 Abdelhafid Nani
Abdelhafid Nani Wafâa Tehami
Wafâa Tehami- Laboratory of Saharan Natural Resources, University of Ahmed Draia, Adrar, Algeria
Obesity, pancreatitis, cardiovascular, gastrointestinal (GI), and liver diseases have all been linked to the Western lifestyle, characterized by increased unhealthy food consumption and decreased physical activity. Besides obesity and pancreatitis, many GI and liver diseases are associated with inflammation. Inflammasomes are multi-protein complexes that mediate acute and restorative inflammatory pathways. However, many aberrations in inflammasome activity originate from shifts in dietary habits. Evidence reveals that dietary polyphenols effectively modulate inflammasome-associated dysfunctions. With a focus on pancreatitis, GI, and liver disorders, this review set out to provide the most relevant evidence for the therapeutic impact of polyphenols via the regulation of the inflammasome pathway. Overall, flavonoid and non-flavonoid polyphenols maintain intestinal eubiosis, downregulate NLRP3 inflammasome canonical pathway, and restore redox status via upregulating Nrf2/HO-1 signaling. These effects at the level of the intestine, the liver, and the pancreas are associated with decreased systemic levels of key pro-inflammatory cytokines, including TNF-α, IL-1β, and IL-6.
1. Introduction
Western (or more broadly “modern”) lifestyle, which is characterized by increased unhealthy food consumption and decreased physical activity, is linked to chronic diseases of civilization, including epithelial cell cancers, obesity, pancreatitis, cardiovascular, gastrointestinal (GI), and liver diseases (1–4). GI diseases encompass one or more of the four typical symptoms and signs: abdominal or chest pain, altered food ingestion, altered bowel movements, and GI tract bleeding (5). Many GI and liver diseases are associated with inflammation, which is a physiological response that can be triggered by invading microbes’ antigens or host inflammatory molecules (6, 7). Inflammasomes are multi-protein complexes that mediate acute and reparative inflammatory pathways (8). However, deregulated inflammasome activities can result in chronic inflammation underlying a series of pathologies, such as GI and liver disorders and pancreatitis (9–13). In addition to the gut microbiota, dietary habit changes are believed to underlie many host inflammatory responses (8, 14–16). Indeed, the release of free fatty acids (FFAs) in the human GI tract may trigger NLRP3 inflammasome-mediated inflammation (16–18). Therefore, a healthy diet rich in nutraceuticals can be an excellent strategy for managing GI and liver disorders, partly through regulating inflammasome activities (19, 20). Polyphenols are dietary nutraceuticals that have been reported to exhibit a number of biological activities, such as antioxidant, antimicrobial, anti-inflammatory, and anticancer properties (18). These bioactive substances are found in many cereals, vegetables, fruits, herbs, and beverages. Accumulating evidence reveals that polyphenols are effective in modulating inflammasome-associated dysfunctions, including pancreatitis, GI and liver disorders (21–24). Pellegrini et al. (25) recently provided an overview of NLRP3 inflammasome pathway modulation by secondary metabolites. Another recent review by Owona et al. (26) highlighted the role of flavonoids, the most abundant phenolic compounds, in modulating numerous inflammasome-associated chronic diseases. Therefore, studies have been expanding on polyphenols-induced inflammasome regulation as a prominent approach for managing and treating inflammation-associated diseases. This review aimed to provide up-to-date evidence for the therapeutic impact of polyphenols via the regulation of the NLRP3 inflammasome pathway, with a focus on pancreatitis, gastrointestinal and liver disorders.
2. Inflammasome activation by Western dietary patterns
Inflammasomes are intracellular multi-protein complexes made up of three components: a sensor molecule consisting of nucleotide-binding oligomerization domain (NOD)-like receptors (NLRs), an adaptor protein ASC (apoptotic speck-like protein containing), and an effector molecule procaspase-1 (12). NLR family pyrin domain-containing protein (NLRP) inflammasomes are innate immune sensors that are assembled following NLR’s recognition of multiple classes of damage-associated molecular patterns (DAMPs), in response to cell injury, and pathogen-associated molecular patterns (PAMPs) of invading microbes (8). Once assembled, NLRPs mediate the caspase-1 activation that results in proteolytic cleavage of pro-interleukin-1β (pro-IL-1β) and IL-18 into bioactive forms leading to a myriad of additional cytokines and chemokines upregulation or initiating pyroptotic cell death (pyroptosis) (27).
Approximately half of all deaths worldwide in the late 19th century was due to infectious diseases. This burden has dropped significantly in the last century, from 85 to 15%, thanks to significant developments such as widespread sanitation improvement. Contrarily, shifts in dietary habits, especially in Western societies, have contributed significantly to the emergence of non-communicable diseases. Furthermore, a Western-style calorie-rich diet is directly or indirectly responsible for over 80% of all deaths (28). Western diet (WD) patterns include a lot of high-glycemic/high-insulinemic carbohydrate foods like refined cereals, corn, potatoes, sugars (mainly sucrose and fructose), dairy products, and a considerable amount of fat (a fair amount of ω-6 polyunsaturated fatty acids “PUFAs”) and plenty of protein (29). Adherence to the Western dietary pattern enhances refined starchy carbohydrate amounts and free fatty acids (FFAs) release. All of which can result in sterile (or non-pathogenic) inflammation associated with alterations in gut microbiota (17, 22, 30–32). Indeed, a sedentary lifestyle and excessive intake of fat, starchy carbohydrates, and free sugars have been suggested to activate inflammasomes through uric acid, resulting in exacerbated oxidative stress and inflammation in the liver (31). It has also been shown that FFAs induce the expression of NLRP3 inflammasome complex-forming proteins in endothelial cells (17). Increasing evidence has established the association of WD with gut microbiota profile alterations and gut mucosal barrier disruption (32, 33). The gut mucosa disruption is a hallmark of inflammatory bowel diseases (IBD) characterized by an upregulated NLRP3 inflammasome pathway, which can be triggered following short chain fatty acids (SCFAs) binding to GPR43 of the enterocytes (19, 22, 32, 34). Moreover, a recent study showed that WD is associated with altered gene expression, resulting in an elevated risk for autoimmune pancreatitis (30). Since the liver is a key metabolic organ, WD patterns, including simple sugars, saturated fatty acids, trans fats, and animal proteins, play a crucial role in the onset and progression of liver pathologies such as steatosis and nonalcoholic fatty liver disease (NAFLD) (35). Indeed, high levels of alcohol absorption can cause hepatocyte death. Likewise, hepatocyte steatosis can be caused by a high energy intake of fat and sugar (31). All these factors can alter the gut microbiota composition, resulting in increased microbial translocation to the portal blood and increased PAMPs exposure in the liver. These PAMPs from the gut activate liver immune cells via PRRs, leading to increased IL-1β and IL-18 production through toll like receptor 4/nuclear factor-kappa B /NLRP3 (TLR4/NF-κB/NLRP3) inflammasome signaling pathway (36).
The main mechanisms underlying NLRP3 inflammasome complex-mediated inflammatory disorders by WD are overviewed in Figure 1.
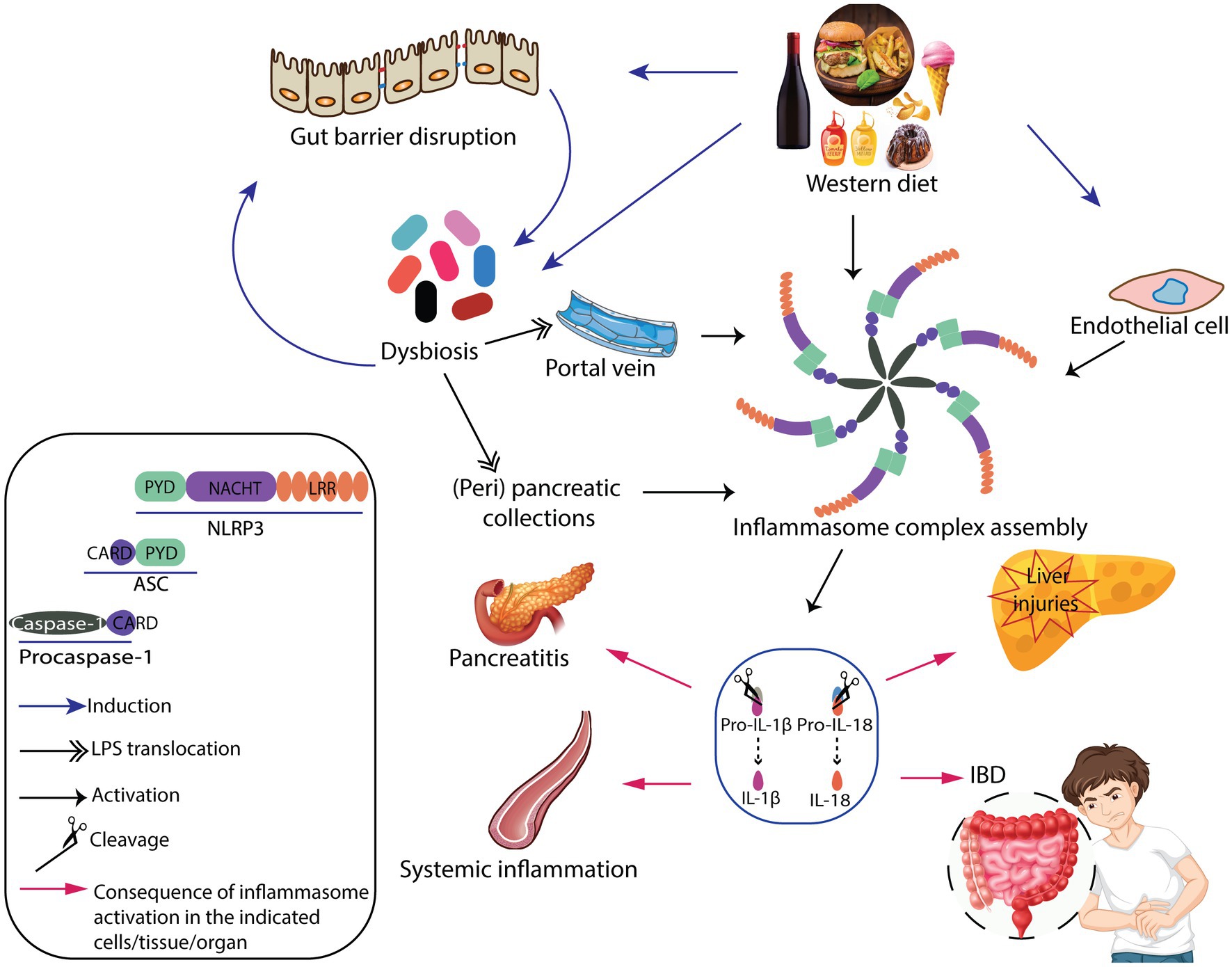
Figure 1. Graphical summary of the mechanisms underlying inflammasome complex-mediated inflammatory disorders by Western diet.
3. Polyphenols impact on inflammasome-mediated diseases
The term polyphenols refers to a series of homologous compounds comprising a hydroxyl group bonded directly to a benzene ring. Thus, polyphenols are substances broadly distributed in the plant kingdom that contain multiple phenyl rings and at least one hydroxyl substituent (37). Their structure may vary from basic compounds like phenolic acids and stilbenes to more complex polymers with a high molecular weight, like tannins (38). Dietary polyphenols are frequently classified into flavonoid and non-flavonoid polyphenols. Plant-based polyphenols are potent anti-inflammatory agents, and this benefit may be mediated by their ability to modulate inflammasome activity (39). Indeed, in silico investigations showed that phenolics such as phenylpropanoids, curcumin, and epigallocatechin-3-gallate (EGCG) exhibited a high affinity with the NLRP3 inflammasome complex (40, 41). Thus, phenolic compounds may be potent inhibitors of NLRP3 inflammasome activation. Therefore, they can modulate NLRP3 inflammasome-associated inflammation and pathologies.
3.1. Pancreatitis
Acute pancreatitis (AP) is the most prevalent pancreatic pathology and the most likely cause of hospitalization among nonmalignant gastrointestinal illnesses (13). Evidence showed that AP may be linked to luminal lipids maldigestion, and the premature activation of the proteolytic proenzymes such as trypsinogen within the pancreatic acinar cells is expected in this pathology (42). It is demonstrated that the NLRP3 inflammasome is incriminated in pancreatitis onset and complications (13). Thus, inhibiting the NLRP3 inflammasome pathway might be an effective treatment for patients with severe pancreatitis. Phytochemicals have been reported to modulate inflammasome-mediated diseases (25, 26, 43). Accordingly, several studies investigated the impact of dietary polyphenols on inflammasome-mediated pancreatitis (21, 44–47). Aruna et al. (44) suggested that rutin, a flavonoid glycoside, curtails pancreatitis through the downregulation of ASC–NLRP3, resulting in reduced caspase-1 activation and decreased IL-1β, IL-18, and tumor necrosis factor-α (TNF-α) pro-inflammatory cytokines expression and production in alcohol and cerulein-induced pancreatitis model. It is well-established that systemic inflammation is common in severe AP (13). Interestingly, EGCG and other phenolic compounds have been shown to decrease systemic levels of key pro-inflammatory cytokines, including TNF-α, IL-1β, and IL-6 (21, 45, 47).
Reactive oxygen species (ROS) are the root cause of cellular oxidative stress, significantly contributing to AP pathogenesis (48). It is well known that endogenous and exogenous antioxidants counteract cellular ROS overproduction. Several phenolic compounds exhibited a preventive effect against AP severity and complications by mitigating mitochondrial ROS-triggered NLRP3 inflammasome activation or upregulating the nuclear factor erythroid 2-related factor 2 (Nrf-2) pathway (21, 44, 45, 49). Indeed, rutin reduced plasma levels of thiobarbituric acid reactive substances (TBARS), lipid peroxidation end by-products, and enhanced glutathione peroxidase (GPx), Superoxide dismutase (SOD), Catalase (CAT) activity in pancreatic tissue of alcohol and cerulein-induced pancreatitis model (44). Moreover, apocynin, a phenolic compound, decreased the ROS level in the lung of rats suffering from severe AP (45). Another study revealed that EGCG co-administration enhanced SOD activation and glutathione (GSH) level concomitant with decreased malondialdehyde (MDA) levels in the lung tissue of Balb/C mice suffering from AP induced with L-arginine (21).
3.2. Gastrointestinal pathologies
The gastrointestinal tract contains many immunocompetent cells and is the largest compartment for food digestion and nutrient absorption. The body’s most significant mucosal layer, the intestinal mucosa, is crucial to maintaining intestinal homeostasis. Nevertheless, nutrition, drugs, microbial stimuli, and exacerbated production of many metabolites normally produced in cells are risk factors for intestinal inflammation that underlies numerous intestinal disorders. Indeed, long-term exposure to inflammatory cytokines such as IL-1, IL-6, IL-8, TNF-α and interferon-gamma (IFN-γ) is linked to several illnesses, including IBD which include ulcerative colitis (UC) and Crohn’s disease (CD). These cytokines disrupt the homeostasis of the digestive system during an imbalanced inflammatory condition, which results in a chronic inflammatory process (50–52). Recent research has shown how crucial canonical NLRP3 inflammasome signaling is for controlling intestinal homeostasis and the risk for IBD (53).
Phenolic compounds have attracted the attention of several researchers due to their pharmacological actions as an antioxidant, an anti-inflammatory, and an antibacterial (18). In vivo studies showed that bergenin administration to rats with 2,4,6-trinitrobenzenesulfonic acid (TNBS)-induced acute colitis significantly downregulated IFN-γ, NLRP3 inflammasome complex-forming proteins, IL-1β, and IL-18 in colonic tissue. Thereby blocking the canonical pathway of the NLRP3 inflammasome (53).
Since the pro-inflammatory macrophage phenotype plays a central role in intestinal mucosal barrier damage, there has been much interest in counteracting macrophage activation by dietary polyphenols to maintain intestinal barrier integrity (54, 55). A recent study showed that ligstroside aglycon (LA), an abundant phenolic compound in extra virgin olive oil (EVOO), inhibited canonical and non-canonical activation of NLRP3 inflammasome and modulated cyclo-oxygenase-2 (COX-2) in lipopolysaccharide (LPS)-stimulated murine peritoneal macrophages. This anti-inflammatory effect was associated with a significant antioxidant activity revealed by a decrease in NO production, inducible nitric oxide synthase (iNOS), and NADPH oxidase-1 (NOX-1) protein expression (56). Besides, rosmarinic acid nanovesicles protected the colonic mucosa from dextran sodium sulfate (DSS)-induced injury by regulating the NLRP3 inflammasome and restoring Nrf2/ heme oxygenase (HO-1) signaling pathway (52). It has also been demonstrated that hydroxytyrosol supplementation exhibits anti-inflammatory effects in murine ulcerative colitis models by promoting eubiosis. Moreover, hydroxytyrosol enhanced the colonic antioxidant capacity concomitant with NLRP3 inflammasome inhibition, revealed by caspase-1, ASC, IL-18, and IL-1β downregulation (22).
3.3. Liver disorders
Clinically, IBD and NAFLD frequently coexist (57). Liver diseases account for more than 45% of fatalities in developed countries and significantly contribute to worldwide morbidity and mortality since the liver is the main metabolically active and detoxification organ (58, 59). Numerous conditions, such as drug abuse, alcoholism, viral infections, metabolic abnormalities, and autoimmune reactions, can result in liver damage. Additionally, liver pathophysiological processes are frequently linked to inflammation and oxidative stress, suggesting a dual control of one another (58). Some hepatic pro-inflammatory cytokines, such as IL-1β, are rapidly produced after liver damage and directly activate hepatic stellate cells (HSCs), causing the conversion of these cells into myofibroblasts. The last secret a large amount of extracellular matrix (ECM) that leads to scar tissue formation. Many studies have associated increased pyroptosis with various illnesses, including liver abnormalities (58). Activation of NLRP3 inflammasome has emerged as the primary trigger of liver fibrosis and cirrhosis (58, 60). Moreover, caspase-1 and ASC are involved in the inflammatory and oxidative stress responses (61). The transcription factor Nrf2 orchestrates the cellular response to redox stress. Once activated, the cytosolic Nrf2 translocates to the nucleus, initiating gene expression of several antioxidant enzymes (62). Therefore, Nrf2 has become a main target for managing oxidative stress-associated disorders. Indeed, in addition to stimulating liver regeneration, Nrf2 has been reported to play intricate roles in the modulation of liver fibrosis, cancer, and inflammation (59). Likewise, NLRP3 upregulates hepatic Kelch-like ECH-related protein 1 (Keap-1), Nrf2 negative regulator, which may cause fibrogenesis as a result of ROS-induced pyroptosis (58). Hence, antioxidant therapy effectively counters ROS-mediated activation of the NLRP3 inflammasome in murine acute liver injury models (61). Another in vivo study demonstrated that chlorogenic acid, a hydroxycinnamic acid, upregulates the expression and the activation of Nrf2-related antioxidant genes, including HO-1, NAD(P)H:quinone oxidoreductase-1 (NQO1), and glutamate-cysteine ligase catalytic subunit (GCLC). Furthermore, this phenolic acid inhibited NLRP3 inflammasome activation revealed by caspase-1 and IL-1β proteins downregulation in Sprague–Dawley rats with carbon tetrachloride (CCl4)-induced acute liver injury (59).
Without excessive alcohol consumption, NAFLD is the most common cause of liver dysfunction in the Western world (63). NAFLD is not a single disorder. Instead, it refers to a variety of hepatic pathologies, from fatty liver (steatosis) to fatty liver with a pronounced inflammation and fibrosis (nonalcoholic steatohepatitis, or NASH) to cirrhosis and possibly hepatocellular cancer (64, 65). The NLRP3 inflammasome plays a crucial role in the development of NASH. Recent findings suggested that cannabidiol, a terpenophenolic compound, may diminish the risk for NASH by downregulating NF-κB and NLRP3 inflammasome signaling pathways in macrophages in high-fat high cholesterol (HFC) diet-fed mice (66).
Although the significant decrease in pathogen-caused foodborne diseases in developed countries, increased exposure to aflatoxin B1 (AFB1), produced by toxigenic Aspergillus fungi, has recently been found in some regions of the United States (67). Prolonged exposure to AFB1 has been suggested to induce hepatocyte pyroptosis and oxidative stress, which further results in liver injury (23). Importantly, curcumin mitigated AFB1-induced TNF-α, IL-6, and IL-1β pro-inflammatory cytokines production and ROS generation, resulting in curtailed necroptosis of chicken liver tissue (68). In line, another study showed that EGCG, a major flavonoid in green tea, inhibited NLRP3 inflammasome activation, which was associated with improved hepatic oxidative stress, cell apoptosis, necrosis, steatosis, and degeneration in CD-1 (ICR) mice (69). Emerging evidence from in vivo models highlights that the protective effects of curcumin against liver injuries and diseases are mainly exerted through Nrf-2 upregulation concomitant with NLRP3 and NF-κB pathways downregulation (20, 23, 70, 71). Data from a case–control study and a randomized controlled trial showed that anthocyanins, a flavonoid subclass, abrogate NLRP3 inflammasome, caspase-1, IL-1β, and IL-18 expression in subjects’ peripheral blood mononuclear cells (PBMCs) and plasma levels of IL-1β and IL-18 in patients with NAFLD (72). It has also been shown that cyanidin-3-O-β-glucoside, an anthocyanin, can attenuate alcoholic steatohepatitis through inducing NF-κB deacetylation and counteracting NLRP3 inflammasome activation (73).
The effects of various polyphenol classes on the studied inflammasome-related pathologies are summarized in Table 1.
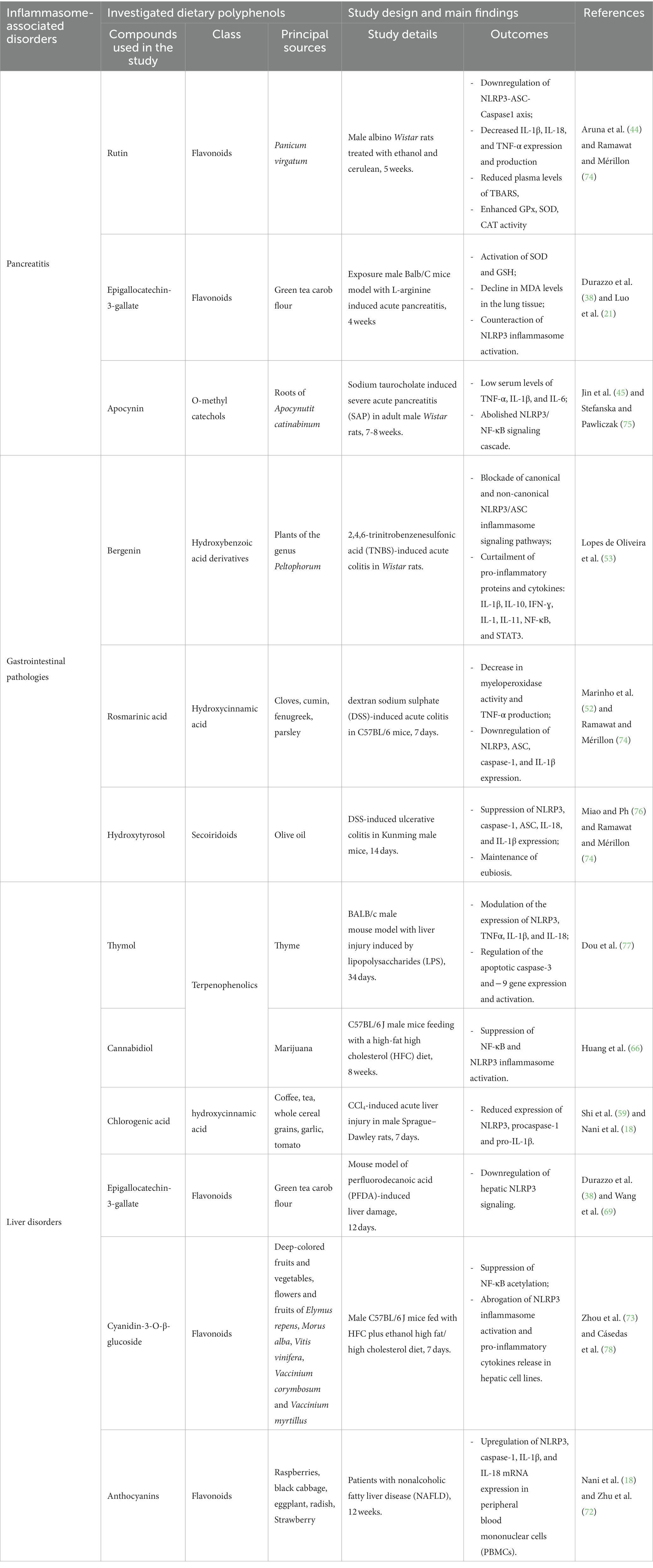
Table 1. A summary of phenolic compounds’ regulation of the inflammasome pathway in pancreatitis, gastrointestinal, and liver disorders.
4. Conclusion
Western lifestyle characterized by nutritional transition is a major risk factor for inflammasome-associated inflammation and illnesses. Nutritional approaches encourage polyphenol-rich foods as a prominent strategy for NLRP3 inflammasome-related disorders prevention and management. The current review sheds light on how dietary polyphenols can regulate NLRP3 inflammasome activation in pancreatitis, gastrointestinal and liver diseases. Overall, polyphenols counteract inflammasome complex assembly, downregulating its downstream substrates, IL-1β and IL-18. Moreover, polyphenols may modulate oxidative stress associated with inflammation in these NLRP3 inflammasome-linked pathologies via upregulating Nrf-2 and abolishing NF-κB pathway.
Author contributions
AN conceptualized the topic, researched and analyzed the literature and wrote the manuscript, and revised the manuscript critically. WT researched and analyzed the literature, wrote the manuscript, and constructed the figures. All authors contributed to the article and approved the submitted version.
Funding
The author(s) received no financial support for this article’s research, authorship, and/or publication.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnut.2023.1157572/full#supplementary-material
References
1. Carrera-Bastos, P, Fontes, O'K, Lindeberg, S, and Cordain, L. The western diet and lifestyle and diseases of civilization. Res Reports Clin Cardiol. (2011) 2:15–35. doi: 10.2147/RRCC.S16919
2. Yadav, D, and Lowenfels, AB. The epidemiology of pancreatitis and pancreatic cancer. Gastroenterology. (2013) 144:1252–61. doi: 10.1053/j.gastro.2013.01.068
3. Mikolasevic, I, Milic, S, Orlic, L, Poropat, G, Jakopcic, I, Franjic, N, et al. Metabolic syndrome and acute pancreatitis. Eur J Intern Med. (2016) 32:79–83. doi: 10.1016/j.ejim.2016.04.004
4. Moszak, M, Szulińska, M, and Bogdański, P. You are what you eat—the relationship between diet, microbiota, and metabolic disorders—a review. Nutrients. (2020) 12:1096. doi: 10.3390/nu12041096
5. Ciorba, MA . “Gastrointestinal Disease.,” Pathophysiology of Disease: An Introduction to Clinical Medicine 8E. McGraw Hill (2018) 369.
6. Feldman, M, Friedman, LS, and Brandt, LJ. Sleisenger and Fordtran’s gastrointestinal and liver disease: pathophysiology, diagnosis, management. USA: Elsevier Health Sciences (2020).
7. Hernandez, J-C, Sirois, CM, and Latz, E. Activation and regulation of the NLRP3 inflammasome In: The inflammasomes : Springer (2011). 197–208.
8. Kieser, KJ, and Kagan, JC. Multi-receptor detection of individual bacterial products by the innate immune system. Nat Rev Immunol. (2017) 17:376–90. doi: 10.1038/nri.2017.25
9. Man, SM . Inflammasomes in the gastrointestinal tract: infection, cancer and gut microbiota homeostasis. Nat Rev Gastroenterol Hepatol. (2018) 15:721–37. doi: 10.1038/s41575-018-0054-1
10. Monteiro, S, Grandt, J, Uschner, FE, Kimer, N, Madsen, JL, Schierwagen, R, et al. Differential inflammasome activation predisposes to acute-on-chronic liver failure in human and experimental cirrhosis with and without previous decompensation. Gut. (2021) 70:379–87. doi: 10.1136/gutjnl-2019-320170
11. Ferrero-Andrés, A, Panisello-Roselló, A, Roselló-Catafau, J, and Folch-Puy, E. NLRP3 inflammasome-mediated inflammation in acute pancreatitis. Int J Mol Sci. (2020) 21:5386. doi: 10.3390/ijms21155386
12. Luan, J, and Ju, D. Inflammasome: a double-edged sword in liver diseases. Front Immunol. (2018) 9:2201. doi: 10.3389/fimmu.2018.02201
13. Sendler, M, van den Brandt, C, Glaubitz, J, Wilden, A, Golchert, J, Weiss, FU, et al. NLRP3 inflammasome regulates development of systemic inflammatory response and compensatory anti-inflammatory response syndromes in mice with acute pancreatitis. Gastroenterology. (2020) 158:253–69. doi: 10.1053/j.gastro.2019.09.040
14. Hansen, NW, and Sams, A. The microbiotic highway to health—new perspective on food structure, gut microbiota, and host inflammation. Nutrients. (2018) 10. doi: 10.3390/nu10111590
15. Gill, V, Kumar, V, Singh, K, Kumar, A, and Kim, J-J. Advanced glycation end products (AGEs) may be a striking link between modern diet and health. Biomol Ther. (2019) 9:888. doi: 10.3390/biom9120888
16. Nie, C, Li, Y, Qian, H, Ying, H, and Wang, L. Advanced glycation end products in food and their effects on intestinal tract. Crit Rev Food Sci Nutr. (2022) 62:3103–15. doi: 10.1080/10408398.2020.1863904
17. Qi, Y, Du, X, Yao, X, and Zhao, Y. Vildagliptin inhibits high free fatty acid (FFA)-induced NLRP3 inflammasome activation in endothelial cells. Artif Cells Nanomed Biotechnol. (2019) 47:1067–74. doi: 10.1080/21691401.2019.1578783
18. Nani, A, Murtaza, B, Sayed Khan, A, Khan, NA, and Hichami, A. Antioxidant and anti-inflammatory potential of polyphenols contained in Mediterranean diet in obesity: molecular mechanisms. Molecules. (2021) 26:985. doi: 10.3390/molecules26040985
19. Macia, L, Tan, J, Vieira, AT, Leach, K, Stanley, D, Luong, S, et al. Metabolite-sensing receptors GPR43 and GPR109A facilitate dietary fibre-induced gut homeostasis through regulation of the inflammasome. Nat Commun. (2015) 6:1–15. doi: 10.1038/ncomms7734
20. Wang, D, Zhang, M, Wang, T, Cai, M, Qian, F, Sun, Y, et al. Green tea polyphenols prevent lipopolysaccharide-induced inflammatory liver injury in mice by inhibiting NLRP3 inflammasome activation. Food Funct. (2019) 10:3898–908. doi: 10.1039/C9FO00572B
21. Luo, Z-L, Sun, H-Y, Wu, X-B, Cheng, L, and Ren, J-D. Epigallocatechin-3-gallate attenuates acute pancreatitis induced lung injury by targeting mitochondrial reactive oxygen species triggered NLRP3 inflammasome activation. Food Funct. (2021) 12:5658–67. doi: 10.1039/D1FO01154E
22. Miao, F . Hydroxytyrosol alleviates dextran sodium sulfate–induced colitis by inhibiting NLRP3 inflammasome activation and modulating gut microbiota in vivo. Nutrition. (2022) 97:111579. doi: 10.1016/j.nut.2021.111579
23. Wang, Y, Liu, F, Liu, M, Zhou, X, Wang, M, Cao, K, et al. Curcumin mitigates aflatoxin B1-induced liver injury via regulating the NLRP3 inflammasome and Nrf2 signaling pathway. Food Chem Toxicol. (2022) 161:112823. doi: 10.1016/j.fct.2022.112823
24. Rodrigues, L, Donado-Pestana, CM, Moura, MHC, e Silva, RR, Pessoa, EVM, and Genovese, MI. Phenolic compounds from Jaboticaba (Plinia Jaboticaba (vell.) berg) ameliorate intestinal inflammation and associated endotoxemia in obesity. Food Res Int. (2021) 141:110139. doi: 10.1016/j.foodres.2021.110139
25. Pellegrini, C, Fornai, M, Antonioli, L, Blandizzi, C, and Calderone, V. Phytochemicals as novel therapeutic strategies for NLRP3 inflammasome-related neurological, metabolic, and inflammatory diseases. Int J Mol Sci. (2019) 20:2876. doi: 10.3390/ijms20122876
26. Owona, BA, Abia, WA, and Moundipa, PF. Natural compounds flavonoids as modulators of inflammasomes in chronic diseases. Int Immunopharmacol. (2020) 84:106498. doi: 10.1016/j.intimp.2020.106498
27. Couillin, I, Pétrilli, V, and Martinon, F. The inflammasomes. Berlin: Springer Science & Business Media (2011).
28. Christ, A, Günther, P, Lauterbach, MAR, Duewell, P, Biswas, D, Pelka, K, et al. Western diet triggers NLRP3-dependent innate immune reprogramming. Cells. (2018) 172:162–75. doi: 10.1016/j.cell.2017.12.013
29. Kopp, W . How western diet and lifestyle drive the pandemic of obesity and civilization diseases. Diabetes Metab Syndr Obes. (2019) 12:2221–36. doi: 10.2147/DMSO.S216791
30. Jaster, R, Gupta, Y, Rohde, S, Ehlers, L, Nizze, H, Vorobyev, A, et al. Impact of diet and genes on murine autoimmune pancreatitis. J Cell Mol Med. (2020) 24:8862–70. doi: 10.1111/jcmm.15540
31. Gao, Y, Hua, R, Hu, K, and Wang, Z. Carbohydrates deteriorate fatty liver by activating the inflammatory response. Nutr Res Rev. (2022) 35:252–67. doi: 10.1017/S0954422421000202
32. García-Montero, C, Fraile-Martínez, O, Gómez-Lahoz, AM, Pekarek, L, Castellanos, AJ, Noguerales-Fraguas, F, et al. Nutritional components in Western diet versus Mediterranean diet at the gut microbiota–immune system interplay. Implications for health and disease. Nutrients. (2021) 13:699. doi: 10.3390/nu13020699
33. Conteh, AR, and Huang, R. Targeting the gut microbiota by Asian and Western dietary constituents: a new avenue for diabetes. Toxicol Res (Camb). (2020) 9:569–77. doi: 10.1093/toxres/tfaa065
34. Martel, J, Chang, S-H, Ko, Y-F, Hwang, T-L, Young, JD, and Ojcius, DM. Gut barrier disruption and chronic disease. Trends Endocrinol Metab. (2022) 4:247–65. doi: 10.1016/j.tem.2022.01.002
35. Berná, G, and Romero-Gomez, M. The role of nutrition in non-alcoholic fatty liver disease: pathophysiology and management. Liver Int. (2020) 40:102–8. doi: 10.1111/liv.14360
36. Szabo, G, and Petrasek, J. Inflammasome activation and function in liver disease. Nat Rev Gastroenterol Hepatol. (2015) 12:387–400. doi: 10.1038/nrgastro.2015.94
37. Baruah, JB . Chemistry of phenolic compounds: State of the art. Hauppauge, NY, USA: Nova Science Publishers (2011).
38. Durazzo, A, Lucarini, M, Souto, EB, Cicala, C, Caiazzo, E, Izzo, AA, et al. Polyphenols: a concise overview on the chemistry, occurrence, and human health. Phyther Res. (2019) 33:2221–43. doi: 10.1002/ptr.6419
39. Wang, T, Xu, H, Dong, R, Wu, S, Guo, Y, and Wang, D. Effectiveness of targeting the NLRP3 inflammasome by using natural polyphenols: a systematic review of implications on health effects. Food Res Int. (2023) 165:112567. doi: 10.1016/j.foodres.2023.112567
40. Kinra, M, Joseph, A, Nampoothiri, M, Arora, D, and Mudgal, J. Inhibition of NLRP3-inflammasome mediated IL-1β release by phenylpropanoic acid derivatives: in-silico and in-vitro approach. Eur J Pharm Sci. (2021) 157:105637. doi: 10.1016/j.ejps.2020.105637
41. Jena, AB, Dash, UC, and Duttaroy, AK. An in silico investigation on the interactions of curcumin and epigallocatechin-3-gallate with NLRP3 inflammasome complex. Biomed Pharmacother. (2022) 156:113890. doi: 10.1016/j.biopha.2022.113890
42. Buchman, AL . Nutritional Care of the Patient with Gastrointestinal Disease. The Netherlands: CRC Press (2015).
43. Özenver, N, and Efferth, T. Phytochemical inhibitors of the NLRP3 inflammasome for the treatment of inflammatory diseases. Pharmacol Res. (2021) 170:105710. doi: 10.1016/j.phrs.2021.105710
44. Aruna, R, Geetha, A, and Suguna, P. Rutin modulates ASC expression in NLRP3 inflammasome: a study in alcohol and cerulein-induced rat model of pancreatitis. Mol Cell Biochem. (2014) 396:269–80. doi: 10.1007/s11010-014-2162-8
45. Jin, H, Yang, X, Zhao, K, Mei, F, Zhou, Y, You, Y, et al. Apocynin alleviates lung injury by suppressing NLRP3 inflammasome activation and NF-κB signaling in acute pancreatitis. Int Immunopharmacol. (2019) 75:105821. doi: 10.1016/j.intimp.2019.105821
46. Yuvaraj, K, and Geetha, A. Effect of Morus alba root bark extract on gene-level expression of inflammatory markers in rats subjected to ethanol and cerulein induced pancreatitis–influence of heat shock protein 70. J Complement Integr Med. (2019) 16:149. doi: 10.1515/jcim-2017-0149
47. Wen, Y, Han, C, Liu, T, Wang, R, Cai, W, Yang, J, et al. Chaiqin chengqi decoction alleviates severity of acute pancreatitis via inhibition of TLR4 and NLRP3 inflammasome: identification of bioactive ingredients via pharmacological sub-network analysis and experimental validation. Phytomedicine. (2020) 79:153328. doi: 10.1016/j.phymed.2020.153328
48. Pădureanu, V, Florescu, DN, Pădureanu, R, Ghenea, AE, Gheonea, DI, and Oancea, CN. Role of antioxidants and oxidative stress in the evolution of acute pancreatitis (review). Exp Ther Med. (2022) 23:1–5. doi: 10.3892/etm.2022.11120
49. Cordaro, M, Fusco, R, D’Amico, R, Siracusa, R, Peritore, AF, Gugliandolo, E, et al. Nuts modulate the Nrf2 and NLRP3 pathways in pancreas and lung after induction of acute pancreatitis by cerulein. Antioxidants. (2020) 9:992. doi: 10.3390/antiox9100992
50. Miao, F, Shan, C, Ma, T, Geng, S, and Ning, D. Walnut oil alleviates DSS–induced colitis in mice by inhibiting NLRP3 inflammasome activation and regulating gut microbiota. Microb Pathog. (2021) 154:104866. doi: 10.1016/j.micpath.2021.104866
51. Barbalho, SM, Bosso, H, Salzedas-Pescinini, LM, and de Alvares, GR. Green tea: a possibility in the therapeutic approach of inflammatory bowel diseases?: Green tea and inflammatory bowel diseases. Complement Ther Med. (2019) 43:148–53. doi: 10.1016/j.ctim.2019.01.015
52. Marinho, S, Illanes, M, Ávila-Román, J, Motilva, V, and Talero, E. Anti-inflammatory effects of rosmarinic acid-loaded nanovesicles in acute colitis through modulation of NLRP3 inflammasome. Biomol Ther. (2021) 11:162. doi: 10.3390/biom11020162
53. Lopes de Oliveira, GA, Alarcón de la Lastra, C, Rosillo, MÁ, Castejon Martinez, ML, Sánchez-Hidalgo, M, Rolim Medeiros, JV, et al. Preventive effect of bergenin against the development of TNBS-induced acute colitis in rats is associated with inflammatory mediators inhibition and NLRP3/ASC inflammasome signaling pathways. Chem Biol Interact. (2019) 297:25–33. doi: 10.1016/j.cbi.2018.10.020
54. You, X-Y, Zhang, H-Y, Han, X, Wang, F, Zhuang, P, and Zhang, Y-J. Intestinal mucosal barrier is regulated by intestinal tract neuro-immune interplay. Front Pharmacol. (2021) 12:659716. doi: 10.3389/fphar.2021.659716
55. Catalkaya, G, Venema, K, Lucini, L, Rocchetti, G, Delmas, D, Daglia, M, et al. Interaction of dietary polyphenols and gut microbiota: microbial metabolism of polyphenols, influence on the gut microbiota, and implications on host health. Food Front. (2020) 1:109–33. doi: 10.1002/fft2.25
56. Castejón, ML, Montoya, T, Ortega-Vidal, J, Altarejos, J, and Alarcón-de-la-Lastra, C. Ligstroside aglycon, an extra virgin olive oil secoiridoid, prevents inflammation by regulation of MAPKs, JAK/STAT, NF-κB, Nrf2/HO-1, and NLRP3 inflammasome signaling pathways in LPS-stimulated murine peritoneal macrophages. Food Funct. (2022) 13:10200–9. doi: 10.1039/D2FO00351A
57. Chen, L, Fan, Z, Sun, X, Qiu, W, Chen, Y, Zhou, J, et al. Mendelian randomization rules out causation between inflammatory bowel disease and non-alcoholic fatty liver disease. Front Pharmacol. (2022) 13:13. doi: 10.3389/fphar.2022.891410
58. Hurtado-Navarro, L, Angosto-Bazarra, D, Pelegrín, P, Baroja-Mazo, A, and Cuevas, S. NLRP3 inflammasome and Pyroptosis in liver pathophysiology: the emerging relevance of Nrf2 inducers. Antioxidants. (2022) 11:1–15. doi: 10.3390/antiox11050870
59. Shi, A, Shi, H, Wang, Y, Liu, X, Cheng, Y, Li, H, et al. Activation of Nrf2 pathway and inhibition of NLRP3 inflammasome activation contribute to the protective effect of chlorogenic acid on acute liver injury. Int Immunopharmacol. (2018) 54:125–30. doi: 10.1016/j.intimp.2017.11.007
60. Ma, X, Zheng, X, Pan, L, and Zhang, X. NLRP3 inflammasome activation in liver cirrhotic patients. Biochem Biophys Res Commun. (2018) 505:40–4. doi: 10.1016/j.bbrc.2018.09.055
61. Chen, Y, Que, R, Lin, L, Shen, Y, Liu, J, and Li, Y. Inhibition of oxidative stress and NLRP3 inflammasome by Saikosaponin-d alleviates acute liver injury in carbon tetrachloride-induced hepatitis in mice. Int J Immunopathol Pharmacol. (2020) 34:2058738420950593. doi: 10.1177/2058738420950593
62. Tehami, W, Nani, A, Khan, NA, and Hichami, A. New insights into the anticancer effects of p-coumaric acid: focus on colorectal cancer. Dose-Response. (2023) 21:15593258221150704. doi: 10.1177/15593258221150704
63. Dooley, JS, Lok, AS, Garcia-Tsao, G, and Pinzani, M. Sherlock’s diseases of the liver and biliary system. India: John Wiley & Sons (2018).
64. Tariq, Z, Green, CJ, and Hodson, L. Are oxidative stress mechanisms the common denominator in the progression from hepatic steatosis towards non-alcoholic steatohepatitis (NASH)? Liver Int. (2014) 34:e180–90. doi: 10.1111/liv.12523
65. Huby, T, and Gautier, EL. Immune cell-mediated features of non-alcoholic steatohepatitis. Nat Rev Immunol. (2022) 22:429–43. doi: 10.1038/s41577-021-00639-3
66. Huang, Y, Wan, T, Pang, N, Zhou, Y, Jiang, X, Li, B, et al. Cannabidiol protects livers against nonalcoholic steatohepatitis induced by high-fat high cholesterol diet via regulating NF-κB and NLRP3 inflammasome pathway. J Cell Physiol. (2019) 234:21224–34. doi: 10.1002/jcp.28728
67. Xue, KS, Tang, L, Shen, CL, Pollock, BH, Guerra, F, Phillips, TD, et al. Increase in aflatoxin exposure in two populations residing in east and West Texas, United States. Int J Hyg Environ Health. (2021) 231:113662. doi: 10.1016/j.ijheh.2020.113662
68. Li, S, Liu, R, Xia, S, Wei, G, Ishfaq, M, Zhang, Y, et al. Ecotoxicology and environmental safety protective role of curcumin on aflatoxin B1-induced TLR4 / RIPK pathway mediated-necroptosis and inflammation in chicken liver. Ecotoxicol Environ Saf. (2022) 233:113319. doi: 10.1016/j.ecoenv.2022.113319
69. Wang, D, Gao, Q, Wang, T, Kan, Z, Li, X, Hu, L, et al. Green tea polyphenols and epigallocatechin-3-gallate protect against perfluorodecanoic acid induced liver damage and inflammation in mice by inhibiting NLRP3 inflammasome activation. Food Res Int. (2020) 127:108628. doi: 10.1016/j.foodres.2019.108628
70. Xie, Y-L, Chu, J-G, Jian, X-M, Dong, J-Z, Wang, L-P, Li, G-X, et al. Curcumin attenuates lipopolysaccharide/d-galactosamine-induced acute liver injury by activating Nrf2 nuclear translocation and inhibiting NF-kB activation. Biomed Pharmacother. (2017) 91:70–7. doi: 10.1016/j.biopha.2017.04.070
71. Ding, X-Q, Wu, W-Y, Jiao, R-Q, Gu, T-T, Xu, Q, Pan, Y, et al. Curcumin and allopurinol ameliorate fructose-induced hepatic inflammation in rats via miR-200a-mediated TXNIP/NLRP3 inflammasome inhibition. Pharmacol Res. (2018) 137:64–75. doi: 10.1016/j.phrs.2018.09.021
72. Zhu, X, Lin, X, Zhang, P, Liu, Y, Ling, W, and Guo, H. Upregulated NLRP3 inflammasome activation is attenuated by anthocyanins in patients with nonalcoholic fatty liver disease: a case-control and an intervention study. Clin Res Hepatol Gastroenterol. (2022) 46:101843. doi: 10.1016/j.clinre.2021.101843
73. Zhou, Y, Wang, S, Wan, T, Huang, Y, Pang, N, Jiang, X, et al. Cyanidin-3-O-β-glucoside inactivates NLRP3 inflammasome and alleviates alcoholic steatohepatitis via SirT1/NF-κB signaling pathway. Free Radic Biol Med. (2020) 160:334–41. doi: 10.1016/j.freeradbiomed.2020.08.006
75. Stefanska, J, and Pawliczak, R. Apocynin: Molecular aptitudes. Mediat Inflamm. (2008) 2008:106507. doi: 10.1155/2008/106507
76. Miao, F, and Ph, D. Hydroxytyrosol alleviates dextran sodium sulfate À induced colitis by inhibiting NLRP3 in fl ammasome activation and modulating gut microbiota in vivo. Nutrition. (2022) 97:111579. doi: 10.1016/j.nut.2021.111579
77. Dou, X, Yan, D, Liu, S, Gao, L, and Shan, A. Thymol alleviates LPS-induced liver inflammation and apoptosis by inhibiting NLRP3 inflammasome activation and the AMPK-mTOR-autophagy pathway. Nutrients. (2022) 14:2809. doi: 10.3390/nu14142809
Keywords: Western diet, NLRP3 inflammasome, polyphenols, pancreatitis, gastrointestinal and liver diseases, oxidative stress
Citation: Nani A and Tehami W (2023) Targeting inflammasome pathway by polyphenols as a strategy for pancreatitis, gastrointestinal and liver diseases management: an updated review. Front. Nutr. 10:1157572. doi: 10.3389/fnut.2023.1157572
Edited by:
Luca Rastrelli, University of Salerno, ItalyReviewed by:
Dorin Dragoş, Carol Davila University of Medicine and Pharmacy, RomaniaRita Celano, University of Salerno, Italy
Zhihui Tong, Nanjing General Hospital of Nanjing Military Command, China
Copyright © 2023 Nani and Tehami. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Abdelhafid Nani, bmFuaS5hYmRlbGhhZmlkQHVuaXYtYWRyYXIuZWR1LmR6