
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Nutr. , 16 February 2023
Sec. Nutritional Epidemiology
Volume 10 - 2023 | https://doi.org/10.3389/fnut.2023.1110536
This article is part of the Research Topic Molecular Mechanisms in Diet-Mediated Inflammatory Diseases View all 9 articles
 Marta Maestri1Francesco Santopaolo1Maurizio Pompili1,2
Marta Maestri1Francesco Santopaolo1Maurizio Pompili1,2 Antonio Gasbarrini1,2
Antonio Gasbarrini1,2 Francesca Romana Ponziani1,2*
Francesca Romana Ponziani1,2*Non-alcoholic fatty liver disease (NAFLD) is frequently associated with metabolic disorders, being highly prevalent in obese and diabetic patients. Many concomitant factors that promote systemic and liver inflammation are involved in NAFLD pathogenesis, with a growing body of evidence highlighting the key role of the gut microbiota. Indeed, the gut-liver axis has a strong impact in the promotion of NAFLD and in the progression of the wide spectrum of its manifestations, claiming efforts to find effective strategies for gut microbiota modulation. Diet is among the most powerful tools; Western diet negatively affects intestinal permeability and the gut microbiota composition and function, selecting pathobionts, whereas Mediterranean diet fosters health-promoting bacteria, with a favorable impact on lipid and glucose metabolism and liver inflammation. Antibiotics and probiotics have been used to improve NAFLD features, with mixed results. More interestingly, medications used to treat NAFLD-associated comorbidities may also modulate the gut microbiota. Drugs for the treatment of type 2 diabetes mellitus (T2DM), such as metformin, glucagon-like peptide-1 (GLP-1) agonists, and sodium-glucose cotransporter (SGLT) inhibitors, are not only effective in the regulation of glucose homeostasis, but also in the reduction of liver fat content and inflammation, and they are associated with a shift in the gut microbiota composition towards a healthy phenotype. Even bariatric surgery significantly changes the gut microbiota, mostly due to the modification of the gastrointestinal anatomy, with a parallel improvement in histological features of NAFLD. Other options with promising effects in reprogramming the gut-liver axis, such as fecal microbial transplantation (FMT) and next-generation probiotics deserve further investigation for future inclusion in the therapeutic armamentarium of NAFLD.
Non-alcoholic fatty liver disease (NAFLD) is considered the liver mirror of systemic metabolic dysfunction, and represents a condition driven by chronic inflammation (1). NAFLD encompasses a wide spectrum of alterations, ranging from non-alcoholic fatty liver (NAFL) to non-alcoholic steatohepatitis (NASH), with fibrosis at different stages up to cirrhosis (2).
Despite less than 10% of people suffering from NAFLD develop liver-related complications, such as cirrhosis and hepatocellular carcinoma (HCC), the economic burden of the disease is heavy. In fact, NAFLD global prevalence is around 25% in the adult population and 7.6% among children. Owing to its high prevalence, NAFLD is the most rapidly increasing cause of end-stage liver disease, HCC and liver transplantation worldwide, being already in USA and Europe the second leading cause of death (3). In addition, NAFLD is associated with an increased long-term risk of fatal and non-fatal cardiovascular events, and the risk of cardiovascular disease (CVD) is proportional to the stage of fibrosis (4).
Thus, the NAFLD umbrella includes a wide heterogeneity of patients, which cannot be defined simply by the absence of alcohol consumption. The term fatty liver disease associated with metabolic dysfunction (MAFLD) has been therefore introduced to define the disease based on its features, precisely the evidence of liver fat accumulation in addition to at least one among: overweight/obesity, type 2 diabetes mellitus (T2DM) or evidence of metabolic dysregulation (such as high blood pressure, altered lipid panel, impaired fasting glucose, or insulin resistance) (5, 6).
Based on these premises, NAFLD is an increasingly emerging global health problem. Understanding the pathophysiology and molecular mechanisms underlying NAFLD is necessary to highlight new therapeutic targets, considering that there are currently no approved drugs for the treatment of NASH and the standard of care is still based on lifestyle modification (2).
In this review, we focus on the impact of the gut microbiota on the molecular mechanisms underlying NAFLD, understanding how current therapeutic approaches used to treat NAFLD/MAFLD and its associated comorbidities may influence the natural history of the disease through gut microbiota modulation. Finally, we point the view to what may become future therapeutic weapons in NAFLD/MAFLD, acting on the gut microbiota.
The most valuable hypothesis on the development of NAFLD concerns the presence of several parallel factors that simultaneously generate and maintain inflammation, promoting liver damage with the accumulation of fibrosis. The main protagonists are high-fat diet (HFD), lipotoxicity, gut barrier dysfunction, and dysbiosis (7, 8).
In the last decades, the importance of the gut microbiota in the pathophysiology of NAFLD has strongly emerged. Several studies have been carried out to understand the gut microbiota composition. The wide variety of bacteria and the multiple factors which can modify the gut microbiota, included genetic and environmental factors, make the issue hard, with discordant results involving phyla, but also families and genera (9). Of note, some studies correlate the severity of the disease with a specific microbial signature. Enterobacteriaceae, including Escherichia coli and Shigella, Bacteroides and Ruminococcus are enriched in patients with moderate-severe fibrosis, while Faecalibacterium prausnitzii and Prevotella decrease (Figure 1) (10, 11). A recent study by Oh et al. (12) identified some bacteria and bacterial metabolic signatures that independently predict NAFLD-cirrhosis. Veillonella spp., Enterobacteriaceae and Acidaminococcus correlated positively with the severity of liver fibrosis, whereas Eubacterium spp. and Faecalibacterium prausnitzii showed opposite trends; in addition, tryptophan, and related metabolites such as indole and kynurenic acid were altered in NAFLD-cirrhosis, with an overall increase in stool tryptophan levels.
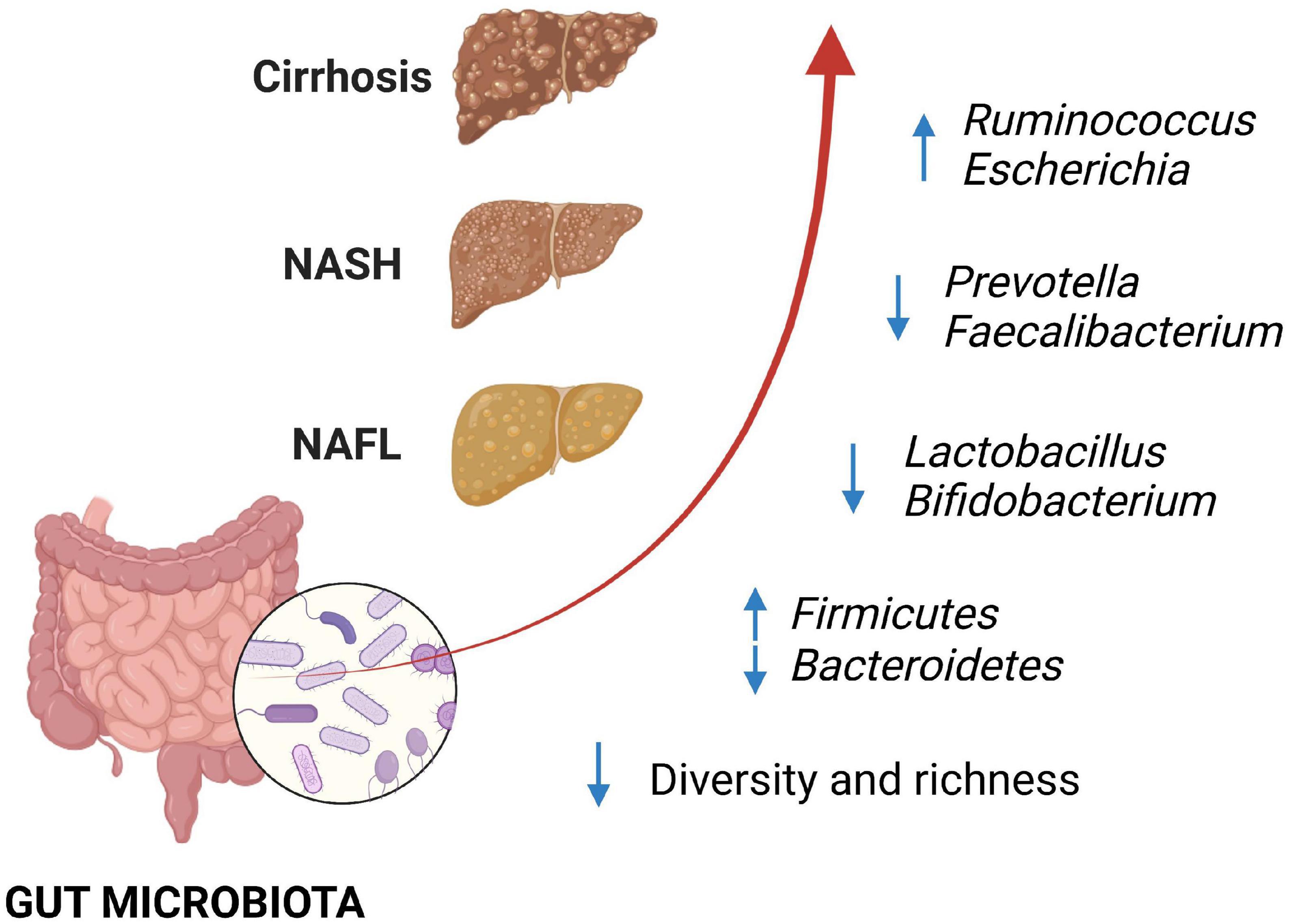
Figure 1. Changes in gut microbiota have been associated with non-alcoholic fatty liver disease (NAFLD), with progressive alteration of some bacterial components, according to the fibrosis degree. Loss of bacterial diversity and depletion of beneficial bacteria such as Lactobacillus and Bifidobacterium have been described with the progression of NAFLD, together with the increase in Ruminococcus and Escherichia in patients with advanced fibrosis. NAFLD, non-alcoholic fatty liver; NASH, non-alcoholic steatohepatitis.
Going beyond the mere association between microbial signatures and NAFLD, several studies showed that the gut microbiota is pivotal in inducing the disease phenotype. Indeed, germ-free mice receiving fecal microbial transplantation (FMT) from NASH-affected mice develop hepatic steatosis and inflammation, compared to those receiving FMT from healthy mice (13), on the contrary, FMT from healthy controls protects mice on HFD from intrahepatic lipid accumulation and inflammation (14). Furthermore, FMT from HFD fed mice into pathogen-free mice fed a standard diet, can induce intestinal epithelial barrier (IEB) and gut-vascular-barrier (GVB) derangement (15). This confirms that it is not the type of diet to induce intestinal barrier alteration with consequent bacterial translocation and establishment of inflammatory damage, but rather its influence on the gut microbiota composition. In addition, this study highlighted that the disruption of the intestinal barrier is an early event in the development of NAFLD (15) explaining why liver fat accumulation and lipotoxicity facilitated by insulin resistance (IR) are only components of a wider and more complicated picture. On the other hand, recent studies showed that allogenic FMT from healthy donors can only improve intestinal permeability in patient with NAFLD, with no effect on metabolic parameters (16). Of note, in a mice model of HFD-induced obesity, FMT was able to transmit the beneficial effects of diet and exercise on gut microbiota and metabolic profiles (17).
Noteworthy, gut microbiota exerts a continuous pressure on the immune system, especially when intestinal permeability and bacterial translocation are increased, as it happens in NAFLD (8, 18). Pathogen associated molecular patterns (PAMPs), such as lipopolysaccharide (LPS) from Gram-negative bacteria, bind toll like receptors (TLRs) expressed on epithelial cells and cells belonging to the innate immune system, modulating the inflammatory response against exogenous antigens (19–22). Some preclinical studies have shown a marked involvement of TLR4 and TLR9 in the development of steatosis, inflammation and fibrosis. In fact, TLR4- or TLR9-deficient mice given HFD or choline-deficient diet were protected from hepatic steatosis and inflammation (23, 24). Therefore, dysbiosis associated with many chronic metabolic diseases, producing a continuous immunological stimulation, can promote a condition of low-grade chronic inflammation called meta-inflammation (25, 26).
Therefore, gut microbiota modulation appears crucial in the future treatment of NAFLD/NASH.
Though several pharmacologic agents have been developed or tested for the treatment of NAFLD, diet still represents the therapeutic cornerstone (2, 27).
It is well-known that the dietary pattern strongly influences the development of NAFLD and other metabolic diseases, but also the gut microbiota (Figure 2). Western diet (WD) rich in refined sugars and saturated fat, mainly based on high red meat consumption and low fish, fruit, vegetables, and fibers intake, has been found to be associated with liver fat deposition (28–32). Conversely Mediterranean diet (MD) based on vegetables and fruit, legumes, aromatic herbs, and extra virgin olive oil as the main source of fat is associated with improvement in metabolic syndrome and intrahepatic fat accumulation (28, 33, 34).
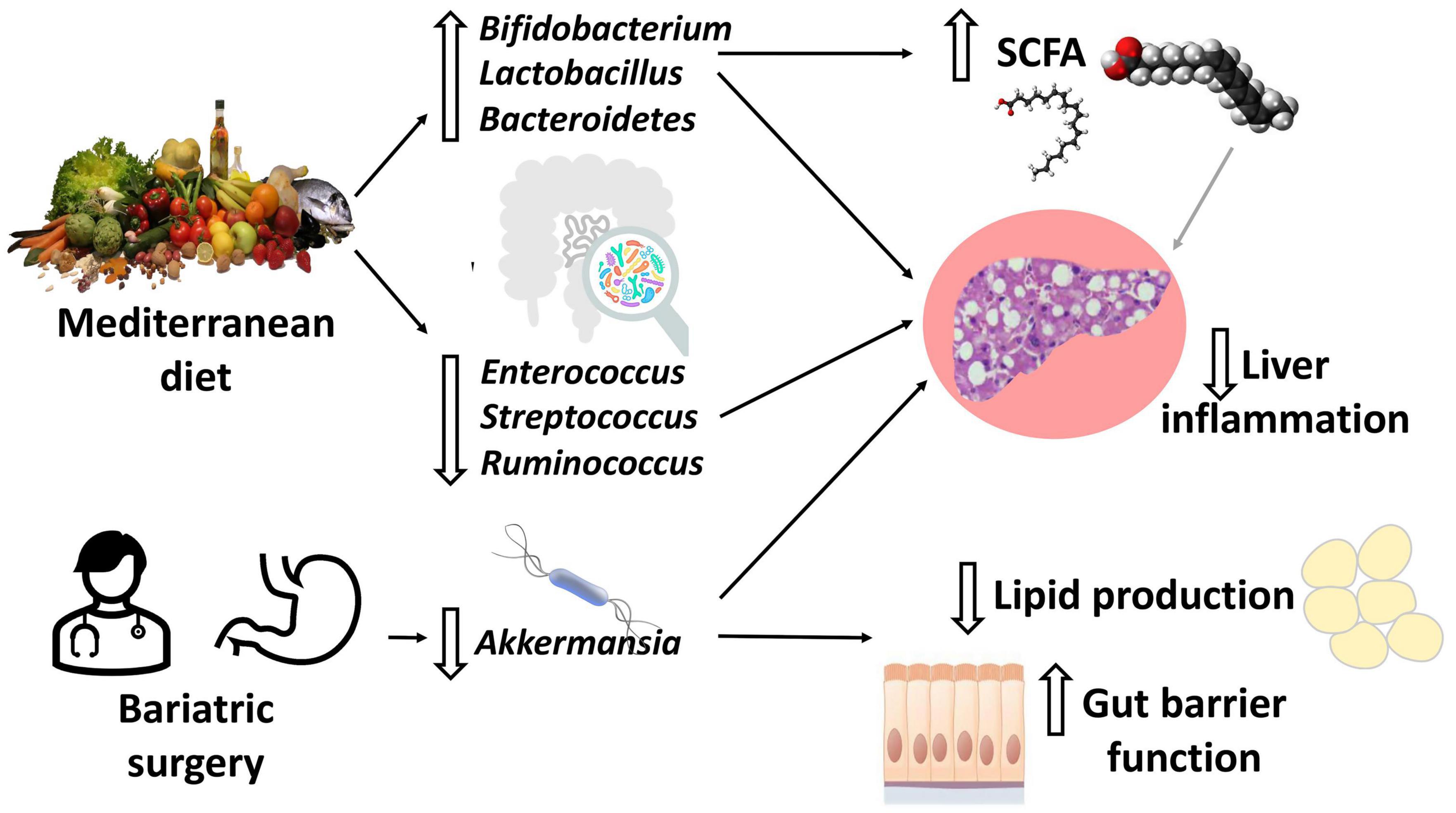
Figure 2. Diet and bariatric surgery effects on the gut microbiota. Adherence to Mediterranean diet (MD) leads to a reduction in intestinal inflammatory bacteria and an increase in beneficial bacteria, including short chain fatty acids (SCFAs)-producing bacteria (mainly Bifidobacterium and Lactobacillus). Most studies agree on the increase in Akkermansia abundance after bariatric surgery (BS). For both MD and BS, the end result is a reduction in inflammation, lipid production and preservation/improvement in the gut barrier function.
Recently, a clinical trial (35) evaluated the effects of the green-MD, a type of MD further restricted in red and processed meats and particularly enriched in green plants and polyphenols, in a population of patients affected by obesity and dyslipidemia; a significant reduction in liver fat was found with a halving of NAFLD prevalence in the study population. An effect on the gut microbiota was also reported; in particular, at the genus level, eight bacteria were significantly associated with changes in the intrahepatic fat content. Long-term adherence to MD is, in addition, protective against cardio-metabolic diseases, and this effect is greatest among Prevotella copri non-carriers in their gut microbiota (36). Fibers, one of the most beneficial and healthy element of MD, improve liver enzymes, lipid panel and fatty liver status in patients with NAFLD, but also improve intestinal permeability (37) and have a great impact on the gut microbiota composition. Overall, fibers intake increases Bifidobacterium and Bacteroidetes, and decreases inflammatory bacteria such as Enterococcus, Streptococcus, and Ruminococcus (38). A specific subtype of fibers is represented by resistant starch, which are found in foods such as banana, potatoes and corn, and enhance the growth of SCFAs producing bacteria, such as Lactobacillus and Bifidobacterium among others (39). SCFAs interact directly with the G protein-coupled receptors (GPR) 41 and 43, also known as free fatty acid receptor (FFAR) 3 and 2, respectively. The main SCFAs agonists of these receptors are acetate, butyrate and propionate, with different affinity (40). Through this interaction, SCFAs trigger anti-inflammatory pathways and the peroxisome proliferator-activated receptor (PPAR)-γ/adenosine monophosphate-activated protein kinase (AMPK) signaling pathway, the latter causing the inhibition of triglycerides and cholesterol production, the increased release of pro-peptide YY (PYY) and glucagon-like peptide 1 (GLP-1), the regulation of appetite and the improvement of intestinal barrier function (39–41). In a preclinical study, inulin fiber administration improved hepatic steatosis and fibrosis in mice, through hepatic free fatty acid receptor 2 (FFAR2)/G-protein-coupled receptor 43 (GPR43) signaling; it should be noted that this effect was mediated by the growth of the SCFAs-producing strains Bacteroides acidifaciens and Blautia producta, resulting in increased acetate content in the intestinal lumen (42). In a randomized, double-blind, cross-over study, colonic infusion of SCFAs mixtures increased fat oxidation, energy expenditure and PYY levels, also decreasing lipolysis in overweight and obese men (43). Another randomized, controlled, cross-over study demonstrated that ingestion of 10 g inulin-propionate ester significantly increased postprandial plasma PYY and GLP-1 secretion and reduced energy intake over a 24-week period; furthermore, hepatic lipid content and visceral fat deposition were reduced, while weight gain and insulin-resistance were prevented (44). In murine models, administration of sodium butyrate, reduced inflammation and liver steatosis, protecting against WD-induced NASH (45).
Fructose is another crucial dietary element involved in the onset of NAFLD, both by damaging the intestinal barrier and inducing dysbiosis (46–53). Indeed, rats fed with high-sugar-diet show an overall decrease in gut microbiota alpha diversity, as well as a reduction in Bifidobacterium, Lactobacillus, and members of Clostridiaceae family, and an increase in Coprococcus, Ruminococcus, Clostridium, and Firmicutes/Bacteroidetes ratio (51, 53, 54).
Meat, yolk, and dairy products are rich in choline, an essential nutrient involved in triglyceride metabolism and necessary for the packaging of very low-density lipoprotein (VLDL) and its export from hepatocytes (55–57). For this reason, a choline deficient diet has been adopted for decades to study NAFLD and NASH in rats (58). As the gut microbiota uses choline for the production of trimethylamine (TMA) (59), contributes to reduce choline bioavailability, mimicking the effects of a choline deficient diet (60). In addition, TMA is oxidized by hepatic flavin monooxygenases to trimethylamine oxide (TMAO) before being released into circulation (61). Noteworthy, higher levels of TMAO were found in NAFLD patients with respect to healthy controls, and correlated with fibrosis stage (62). TMAO reduces cholesterol conversion into bile acids (BAs) altering lipid homeostasis (63), promotes inflammation in adipose tissue and leads to insulin resistance (64), thus favoring the development of NAFLD and NASH. Choline deficiency has been recently linked not only to NASH development, but also to gut microbiota dysbiosis in mice; decreased abundance of Alistipes, Ruminococcaceae, Bifidobacterium, Lactobacillus, and Akkermansia, and increased abundance of Bacteroides and Ruminococcus were found (65, 66). As mentioned above, Bifidobacterium and Akkermansia usually have beneficial effects through SCFAs production and modulation of inflammatory response, while Ruminococcus is associated with fibrosis in patients with NASH.
Besides diet, physical activity has been proven to be effective in NAFLD treatment, even without weight loss or any dietary change (67–69). Eight weeks of individualized exercise reduce transaminases, markers of inflammation, and improve surrogate scores of steatosis and fibrosis. Furthermore, exercise modifies the gut microbiota, increasing the abundance of Bacteroidetes and Euryarchaeota, decreasing Actinobacteria, and improving richness (70). Both moderate-continuous and sprint-interval training reduce systemic and intestinal inflammation, and improve the gut microbiota profile by reducing Firmicutes/Bacteroidetes ratio, and decreasing Clostridium and Blautia abundance (71). Responders to exercise exhibit gut microbiota enhanced capacity for SCFAs biosynthesis and catabolism of branched-chain amino acids (72). In a 1-year lifestyle intervention with energy-restricted MD plus physical activity and behavioral support, a decrease in several members of Firmicutes and a selective increase in SCFAs producers was observed, which was paralleled by weight loss and improved CVD risk (73).
Bariatric surgery (BS) is the most effective treatment for long-term weight control in obese people (74, 75), and to effectively improve obesity-related comorbidities (74). Epidemiological data report that almost all obese patients and about 75% of overweight people are affected by NAFLD (76). Several studies and meta-analyses demonstrated a significant improvement or even resolution of NAFLD histological features, liver enzymes, glucose tolerance, and lipid panel after BS (76, 77). This beneficial effect results not only from the metabolic consequences of weight loss and visceral adipose tissue reduction, but also from the inhibition of pro-inflammatory cytokines release from adipose tissue, and the reduced supply of free fatty acids (FFAs) to the liver, with a consequent modulation of lipids and glucose metabolism (41). Roux-en-Y Gastric Bypass (RYGB), inducing anatomical changes with hepato-biliary diversion, lowers the concentration of BAs delivered to the colon; while some studies reported either an increase or a reduction in both primary and secondary BAs (78–82), overall it seems that the ratio of primary to secondary BAs decreases regardless of weight loss (83). Furthermore, BS is associated with changes in the release of gastrointestinal hormones, such as GLP-1, gastric inhibitory polypeptide, leptin, PYY, and ghrelin, which are implied in the reduction of appetite and increase in energy expenditure (84).
Changes in the gut microbiota and its related metabolites have been observed after BS, potentially being crucial in NAFLD/NASH improvement or resolution (Figure 2). Published data agree on increased microbial richness after BS, mostly Roux-en-Y Gastric Bypass (RYGB) and Sleeve Gastrectomy (SG) (9, 72, 85–87). There is also broad consensus on the increased abundance of Akkermansia muciniphila after both SG and RYGB, and, generally, after weight loss (76, 87–89). Akkermansia has been demonstrated to prevent the development of fatty liver disease in mice, reducing the expression of interleukin (IL)-6 and sterol regulatory element-binding protein (SREBP), which is involved in triglycerides synthesis in the liver (90). Akkermansia is also a mucin degrader, with the ability to reinforce epithelial barrier, and has been linked with reduction in fat deposition, and protection against insulin resistance and obesity in humans (91). While after RYGB abundance of Proteobacteria seems always to increase (78, 88), data about Firmicutes and Bacteroidetes after both RYGB and SG are discordant (76, 88, 89). A recent meta-analysis investigating human studies and animal experiments involving six different BS techniques, found a reduction in Firmicutes abundance with concomitant increase in Bacteroidetes, Proteobacteria, Verrucomicrobia, and Fusobacteria (92). Also during the long-term follow-up of nine severe obese patients who underwent biliopancreatic diversion, gastric bypass, or SG, Enterobacteriaceae enrichment was observed, while Clostridiaceae and Lachnospiraceae decreased (86). Interestingly, Tremaroli et al. (93) showed that changes in the gut microbiome did not depend on body mass index (BMI) variation.
However, it is not clear whether changes in the gut microbiota are a mere consequence of the anatomical, hormonal, and metabolic changes, or take part to these modifications contributing to the beneficial effects of BS. The most important proof of concept that the gut microbiota is a main actor in this context is provided by FMT models. Indeed, FMT from mice treated with RYGB to non-operated germ-free mice or from obese patients treated with RYBG or vertical banded gastroplasty to germ-free mice resulted in weight loss and reduced fat mass (93, 94).
Due to the strong association between NAFLD and diabetes (6, 95) in absence of drugs specifically approved for the treatment of NAFLD the effect of anti-diabetes drugs on this disease have been object of great interest. Intriguingly, the effect on gut microbiota composition, which is a crucial element in NAFLD pathogenesis and progression, may contribute to their benefit in this setting (Figure 3 and Table 1).
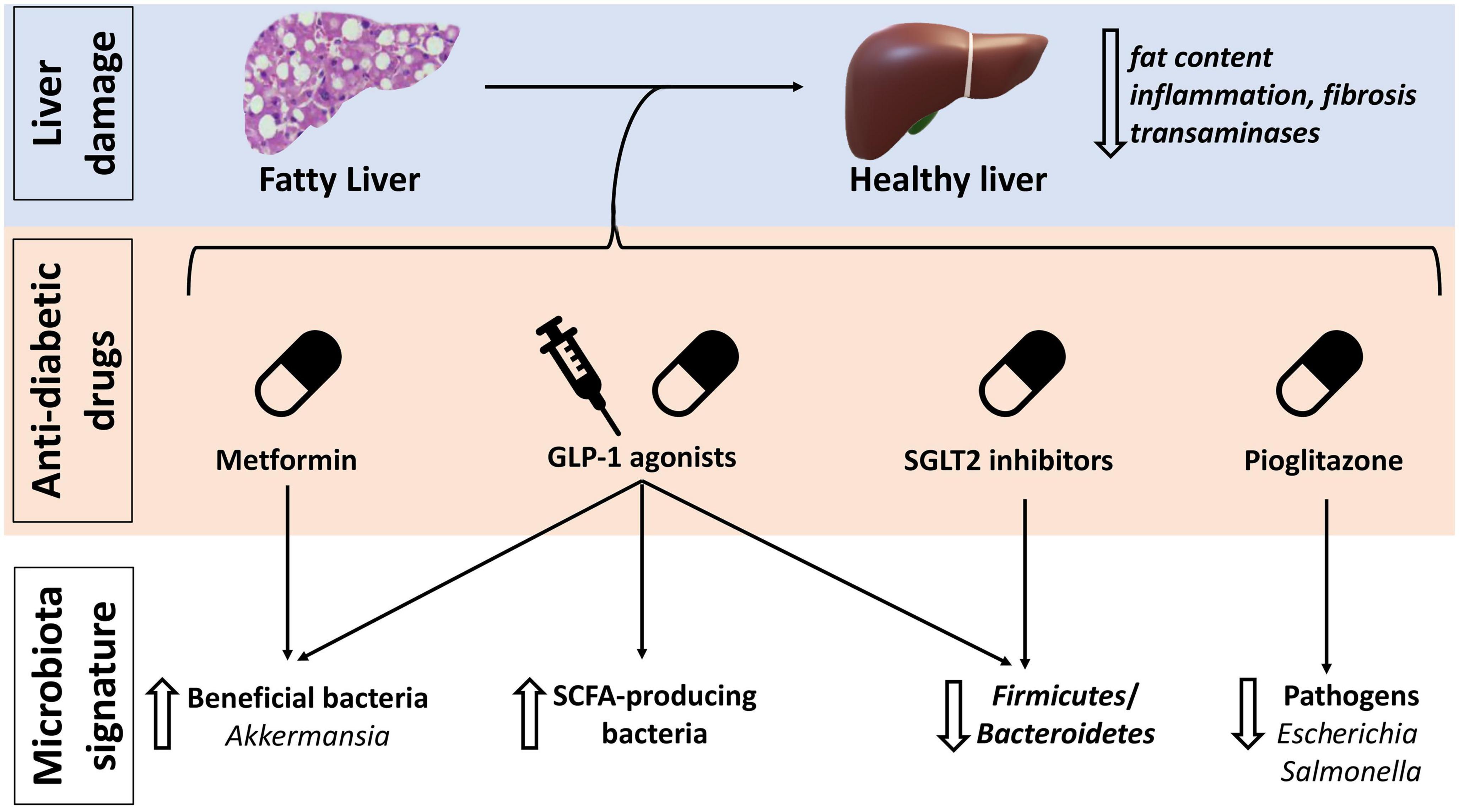
Figure 3. Anti-diabetic drugs effects on non-alcoholic fatty liver disease (NAFLD) and gut microbiota. GLP-1, glucagon-like peptide 1; SCFA, short-chain fatty acids; SGLT, sodium-glucose cotransporter inhibitors.
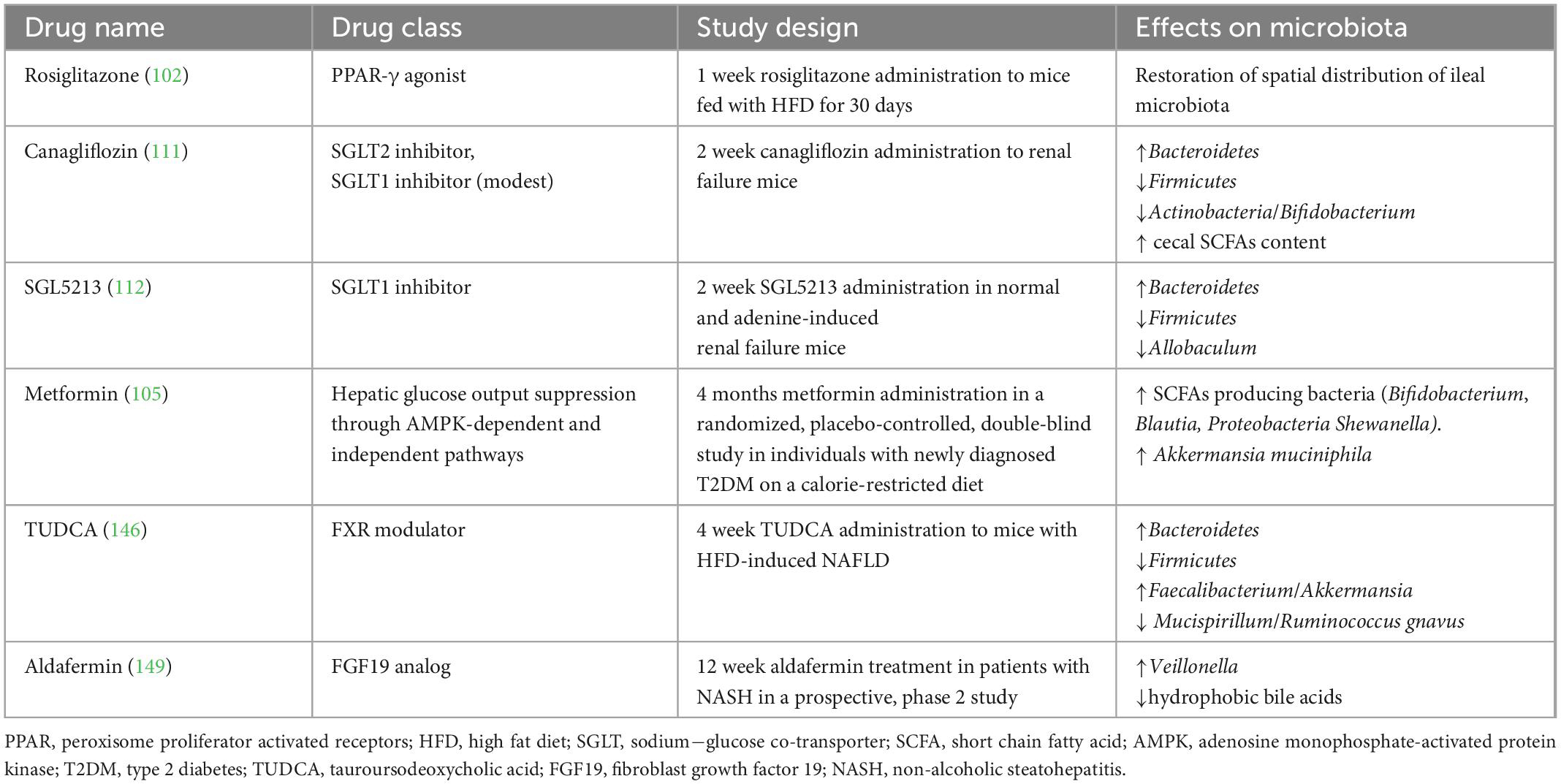
Table 1. Studies investigating the effect of anti-diabetic drugs and farnesoid X receptor (FXR) modulators on the gut microbiota.
Pioglitazone appears to be especially effective in reducing liver fat content, fibrosis and liver enzymes, despite causing an increase in BMI (96, 97). Pioglitazone belongs to PPARs agonists. PPARs are a group of receptors involved in glucose and lipid metabolism and in the anti-inflammatory response in NAFLD/NASH (98–100). They also take part in gut microbiota modulation, being involved in commensal bacteria homeostasis and prevention of the growth of pathobionts such as Escherichia and Salmonella (99–101). As demonstrated in HFD fed mice, rosiglitazone restores a healthy gut microbiota and improves intestinal permeability after only 1 week of treatment (102).
Metformin has shown some improvement in glucose tolerance, liver function and steatosis, in patients with NAFLD associated or not whit diabetes (96, 103, 104). Its effects are mediated, at least partially, by gut microbiota modulation, as metformin is able to select SCFAs producing bacteria such as Bifidobacterium, Blautia, and Shewanella, increasing also the abundance of Akkermansia muciniphila (105).
GLP-1 agonists are another relatively new class of drugs effective in diabetes and also able to ameliorate BMI, liver enzymes and liver fat content in NAFLD/NASH patients (96, 97, 106–109). A recent phase 2 trial on Semaglutide reported 40% of NASH resolution in patients with or without type 2 diabetes (107). Remarkably, in a pre-clinical study, a GLP-1/GLP-2 receptor dual agonist improved BMI, glucose homeostasis, liver triglycerides, liver fibrosis, and intestinal barrier permeability in NASH murine models. In addition, the abundance of SCFAs producing bacteria, in particular Bifidobacterium, increased, together with that of several bacteria associated with a healthy phenotype such as Prevotella, Lactobacillus, and Akkermansia; on the contrary Firmicutes, implied in obesity, were decreased (Figure 3) (110).
Lastly, sodium-glucose cotransporter 2 (SGLT2) inhibitors have also shown promising results in patients with NAFLD, liver fibrosis and steatosis (98). Canagliflozin, a SGLT2 inhibitor with also a modest inhibitory effect on SGLT1, was found to tendentially increase Bacteroidetes and decrease Firmicutes abundance, and to increase cecal SCFAs content in mice (111). Selective inhibition of SGLT1 has demonstrated to restore gut dysbiosis in renal failure mice (112, 113).
Systemic antibiotics have been one of the first experimented pharmacologic treatment for NAFLD, with beneficial effects on insulin resistance and liver fat accumulation in humans and mice models (48, 114). More recently, broad spectrum antibiotic therapy with metronidazole 1,000 mg per day plus ciprofloxacin 500 mg daily for 1 week, has been proven to reduce plasma levels of TMAO in healthy participants exposed to phosphatidylcholine challenge (115). This supports a possible use of antibiotics as disease modifiers by correction of NAFLD-associated dysbiosis via the TMAO metabolic pathway.
Rifaximin, a poorly absorbed antibiotic with eubiotic properties (116), also showed beneficial effects in patients with NAFLD. Some clinical trials in biopsy-proven NAFLD and NASH patients treated with rifaximin 1,100/1,200 mg daily have shown a significant reduction in endotoxin and liver enzymes serum levels, while reduction in BMI was only mild and no changes in the lipid profile were observed (117, 118). In addition, a NAFLD-liver fat score improvement occurred (117).
Rifaximin is currently approved for the treatment of hepatic encephalopathy, and its use is supported by data showing an overall improvement in intestinal permeability, bacterial translocation, and endotoxemia (119, 120). Preclinical studies have investigated the possible mechanisms. Rifaximin directly upregulates the expression of tight junction proteins, mainly zonula occludin-1 (ZO-1), thus lowering intestinal permeability (121, 122). A recent clinical study also showed that rifaximin improves hepatic encephalopathy by suppressing the enrichment of living from the oral cavity mucin-degrading bacteria in the colonic microbiota (i.e., Veillonella, Streptococcus, Akkermansia, and Hungatella) (120). Among these bacteria, Akkermansia is usually known for its beneficial effects including the control of host mucus turnover, layer thickness, and gut barrier preservation (123). However, it is possible that excessive mucus-degrading activity exerted by multiple elements of the gut microbiota can be potentially harmful, and may damage the intestinal barrier.
In mice model of NASH, Jian et al. (124) found that rifaximin modulates gut microbiota and reduces ileal deoxycholic acid, whereas Enomoto et al. (125) found that combination of rifaximin and the pro-kinetic lubiprostone ameliorated intestinal permeability via restoring gut epithelial tight junction proteins and counteracting LPS-induced intestinal barrier dysfunction; in addition, the abundance of Bacteroides, Lactobacillus, and Faecalibacterium increased while that of Veillonella decreased, resulting in higher levels of SCFAs.
In the last few decades, several clinical trials investigated the effects of probiotics in NAFLD. Despite good expectations, a recent meta-analysis by Tang et al. (126) showed poor efficacy in reducing body weight, and minor results on the degree of liver fat infiltration, liver enzymes, lipid panel, glucose homeostasis, and pro-inflammatory cytokines. Another meta-analysis reported the superiority of probiotics over placebo in patients with NAFLD on the improvement of BMI, liver tests, and hyperglycemia; however, probiotics failed to ameliorate liver fibrosis and did not seem as beneficial as previously suggested on lipid profile, lacking strong evidence to support a positive effect on inflammation (127). In both meta-analyses Lactobacillus spp. and Bifidobacterium spp. were the predominant strains investigated. Apart from the modest results, there was a large heterogeneity among studies, with the heavy limitation of the absence of standardization of the currently available probiotic supplements, and no guidance on the best formulations or duration of treatment to adopt, which makes it difficult to interpret the data. A further confusing element are technical limitations; in fact, commercially available probiotics and those tested in previous studies basically have an aerobic metabolism, whereas most beneficial probiotics are anaerobic (128, 129). For these reasons, numerous conflicting data on probiotics can be found in literature.
Nevertheless, research in this field is still ongoing. Starting from the evidence of a lower abundance of Akkermansia muciniphila in overweight/obesity untreated type 2 diabetes mellitus or hypertension (130), a randomized, double-blind, placebo-controlled pilot study of daily oral supplementation of 1010 A. muciniphila bacteria, either live or pasteurized, for 3 months in overweight and obese insulin-resistant subjects was performed (131). No safety issue was reported; metabolic parameters such as insulin sensitivity, plasma total cholesterol, insulin, and BMI improved; in addition, pasteurized A. muciniphila led to the reduction of liver enzymes and lipopolysaccharides (LPS) plasma levels, suggesting a strengthening effect on the intestinal barrier. However, no significant change on the overall gut microbiota community was observed, except for the enrichment of A. muciniphila abundance. Further studies are needed to bring this new probiotic in clinical practice.
FMT aims to restore intestinal homeostasis through the administration of a healthy gut microbiota. It can be performed with different techniques, mainly by endoscopic infusion or oral capsules and it is currently approved for the treatment of C. difficile infection, although potential application fields are multiple, including liver disorders (132–134).
Benefits of FMT in mice models of HFD-induced steatohepatitis has been demonstrated at many levels. A reduction in liver fat content and intrahepatic pro-inflammatory cytokines, together with improvement in NAS score has been reported after FMT; higher abundance of beneficial bacteria such as Christensenellaceae and Lactobacillus, increased butyrate cecal content and higher expression of ZO-1 were also documented, along with reduced endotoxemia (14). Histological amelioration of necro-inflammatory features, pro-inflammatory cytokines, and lipid metabolism were found after repeated FMT in humans as well (Table 2) (135). Only 6 weeks after FMT from lean donors to male recipients with metabolic syndrome, a significant increase in gut microbiota diversity and in butyrate-producing intestinal bacteria, as well as an improvement in peripheral insulin resistance was observed (136).
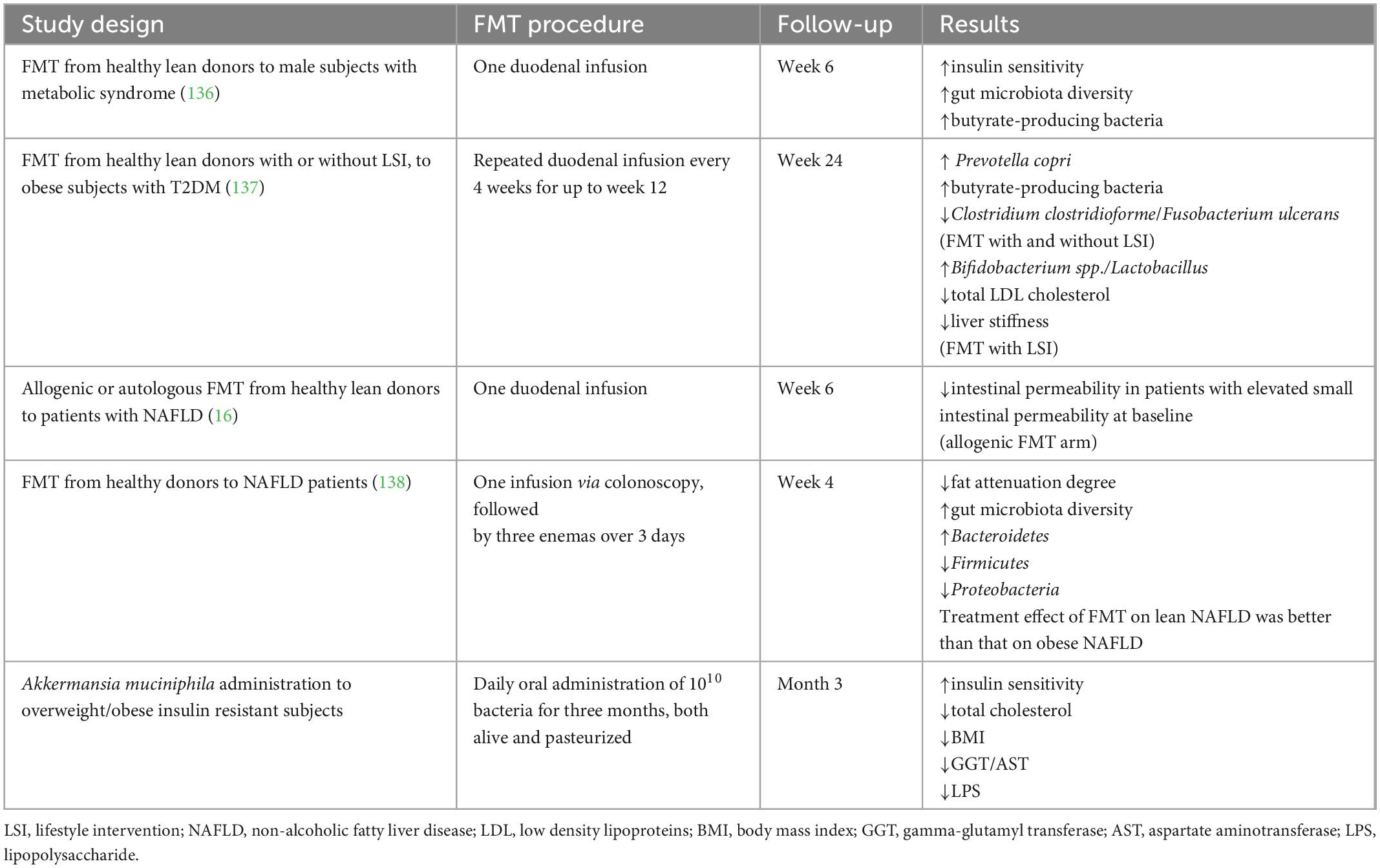
Table 2. Human studies evaluating the efficacy of fecal microbial transplantation (FMT) or next generation probiotics for the treatment of NAFLD.
A recent randomized, double-blind, placebo-controlled trial including obese subjects with type 2 diabetes demonstrated that FMT via oesophago-gastro-duodenoscopy repeated every 4 weeks for up to 12 weeks was safe and enhanced lean gut microbiota engraftment (88.2%) in this population, although better results (100%) were obtained by FMT plus lifestyle intervention (LSI). Notably, only FMT plus LSI led to lipid panel and liver stiffness improvement at week 24 (137).
Another double-blinded randomized controlled trial, conducted in patients with NAFLD, evaluated the effects of autologous or allogenic FMT from healthy donors on insulin resistance, hepatic fat content, and intestinal permeability. After 6 months, no significant benefits were observed, except for an improvement in small intestinal permeability (16). Of note, a recent randomized controlled trial conducted in patients with NAFLD, demonstrated FMT superiority over probiotics in improving liver fat content and gut microbiota composition; a superior clinical efficacy of FMT in lean NAFLD than in obese NAFLD patients was observed (138).
Given the key role of the gut barrier and gut microbiota in the pathogenesis of NAFLD, ongoing studies (Table 3) are trying to better understand the reason for the failures of FMT in this setting and to maximize its efficacy in NALFD patients.
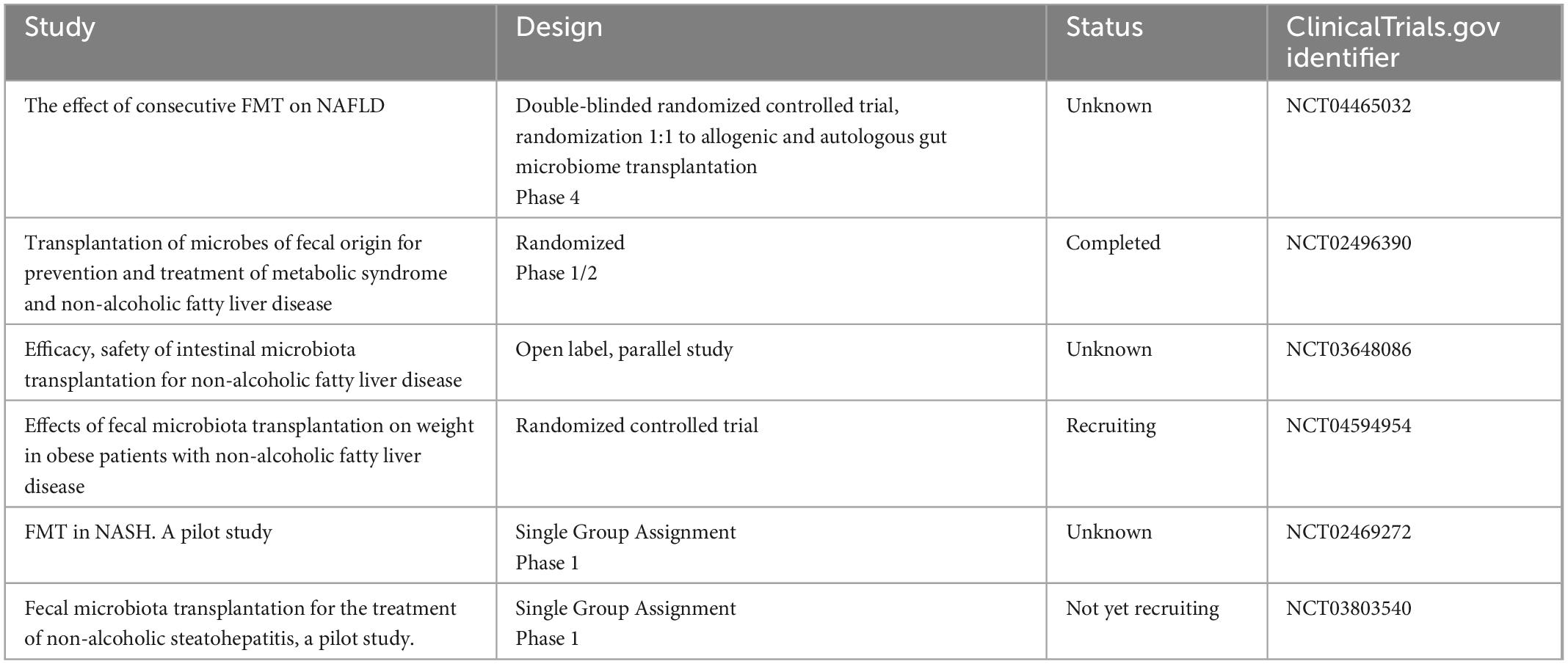
Table 3. Ongoing trials investigating the application of fecal microbial transplantation (FMT) in non-alcoholic fatty liver disease (NAFLD)/non-alcoholic steatohepatitis (NASH) treatment.
According to several preclinical studies, BAs metabolism is involved in NAFLD pathogenesis and progression through different mechanisms, including modulation of the farnesoid X receptor (FXR) signaling (139). FXR, with the downstream expression of fibroblast growth factor 19 (FGF19) in human intestine, is implicated in the negative feedback of BAs synthesis (139, 140). Beyond their toxic effect on the liver, BAs modulate the activation of Takeda G protein-coupled receptor 5 (TGR5) which regulates inflammation and glucose homeostasis, by inducing the release of GLP-1. FXR is also involved in glucose and lipid homeostasis, so that its activation promotes glucose uptake, inhibits lipogenesis, and favors fatty acids oxidation (139).
A progressive decrease in FXR expression was found in the liver of healthy controls, patients with NAFLD and patients with NASH, respectively (141, 142); notably, NAFLD-associated dysbiosis is characterized by the overabundance of bacteria producing secondary BAs, which inhibit FXR signaling (8). Indeed, BAs pool presents some differences between NAFLD/NASH and controls. Serum concentration of both primary and secondary BA is increased in NAFLD (143) as well as in NASH, both in fasting and postprandial conditions (144). A four-fold increase in glycocholate and taurocholate, and a two-fold increase in glycochenodeoxycholate were found in NASH, with a tendency for these two BAs to increase also in simple steatosis (145). Remarkably, these differences are associated with changes in the gut microbiota composition. Patients with NASH have a higher abundance of taurine and glycine metabolizing bacteria, compared with healthy patients; moreover, the FXR antagonist DCA was increased, while the agonist CDCA was decreased in NAFLD, explaining, at least partially, the altered FXR-signaling mechanism (143). In mice models of HFD-induced liver steatosis, supplementation with tauroursodeoxycholic acid (TUDCA) counteracts intestinal inflammation and intestinal barrier disruption by increasing the expression of tight junction molecules, antimicrobial peptides, lysozymes, and mucopolysaccharide, reducing serum inflammatory cytokines and intestinal lipid absorption (146). Of note, HFD fed mice showed a gut microbiota composition similar to that associated with obesity and NAFLD, with increased Firmicutes and decreased Bacteroidetes, whereas HFD fed mice treated with TUDCA presented a gut microbiota composition similar to that of normal diet fed mice, with inverted proportion of Firmicutes and Bacteroidetes and reduction in Proteobacteria (146). In addition, anti-inflammatory taxa such as Faecalibacterium and Akkermansia were increased, and pro-inflammatory taxa such as Mucispirillum and Ruminococcus gnavus were reduced in TUDCA treated mice (146), proving the strict connection between BAs, microbiota and liver inflammation.
Positive evidences in mice have led to test FXR modulating agents for the treatment of NAFLD/NASH. In the multicenter, randomized, placebo-controlled phase 3 trial REGENERATE, the FXR agonist obeticholic acid, has been shown to improve the histological features of NASH. According to other previous studies, pruritus and increased LDL cholesterol were the most commonly reported adverse events (147). Another FXR agonist, the non-bile acid agonist MET409, in a 12-week, randomized, placebo-controlled study, ameliorated liver fat content in NASH patients, showing a better and more tolerable profile then others FXR agonists (148). A recent phase 2 study evaluated the effect of aldafermin, an analogue of the intestinal hormone FGF19, on the gut microbiota in patients with NASH. Aldafermin increased the abundance of Veillonella, a commensal with lactate-degrading properties, in a dose-dependent manner, and this inversely correlated with serum levels of toxic, hydrophobic BAs. As both the richness and the diversity of the gut microbiota were substantially stable during aldafermin treatment, and only Veillonella abundance changed significantly, Veillonella has been proposed as a biomarker of treatment response (149).
NAFLD is a worldwide high-prevalent disease, nowadays one of the main causes of chronic liver disease. It is strongly associated with metabolic disorders, and heavily influenced by the gut microbiota for both onset and progression.
A healthy gut microbiota seems to have features that are lost in NAFLD. Indeed, bacterial diversity and richness are reduced, and the proportion of Firmicutes increases over Bacteroidetes; bacteria able to produce SCFAs from fermentation of dietary fibers, such as Lactobacillus and Bifidobacterium, are depleted as well as those with anti-inflammatory properties, such as Faecalibacterium and Akkermansia. Conversely, pro-inflammatory pathobionts such as Ruminococcus, Streptococcus, Enterococcus, Shigella, Escherichia, and Clostridium are well-represented. The imbalance in BAs pool, in particular the overall increase in BAs and the relative prevalence of secondary BAs, is strictly associated with gut microbiota composition and involved in the alteration of FXR signaling, with detrimental metabolic and toxic effects. Intestinal barrier impairment is another hallmark of NAFLD, being documented since the early stages of the disease.
Several NAFLD features are similar to that induced by WD regimens, while MD is associated with a healthy gut microbiota. Although weight loss and MD are still the cornerstones of NAFLD treatment, other promising treatment opportunities are landing in the NAFLD scenario, and base their efficacy on gut microbiota modulation. BS can lead even to a complete resolution of histological features of NAFLD/NASH; it is associated with an increase in gut microbiota richness and in the abundance of Akkermansia, which has anti-inflammatory and positive metabolic properties. However, little is known about the effects of increasing other bacterial phyla, such as Proteobacteria, so further studies are necessary to understand the clinical significance. Furthermore, diabetes therapeutic armamentarium is part of NAFLD management, as NAFLD and diabetes share a common metabolic background, and are strictly connected. Diabetes drugs not only ameliorate glucose homeostasis, but also influence the gut microbiota, promoting the growth of healthy bacteria. Other agents such as next-generation probiotics, mainly A. muciniphila, or FMT could be valid additional or alternative options to restore a healthy microbiota and modify the course of the disease. Finally, intestinal microbes may also be used as biomarkers of treatment response, such as Veillonella during aldafermin treatment.
The evidence discussed in this paper stems from the effort to find a common thread between gut microbiota, NAFLD, and possible therapeutic implications. However, as a final remark, it should be noted that the scientific literature is overflowing with conflicting data. This is because the gut microbiota is extremely complex in its organization, and equally variable from person to person, being strongly influenced by different pathological conditions and multiple environmental as well as socio-cultural factors. Furthermore, the taxonomic identification of bacteria reported by different studies is strictly dependent on the methodology and bioinformatics pipeline used (Supplementary Table 1). Therefore, results derived from similar studies but using different analysis methodologies may produce non-uniform data. Another element of difficulty in bringing order to this vast landscape is the fact that bacteria are often analyzed at high levels in the taxonomic scale, while at the genus and species level they may have opposite behaviors. Each bacterium is also capable of exerting different functions even though it belongs to the same genus or species, making it extremely complex to understand what metabolic results may derive from apparently taxonomically similar bacterial populations. Last but not least, given the high complexity of the microbiota-host-external factors interaction, the end result is made even more fluid and variable. As a function of this, the concepts of “healthy gut microbiota” and “disease signature” are being questioned, as it is not possible to make generalizations that can fit everyone. However, certain ecological features (e.g., a reduced alpha diversity) and specific microbial elements (such as increase of Enterobacteriaceae abundance in inflammatory conditions) are recurrent in many studies, and suggest that there is a common denominator or “core” features of the gut microbiota associated with human disease.
MM reviewed and interpreted literature data and wrote the manuscript. FS and FP interpreted literature data and wrote and revised the manuscript. AG and MP revised the manuscript. All authors read and agreed to the published version of the manuscript.
Publication cost granted by the Italian MoH RC 2022.
We thank Fondazione Roma for the continuous support to our research.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnut.2023.1110536/full#supplementary-material
1. VanWagner L, Rinella M. Extrahepatic manifestations of nonalcoholic fatty liver disease. Curr Hepatol Rep. (2016) 15:75–85. doi: 10.1007/s11901-016-0295-9
2. European Association for the Study of the Liver [EASL], European Association for the Study of Diabetes [EASD], European Association for the Study of Obesity [EASO]. EASL-EASD-EASO clinical practice guidelines for the management of non-alcoholic fatty liver disease. J Hepatol. (2016) 64:1388–402. doi: 10.1016/j.jhep.2015.11.004
3. Powell E, Wong V, Rinella M. Non-alcoholic fatty liver disease. Lancet. (2021) 397:2212–24. doi: 10.1016/S0140-6736(20)32511-3
4. Mantovani A, Csermely A, Petracca G, Beatrice G, Corey K, Simon T, et al. Non-alcoholic fatty liver disease and risk of fatal and non-fatal cardiovascular events: an updated systematic review and meta-analysis. Lancet Gastroenterol Hepatol. (2021) 6:903–13. doi: 10.1016/S2468-1253(21)00308-3
5. Eslam M, Sanyal A, George J. International consensus panel. MAFLD: a consensus-driven proposed nomenclature for metabolic associated fatty liver disease. Gastroenterology. (2020) 158:1999–2014.e1. doi: 10.1053/j.gastro.2019.11.312
6. Eslam M, Newsome P, Sarin S, Anstee Q, Targher G, Romero-Gomez M, et al. A new definition for metabolic dysfunction-associated fatty liver disease: an international expert consensus statement. J Hepatol. (2020) 73:202–9. doi: 10.1016/j.jhep.2020.07.045
7. Tilg H, Adolph T, Moschen A. Multiple parallel hits hypothesis in nonalcoholic fatty liver disease: revisited after a decade. Hepatology. (2021) 73:833–42. doi: 10.1002/hep.31518
8. Albillos A, de Gottardi A, Rescigno M. The gut-liver axis in liver disease: pathophysiological basis for therapy. J Hepatol. (2020) 72:558–77. doi: 10.1016/j.jhep.2019.10.003
9. Aron-Wisnewsky J, Vigliotti C, Witjes J, Le P, Holleboom A, Verheij J, et al. Gut microbiota and human NAFLD: disentangling microbial signatures from metabolic disorders. Nat Rev Gastroenterol Hepatol. (2020) 17:279–97. doi: 10.1038/s41575-020-0269-9
10. Loomba R, Seguritan V, Li W, Long T, Klitgord N, Bhatt A, et al. Gut microbiome-based metagenomic signature for non-invasive detection of advanced fibrosis in human nonalcoholic fatty liver disease. Cell Metab. (2017) 25:1054–62.e5. doi: 10.1016/j.cmet.2017.04.001
11. Shen F, Zheng R, Sun X, Ding W, Wang X, Fan J. Gut microbiota dysbiosis in patients with non-alcoholic fatty liver disease. Hepatobiliary Pancreat Dis Int. (2017) 16:375–81. doi: 10.1016/S1499-3872(17)60019-5
12. Oh T, Kim S, Caussy C, Fu T, Guo J, Bassirian S, et al. A universal gut-microbiome-derived signature predicts cirrhosis. Cell Metab. (2020) 32:878–88.e6. doi: 10.1016/j.cmet.2020.06.005
13. Chiu C, Ching Y, Li Y, Liu J, Huang Y, Huang Y, et al. Nonalcoholic fatty liver disease is exacerbated in high-fat diet-fed gnotobiotic mice by colonization with the gut microbiota from patients with nonalcoholic steatohepatitis. Nutrients. (2017) 9:1220. doi: 10.3390/nu9111220
14. Zhou D, Pan Q, Shen F, Cao H, Ding W, Chen Y, et al. Total fecal microbiota transplantation alleviates high-fat diet-induced steatohepatitis in mice via beneficial regulation of gut microbiota. Sci Rep. (2017) 7:1529. doi: 10.1038/s41598-017-01751-y
15. Mouries J, Brescia P, Silvestri A, Spadoni I, Sorribas M, Wiest R, et al. Microbiota-driven gut vascular barrier disruption is a prerequisite for non-alcoholic steatohepatitis development. J Hepatol. (2019) 71:1216–28. doi: 10.1016/j.jhep.2019.08.005
16. Craven L, Rahman A, Nair Parvathy S, Beaton M, Silverman J, Qumosani K, et al. Allogenic fecal microbiota transplantation in patients with nonalcoholic fatty liver disease improves abnormal small intestinal permeability: a randomized control trial. Am J Gastroenterol. (2020) 115:1055–65. doi: 10.14309/ajg.0000000000000661
17. Lai Z, Tseng C, Ho H, Cheung C, Lin J, Chen Y, et al. Fecal microbiota transplantation confers beneficial metabolic effects of diet and exercise on diet-induced obese mice. Sci Rep. (2018) 8:15625. doi: 10.1038/s41598-018-33893-y
18. Ponziani F, Zocco M, Cerrito L, Gasbarrini A, Pompili M. Bacterial translocation in patients with liver cirrhosis: physiology, clinical consequences, and practical implications. Expert Rev Gastroenterol Hepatol. (2018) 12:641–56. doi: 10.1080/17474124.2018.1481747
19. Scheithauer T, Rampanelli E, Nieuwdorp M, Vallance B, Verchere C, van Raalte D, et al. Gut microbiota as a trigger for metabolic inflammation in obesity and type 2 diabetes. Front Immunol. (2020) 11:571731. doi: 10.3389/fimmu.2020.571731
20. Gao B, Jeong W, Tian Z. Liver: an organ with predominant innate immunity. Hepatology. (2008) 47:729–36. doi: 10.1002/hep.22034
21. Di Tommaso N, Gasbarrini A, Ponziani F. Intestinal barrier in human health and disease. Int J Environ Res Public Health. (2021) 18:12836. doi: 10.3390/ijerph182312836
22. Nicoletti A, Ponziani F, Biolato M, Valenza V, Marrone G, Sganga G, et al. Intestinal permeability in the pathogenesis of liver damage: from non-alcoholic fatty liver disease to liver transplantation. World J Gastroenterol. (2019) 25:4814–34. doi: 10.3748/wjg.v25.i33.4814
23. Rivera C, Adegboyega P, van Rooijen N, Tagalicud A, Allman M, Wallace M. Toll-like receptor-4 signaling and Kupffer cells play pivotal roles in the pathogenesis of non-alcoholic steatohepatitis. J Hepatol. (2007) 47:571–9. doi: 10.1016/j.jhep.2007.04.019
24. Miura K, Kodama Y, Inokuchi S, Schnabl B, Aoyama T, Ohnishi H, et al. Toll-like receptor 9 promotes steatohepatitis by induction of interleukin-1beta in mice. Gastroenterology. (2010) 139:323–34.e7.. doi: 10.1053/j.gastro.2010.03.052
25. Esser N, Legrand-Poels S, Piette J, Scheen A, Paquot N. Inflammation as a link between obesity, metabolic syndrome and type 2 diabetes. Diabetes Res Clin Pract. (2014) 105:141–50. doi: 10.1016/j.diabres.2014.04.006
26. Calder P, Ahluwalia N, Albers R, Bosco N, Bourdet-Sicard R, Haller D, et al. A consideration of biomarkers to be used for evaluation of inflammation in human nutritional studies. Br J Nutr. (2013) 109(Suppl. 1):S1–34. doi: 10.1017/S0007114512005119
27. Ghetti F, De Oliveira D, De Oliveira J, Ferreira L, Cesar D, Moreira A. Effects of dietary intervention on gut microbiota and metabolic-nutritional profile of outpatients with non-alcoholic steatohepatitis: a randomized clinical trial. J Gastrointest Liver Dis. (2019) 28:279–87. doi: 10.15403/jgld-197
28. Merra G, Noce A, Marrone G, Cintoni M, Tarsitano M, Capacci A, et al. Influence of mediterranean diet on human gut microbiota. Nutrients. (2020) 13:7. doi: 10.3390/nu13010007
29. Mollard R, Sénéchal M, MacIntosh A, Hay J, Wicklow B, Wittmeier K, et al. Dietary determinants of hepatic steatosis and visceral adiposity in overweight and obese youth at risk of type 2 diabetes. Am J Clin Nutr. (2014) 99:804–12. doi: 10.3945/ajcn.113.079277
30. Sevastianova K, Santos A, Kotronen A, Hakkarainen A, Makkonen J, Silander K, et al. Effect of short-term carbohydrate overfeeding and long-term weight loss on liver fat in overweight humans. Am J Clin Nutr. (2012) 96:727–34. doi: 10.3945/ajcn.112.038695
31. Jensen V, Hvid H, Damgaard J, Nygaard H, Ingvorsen C, Wulff E, et al. Dietary fat stimulates development of NAFLD more potently than dietary fructose in Sprague-Dawley rats. Diabetol Metab Syndr. (2018) 10:4. doi: 10.1186/s13098-018-0307-8
32. Parry S, Rosqvist F, Mozes F, Cornfield T, Hutchinson M, Piche M, et al. Intrahepatic fat and postprandial glycemia increase after consumption of a diet enriched in saturated fat compared with free sugars. Diabetes Care. (2020) 43:1134–41. doi: 10.2337/dc19-2331
33. Montemayor S, Mascaró C, Ugarriza L, Casares M, Llompart I, Abete I, et al. Adherence to mediterranean diet and NAFLD in patients with metabolic syndrome: the FLIPAN study. Nutrients. (2022) 14:3186. doi: 10.3390/nu14153186
34. Marin-Alejandre B, Abete I, Cantero I, Monreal J, Elorz M, Herrero J, et al. The metabolic and hepatic impact of two personalized dietary strategies in subjects with obesity and nonalcoholic fatty liver disease: the fatty liver in obesity (FLiO) randomized controlled trial. Nutrients. (2019) 11:2543. doi: 10.3390/nu11102543
35. Yaskolka Meir A, Rinott E, Tsaban G, Zelicha H, Kaplan A, Rosen P, et al. Effect of green-Mediterranean diet on intrahepatic fat: the DIRECT PLUS randomised controlled trial. Gut. (2021) 70:2085–95. doi: 10.1136/gutjnl-2020-323106
36. Wang D, Nguyen L, Li Y, Yan Y, Ma W, Rinott E, et al. The gut microbiome modulates the protective association between a Mediterranean diet and cardiometabolic disease risk. Nat Med. (2021) 27:333–43. doi: 10.1038/s41591-020-01223-3
37. Krawczyk M, Maciejewska D, Ryterska K, Czerwińka-Rogowska M, Jamioł-Milc D, Skonieczna-Żydecka K, et al. Gut permeability might be improved by dietary fiber in individuals with nonalcoholic fatty liver disease (NAFLD) undergoing weight reduction. Nutrients. (2018) 10:1793. doi: 10.3390/nu10111793
38. Pérez-Montes de Oca A, Julián M, Ramos A, Puig-Domingo M, Alonso N. Microbiota, fiber, and NAFLD: is there any connection? Nutrients. (2020) 12:3100. doi: 10.3390/nu12103100
39. Zhu W, Zhou Y, Tsao R, Dong H, Zhang H. Amelioratory effect of resistant starch on non-alcoholic fatty liver disease via the gut-liver axis. Front Nutr. (2022) 9:861854. doi: 10.3389/fnut.2022.861854
40. Tan J, McKenzie C, Mariño E, Macia L, Mackay C. Metabolite-sensing G protein-coupled receptors-facilitators of diet-related immune regulation. Annu Rev Immunol. (2017) 35:371–402. doi: 10.1146/annurev-immunol-051116-052235
41. Dai X, Hou H, Zhang W, Liu T, Li Y, Wang S, et al. Microbial metabolites: critical regulators in NAFLD. Front Microbiol. (2020) 11:567654. doi: 10.3389/fmicb.2020.567654
42. Aoki R, Onuki M, Hattori K, Ito M, Yamada T, Kamikado K, et al. Commensal microbe-derived acetate suppresses NAFLD/NASH development via hepatic FFAR2 signalling in mice. Microbiome. (2021) 9:188. doi: 10.1186/s40168-021-01125-7
43. Canfora E, van der Beek C, Jocken J, Goossens G, Holst J, Olde Damink S, et al. Colonic infusions of short-chain fatty acid mixtures promote energy metabolism in overweight/obese men: a randomized crossover trial. Sci Rep. (2017) 7:2360. doi: 10.1038/s41598-017-02546-x
44. Chambers E, Viardot A, Psichas A, Morrison D, Murphy K, Zac-Varghese S, et al. Effects of targeted delivery of propionate to the human colon on appetite regulation, body weight maintenance and adiposity in overweight adults. Gut. (2015) 64:1744–54. doi: 10.1136/gutjnl-2014-307913
45. Jin C, Sellmann C, Engstler A, Ziegenhardt D, Bergheim I. Supplementation of sodium butyrate protects mice from the development of non-alcoholic steatohepatitis (NASH). Br J Nutr. (2015) 114:1745–55. doi: 10.1017/S0007114515003621
46. Lambertz J, Weiskirchen S, Landert S, Weiskirchen R. Fructose: a dietary sugar in crosstalk with microbiota contributing to the development and progression of non-alcoholic liver disease. Front Immunol. (2017) 8:1159. doi: 10.3389/fimmu.2017.01159
47. Jensen T, Abdelmalek M, Sullivan S, Nadeau K, Green M, Roncal C, et al. Fructose and sugar: a major mediator of non-alcoholic fatty liver disease. J Hepatol. (2018) 68:1063–75. doi: 10.1016/j.jhep.2018.01.019
48. Bergheim I, Weber S, Vos M, Krämer S, Volynets V, Kaserouni S, et al. Antibiotics protect against fructose-induced hepatic lipid accumulation in mice: role of endotoxin. J Hepatol. (2008) 48:983–92. doi: 10.1016/j.jhep.2008.01.035
49. Jegatheesan P, Beutheu S, Ventura G, Sarfati G, Nubret E, Kapel N, et al. Effect of specific amino acids on hepatic lipid metabolism in fructose-induced non-alcoholic fatty liver disease. Clin Nutr. (2016) 35:175–82. doi: 10.1016/j.clnu.2015.01.021
50. Volynets V, Louis S, Pretz D, Lang L, Ostaff M, Wehkamp J, et al. Intestinal barrier function and the gut microbiome are differentially affected in mice fed a western-style diet or drinking water supplemented with fructose. J Nutr. (2017) 147:770–80. doi: 10.3945/jn.116.242859
51. Jegatheesan P, Beutheu S, Freese K, Waligora-Dupriet A, Nubret E, Butel M, et al. Preventive effects of citrulline on Western diet-induced non-alcoholic fatty liver disease in rats. Br J Nutr. (2016) 116:191–203. doi: 10.1017/S0007114516001793
52. Ritze Y, Bárdos G, Claus A, Ehrmann V, Bergheim I, Schwiertz A, et al. Lactobacillus rhamnosus GG protects against non-alcoholic fatty liver disease in mice. PLoS One. (2014) 9:e80169. doi: 10.1371/journal.pone.0080169
53. Crescenzo R, Mazzoli A, Di Luccia B, Bianco F, Cancelliere R, Cigliano L, et al. Dietary fructose causes defective insulin signalling and ceramide accumulation in the liver that can be reversed by gut microbiota modulation. Food Nutr Res. (2017) 61:1331657. doi: 10.1080/16546628.2017.1331657
54. Sen T, Cawthon C, Ihde B, Hajnal A, DiLorenzo P, de La Serre C, et al. Diet-driven microbiota dysbiosis is associated with vagal remodeling and obesity. Physiol Behav. (2017) 173:305–17. doi: 10.1016/j.physbeh.2017.02.027
55. Institute of Medicine (US) Standing Committee on the Scientific Evaluation of Dietary Reference Intakes and its Panel on Folate, Other B Vitamins, and Choline. Dietary Reference Intakes for Thiamin, Riboflavin, Niacin, Vitamin B6, Folate, Vitamin B12, Pantothenic Acid, Biotin, and Choline. Washington, DC: National Academies Press (US) (1998).
56. Vance D. Role of phosphatidylcholine biosynthesis in the regulation of lipoprotein homeostasis. Curr Opin Lipidol. (2008) 19:229–34. doi: 10.1097/MOL.0b013e3282fee935
57. Yao Z, Vance D. Reduction in VLDL, but not HDL, in plasma of rats deficient in choline. Biochem Cell Biol Biochim Biol Cell. (1990) 68:552–8. doi: 10.1139/o90-079
58. Hebbard L, George J. Animal models of nonalcoholic fatty liver disease. Nat Rev Gastroenterol Hepatol. (2011) 8:35–44. doi: 10.1038/nrgastro.2010.191
59. Zhu L, Baker S, Gill C, Liu W, Alkhouri R, Baker R, et al. Characterization of gut microbiomes in nonalcoholic steatohepatitis (NASH) patients: a connection between endogenous alcohol and NASH. Hepatolology. (2013) 57:601–9. doi: 10.1002/hep.26093
60. Dumas M, Barton R, Toye A, Cloarec O, Blancher C, Rothwell A, et al. Metabolic profiling reveals a contribution of gut microbiota to fatty liver phenotype in insulin-resistant mice. Proc Natl Acad Sci USA. (2006) 103:12511–6. doi: 10.1073/pnas.0601056103
61. Janeiro M, Ramírez M, Milagro F, Martínez J, Solas M. Implication of trimethylamine N-oxide (TMAO) in disease: potential biomarker or new therapeutic target. Nutrients. (2018) 10:1398. doi: 10.3390/nu10101398
62. Chen Y, Liu Y, Zhou R, Chen X, Wang C, Tan X, et al. Associations of gut-flora-dependent metabolite trimethylamine-N-oxide, betaine and choline with non-alcoholic fatty liver disease in adults. Sci Rep. (2016) 6:19076. doi: 10.1038/srep19076
63. Koeth R, Wang Z, Levison B, Buffa J, Org E, Sheehy B, et al. Intestinal microbiota metabolism of L-carnitine, a nutrient in red meat, promotes atherosclerosis. Nat Med. (2013) 19:576–85. doi: 10.1038/nm.3145
64. Gao X, Liu X, Xu J, Xue C, Xue Y, Wang Y. Dietary trimethylamine N-oxide exacerbates impaired glucose tolerance in mice fed a high fat diet. J Biosci Bioeng. (2014) 118:476–81. doi: 10.1016/j.jbiosc.2014.03.001
65. Ye J, Li Y, Wu W, Shi D, Fang D, Yang L, et al. Dynamic alterations in the gut microbiota and metabolome during the development of methionine-choline-deficient diet-induced nonalcoholic steatohepatitis. World J Gastroenterol. (2018) 24:2468–81. doi: 10.3748/wjg.v24.i23.2468
66. Schneider K, Mohs A, Kilic K, Candels L, Elfers C, Bennek E, et al. Intestinal microbiota protects against MCD diet-induced steatohepatitis. Int J Mol Sci. (2019) 20:308. doi: 10.3390/ijms20020308
67. Babu A, Csader S, Lok J, Gómez-Gallego C, Hanhineva K, El-Nezami H, et al. Positive effects of exercise intervention without weight loss and dietary changes in NAFLD-related clinical parameters: a systematic review and meta-analysis. Nutrients. (2021) 13:3135. doi: 10.3390/nu13093135
68. Guevara-Cruz M, Flores-López A, Aguilar-López M, Sánchez-Tapia M, Medina-Vera I, Díaz D, et al. Improvement of lipoprotein profile and metabolic endotoxemia by a lifestyle intervention that modifies the gut microbiota in subjects with metabolic syndrome. J Am Heart Assoc. (2019) 8:e012401. doi: 10.1161/JAHA.119.012401
69. Wong V, Wong G, Chan R, Shu S, Cheung B, Li L, et al. Beneficial effects of lifestyle intervention in non-obese patients with non-alcoholic fatty liver disease. J Hepatol. (2018) 69:1349–56. doi: 10.1016/j.jhep.2018.08.011
70. Huber Y, Pfirrmann D, Gebhardt I, Labenz C, Gehrke N, Straub B, et al. Improvement of non-invasive markers of NAFLD from an individualised, web-based exercise program. Aliment Pharmacol Ther. (2019) 50:930–9. doi: 10.1111/apt.15427
71. Motiani K, Collado M, Eskelinen J, Virtanen K, Löyttyniemi E, Salminen S, et al. Exercise training modulates gut microbiota profile and improves endotoxemia. Med Sci Sports Exerc. (2020) 52:94–104. doi: 10.1249/MSS.0000000000002112
72. Liu Y, Wang Y, Ni Y, Cheung C, Lam K, Wang Y, et al. Gut microbiome fermentation determines the efficacy of exercise for diabetes prevention. Cell Metab. (2020) 31:77–91.e5. doi: 10.1016/j.cmet.2019.11.001
73. Muralidharan J, Moreno-Indias I, Bulló M, Lopez J, Corella D, Castañer O, et al. Effect on gut microbiota of a 1-y lifestyle intervention with Mediterranean diet compared with energy-reduced Mediterranean diet and physical activity promotion: PREDIMED-plus Study. Am J Clin Nutr. (2021) 114:1148–58. doi: 10.1093/ajcn/nqab150
74. Maggard M, Shugarman L, Suttorp M, Maglione M, Sugerman H, Livingston E, et al. Meta-analysis: surgical treatment of obesity. Ann Intern Med. (2005) 142:547–59. doi: 10.7326/0003-4819-142-7-200504050-00013
75. Colquitt J, Picot J, Loveman E, Clegg A. Surgery for obesity. Cochrane Database Syst Rev. (2009) 2:CD003641. doi: 10.1002/14651858.CD003641.pub3
76. Cerreto M, Santopaolo F, Gasbarrini A, Pompili M, Ponziani F. Bariatric surgery and liver disease: general considerations and role of the gut-liver axis. Nutrients. (2021) 13:2649. doi: 10.3390/nu13082649
77. Lee Y, Doumouras A, Yu J, Brar K, Banfield L, Gmora S, et al. Complete resolution of nonalcoholic fatty liver disease after bariatric surgery: a systematic review and meta-analysis. Clin Gastroenterol Hepatol. (2019) 17:1040–60.e11. doi: 10.1016/j.cgh.2018.10.017
78. Ilhan Z, DiBaise J, Dautel S, Isern N, Kim Y, Hoyt D, et al. Temporospatial shifts in the human gut microbiome and metabolome after gastric bypass surgery. NPJ Biofilms Microbiomes. (2020) 6:12. doi: 10.1038/s41522-020-0122-5
79. de Siqueira Cardinelli C, Torrinhas R, Sala P, Pudenzi M, Fernando F Angolini C, Marques da Silva M, et al. Fecal bile acid profile after Roux-en-Y gastric bypass and its association with the remission of type 2 diabetes in obese women: a preliminary study. Clin Nutr. (2019) 38:2906–12. doi: 10.1016/j.clnu.2018.12.028
80. Sachdev S, Wang Q, Billington C, Connett J, Ahmed L, Inabnet W, et al. FGF 19 and bile acids increase following Roux-en-Y gastric bypass but not after medical management in patients with type 2 diabetes. Obes Surg. (2016) 26:957–65. doi: 10.1007/s11695-015-1834-0
81. Gerhard G, Styer A, Wood G, Roesch S, Petrick A, Gabrielsen J, et al. A role for fibroblast growth factor 19 and bile acids in diabetes remission after Roux-en-Y gastric bypass. Diabetes Care. (2013) 36:1859–64. doi: 10.2337/dc12-2255
82. Steinert R, Peterli R, Keller S, Meyer-Gerspach A, Drewe J, Peters T, et al. Bile acids and gut peptide secretion after bariatric surgery: a 1-year prospective randomized pilot trial. Obesity. (2013) 21:E660–8. doi: 10.1002/oby.20522
83. Talavera-Urquijo E, Beisani M, Balibrea J, Alverdy J. Is bariatric surgery resolving NAFLD via microbiota-mediated bile acid ratio reversal? A comprehensive review. Surg Obes Relat Dis. (2020) 16:1361–9. doi: 10.1016/j.soard.2020.03.013
84. Tabasi M, Ashrafian F, Khezerloo J, Eshghjoo S, Behrouzi A, Javadinia S, et al. Changes in gut microbiota and hormones after bariatric surgery: a bench-to-bedside review. Obes Surg. (2019) 29:1663–74. doi: 10.1007/s11695-019-03779-7
85. Chen G, Zhuang J, Cui Q, Jiang S, Tao W, Chen W, et al. Two bariatric surgical procedures differentially alter the intestinal microbiota in obesity patients. Obes Surg. (2020) 30:2345–61. doi: 10.1007/s11695-020-04494-4
86. Juárez-Fernández M, Román-Sagüillo S, Porras D, García-Mediavilla M, Linares P, Ballesteros-Pomar M, et al. Long-term effects of bariatric surgery on gut microbiota composition and faecal metabolome related to obesity remission. Nutrients. (2021) 13:2519. doi: 10.3390/nu13082519
87. Koutoukidis D, Jebb S, Zimmerman M, Otunla A, Henry J, Ferrey A, et al. The association of weight loss with changes in the gut microbiota diversity, composition, and intestinal permeability: a systematic review and meta-analysis. Gut Microbes. (2022) 14:2020068. doi: 10.1080/19490976.2021.2020068
88. Martínez-Montoro J, Kuchay M, Balaguer-Román A, Martínez-Sánchez M, Frutos M, Fernández-García J, et al. Gut microbiota and related metabolites in the pathogenesis of nonalcoholic steatohepatitis and its resolution after bariatric surgery. Obes Rev. (2022) 23:e13367. doi: 10.1111/obr.13367
89. Xu G, Song M. Recent advances in the mechanisms underlying the beneficial effects of bariatric and metabolic surgery. Surg Obes Relat Dis. (2021) 17:231–8. doi: 10.1016/j.soard.2020.08.028
90. Kim S, Lee Y, Kim Y, Seo Y, Lee H, Ha J, et al. Akkermansia muciniphila prevents fatty liver disease, decreases serum triglycerides, and maintains gut homeostasis. Appl Environ Microbiol. (2020) 86:e3004–19. doi: 10.1128/AEM.03004-19
91. Dao M, Everard A, Aron-Wisnewsky J, Sokolovska N, Prifti E, Verger E, et al. Akkermansia muciniphila and improved metabolic health during a dietary intervention in obesity: relationship with gut microbiome richness and ecology. Gut. (2016) 65:426–36. doi: 10.1136/gutjnl-2014-308778
92. Guo Y, Huang Z, Liu C, Qi L, Sheng Y, Zou D. Modulation of the gut microbiome: a systematic review of the effect of bariatric surgery. Eur J Endocrinol. (2018) 178:43–56. doi: 10.1530/EJE-17-0403
93. Tremaroli V, Karlsson F, Werling M, Ståhlman M, Kovatcheva-Datchary P, Olbers T, et al. Roux-en-Y gastric bypass and vertical banded gastroplasty induce long-term changes on the human gut microbiome contributing to fat mass regulation. Cell Metab. (2015) 22:228–38. doi: 10.1016/j.cmet.2015.07.009
94. Liou A, Paziuk M, Luevano J, Machineni S, Turnbaugh P, Kaplan L. Conserved shifts in the gut microbiota due to gastric bypass reduce host weight and adiposity. Sci Transl Med. (2013) 5:178ra41. doi: 10.1126/scitranslmed.3005687
95. Mantovani A, Byrne C, Bonora E, Targher G. Nonalcoholic fatty liver disease and risk of incident type 2 diabetes: a meta-analysis. Diabetes Care. (2018) 41:372–82. doi: 10.2337/dc17-1902
96. Mantovani A, Byrne C, Scorletti E, Mantzoros C, Targher G. Efficacy and safety of anti-hyperglycaemic drugs in patients with non-alcoholic fatty liver disease with or without diabetes: an updated systematic review of randomized controlled trials. Diabetes Metab. (2020) 46:427–41. doi: 10.1016/j.diabet.2019.12.007
97. Kumar J, Memon R, Shahid I, Rizwan T, Zaman M, Menezes R, et al. Antidiabetic drugs and non-alcoholic fatty liver disease: a systematic review, meta-analysis and evidence map. Dig Liver Dis. (2021) 53:44–51. doi: 10.1016/j.dld.2020.08.021
98. Xu X, Poulsen K, Wu L, Liu S, Miyata T, Song Q, et al. Targeted therapeutics and novel signaling pathways in non-alcohol-associated fatty liver/steatohepatitis (NAFL/NASH). Signal Transduct Target Ther. (2022) 7:287. doi: 10.1038/s41392-022-01119-3
99. Yu Q, Wu L, Ji J, Feng J, Dai W, Li J, et al. Gut microbiota, peroxisome proliferator-activated receptors, and hepatocellular carcinoma. J Hepatocell Carcinoma. (2020) 7:271–88. doi: 10.2147/JHC.S277870
100. Wu L, Li J, Feng J, Ji J, Yu Q, Li Y, et al. Crosstalk between PPARs and gut microbiota in NAFLD. Biomed Pharmacother. (2021) 136:111255. doi: 10.1016/j.biopha.2021.111255
101. Byndloss M, Olsan E, Rivera-Chávez F, Tiffany C, Cevallos S, Lokken K, et al. Microbiota-activated PPAR-γ signaling inhibits dysbiotic Enterobacteriaceae expansion. Science. (2017) 357:570–5. doi: 10.1126/science.aam9949
102. Tomas J, Mulet C, Saffarian A, Cavin J, Ducroc R, Regnault B, et al. High-fat diet modifies the PPAR-γ pathway leading to disruption of microbial and physiological ecosystem in murine small intestine. Proc Natl Acad Sci USA. (2016) 113:E5934–43. doi: 10.1073/pnas.1612559113
103. Jalali M, Rahimlou M, Mahmoodi M, Moosavian S, Symonds M, Jalali R, et al. The effects of metformin administration on liver enzymes and body composition in non-diabetic patients with non-alcoholic fatty liver disease and/or non-alcoholic steatohepatitis: an up-to date systematic review and meta-analysis of randomized controlled trials. Pharmacol Res. (2020) 159:104799. doi: 10.1016/j.phrs.2020.104799
104. Komorizono Y, Hosoyamada K, Imamura N, Kajiya S, Hashiguchi Y, Ueyama N, et al. Metformin dose increase versus added linagliptin in non-alcoholic fatty liver disease and type 2 diabetes: an analysis of the J-LINK study. Diabetes Obes Metab. (2021) 23:832–7. doi: 10.1111/dom.14263
105. Wu H, Esteve E, Tremaroli V, Khan M, Caesar R, Mannerås-Holm L, et al. Metformin alters the gut microbiome of individuals with treatment-naive type 2 diabetes, contributing to the therapeutic effects of the drug. Nat Med. (2017) 23:850–8.
106. Armstrong M, Hull D, Guo K, Barton D, Hazlehurst J, Gathercole L, et al. Glucagon-like peptide 1 decreases lipotoxicity in non-alcoholic steatohepatitis. J Hepatol. (2016) 64:399–408. doi: 10.1016/j.jhep.2015.08.038
107. Newsome P, Buchholtz K, Cusi K, Linder M, Okanoue T, Ratziu V, et al. A placebo-controlled trial of subcutaneous semaglutide in nonalcoholic steatohepatitis. N Engl J Med. (2021) 384:1113–24. doi: 10.1056/NEJMoa2028395
108. Hartman M, Sanyal A, Loomba R, Wilson J, Nikooienejad A, Bray R, et al. Effects of novel dual GIP and GLP-1 receptor agonist tirzepatide on biomarkers of nonalcoholic steatohepatitis in patients with type 2 diabetes. Diabetes Care. (2020) 43:1352–5. doi: 10.2337/dc19-1892
109. Kuchay M, Krishan S, Mishra S, Choudhary N, Singh M, Wasir J, et al. Effect of dulaglutide on liver fat in patients with type 2 diabetes and NAFLD: randomised controlled trial (D-LIFT trial). Diabetologia. (2020) 63:2434–45. doi: 10.1007/s00125-020-05265-7
110. Kim E, Park J, Kim J, Oh J, Oh I, Choi D, et al. A GLP-1/GLP-2 receptor dual agonist to treat NASH: targeting the gut-liver axis and microbiome. Hepatology. (2022) 75:1523–38. doi: 10.1002/hep.32235
111. Mishima E, Fukuda S, Kanemitsu Y, Saigusa D, Mukawa C, Asaji K, et al. Canagliflozin reduces plasma uremic toxins and alters the intestinal microbiota composition in a chronic kidney disease mouse model. Am J Physiol Renal Physiol. (2018) 315:F824–33. doi: 10.1152/ajprenal.00314.2017
112. Ho H, Kikuchi K, Oikawa D, Watanabe S, Kanemitsu Y, Saigusa D, et al. SGLT-1-specific inhibition ameliorates renal failure and alters the gut microbial community in mice with adenine-induced renal failure. Physiol Rep. (2021) 9:e15092. doi: 10.14814/phy2.15092
113. Anhê F, Zlitni S, Zhang S, Choi B, Chen C, Foley K, et al. Human gut microbiota after bariatric surgery alters intestinal morphology and glucose absorption in mice independently of obesity. Gut. (2022) 72:460–71. doi: 10.1136/gutjnl-2022-328185
114. Sajjad A, Mottershead M, Syn W, Jones R, Smith S, Nwokolo C. Ciprofloxacin suppresses bacterial overgrowth, increases fasting insulin but does not correct low acylated ghrelin concentration in non-alcoholic steatohepatitis. Aliment Pharmacol Ther. (2005) 22:291–9. doi: 10.1111/j.1365-2036.2005.02562.x
115. Tang W, Wang Z, Levison B, Koeth R, Britt E, Fu X, et al. Intestinal microbial metabolism of phosphatidylcholine and cardiovascular risk. N Engl J Med. (2013) 368:1575–84. doi: 10.1056/NEJMoa1109400
116. Ponziani F, Zocco M, D’Aversa F, Pompili M, Gasbarrini A. Eubiotic properties of rifaximin: disruption of the traditional concepts in gut microbiota modulation. World J Gastroenterol. (2017) 23:4491–9. doi: 10.3748/wjg.v23.i25.4491
117. Abdel-Razik A, Mousa N, Shabana W, Refaey M, Elzehery R, Elhelaly R, et al. Rifaximin in nonalcoholic fatty liver disease: hit multiple targets with a single shot. Eur J Gastroenterol Hepatol. (2018) 30:1237–46. doi: 10.1097/MEG.0000000000001232
118. Gangarapu V, Ince A, Baysal B, Kayar Y, Kılıç U, Gök Ö, et al. Efficacy of rifaximin on circulating endotoxins and cytokines in patients with nonalcoholic fatty liver disease. Eur J Gastroenterol Hepatol. (2015) 27:840–5. doi: 10.1097/MEG.0000000000000348
119. European Association for the Study of the Liver. Electronic address:ZWFzbG9mZmljZUBlYXNsb2ZmaWNlLmV1LA== European Association for the Study of the Liver. EASL clinical practice guidelines on the management of hepatic encephalopathy. J Hepatol. (2022) 77:807–24.
120. Patel V, Lee S, McPhail M, Da Silva K, Guilly S, Zamalloa A, et al. Rifaximin-α reduces gut-derived inflammation and mucin degradation in cirrhosis and encephalopathy: RIFSYS randomised controlled trial. J Hepatol. (2022) 76:332–42. doi: 10.1016/j.jhep.2021.09.010
121. Jin Y, Ren X, Li G, Li Y, Zhang L, Wang H, et al. Beneficial effects of rifaximin in post-infectious irritable bowel syndrome mouse model beyond gut microbiota. J Gastroenterol Hepatol. (2018) 33:443–52. doi: 10.1111/jgh.13841
122. Fujinaga Y, Kawaratani H, Kaya D, Tsuji Y, Ozutsumi T, Furukawa M, et al. Effective combination therapy of angiotensin-II receptor blocker and rifaximin for hepatic fibrosis in rat model of nonalcoholic steatohepatitis. Int J Mol Sci. (2020) 21:5589. doi: 10.3390/ijms21155589
123. Everard A, Belzer C, Geurts L, Ouwerkerk J, Druart C, Bindels L, et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc Natl Acad Sci USA. (2013) 110:9066–71. doi: 10.1073/pnas.1219451110
124. Jian J, Nie M, Xiang B, Qian H, Yin C, Zhang X, et al. Rifaximin ameliorates non-alcoholic steatohepatitis in mice through regulating gut microbiome-related bile acids. Front Pharmacol. (2022) 13:841132. doi: 10.3389/fphar.2022.841132
125. Enomoto M, Kaji K, Nishimura N, Fujimoto Y, Murata K, Takeda S, et al. Rifaximin and lubiprostone mitigate liver fibrosis development by repairing gut barrier function in diet-induced rat steatohepatitis. Dig Liver Dis. (2022) 54:1392–402. doi: 10.1016/j.dld.2022.04.012
126. Tang Y, Huang J, Zhang W, Qin S, Yang Y, Ren H, et al. Effects of probiotics on nonalcoholic fatty liver disease: a systematic review and meta-analysis. Ther Adv Gastroenterol. (2019) 12:1756284819878046. doi: 10.1177/1756284819878046
127. Xiao M, Lin S, Shen Z, Luo W, Wang X. Systematic review with meta-analysis: the effects of probiotics in nonalcoholic fatty liver disease. Gastroenterol Res Pract. (2019) 2019:1484598. doi: 10.1155/2019/1484598
128. El Enshasy H, Malik K, Malek R, Othman N, Elsayed E, Wadaan M. Anaerobic probiotics: the key microbes for human health. Adv Biochem Eng Biotechnol. (2016) 156:397–431. doi: 10.1007/10_2015_5008
129. Van Immerseel F, Ducatelle R, De Vos M, Boon N, Van De Wiele T, Verbeke K, et al. Butyric acid-producing anaerobic bacteria as a novel probiotic treatment approach for inflammatory bowel disease. J Med Microbiol. (2010) 59(Pt 2):141–3. doi: 10.1099/jmm.0.017541-0
130. Yassour M, Lim M, Yun H, Tickle T, Sung J, Song Y, et al. Sub-clinical detection of gut microbial biomarkers of obesity and type 2 diabetes. Genome Med. (2016) 8:17. doi: 10.1186/s13073-016-0271-6
131. Depommier C, Everard A, Druart C, Plovier H, Van Hul M, Vieira-Silva S, et al. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: a proof-of-concept exploratory study. Nat Med. (2019) 25:1096–103. doi: 10.1038/s41591-019-0495-2
132. Bhutiani N, Schucht J, Miller K, McClave S. Technical aspects of fecal microbial transplantation (FMT). Curr Gastroenterol Rep. (2018) 20:30. doi: 10.1007/s11894-018-0636-7
133. Ianiro G, Segal J, Mullish B, Quraishi M, Porcari S, Fabiani G, et al. Fecal microbiota transplantation in gastrointestinal and extraintestinal disorders. Future Microbiol. (2020) 15:1173–83. doi: 10.2217/fmb-2020-0061
134. Paratore M, Santopaolo F, Cammarota G, Pompili M, Gasbarrini A, Ponziani F. Fecal microbiota transplantation in patients with HBV infection or other chronic liver diseases: update on current knowledge and future perspectives. J Clin Med. (2021) 10:2605. doi: 10.3390/jcm10122605
135. Witjes J, Smits L, Pekmez C, Prodan A, Meijnikman A, Troelstra M, et al. Donor fecal microbiota transplantation alters gut microbiota and metabolites in obese individuals with steatohepatitis. Hepatol Commun. (2020) 4:1578–90. doi: 10.1002/hep4.1601
136. Vrieze A, Van Nood E, Holleman F, Salojärvi J, Kootte R, Bartelsman J, et al. Transfer of intestinal microbiota from lean donors increases insulin sensitivity in individuals with metabolic syndrome. Gastroenterology. (2012) 143:913–6.e7. doi: 10.1053/j.gastro.2012.06.031
137. Ng S, Xu Z, Mak J, Yang K, Liu Q, Zuo T, et al. Microbiota engraftment after faecal microbiota transplantation in obese subjects with type 2 diabetes: a 24-week, double-blind, randomised controlled trial. Gut. (2022) 71:716–23.
138. Xue L, Deng Z, Luo W, He X, Chen Y. Effect of fecal microbiota transplantation on non-alcoholic fatty liver disease: a randomized clinical trial. Front Cell Infect Microbiol. (2022) 12:759306. doi: 10.3389/fcimb.2022.759306
139. Chu H, Duan Y, Yang L, Schnabl B. Small metabolites, possible big changes: a microbiota-centered view of non-alcoholic fatty liver disease. Gut. (2019) 68:359–70. doi: 10.1136/gutjnl-2018-316307
140. Sayin S, Wahlström A, Felin J, Jäntti S, Marschall H, Bamberg K, et al. Gut microbiota regulates bile acid metabolism by reducing the levels of tauro-beta-muricholic acid, a naturally occurring FXR antagonist. Cell Metab. (2013) 17:225–35. doi: 10.1016/j.cmet.2013.01.003
141. Nobili V, Alisi A, Mosca A, Della Corte C, Veraldi S, De Vito R, et al. Hepatic farnesoid X receptor protein level and circulating fibroblast growth factor 19 concentration in children with NAFLD. Liver Int. (2018) 38:342–9. doi: 10.1111/liv.13531
142. Ponziani F, Nesci A, Caputo C, Salvatore L, Picca A, Del Chierico F, et al. High prevalence of lower limb atherosclerosis is linked with the gut-liver axis in patients with primary biliary cholangitis. Liver Int. (2022) 43:370–80. doi: 10.1111/liv.15463
143. Jiao N, Baker S, Chapa-Rodriguez A, Liu W, Nugent C, Tsompana M, et al. Suppressed hepatic bile acid signalling despite elevated production of primary and secondary bile acids in NAFLD. Gut. (2018) 67:1881–91. doi: 10.1136/gutjnl-2017-314307
144. Ferslew B, Xie G, Johnston C, Su M, Stewart P, Jia W, et al. Altered bile acid metabolome in patients with nonalcoholic steatohepatitis. Dig Dis Sci. (2015) 60:3318–28. doi: 10.1007/s10620-015-3776-8
145. Kalhan S, Guo L, Edmison J, Dasarathy S, McCullough A, Hanson R, et al. Plasma metabolomic profile in nonalcoholic fatty liver disease. Metabolism. (2011) 60:404–13. doi: 10.1016/j.metabol.2010.03.006
146. Wang W, Zhao J, Gui W, Sun D, Dai H, Xiao L, et al. Tauroursodeoxycholic acid inhibits intestinal inflammation and barrier disruption in mice with non-alcoholic fatty liver disease. Br J Pharmacol. (2018) 175:469–84. doi: 10.1111/bph.14095
147. Younossi Z, Ratziu V, Loomba R, Rinella M, Anstee Q, Goodman Z, et al. Obeticholic acid for the treatment of non-alcoholic steatohepatitis: interim analysis from a multicentre, randomised, placebo-controlled phase 3 trial. Lancet. (2019) 394:2184–96.
148. Harrison S, Bashir M, Lee K, Shim-Lopez J, Lee J, Wagner B, et al. A structurally optimized FXR agonist, MET409, reduced liver fat content over 12 weeks in patients with non-alcoholic steatohepatitis. J Hepatol. (2021) 75:25–33. doi: 10.1016/j.jhep.2021.01.047
Keywords: MAFLD, NAFLD, gut microbiota, metabolomics, diabetes, diet, bariatric surgery, probiotics
Citation: Maestri M, Santopaolo F, Pompili M, Gasbarrini A and Ponziani FR (2023) Gut microbiota modulation in patients with non-alcoholic fatty liver disease: Effects of current treatments and future strategies. Front. Nutr. 10:1110536. doi: 10.3389/fnut.2023.1110536
Received: 28 November 2022; Accepted: 16 January 2023;
Published: 16 February 2023.
Edited by:
Enrico Dainese, University of Teramo, ItalyReviewed by:
Tien Sy Dong, UCLA Health System, United StatesCopyright © 2023 Maestri, Santopaolo, Pompili, Gasbarrini and Ponziani. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Francesca Romana Ponziani,  ZnJhbmNlc2NhLnBvbnppYW5pQGdtYWlsLmNvbQ==
ZnJhbmNlc2NhLnBvbnppYW5pQGdtYWlsLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.