 Alfredo Fernandez-Quintela1,2,3†María Teresa Macarulla1,2,3†
Alfredo Fernandez-Quintela1,2,3†María Teresa Macarulla1,2,3† Saioa Gómez-Zorita1,2,3*Marcela González4
Saioa Gómez-Zorita1,2,3*Marcela González4 Iñaki Milton-Laskibar1,2,3*
Iñaki Milton-Laskibar1,2,3* María P. Portillo1,2,3
María P. Portillo1,2,3- 1Nutrition and Obesity Group, Department of Nutrition and Food Science, University of the Basque Country (UPV/EHU) and Lucio Lascaray Research Institute, Vitoria-Gasteiz, Spain
- 2Bioaraba Health Research Institute, Vitoria-Gasteiz, Spain
- 3CIBERobn Physiopathology of Obesity and Nutrition, Institute of Health Carlos III, Vitoria-Gasteiz, Spain
- 4Nutrition and Food Science Department, Faculty of Biochemistry and Biological Sciences, National University of Litoral and National Scientific and Technical Research Council (CONICET), Santa Fe, Argentina
Although a general healthy gut microbiota cannot be defined due to numerous internal and external individual factors, such as sex, age, ethnicity, genetics, environment, diet and drugs affect its composition, certain microbial species and gut microbiota compositions seem to be related to the progression of insulin resistance to type 2 diabetes, as well as the development of microvascular and macrovascular complications of diabetes. The present review aimed at gathering the reported information describing how resveratrol induced changes in microbiota composition can mediate the positive effects of this polyphenol on glucose homeostasis under type 2 diabetic conditions, both in animals and humans. Based on the fact that some changes observed in the gut microbiota of type 2 diabetic animals and patients are reversed by resveratrol treatment, and taking into account that some resveratrol mediated changes in gut microbiota composition are similar to those induced by anti-diabetic drugs such as metformin, it can be proposed that four genera, Alistipes, Allobaculum, Desulfovibrio and Blautia could be involved in the benefits of resveratrol on glycameic control. Nevertheless some limitations are observed in this research field: (a) the number of studies analyzing both the effects of resveratrol on glucose homeostasis and microbiota composition in the same cohort of animals, in order to know the potential involvement of microbiota in the anti-diabetic effects of this phenolic compound, are very scarce and practically inexistent in the case of humans., (b) the studies present inconsistencies concerning the effects of resveratrol on gut microbiota changes, (c) the experimental design used do not allow the researchers to establish a causal relationship between the changes in microbiota and the anti-diabetic effect, in the vast majority of the studies, (d) the knowledge about the role of each type of bacteria on glycaemic control is not sufficient so far.
Introduction
Type 2 diabetes mellitus is a non-communicable disease characterized by an elevated blood glucose level or hyperglycaemia. Several risk factors such as a family history of diabetes, poor eating habits and being overweight or obese have been identified. In 2019, about 463 million people suffered diabetes worlwide, and future estimates predict that by 2045, the number of diabetic patients will exceed 700 million (1).
The intestinal microbiota could be defined as the complex community of microorganisms that inhabit our gastrointestinal tract (2, 3). It is mainly composed of bacteria, with the most common being the phyla Bacteroidetes, Clostridium, Prevotella, Eubacterium, Ruminococcus, Fusobacterium, Peptococcus and Bifidobacterium (4). In addition, other microorganisms, such as viruses and fungi are also part of the intestinal microbiota, although in a much smaller proportion. It is known that gut microbiota plays a major role in various biological processes as well as being involved in the development of several diseases (5). Indeed, although a general healthy gut microbiota cannot be defined due to the fact that numerous internal and external individual factors, including sex, age, ethnicity, genetics, environment, diet and drugs affect its composition (6–9), certain species and combinations of them seem to be related to, or associated with specific diseases (10).
Regarding type 2 diabetes mellitus and insulin resistance, some studies have revealed a significant influence of gut microbiota on insulin signaling, inflammation, and glucose homeostasis (11–13). In addition, it has been reported that dysbiosis (the impairment of gut microbiota composition and gut barrier function) accompanies the progression of insulin resistance to type 2 diabetes, as well as the development of microvascular (retinopathy, nephropathy, and neuropathy) and macrovascular (atherosclerosis) complications of diabetes (14). The dysbiosis found in type 2 diabetic subjects is characterized by a decrease in butyrate-producing bacteria and an increase in opportunistic pathogens (11). Dysbiosis also involves the modification of intestinal integrity, islet inflammation and insulin signaling, at least in part, by means of metabolites produced by bacteria (short-chain fatty acids or secondary bile acids), which interact with receptors on epithelial, fat, muscle, liver, pancreatic and cardiac cells.
Very recently, Letchumanan et al. (13) have published a systematic review devoted to analyzing the existing evidence related to gut microbiota composition and diversity in individuals with pre-diabetes and newly diagnosed individuals with type 2 diabetes mellitus, in comparison to control subject. In this review, the authors pointed out that, despite the studies found significant associations between clinical biomarkers and the abundance of specific bacterial groups, a great heterogeneity in methodology and inconsistencies in the results were found among the eighteen studies gathered, with the exception of the correlation between glycaemic markers and Lactobacillus abundance. By focusing on changes that were similarly reported in two or more studies, they observed that the abundance of certain bacteria was increased (Lactobacillus, Streptococcus, Escherichia, Veillonella and Collinsella) or decreased (Roseburia, Dialister, Flavonifractor, Alistipes, Haemophilus and Akkermansia muciniphila) in prediabetic and diabetic subjects. In other studies, high relative abundances of some opportunistic pathogens, such as Bacteroides caccae, Clostridium hathewayi, Clostridium ramosum, Clostridium symbiosum, and Escherichia coli, as well as low relative abundances of butyrate-producing bacteria (Roseburia intestinalis and Faecalibacterium prausnitzii) have been observed (15, 16). Moreover, some studies carried out in rodents have confirmed that the abundances of Allobaculum and Lactobacillus diminish whereas those of Bacteroides, Veillonella, Lachnoclostridium, Parasutterella, Bifidobacterium, Helicobacter, Cupriavidus, Halomonas, Odoribacter, Enterococcus and Shigella increase in type 2 diabetic animals (17–19).
As previously indicated in this section, microbiota composition very much depends on the diet. In particular, polyphenols, which are chemical compounds naturally present in some foodstuffs, such as fruits, vegetables and nuts, among others, have been demonstrated to strongly interact with microbiota, thus leading to significant changes on its composition. In addition, these compounds, among which resveratrol (3,5,4’-trihydroxy-trans-stilbene) stands out, have shown beneficial effects on health in numerous pre-clinical studies, as well as in epidemiological studies and clinical trials.
Regarding type 2 diabetes mellitus, it has been reported that resveratrol improves insulin sensitivity and reduces blood glucose in cell cultures and animal models (20). The recent meta-analysis published by García-Martínez et al. (21) shows that in several clinical trials, resveratrol increases insulin sensitivity and decreases blood glucose levels either in subjects featuring insulin resistance or type 2 diabetes mellitus (22, 23; Figure 1). However, a general consensus has not been achieved since there are also some clinical trials in which these positive effects of resveratrol have not been observed. These discrepancies have been attributed to the wide range of doses used (5–5,000 mg/day), as well as to other factors, such as the age and the metabolic status of the participants. The general conclusion of this meta-analysis is that resveratrol has a significant dose-response effect on glucose concentrations, HbA1c percentage and insulin levels in subjects with type 2 diabetes mellitus, aged 45–59 years.
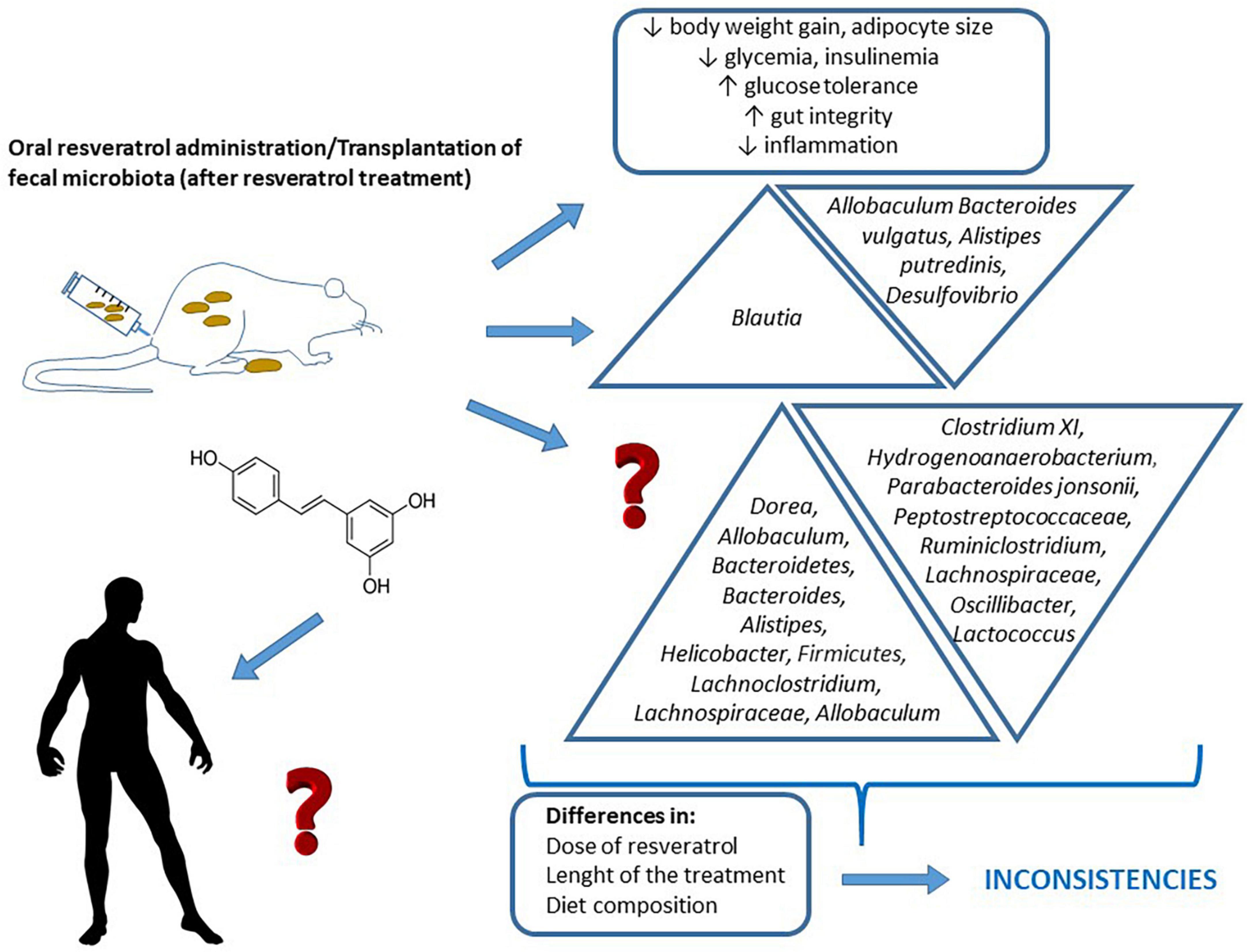
Figure 1. Changes induced by resveratrol administration, or fecal matter transplantations from animals treated with resveratrol, on glycaemic control and gut microbiota composition. The figure also shows the differences among experimental designs that sometimes lead to insistencies in the obtained results, and the lack of studies addressed in humans.
Among the mechanisms underlying the anti-diabetic effects of resveratrol, increased insulin sensitivity, activation of 5′-AMP-activated protein kinase (AMPK), epigenetic modifications of DNA sequence (including methylation and histone modifications) leading to changes in gene expression, and improvement of pancreatic β-cell functionality, due to the protection against oxidative damage, have been described (24–27). In addition, several authors have proposed that changes induced by resveratrol in gut microbiota composition may well be involved on its anti-diabetic effect.
In this context, the aim of the present review is to gather the information reported so far describing how the changes induced by resveratrol in gut microbiota composition can mediate the positive effects of this polyphenol on glucose homeostasis under type 2 diabetic conditions, both in animals and humans.
Animal studies addressing the effects of resveratrol administration on glycemic control and gut microbiota
Although there are a great number of studies aimed at analyzing the effect of resveratrol on glycaemic control or to determine how resveratrol administration modifies gut microbiota, only a reduced number of such studies have analyzed both aspects at the same time (in the same cohort of animals), in order to shed light onto the involvement of microbiota changes in the anti-diabetic effect of this polyphenol. These studies have been carried out in both mice and rats (Table 1 and Figure 1).
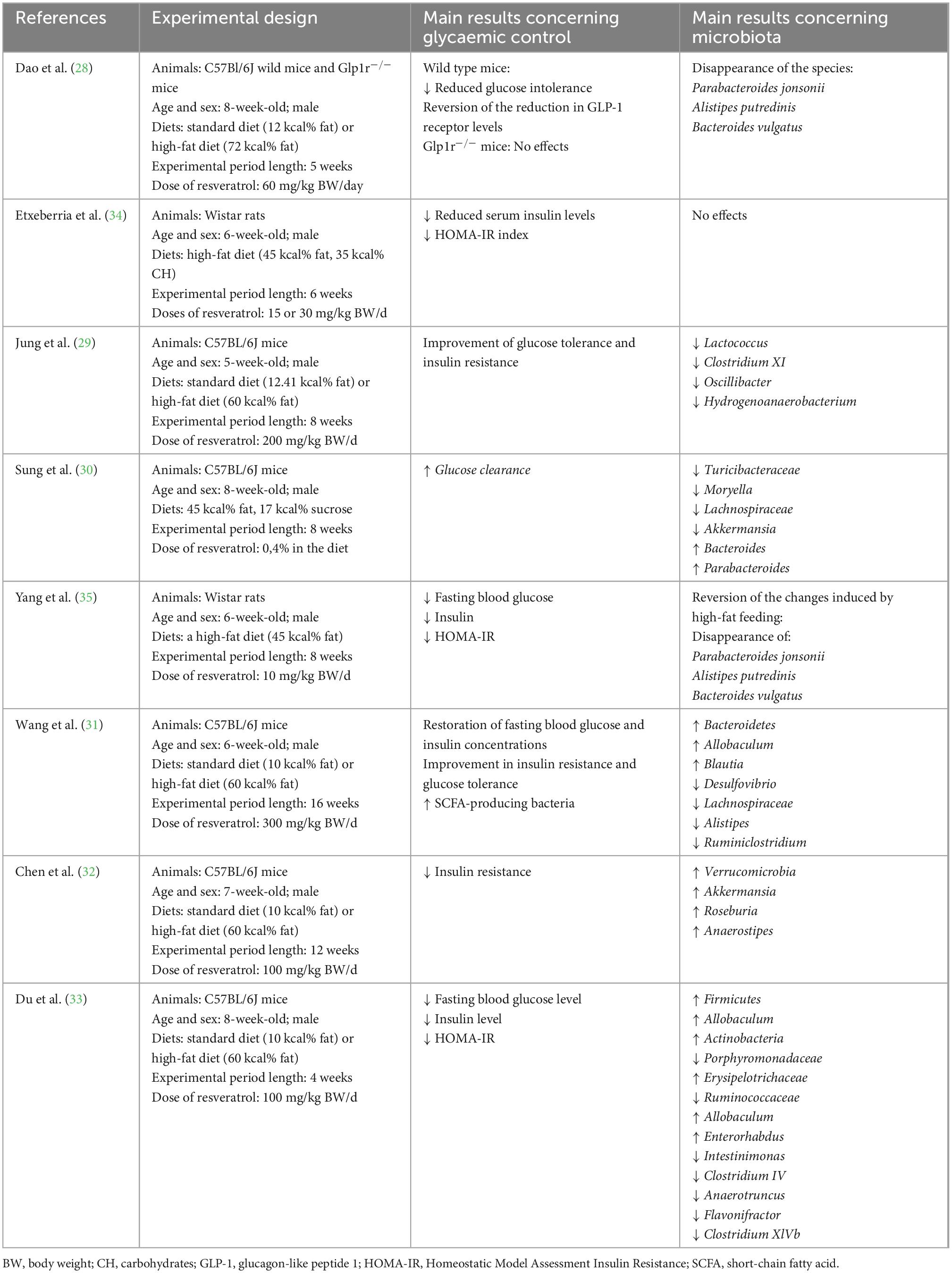
Table 1. Effects of resveratrol administration on glycaemic control and gut microbiota composition.
As far as the studies conducted in mice, Dao et al. (28) studied the involvement of glucagon-like peptide-1 (GLP-1) and its receptor on the effect of resveratrol on glycaemia. For this purpose, they worked with two mice models, wild-type C57Bl/6J and Glp1r-/- (mice with a functional disruption of the Glp1 receptor). One group of each model was fed a standard diet (12% of energy from fat) and another a high-fat diet (72% of energy from fat), which induced glucose intolerance before the onset of obesity, and reduced plasma GLP-1 levels. Half of the animals fed the high-fat diet were treated with resveratrol (60 mg/kg body weight/d), while the other half did not receive the polyphenol. In the wild-type model fed the high-fat diet, the authors observed that resveratrol treatment reduced glucose intolerance (as determined by a glucose tolerance test), tripled the concentration of active GLP-1 in the portal vein, increased intestinal glucagon and active GLP-1 and increased insulin concentration during the glucose tolerance test. In contrast, Glp1r-/- mice were insensitive to resveratrol treatment, revealing an essential role of GLP-1 receptor in the control of glucose tolerance by this polyphenol. In addition, in the peripheral organs, resveratrol reduced inflammation, in part by increasing interleukin 10 (IL-10) production in the colon, liver and muscle.
In terms of gut microbiota, the high-fat diet induced great alterations on its composition, while resveratrol treatment normalized them. In particular, the species Parabacteroides jonsonii DMS 18315, Alistipes putredinis DMS 17216 and Bacteroides vulgatus ATCC 8482, which were present in the animals fed the high-fat diet, disappeared in the resveratrol-treated animals. The authors concluded that the GLP-1 receptor is required to mediate resveratrol anti-diabetic effect, and that the mechanism(s) by which GLP-1 secretion is restored may be related to a change in gut microbiota and inflammation.
In another study, Jung et al. (29) fed C57BL/6J mice either a standard (12.41% of energy from fat) or a high-fat diet (60% of energy from fat) and treated or not with resveratrol (200 mg/kg body weight/d), for 8 weeks. The authors studied the involvement of mammalian target of rapamycin (mTOR), which acts as a tyrosine protein kinase that promotes the activation of insulin receptors and insulin-like growth factor 1 receptors. The results showed that the polyphenol prevented glucose intolerance in mice fed the high-fat diet. Regarding gut microbiota, they observed that the high-fat diet increased the relative abundances of the genera Lactococcus, Oscillibacter, Hydrogenoanaerobacterium and Clostridium cluster XI. After resveratrol treatment, the abundance of these bacteria decreased. The authors observed that changes in gut microbiota composition caused by resveratrol treatment correlated with alterations in insulin resistance and gut inflammation, concluding that mTOR signaling appeared to be a key component of the regulation of gut microbiota composition.
In the study reported by Sung et al. (30), C57BL/6N mice were divided into four groups: two groups were fed a standard diet, supplemented or not with resveratrol (0.4% in the diet), and two groups were fed a diet rich in fat and sucrose (HFHS, 45% of energy from fat and 17% of energy from sucrose) supplemented or not with resveratrol (0.4% in the diet). The experimental period was of 8 weeks. Mice fed the HFHS diet showed lower glucose clearance in the glucose tolerance test than animals fed the standard diet, while mice fed the HSHS diet and treated with resveratrol showed an improved glucose clearance (represented by the AUC in glucose tolerance test) than non treated HFHS fed mice. With regard to microbiota composition, the authors observed that resveratrol administration produced several changes: a decreased abundance of the families Turicibacteraceae, Lachnospiraceae and the genera Moryella and Akkermansia, and an increased abundance of the genera Bacteroides and Parabacteroides.
Also using C57BL/6 J male mice Wang et al. (31) carried out an experiment feeding the animals either a standard diet (SD group) or a high-fat diet (HFD group; 60% of energy from fat), and administering (by gavage) resveratrol at a dose of 300 mg/kg body weight/d to the treated groups (groups SDR and HFDR), for 16 weeks. The authors observed that resveratrol supplementation improved insulin resistance in high-fat diet fed mice by restoring fasting blood glucose and insulin concentrations to nearly normal levels. Further, and as expected, microbiota composition also changed after the polyphenol administration. Relative abundances of the phylum Bacteroidetes and the genus Bacteroides were increased in both resveratrol-treated groups (SDR and HFDR groups). Moreover, mice treated with resveratrol showed lower abundances of Allobaculum, Desulfovibrio, Lachnospiraceae_NK4A136_group and Ruminiclostridium_9 and higher abundances of Helicobacter, Blautia and Lachnoclostridium.
More recently, Chen et al. (32) determined the effects of resveratrol on gut microbiota composition and intestinal tight-junction proteins in 6-week-old male C57BL/6J mice fed a standard or a high-fat diet (60% of energy from fat). Half of the animals received resveratrol at a dose of 100 mg/kg body weight/day for 12 weeks. Consumption of the high-fat diet induced hyperglycemia and insulin resistance, which were prevented by resveratrol administration. Regarding inflammation, resveratrol improved endotoxemia and secondary systemic inflammation by decreasing bacterial lipopolysaccharide, tumor necrosis factor α (TNF-α) and interleukin 1b (IL-1b). On the other hand, in relation to the structure of the tight junctions, the authors observed a decrease in the expression of the tight junction proteins claudin-1, occludin and Zo-1 in the colon of mice fed high-fat diet, which may contribute to the increased passage of bacterial endotoxins into the circulation. Concerning gut microbiota composition, at the phylum level, feces from the group fed the high-fat diet and treated with resveratrol showed increased abundance of Verrucomicrobia and at the genus level increased abundance of Akkermansia, Roseburia and Anaerostipes.
Du et al. (33), carried out a study to test the effects of resveratrol in a non-alcoholic fatty liver disease (NAFLD) mouse model. For this purpose, they used male C57BL/6J mice randomly assigned to the control group (NC, fed a normal chow diet), to the HFD group (fed a high-fat diet providing 60% of energy from fat) or to the RSV group (fed the same high-fat diet and treated with 100 mg/kg body weight/d of resveratrol), for 4 weeks. Resveratrol improved glucose homeostasis by reducing insulin levels, fasting blood glucose, and HOMA-IR. Regarding gut microbiota composition, several changes were induced by the high-fat feeding. Thus, at the phylum level, the relative abundance of Firmicutes was significantly increased and that of Bacteroidetes significantly decreased. At the family level, Porphyromonadaceae was decreased and Lachnospiraceae and Ruminococcaceae were elevated. At the genus level, decreases in Barnesiella and Parasutterella, and increases in Intestinimonas, Enterorhabdus, Oscillibacter, Clostridium IV, Pseudoflavonifractor, Anaerotruncus, Clostridium XlVb, and Peptococcus relative abundances were observed. When comparing gut microbiota composition between HFD and RSV groups, it was observed that resveratrol increased Firmicutes abundance at the phylum level and Allobaculum abundance at the genus level. Correlation analysis revealed that Firmicutes, Lachnospiraceae, Pseudoflavonifractor, Intestinimonas, Ruminococcaceae, Flavonifractor, Clostridium XlVb, Peptococcus, Oscillibacter, Anaerotruncus, and Hydrogenoanaerobacterium were positively correlated to fasting blood glucose and HOMA-IR index, while Barnesiella, Bacteroidetes, Porphyromonadaceae, Parasutterella, and Allobaculum were negatively correlated.
In addition, other studies have also been addressed in rats. In our laboratory a study was carried out to analyze the effect of two resveratrol doses (15 or 30 mg/kg body weight/d) on glycaemic control in rats fed a high-fat high-sucrose diet (45% of energy as fat, 17 g sucrose/100 g diet) for 6 weeks. Resveratrol administration effectively reduced serum insulin levels and improved insulin resistance (determined by the HOMA-IR index), but no changes in microbiota composition were observed (34).
In the study reported by Yang et al. (35) using rats, the animals were randomly distributed in three experimental groups: a control group fed a standard diet (10% of energy from fat); a group fed a high-fat diet (HF; 45% of energy from fat) and a group, fed a high-fat diet and supplemented with resveratrol (HFR; 10 mg/kg body weight/day). The experimental period length was of 8 weeks. Plasma glucose and insulin levels were reduced after resveratrol supplementation as compared to the non-supplemented group, meaning that insulin resistance was reduced.
Regarding gut microbiota composition, alpha diversity indexes were significantly increased in resveratrol-treated rats. Due to the decline of Bacteroides, the relative ratio of Firmicutes to Bacteroidetes showed an increase in the group treated with resveratrol, when compared to the control and HF groups. At the family level, resveratrol-supplemented rats (HFR group) showed the lowest S24-7 and Peptostreptococcaceae abundances. No significant changes in the relative abundance of Ruminococcaceae, Clostridiaceae, Bacteroidaceae or Desulfovibrionaceae were detected among the experimental groups. At the genus level, the HFR group was enriched in the butyrate producer Blautia and Dorea when compared to the HF group, which contributed to a sharp increase in the Lachnospiraceae family.
Human studies addressing the effects of resveratrol administration on glycemic control and gut microbiota
Walker et al. (36) investigated whether the administration of high doses of resveratrol to obese individuals featuring insulin resistance and metabolic syndrome could improve certain parameters such as insulin sensitivity and glucose tolerance. Moreover, they also analyzed intestinal microbiota composition. For this purpose, obese men with a body mass index of 30–40 kg/m2 and aged between 30 and 70 years were studied. The work was conducted in two experimental groups using a placebo-controlled, double blind, randomized, parallel-group pilot study design. The patients were fed for 4 days an isocaloric Western diet, providing the calories needed to maintain a stable weight. On day four and for 30 days, twice daily, subjects in the resveratrol group began receiving two 500 mg capsules of Mega-RES 99% (made exclusively from organically grown Japanese root) and those in the placebo group, two 500 mg placebo capsules. Only minor overall effects of resveratrol were demonstrated in obese men with insulin resistance and metabolic syndrome. Resveratrol did not induce changes in insulin resistance, but it did reduce the 120-min time point and the area under the 2-h glucose tolerance test glucose concentration curve. Several changes in gut microbiota composition were also induced by the polyphenol. Thus, a drop in the relative abundances of family Rikenellaceae and the genera Ruminococcus, Oscillospira, Clostridium, Alistipes, Odoribacter and Butyricimonas were found. Moreover, the relative abundances of the phylum Gammaproteobacteria, the family Gemellaceae and the genera Turicibacter and Atopobium were increased.
Animal studies addressing the effects of fecal transplantation from resveratrol-treated animals on glycemic control and gut microbiota
The studies described in the previous section do not establish a causal relationship between changes in microbiota and changes in glucose homeostasis induced by resveratrol. However, in the literature, there are some studies in which fecal matter transplantations from animals treated with resveratrol to non- treated animals, showing alterations in glucose homeostasis, have been carried out to gain more insight about the potential involvement of microbiota in the anti-diabetic effects of resveratrol (Table 2 and Figure 1).
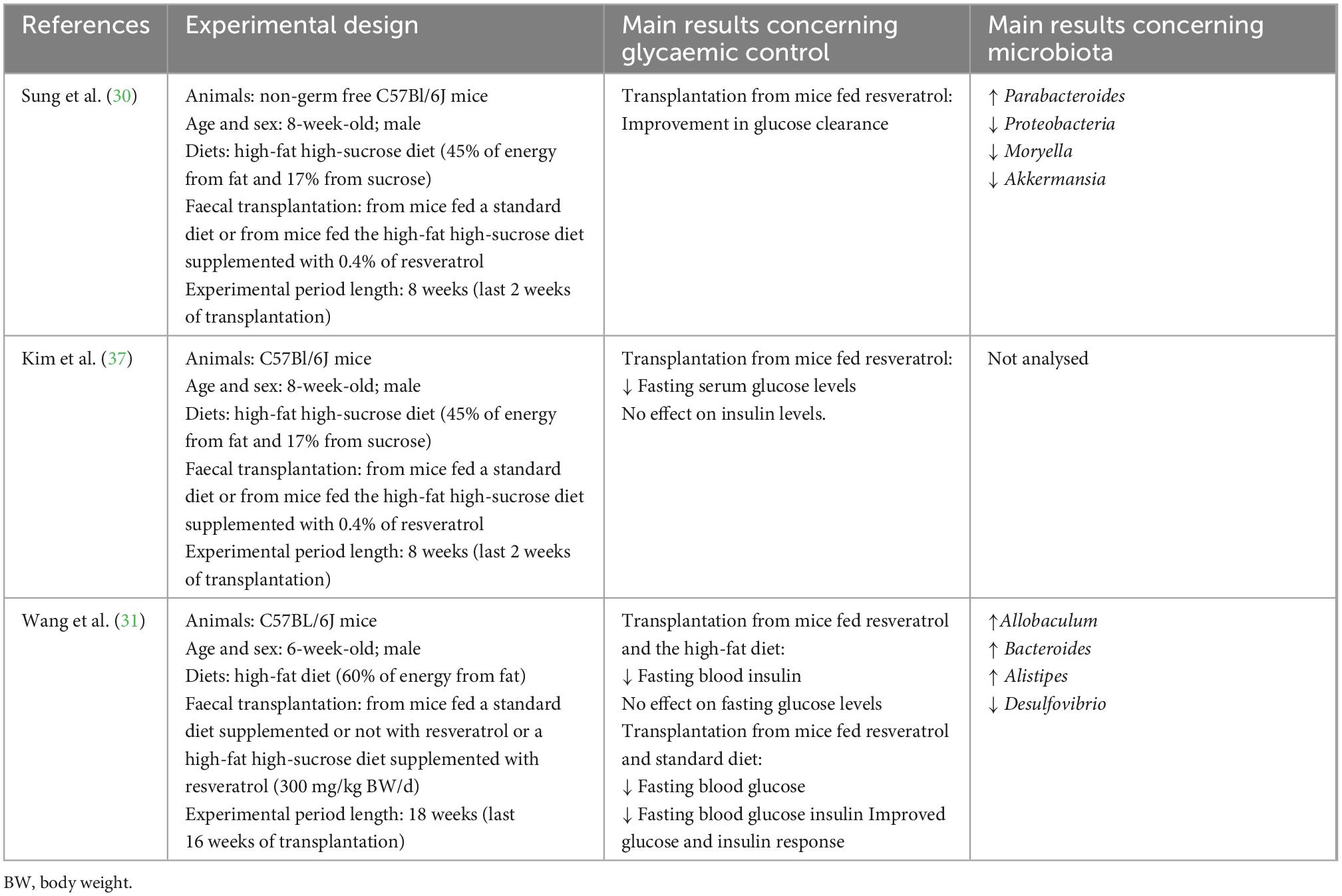
Table 2. Effects induced by fecal matter transplantations from animals treated with resveratrol on gut microbiota composition.
In this line, Sung et al. (30) fed a cohort of C57BL/6N mice a high-fat high-sucrose diet (45% of energy from fat and 17% from sucrose) for 5 weeks and then, by oral gavage, they administered three fecal matter transplantations (FMTs) during another 2 weeks, collected from donor mice fed either a chow diet (without resveratrol) or a high-fat high-sucrose diet supplemented with resveratrol (0.4%) for 8 weeks. One week after the final resveratrol-FMT dose, animals showed clear improvements in glucose clearance, whereas no effects were found in the control-FMTs animals. An improvement in glucose clearance was also observed in obese mice receiving FMT from donor mice maintained on a high-fat high-sucrose diet supplemented with resveratrol. This improvement in glucose homeostasis occurred in the absence of a reduction in body weight. Moreover, the obese mice that received the FMT showed higher levels of bacteria from the genus Parabacteroides and lower relative abundance of the phylum Proteobacteria and the genera Moryella and Akkermansia, which was similar to the results obtained with oral resveratrol administration.
In the study reported by Kim et al. (37), the authors fed C57BL/6N mice a high-fat high-sucrose diet (45% of energy from fat and 17% from sucrose) for 5 weeks. These mice were assigned either to a group receiving fecal microbiome fecal matter transplantations from donor mice fed a chow diet (FMTs) or a group receiving fecal microbiome fecal matter transplantations from donor mice fed a high-fat high-sucrose diet supplemented with 4 g of resveratrol/kg diet (Resv-FMT) by oral gavage, on days 1, 3, and 5 after an overnight fast. Mice receiving Resv-FMTs showed a marked decrease in fasting serum glucose levels, but not in insulin level, in the absence of differences in body weight. Together, these data suggest that Resv-FMTs improved glucose homeostasis by improving glucose uptake in peripheral tissues through increased sensitivity to insulin and that this effect was not secondary to weight loss. The authors tested the influence of Resv-FMTs transplant on colon inflammatory parameters in order to propose a mechanism underlying its beneficial effect on insulin sensitivity. Resv-FMT recipients showed significantly lower levels of TNF-α, CXCL1/KC (murine IL-8 homologue), and IL-1β, demonstrating reduced colon inflammation. In addition, the values obtained in a glucose tolerance test were positively correlated with the levels of TNF-α. Furthermore, the authors analyzed 23 short-chain fatty acids (SCFA) in the fecal matter of donors. This analysis revealed that the concentration of 4-hydroxyphenylacetate was significantly higher in the feces of resveratrol-fed donor mice than in the feces of the chow-fed donors. In addition, the levels of formate, isobutyrate, and isovalerate were significantly lower in the feces of resveratrol-fed donor mice. Thus, the presence or lack of these SCFAs may potentially be associated with the observed physiological benefits in obese Resv-FMT recipient mice. In this study the authors did not analyzed microbiota composition.
In a recent study, Wang et al. (31) used the feces obtained from mice treated with resveratrol at a dose of 300 mg/kg body weight/d (experiment described in the previous section of this review) to make a fecal matter transplantation to mice fed a high-fat diet. The fasting blood glucose levels were significantly lower after the transplantation from mice fed a standard diet supplemented with resveratrol. Insulin concentration was significantly lower after the transplantation from mice fed a standard diet (supplemented or not with resveratrol) and the high-fat diet supplemented with resveratrol. Moreover, glucose and insulin responses improved after the transplantation from mice receiving the standard diet (supplemented or not). Additionally, the authors confirmed a positive effect on the intestinal barrier function after microbial transplantation. When the authors analyzed microbiota composition they observed that after transplantation, the microbiota of the recipient mice resembled, to a certain degree, that of their corresponding donor groups. Thus, at the genus level, fecal-treatment increased Allobaculum, Bacteroides and Alistipes abundances and decreased those of Desulfovibrio in mice.
Discussion
Resveratrol shows a low bioavailability, due to its low solubility in the intestine and its extensive phase II metabolism in the intestine and liver. For that reason, different strategies to increase its bioavailability have been studied. Solubility can be increased by using micronized resveratrol. In order to prevent resveratrol metabolism, this compound can be administered along with other molecules that inhibit glucouronidation and/or sulphation, such as piperine or quercetine. Another approach is to mask the prime targets of metabolism, that is the hydrophilic hydroxyl groups of resveratrol. Finally, another possibility is to use resveratrol nanoformulations (38). But, in spite of its poor biavailability, a large number of studies have shown the beneficial effects of resveratrol. One of the possible reasons is that its metabolites have biological activity, and another reason may be that resveratrol exerts its effects, in part, through the modulation of the intestinal microbiota.
In this context, and focusing the review on the beneficial effects of resveratrol on type 2 diabetes, the number of studies analyzing the effects of resveratrol on glucose homeostasis or microbiota composition are numerous. However, only a reduced number of them have addressed both issues in the same cohort of animals or humans, in order to know the potential involvement of microbiota in the anti-diabetic effects of this phenolic compound. All the studies included in this review, with the exception of that reported by Kim et al. (37) showed beneficial effects of oral resveratrol administration on glycaemic control, which is in good accordance with the vast majority of the reported studies addressing this issue (20). In the case of Kim’s work, the lack of effects could have been due to the instability of resveratrol, which, in contact with the diet, is degraded to a large extent (39). In contrast, the studies present inconsistencies in the findings concerning the effects of resveratrol on gut microbiota changes. Indeed, the changes observed in the studies are very different.
By analyzing the experiment designs used in the studies carried out in rodents, it is evident that important differences exist among parameters that play a crucial role in the influence of polyphenols on gut microbiota. One of them is the experimental period length, which is in the range 3–16 weeks. On the other hand, the used resveratrol doses and ways of administration are also different. Regarding the dose, this ranged from 15 to 200 mg/kg body weight/day of resveratrol. Moreover, resveratrol was administered either by oral gavage or included in the diet. As explained previously in this review, the diet is an important factor affecting gut microbiota composition; thus, the experimental diet may have conditioned the effect of resveratrol in this regard. The diets used in the studies were mainly high-fat diets providing 60% of energy as fat, but other types of diets, such as high-fat diets providing 45 or 72% of energy as fat, and diets rich in fat and sucrose have also been used. In addition, no information about the fatty acid composition of the dietary fat component was included in the articles included in this review. Thus, the potential differences among studies and their influence on the observed effects cannot be considered to discuss the differences observed among all the studies.
An important aspect to be considered is that, in some of these works, the authors limited the study to the description of changes in gut microbiota composition, without establishing a relationship between changes in microbiota and the improvement in glycaemic control. In other works, the authors analyzed the correlations between changes in gut microbiota and parameters related to glucose homeostasis control, but this analysis did not allow them to know if the changes observed in gut microbiota composition were a causal factor of the improvement in glucose homeostasis. Thus, unfortunately, a deeper discussion to explain which metabolic changes could be expected, based on the changes in gut microbiota, which could serve to explain the improvement in glycaemic control, was not provided.
In order to give more insight on this topic some researchers have addressed experiments based on FMT from animals treated with resveratrol to animals showing either insulin resistance or type 2 diabetes mellitus. This approach provides a stronger scientific evidence about the potential causality of microbiota composition changes. Concerning this type of studies, in that reported by Kim et al. (37) the authors did not characterize fecal microbiota composition, and thus no information concerning the potential bacteria responsible for the improvement of glucose homeostasis in mice receiving the FMTs was provided. In contrast, they analyzed 23 SCFA in the fecal matter of donors. According to this analysis, they suggested that the increased amount of 4-hydroxyphenylacetate and the decreased amounts of formate, isobutyrate and isovalerate may be responsible of the beneficial effects of Resv-FMT. In the study reported by Sung et al. (30) an increase in the genus Parabacteroides and a decrease in the phylum Proteobacteria and the genera Moryella and Akkermansia seem to be related to the improvement of glucose homeostasis induced by resveratrol. Wang et al. (31), analyzed microbiota composition and observed that after transplantation, the microbiota of the recipient mice partially resembled that of their corresponding donor groups. In this case, the changes in microbiota compostion related to the reductions observed in serum glucose and insulin levels were an increase in Allobaculum, Bacteroides and Alistipes abundances and a decrease in Desulfovibrio abundance.
As indicated in the Introduction section, certain bacteria are increased (Lactobacillus, Streptococcus, Escherichia, Veillonella, Collinsella, Bacteroides, Lachnoclostridium, Parasutterella and Bifidobacterium) and other are decreased (Faecalibacterium prausnitzii, Roseburia, Dialister, Flavonifractor, Allobaculum, Alistipes, Haemophilus and Akkermansia muciniphila) in pre-diabetic and diabetic subjects (human and/or rodents). Among these bacteria, Alistipes (31) and Allobaculum (31, 33) are reverse modified by resveratrol in two of the studies included in this review.
As far as Alistipes is concerned, although its role on glycaemic control has not been reported so far, in a study addressed in women showing gestational diabetes, statistical and computational analyses of the metagenomic data identified four potential bacterial markers associated with gestational diabetes, being Alistipes putredinis among them. Thus, women showing gestational diabetes presented lower abundance of this bacterium (40). The reduced abundance of Alistipes in two different types of diabetes, type 2 diabetes mellitus and gestational diabetes, reinforce the idea of its involvement in diabetes onset. On the other hand, it is well known that some species of this genus, such as Alistipes onderdonkii, seem to play an anti-inflammatory role in some diseases (41, 42). In fact, some authors observed that Alistipes was negatively correlated to TNF-α and lipopolysaccharide in type 2 diabetic rats, suggesting that it may suppress diabetic inflammation (43). Regarding the genus Allobaculum, it has been reported that calorie restriction treatment, a dietary intervention used to reduce body weight and improve insulin resistance and glycaemic control, increases its abundance. Other treatments that also improve glycaemic control, such as berberine and metformin administration (44), have also been reported to increase the abundance of this genus. Taken together, these data and those gathered in the present review, suggest that treatments able to ameliorate glycaemic control could act, at least in part, by increasing Allobaculum abundance.
There are two additional genera, Desulfovibrio and Blautia that decreased and increased, respectively, in some of the studies included in the present review (31, 35). Desulfovibrio, which is a genus of the phylum Proteobacteria, is known for its sulfate-reducing capacity, thereby producing toxic hydrogen sulfide, which can permeate the gut mucus barrier and increase inflammation levels (45). Taking that into account, and considering that a low-grade inflammation is associated with type 2 diabetes, the involvement of Desulfovibrio in the anti-diabetic effect of resveratrol could be proposed. Moreover, mice treated with liraglutide, an injectable GLP-1 receptor agonist used in the treatment of type 2 diabetes, has been shown to induce a recruitment of the genus Desulfovibrio (46). These findings support the potential involvement of Desulfovibrio in glucose homeostasis through the production of H2S.
Finally, it has been reported that the treatment with metformin in rodents leads to an improvement in glycaemic control, accompanied by an increase in the intestinal abundance of the genus Blautia in rats (44). This result has been confirmed in type 2 diabetic patients treated with this anti-diabetic drug (47). Moreover, the administration of a quality controlled herbal formula composed of eight herbs, namely, Rhizoma Anemarrhenae, Momordica charantia, Coptis chinensis, Salvia miltiorrhiza, red yeast rice, Aloe vera, Schisandra chinensis and dried ginger (Jiangyin Tianjiang Pharmaceutical Co., Ltd.), which induced significant amelioration in fasting glucose, HbA1c and HOMA in diabetic patients, also increased gut Blautia abundance (47). These results suggest that Blautia might be a target for the management of diabetes. In addition, it has been reported that Blautia is a common acetic acid producer in the intestine, which may inhibit insulin signaling in adipocytes by activating the G protein-coupled receptors GPR41 and GPR43, thus promoting the metabolism of unbound lipids and glucose in other tissues, and consequently alleviating obesity-related diseases such as type 2 diabetes. A cross-sectional study showed that Blautia, especially Blautia luti and Blautia wexlerae, probably help to reduce the inflammation associated with obesity-related complications. The beneficial effects of Blautia wexlerae are correlated with the production of S-adenosylmethionine, acetylcholine and l-ornithine and carbohydrate metabolism, resulting in the accumulation of amylopectin and production of succinate, lactate, and acetate. Consequently, Blautia wexlerae, modifies the gut environment, including the bacterial and SCFA composition of the gut microbiota (48).
Concluding remarks and perspectives
In view of the results presented in the studies included in the present review, it can be proposed that four genera, Alistipes, Allobaculum, Desulfovibrio and Blautia could be involved in the positive effects of resveratrol on glycameic control. Nevertheless, it is important to emphasize that this is mainly based on studies addressed in rodents. Moreover, some limitations are observed in this field of research. First of all, the number of studies analyzing both the effects of resveratrol on glucose homeostasis and microbiota composition in the same cohort of animals, in order to know the potential involvement of microbiota in the anti-diabetic effects of this phenolic compound, are very scarce and practically inexistent in the case of humans. Moreover, the studies present inconsistencies in the findings concerning the effects of resveratrol on gut microbiota changes. In addition, the experimental designs used do not allow the researchers to establish a causal relationship between the changes in microbiota and the anti-diabetic effect, in the vast majority of the studies. Finally, the knowledge about the role of each type of bacteria on glycaemic control is not sufficient so far.
Consequently, there is a clear need for further investigation to define the actual involvement of microbiota in the anti-diabetic effect of resveratrol. The first step should be to well establish the role of different types of bacteria in glucose metabolism and other metabolic processes related to insulin resistance and type 2 diabetes mellitus such as inflammation. Moreover, a more accurate characterization of the alterations in gut microbiota composition accompanying type 2 diabetes mellitus should be addressed. In addition, large scale randomized clinical trials devoted to analyzing the effects of resveratrol on glucose homeostasis and gut microbiota in very well defined cohorts, in order to limit inconsistencies among different studies, are required.
Author contributions
MP and IM-L: conceptualization. MP: supervision and funding acquisition. All authors: writing – original draft preparation, writing – reviewing and editing, and have read and agreed to the published version of the manuscript.
Funding
This study was supported by Instituto de Salud Carlos III (CIBERobn) under Grant CB12/03/30007 and the Government of the Basque Country (IT1482-22).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Saeedi P, Petersohn I, Salpea P, Malanda B, Karuranga S, Unwin N, et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: results from the international diabetes federation diabetes atlas, 9th edition. Diabetes Res Clin Pract. (2019) 157:107843. doi: 10.1016/j.diabres.2019.107843
2. Etxeberria U, Fernández-Quintela A, Milagro F, Aguirre L, Martínez J, Portillo M, et al. Impact of polyphenols and polyphenol-rich dietary sources on gut microbiota composition. J Agric Food Chem. (2013) 61:9517–33. doi: 10.1021/jf402506c
3. Lagier J, Khelaifia S, Alou M, Ndongo S, Dione N, Hugon P, et al. Culture of previously uncultured members of the human gut microbiota by culturomics. Nat Microbiol. (2016) 1:16203. doi: 10.1038/nmicrobiol.2016.203
4. O’Hara A, Shanahan F. The gut flora as a forgotten organ. EMBO Rep. (2006) 7:688–93. doi: 10.1038/sj.embor.7400731
5. Rowland I, Gibson G, Heinken A, Scott K, Swann J, Thiele I, et al. Gut microbiota functions: metabolism of nutrients and other food components. Eur J Nutr. (2018) 57:1–24. doi: 10.1007/s00394-017-1445-8
6. Lynch S, Pedersen O. The human intestinal microbiome in health and disease. N Engl J Med. (2016) 375:2369–79. doi: 10.1056/NEJMra1600266
7. Falony G, Joossens M, Vieira-Silva S, Wang J, Darzi Y, Faust K, et al. Population-level analysis of gut microbiome variation. Science. (2016) 352:560–64. doi: 10.1126/science.aad3503
8. Deschasaux M, Bouter K, Prodan A, Levin E, Groen A, Herrema H, et al. Depicting the composition of gut microbiota in a population with varied ethnic origins but shared geography. Nat Med. (2018) 24:1526–31. doi: 10.1038/s41591-018-0160-1
9. Simões C, Maganinho M, Sousa A. FODMAPs, inflammatory bowel disease and gut microbiota: updated overview on the current evidence. Eur J Nutr. (2022) 61:1187–98. doi: 10.1007/s00394-021-02755-1
10. Bresser L, de Goffau M, Levin E, Nieuwdorp M. Gut microbiota in nutrition and health with a special focus on specific bacterial clusters. Cells. (2022) 11:3091. doi: 10.3390/cells11193091
11. Baothman O, Zamzami M, Taher I, Abubaker J, Abu-Farha M. The role of gut microbiota in the development of obesity and diabetes. Lipids Health Dis. (2016) 15:108. doi: 10.1186/s12944-016-0278-4
12. Cani P. Human gut microbiome: hopes, threats and promises. Gut. (2018) 67:1716–25. doi: 10.1136/gutjnl-2018-316723
13. Letchumanan G, Abdullah N, Marlini M, Baharom N, Lawley B, Omar M, et al. Gut microbiota composition in prediabetes and newly diagnosed type 2 diabetes: a systematic review of observational studies. Front Cell Infect Microbiol. (2022) 12:943427. doi: 10.3389/fcimb.2022.943427
14. Martínez-López YE, Esquivel-Hernández DA, Sánchez-Castañeda JP, Neri-Rosario D, Guardado-Mendoza R, Resendis-Antonio O, et al. Type 2 diabetes, gut microbiome, and systems biology: a novel perspective for a new era. Gut Microbes. (2022) 14:2111952. doi: 10.1080/19490976.2022.2111952
15. Qin J, Li Y, Cai Z, Li S, Zhu J, Zhang F, et al. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature. (2012) 490:55–60. doi: 10.1038/nature11450
16. Karlsson F, Tremaroli V, Nookaew I, Bergström G, Behre C, Fagerberg B, et al. Gut metagenome in European women with normal, impaired and diabetic glucose control. Nature. (2013) 498:99–103. doi: 10.1038/nature12198
17. Beli E, Prabakaran S, Krishnan P, Evans-Molina C, Grant M. Loss of diurnal oscillatory rhythms in gut microbiota correlates with changes in circulating metabolites in type 2 diabetic db/db mice. Nutrients. (2019) 11:2310. doi: 10.3390/nu11102310
18. Zhang L, Zhang T, Sun J, Huang Y, Liu T, Ye Z, et al. Calorie restriction ameliorates hyperglycemia, modulates the disordered gut microbiota, and mitigates metabolic endotoxemia and inflammation in type 2 diabetic rats. J Endocrinol Invest. (2022). [Epub ahead of print]. doi: 10.1007/s40618-022-01914-3
19. Chen X, Chen C, Fu X. Hypoglycemic effect of the polysaccharides from Astragalus membranaceus on type 2 diabetic mice based on the “gut microbiota-mucosal barrier”. Food Funct. (2022) 13:10121–33. doi: 10.1039/d2fo02300h
20. Bagul P, Banerjee S. Application of resveratrol in diabetes: rationale, strategies and challenges. Curr Mol Med. (2015) 15:312–30. doi: 10.2174/1566524015666150505155702
21. García-Martínez B, Ruiz-Ramos M, Pedraza-Chaverri J, Santiago-Osorio E, Mendoza-Núñez V. Influence of age and dose on the effect of resveratrol for glycemic control in type 2 diabetes mellitus: systematic review and meta-analysis. Molecules. (2022) 27:5232. doi: 10.3390/molecules27165232
22. Oyenihi O, Oyenihi A, Adeyanju A, Oguntibeju O. Antidiabetic effects of resveratrol: the way forward in its clinical utility. J Diabetes Res. (2016) 2016:9737483. doi: 10.1155/2016/9737483
23. Öztürk E, Karaboğa Arslan A, Betül Yerer M, Bishayee A. Resveratrol and diabetes: a critical review of clinical studies. Biomed Pharmacother. (2017) 95:230–34. doi: 10.1016/j.biopha.2017.08.070
24. Szkudelski T, Szkudelska K. Anti-diabetic effects of resveratrol. Ann N Y Acad Sci. (2011) 1215:34–9. doi: 10.1111/j.1749-6632.2010.05844.x
25. Rouse M, Younès A, Egan J. Resveratrol and curcumin enhance pancreatic β-cell function by inhibiting phosphodiesterase activity. J Endocrinol. (2014) 223:107–17. doi: 10.1530/JOE-14-0335
26. Trepiana J, Milton-Laskibar I, Gómez-Zorita S, Eseberri I, González M, Fernández-Quintela A, et al. Involvement of 5′-activated protein kinase (AMPK) in the effects of resveratrol on liver steatosis. Int J Mol Sci. (2018) 19:3473. doi: 10.3390/ijms19113473
27. Gowd V, Kang Q, Wang Q, Wang Q, Chen F, Cheng K, et al. Resveratrol: evidence for its nephroprotective effect in diabetic nephropathy. Adv Nutr. (2020) 11:1555–68. doi: 10.1093/advances/nmaa075
28. Dao T, Waget A, Klopp P, Serino M, Vachoux C, Pechere L, et al. Resveratrol increases glucose induced GLP-1 secretion in mice: a mechanism which contributes to the glycemic control. PLoS One. (2011) 6:e20700. doi: 10.1371/journal.pone.0020700
29. Jung M, Lee J, Shin N, Kim M, Hyun D, Yun J, et al. Chronic repression of mTOR complex 2 induces changes in the gut microbiota of diet-induced obese mice. Sci Rep. (2016) 29:30887. doi: 10.1038/srep30887
30. Sung M, Kim T, Denou E, Soltys C, Hamza S, Byrne N, et al. Improved glucose homeostasis in obese mice treated with resveratrol is associated with alterations in the gut microbiome. Diabetes. (2017) 66:418–25. doi: 10.2337/db16-0680
31. Wang P, Gao J, Ke W, Wang J, Li D, Liu R, et al. Resveratrol reduces obesity in high-fat diet-fed mice via modulating the composition and metabolic function of the gut microbiota. Free Radic Biol Med. (2020) 156:83–98. doi: 10.1016/j.freeradbiomed.2020.04.013
32. Chen K, Zhao H, Shu L, Xing H, Wang C, Lu C, et al. Effect of resveratrol on intestinal tight junction proteins and the gut microbiome in high-fat diet-fed insulin resistant mice. Int J Food Sci Nutr. (2020) 71:965–78. doi: 10.1080/09637486.2020.1754351
33. Du F, Huang R, Lin D, Wang Y, Yang X, Huang X, et al. Resveratrol improves liver Steatosis and Insulin resistance in non-alcoholic fatty liver disease in association with the gut microbiota. Front Microbiol. (2021) 12:611323. doi: 10.3389/fmicb.2021.611323
34. Etxeberria U, Arias N, Boqué N, Macarulla M, Portillo M, Martínez J, et al. Reshaping faecal gut microbiota composition by the intake of trans-resveratrol and quercetin in high-fat sucrose diet-fed rats. J Nutr Biochem. (2015) 26:651–60. doi: 10.1016/j.jnutbio.2015.01.002
35. Yang C, Deng Q, Xu J, Wang X, Hu C, Tang H, et al. Sinapic acid and resveratrol alleviate oxidative stress with modulation of gut microbiota in high-fat diet-fed rats. Food Res Int. (2019) 116:1202–11. doi: 10.1016/j.foodres.2018.10.003
36. Walker J, Eckardt P, Aleman J, da Rosa J, Liang Y, Iizumi T, et al. The effects of trans-resveratrol on insulin resistance, inflammation, and microbiota in men with the metabolic syndrome: a pilot randomized, placebo-controlled clinical trial. J Clin Transl Res. (2018) 4:122–35
37. Kim T, Parajuli N, Sung N, Bairwa S, Levasseur J, Soltys C, et al. Fecal transplant from resveratrol-fed donors improves glycaemia and cardiovascular features of the metabolic syndrome in mice. Am J Physiol Endocrinol Metab. (2018) 315:E511–19. doi: 10.1152/ajpendo.00471.2017
38. Pannu N, Bhatnagar A. Resveratrol: from enhanced biosynthesis and bioavailability to multitargeting chronic diseases. Biomed Pharmacother. (2019) 109:2237–51. doi: 10.1016/j.biopha.2018.11.075
39. Macarulla M, Alberdi G, Gómez S, Tueros I, Bald C, Rodríguez V, et al. Effects of different doses of resveratrol on body fat serum parameters in rats fed a hypocaloric diet. J Physiol Biochem. (2015) 65:369–76. doi: 10.1007/BF03185932
40. Wu Y, Bible P, Long S, Ming W, Ding W, Long Y, et al. Metagenomic analysis reveals gestational diabetes mellitus-related microbial regulators of glucose tolerance. Acta Diabetol. (2020) 57:569–81. doi: 10.1007/s00592-019-01434-2
41. Saulnier D, Riehle K, Mistretta T, Mandal D, Raza S, Weidler E, et al. Gastrointestinal microbiome signatures of pediatric patients with irritable bowel syndrome. Gastroenterlogy. (2011) 141:1782–91. doi: 10.1053/j.gastro.2011.06.072
42. Parker B, Wearsch P, Veloo A, Rodriguez-Palacios A. The genus Alistipes: gut bacteria with emerging implications to inflammation, cancer and mental health. Front Immunol. (2020) 11:906. doi: 10.3389/fimmu.2020.00906
43. Zhu L, Sha L, Li K, Wang Z, Wang T, Li Y, et al. Dietary flaxseed oil rich in omega-3 suppresses severity of type 2 diabetes mellitus via anti-inflammation and modulating gut microbiota in rats. Lipids Health Dis. (2020) 19:20–36. doi: 10.1186/s12944-019-1167-4
44. Zhang X, Zhao Y, Xu J, Xue Z, Zhang M, Pang X, et al. Modulation of gut microbiota by berberine and metformin during the treatment of high-fat diet-induced obesity in rats. Sci Rep. (2015) 5:14405–15. doi: 10.1038/srep14405
45. Braccia D, Jiang X, Pop M, Hall A. The capacity to produce hydrogen sulfide (H2S) via cysteine degradation is ubiquitous in the human gut microbiome. Front Microbiol. (2020) 12:705583. doi: 10.3389/fmicb.2021.705583
46. Liu Q, Cai B, Zhu L, Xin X, Wang X, Ming Z, et al. Liraglutide modulates gut microbiome and attenuates nonalcoholic fatty liver in db/db mice. Life Sci. (2020) 261:118457. doi: 10.1016/j.lfs.2020.118457
47. Tong X, Xu J, Lian F, Yu X, Zhao Y, Xu L, et al. Structural alteration of gut microbiota during the amelioration of human type 2 diabetes with hyperlipidemia by metformin and a traditional Chinese herbal formula: a multicenter, randomized, open label clinical trial. mBio. (2018) 9:e02392–17. doi: 10.1128/mBio.02392-17
Keywords: resveratrol, type 2 diabetes, insulin resistance, anti-diabetic effect, gut microbiota
Citation: Fernandez-Quintela A, Macarulla MT, Gómez-Zorita S, González M, Milton-Laskibar I and Portillo MP (2023) Relationship between changes in microbiota induced by resveratrol and its anti-diabetic effect on type 2 diabetes. Front. Nutr. 9:1084702. doi: 10.3389/fnut.2022.1084702
Received: 30 October 2022; Accepted: 12 December 2022;
Published: 06 January 2023.
Edited by:
Sakineh Shab-Bidar, Tehran University of Medical Sciences, IranReviewed by:
Pamela Senesi, University of Milan, ItalyMonica Gulati, Lovely Professional University, India
Copyright © 2023 Fernandez-Quintela, Macarulla, Gómez-Zorita, González, Milton-Laskibar and Portillo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Saioa Gómez-Zorita,  c2Fpb2EuZ29tZXpAZWh1LmV1cw==; Iñaki Milton-Laskibar, aW5ha2kubWlsdG9uQGVodS5ldXM=
c2Fpb2EuZ29tZXpAZWh1LmV1cw==; Iñaki Milton-Laskibar, aW5ha2kubWlsdG9uQGVodS5ldXM=
†These authors share first authorship