR4, and the inhibition of TLR2 were achieved for some pectins regardless of the DM (69, 71, 79, 81, 82). Pectins with a low DM of 50 can penetrate the mucin layer and interact with IEC (24). Treating polarized monolayers of human T84 intestinal epithelial cells with lemon pectin, especially pectins with DM30 and DM74, can also improve their transepithelial electrical resistance (81). Moreover, low-methoxyl pectin from lemon can restore epithelial barrier integrity by increasing TJ protein expression such as occluding and zonula occludens (ZO-1) (86). The immunomodulatory effects of pectins in in vitro studies has been summarized in Table 1.
An increasing amount of evidence on in vivo models (Table 2) suggests that pectins from apple, artichoke, citrus, lemon, orange, and pear can have anti-inflammatory properties, including the capacity to regulate cytokine production, macrophage activity, and TLR expression (83, 85, 87, 88). Apple pectin treatment decreased the production of TNF-α and inflammation in the colon in a DMH/ DSS-induced colitis model (77). Native artichoke pectin and modified artichoke pectin reduced the expression of IL-6, TNF-α, iNOS, and ICAM and also increased the expression of TJ proteins MUC1 and occludin (84). Citrus and lemon pectins with low DM reduced the production of IL-1β, IL-6, and TNF-α, reduced neutrophil migration, improved epithelial barrier integrity, and increased expression of occludin in doxorubicin-induced ileitis (70, 82) and carerulein-induced pancreatitis (86). A compilation of the immunomodulatory effects of different pectins in animal models is shown in Table 2.
It is essential to acknowledge that many referenced in vivo studies (Table 2) were conducted on mice or rats a few weeks old, not neonates. While in vitro experiments (Table 1) provide some insights into cell responses, the question remains whether preterm and neonatal cells will exhibit similar responses as the established cell lines. Hence, there is a strong rationale for developing advanced models, like neonatal gut organoids cultured under anaerobic conditions, which can accurately replicate the interactions between pectin, neonatal gut cells, and microbiota to validate and extend these findings.
Pectin consumption stimulates the production of SCFA by the gut microbiota, which has beneficial effects on gut health (24, 25). SCFA, such as butyrate, stimulates mucin secretion in vitro through the upregulation of MUC3, MUC4, and MUC12 genes in the LS174T human colorectal cancer cell line (89). A low-fiber diet causes a shift in the gut microbiota to mucin degraders bacteria (e.g., A muciniphila, B. thetaiotaomicron) to fulfill their energy requirements (90). When pectins and other soluble fibers like inulin and β-glucan are fermented in the large intestine, they produce SCFAs due to their higher viscosity and solubility. This fermentation process stimulates the growth of healthy bacteria and reduces the growth of pathogenic bacteria (91). SCFAs, particularly acetate, propionate, and butyrate, serve as a primary energy source for colonocytes and play a crucial role in maintaining normal colonic function (92). They help in lowering intestinal pH, stimulating electrolyte and fluid absorption, increasing blood flow, and preventing pathogen overgrowth and intestinal inflammation (1, 91, 93, 94).
The production of SCFAs also contributes to reducing inflammation through different mechanisms, including the activation of G-protein-coupled receptors (GPRs), which inactivate the NF-κB pathway in immune and intestinal cells (91, 95). GPR41 and GPR43 are important for immune surveillance in the colon, stimulating the secretion of cytokines IL-1β and IL-18 (91). GPR43 is mainly expressed in innate immune cells, such as neutrophils and macrophages (96). GPR109, activated by butyrate, inhibits the pro-inflammatory NF-κB pathway (91, 96). Furthermore, SCFAs inhibit the production of pro-inflammatory cytokines IL-8, IL-12, IL-1, and TNF-α, and decrease NF-κB expression (95). The main three GPRs activated by SCFA are expressed in the enteroendocrine cells of the colonic epithelium, the polymorphonuclear immune cells, and smooth muscle cells (91).
Butyrate, in particular, modulates immune cells, such as macrophages, dendritic cells, and lymphocytes, inhibiting the production of cytokines IL-12p70 and IL-23 (91). It also regulates the proliferation of stem cells from the intestinal crypts (97). SCFAs are crucial for regulating intestinal inflammation by controlling the migration of immune cells to sites of injury and modulating their activation state (95). Additionally, they inhibit histone deacetylases (HDACs) in the IECs and immune cells (92). HDCA inhibition reduces the expression of NF-κB in immune cells and, as a result, decreases the production of pro-inflammatory cytokines (92). Studies have shown that SCFA can reduce the pro-inflammatory response by modulating TLR4 signaling pathway, reducing leukocyte infiltration, increasing the production of the anti-inflammatory cytokine IL-10, and reducing the pro-inflammatory cytokines IL-6, IL-12, and TNF-α (92).
It is crucial to emphasize that neonates, unlike full-term infants, do not consume pectins through a regular diet. Nonetheless, there are two potential pathways through which pectins could inhibit NEC development. Firstly, direct immunomodulatory effects of pectins have been demonstrated both in vitro (Table 1) and in vivo studies (Table 2). Secondly, pectins may indirectly affect NEC by modulating the gut microbiota. Considering these pathways, pectins, which are natural compounds found in vegetables, could be added as a supplement to breast milk to help reduce inflammation in neonates with NEC or even mitigate the risk of NEC in preterm neonates within neonatal intensive care units.
Challenges and outstanding questions.
1. What are the best sources of pectin? Natural or “modified” pectins?
2. What is the minimum amount of pectin to observe anti-inflammatory effects?
3. Are the pectin anti-inflammatory effects age-related?
JD: Funding acquisition, Writing – original draft, Visualization. JF: Supervision, Writing – review & editing. MS: Writing – review & editing, Funding acquisition. RS-G: Funding acquisition, Supervision, Writing – original draft.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported, in part, by Richard Schwartz Award for Excellence in Children’s Health Research, Department of Pediatrics, UMB (JD and RS-G), NIAID, NIH, grants R01-AI036525, U19-AI082655 [Cooperative Center for Human Immunology (CCHI)], and U19-AI142725 to MS, and FAPESP (JP), São Paulo Research Foundation (FAPESP) scholarships to JD (2019/018794-0 and 2021/07292-3). The content is solely the authors’ responsibility and does not necessarily represent the official views of the National Institute of Allergy and Infectious Diseases, the National Institutes of Health, the National Health Service, the National Institute for Health Research (NIHR).
We recognize the generous financial assistance provided by the Richard Schwartz Award for Excellence in Children’s Health Research, the São Paulo Research Foundation (FAPESP), and the National Institute of Allergy and Infectious Diseases at the National Institutes of Health (NIH).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Naqash, F , Masoodi, FA , Rather, SA , Wani, SM , and Gani, A . Emerging concepts in the nutraceutical and functional properties of pectin—a review. Carbohydr Polym. (2017) 168:227–39. doi: 10.1016/j.carbpol.2017.03.058
2. Cronin, P , Joyce, SA , O'Toole, PW , and O'Connor, EM . Dietary fibre modulates the gut microbiota. Nutrients. (2021) 13:1655. doi: 10.3390/nu13051655
3. Dupont, C , and Vandenplas, Y . Different thickening complexes with pectin in infant anti-regurgitation formula. Acta Paediatr. (2020) 109:471–80. doi: 10.1111/apa.15015
4. Vandenplas, Y , De Greef, E , Xinias, I , Vrani, O , Mavroudi, A , Hammoud, M, et al. Safety of a thickened extensive casein hydrolysate formula. Nutrition. (2016) 32:206–12. doi: 10.1016/j.nut.2015.08.008
5. Vandenplas, Y , De Greef, E , and Hauser, B . Safety and tolerance of a new extensively hydrolyzed rice protein-based formula in the management of infants with cow's milk protein allergy. Eur J Pediatr. (2014) 173:1209–16. doi: 10.1007/s00431-014-2308-4
6. Dupont, C , Kalach, N , Soulaines, P , Bradatan, E , Lachaux, A , Payot, F, et al. Safety of a new amino acid formula in infants allergic to Cow's Milk and intolerant to hydrolysates. J Pediatr Gastroenterol Nutr. (2015) 61:456–63. doi: 10.1097/MPG.0000000000000803
7. Dupont, C , Bradatan, E , Soulaines, P , Nocerino, R , and Berni-Canani, R . Tolerance and growth in children with cow's milk allergy fed a thickened extensively hydrolyzed casein-based formula. BMC Pediatr. (2016) 16:96. doi: 10.1186/s12887-016-0637-3
8. Rabbani, GH , Teka, T , Zaman, B , Majid, N , Khatun, M , and Fuchs, GJ . Clinical studies in persistent diarrhea: dietary management with green banana or pectin in Bangladeshi children. Gastroenterology. (2001) 121:554–60. doi: 10.1053/gast.2001.27178
9. Oppen, RL . Pectin and its therapeutic use: With special reference to the treatment of diarrhea in infants and children University of Nebraska Medical Center (1940).
10. Salomov, IT , Ashurov, AR , Oblakulov Kh, M , Salomov, UI , and Dzhalilov, BM . Effect of pectin additives from cotton on the physical development, nutrition and erythrocytes in infants. Vopr Pitan. (1994):16–8.
11. Binder, HJ . Role of colonic short-chain fatty acid transport in diarrhea. Annu Rev Physiol. (2010) 72:297–313. doi: 10.1146/annurev-physiol-021909-135817
12. Dhingra, D , Michael, M , Rajput, H , and Patil, RT . Dietary fibre in foods: a review. J Food Sci Technol. (2012) 49:255–66. doi: 10.1007/s13197-011-0365-5
13. Blanco-Pérez, F , Steigerwald, H , Schülke, S , Vieths, S , Toda, M , and Scheurer, S . The dietary Fiber pectin: health benefits and potential for the treatment of allergies by modulation of gut microbiota. Curr Allergy Asthma Rep. (2021) 21:43. doi: 10.1007/s11882-021-01020-z
14. Bode, L . Human milk oligosaccharides: every baby needs a sugar mama. Glycobiology. (2012) 22:1147–62. doi: 10.1093/glycob/cws074
15. Rastall, RA , Diez-Municio, M , Forssten, SD , Hamaker, B , Meynier, A , Moreno, FJ, et al. Structure and function of non-digestible carbohydrates in the gut microbiome. Benefic Microbes. (2022) 13:95–168. doi: 10.3920/BM2021.0090
16. Akkerman, R , Faas, MM , and de Vos, P . Non-digestible carbohydrates in infant formula as substitution for human milk oligosaccharide functions: effects on microbiota and gut maturation. Crit Rev Food Sci Nutr. (2019) 59:1486–97. doi: 10.1080/10408398.2017.1414030
17. Yi, DY , and Kim, SY . Human breast Milk composition and function in human health: from nutritional components to microbiome and MicroRNAs. Nutrients. (2021) 13:3094. doi: 10.3390/nu13093094
18. Coppa, GV , Pierani, P , Zampini, L , Carloni, I , Carlucci, A , and Gabrielli, O . Oligosaccharides in human milk during different phases of lactation. Acta Paediatr. (1999) 88:89–94. doi: 10.1111/j.1651-2227.1999.tb01307.x
19. Morris, VJ , Belshaw, NJ , Waldron, KW , and Maxwell, EG . The bioactivity of modified pectin fragments. Bioact Carbohydr Diet Fibre. (2013) 1:21–37. doi: 10.1016/j.bcdf.2013.02.001
20. Mohnen, D . Pectin structure and biosynthesis. Curr Opin Plant Biol. (2008) 11:266–77. doi: 10.1016/j.pbi.2008.03.006
21. O'Neill, MA , Ishii, T , Albersheim, P , and Darvill, AG . Rhamnogalacturonan II: structure and function of a borate cross-linked cell wall pectic polysaccharide. Annu Rev Plant Biol. (2004) 55:109–39. doi: 10.1146/annurev.arplant.55.031903.141750
22. Prado, S , Ferreira, GF , Harazono, Y , Shiga, TM , Raz, A , Carpita, NC, et al. Ripening-induced chemical modifications of papaya pectin inhibit cancer cell proliferation. Sci Rep. (2017) 7:16564. doi: 10.1038/s41598-017-16709-3
23. Eliaz, I , and Raz, A . Pleiotropic effects of modified citrus pectin. Nutrients. (2019) 11:1–18. doi: 10.3390/nu11112619
24. Beukema, M , Faas, MM , and de Vos, P . The effects of different dietary fiber pectin structures on the gastrointestinal immune barrier: impact via gut microbiota and direct effects on immune cells. Exp Mol Med. (2020) 52:1364–76. doi: 10.1038/s12276-020-0449-2
25. Hotchkiss, AT Jr, Renye, JA Jr, White, AK , Nunez, A , Guron, GKP , Chau, H, et al. Cranberry Arabino-xyloglucan and Pectic oligosaccharides induce Lactobacillus growth and short-chain fatty acid production. Microorganisms. (2022) 10:1346. doi: 10.3390/microorganisms10071346
26. Beukema, M , Akkerman, R , Jermendi, É , Koster, T , Laskewitz, A , Kong, C, et al. Pectins that structurally differ in the distribution of methyl-esters attenuate Citrobacter rodentium-induced colitis. Mol Nutr Food Res. (2021) 65:e2100346. doi: 10.1002/mnfr.202100346
27. Hunter, CJ , Upperman, JS , Ford, HR , and Camerini, V . Understanding the susceptibility of the premature infant to necrotizing enterocolitis (NEC). Pediatr Res. (2008) 63:117–23. doi: 10.1203/PDR.0b013e31815ed64c
28. Dominguez-Bello, MG , Costello, EK , Contreras, M , Magris, M , Hidalgo, G , Fierer, N, et al. Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc Natl Acad Sci U S A. (2010) 107:11971–5. doi: 10.1073/pnas.1002601107
29. Mueller, NT , Bakacs, E , Combellick, J , Grigoryan, Z , and Dominguez-Bello, MG . The infant microbiome development: mom matters. Trends Mol Med. (2015) 21:109–17. doi: 10.1016/j.molmed.2014.12.002
30. Secher, T , Brehin, C , and Oswald, E . Early settlers: which E. coli strains do you not want at birth? Am J Physiol Gastrointest Liver Physiol. (2016) 311:G123–9. doi: 10.1152/ajpgi.00091.2016
31. Falk, PG , Hooper, LV , Midtvedt, T , and Gordon, JI . Creating and maintaining the gastrointestinal ecosystem: what we know and need to know from gnotobiology. Microbiol Mol Biol Rev. (1998) 62:1157–70. doi: 10.1128/MMBR.62.4.1157-1170.1998
32. Mackie, RI , Sghir, A , and Gaskins, HR . Developmental microbial ecology of the neonatal gastrointestinal tract. Am J Clin Nutr. (1999) 69:1035s–45s. doi: 10.1093/ajcn/69.5.1035s
33. Patel, RM , and Denning, PW . Intestinal microbiota and its relationship with necrotizing enterocolitis. Pediatr Res. (2015) 78:232–8. doi: 10.1038/pr.2015.97
34. Fajardo, C , Alshaikh, B , and Harabor, A . Prolonged use of antibiotics after birth is associated with increased morbidity in preterm infants with negative cultures. J Matern Fetal Neonatal Med. (2019) 32:4060–6. doi: 10.1080/14767058.2018.1481042
35. Yu, Y , Lu, L , Sun, J , Petrof, EO , and Claud, EC . Preterm infant gut microbiota affects intestinal epithelial development in a humanized microbiome gnotobiotic mouse model. Am J Physiol Gastrointest Liver Physiol. (2016) 311:G521–32. doi: 10.1152/ajpgi.00022.2016
36. Belkaid, Y , and Hand, TW . Role of the microbiota in immunity and inflammation. Cells. (2014) 157:121–41. doi: 10.1016/j.cell.2014.03.011
37. Macpherson, AJ , Slack, E , Geuking, MB , and McCoy, KD . The mucosal firewalls against commensal intestinal microbes. Semin Immunopathol. (2009) 31:145–9. doi: 10.1007/s00281-009-0174-3
38. Mahdally, SM , Izquierdo, M , Viscardi, RM , Magder, LS , Crowley, HM , Bafford, AC, et al. Secretory-IgA binding to intestinal microbiota attenuates inflammatory reactions as the intestinal barrier of preterm infants matures. Clin Exp Immunol. (2023) 213:339–56. doi: 10.1093/cei/uxad042
39. Salerno-Goncalves, R , Safavie, F , Fasano, A , and Sztein, MB . Free and complexed-secretory immunoglobulin a triggers distinct intestinal epithelial cell responses. Clin Exp Immunol. (2016) 185:338–47. doi: 10.1111/cei.12801
40. Slack, E , Balmer, ML , Fritz, JH , and Hapfelmeier, S . Functional flexibility of intestinal IgA - broadening the fine line. Front Immunol. (2012) 3:100. doi: 10.3389/fimmu.2012.00100
41. Macpherson, AJ , Gatto, D , Sainsbury, E , Harriman, GR , Hengartner, H , and Zinkernagel, RM . A primitive T cell-independent mechanism of intestinal mucosal IgA responses to commensal bacteria. Science. (2000) 288:2222–6. doi: 10.1126/science.288.5474.2222
42. Shroff, KE , and Cebra, JJ . Development of mucosal humoral immune responses in germ-free (GF) mice. Adv Exp Med Biol. (1995) 371A:441–6.
43. Cebra, JJ , Periwal, SB , Lee, G , Lee, F , and Shroff, KE . Development and maintenance of the gut-associated lymphoid tissue (GALT): the roles of enteric bacteria and viruses. Dev Immunol. (1998) 6:13–8. doi: 10.1155/1998/68382
44. Lim, BO , Lee, SH , Park, DK , and Choue, RW . Effect of dietary pectin on the production of immunoglobulins and cytokines by mesenteric lymph node lymphocytes in mouse colitis induced with dextran sulfate sodium. Biosci Biotechnol Biochem. (2003) 67:1706–12. doi: 10.1271/bbb.67.1706
45. Popov, SV , Golovchenko, VV , Ovodova, RG , Smirnov, VV , Khramova, DS , Popova, GY, et al. Characterisation of the oral adjuvant effect of lemnan, a pectic polysaccharide of Lemna minor L. Vaccine. (2006) 24:5413–9. doi: 10.1016/j.vaccine.2006.03.076
46. Sori, N , Kunnummal, SP , Peddha, MS , and Khan, M . Prophylactic effect of pectic oligosaccharides against poly I: C- induced virus-like infection in BALB/c mice. J Food Biochem. (2022) 46:e14459. doi: 10.1111/jfbc.14459
47. Lim, BO , Yamada, K , Nonaka, M , Kuramoto, Y , Hung, P , and Sugano, M . Dietary fibers modulate indices of intestinal immune function in rats. J Nutr. (1997) 127:663–7. doi: 10.1093/jn/127.5.663
48. Cuna, A , Morowitz, MJ , Ahmed, I , Umar, S , and Sampath, V . Dynamics of the preterm gut microbiome in health and disease. Am J Physiol Gastrointest Liver Physiol. (2021) 320:G411–9. doi: 10.1152/ajpgi.00399.2020
49. Chi, C , Buys, N , Li, C , Sun, J , and Yin, C . Effects of prebiotics on sepsis, necrotizing enterocolitis, mortality, feeding intolerance, time to full enteral feeding, length of hospital stay, and stool frequency in preterm infants: a meta-analysis. Eur J Clin Nutr. (2019) 73:657–70. doi: 10.1038/s41430-018-0377-6
50. Neu, J , and Pammi, M . Pathogenesis of NEC: impact of an altered intestinal microbiome. Semin Perinatol. (2017) 41:29–35. doi: 10.1053/j.semperi.2016.09.015
51. Neu, J , and Walker, AW . Necrotizing enterocolitis. N Engl J Med. (2011) 364:255–64. doi: 10.1056/NEJMra1005408
52. Elshahed, MS , Miron, A , Aprotosoaie, AC , and Farag, MA . Pectin in diet: interactions with the human microbiome, role in gut homeostasis, and nutrient-drug interactions. Carbohydr Polym. (2021) 255:117388. doi: 10.1016/j.carbpol.2020.117388
53. Mao, G , Li, S , Orfila, C , Shen, X , Zhou, S , Linhardt, RJ, et al. Depolymerized RG-I-enriched pectin from citrus segment membranes modulates gut microbiota, increases SCFA production, and promotes the growth of Bifidobacterium spp., Lactobacillus spp. and Faecalibaculum spp. Food Funct. (2019) 10:7828–43. doi: 10.1039/C9FO01534E
54. Larsen, N , Bussolo de Souza, C , Krych, L , Barbosa Cahú, T , Wiese, M , Kot, W, et al. Potential of Pectins to beneficially modulate the gut microbiota depends on their structural properties. Front Microbiol. (2019) 10:223. doi: 10.3389/fmicb.2019.00223
55. Denning, TW , Bhatia, AM , Kane, AF , Patel, RM , and Denning, PW . Pathogenesis of NEC: role of the innate and adaptive immune response. Semin Perinatol. (2017) 41:15–28. doi: 10.1053/j.semperi.2016.09.014
56. Lu, P , Sodhi, CP , and Hackam, DJ . Toll-like receptor regulation of intestinal development and inflammation in the pathogenesis of necrotizing enterocolitis. Pathophysiology. (2014) 21:81–93. doi: 10.1016/j.pathophys.2013.11.007
57. Neu, J . Necrotizing enterocolitis: the future. Neonatology. (2020) 117:240–4. doi: 10.1159/000506866
58. Kaminsky, LW , Al-Sadi, R , and Ma, TY . IL-1β and the intestinal epithelial tight junction barrier. Front Immunol. (2021) 12:767456. doi: 10.3389/fimmu.2021.767456
59. Peterson, LW , and Artis, D . Intestinal epithelial cells: regulators of barrier function and immune homeostasis. Nat Rev Immunol. (2014) 14:141–53. doi: 10.1038/nri3608
60. Xie, J , Yu, R , Qi, J , Zhang, G , Peng, X , and Luo, J . Pectin and inulin stimulated the mucus formation at a similar level: An omics-based comparative analysis. J Food Sci. (2020) 85:1939–47. doi: 10.1111/1750-3841.15163
61. Mantis, NJ , and Forbes, SJ . Secretory IgA: arresting microbial pathogens at epithelial borders. Immunol Investig. (2010) 39:383–406. doi: 10.3109/08820131003622635
62. Mantis, NJ , Rol, N , and Corthesy, B . Secretory IgA's complex roles in immunity and mucosal homeostasis in the gut. Mucosal Immunol. (2011) 4:603–11. doi: 10.1038/mi.2011.41
63. Elphick, DA , and Mahida, YR . Paneth cells: their role in innate immunity and inflammatory disease. Gut. (2005) 54:1802–9. doi: 10.1136/gut.2005.068601
64. Hansson, GC . Role of mucus layers in gut infection and inflammation. Curr Opin Microbiol. (2012) 15:57–62. doi: 10.1016/j.mib.2011.11.002
65. McElroy, SJ , Prince, LS , Weitkamp, JH , Reese, J , Slaughter, JC , and Polk, DB . Tumor necrosis factor receptor 1-dependent depletion of mucus in immature small intestine: a potential role in neonatal necrotizing enterocolitis. Am J Physiol Gastrointest Liver Physiol. (2011) 301:G656–66. doi: 10.1152/ajpgi.00550.2010
66. Martin, NA , Mount Patrick, SK , Estrada, TE , Frisk, HA , Rogan, DT , Dvorak, B, et al. Active transport of bile acids decreases mucin 2 in neonatal ileum: implications for development of necrotizing enterocolitis. PLoS One. (2011) 6:e27191. doi: 10.1371/journal.pone.0027191
67. Jantscher-Krenn, E , Zherebtsov, M , Nissan, C , Goth, K , Guner, YS , Naidu, N, et al. The human milk oligosaccharide disialyllacto-N-tetraose prevents necrotising enterocolitis in neonatal rats. Gut. (2012) 61:1417–25. doi: 10.1136/gutjnl-2011-301404
68. Meldrum, OW , Yakubov, GE , Gartaula, G , McGuckin, MA , and Gidley, MJ . Mucoadhesive functionality of cell wall structures from fruits and grains: electrostatic and polymer network interactions mediated by soluble dietary polysaccharides. Sci Rep. (2017) 7:15794. doi: 10.1038/s41598-017-16090-1
69. Prado, SBR , Beukema, M , Jermendi, E , Schols, HA , de Vos, P , and Fabi, JP . Pectin interaction with immune receptors is modulated by ripening process in papayas. Sci Rep. (2020) 10:1690. doi: 10.1038/s41598-020-58311-0
70. Sahasrabudhe, NM , Beukema, M , Tian, L , Troost, B , Scholte, J , Bruininx, E, et al. Dietary Fiber pectin directly blocks toll-like receptor 2-1 and prevents doxorubicin-induced ileitis. Front Immunol. (2018) 9:383. doi: 10.3389/fimmu.2018.00383
71. Beukema, M , Jermendi, É , van den Berg, MA , Faas, MM , Schols, HA , and de Vos, P . The impact of the level and distribution of methyl-esters of pectins on TLR2-1 dependent anti-inflammatory responses. Carbohydr Polym. (2021) 251:117093. doi: 10.1016/j.carbpol.2020.117093
72. Hyun, GH , Cho, IH , Yang, YY , Jeong, DH , Kang, YP , Kim, YS, et al. Mechanisms of interactions in pattern-recognition of common glycostructures across pectin-derived heteropolysaccharides by toll-like receptor 4. Carbohydr Polym. (2023) 314:120921. doi: 10.1016/j.carbpol.2023.120921
73. Hunter, CJ , and De Plaen, IG . Inflammatory signaling in NEC: role of NF-κB, cytokines and other inflammatory mediators. Pathophysiology. (2014) 21:55–65. doi: 10.1016/j.pathophys.2013.11.010
74. El-Zayat, SR , Sibaii, H , and Mannaa, FA . Toll-like receptors activation, signaling, and targeting: an overview. Bull Natl Res Centre. (2019) 43:187. doi: 10.1186/s42269-019-0227-2
75. Ramachandran, C , Wilk, BJ , Hotchkiss, A , Chau, H , Eliaz, I , and Melnick, SJ . Activation of human T-helper/inducer cell, T-cytotoxic cell, B-cell, and natural killer (NK)-cells and induction of natural killer cell activity against K562 chronic myeloid leukemia cells with modified citrus pectin. BMC Complement Altern Med. (2011) 11:59. doi: 10.1186/1472-6882-11-59
76. Ishisono, K , Mano, T , Yabe, T , and Kitaguchi, K . Dietary Fiber pectin ameliorates experimental colitis in a neutral sugar side chain-dependent manner. Front Immunol. (2019) 10:2979. doi: 10.3389/fimmu.2019.02979
77. Liu, L , Li, YH , Niu, YB , Sun, Y , Guo, ZJ , Li, Q, et al. An apple oligogalactan prevents against inflammation and carcinogenesis by targeting LPS/TLR4/NF-κB pathway in a mouse model of colitis-associated colon cancer. Carcinogenesis. (2010) 31:1822–32. doi: 10.1093/carcin/bgq070
78. Amorim, JC , Vriesmann, LC , Petkowicz, CLO , Martinez, GR , and Noleto, GR . Modified pectin from Theobroma cacao induces potent pro-inflammatory activity in murine peritoneal macrophage. Int J Biol Macromol. (2016) 92:1040–8. doi: 10.1016/j.ijbiomac.2016.08.015
79. Chen, CH , Sheu, MT , Chen, TF , Wang, YC , Hou, WC , Liu, DZ, et al. Suppression of endotoxin-induced proinflammatory responses by citrus pectin through blocking LPS signaling pathways. Biochem Pharmacol. (2006) 72:1001–9. doi: 10.1016/j.bcp.2006.07.001
80. Salman, H , Bergman, M , Djaldetti, M , Orlin, J , and Bessler, H . Citrus pectin affects cytokine production by human peripheral blood mononuclear cells. Biomed Pharmacother. (2008) 62:579–82. doi: 10.1016/j.biopha.2008.07.058
81. Vogt, LM , Sahasrabudhe, NM , Ramasamy, U , Meyer, D , Pullens, G , Faas, MM, et al. The impact of lemon pectin characteristics on TLR activation and T84 intestinal epithelial cell barrier function. J Funct Foods. (2016) 22:398–407. doi: 10.1016/j.jff.2016.02.002
82. Beukema, M , Jermendi, É , Koster, T , Kitaguchi, K , de Haan, BJ , van den Berg, MA, et al. Attenuation of doxorubicin-induced small intestinal mucositis by Pectins is dependent on Pectin's methyl-Ester number and distribution. Mol Nutr Food Res. (2021) 65:e2100222. doi: 10.1002/mnfr.202100222
83. Ye, MB , and Lim, BO . Dietary pectin regulates the levels of inflammatory cytokines and immunoglobulins in interleukin-10 knockout mice. J Agric Food Chem. (2010) 58:11281–6. doi: 10.1021/jf103262s
84. Sabater, C , Molina-Tijeras, JA , Vezza, T , Corzo, N , Montilla, A , and Utrilla, P . Intestinal anti-inflammatory effects of artichoke pectin and modified pectin fractions in the dextran sulfate sodium model of mice colitis. Artificial neural network modelling of inflammatory markers. Food Funct. (2019) 10:7793–805. doi: 10.1039/c9fo02221j
85. Merheb, R , Abdel-Massih, RM , and Karam, MC . Immunomodulatory effect of natural and modified Citrus pectin on cytokine levels in the spleen of BALB/c mice. Int J Biol Macromol. (2019) 121:1–5. doi: 10.1016/j.ijbiomac.2018.09.189
86. Sun, Y , He, Y , Wang, F , Zhang, H , de Vos, P , and Sun, J . Low-methoxyl lemon pectin attenuates inflammatory responses and improves intestinal barrier integrity in caerulein-induced experimental acute pancreatitis. Mol Nutr Food Res. (2017) 61:1600885. doi: 10.1002/mnfr.201600885
87. Xu, GR , Zhang, C , Yang, HX , Sun, JH , Zhang, Y , Yao, TT, et al. Modified citrus pectin ameliorates myocardial fibrosis and inflammation via suppressing galectin-3 and TLR4/MyD88/NF-κB signaling pathway. Biomed Pharmacother. (2020) 126:110071. doi: 10.1016/j.biopha.2020.110071
88. Lee, JC , Pak, SC , Lee, SH , Na, CS , Lim, SC , Song, CH, et al. Asian pear pectin administration during presensitization inhibits allergic response to ovalbumin in BALB/c mice. J Altern Complement Med. (2004) 10:527–34. doi: 10.1089/1075553041323867
89. Jung, TH , Park, JH , Jeon, WM , and Han, KS . Butyrate modulates bacterial adherence on LS174T human colorectal cells by stimulating mucin secretion and MAPK signaling pathway. Nutr Res Pract. (2015) 9:343–9. doi: 10.4162/nrp.2015.9.4.343
90. Daniel, N , Lécuyer, E , and Chassaing, B . Host/microbiota interactions in health and diseases-time for mucosal microbiology! Mucosal Immunol. (2021) 14:1006–16. doi: 10.1038/s41385-021-00383-w
91. Gasaly, N , de Vos, P , and Hermoso, MA . Impact of bacterial metabolites on gut barrier function and host immunity: a focus on bacterial metabolism and its relevance for intestinal inflammation. Front Immunol. (2021) 12:658354. doi: 10.3389/fimmu.2021.658354
92. van der Beek, CM , Dejong, CHC , Troost, FJ , Masclee, AAM , and Lenaerts, K . Role of short-chain fatty acids in colonic inflammation, carcinogenesis, and mucosal protection and healing. Nutr Rev. (2017) 75:286–305. doi: 10.1093/nutrit/nuw067
93. Aune, D , Chan, DS , Lau, R , Vieira, R , Greenwood, DC , Kampman, E, et al. Dietary fibre, whole grains, and risk of colorectal cancer: systematic review and dose-response meta-analysis of prospective studies. BMJ. (2011) 343:d6617. doi: 10.1136/bmj.d6617
94. Fung, KY , Ooi, CC , Zucker, MH , Lockett, T , Williams, DB , Cosgrove, LJ, et al. Colorectal carcinogenesis: a cellular response to sustained risk environment. Int J Mol Sci. (2013) 14:13525–41. doi: 10.3390/ijms140713525
95. Iddir, M , Brito, A , Dingeo, G , Fernandez Del Campo, SS , Samouda, H , La Frano, MR, et al. Strengthening the immune system and reducing inflammation and oxidative stress through diet and nutrition: considerations during the COVID-19 crisis. Nutrients. (2020) 12:1562. doi: 10.3390/nu12061562
96. Dalile, B , Van Oudenhove, L , Vervliet, B , and Verbeke, K . The role of short-chain fatty acids in microbiota-gut-brain communication. Nat Rev Gastroenterol Hepatol. (2019) 16:461–78. doi: 10.1038/s41575-019-0157-3
97. Schulthess, J , Pandey, S , Capitani, M , Rue-Albrecht, KC , Arnold, I , Franchini, F, et al. The short chain fatty acid butyrate imprints an antimicrobial program in macrophages. Immunity. (2019) 50:432–445.e7. doi: 10.1016/j.immuni.2018.12.018

Keywords: pectin, inflammation, intestine, newborn, preterm
Citation: Donadio JLS, Fabi JP, Sztein MB and Salerno-Gonçalves R (2024) Dietary fiber pectin: challenges and potential anti-inflammatory benefits for preterms and newborns. Front. Nutr. 10:1286138. doi: 10.3389/fnut.2023.1286138
Edited by:
Bin Du, Hebei Normal University of Science and Technology, ChinaReviewed by:
Alma Cruz-Guerrero, Metropolitan Autonomous University, MexicoCopyright © 2024 Donadio, Fabi, Sztein and Salerno-Gonçalves. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rosângela Salerno-Gonçalves, cm1lemdoYW5Ac29tLnVtYXJ5bGFuZC5lZHU=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.