
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Nutr. , 12 October 2022
Sec. Nutrition and Food Science Technology
Volume 9 - 2022 | https://doi.org/10.3389/fnut.2022.993744
This article is part of the Research Topic Food-derived Bioactive Peptides and its Application in Treatment of Chronic Diseases View all 5 articles
 Xinchang Gao1,2
Xinchang Gao1,2 Fan Bu3Dalong Yi4Huaigao Liu4Zhiying Hou2Chaoying Zhang1Chang Wang1Jin-Ming Lin1
Fan Bu3Dalong Yi4Huaigao Liu4Zhiying Hou2Chaoying Zhang1Chang Wang1Jin-Ming Lin1 Yali Dang3*
Yali Dang3* Yufen Zhao1,2*
Yufen Zhao1,2*A novel angiotensin-converting enzyme (ACE) inhibitory peptide ser-ala-ser-val-ile-pro-val-ser-ala-val-arg-ala (SASVIPVSAVRA) was purified and identified from yak bone by Electrospray Ionization-Time of Flight-Mass Spectrometry (ESI-TOF-MS). Results in vitro showed that the peptide exhibited strong ACE inhibition activities with an IC50 of 54.22 μM. Molecular docking results showed the binding between the peptide SASVIPVSAVRA and ACE mainly driven by van der Waals forces, hydrogen bonds and metal receptor. Interestingly, the ACE inhibition activities of the peptide increased about 19% after digestion, but none of its metabolites showed stronger activity than it. The in vivo experiment showed that the antihypertensive effect of peptide SASVIPVSAVRA at dose of 30 mg/kg is nearly equal to Captopril at dose of 10 mg/kg to spontaneously hypertensive rats (SHRs). The antihypertensive effect mechanism of SASVIPVSAVRA should be further studied through plasma metabolomics and bioanalysis. Structure analysis of amino acids and peptides produced during digestion may help better understand the antihypertensive effect of peptides.
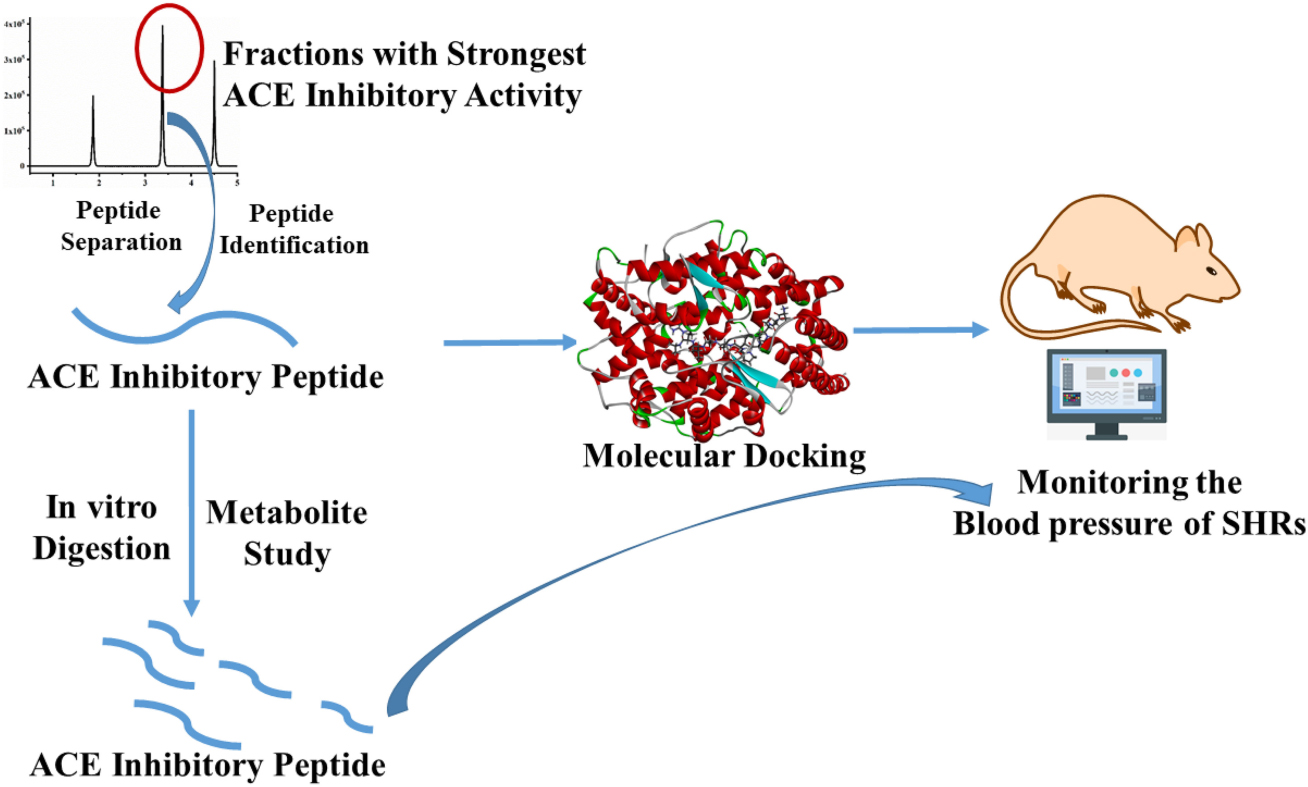
GRAPHICAL ABSTRACT. Study on the peptide ser-ala-ser-val-ile-pro-val-ser-ala-val-arg-ala (SASVIPVSAVRA) in vitro and in vivo.
Hypertension is the most common chronic disease and a significant threat to cerebrovascular diseases. It usually causes headaches, inattention, memory loss, chest tightness, fatigue, and other symptoms (1).
With the continuous increase in the number of people suffering from hypertension, researchers have paid more attention to the prevention, and the treatment of hypertension. The renin-angiotensin system (RAS) and kallikrein-kinin system (KKS) play an important role in regulating blood pressure in vivo (2). In RAS, Renin cleaves angiotensinogen to produce the inactive Angiotensin I, and through the action of ACE, which generated Angiotensin II with vascular smooth muscle activity, thereby causing the blood pressure spiked. In KKS, ACE deactivates bradykinin, and bradykinin stabilizes blood pressure by relaxation blood vessels and regulating electrolytes. Unfortunately, synthetic ACE inhibitors (captopril and enalapril) occasionally cause some adverse reactions, such as coughing, allergic reactions, and elevated blood potassium levels (3). More and more attention has been focused on seeking food-derived ACE inhibitory peptides that could lower blood pressure and reduce side effects in recent years. Interestingly, more and more reports appeared about the natural polypeptides with potential ACE inhibitory activity in vitro and in vivo, and they are structurally similar to synthetic drugs (4). These ACE inhibitors are safe without side effects.
Animal bones are an important source of functional peptide (5, 6). Researchers found many food-derived antihypertensive peptides in animals and plants, including fungi, milk, yeast, cheese, fish, and soybeans (7, 8). The ACE inhibitory activities of peptides are nearly related to their unique amino acid composition, such as Pro, Ala, and Val (9). Interestingly, yak bones are also rich in these unique amino acids. Chen et al. (10) reported that yak bones were rich in glycine, alanine, proline and glutamic acid (10). However, there are few reports on the isolation of ACE inhibitory peptide sequences from yak bones. Sun et al. (11) reported an antioxidant peptide GPHGAAGVA from yak bone. Ye et al. (12) reported an peptide GPSGPAGKDGRIGQPG from Yak bone with osteoblast-proliferating activity.
This study aims to purify and identified the bioactive peptides from Yak bone, evaluate the antihypertensive effects in vivo, explain the mechanism between peptide and ACE.
Yak bone was provided by Anhui Guotai Biotechnology Co., Ltd., Papain (800 U/g proteins), pepsin (25,000 U/mL), pancreatin (800 U/mL), ACE, Hippuryl-histidyl-leucine (HHL), BCA Protein Assay Kit and Sephadex G-25 were purchased from Sigma-Aldrich (St. Louis, MO, USA). All the reagents were of analytical grade.
One hundred and fifty gram of yak bone powder was dispersed in distilled water with the mass ratio of 1:30, and the content of yak bone protein was 197 g/kg. The hydrolyzed were mixed with papain (800 U/g proteins) at 60°C, 80 r/min for 6 h. After inactiving the enzyme by boiling for 10 min, The mixture was filtered and lyophilized (13).
The method of Separation and Purification was according to the method of Bu et al. (14) with minor modification (14). The lyophilized samples of the hydrolysate product were dissolved in distilled water to reach 20 mg/mL concentration.
ESI-TOF-MS was used for the identification of peptides from HPLC Fractions as described by Dang et al. (15). Electrospray ionization (ESI) was used in positive ion mode, source temperature was set at 150°C, desolvation temperature was set at 550°C, scan range was m/z 50–1200. The most active fraction (F6) was resuspended in water at a concentration of 1 mg/mL. The filtrate was then analyzed with the ESI-TOF mass spectrometer of the ACQUITY UPLC H-Class HPLC system (Waters, USA). The column used was BEH300 C18 (2.1 × 100 mm; 1.7 μm) (Waters, USA), the flow rate was 0.2 mL/min, and the injection volume was 10 μL. All MS and MS/MS spectra were collected and analyzed using Mass Lynx (Waters version 4.1). The PepSeq program of Biolynx (Waters Corp.) software was used for sequencing. These peptides were custom-synthesized by Sangon Biotech Co., Ltd., (Shanghai, China) (16).
The purity of these peptides was above 99%.
The original ACE Inhibitory Activity method was developed by Cushing and Cheung (17). The ACE inhibitory activity assay used here were performed according to the method of Dang et al. (15). 80 μL of 5 mM HHL (in HEPES buffer containing 300 mM NaCl at pH 8.3) and 30 μL samples were mixed in 37°C water bath for 5 min, 40 μL ACE (0.025 U/mL) was added for incubation for 1 h, The reaction was stopped by adding 250 μL of 1 M HCl. they were filtered and measured the ACE inhibitory activity by an Agilent 1260 HPLC system (Agilent Technologies, Karlsruhe, Germany).
Gastrointestinal digestion was evaluated according to the method of Dang et al. (15) and Mulet-Cabero et al. (18) with minor modification. Simulated gastric juice (pH 2.0) and simulated intestinal fluid mixture (pH 7.6) were added to peptide solution and incubated at 37°C for 2 h, respectively.
The systolic blood pressure (SBP) changes in the SHRs were measured by the tail-cuff method to determine the antihypertensive effect of ser-ala-ser-val-ile-pro-val-ser-ala-val-arg-ala (SASVIPVSAVRA) in vivo. Twenty female SHRs, weighing 230 ± 15 g and the SBP exceeding 190 mmHg, were obtained from Beijing Vital River Laboratory Animal Technology Co., Ltd. (China).
The SHRs were free to eat and drink at 25 ± 4°C and a 40 ± 6% humidity. To get the rats to adapt to the environment, the systolic blood pressure was measured once a day for 1 week. After a week of adaptation, the SHRs were randomly divided into three groups with each group containing 6 SHRs: the control group (sterilized water), captopril group (10 mg/kg captopril BW), SASVIPVSAVRA (30 mg/kg SASVIPVSAVRA BW). The effect of ACE inhibitors on SBP were measured by the Softron BP non-invasive blood pressure monitor (Softron BP-2000, Tokyo, Japan) at 0, 2, 4, and 6 h after oral administration (1), The measurements were repeated five times, the recordings were reported as mean ± standard deviation (SD). All animal procedures used in this trial were approved by the Institutional Animal Care and Use Committee of Hangzhou Medical College (Approval No. ZJCLA-IACUC-20010014).
The ACE inhibition pattern was similar to that used by Zhang et al. (19) with slight modifications. The concentration of HHL was changed to 0.5, 1, and 2 mmol/L, and the concentration of the purified peptide was changed to 0.1 and 0.2 mg/mL.
The molecular docking was performed using the Discovery Studio software package (Neo Trident Technology Ltd., China) according to the method of Chen et al. (20) with some modifications. The crystal structure of the human ACE-lisinopril complex (1O8A.pdb) was from RCSB PDB Protein Data Bank.1 SBD_Site_Sphere was X: 37.8546, Y: 38.1941, Z: 47.2396 and the docking radius was 20 Å.
All data were analyzed using SPSS for Windows Version 25.0. The results were based on the mean ± standard deviation of at least three determinations. A P-value of less than 0.05 was considered statistically significant.
Results showed that the yak bone peptide prepared by section “Preparation of yak bone peptide” had ACE inhibitory activity and its IC50 was 1.47 mg/mL. So the ACE fractions were further separated and purified. Four components of Yak bone peptide were separated by Sephadex G-25 column (Figure 1A). G2 had the highest ACE inhibitory activity with IC50 value of 0.181 mg/mL, while IC50 values of other fractions G1, G3, and G4 were 0.258, 0.459, and 0.563 mg/mL, respectively. So, G2 was chosen for further purification, and the peptides were separated using RP-HPLC (Figure 1B). Finally, the 7 fractions were collected, and F6 had the highest ACE inhibitory activity among them(IC50 = 0.12 mg/mL). So F6 was collected for further experiment.
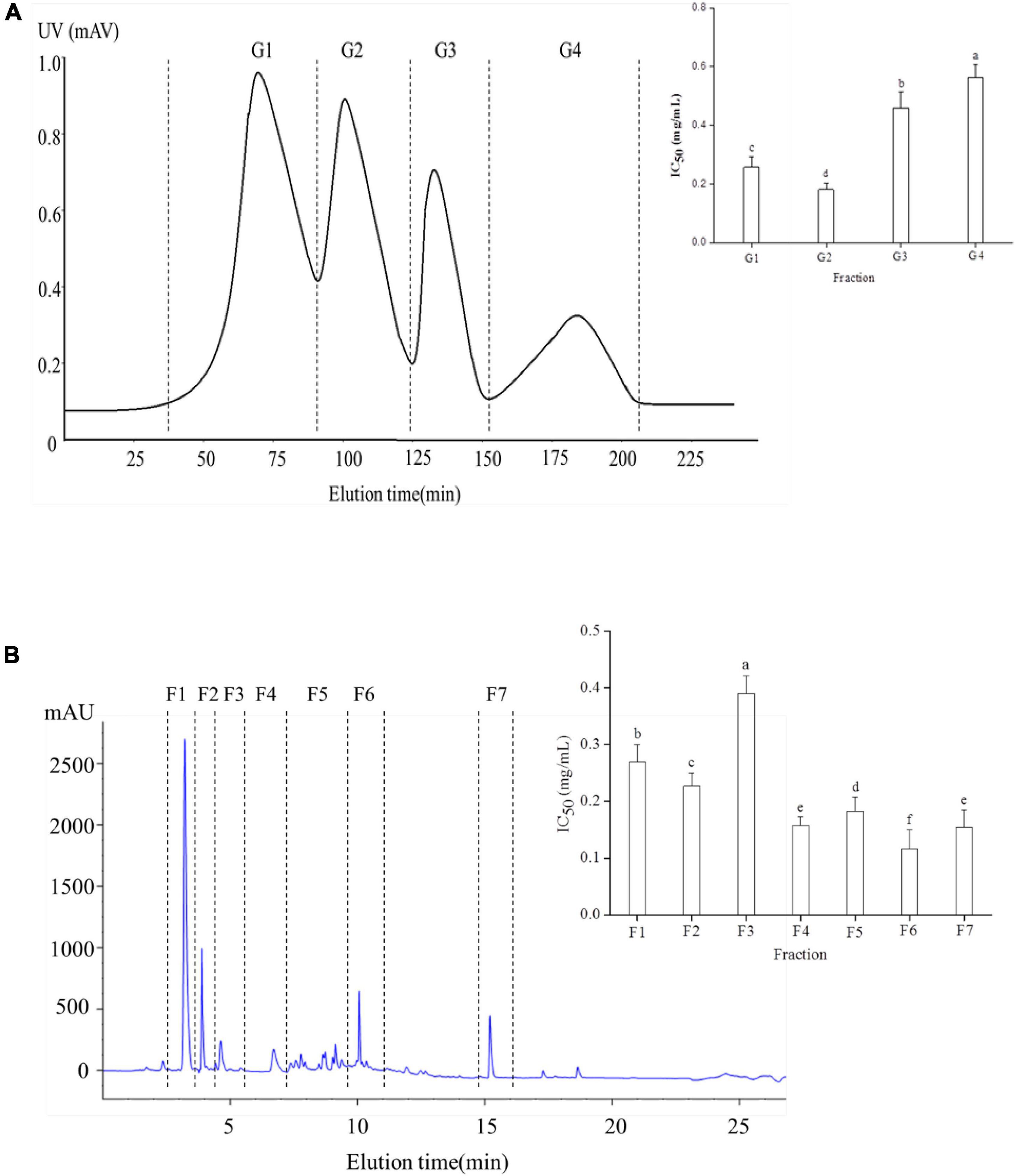
Figure 1. Separation and purification of a novel ACE inhibitory peptide. (A) Sephadex G-25 column chromatography separation diagram of Yak bone hydrolysate. (B) The fraction G2 purified using RP-HPLC. Different letters indicate significant differences in peptide activity (p < 0.05).
Peptides from F6 was identified by ESI-TOF-MS and synthesized by the Sangon Biotech Co., Ltd. The sequence was SASVIPVSAVRA, The IC50 of ACE inhibitory activity was 54.22 μM, The mass spectrum of this peptide was shown in Figures 2A,B.
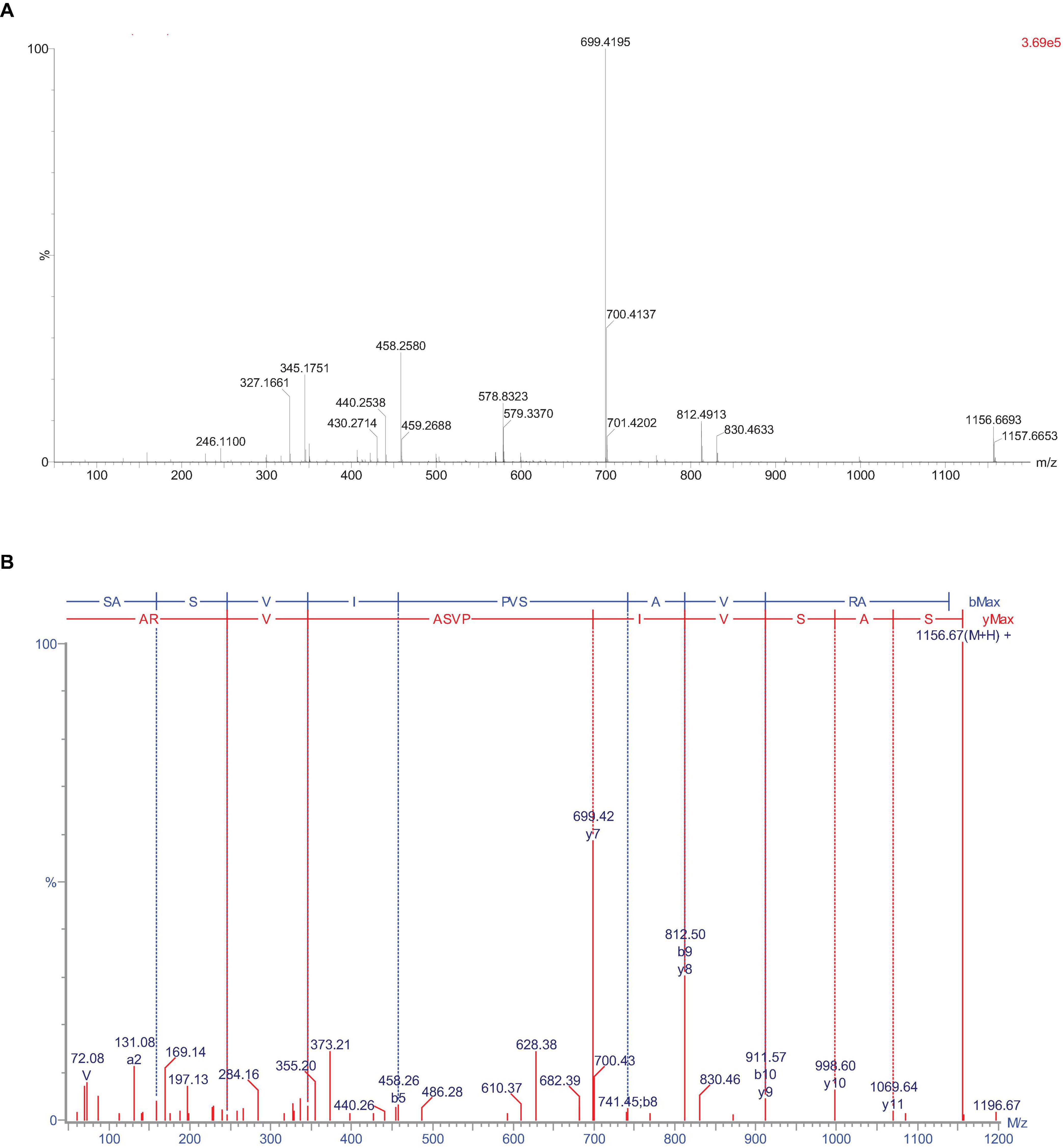
Figure 2. The mass spectrum of the peptide SASVIPVSAVRA. (A) MS spectrum (B) MS/MS spectrum.
It was found that the IC50 value of SASVIPVSAVRA (54.22 μM) was much higher than the reported ACE peptide SGGSYADELVSTAK (0.093 μM) (21) while lower than other peptides, such as QLLLQQ (IC50, 75.0 μM) and IVVE (IC50, 315.3 μM) (22, 23). Therefore, it was meaningful to study the ACE inhibitory activity of SASVIPVSAVRA.
The ACE inhibitory Activity of peptides was closely related to hydrophobic amino acid residues, such as Proline, Valine, Serine, Alanine, Leucine and Isoleucine (24). Therefore, the rich hydrophobic amino acids in the peptide SASVIPVSAVRA may be the main reason for the ACE inhibitory activity. The presence of proline in the sequence may allowed the carboxyl group to interact with the positively charged residues in the active site of the enzymec to contribute to its ACE activity (25).
Generally, the presence of amino acids (Val, Ile) with hydrophobic branches would increase the ACE inhibitory activity (26). Therefore, the Val residue and Ile residue in SASVIPVSAVRA may contribute to the ACE activity of the peptide. Besides, Matsumura et al. (27) reported that the tripeptide VIP extracted from Bonito Bowels Autolysate had strong ACE inhibitory activity. The peptide SASVIPVSAVRA isolated from the yak bone hydrolysate also contained the tripeptide above-mentioned (VIP). The ACE inhibitory activity may also be due to the presence of ACE inhibitor VIP in its structure. However, this peptide still needed further tests on SHRs to determine whether its activity remained in vivo. It was also necessary to determine the digestion stability and the activity of the peptide after digestion. Therefore, it was discussed about the blood pressure inhibitory effect of the peptide in vivo and the simulated digestion results as follows.
More and more evidence showed that some bioactive peptides were unstable after digestion (28). Therefore, it is also very important to evaluate the digestive stability of bioactive peptides.
The IC50 of SASVIPVSAVRA before and after digestion were 54.22 and 44.06 μM, respectively. The ACE inhibitory activity after digestion increased about 19%. SASVIPVSAVR, SASVIPVSA, SASVIPV, and AVR were found in the digestion sample (Figure 3). The IC50 were 102.3, 144.6, 167.6, 213.2 μM.
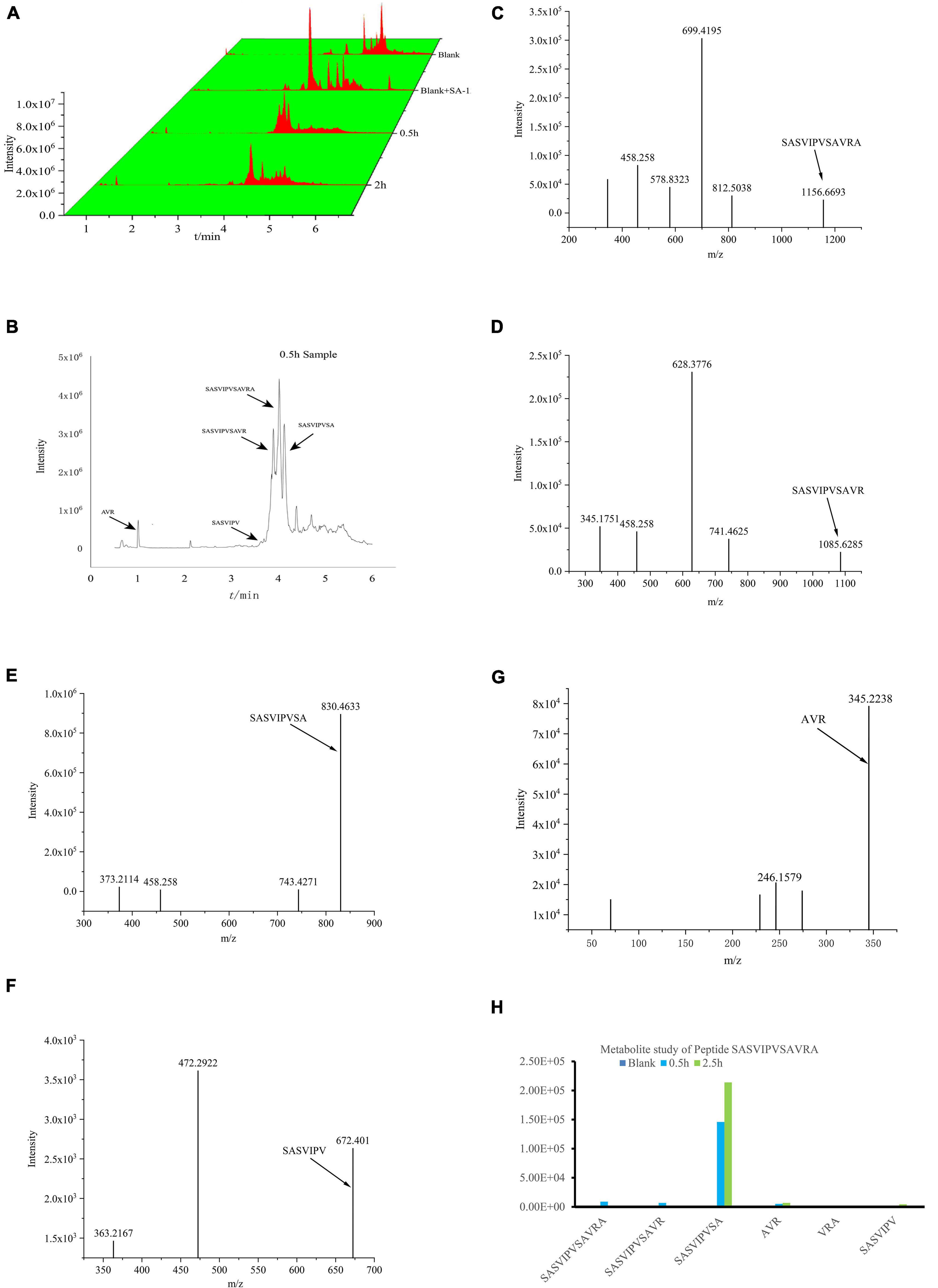
Figure 3. Analysis of peptides SASVIPVSAVRA after digestion. (A) 3D waterfall Tic chromatography of the samples. (B) Tic chromatography of the 0.5 h samples. (C) Ms spectrum of peptide SASVIPVSAVRA. (D) Ms spectrum of peptide SASVIPVSAVR. (E) Ms spectrum of peptide SASVIPVSA. (F) Ms spectrum of peptide SASVIPV. (G) Ms spectrum of peptide AVR. (H) Peak area of Metabolites in digestion samples.
There is no significant peak of SASVIPVSAVRA found in the digested samples, indicating SASVIPVSAVRA was easily digested by gastrointestinal tract. SASVIPVSA was the main metabolite. The reason for the increased ACE inhibitory activity after the peptide SASVIPVSAVRA digestion needs further investigation.
As reported by Katayama et al. (29), certain peptides had certain resistance to digestive proteases in vivo and could still show strong biological activity. In addition, Long chain peptides are more easily digested into many short peptides, and active peptides are more likely to appear (30). Therefore, in the future, we will further study the transport of peptide in human intestinal CaCo-2 cell monolayers to determine the absorption of the peptide in vitro.
Most peptides are in a competitive ACE inhibition pattern, while a small part of them are in a non-competitive ACE inhibition pattern (31). According to the Lineweaver-Burk plots, the ACE inhibition pattern of the peptide was estimated (Figure 4). The increased Km and constant Vmax indicated that SASVIPVSAVRA acted as a competitive ACE inhibitor. This suggested that SASVIPVSAVRA might competitively bind to the catalytic site of ACE to prevent ACE from binding to the substrate (32). Also, Cheung et al. (33) found those peptides containing hydrophobic C-terminal amino acids (such as alanine, phenylalanine and tyrosine) generally performed a competitive inhibition pattern. From our results, the peptide SASVIPVSAVRA had alanine residues at the C-terminal, respectively. This may offer an explanation of why SASVIPVSAVRA exhibited a competitive inhibitor.
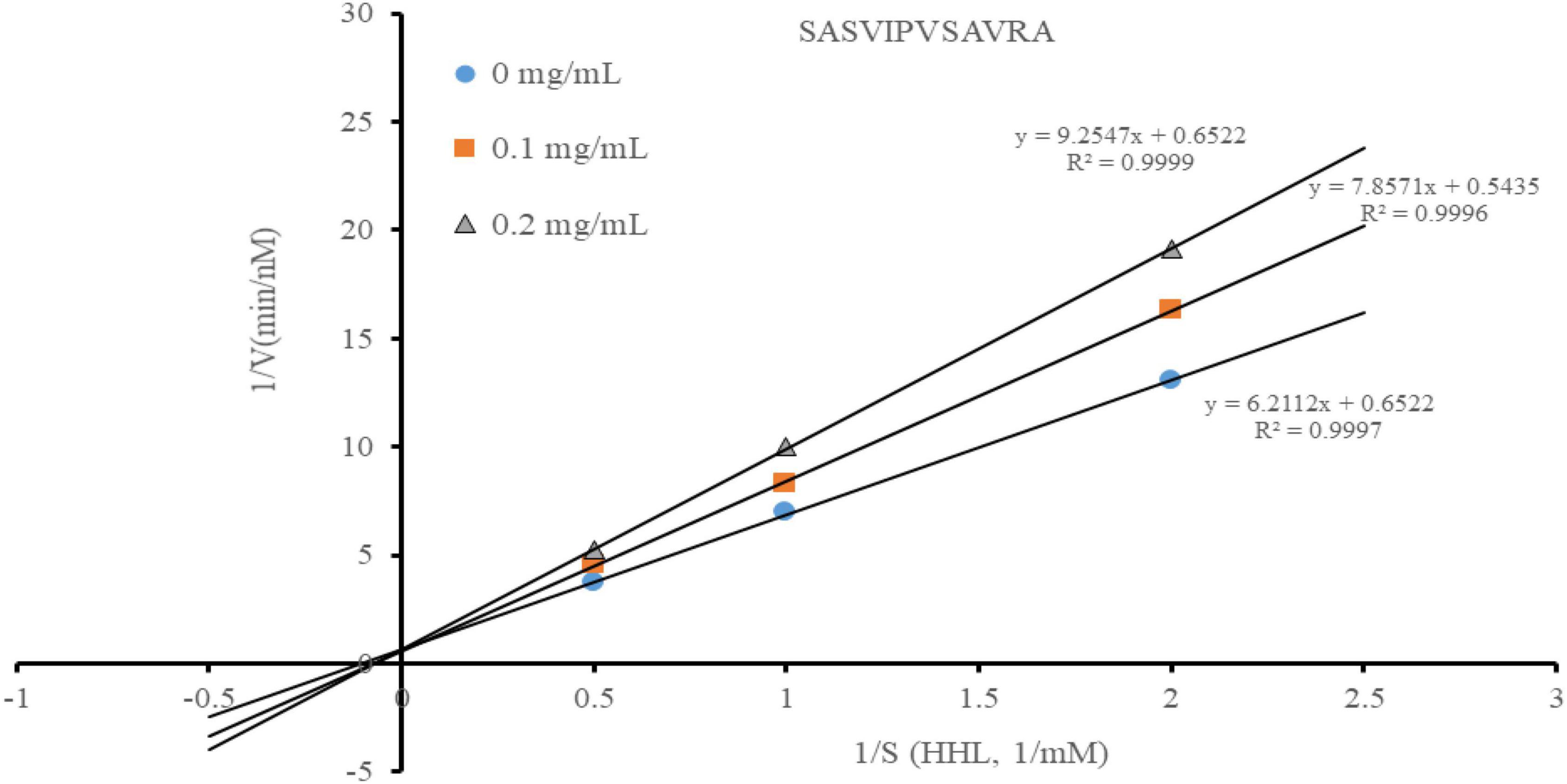
Figure 4. Line weaver-Burke plot of ACE activity in the presence of the peptide.
Molecular docking is to explore the binding interactions effect of peptide on ACE. Figure 5A showed the best posture for docking peptide SASVIPVSAVRA at the active site of ACE through docking simulation. The mutual binding energy was 134.935 kJ/mol. The docking structure Figure 5B showed that Van der Waals forces, hydrogen bonds and metal receptor were the major binding forces in the interaction between SASVIPVSAVRA and ACE.
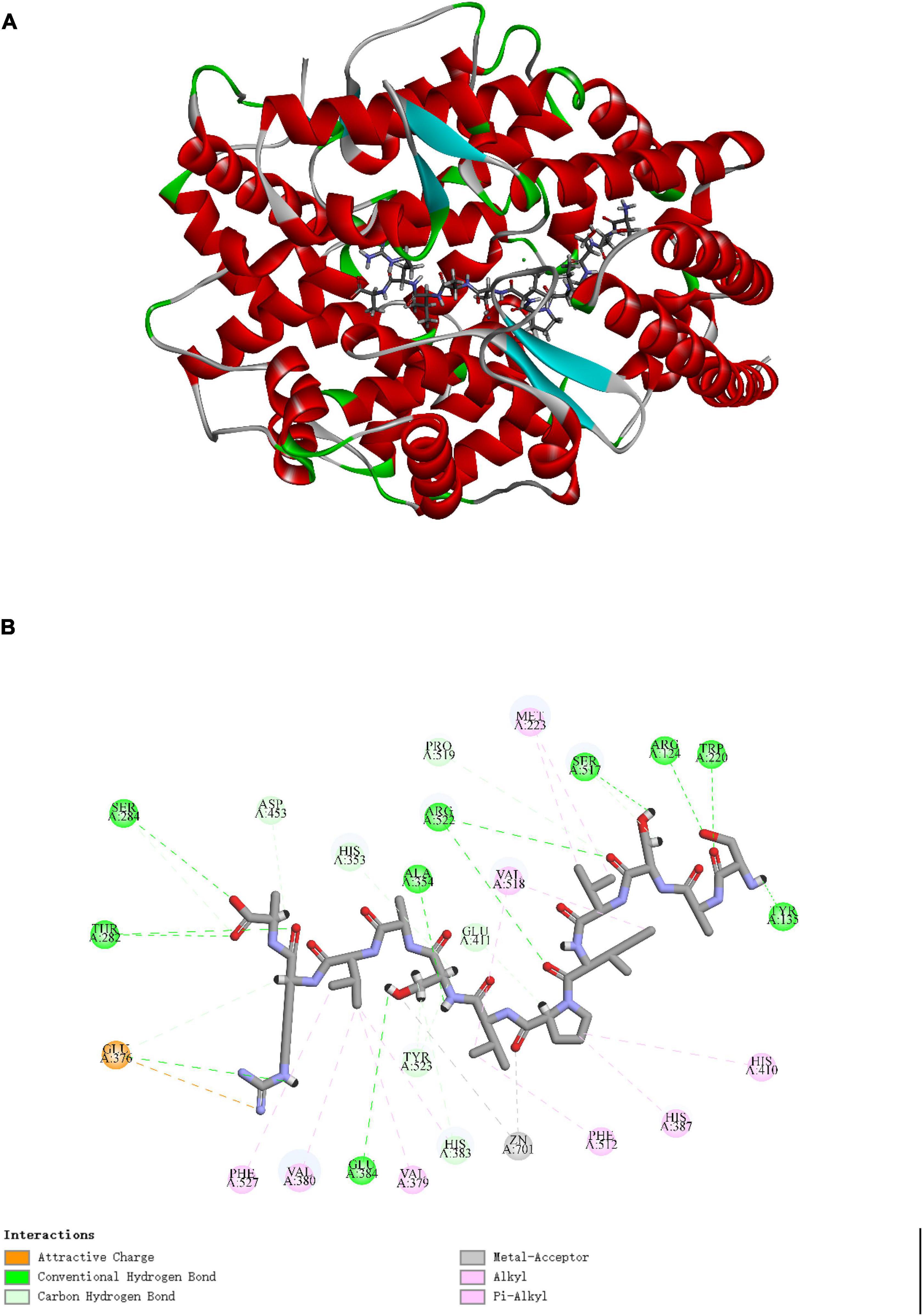
Figure 5. Diagram of SASVIPVSAVRA binding with ACE. (A) The optimal docking structure diagram of SASVIPVSAVRA binding with ACE (PDB: 1O8A). (B) The 2D diagram of the interaction between SASVIPVSAVRA and ACE amino acid residues.
The peptide existed in the ACE active site’s narrow channel and formed 9 hydrogen bonds with ACE. It indicated that the peptide had strong interaction forces with ACE (34). Specifically, the peptide established hydrogen bonds with ACE residues Arg124, Tyr135, Trp220, Thr282, Ser284, Ala354, Glu384, Ser517, and Arg522, which may be the key residues in ACE binding (35). The active center of ACE comprises zinc ions and three primary active capsules (S1, S2, S3). More studies have shown that high ACE inhibitory activity peptides can directly interact with the ACE Zn(II) (36). This result may offer an clarification of why SASVIPVSAVRA displayed high ACE inhibitory activity.
The antihypertensive effect of peptide SASVIPVSAVRA was evaluated by measuring the changes in SBP in the SHRS within 8 h after administration.
The systolic blood pressure of the negative control group (sterilized water) had no significant change after administration (Figure 6). The SASVIPVSAVRA group and the captopril group significantly reduced the SBP of SHRs (Figure 6). A significant antihypertensive effect had been observed within 8 h of administration of the SASVIPVSAVRA peptide. Antihypertensive activity peaked at the 6th hour after administration. When the oral dose was 30 mg/kg, the maximum reduction of systolic blood pressure in the sample group and the positive control group was 15.3 and 14.9%, respectively. It was shown that ACE peptide and captopril have similar antihypertensive effects in vivo, suggesting that ACE peptide and captopril were not directly proportional to their antihypertensive abilities in vitro and in vivo, This was consistent with the study by Cao et al. (1). Some chemically synthesized drugs have a substantial effect on lowering the blood pressure in vitro, but their effects in vivo will be weakened (37, 38). Remarkably, Captopril is a chemically designed and commercially used drug, The peptide SASVIPVSAVRA also had a blood pressure lowering effect in vivo, so it could provide a reference for the development of new blood pressure lowering drugs. Furthermore, the ACEI peptide IVGRPRHQG from chicken muscle showed similar antihypertensive effect as SASVIPVSAVRA, but required a higher dose of 60 mg/kg BW (39).
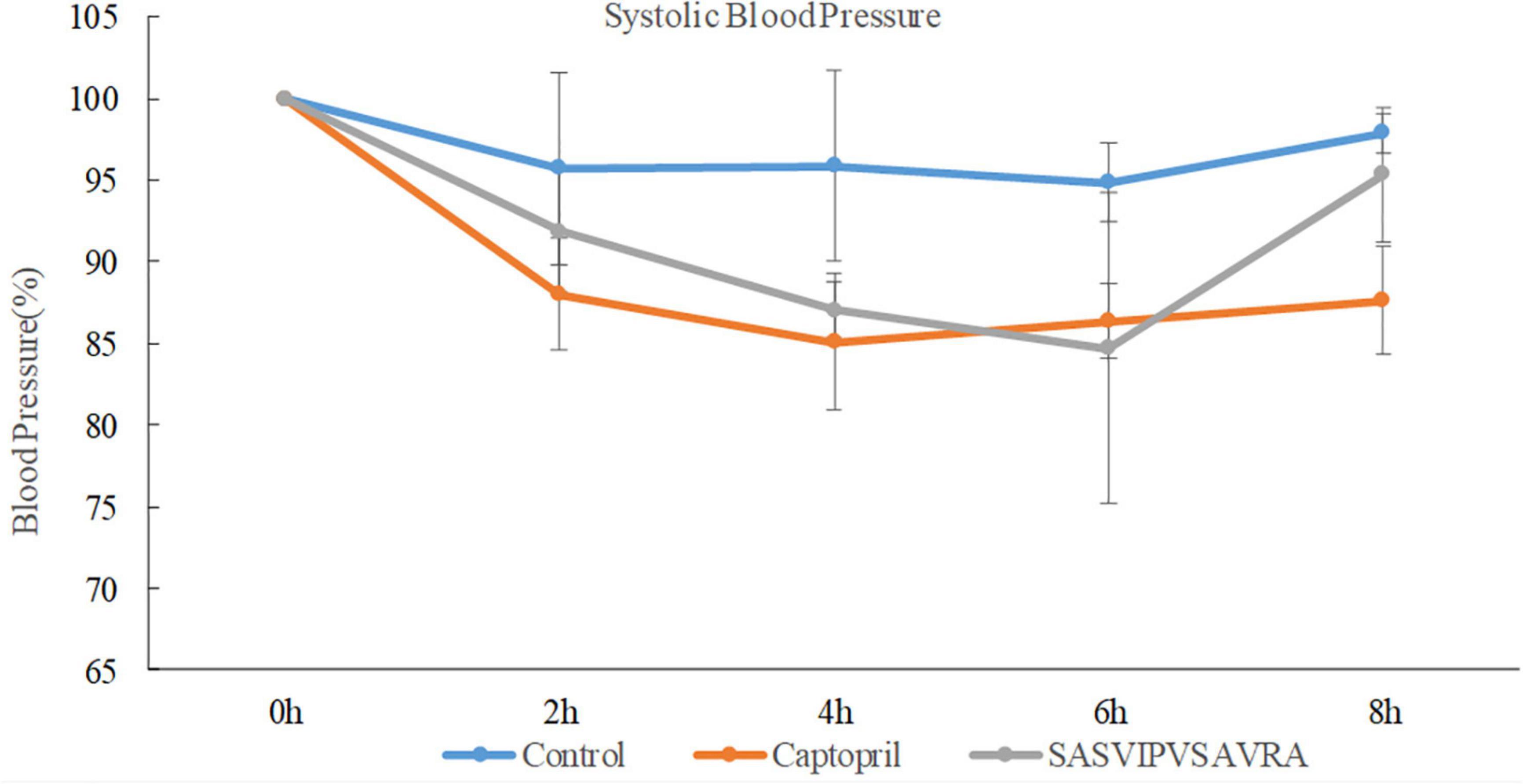
Figure 6. The effects of distilled water, captopril and SASVIPVSAVRA on systolic blood pressure of SHRs.
The peptide SASVIPVSAVRA had high ACE inhibitory activity both in vitro and in vivo. The ACE inhibitory activity increased about 19% after digestion, but none of its metabolites showed stronger activity than it, Some hydrophobic amino acids produced during this digestion are worth considering. The relationship between the metabolites of the peptide in blood and effect of antihypertensive effect in vivo should be deeply studied through the plasma metabonomics and bioanalysis.
A novel angiotensin converting enzyme (ACE) inhibitory peptide SASVIPVSAVRA was purified and identified from yak bone in this study. The results show that the peptide has strong ACE inhibitory activity in vitro and hypotensive effect in vivo, and has the potential to be developed into antihypertensive drugs.
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/supplementary material.
All animal procedures used in this trial were approved by the Institutional Animal Care and Use Committee of Hangzhou Medical College (Approval No. ZJCLA-IACUC-20010014).
YZ and YD contributed to the conception of the study. XG and FB performed the experiment. DY and HL contributed significantly to analysis and manuscript preparation. ZH, CZ, and CW performed the data analyses and wrote the manuscript. XG, J-ML, and YD helped perform the analysis with constructive discussions. All authors contributed to the article and approved the submitted version.
This work was supported by the Scientific Research Grant of Ningbo University (215-432000282), the Ningbo Top Talent Project (215-432094250), and the Zhejiang Provincial Administration for Market Regulation (CY202234).
Authors DY and HL were employed by Anhui Guotai Biotechnology Co., Ltd.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Cao S, Wang Y, Hao Y, Zhang W, Zhou G. Antihypertensive effects in vitro and in vivo of novel angiotensin-converting enzyme inhibitory peptides from bovine bone gelatin hydrolysate. J Agric Food Chem. (2020) 68:759–68. doi: 10.1021/acs.jafc.9b05618
2. Fitzgerald RJ, Murray BA, Walsh DJ. Hypotensive peptides from milk proteins. J Nutr. (2004) 134:980S–8S.
3. Liu C, Fang L, Min W, Liu J, Li HJ. Exploration of the molecular interactions between angiotensin-I-converting enzyme (ACE) and the inhibitory peptides derived from hazelnut (Corylus heterophylla Fisch.). Food Chem. (2018) 245:471–80. doi: 10.1016/j.foodchem.2017.10.095
4. Liao W, Fan H, Wu J. Egg white-derived antihypertensive peptide IRW (Ile-Arg-Trp) inhibits angiotensin II-stimulated migration of vascular smooth muscle cells via angiotensin type i receptor. J Agric Food Chem. (2018) 66:5133–8. doi: 10.1021/acs.jafc.8b00483
5. Bao Y, Yang H, Jing L, Li Z, Jiang YJ. Amino acid composition, molecular weight distribution and antioxidant activity of protein hydrolysates of soy sauce lees. Food Chem. (2010) 124:551–5.
6. Li F, Jia D, Yao K. Amino acid composition and functional properties of collagen polypeptide from Yak (Bos grunniens) bone. LWT Food Sci Technol. (2009) 42:945–9. doi: 10.1016/j.lwt.2008.12.005
7. Gallego M, Mora L, Science FT. Health relevance of antihypertensive peptides in foods. Curr Opin Food Sci. (2017) 19:8–14.
8. Shen S, Chahal B, Majumder K, You SJ, Wu J. Identification of novel antioxidative peptides derived from a thermolytic hydrolysate of ovotransferrin by LC-MS/MS. J Agric Food Chem. (2010) 58:7664–72. doi: 10.1021/jf101323y
9. Taga Y, Hayashida O, Ashour A, Amen Y, Kusubata M, Ogawa-Goto K, et al. Characterization of angiotensin-converting enzyme inhibitory activity of X-Hyp-Gly-type tripeptides: importance of collagen-specific prolyl hydroxylation. J Agric Food Chem. (2018) 66:8737–43. doi: 10.1021/acs.jafc.8b03648
10. Chen G, Cheng L, Xu H, Song H, Lv Y, Yang C, et al. Functions of different yak bone peptides. Int J Food Propert. (2011) 14:1136–41. doi: 10.1080/10942911003592753
11. Sun X, Wang K, Gao S, Hong H, Zhang L, Liu H, et al. Purification and characterization of antioxidant peptides from yak (Bos grunniens) bone hydrolysates and evaluation of cellular antioxidant activity. J Food Sci Technol. (2021) 58:3106–19. doi: 10.1007/s13197-020-04814-7
12. Ye M, Jia W, Zhang C, Shen Q, Zhu L, Wang L. Preparation, identification and molecular docking study of novel osteoblast proliferation-promoting peptides from yak (Bos grunniens) bones. RSC Adv. (2019) 9:14627–37. doi: 10.1039/c9ra00945k
13. Guo Z, Liu C, Hu B, Zhu L, Yang Y, Liu F, et al. Simulated gastrointestinal digestion of yak bone collagen hydrolysates and insights into its effects on gut microbiota composition in mice. Food Biosci. (2021) 44:101463. doi: 10.1016/j.fbio.2021.101463
14. Bu F, Yi D, Zhang H, Li Q, Pan D, Dang Y. Isolation and identification of antioxidant and DPP-IV inhibitory peptide PYPYEPYEPYPY from yak bone hydrolysate. Food Sci Technol Res. (2021) 27:441–52. doi: 10.3136/fstr.27.441
15. Dang Y, Zhou T, Hao L, Cao J, Sun Y, Pan D. In vitro and in vivo studies on the angiotensin-converting enzyme inhibitory activity peptides isolated from broccoli protein hydrolysate. J Agric Food Chem. (2019) 67:6757–64. doi: 10.1021/acs.jafc.9b01137
16. Dang Y, Gao X, Ma F, Wu X. Comparison of umami taste peptides in water-soluble extractions of Jinhua and Parma hams. LWT Food Sci Technol. (2015) 60:1179–86.
17. Cushman DW, Cheung HC. Spectrophotometric assay and properties of angiotensin-converting enzyme of rabbit lung. Biochem Pharmacol. (1971) 20:1637–48.
18. Mulet-Cabero AI, Egger L, Portmann R, Menard O, Marze S, Minekus M, et al. A standardised semi-dynamic in vitro digestion method suitable for food – an international consensus. Food Funct. (2020) 11:1702–20. doi: 10.1039/c9fo01293a
19. Zhang Y, Pan D, Yang Z, Gao X, Dang Y. Angiotensin I-Converting enzyme (ACE) inhibitory and dipeptidyl Peptidase-4 (DPP-IV) inhibitory activity of umami peptides from Ruditapes philippinarum. Lwt. (2021) 144:111265. doi: 10.1016/j.lwt.2021.111265
20. Chen M, Pan D, Zhou T, Gao X, Dang Y. Novel umami peptide IPIPATKT with dual dipeptidyl peptidase-IV and angiotensin I-converting enzyme inhibitory activities. J Agric Food Chem. (2021) 69:5463–70. doi: 10.1021/acs.jafc.0c07138
21. Karami Z, Peighambardoust SH, Hesari J, Akbari-Adergani B, Andreu D. Antioxidant, anticancer and ACE-inhibitory activities of bioactive peptides from wheat germ protein hydrolysates. Food Biosci. (2019) 32:100450. doi: 10.1016/j.fbio.2019.100450
22. Suetsuna K, Chen JR. Identification of antihypertensive peptides from peptic digest of two microalgae, Chlorella vulgaris and Spirulina platensis. Mar Biotechnol. (2001) 3:305–9. doi: 10.1007/s10126-001-0012-7
23. Bhaskar B, Ananthanarayan L, Jamdar S. Purification, identification, and characterization of novel angiotensin I-converting enzyme (ACE) inhibitory peptides from alcalase digested horse gram flour. Lwt. (2019) 103:155–61. doi: 10.1016/j.lwt.2018.12.059
24. Hao L, Gao X, Zhou T, Cao J, Sun Y, Dang Y, et al. Angiotensin I-Converting Enzyme (ACE) inhibitory and antioxidant activity of umami peptides after in vitro gastrointestinal digestion. J Agric Food Chem. (2020) 68:8232–41. doi: 10.1021/acs.jafc.0c02797
25. Mine, Y Shahidi F. Nutraceutical Proteins And Peptides In Health And Disease. New York, NY: Taylor & Francis Group (2006). p. 269–315.
26. Wu J, Aluko RE, Nakai S. Structural requirements of angiotensin I-converting enzyme inhibitory peptides: quantitative structure-activity relationship modeling of peptides containing 4-10 amino acid residues. QSAR Combinat Sci. (2006) 25:873–80. doi: 10.1002/qsar.200630005
27. Matsumura N, Fujii M, Takeda Y, Sugita K, Shimizu TJ. Angiotensin I-converting enzyme inhibitory peptides derived from bonito bowels autolysate. Biosci Biotechnol Biochem. (1993) 57:695–7. doi: 10.1271/bbb.57.695
28. Yu Y, Jin Y, Wang F, Yan J, Qi Y, Ye M. Protein digestomic analysis reveals the bioactivity of deer antler velvet in simulated gastrointestinal digestion. Food Res Int. (2017) 96:182–90. doi: 10.1016/j.foodres.2017.04.002
29. Katayama K, Mori JT, Kawahara S, Miake K, Kodama Y, Sugiyama M, et al. Angiotensin-I converting enzyme inhibitory peptide derived from porcine skeletal muscle myosin and its antihypertensive activity in spontaneously hypertensive rats. J Food Sci. (2007) 72:S702–6. doi: 10.1111/j.1750-3841.2007.00571.x
30. Ye Y, Engholm-Keller K, Fang Y, Nielsen CF, Jorda A, Lund MN, et al. UHT treatment and storage of liquid infant formula affects protein digestion and release of bioactive peptides. Food Funct. (2022) 13:344–55. doi: 10.1039/d1fo02619d
31. Li GH, Le GW, Shi YH, Shrestha S. Angiotensin I-converting enzyme inhibitory peptides derived from food proteins and their physiological and pharmacological effects. Nutr Res. (2004) 24:469–86.
32. Ye S, Chen Q, Li D, Zhou H, Chen Y, Meng C, et al. Isolation and identification of novel angiotensin I-converting enzyme (ACE) inhibitory peptides from Pony Seed and evaluation of the inhibitory mechanisms. J Funct Foods. (2022) 95:105151. doi: 10.1016/j.jff.2022.105151
33. Cheung HS, Wang FL, Ondetti MA, Sabo EF, Cushman DW. Binding of peptide substrates and inhibitors of angiotensin-converting enzyme. Importance of the COOH-terminal dipeptide sequence. J Biol Chem. (1980) 255:401–7. doi: 10.1016/s0021-9258(19)86187-2
34. Chaudhary S, Vats ID, Chopra M, Biswas P, Pasha S. Effect of varying chain length between P(1) and P(1’) position of tripeptidomimics on activity of angiotensin-converting enzyme inhibitors. Bioorg Med Chem Lett. (2009) 19:4364–6. doi: 10.1016/j.bmcl.2009.05.079
35. Wu Q, Du J, Jia J, Kuang C. Production of ACE inhibitory peptides from sweet sorghum grain protein using alcalase: hydrolysis kinetic, purification and molecular docking study. Food Chem. (2016) 199:140–9. doi: 10.1016/j.foodchem.2015.12.012
36. Jimsheena VK, Gowda LR. Arachin derived peptides as selective angiotensin I-converting enzyme (ACE) inhibitors: structure-activity relationship. Peptides. (2010) 31:1165–76. doi: 10.1016/j.peptides.2010.02.022
37. Hernández-Ledesma B, Contreras M, Recio I. Antihypertensive peptides: production, bioavailability and incorporation into foods. Adv Colloid Interface Sci. (2010) 165:23–35.
38. Fu Y, Therkildsen M, Aluko RE, Lametsch R. Exploration of collagen recovered from animal by-products as a precursor of bioactive peptides: successes and challenges. Crit Rev Food Sci Nutr. (2019) 59:2011–27. doi: 10.1080/10408398.2018.1436038
Keywords: molecular docking, antihypertensive effects, yak bone, ACE inhibitory peptide, digestion
Citation: Gao X, Bu F, Yi D, Liu H, Hou Z, Zhang C, Wang C, Lin J-M, Dang Y and Zhao Y (2022) Molecular docking and antihypertensive effects of a novel angiotensin-I converting enzyme inhibitory peptide from yak bone. Front. Nutr. 9:993744. doi: 10.3389/fnut.2022.993744
Received: 14 July 2022; Accepted: 05 September 2022;
Published: 12 October 2022.
Edited by:
Acácio Antonio Ferreira Zielinski, Federal University of Santa Catarina, BrazilReviewed by:
Sutapa Biswas Majee, NSHM College of Pharmaceutical Technology (NCPT), IndiaCopyright © 2022 Gao, Bu, Yi, Liu, Hou, Zhang, Wang, Lin, Dang and Zhao. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yali Dang, ZGFuZ3lhbGlAbmJ1LmVkdS5jbg==; Yufen Zhao, emhhb3l1ZmVuQG5idS5lZHUuY24=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.