 Begoña Ayuda-Durán1†
Begoña Ayuda-Durán1† Eva Sánchez-Hernández2†
Eva Sánchez-Hernández2† Susana González-Manzano1
Susana González-Manzano1 Celestino Santos-Buelga1
Celestino Santos-Buelga1 Ana M. González-Paramás1*
Ana M. González-Paramás1*- 1Grupo de Investigación en Polifenoles (GIP-USAL), Universidad de Salamanca, Salamanca, Spain
- 2Department of Agricultural and Forestry Engineering, ETSIIAA, University of Valladolid, Palencia, Spain
Introduction: Increasing evidence supports the role of gut microbiota in many aspects of human health, including immune, metabolic and neurobehavioral traits. Several studies have focused on how different components of the diet, such as polyphenols, can modulate the composition and function of the gut microbiota leading to health benefits.
Methods: The effects on the resistance against thermally induced oxidative stress of C. elegans grown in the presence of flavonoids (quercetin or epicatechin) and fed different probiotic strains, namely Lactobacillus plantarum CLC17, Bifidobacterium longum NCIMB 8809 and Enterococcus faecium CECT 410, were explored.
Results: Feeding C. elegans with the assayed bacteria in the absence of flavonoids did not significantly affect body size and fertility of the worms neither improve their resistance against oxidative stress compared to E. coli controls. However, increased resistance to stress was found when C. elegans was cultivated in the presence of both L. plantarum and flavonoids, but not with B. longum or E. faecium. An exploratory study revealed the presence of glycosylated and sulfated metabolites together with the aglycone in worms treated with quercetin and fed any of the different assayed LAB strains. However, in the assays with epicatechin a differential metabolite, tentatively identified as 5-(4′-hydroxyphenyl)-γ-valerolactone 3′-O-glucoside, was detected in the worms fed L. plantarum but not with the other bacteria.
Conclusion: The obtained results indicated that the interactions bacteria/polyphenol play a key role in the effects produced in C. elegans regarding resistance against oxidative stress, although those effects cannot be only explained by the ability of bacteria to metabolize polyphenols, but other mechanisms should also be involved.
Introduction
Probiotic bacteria have been linked with beneficial effects on the health of the host (1). Several clinical studies have attributed to probiotics a high therapeutic potential in the treatment of diarrhea associated with antibiotics (2), inflammatory bowel disease (3), obesity and diabetes (4), respiratory tract infections in children (5), anti-carcinogenic effects (6) and prevention of urinary tract infections (7). Probiotics may exert their action on the host in different ways, including modulation of the composition of the intestinal microbiota, maintenance of the integrity of the intestinal barrier or improvement of the human immune system (8, 9). The most commonly used probiotic microorganisms are Lactic Acid Bacteria (LAB) such as Lactobacillus, Bifidobacterium and Enterococcus species, the latter harboring both pathogenic and commensal microorganisms, although only in rare occasions they are associated with a risk of infection for humans (10).
Lactobacillus plantarum, one of the most important species of the genus Lactobacillus, has been reported to decrease cholesterol levels and reduce symptoms of gastrointestinal disorders (11). Different strains of L. plantarum have been shown to be able to metabolize polyphenols, exerting antimicrobial activity against a wide range of pathogenic bacteria, owing to their ability to produce low molecular weight compounds with bactericidal potential (12, 13). In particular, L. plantarum CLC17 strain, isolated from breast milk (14), can degrade polyphenols to a variety of end products of phenolic catabolism, like syringic, vanillic, 3-O-methyl gallic and gallic acids, being its growth in turn stimulated by the presence of phenolic compounds and their metabolites (15). Furthermore, this strain can provide protection to the intestinal ecosystem, reducing the content of bacteria belonging to the Enterobacteriaceae family (16). These effects have led to postulate L. plantarum CLC17 to be used as a food ingredient to improve the metabolism of dietary polyphenols (16). The genus Bifidobacterium possesses recognized ability to metabolize dietary carbohydrates and is believed to allow efficient and persistent colonization in the gut (17). Bifidobacterium longum subsp. longum, commonly found in the human gut microbiota and with perceived positive health effects (18), also demonstrated capable of metabolizing hydroxycinnamic acids (19). The administration of B. longum has been proposed to contribute to prevent gastrointestinal disturbances such as diarrhea, constipation and bowel cancer (6). Enterococcus strains are highly adapted to various food systems due to their good tolerance to salts and acids, being involved in cheese and sausage fermentation processes (20). Different strains of E. faecium and E. faecalis have been proposed as probiotics (21), being shown to exhibit desirable features, such as high adhesion and low cytotoxicity, as well as cytoprotective and anti-inflammatory properties in in vitro studies (22). In particular, E. faecium CECT 410 strain was reported to be able to colonize pig intestinal microbiota and to reduce coliforms in their feces (23).
The phenolic compounds, commonly referred to as polyphenols, are a group of plant secondary metabolites widespread in higher plants, where they contribute to the mechanisms of natural resistance. They are also widely distributed in the human diet through grains, fruits and vegetables, and derived products, such as wine, tea or chocolate, in which they contribute to sensory, technological and health properties. Based on their chemical structures, they can be divided into different subgroups. Among them, flavonoids constitute the largest group in nature, being broadly distributed in the human diet. The consumption of a diet rich in flavonoids has been associated with a decreased incidence of the most common chronic diseases in developed countries, like type 2 diabetes, different types of cancer, neurodegenerative illnesses and cardiovascular disorders (24).
Most dietary polyphenols are not absorbed by the small intestine and reach the gut, where they interact with the intestinal microbiota to be metabolized to a range of metabolites, some of which can be bioactive and contribute to the potential health effects associated to polyphenol consumption. The composition of the intestinal microbiota varies among people, which may affect the metabolism of the ingested polyphenols (25). For their part, polyphenols and their metabolites might influence the composition and function of the gut microbiota acting on their growth or metabolism, as well as avoiding the growth of pathogens (25, 26). Actually, phenolic compounds have been attributed prebiotic properties associated with their antimicrobial activity and capacity to modulate the intestinal microbiota (27). Thus, two-way interactions between polyphenols and microbiota could contribute to their putative effects in human health.
In order to clarify the interactions polyphenols-microbiota, the use of simple organisms like C. elegans can be very useful. This worm is a free-living bacterivore that grows optimally in a wide variety of microorganisms. Zhang et al. (28) denoted significant differences in the worm’s microbial populations depending on the area and environment where they have grown. However, in the laboratory the nematode is fed routinely with Escherichia coli strain OP50 in a monoxenic culture. This and the synchronization of cultures by bleaching make worms to lose their innate microbiota. Several studies have explored feeding the nematode with different bacterial strains. Sánchez-Blanco et al. (29) found that C. elegans fed Bacillus subtilis (PY79, 3610, and 168 strains), present in its usual soil habitat, lived longer (43–58%) than those fed the standard E. coli diet. The authors showed that this difference was not due to the nutritive quality of the diet, but to factors contributed by the bacterium, namely the presence of coenzyme Q (CoQ) synthesized by E. coli but not by B. subtilis. The authors proposed that CoQ supplementation provided by E. coli would alter the worm cellular redox homeostasis leading to a decreased longevity, exemplifying the relevance of the microbiome on life expectancy. Regarding LAB, it has been observed that C. elegans fed heat-killed L. fermentum LA12 and L. plantarum CJLP133 presented longer lifespan compared with E. coli controls (30). Similarly, worms cultured with L. salivarus FDB89 improved the lifespan by a caloric restriction mechanism (31), while E. faecium L11 increased worm longevity triggering the expression of genes related to aging and innate immunity in C. elegans (32). Feeding with L. rhamnosus improved worm resistance against inflammation and oxidative stress (33), whereas B. animalis subsp. lactis CECT 8145 increased oxidative stress resistance in C. elegans and decreased worm body fat and triglycerides (34).
Some studies have focused on the ability of LAB to protect C. elegans from pathogenic bacteria, such as Salmonella enterica serovar Enteritis (35), Salmonella enterica serovar Typhimurium (32), Yersinia enterocolitica (36), Legionella pneumophila (37), E. coli (38), and Staphylococcus aureus and E. coli O157:H7 (39). Quite recently, Kumar et al. (40), employed C. elegans to explore the probiotic potential of a Lactobacillus plantarum strain (LPJBC5). Those authors concluded that this strain was able to increase worm lifespan, improve stress resistance and promote a series of traits associated with healthy aging, such as physical and cognition performance, fat accumulation, gut integrity, or mitochondrial function. The observed effects were suggested to be mediated by the activation of the p38 MAPK signaling pathway and downstream targets, such as the SKN-1 transcription factor, upregulating the expression of stress resistance genes. Furthermore, LPJBC5 was also able to downregulate fat-5 and fat-7 genes modulating fat metabolism, as well as to upregulate genes involved in serotonin signaling (ser-1, tph-1, and mod-1) related to improved cognitive function (40).
The aim of the present work is to evaluate the ability of three LAB strains, Lactobacillus plantarum CLC17, Bifidobacterium longum NCIMB 8809 and Enterococcus faecium CECT 410, to modulate C. elegans resistance against oxidative stress by themselves and when combined with polyphenols. Epicatechin and quercetin, belonging to the flavan-3-ol and flavonol classes, respectively, and that are two of the best-represented flavonoids in the human diet, were employed as model polyphenols.
Materials and methods
Standards and reagents
(-)-Epicatechin (EC), quercetin (Quer), ampicillin sodium salt, nistatine, agar, yeast extract, cholesterol, 5-fluoro-2′-deoxyuridine (FUdR), and bovine serum albumin were purchased from Sigma-Aldrich (Madrid, Spain). Sodium chloride, calcium chloride, 10% w/v sodium hypochlorite solution, hydrogen chloride, sodium hydroxide and dimethyl sulfoxide (DMSO) were obtained from Panreac (Barcelona, Spain). Potassium dihydrogen phosphate, potassium monohydrogen phosphate, sodium monohydrogen phosphate, acetic acid and magnesium sulfate were from Merck (Darmstadt, Germany). Petri plates Ø 35 and 60 mm were from Brand GMBH (Wertheim, Germany) and Petri plates Ø 90 mm from Francisco Soria Melguizo (Valdemoro, Madrid). MRS Broth was from Fisher Scientific (Madrid, Spain) and tryptone medium from Fluka Analytical (Madrid, Spain).
Strains and maintenance conditions
The C. elegans wild-type strain N2 and E. coli OP50 bacterial strain were obtained from the Caenorhabditis Genetics Center at the University of Minnesota (Minneapolis, USA). Probiotic strains L. plantarum CLC17, B. longum NCIMB 8809 and E. faecium CECT 410 were provided by the Department of Food Biotechnology and Microbiology of the Food Science Research Institute (CIAL-CSIC, Madrid, Spain).
LAB strains were cultured in De Man, Rogosa, and Sharpe (MRS) broth supplemented with L-cysteine (0.05%) at 37°C for 24 h in anaerobic conditions. In order to be used as standard feed for nematodes, E. coli OP50 was grown in LB broth at 37°C for 24 h. Bacteria were collected by centrifugation at 16,300 g for 1 min, washed twice with sterile M9 buffer (3 g KH2PO4, 6 g Na2HPO4, 5 g NaCl, 1 ml 1 M MgSO4, H2O to 1 L) and centrifuged again at 16,300 g for 1 min to remove the supernatant. The bacteria were adjusted to a final concentration of 0.10 mg (wet weight) per μL in M9 buffer. The suspended cells were placed on nematode growth medium (NGM) plates and dried.
EC and Quer solutions (200 mM) in DMSO were added to the nematode growth medium during its preparation to get a 200 μM final concentration of the flavonoids on the plates. Control plates were also prepared without the compounds but containing the same volume of DMSO (0.1% DMSO, v/v).
Wild worms were routinely propagated at 20°C on NGM plates containing E. coli OP50 as a food source. Synchronization of worm cultures was achieved by treating gravid hermaphrodites with sodium hypochlorite-sodium hydroxide solution 5 N (50:50). Eggs are resistant whereas worms are dissolved in the bleach solution. The suspension was vortex shaken for one min, kept a further minute on rest and then centrifuged (2 min, 8,700 g); this process was repeated five times. The final pellet containing the eggs was washed six times with an equal volume of buffer M9. A volume of 100–300 μL of the M9 buffer with eggs (depending on eggs concentration) was transferred and incubated on NGM agar plates in the presence of the flavonoids (EC or Quer) or in their absence. The synchronized L1 larvae were fed with E. coli OP50 until they reached the larval state L4. Then, they were moved to new plates covered with a lawn of each of the different bacteria, i.e., E. coli OP50, as a control, L. plantarum CLC17, B. longum NCIMB 8809, or E. faecium CECT 410, and supplemented or not with EC or Quer. The worms were transferred every 2 days to fresh plates until they reached the day of the assay. In all cases, the plates also contained FUdR at a concentration of 150 μM to prevent reproduction and progeny overgrowth.
Body length measurement
Age-synchronous worms were grown to the L4 stage on NGM plates seeded with E. coli OP50, then transferred individually to plates containing either OP50 or the probiotic strain and maintained at 20°C for 6 days. Nematode length was measured every 24 h alive with a microscope (Leica M205 FA, Germany) equipped with a camera (Leica DFC 420), coupled with a Leica Application Suite V3 data processing software. The images were analyzed using the Image J software. Nematode length was measured in triplicate from about 10 individuals per treatment and assay.
Reproduction assays
Synchronized L4 worms fed E. coli OP50 were individually transferred to plates with the different bacterial strains and stored at 20°C for 6 days until their reproductive stage ceased. During that time, the worms were moved every day to fresh plates with the corresponding assayed bacteria. The offspring of each animal was counted to verify that eggs were fertile. For each of the studied LAB, assays were performed three times with 10 worms per assay.
Thermal stress assays
L1 nematodes were incubated on OP50 plates with or without the assayed flavonoid. At the L4 stage, the nematodes were placed onto the different bacterial lawns (L. plantarum CLC17, B. longum NCIMB 8809, E. faecium CECT 410 or E. coli OP50), and, in the assays with the dual bacteria/flavonoid system, also in the presence of EC or Quer. Worms from the different treatments were transferred with a platinum wire to agar plates (Ø 35 mm, 20 worms per plate) at two stages of development: reproduction (2nd day of adulthood) and post-reproduction stage (9th day of adulthood), at which they were submitted to thermally induced oxidative stress (35°C, 8 h). Afterward, the number of dead and alive nematodes was counted. The relative rates of survival of worms after being subjected to thermal stress were expressed in relation to the untreated controls. Assays were carried out with approximately 100 nematodes per treatment, performing three independent trials per assay.
Assessment of flavonoids uptake by Caenorhabditis elegans
Synchronized worms were grown until L4 stage with OP50 in the presence or not of the assayed flavonoids. Then, animals were washed twice with M9 to remove the E. coli as much as possible and moved to fresh plates that contained E. coli or the different bacteria (L. plantarum CLC17, B. longum NCIMB 8809, E. faecium CECT 410) with or without the flavonoid (EC or Quer). After 4 days, the worm biomass was washed successively once with M9, three times with PBST (PBS + 0.01% Tween 20), once with PBS and finally with M9 buffer.
The remaining worm pellet was resuspended in 1 ml of 30% methanol, the mixture was vortex shaken vigorously and further homogenized in a Thermo Savant FastPrep 120 Cell Disrupter System at a speed of 5.5 m/s using 7 cycles of 10 s. The supernatant was then collected and the pellet washed and centrifuged further two times with 1 ml of 30% methanol. The combined methanolic supernatants were dried in a centrifugal concentrator micVac (GeneVac, Ipswich, UK) and the residue dissolved in 500 μL of acetonitrile 20% for high-performance liquid chromatography (HPLC)-DAD/MS analysis. Two independent experiments were performed.
Total protein was also determined by the Bradford method after digestion of the worm homogenates, in order to express compound concentration in relation to worm protein.
Analysis by high-performance liquid chromatography-diode array detection-mass spectrometry
Analyses were performed in a Hewlett-Packard 1100 chromatograph (Agilent Technologies, Waldbronn, Germany) equipped with a quaternary pump and a diode array detector (DAD) coupled to an HP Chem Station (rev. A.05.04) data-processing station. A Waters Spherisorb S3 ODS-2C8, 3 μm (4.6 × 150 mm) column thermostatted at 30°C was used. The solvents used were: (A) 0.1% formic acid, and (B) acetonitrile. The elution gradient was 100% A to 15% B in 35 min, and from 15 to 40% B over 10 min, at a flow rate of 0.5 ml min–1. Double online detection was carried out in the DAD at 280 nm and in a mass spectrometer (MS) connected to the HPLC system via the DAD cell outlet. MS detection was performed in an API 3200 Qtrap (Applied Biosystems, Darmstadt, Germany) equipped with an ESI source and a triple quadrupole-ion trap mass analyzer, which was controlled by the Analyst 5.1 software. Zero grade air served as the nebulizer gas (30 psi) and as turbo gas (400°C) for solvent drying (40 psi). Nitrogen was used as the curtain (20 psi) and collision gas (medium). Both quadrupoles were set at unit resolution. The ion spray voltage was operated at –4,500 V in the negative mode. Method settings were: declustering potential (DP), –40 V; entrance potential (EP), –10 V; collision energy (CE), –50 V; and cell exit potential (CXP) –3 V. Fragmentation of the parent ions was obtained in enhanced product ion (EPI) mode using the following settings: declustering potential (DP), –50 V; entrance potential (EP), –6 V; collision energy (CE), –25 V; and collision energy spread (CES) 0 V.
Statistical analyses
Statistical analyses were performed using the SPSS PC software package (version 25.0, SPSS Inc., Chicago). ANOVA was used for multiple comparisons of values to determine possible significant differences between the treated and control groups in the phenotypic characterization tests: body length and reproduction. For thermal stress survival trials, contingency tables were made, and statistical significance was calculated using the Chi-Square Test. In each analysis, the differences were considered statistically significant at the level of p < 0.05.
Results and discussion
Assessment of Caenorhabditis elegans viability
Phenotypical characteristics (body length and reproduction rate) of the worms cultured with the LAB bacteria (L. plantarum CLC17, B. longum NCIMB 8809, and E. faecium CECT410) were checked and compared with those fed E. coli OP50, in order to test if C. elegans viability could be affected by the bacterial strain. It must be noted that cultivation had to be made using E. coli OP50 as a feed during the larval stage, and only after reaching the larval state L4 worms were transferred to plates seeded with the different bacteria. This practical approach is required since probiotic bacteria like Lactobacillus cause hatched eggs to be arrested as L1 larvae (41).
Figure 1 shows the changes in the body length of worms grown in the presence of the different bacteria for 6 days. It was observed that C. elegans fed L. plantarum and E. faecium showed a developmental delay leading to a significantly shorter body length compared to the control (E. coli OP50) in the first and second day of adulthood. However, from day 3 onward no significant differences were found in body length among individuals cultivated with the different bacteria. This developmental delay could be explained by the change of food from E. coli (larval phase) to LAB strains (L4 onward), as worms need some period of adaptation to the distinct bacteria.
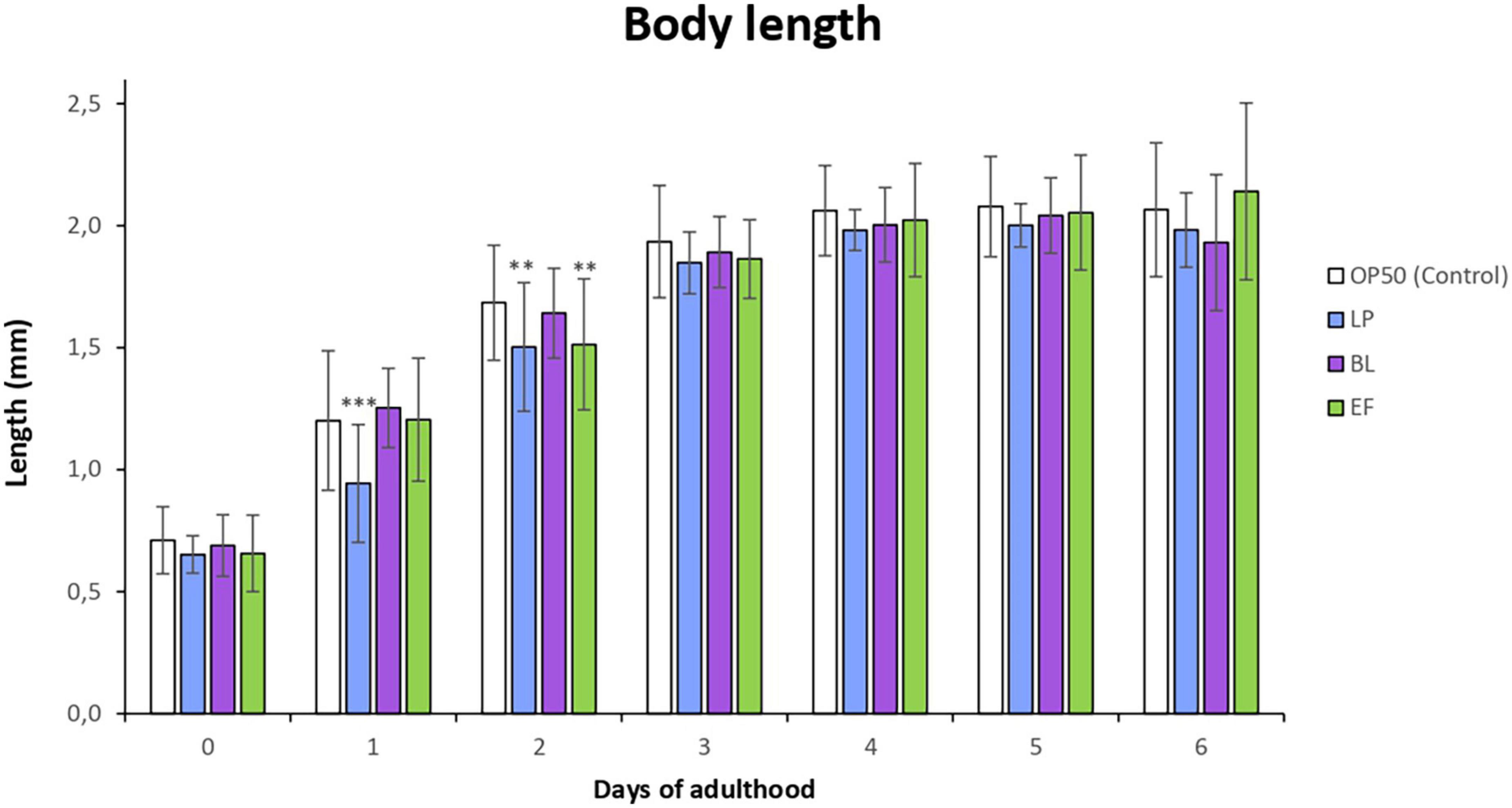
Figure 1. Changes in body length in C. elegans fed Lactobacillus plantarum CLC17 (LP), Bifidobacterium longum NCIMB 8809 (BL), Enterococcus faecium CECT 410 (EF) and E. coli OP50 (control) from 1st to 6th day of worm adulthood. The error bars represent standard deviation (n = 30). Differences in body length in worms fed LAB bacteria vs. E. coli OP50 were considered significant at ***p < 0.001, **p < 0.01.
The rate of reproduction of C. elegans cultivated in the presence of the different assayed bacteria was examined by scoring total progeny eggs from 30 worms for the first 6 days of adulthood. The results are depicted in Figure 2. Cultivation with B. longum and E. faecium led to a smaller number of viable eggs compared with E. coli control, although the differences were no significant. In all cases, worms started to lay eggs the first day of adult and until approximately the fourth day. Nevertheless, in the case of those grown in presence of B. longum a delay of some hours at the beginning of reproduction was observed with respect to the E. coli OP50 control, which might explain the lower rate of egg production compared with the other bacteria.
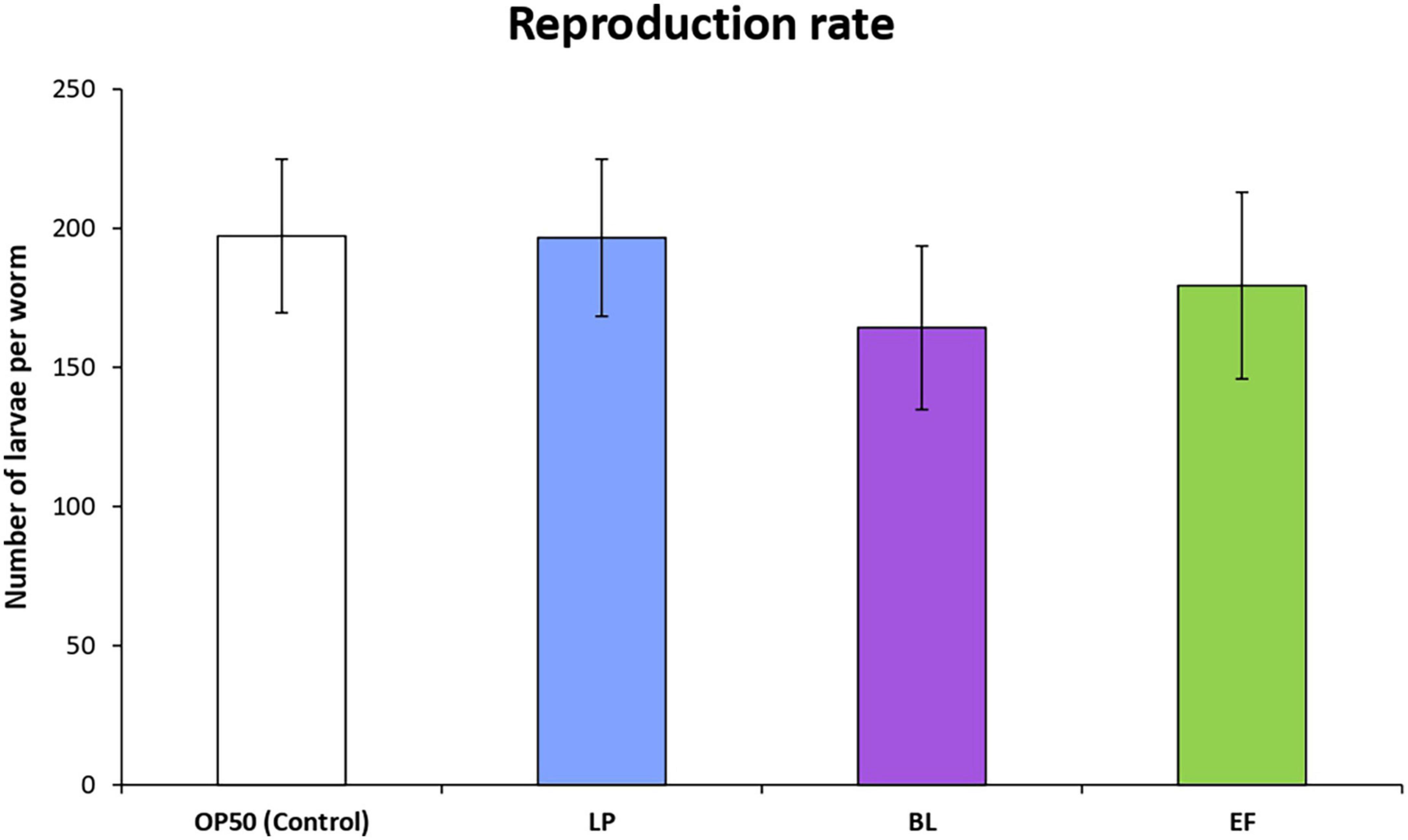
Figure 2. Influence of Lactobacillus plantarum CLC17 (LP), Bifidobacterium longum NCIMB 8809 (BL), Enterococcus faecium CECT 410 (EF) and E. coli OP50 (control) in the reproduction rate of C. elegans. The error bars represent standard deviation (n = 30 worms). No significant differences among worms cultivated with the different bacteria were observed (p < 0.05).
Several studies have observed a smaller body size and a reduction in the reproduction capacity in worms fed with different probiotic strains. Schifano et al. (42) found that Lactobacillus fermentum MBC2 affected C. elegans fertility reducing the reproduction rate of the progeny by 40% compared to E. coli OP50 fed animals, and the body length of worms was also shorter. Zhao et al. (31) observed that worms grown in the presence of L. salivarus were remarkably smaller and showed a decrease in their reproduction rate compared to those fed E. coli, which was explained by a possible reduction in the caloric intake of the worm. Ikeda et al. (35) reported that worms fed B. infantis presented a significant decrease in their body size in the first 3 days after the change of food from E. coli OP50 to the probiotic to equal the control in the following days, an observation similar to the one made in the present study.
The obtained results confirmed that the assayed LAB can be used to feed C. elegans without affecting significantly worm viability and phenotypic and reproductive characteristics. Therefore, we could proceed to check whether LAB could provide advantages on worm resistance against oxidative stress, either by themselves or in combination with polyphenols.
Assessment of the antioxidant effect of bacterial strains in Caenorhabditis elegans
The three assayed probiotic bacteria were assessed to check whether they could influence the resistance of C. elegans against oxidative damage. With this aim, worms grown from L4 larval stage in the presence of L. plantarum CLC17, B. longum NCIMB 8809 or E. faecium CECT410 were subjected to thermally induced oxidative stress (35°C, 8 h) applied on the 2nd and 9th day of adulthood and compared to those fed E. coli OP50 as a control. These times were chosen to verify if the effects could be different according to the age of the nematode, i.e., young reproductive adults or older adults in post-reproductive stage.
As it can be seen in Figure 3, despite some variations were observed in the percentages of survival of C. elegans fed L. plantarum and B. longum, the differences among them and with the E. coli control were not significant, either at days 2 or 9 of adulthood. By contrast, a significant decrease was produced in the survival of worms cultivated with E. faecium CECT410 after the stress applied on day 2, although no differences were found when the stress was applied on day 9. These findings seem coherent with the variation in the body length (Figure 1), which was also decreased at day 2 compared to control worms. None of the assayed bacteria seemed to provide additional advantages against oxidative stress compared to usual E. coli feed, and all the effects observed after day 2 could be explained by the adaptation of the worms to a new food source. These results do not agree with those obtained by some authors with other probiotic strains, like Lactobacillus rhamnosus CNCM I-3690 (33), Lactobacillus gasseri SBT2055 (43), Bifidobacterium animalis subsp. lactis CECT 8145 (34) and Bifidobacterium longum CECT 7347 (44), all of them providing C. elegans enhanced resistance against acute oxidative stress damage.
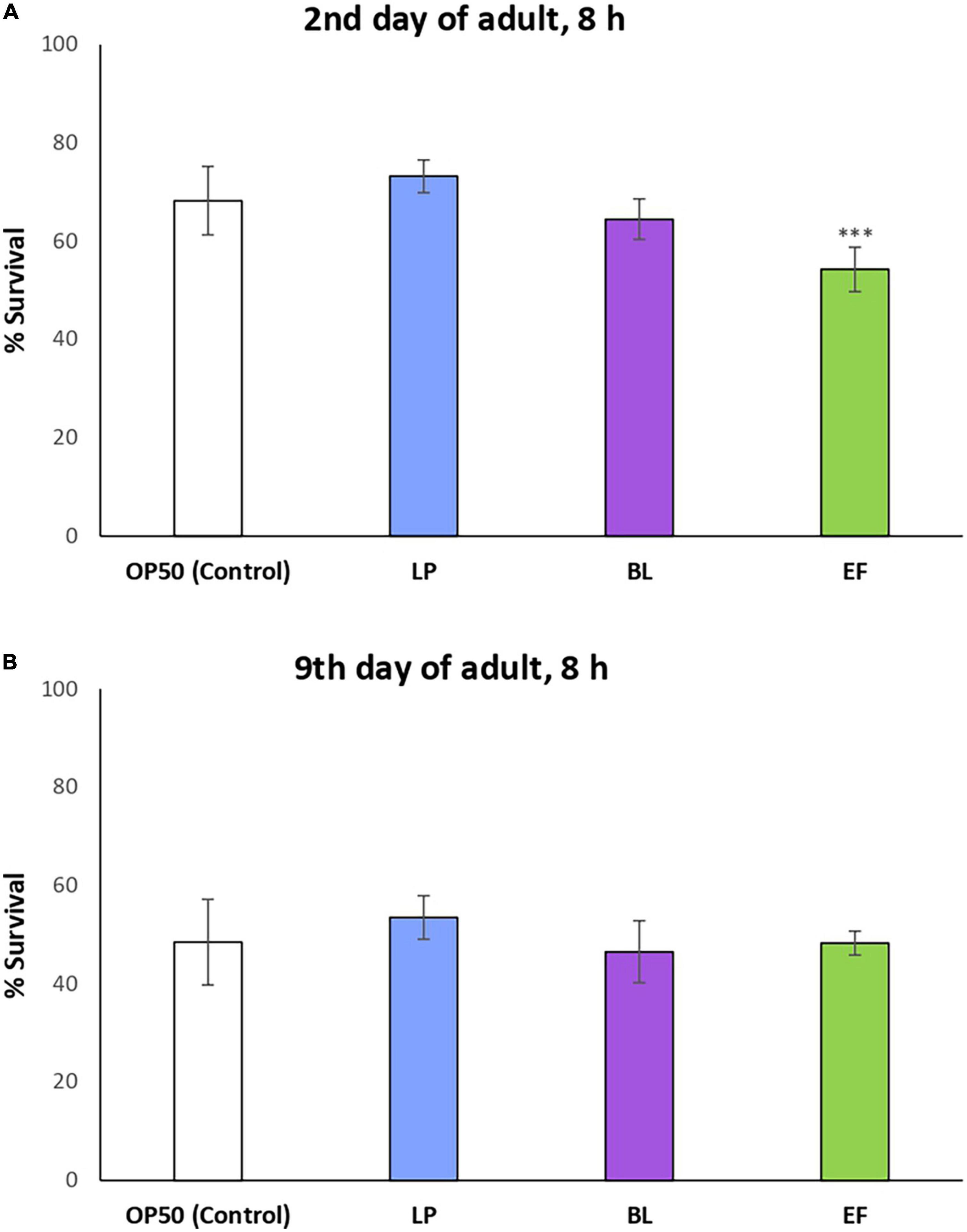
Figure 3. Percentages of survival following thermal stress (35°C, 8 h) applied at days 2 (A) and 9 of adulthood (B) in N2 wild type C. elegans grown in presence of different bacteria strains: Lactobacillus plantarum CLC17 (LP), Bifidobacterium longum NCIMB 8809 (BL), Enterococcus faecium CECT 410 (EF) and E. coli OP50 (control). Statistical significance was calculated using the Chi-Square Test. The differences were considered significant at ***p < 0.001.
Effect of bacteria/polyphenol systems on Caenorhabditis elegans viability and resistance to oxidative stress
The effects on resistance to oxidative stress of dual bacteria/polyphenol systems was assessed in worms cultured in the presence of each of the assayed probiotics and two highly consumed flavonoids. In particular, epicatechin (EC), a monomeric flavan-3-ol, and quercetin (Quer), a flavonol, compounds that are widely distributed in the human diet through a variety of fruits, vegetables and derived products, whose consumption has been related to health-promoting effects (24). The assays were carried out using concentrations of EC and Quer in the culture medium of 200 μM, a level that was chosen based on previous studies of our group where a range of concentrations was checked (10–500 μM). In those studies, a hormetic response of C. elegans to polyphenols was observed, leading to beneficial effects on longevity and resistance to oxidative stress up to 200 μM, while detrimental effects were produced at higher concentrations (45, 46). Similar results regarding improvement in lifespan and stress resistance were also observed by other authors at that concentration (47, 48).
Significant increases in survival were found in C. elegans cultivated with L. plantarum and B. longum in the presence of EC (200 μM) after being subjected to thermal stress (35°C, 8 h) at day 2 of adulthood compared to those grown with the probiotics alone (Figure 4A). However, in older worms (day 9) the increase was only significant for the system L. plantarum + EC but not for B. longum (Figure 4B). No significant differences were found in worms fed E. faecium at any of the assayed days.
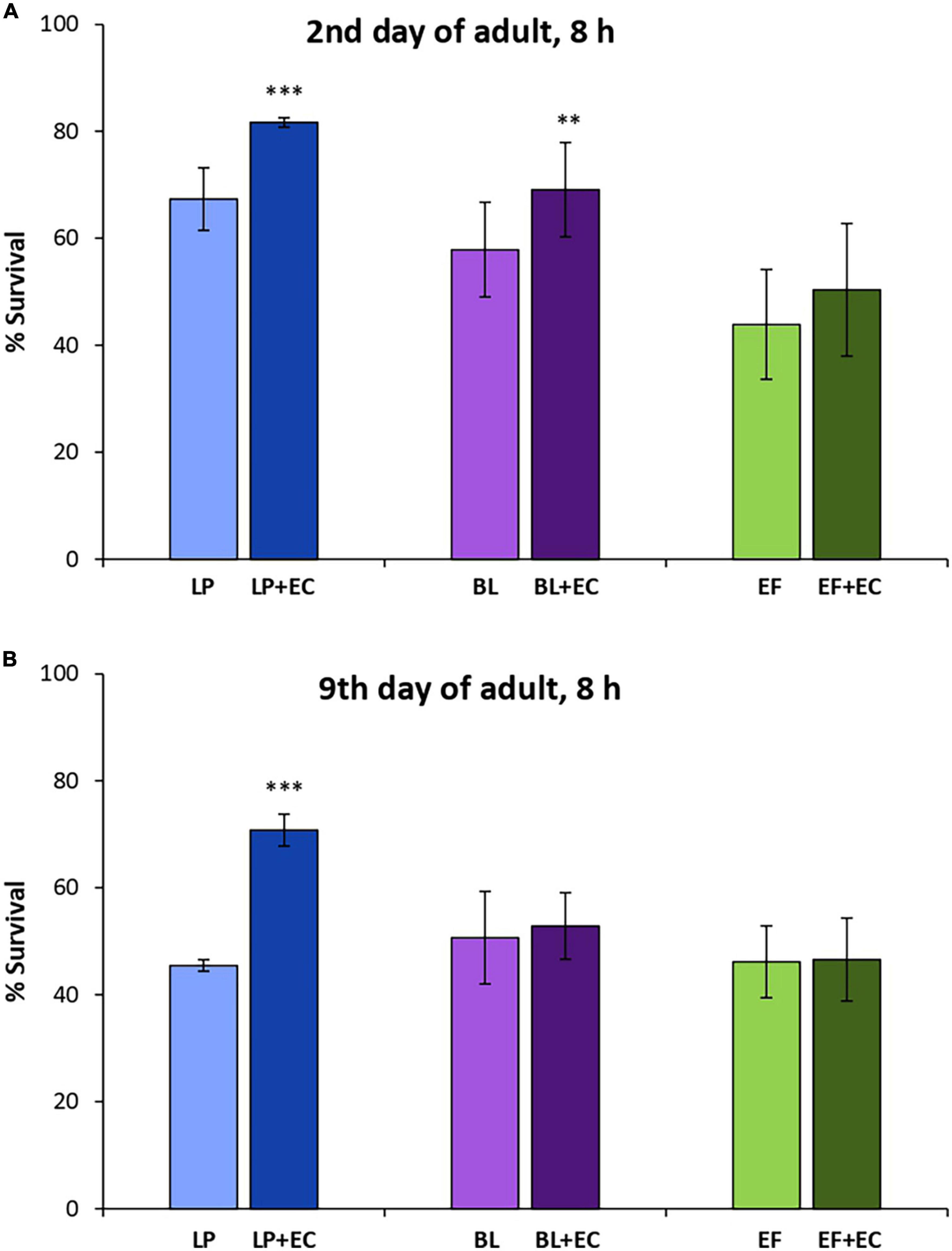
Figure 4. Survival rates of N2 wild type C. elegans after being submitted to thermal stress (35°C, 8 h) applied at days 2 (A) or 9 of adulthood (B) following cultivation with and without epicatechin (EC, 200 μM) in the presence of Lactobacillus plantarum CLC17 (LP), Bifidobacterium longum NCIMB 8809 (BL) or Enterococcus faecium CECT 410 (EF). Differences between worms grown without and with EC were calculated for each strain using the Chi-Square Test. The differences were considered significant at ***p < 0.001 and **p < 0.01.
Similar observations as for EC were made in the assays carried out in the presence of Quer (200 μM), where significant increases in the survival rate after stress were only observed in worms fed L. plantarum, both in young and older animals. In this case, no improvement in the survival was found in the systems B. longum + Quer and E. faecium + Quer, either in young or older worms (Figure 5).
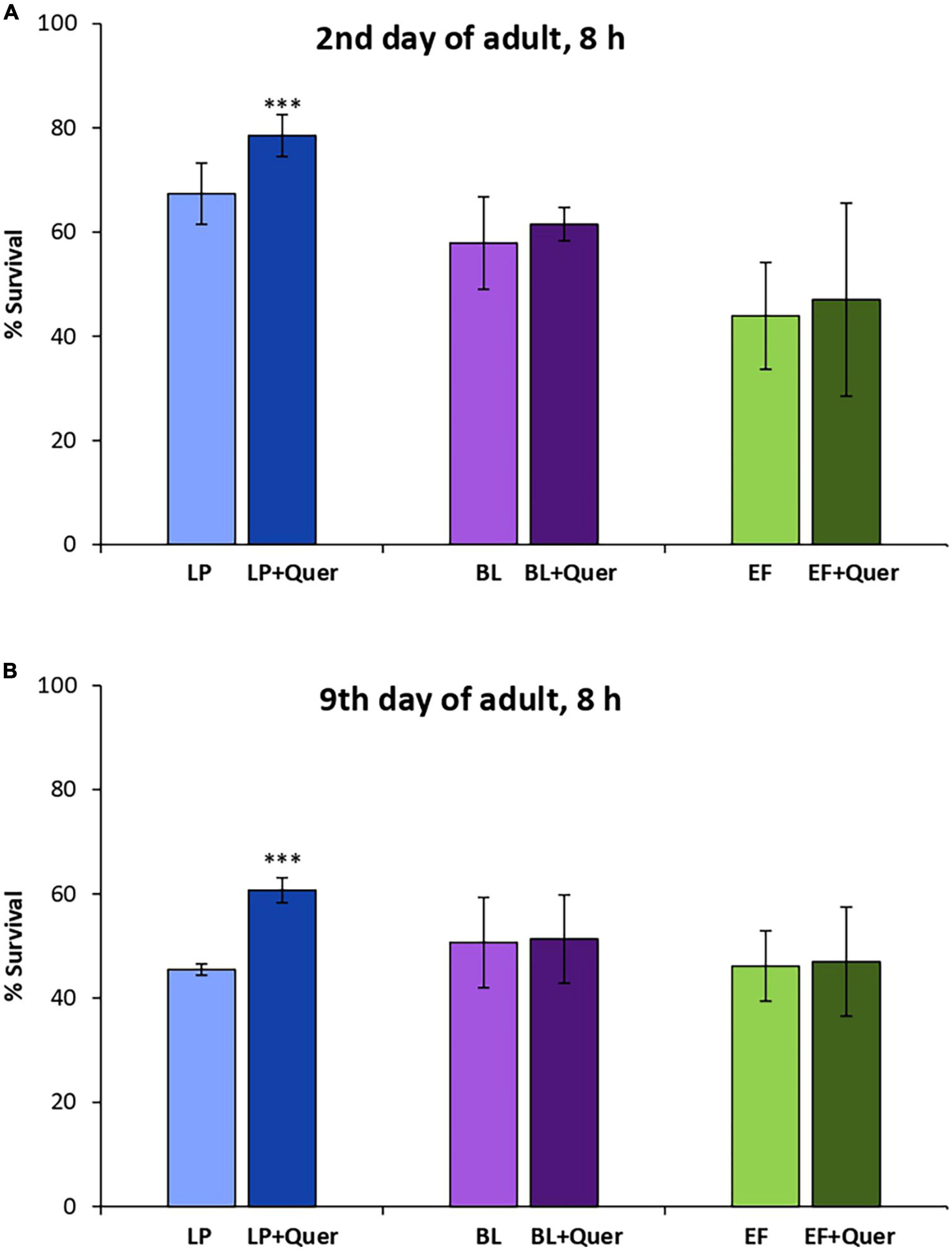
Figure 5. Survival rates of N2 wild type C. elegans after being submitted to thermal stress (35°C, 8 h) applied at days 2 (A) and 9 of adulthood (B) following cultivation with and without quercetin (Quer, 200 μM) in the presence of Lactobacillus plantarum CLC17 (LP), Bifidobacterium longum NCIMB 8809 (BL) or Enterococcus faecium CECT 410 (EF). Differences between worms grown without and with Quer were calculated for each strain using the Chi-Square Test. The differences were considered significant at ***p < 0.001.
The results obtained for L. plantarum are similar to those previously obtained by our group in studies with C. elegans fed E. coli OP50, where significant increases in the resistance against thermal and oxidative stress were found in worms cultivated in the presence of EC or Quer (49–51). Positive effects against oxidative stress for these compounds have also been reported by other authors using the canonical E. coli OP50 to feed C. elegans (47, 52, 53). Similar results were also obtained by our group with other related flavonoids, such as isorhamnetin, tamarixetin, catechin, 4′-O-methylepicatechin and 3′-O-methylepicatechin (49–51, 54).
Different observations were made in the assays with B. longum, where positive effects were only observed when combined with EC in young reproductive animals (Figure 4A), but not for Quer neither in older individuals. In the case of E. faecium no benefits in the protection against oxidative stress were obtained when combined with EC or Quer (Figures 4, 5).
The results herein obtained with the LAB strains are compared with those previously achieved using E. coli OP50 (49–51) in Supplementary Figures 1, 2 included as Supplementary Information. The same experimental conditions have been used in all cases, but in the assays with E. coli + EC the stress was applied in days 1 and 10 instead of days 2 and 9, corresponding in either case to young reproductive worms (days 1–2) and non-reproductive adults (days 9–10). In Supplementary Figures 1, 2 it can be verified that the survival rates of C. elegans at each development stage are fairly consistent in the current assays with the distinct probiotics and in the previous ones with E. coli OP50, despite they were obtained time ago.
Polyphenols uptake and metabolism
In order to try to evaluate the contribution of the different LAB to flavonoid metabolism, worms were cultured for 4 days in presence of the different bacteria in NGM plates containing Quer, EC or DMSO (control). Afterward, the worm biomass from 2 plates of every assay was collected and homogenized to be analyzed by HPLC-DAD-MS. After a careful study of the complex chromatograms obtained, only those peaks that appeared in any of the treated groups and did not appear in the control group without flavonoids were considered as differential.
The metabolite profile in all worms fed the different bacteria in the assays carried out with Quer was similar in all cases, identifying the quercetin aglycone and some quercetin hexosides and quercetin hexoside sulfates. Although these types of metabolites were unequivocally identified by mass and absorption spectra, it was not possible to quantify them since the concentrations present in the samples were very low and their peaks small, close to the chromatogram baseline and/or overlapped by other peaks. Only the concentration of Quer aglycone could be quantified from the area of their chromatographic peak recorded at 360 nm, and was expressed as μg mg–1 of worm protein. The results obtained in the worms fed the different bacteria showed concentrations of Quer aglycone of 0.67 ± 0.10 (OP50), 0.29 ± 0.04 (LP), 4.00 ± 0.44 (BL) and 1.27 ± 0.93 (EF) μg mg–1 protein. No direct correlation could be established between these concentrations and the beneficial effects observed regarding resistance to thermal stress in the presence of the different bacteria. It must be indicated that a targeted search was also performed in all MS chromatograms looking for specific masses corresponding to commonly reported quercetin catabolites from degradation by the colonic microbiota, including benzoic, phenylacetic and phenylpropionic acid derivatives. However, none of the searched compounds was specifically detected in the treated worms. In the end, the only clear conclusion is that quercetin was taken up by the worms and that different amounts of this flavonoid were accumulated by them depending on the type of bacteria used for cultivation, although the same concentration of quercetin was present in the plates.
Different observations were made in the studies with EC. Firstly, contrary to Quer, EC and, conjugated EC metabolites (i.e., glycosylated/sulfated derivatives) could not be found in the chromatograms obtained from the assays with any of the different bacteria. This might suggest that a complete degradation of EC was produced, either by worms or bacteria. Despite this, it was apparent is that, in the cases where beneficial effects were produced, they should be explained by the combined system flavonoid + bacteria, as they were not observed in the assays where only bacteria were present. Another relevant observation was that in the case of L. plantarum a metabolite from EC degradation could be specifically detected in the MS chromatograms, which was tentatively identified as 5-(4′-hydroxyphenyl)-γ-valerolactone 3′-O-glucoside, owing to its previous description as a characteristic metabolite from flavan-3-ol microbiota degradation (55). It is possible that the positive effects observed in worms cultivated in the presence of L. plantarum + EC could be related, at least in part, to the apparent ability of this strain to catabolize the flavan-3-ol yielding valerolactones as potential bioactive metabolites, as they were not observed in the assays with B. longum and E. faecalis strains.
As an example, mass chromatograms obtained for the assays performed with C. elegans grown in the presence of L. plantarum and Quer or EC, indicating the peaks identified as flavonoid metabolites, are included in Supplementary Figure 3.
The whole of the observations made strongly suggest that the beneficial effects that flavonoids may have in counteracting oxidative damage in C. elegans are determined by the type of bacteria. Indeed, if the observed effects were due to the antioxidant activity of the flavonoids, similar results regarding stress resistance should have been expected in the presence of the distinct bacteria. Most of the authors that have studied the possible beneficial effect of LAB in combination with polyphenols attribute the observed benefits to the ability of the bacteria to metabolize the phenolic compounds (12, 13, 15). Our data, especially for Quer, do not provide sufficient basis for concluding that a differential metabolization profile by the distinct bacteria used in our study could exist that might explain the different effects. Nevertheless, although no correlation could be concluded between metabolites and beneficial effects, the obtained results confirm that flavonoids are taken up by C. elegans and that the combination of bacteria + polyphenol is determining for the observed effect.
In earlier studies, it was shown that rather than a direct antioxidant effect, the ability of EC and Quer to improve the resistance against oxidative stress in C. elegans might involve the modulation of transcription factors and genes in molecular pathways related to the endogenous mechanisms of defense, such as the insulin/IGF-1 signaling and MAPK pathways (48, 50, 51). In a recent paper, Kumar et al. (40) reported that L. plantarum LPJBC5 strain was able to increase the lifespan and improve stress resistance in C. elegans through the activation of the p38 MAPK signaling pathway, upregulating the expression of stress resistance genes. The results obtained in the present work seem to suggest that not only the potential ability of L. plantarum to interfere on key molecular pathways but also its interaction with flavonoids could be determining to interpret the effects observed herein regarding resistance against oxidative stress. This might explain the differential effects observed for this probiotic strain and the other two assayed LAB.
Conclusion
Feeding C. elegans with the assayed LAB was found not to affect significantly the viability of the worm, despite some delay in its growth was observed in the first days of cultivation after changing from E. coli to the other bacteria as a food source. Similarly, cultivation in the presence of the probiotics did not modify the resistance of the worm against thermally induced oxidative stress in relation to E. coli control. However, an increase in the stress resistance was found when C. elegans was cultivated in the presence of both Lactobacillus plantarum CLC17 and flavonoids (quercetin or epicatechin), but not in the combination of Bifidobacterium longum or Enterococcus faecium with the same flavonoids. These results reveal that different bacteria may distinctively affect the way the worm responds to flavonoids, highlighting the importance of considering the interaction bacteria + flavonoid to explain the effects on the resistance to stress, a point that was not evident in the previous assays when only E. coli OP50 was used to feed the worm. Indeed, in that case the effect could be interpreted as directly due to the flavonoids. It seems now evident that it is the combined system bacteria + flavonoid that accounts for the effects. On the other hand, the resistance to thermal stress in worms cultivated in presence of L. plantarum + EC might be explained by the differential production of potential bioactive metabolites, such as valerolactones, while no clear relationship with any particular metabolite could be established in the assays with Quer. Anyway, what seems evident is that flavonoids are taken up by C. elegans and the beneficial effects that they may have in counteracting oxidative damage in the worm are determined by the concomitant presence of particular types of bacteria.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
CS-B and AG-P: conceptualization, resources, project administration, and funding acquisition. BA-D, SG-M, and ES-H: methodology and formal analysis. BA-D, ES-H, and CS-B: writing—original draft preparation. BA-D, CS-B, and AG-P: writing—review and editing. CS-B, SG-M, and AG-P: supervision. All authors contributed to the article and approved the submitted version.
Funding
The GIP-USAL was financially supported by the Spanish Ministerio de Ciencia e Innovación (Project PID2019-106167RB-I00/AEI/10.13039/501100011033), Consejería de Educación (Project SA093P20), and the Strategic Research Programs for Units of Excellence from Junta de Castilla y León (ref. CLU-2018-04).
Acknowledgments
We thank Spanish Government (Grant PID2019-106167RB-I00/AEI/10.13039/501100011033), Junta de Castilla y León (Grants SA093P20 and CLU-2018-04) for financial support, and Dr. Begoña Bartolomé and Victoria Moreno-Arribas from CIAL-CSIC (Madrid, Spain) for providing the LAB strains.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnut.2022.989427/full#supplementary-material
References
1. Food and Agriculture Organization [FAO], World Health Organization [WHO]. Guidelines for the evaluation of probiotics in food. In: Food and Agriculture Organization [FAO], World Health Organization [WHO] editors. Probiotics in Food. Health and Nutritional Properties and Guidelines for Evaluation. London: FAO. (2002). p. 1–56.
2. Guarino A, Guandalini S, Lo Vecchio A. Probiotics for prevention and treatment of diarrhea. J Clin Gastroenterol. (2011) 45:S37–45. doi: 10.1097/MCG.0000000000000349
3. Sheil B, Shanahan F, O’Mahony L. Probiotic effects on inflammatory bowel disease. J Nutr. (2007) 137:819S–24. doi: 10.1093/jn/137.3.819S
4. Markowiak P, Śliżewska K. Effects of probiotics, prebiotics, and synbiotics on human health. Nutrients. (2017) 9:1021. doi: 10.3390/nu9091021
5. Wang Y, Li X, Ge T, Xiao Y, Liao Y, Cui Y, et al. Probiotics for prevention and treatment of respiratory tract infections in children: a systematic review and meta-analysis of randomized controlled trials. Medicine. (2016) 95:e4509. doi: 10.1097/MD.0000000000004509
6. Eslami M, Yousefi B, Kokhaei P, Hemati M, Nejad ZR, Arabkari V, et al. Importance of probiotics in the prevention and treatment of colorectal cancer. J Cell Physiol. (2019) 234:17127–43. doi: 10.1002/jcp.28473
7. Reid G. The development of probiotics for women’s health. Can J Microbiol. (2017) 63:269–77. doi: 10.1139/cjm-2016-0733
8. Teitelbaum JE, Walker WA. Nutritional impact of pre and probiotics as protective gastrointestinal organisms. Annu Rev Nutr. (2002) 22:107–38. doi: 10.1146/annurev.nutr.22.110901.145412
9. Cerdó T, García-Santos JA, Bermúdez MG, Campony C. The role of probiotics and prebiotics in the prevention and treatment of obesity. Nutrients. (2019) 11:635. doi: 10.3390/nu11030635
10. Sanders ME, Akkermans LMA, Haller D, Hammerman C, Heimbach J, Hörmannsperger G, et al. Safety assessment of probiotics for human use. Gut Microbes. (2010) 1:164–85. doi: 10.4161/gmic.1.3.12127
11. Seddik HA, Bendali F, Gancel F, Fliss I, Spano G, Drider D. Lactobacillus plantarum and its probiotic and food potentialities. Probiotics Antimicrob Proteins. (2017) 9:111–22. doi: 10.1007/s12602-017-9264-z
12. Barroso E, Sánchez-Patán F, Martín-Alvarez PJ, Bartolomé B, Moreno-Arribas MV, Peláez C, et al. Lactobacillus plantarum IFPL935 favors the initial metabolism of red wine polyphenols when added to a colonic microbiota. J Agric Food Chem. (2013) 61:10163–72. doi: 10.1021/jf402816r
13. Barroso E, Van de Wiele T, Jiménez-Girón A, Muñoz-González I, Martín-Alvarez PJ, Moreno-Arribas MV, et al. T. Lactobacillus plantarum IFPL935 impacts colonic metabolism in a simulator of the human gut microbiota during feeding with red wine polyphenols. Appl Microbiol Biotechnol. (2014) 98:6805–15. doi: 10.1007/s00253-014-5744-1
14. Martín R, Olivares M, Marín ML, Fernández L, Xaus J, Rodríguez JM. Probiotic potential of 3 lactobacilli strains isolated from breast milk. J Hum Lact. (2005) 21:8–17. doi: 10.1177/0890334404272393
15. González de Llano D, Gil-Sánchez I, Esteban-Fernández A, Ramos AM, Cueva C, Moreno-Arribas MV, et al. Some contributions to the study of oenological lactic acid bacteria through their interaction with polyphenols. Beverages. (2016) 2:27. doi: 10.3390/beverages2040027
16. Gil-Sánchez I, Cueva C, Tamargo A, Quintela JC, de la Fuente E, Walker AW, et al. Application of the dynamic gastrointestinal simulator (simgi®) to assess the impact of probiotic supplementation in the metabolism of grape polyphenols. Food Res Int. (2020) 129:108790. doi: 10.1016/j.foodres.2019.108790
17. Pokusaeva K, Fitzgerald GF, van Sinderen D. Carbohydrate metabolism in Bifidobacteria. Genes Nutr. (2011) 6:285–306. doi: 10.1007/s12263-010-0206-6
18. Gueimonde M, Tölkkö S, Korpimäki T, Salminen S. New real-time quantitative PCR procedure for quantification of Bifidobacteria in human fecal samples. Appl Environ Microbiol. (2004) 70:4165–9. doi: 10.1128/AEM.70.7.4165-4169.2004
19. Kelly SM, O’Callaghan J, Kinsella M, van Sinderen D. Characterisation of a hydroxycinnamic acid esterase from the Bifidobacterium longum subsp. longum taxon. Front Microbiol. (2018) 9:2690. doi: 10.3389/fmicb.2018.02690
20. Hanchi H, Mottawea W, Sebei K, Hammami R. The genus enterococcus: between probiotic potential and safety concerns—an update. Front Microbiol. (2018) 9:1791. doi: 10.3389/fmicb.2018.01791
21. Franz CMAP, Van Belkum MJ, Holzapfel WH, Abriouel H, Gálvez A. Diversity of enterococcal bacteriocins and their grouping in a new classification scheme. FEMS Microbio Rev. (2007) 31:293–310. doi: 10.1111/j.1574-6976.2007.00064.x
22. Dowdell P, Chankhamhaengdecha S, Panbangred W, Janvilisri T, Aroonnual A. Probiotic activity of Enterococcus faecium and Lactococcus lactis isolated from thai fermented sausages and their protective effect against Clostridium difficile. Probiotics Antimicrob Proteins. (2019) 12:641–8. doi: 10.1007/s12602-019-09536-7
23. Guerra NP, Fajardo-Bernárdez P, Méndez J, Cachaldora P, Pastrana-Castro L. Production of four potentially probiotic lactic acid bacteria and their evaluation as feed additives for weaned piglets. Ani Feed Sci Tech. (2007) 134:89–107. doi: 10.1016/j.anifeedsci.2006.05.010
24. Del Rio D, Rodriguez-Mateos A, Spencer JP, Tognolini M, Borges G, Crozier A. Dietary (poly)phenolics in human health: structures, bioavailability, and evidence of protective effects against chronic diseases. Antiox Redox Signal. (2013) 18:1818–92. doi: 10.1089/ars.2012.4581
25. Filosa S, Di Meo F, Crispi S. Polyphenols-gut microbiota interplay and brain neuromodulation. Neural Regen Res. (2018) 13:2055–9. doi: 10.4103/1673-5374.241429
26. Cardona F, Andres-Lacueva C, Tulipani S, Tinahones FJ, Queipo-Ortuno MI. Benefits of polyphenols on gut microbiota and implications in human health. J Nutr Biochem. (2013) 24:1415–22. doi: 10.1016/j.jnutbio.2013.05.001
27. Kawabata K, Yoshioka Y, Terao J. Role of intestinal microbiota in the bioavailability and physiological functions of dietary polyphenols. Molecules. (2019) 24:370. doi: 10.3390/molecules24020370
28. Zhang F, Berg M, Dierking K, Félix MA, Shapira M, Samuel BS, et al. Caenorhabditis elegans as a model for microbiome research. Front Microbiol. (2017) 8:485. doi: 10.3389/fmicb.2017.00485
29. Sánchez-Blanco A, Rodríguez-Matellán A, González-Paramás A, González-Manzano S, Kim SK, Mollinedo F. Dietary and microbiome factors determine longevity in Caenorhabditis elegans. Aging. (2016) 8:1513–39. doi: 10.18632/aging.101008
30. Lee J, Yun HS, Cho KW, Oh S, Kim SH, Chun T, et al. Evaluation of probiotic characteristics of newly isolated Lactobacillus spp.: immune modulation and longevity. Int J Food Microbiol. (2011) 148:80–6. doi: 10.1016/j.ijfoodmicro.2011.05.003
31. Zhao Y, Zhao L, Zheng X, Fu T, Guo H, Ren F. Lactobacillus salivarius strain FDB89 induced longevity in Caenorhabditis elegans by dietary restriction. J Microbiol. (2013) 51:183–8. doi: 10.1007/s12275-013-2076-2
32. Sim I, Park KT, Kwon G, Koh JH, Lim YH. Probiotic potential of Enterococcus faecium isolated from chicken cecum with immunomodulating activity and promoting longevity in Caenorhabditis elegans. J Microbiol Biotechnol. (2018) 28:883–92. doi: 10.4014/jmb.1802.02019
33. Grompone G, Martorell P, Llopis S, González N, Genovés S, Mulet AP, et al. Anti-inflammatory Lactobacillus rhamnosus CNCM I-3690 strain protects against oxidative stress and increases lifespan in Caenorhabditis elegans. PLoS One. (2012) 7:52493. doi: 10.1371/journal.pone.0052493
34. Martorell P, Llopis S, González N, Chenoll E, López-Carreras N, Aleixandre A, et al. Probiotic strain bifidobacterium animalis subsp. lactis CECT 8145 reduces fat content and modulates lipid metabolism and antioxidant response in Caenorhabditis elegans. J Agric Food Chem. (2016) 64:3462–72. doi: 10.1021/acs.jafc.5b05934
35. Ikeda T, Yasui C, Hoshino K, Arikawa K, Nishikawa Y. Influence of lactic acid bacteria on longevity of Caenorhabditis elegans and host defense against Salmonella enterica serovar Enteritidis. Appl Environ Microbiol. (2007) 73:6404–9. doi: 10.1128/AEM.00704-07
36. Lee J, Choe J, Kim J, Oh S, Park S, Kim S, et al. Heat-killed Lactobacillus spp cells enhance survivals of Caenorhabditis elegans against Salmonella and Yersinia infections. Lett Appl Microbiol. (2015) 61:523–30. doi: 10.1111/lam.12478
37. Komura T, Yasui C, Miyamoto H, Nishikawa Y. Caenorhabditis elegans as an alternative model host for Legionella pneumophila, and protective effects of bifidobacterium infantis. Appl Environ Microbiol. (2010) 76:4105–8. doi: 10.1128/AEM.03021-09
38. You YS, Cha YJ, Choi SS. Effect of lactic acid bacteria on intestinal E. coli in Caenorhabditis elegans. Food Sci Biotechnol. (2015) 24:1853–8. doi: 10.1007/s10068-015-0242-5
39. Park MR, Ryu S, Maburutse BE, Oh NS, Kim SH, Oh S, et al. Probiotic lactobacillus fermentum strain JDFM216 stimulates the longevity and immune response of Caenorhabditis elegans through a nuclear hormone receptor. Sci Rep. (2018) 8:7441. doi: 10.1038/s41598-018-25333-8
40. Kumar A, Joishy T, Das S, Kalita MC, Mukherjee AK, Khan MR. A potential probiotic Lactobacillus plantarum JBC5 improves longevity and healthy aging by modulating antioxidative, innate immunity and serotonin-signaling pathways in Caenorhabditis elegans. Antioxidants. (2022) 11:268. doi: 10.3390/antiox11020268
41. Fasseas MK, Fasseas C, Mountzouris KC, Syntichaki P. Effects of Lactobacillus salivarius, Lactobacillus reuteri, and Pediococcus acidilactici on the nematode Caenorhabditis elegans include possible antitumor activity. Appl Microbiol Biotechnol. (2013) 97:2109–18. doi: 10.1007/s00253-012-4357-9
42. Schifano E, Zinno P, Guantario B, Roselli M, Marcoccia S, Devirgiliis C, et al. The foodborne strain lactobacillus fermentum MBC2 triggers pept-1-dependent pro-longevity effects in Caenorhabditis elegans. Microorganisms. (2019) 7:45. doi: 10.3390/microorganisms7020045
43. Nakagawa H, Shiozaki T, Kobatake E, Hosoya T, Moriya T, Sakai F, et al. Effects and mechanisms of prolongevity induced by Lactobacillus gasseri SBT2055 in Caenorhabditis elegans. Aging Cell. (2016) 15:227–36. doi: 10.1111/acel.12431
44. Martorell P, Alvarez B, Llopis S, Navarro V, Ortiz P, Gonzalez N, et al. Heat-treated Bifidobacterium longum CECT-7347: a whole-cell postbiotic with antioxidant, anti-inflammatory, and gut-barrier protection properties. Antioxidants. (2021) 10:536. doi: 10.3390/antiox10040536
45. Surco-Laos F. Evaluación De La Actividad De Flavonoides Y Sus Metabolitos En El Organismo Modelo Caenorhabditis Elegans. Ph.D. thesis. Salamanca: Universidad de Salamanca (2011).
46. Dueñas M, Surco-Laos F, González-Manzano S, González-Paramás AM, Gómez-Orte E, Cabello J, et al. Deglycosylation is a key step in biotransformation and lifespan effects of quercetin-3-O-glucoside in Caenorhabditis elegans. Pharmacol Res. (2013) 76:41–8. doi: 10.1016/j.phrs.2013.07.0011
47. Saul N, Pietsch K, Menzel R, Stürzenbaum SR, Steinberg CE. Catechin induced longevity in C. elegans: from key regulator genes to disposable soma. Mech Ageing Dev. (2009) 130:447–86. doi: 10.1016/j.mad.2009.05.005
48. Pietsch K, Saul N, Menzel R, Stürzenbaum SR, Steinberg CE. Quercetin mediated lifespan extension in Caenorhabditis elegans is modulated by age-1, daf-2, sek-1 and unc-43. Biogerontology. (2009) 10:565–78. doi: 10.1007/s10522-008-9199-6
49. Surco-Laos F, Dueñas M, González-Manzano S, Cabello J, Santos-Buelga C, González-Paramás AM. Influence of catechins and their methylated metabolites on lifespan and resistance to oxidative and thermal stress of Caenorhabditis elegans and epicatechin uptake. Foos Res Int. (2012) 46:514–21.
50. Ayuda-Durán B, González-Manzano S, Miranda-Vizuete A, Dueñas M, Santos-Buelga C, González-Paramás AM. Epicatechin modulates stress-resistance in C. elegans via insulin/IGF-1 signaling pathway. PLoS One. (2019) 14:e0199483. doi: 10.1371/journal.pone.0199483
51. Ayuda-Durán B, González-Manzano S, Miranda-Vizuete A, Sánchez-Hernández E, Romero MR, Dueñas M, et al. Exploring target genes involved in the effect of quercetin on the response to oxidative stress in Caenorhabditis elegans. Antioxidants. (2019) 8:585. doi: 10.3390/antiox8120585
52. Saul N, Pietsch K, Menzel R, Steinberg CE. Quercetin- mediated longevity in C. elegans: is DAF-16 involved? Mech Ageing Dev. (2008) 129:611–3. doi: 10.1016/j.mad.2008.07.001
53. Kampkötter A, Timpel C, Zurawski RF, Ruhl S, Chovolou Y, Proksch P, et al. Increase of stress resistance and lifespan of Caenorhabditis elegans by quercetin. Comp Biochem Physiol B Biochem Mol Biol. (2008) 149:314–23. doi: 10.1016/j.cbpb.2007.10.004
54. Surco-Laos F, Cabello J, Gómez-Orte E, González-Manzano S, González-Paramás AM, Santos-Buelga C, et al. Effects of O-methylated metabolites of quercetin on oxidative stress, thermotolerance, lifespan and bioavailability on Caenorhabditis elegans. Food Funct. (2011) 2:445–56.
Keywords: gut microbiota, epicatechin, quercetin, probiotics, Lactobacillus, Bifidobacterium, Enterococcus
Citation: Ayuda-Durán B, Sánchez-Hernández E, González-Manzano S, Santos-Buelga C and González-Paramás AM (2022) The effects of polyphenols against oxidative stress in Caenorhabditis elegans are determined by coexisting bacteria. Front. Nutr. 9:989427. doi: 10.3389/fnut.2022.989427
Received: 08 July 2022; Accepted: 10 November 2022;
Published: 01 December 2022.
Edited by:
J. Fernando Ayala-Zavala, Consejo Nacional de Ciencia y Tecnología (CONACYT), MexicoReviewed by:
Danyue Daisy Zhao, The Hong Kong Polytechnic University, Hong Kong SAR, ChinaYessica Enciso-Martínez, University of Sonora, Mexico
Melvin R. Tapia-Rodriguez, Instituto Tecnológico de Sonora (ITSON), Mexico
Copyright © 2022 Ayuda-Durán, Sánchez-Hernández, González-Manzano, Santos-Buelga and González-Paramás. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ana M. González-Paramás, cGFyYW1hc0B1c2FsLmVz
†These authors have contributed equally to this work