 Ali Afzal
Ali Afzal Tarique Hussain
Tarique Hussain Amjad Hameed
Amjad Hameed
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Nutr. , 23 July 2021
Sec. Nutrition and Metabolism
Volume 8 - 2021 | https://doi.org/10.3389/fnut.2021.700957
This study investigated the effects of supplementing different levels of Moringa oleifera leaf powder (MOLP) on antioxidant status and blood biochemical indices during early gestation in Beetal goats. A total of 30 goats were randomly divided into three groups (n = 10) comprising control (basal diet without MOLP), the 1.6% MOLP supplemented group (basal diet + 1.6% MOLP), and the 3.2% MOLP supplemented group (basal diet + 3.2% MOLP). The experiment started 5 days before estrus synchronization and lasted till day 60 of gestation. The MOLP significantly increased plasma flavonoids in 1.6% as well as 3.2% supplemented group on days 40 and 60 of pregnancy, while total phenolic contents were observed to be higher in the 3.2% MOLP supplemented group throughout the experiment in comparison with the control group. The supplementation improved plasma total antioxidant capacity (TAC) by decreasing malondialdehyde (MDA) and total oxidant status (TOS) in contrast to the control group. The activities of superoxide dismutase (SOD) and peroxidase (POD) were enhanced in both supplemented groups, whereas catalase (CAT) activity was noticed to be significantly high in the 3.2% MOLP supplemented group. The protein contents were significantly elevated with 1.6 and 3.2% supplementation levels from day 40 to day 60 of the experiment. Plasma sugar level, carotenoids, progesterone profile, and hydrolytic (protease and amylase) enzymes activities were improved only when supplemented with 3.2% MOLP. The findings suggest that supplementing with 3.2% MOLP provides beneficial effects on early pregnancy stress in Beetal goats.
Pregnancy is a physiological process characterized by an intense metabolic burden that interrupts antioxidant stability and energy balance. It is common for about 25% of the embryos die or be reabsorbed within 2 weeks of pregnancy before implantation (1). The removal of zona pellucida of the embryo is followed by an increase in reactive oxygen species (ROS) production (2). The overloading of ROS promotes the activity of nicotinamide adenine dinucleotide phosphate hydrogen (NADPH) oxidase and suppresses antioxidant enzymes such as glutathione peroxidase (POD), catalase (CAT), and superoxide dismutase (SOD) (3). The disturbance in oxidant and antioxidant balance during early pregnancy may cause severe damage to lipid membranes, proteins, and DNAs, which arrests the further development and growth of the embryo (4).
The early pregnancy nutritional requirements are more than normal maintenance because goats need more energy to combat oxidative stress and maintain a healthy pregnancy (5). However, maternal malnutrition is very common around the world in small ruminants due to the high cost of feed, particularly in developing countries. Therefore, the small ruminants are mainly kept on conventional grazing pastures, low-quality hay, and crop by-products. These feedstuffs lack an adequate amount of protein, vitamins, and minerals to fulfill the nutritional requirements of pregnant goats (6). Maternal malnutrition has a negative impact on the conception rate and also adversely affects the fetus growth (7). Supplementation of the diet with appropriate phytobiotics (plant-derived feed additives) having suitable nutritional values has been proposed as one of the potential approaches for improving the antioxidant and nutritional status of goats during pregnancy (8–10). Phytobiotics from herbal plants are a balanced source of different nutrients that have been explored to have high antioxidant and immunomodulatory properties and may serve as a potential feed supplement for ruminants (11, 12).
Moringa oleifera (MO) is an evergreen multipurpose tree of high economic importance and nutritional values (13). Leaves of MO, the most nutritious and utilized part of the plant, are a rich source of proteins, amino acids, minerals, and vitamins (14). Generally, the dried leaves contain 19 different amino acids and an abundance of bioactive compounds, especially antioxidant substances such as flavonoids (myricetin, quercetin, and kaempferol); phenolic acids (gallic, chlorogenic, and ellagic acid); proanthocyanidins; vitamin E; vitamin C; selenium, zinc; and β carotene. These compounds are present separately in different plants but MO is unique in having them all in appreciable amounts, and they have also been reported to possess strong antioxidant potential than synthetic antioxidants such as rutin and butylated hydroxytoluene (15). The utilization of MO as an animal feed supplement in tropical and subtropical zones has several advantages over other plants, such as its potential to tolerate both drought and mild frost conditions, to be a multicut fodder with short cutting interval, to be easily adapted and digested by the animals, and to have high mineral (Ca and Fe) and protein contents, essential fatty acids, and negligible amounts of antinutritional factors (16).
The MO has been reported to improve reproductive performance and serum antioxidant status during pregnancy in mice (17). Previous studies provided information about the impact of MO on lactation performance and milk quality in dairy cows and goats (9, 18). However, there is little information about dietary supplementation impacts of MO on early pregnancy stress in goats. Therefore, the study was designed to investigate the effects of Moringa oleifera leaf powder (MOLP) supplementation on plasma antioxidant status and different biochemical indices during early pregnancy in Beetal goats.
The experimental plan and ethical clearance were approved and granted by the Institutional Research Committee for Animal Use and Care at Nuclear Institute for Agriculture and Biology (NIAB), Faisalabad, Pakistan.
The experiment was carried out at the goat farm of the Nuclear Institute for Agriculture and Biology (NIAB) located at about 7 km from the center of Faisalabad city, Pakistan (latitude 31.4287°N and longitude 73.0791°E) with an altitude of 184 m above sea level. The average temperature and rainfall were 24°C and 16.66 mm, respectively, during the months of January to March 2020 of the experiment.
A total of 30 multiparous, clinically healthy, non-pregnant Beetal goats about 2–3 of years of age, weighing about 40 ± 2.8 kg, and with a body condition score of 3.12 ± 1.20 were selected. The animals were randomly (n = 10) divided into control group (250 g basal diet without MOLP per animal per day), 1.6% MOLP supplemented group (250 g basal diet with 4 g MOLP per animal per day), and 3.2% MOLP supplemented group (250 g basal diet with 8 g MOLP per animal per day). The basal diet consisted of wheat, corn, rice bran, soybean meal, sugarcane molasses, and minerals formulated to fulfill the nutritional requirements of the goats (19) as shown in Table 1. The feeding levels were adjusted according to the previous studies conducted on mice with some modification in the goats (17, 20). The does were offered free pasture (Chloris gayana, Carduus nutans, Leptochloa fusca, Cirsium arvense, and Chenopodium album) grazing two times during the morning and evening schedule and were provided free access to clean drinking water. The animals were kept in well-ventilated semiopen sheds during the experiment. The experiment started 5 days prior to estrus synchronization and lasted till day 60 of pregnancy. All the animals were provided an adaptation period of 1 week for experimental diets. Before the commencement of the experiment, deworming of the does was done with Albendazole (Zoben, Prix, Lahore, Pakistan) at a dosage of 2.5 mg/5 kg of the body weight. Blood samples (5 ml) were collected from the jugular vein in sterile EDTA tubes (Vacutainer, Xinle) with a 20-day interval except for hormone (progesterone) analysis, which was collected after a 15-day interval from the day of conception (day 0). Plasma separation was done by centrifugation at 3,000 rpm/4°C and stored at −20°C for biochemical analysis. The graphical abstract and schematic diagram of experiment protocols are shown in Figures 1, 2.
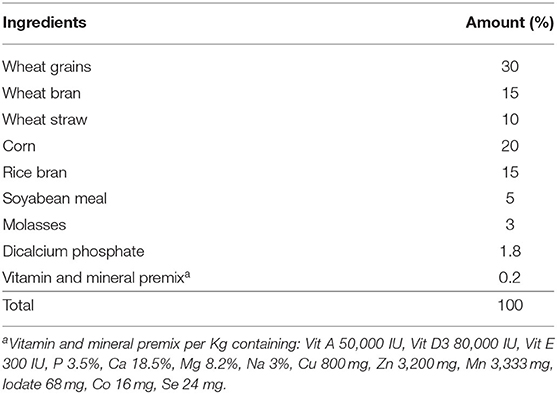
Table 1. Basal diet formulation (% DM).

Figure 1. Graphical abstract.
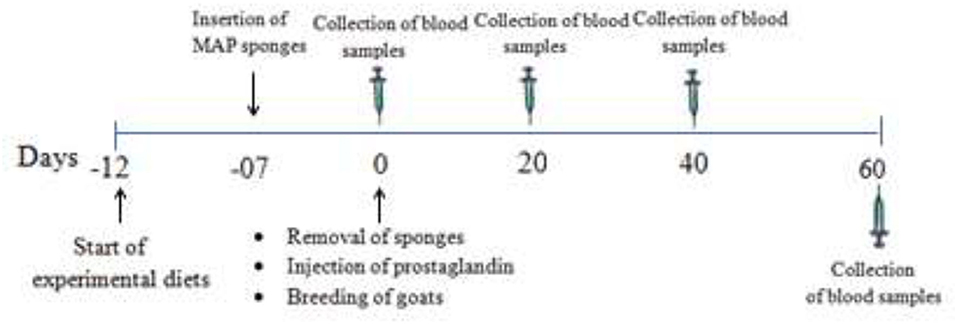
Figure 2. Schematic diagram of experiment protocols.
Estrus synchronization of the does was done using medroxyprogesterone acetate polyurethane (MAP) sponges. The sponges were made using the previous protocol developed for ewes with some modifications for goats (21, 22). Briefly, polyurethane sponges (length 3 cm and diameter 2 cm) were impregnated with 60 mg medroxyprogesterone acetate (Depo Provera; The Pfizer, Belgium), dissolved in 3 ml ethanol, and then left overnight in the fume hood for drying. The sponges were dusted with an antibiotic powder (streptomycin/penicillin) just before their insertion into the cranial part of the vagina with the help of a speculum and a sterilized applicator lubricated with paraffin oil. The sponges were placed for 7 days, and on the day of removal, all the does were injected with prostaglandin (Cloprostinol; Synchromate® @ 125 μg, Bremer, Germany). After showing behavioral estrus signs, a natural breeding service was provided using fertile bucks. Pregnancy determination was based on progesterone profile and non-return to estrus.
The fresh, green MO (PKM1) leaves were obtained from a breeder farm (Lahore, Pakistan), and their authenticity was confirmed by a concerned botanist at NIAB. A representative sample was submitted to the herbarium, Department of Botany, University of Agriculture, Faisalabad, Pakistan for future reference. The leaves were washed properly and dried under shade till constant weight. The dried leaves were ground to make a fine powder and stored in air-tight containers for future use.
The basal diet and MOLP were analyzed for dry matter, crude protein, crude fat, ash, nitrogen-free extract, neutral detergent fiber, and acid detergent fiber by using the standardized protocols of analytical chemists (23). The chemical composition of the basal diet is shown in Table 2.
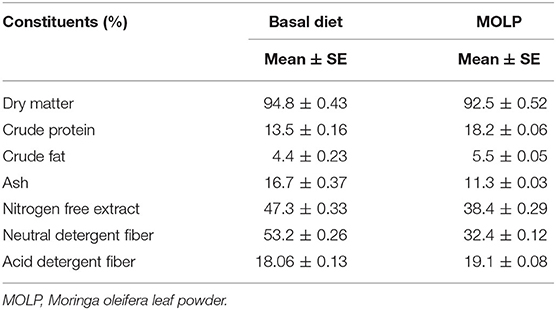
Table 2. Chemical composition of basal diet and MOLP (Dry matter basis).
The MOLP was analyzed for its various biochemical components as shown in Table 3.
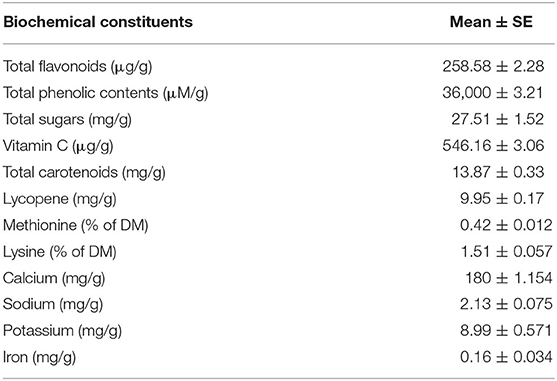
Table 3. Biochemical/nutritional constituents of Moringa oleifera leaf powder.
The amount of total flavonoids (TF) was determined as described by Lin and Tang (24). The MOLP methanol extract (2 mL) was homogenized with 0.1 mL of AlCl3 (10%), 2.8 mL of deionized water, and 0.1 mL of potassium acetate (1 M). After an incubation of 40 min at 37°C, the absorbance of the assay mixture was measured at 415 nm by a spectrophotometer (UV-VIS U2800, Hitachi, Japan). Rutin was used as the standard, and the TF content was measured as microgram RE gram−1 of the sample.
The total phenolic contents (TPC) were estimated by a micro colorimetric assay as described by Ainsworth and Gillespie (25) with some modifications. The MOLP (0.5 g) was dissolved in 500 μL ice-cold methanol (95%) and incubated at 37°C for 48 h in the dark. Then, its centrifugation was done for 5 min at 14,462 × g. The supernatants were separated and used for the estimation of TPC. For this purpose, 100 μL of supernatant was mixed with 100 μL of F-C reagent 10% (v/v), was vortexed thoroughly, and then was incubated at 37°C for 1 h after adding 800 μL of sodium carbonate (Na2CO3) (700 mM). The absorbance was read at 765 nm, and the amount of TPC was estimated by using a standard curve.
The leaf powder was mixed with concentrated sulfuric acid and then neutralized by using Na2CO3 for the determination of total sugars. The contents were filtered and used to measure the optical density (OD) at 415 nm (26).
Vitamin C in the leaf powder was estimated by using the 2,6-dichlorophenolindophenol (DCPIP) method as described by Hussein et al. (27). Briefly, each molecule of vitamin C causes reduction of DCPIP into DCPIPH2 molecule, and this reduction was measured as a decreasing trend in absorbance at 520 nm. A standard curve of ascorbic acid (ASA) was used to measure the vitamin C concentration in the samples.
The amounts of total carotenoids and lycopene were measured by using the protocol of Nagata and Yamashita (28). The leaf powder sample (1 g) was homogenized with 10 mL of hexane–acetone mixture (6:4) and incubated at 37°C for 5 min. The contents were then filtered and their absorbance was measured at 453, 505, and 663 nm. The amounts of carotenoids and lycopene were computed by using the following equations:
Amino acid analysis was done by ion-exchange chromatography with the help of an amino acid analyzer (Biochrom 30+; Biochrome Ltd, Cambridge, England), and the concentration of different minerals was determined by inductively coupled plasma optical emission spectroscopy (ICP-OES; Optima 2100-DV, Perkin Elmer, Massachusetts, USA).
Total flavonoids were determined by adopting an AlCl3 colorimetric approach (24). The plasma samples were mixed with 100 μL of potassium acetate (1 M), 100 μL of AlCl3 (10%), and 275 μL of water. After an incubation period of 40 min at room temperature, the absorbance of the assay solution was measured at 415 nm spectrophotometrically (UV-VIS, U2800, Hitachi, Japan). The TFs were manifested as microgram rutin equivalent (RE) mL−1 of the sample.
Total phenolic contents were determined by using the Folin–Ciocalteu reagent method (25). Each plasma sample (100 μL) was mixed with a 10% Folin–Ciocalteu reagent (100 μL), and homogenized thoroughly by a vortex. Then samples were incubated at room temperature for 1 h after adding 800 μL Na2CO3 (700 mM), and the absorbance was measured at 765 nm. The amount of TPC was calculated as Gallic acid equivalents (GAE) by using a standard Gallic acid linear regression curve.
Vitamin C contents were measured as ascorbic acid by following the standard protocol of Hussein et al. (27) with some necessary modifications. Ascorbic acid causes the reduction of a blue color compound DCPIP into a colorless compound DCPIPH2, and this transformation can be measured by the fall-off absorbance at 520 nm. Vitamin C contents were measured by using a standard calibration curve of ascorbic acid.
Malondialdehyde (MDA) in blood plasma samples was assayed in terms of lipid peroxidation, which is measured through the thiobarbituric acid (TBA) reaction according to the method of Dhindsa et al. (29). For estimation of MDA, plasma samples were homogenized with TCA (0.1%) and centrifuged for 5 min at 14,462 × g. Then, 1 mL of the supernatant was mixed with TCA (20%) having 0.05% TBA. The reaction solution was heated for 30 min at 95°C and cooled quickly in an ice bath. The absorbance of the supernatant was read at 532 nm after centrifugation for 10 min at 14,462 x g, and the value of non-specific absorption at 600 nm was subtracted from it. The MDA value was calculated by using a coefficient of extinction 155 mM−1cm−1.
Total antioxidant capacity (TAC) was calculated by the using 2,2-azino-bis (3-ethylbenzothiazoline-6-sulfonate) (ABTS) method as described by Dikilitas et al. (30). The assay is based on the reduction of the ABTS radical cation (ABTS·+) that is blue-green in color by antioxidants to its original colorless ABTS form. The assay solution composed of reagent R1 (mixture of glacial acetic acid and sodium acetate buffer solution), reagent R2 (mixture of glacial acetic acid, hydrogen peroxide, sodium phosphate buffer and ABTS), and the sample. The absorbance was read at 660 nm after mixing the contents of the tube and allowing them to stand for 6 min. The TAC was finally calculated from the standard curve of ascorbic acid and expressed as μM ascorbic acid equivalent mL−1.
The protocol for the estimation of total oxidant status (TOS) is based on the oxidation of iron with valence 2+ to an iron complex with valence 3+ in an acidic medium (30). Ferric ions react with xylenol orange and form a specific-colored complex. The color intensity is related to oxidant molecule quantity and can be measured spectrophotometrically. The assay mixture for determination of TOS comprised of reagent R1; xylenol orange solution (0.38 g in 500 μL of 25 Mm H2SO4), reagent R2; (o-dianisidine 0.0317 g, ferrous ammonium sulfate (II) 0.0196 g, NaCl 0.4 g, glycerol 500 μL); and the sample. The absorbance value was read at 520 nm after 5 min. The calibration curve of hydrogen peroxide (H2O2) was used to calculate TOS and is expressed in μmol H2O2 equivalent mL−1.
Superoxide dismutase enzyme activity of blood plasma was measured by an inhibition assay with some modifications (31). The assay is based on the ability of SOD to inhibit the photoreduction of nitroblue tetrazolium (NBT) into formazan. Assay solution to study the inhibition of NBT consisted of 50 mM phosphate buffer (pH 7.8), 57 μM NBT, 13 mM L-methionine, riboflavin (0.004%), triton 100X (0.025%), and plasma sample (50 μL) in a total volume of 3 mL. The photoreaction was carried out in a box lined with aluminum (Al) foil and fitted with a 15-W lamp as a source of luminescence. The absorbance during photo-reduction of NBT to formazan was measured at 560 nm spectrophotometrically. One unit of SOD activity was defined as the amount of enzyme required to cause 50% inhibition of photo-chemical reduction of NBT.
Peroxidase enzyme activity in blood plasma was assayed as described by Agostini et al. (32) using guaiacol as the substrate. The reaction solution for the analysis of POD activity comprised phosphate buffer (pH 7.0, 200 mM), guaiacol (200 mM), 545 μL distilled water, H2O2 (400 mM), and 15 μL sample. The reaction was started immediately after adding the sample, and the change in absorbance of the assay mixture was read at 470 nm for 1 min after every 20 s. One unit of enzyme (POD) activity was defined as the amount of enzyme required to catalyze the oxidation of guaiacol (1 μM) at 25°C and pH 7.0 in 1 min.
The blood plasma sample for estimation of CAT activity was homogenized in a medium containing dithiothreitol (1 mM) and potassium phosphate buffer (pH 7.0, 50 mM). The CAT activity was assayed according to the method of Beers and Sizer (33). The assay mixture for measurement of enzyme activity was composed of H2O2 (59 mM), potassium phosphate buffer (pH 7.0, 50 mM), and 100 μl plasma sample. The change in absorbance of the assay solution was measured at 240 nm for 1 min after every 20 s, and a decrease in absorbance in 0.01 min was defined as one unit of CAT activity.
Protein estimation in plasma samples was done by the dye-binding protocol of Bradford (34). For estimation of protein, 5 μL of the sample was homogenized with NaCl (0.1 N) and then 1.0 mL Bradford dye was added and mixed thoroughly. The reaction mixture was incubated at room temperature for 5 min to form a dye–protein complex and then its absorbance was read at 420 nm.
Sugar contents of blood plasma samples were executed by the method of Folin (26) with some modifications. Briefly, the plasma sample was mixed with sulfuric acid and then neutralized with Na2CO3. Thereafter, the contents were filtered and their absorbance was measured at 415 nm.
Plasma carotenoids and lycopene were calculated by using the method of Nagata and Yamashita (28). For estimation of carotenoids and lycopene, 1 mL of plasma sample was mixed with 10 mL of hexane acetone mixture (6:4) and incubated at 37°C for 5 min. The reaction mixture was then filtered and its absorbance was read at 453 nm, 505 nm, and 663 nm. The amounts of carotenoids and lycopene were computed by using the following formulae:
The progesterone concentration of plasma samples was measured by radioimmunoassay (Video gamma/Rack, I'acn, Italy) using commercially available kits (DIAsource, S.A, Belgium). The sensitivity of immunoassay was 0.05 ng mL−1, and the intra-assay and inter-assay variation coefficients were 6.5 and 8.2%, respectively. The cross-reactivity with other steroid hormones was <0.01–15%.
The protease enzyme activity was estimated by using a casein substrate (35). The assay is based on the digestion of casein by protease enzyme and the release of an amino acid “tyrosine” that reacts with Folin's reagent to produce a blue-colored product which is measured and quantified at 660 nm. A calibration curve of tyrosine was drawn by using its standard solutions to measure the protease activity from blood plasma samples.
The esterase (α & β) activity was determined by using α-naphthyl acetate and β-naphthyl acetate as substrates (36). The reaction solution comprised of enzyme extract (plasma) and substrate (1% acetone, 30 mM naphthyl acetate (α & β), and 0.04 M potassium phosphate buffer) was incubated at room temperature for 15 min in the dark. Then, 1 mL of staining solution [sodium dodecyl sulfate (5%) and fast blue BB (1%) in the ratio of 5:2] was added in both the reaction mixture and the blank control (substrate solution and phosphate buffer) and incubated again in the dark for 15 min at room temperature. The absorbance of the reaction mixture was read at 590 nm, and the enzyme activity was calculated in μM/min per mL of plasma by using a standard curve.
Plasma amylase activity was assessed by using tris-malate buffer (0.2 M, pH 7.2), which served as an assay cum extraction medium (37). For the measurement of amylase activity, 1 mL of plasma was mixed with 1 mL of substrate solution (0.15% starch) and incubated for 10 min at 37°C. Then, OD was read at 620 nm after adding quenching reagent. The activity of the enzyme was expressed in milligrams of starch degraded at min−1per mL of blood plasma.
The statistical analyses were performed with SPSS version 20. All the experimental procedures were performed in triplicates, and data were analyzed by one way-ANOVA with repeated measures to access the differences among various treatments on specific days. The results are presented in the tables as means ± SE, and the treatments with p < 0.05 were regarded statistically significant.
The impact of MOLP supplementation on plasma non-enzymatic antioxidant indices is shown in Table 4. The MOLP significantly increased plasma TF in the 1.6% MOLP supplemented group as well as in the 3.2% MOLP supplemented group in comparison with the control group on day 40 and day 60 of the experiment (p < 0.05). The highest value of TF was observed on day 60 in the 3.2% MOLP supplemented group. The TPC was significantly high throughout the experiment in the 3.2% MOLP supplemented group (p < 0.05), and the impact of 1.6% supplementation remained non-significant as compared with the control group (p > 0.05). Plasma vitamin C level increased significantly with 3.2% supplementation from the day of conception (day 0), whereas treatment with 1.6% MOLP exhibited a significant impact from day 20 to 60 of the gestation (p < 0.05). The MDA concentration declined significantly after MOLP supplementation from day 20 of the gestation in response to the control group (p < 0.05). Plasma TAC increased significantly during the experiment in supplemented groups as compared with the control group (p < 0.05) till it reached its peak on day 60 in the 3.2% MOLP supplemented group. The TOS value decreased significantly with the advancement in pregnancy in supplemented groups from day 20 till the end of the experiment on day 60 (p < 0.05).
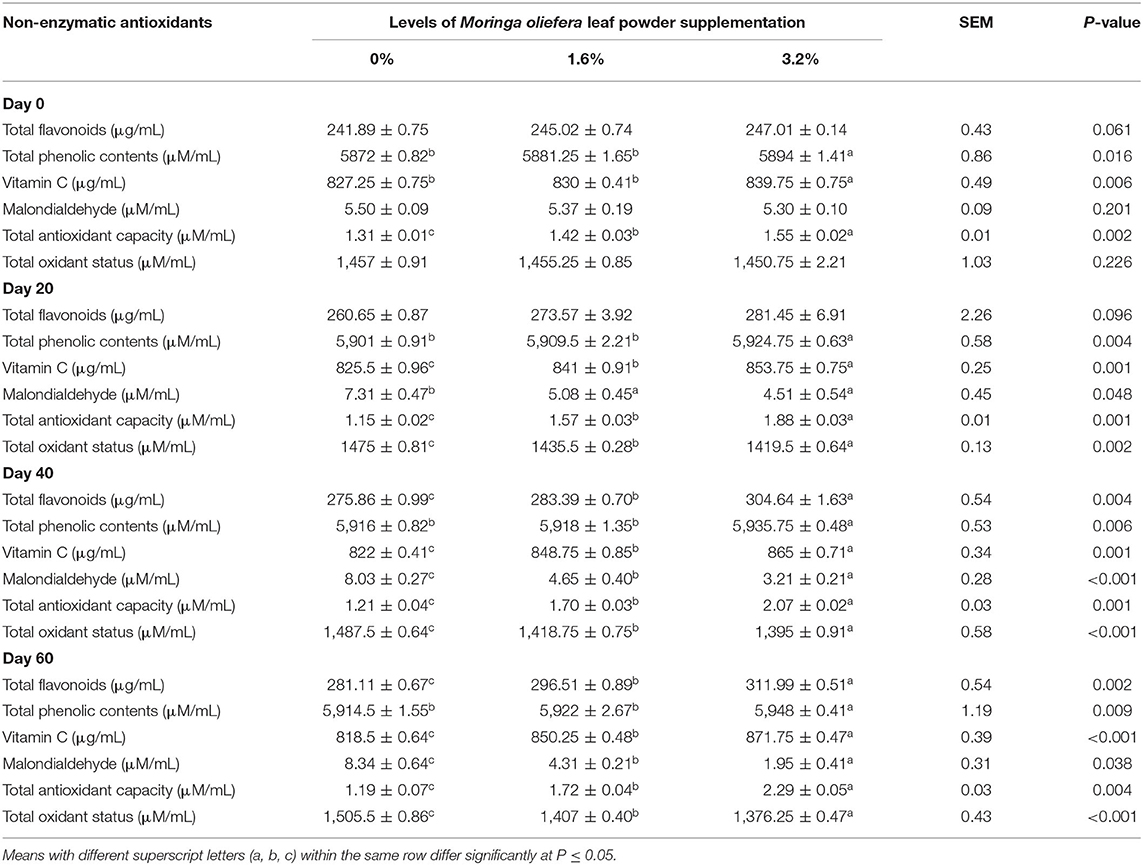
Table 4. Plasma non-enzymatic antioxidant indices of pregnant beetal goats.
Enzymatic antioxidant activities in the blood plasma of control and supplemented groups are shown in Table 5. Plasma SOD activity increased significantly in supplemented groups as compared with the control group (p < 0.05) throughout the experiment; however the increase was nonsignificant within supplemented groups on the day of conception (day 0) and on day 20 of gestation (p > 0.05). The increase in plasma POD activity remained nonsignificant between both (1.6 and 3.2%) treatments (p > 0.05) but remained significant in comparison with the control group during the entire experiment (p < 0.05). The MOLP supplementation did not exhibit any significant impact on CAT activity till day 20 (p > 0.05). After that, 3.2% treatment showed a significant effect on day 40 and day 60 in response to the control group (p < 0.05).
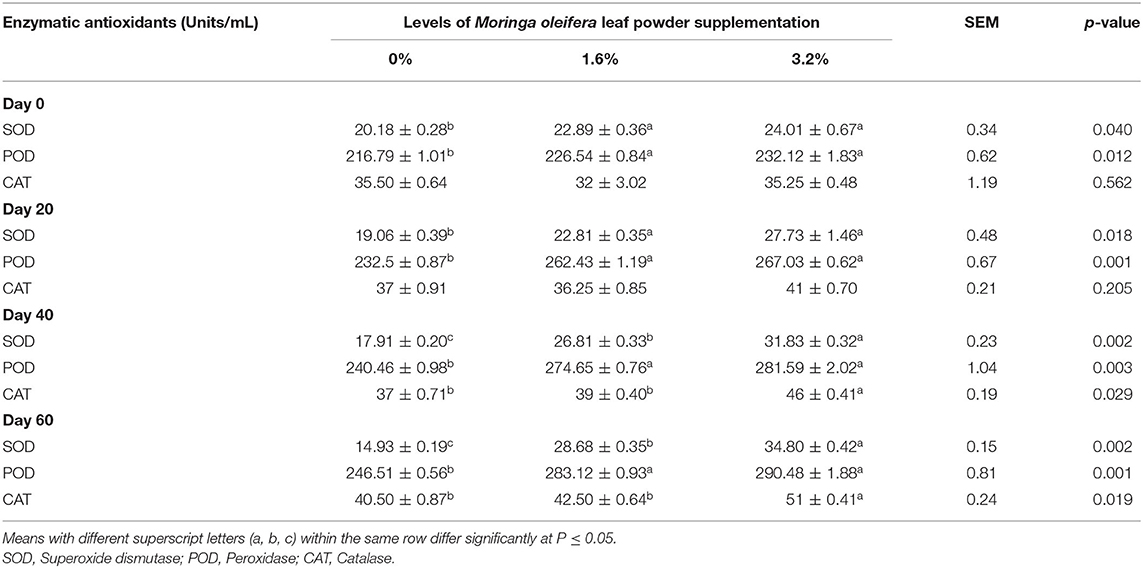
Table 5. Plasma enzymatic antioxidant indices of pregnant beetal goats.
The plasma biochemical profile of different parameters observed during the experiment is depicted in Table 6. The supplementation of MOLP significantly increased plasma protein contents from day 40 to day 60 of gestation in the 1.6% and 3.2% MOLP supplemented groups when compared with the control group (p < 0.05). A significant increase in plasma total sugars and carotenoids was observed throughout the experiment with 3.2% supplementation (p < 0.05), whereas 1.6% supplementation showed significant results on day 60 (p < 0.05). The amount of lycopene in plasma elevated significantly after feeding MOLP from day 20 in the 3.2% MOLP supplemented group and from day 40 in the 1.6% MOLP supplemented group (p < 0.05).

Table 6. Plasma biochemical indices of pregnant beetal goats.
The beneficial effect of MOLP supplementation on plasma progesterone concentration in early pregnant goats is displayed in Table 7. The exposure of the animals with 3.2% supplementation showed significant effect and more effectively enhanced plasma progesterone level throughout the experiment than that of the animals with the 1.6% MOLP supplementation and the control group (p < 0.05).
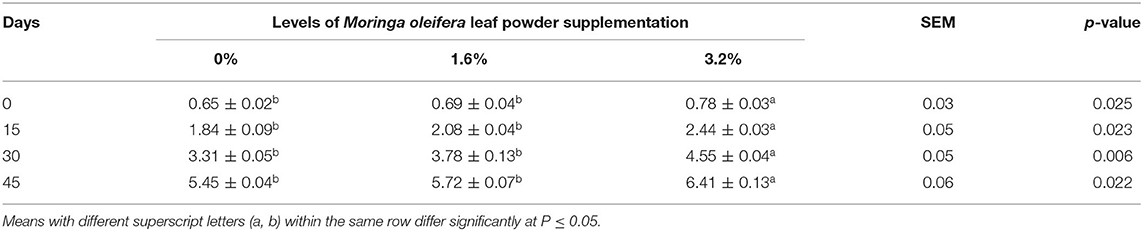
Table 7. Plasma progesterone (ng/mL) profile of pregnant beetal goats.
The activities of hydrolytic enzymes of both supplemented and control groups are illustrated in Table 8. The protease activity was significantly higher in the 3.2% MOLP supplemented group from day 20 till the completion of the experiment on day 60 of gestation as compared with the control group (p < 0.05). However, MOLP supplementation did not show any significant impact on esterase activity across the experiment in both the supplemented groups (p > 0.05). The amylase activity was not influenced by 1.6% supplementation when compared with the control group (p > 0.05), whereas 3.2% supplementation slightly increased the amylase activity and produced a significant impact on day 60 of the experiment (p < 0.05).
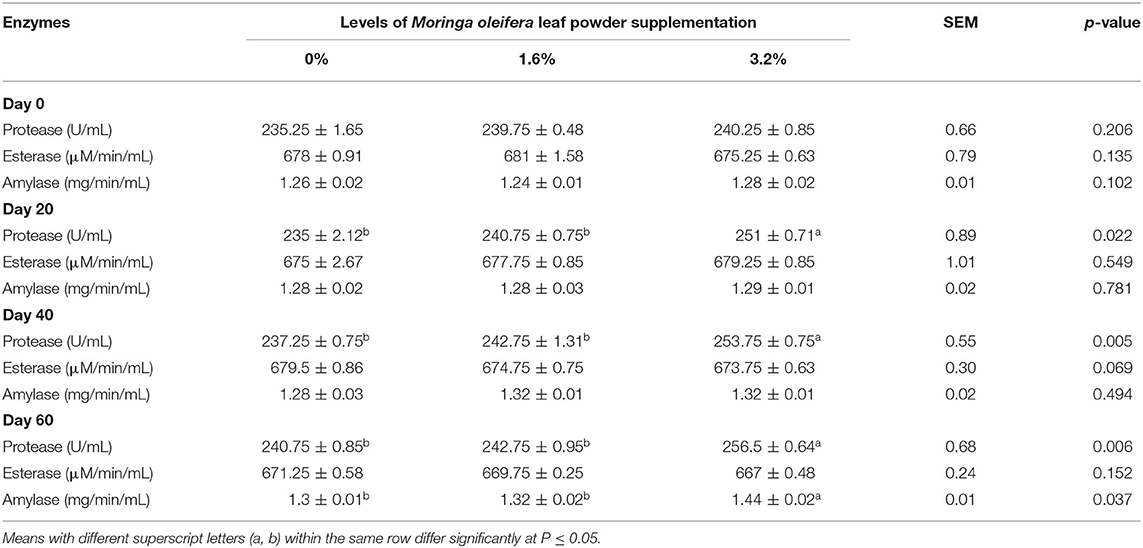
Table 8. Plasma hydrolytic enzyme activities of pregnant beetal goats.
Animals are more vulnerable to oxidative stress during pregnancy, especially in early gestational stages due to certain physiological changes in the maternal body. The quality of feedstuff plays an important role during this critical stage of pregnancy (38). The MO leaves possess several nutritional and medicinal properties that have been ascribed to the presence of different phytochemicals (39, 40). It is evident from the results of this study that supplementation of the diet with MOLP improved the plasma non-enzymatic antioxidants status during early gestation in goats. The elevated level of plasma flavonoids and phenolics enhanced the survival and growth rate of developing embryos by suppressing the immense production of nitric oxide (NO) and hydroxyl (OH) radicals as a result of early pregnancy stress (41). The increase in plasma flavonoids and phenolic contents in response to MOLP supplementation was in accordance with the findings of another study, in which MO leaf extract modified the antioxidant capacity in rats by improving the plasma flavonoids and phenolic contents under different oxidative stress conditions (42).
The plasma vitamin C level drops promptly with the advancement in pregnancy, which makes the mother as well as the developing fetus more prone to detrimental effects of oxidative stress (43). However, a significant improvement in plasma vitamin C contents in the supplemented groups demonstrated that MOLP, being a rich source of vitamin C, has the potential to provide protection from the harmful effects of oxidative stress during pregnancy. Our results were in agreement with the findings of Babiker et al. (44), who reported the positive impacts of MO supplementation on plasma and milk vitamin C contents in ewes.
Furthermore, MOLP supplementation also improved plasma TAC by limiting the production of MDA, a product of lipid peroxidation (LPO). MDA produced during pregnancy is more stable and highly reactive for biomolecules (proteins or DNA); thus, it leads to the development of certain pathological conditions that finally cause abortion (45, 46). The results of this study evidenced that MOLP supplementation also prevented the elevation of plasma MDA to promote the activities of enzymatic (SOD, POD, and CAT) antioxidants. Both the enzymatic and non-enzymatic antioxidant defense systems work side by side in attenuating oxidative stress to make conditions more favorable for the growth of the developing fetus (47). The SOD enzyme during the process of embryo implantation neutralizes ROS by stimulating the transformation of superoxide anion to hydrogen peroxide (H2O2), which is further metabolized into water and oxygen by POD and CAT (48). The successful implantation of an embryo on the endometrium of the uterus is dependent on SOD, POD, and CAT activities (49). The improvement in the enzymatic antioxidant defense system was also observed in rabbits, dairy cows, and sows following Moringa leaf supplementation in different proportions (18, 50, 51).
The excessive production of free radicals (ROS and RNS) during early gestation as a result of physiological changes increased the plasma TOS (52, 53). The high level of TOS is an indicator of oxidative stress. The results of this study depicted that MOLP supplementation decreased TOS values in the supplemented groups by restricting the production of ROS and RNS, as reported in some previous studies performed in ruminants (54, 55).
Blood biochemical analysis is a definite method for predicting the health status of animals (56). The MO supplementation has a significant impact on blood biochemistry (57). The increase in plasma protein level after MOLP supplementation indicated that MOLP was a lavish source of proteins, and its supplementation increased the plasma protein level by decreasing blood urea nitrogen (BUN) contents (50). The BUN is considered as an indicator of the balance between amino acid metabolism and protein synthesis (58). Pregnancy is considered to be a period of severe metabolic and protein alterations in the body (59). The decrease in plasma protein level with the advancement of gestation in the control group was due to the high demand for amino acids, which were transferred from the mother to the developing fetus for the synthesis of numerous proteins required for the proper growth of the fetus and for the maintenance of a healthy pregnancy (60). The high protein profile of MOLP makes it an ideal substitute for rapeseed and soybean meals for ruminants.
The significant increase in plasma sugar contents by dietary supplementation with MOLP specified its positive effects in improving the energy status of the body. During gestation, there is a rapid increase in metabolic rate to fulfill the high energy demands of pregnancy. However, if the provision of feed is not according to the requirements, the mobilization of body fat starts, which results in the production of non-esterified fatty acids (NEFAs). NEFAs are responsible for the development of ketosis and some other metabolic diseases that may adversely affect the pregnancy and future health of the offspring (61, 62). The provision of MOLP met the energy requirements more efficiently in early pregnancy by improving the plasma sugar contents. Our findings were in line with the results of a previous study on Jersey cows (63). In addition, MOLP supplementation also improved the bioavailability of carotenoids and lycopene to ameliorate metabolic and hematological perturbations by reducing LPO and inflammatory cytokines (64).
The progesterone hormone plays a key role in maintaining pregnancy (65). The ROS produced during early gestation prevents the implantation of the embryo by suppressing the production of progesterone from the corpus luteum (66). The high level of plasma progesterone concentration in treatment groups indicates a strong correlation with MOLP supplementation during early pregnancy. In this experiment, a 100% pregnancy rate was observed in the 3.2% MOLP supplemented group, while 80 and 70% pregnancy rates were noticed in the 1.6% M OLP supplemented and control groups, respectively. The results of our experiment are in accordance with the findings in rabbits supplemented with Moringa leaf. An increase in conception rate along with plasma progesterone profile was observed (67).
The protease enzymes from MO leaves help in maintaining body hemostasis during gestational stress by acting as an important component of the complement system. These enzymes also help in hydrolyzing and removing apoptotic products of oxidative stress from the body (68, 69). The high protease activity in supplemented groups indicate the beneficial impact of MOLP in maintaining body conditions by modifying and stabilizing plasma protease activity during early gestation in pregnant goats. Amylase enzyme maintains blood glucose level according to the energy requirements of the body by converting carbohydrates into glucose. The results of an in-vitro study indicated that MO leaf extract inhibited amylase activity at a concentration of 220 μg/mL of blood plasma, and below this level, it potentiated the activity of the enzyme to a certain extent (70). The MOLP used within the limit to increase the amylase activity denotes its supportive role in potentiating enzyme activity.
The supplementation of the basal diet with 3.2% MOLP improved the antioxidant defense system and blood biochemical indices during early gestation in Beetal goats. In addition, it also enhanced the progesterone profile by suppressing the production of reactive oxygen species. These findings suggest that MOLP is a promising agent to improve the reproductive performance of goats during early pregnancy. However, further studies are required on a large flock of animals with different concentrations of MOLP to dig out the molecular aspects of attenuating early pregnancy stress and improving fertility.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
AA: conduct experiment, data aggregation, statistical analysis, and wrote manuscript. AH: lab analysis. TH: design experiment, monitoring experiment, and revised manuscript. All authors contributed to the article and approved the submitted version.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The authors pay their special thanks to Mr. Muhammad Shahzad and Mr. Usman Mazhar for their help in analytical procedures to complete the research work.
1. Spencer TE, Johnson GA, Bazer FW, Burghardt RC. Implantation mechanisms: insights from the sheep. Reproduction. (2004) 128:657–68. doi: 10.1530/rep.1.00398
2. Al-Gubory KH, Faure P, Garrel C. Different enzymatic antioxidative pathways operate within the sheep caruncular and intercaruncular endometrium throughout the estrous cycle and early pregnancy. Theriogenology. (2017) 99:111–18. doi: 10.1016/j.theriogenology.2017.05.017
3. Duhig K, Chappell LC, Shennan AH. Oxidative stress in pregnancy and reproduction. Obstet Med. (2016) 9:113–16. doi: 10.1177/1753495X16648495
4. Eick SM, Barrett ES, Van ‘t Erve TJ, Nguyen RH, Bush NR, Milne G, et al. Association between prenatal psychological stress and oxidative stress during pregnancy. Paediatr Perinat Epidemiol. (2018) 32:318–26. doi: 10.1111/ppe.12465
5. Nawito MF, Abd El Hameed AR, Sosa AS, Mahmoud KG. Impact of pregnancy and nutrition on oxidant/antioxidant balance in sheep and goats reared in South Sinai, Egypt. Vet World. (2016) 9:801–5. doi: 10.14202/vetworld.2016.801-805
6. Härter CJ, Lima LD, Silva HGO, Castagnino DS, Rivera AR, Resende KT, et al. Energy and protein requirements for maintenance of dairy goats during pregnancy and their efficiencies of use. J Anim Sci. (2017) 95:4181–93. doi: 10.2527/jas.2017.1490
7. Sharma D, Shastri S, Sharma P. Intrauterine growth restriction: antenatal and postnatal aspects. Clin Med Insights Pediatr. (2016) 10:67–83. doi: 10.4137/CMPed.S40070
8. Kholif AE, Morsy TA, Gouda GA, Anele UY, Galyean ML. Effect of feeding diets with processed Moringa oleifera meal as protein source in lactating Anglo-Nubian goats. Anim Feed Sci Technol. (2016) 217:45–55. doi: 10.1016/j.anifeedsci.2016.04.012
9. Al-Juhaimi FY, Alsawmahi ON, Abdoun KA, Ghafoor K, Babiker EE. Antioxidant potential of moringa leaves for improvement of milk and serum quality of aardi goats. S Afr J Bot. (2020) 129:134–37. doi: 10.1016/j.sajb.2019.03.022
10. Abou-Elkhair R, Mahboub H, Sadek K, Ketkat S. Effect of prepartum dietary energy source on goat maternal metabolic profile, neonatal performance, and economic profitability. J Adv Vet Anim Res. (2020) 7:566–74. doi: 10.5455/javar.2020.g454
11. Bampidis VA, Christodoulou V, Christaki E, Florou-Paneri P, Spais AB. Effect of dietary garlic bulb and garlic husk supplementation on performance and carcass characteristics of growing lambs. Anim Feed Sci Technol. (2005) 121:273–83. doi: 10.1016/j.anifeedsci.2005.02.003
12. Patra AK, Amasheh S, Aschenbach JR. Modulation of gastrointestinal barrier and nutrient transport function in farm animals by natural plant bioactive compounds–a comprehensive review. Crit Rev Food Sci Nutr. (2019) 59:3237–66. doi: 10.1080/10408398.2018.1486284
13. Padayachee B, Baijnath H. An updated comprehensive review of the medicinal, phytochemical and pharmacological properties of Moringa oleifera. S Afr J Bot. (2020) 129:304–16. doi: 10.1016/j.sajb.2019.08.021
14. Abbas RK, Elsharbasy FS, Fadlelmula AA. Nutritional values of moringa oleifera, total protein. Amino acid, vitamins, minerals, carbohydrates, total fat and crude fiber, under the semi-arid conditions of Sudan. J Microb Biochem Technol. (2018) 10:56–8. doi: 10.4172/1948-5948.1000396
15. Saleem A, Saleem M, Akhtar MF. Antioxidant, anti-inflammatory and antiarthritic potential of Moringa oleifera lam: an ethnomedicinal plant of Moringaceae family. S Afr J Bot. (2020) 128:246–56. doi: 10.1016/j.sajb.2019.11.023
16. Olusanya RN, Kolanisi U, Van Onselen A, Ngobese NZ, Siwela M. Nutritional composition and consumer acceptability of Moringa oleifera leaf powder (MOLP)-supplemented mahewu. S Afr J Bot. (2020) 129:175–80. doi: 10.1016/j.sajb.2019.04.022
17. Zeng B, Luo J, Wang P, Yang L, Chen T, Sun J, et al. The beneficial effects of Moringa oleifera leaf on reproductive performance in mice. Food Sci Nutr. (2019) 7:738–46. doi: 10.1002/fsn3.918
18. Kekana TW, Marume U, Muya MC, Nherera-Chokuda FV. Periparturient antioxidant enzymes, haematological profile and milk production of dairy cows supplemented with Moringa oleifera leaf meal. Anim Feed Sci Technol. (2020) 268:114606. doi: 10.1016/j.anifeedsci.2020.114606
19. NRC. Nutrient Requirements of Small Ruminants: Sheep, Goats, Cervids, and New World Camelids. Washington, DC: National Academy Press (2007).
20. Agrawal SS, Vishal D, Sumeet G, Shekhar C, Ashish N, Parul D, et al. Antifertility activity of ethanol leaf extract of Moringa oleifera lam in female wistar rats. Indian J Pharm Sci. (2018) 80:565–70. doi: 10.4172/pharmaceutical-sciences.1000394
21. Kausar R, Khanum SA, Hussain M, Shah MS. Estrus synchronization with medroxyprogesterone acetate impregnated sponges in goats (Capra hircus). Pak Vet J. (2009) 29:16–8.
22. Simonetti L, Blanco MR, Gardón JC. Estrus synchronization in ewes treated with sponges impregnated with different doses of medroxyprogesterone acetate. Small Rumin Res. (2000) 38:243–47. doi: 10.1016/S0921-4488(00)00160-7
23. AOAC. Association of Official Analytical Chemist. Official Methods of Analysis, Vol. 222, 21 ed. Washington, DC: Association of Official Analytical Chemists (2005).
24. Lin JY, Tang CY. Determination of total phenolic and flavonoid contents in selected fruits and vegetables, as well as their stimulatory effects on mouse splenocyte proliferation. Food Chem. (2007) 101:140–47. doi: 10.1016/j.foodchem.2006.01.014
25. Ainsworth EA, Gillespie KM. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin–Ciocalteu reagent. Nat protoc. (2007) 2:875–77. doi: 10.1038/nprot.2007.102
26. Folin O. Two revised copper methods for blood sugar determination. J Biol Chem. (1929) 82:83–93. doi: 10.1016/S0021-9258(18)63690-7
27. Hussein SA, Barakat HH, Merfort I, Nawwar MA. Tannins from the leaves of Punica granatum. Phytochemistry. (1997) 45:819–23. doi: 10.1016/S0031-9422(96)00888-6
28. Nagata M, Yamashita I. Simple method for simultaneous determination of chlorophyll and carotenoids in tomato fruit. Nippon Shokuhin Kogyo Gakkaishi. (1992) 39:925–28. doi: 10.3136/nskkk1962.39.925
29. Dhindsa RS, Plumb-Dhindsa PA, Thorpe TA. Leaf senescence: correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J Exp Bot. (1981) 32:93–101. doi: 10.1093/jxb/32.1.93
30. Dikilitas M, Guldur ME, Deryaoglu A, Ozcan ER. Antioxidant and oxidant levels of pepper (Capsicum annuum cv. â€~ Charlee’) infected with pepper mild mottle virus. Not Bot Horti Agrobo. (2011) 39:58–63. doi: 10.15835/nbha3925881
31. Beyer Jr WF, Fridovich I. Assaying for superoxide dismutase activity: some large consequences of minor changes in conditions. Anal Biochem. (1987) 161:559–66. doi: 10.1016/0003-2697(87)90489-1
32. Agostini E, Medina MI, Milrad de Forchetti SR, Tigier H. Properties of two anionic peroxidase isoenzymes from turnip (Brassica napus L.) roots. J Agric Food Chem. (1997) 45:596–98. doi: 10.1021/jf960259u
33. Beers RF, Sizer IW. A spectrophotometric method for measuring the breakdown of hydrogen peroxide by catalase. J Biol Chem. (1952) 195:133–40. doi: 10.1016/S0021-9258(19)50881-X
34. Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. (1976) 72:248–54. doi: 10.1016/0003-2697(76)90527-3
35. Peterson GL. A simplification of the protein assay method of Lowry et al. which is more generally applicable. Anal Biochem. (1977) 83:346–56. doi: 10.1016/0003-2697(77)90043-4
36. Van Asperen K. A study of housefly esterases by means of a sensitive colorimetric method. J Insect Physiol. (1962) 8:401–16. doi: 10.1016/0022-1910(62)90074-4
37. Filner P, Varner JE. A test for de novo synthesis of enzymes: density labeling with H2O18 of barley alpha-amylase induced by gibberellic acid. Proc Natl Acad Sci USA. (1967) 58:1520–6. doi: 10.1073/pnas.58.4.1520
38. Li X, Li H, He Z, Tan Z, Yan Q. Effects of maternal intake restriction during early pregnancy on fetal growth and bone metabolism in goats. Small Rumin Res. (2020) 183:106027. doi: 10.1016/j.smallrumres.2019.106027
39. Verma AR, Vijayakumar M, Mathela CS, Rao CV. In vitro and in vivo antioxidant properties of different fractions of Moringa oleifera leaves. Food Chem Toxicol. (2009) 47:2196–201. doi: 10.1016/j.fct.2009.06.005
40. Vongsak B, Sithisarn P, Mangmool S, Thongpraditchote S, Wongkrajang Y, Gritsanapan W. Maximizing total phenolics, total flavonoids contents and antioxidant activity of Moringa oleifera leaf extract by the appropriate extraction method. Ind Crops Prod. (2013) 44:566–71. doi: 10.1016/j.indcrop.2012.09.021
41. Ismail HF, Hashim Z, Soon WT, Ab Rahman NS, Zainudin AN, Majid FA. Comparative study of herbal plants on the phenolic and flavonoid content, antioxidant activities and toxicity on cells and zebrafish embryo. J Tradit Complement Med. (2017) 7:452–65. doi: 10.1016/j.jtcme.2016.12.006
42. Aekthammarat D, Pannangpetch P, Tangsucharit P. Moringa oleifera leaf extract lowers high blood pressure by alleviating vascular dysfunction and decreasing oxidative stress in L-NAME hypertensive rats. Phytomedicine. (2019) 54:9–16. doi: 10.1016/j.phymed.2018.10.023
43. Rao GM, Sumita P, Roshni M, Ashtagimatt MN. Plasma antioxidant vitamins and lipid peroxidation products in pregnancy induced hypertension. Indian J Clin Biochem. (2005) 20:198–200. doi: 10.1007/BF02893070
44. Babiker EE, Juhaimi FA, Ghafoor K, Mohamed HE, Abdoun KA. Effect of partial replacement of alfalfa hay with Moringa species leaves on milk yield and composition of Najdi ewes. Trop Anim Health Prod. (2016) 48:1427–33. doi: 10.1007/s11250-016-1111-9
45. Zarkovic N, Cipak A, Jaganjac M, Borovic S, Zarkovic K. Pathophysiological relevance of aldehydic protein modifications. J Proteom. (2013) 92:239–47. doi: 10.1016/j.jprot.2013.02.004
46. Blair IA. DNA adducts with lipid peroxidation products. J Biol Chem. (2008) 283:15545. doi: 10.1074/jbc.R700051200
47. Kharrazi H, Vaisi-Raygani A, Rahimi Z, Tavilani H, Aminian M, Pourmotabbed T. Association between enzymatic and non-enzymatic antioxidant defense mechanism with apolipoprotein E genotypes in Alzheimer disease. Clin Biochem. (2008) 41:932–36. doi: 10.1016/j.clinbiochem.2008.05.001
48. Liu G, Dong Y, Wang Z, Cao J, Chen Y. Restraint stress alters immune parameters and induces oxidative stress in the mouse uterus during embryo implantation. Stress. (2014) 17:494–503. doi: 10.3109/10253890.2014.966263
49. Al-Gubory KH, Bolifraud P, Germain G, Nicole A, Ceballos-Bicot I. Antioxidant enzymatic defence systems in sheep corpus luteum throughout pregnancy. Reproduction. (2004) 128:767–74. doi: 10.1530/rep.1.00389
50. Sun JJ, Wang P, Chen GP, Luo JY, Xi QY, Cai GY, et al. Effect of Moringa oleifera supplementation on productive performance, colostrum composition and serum biochemical indexes of sow. J Anim Physiol Anim Nutr. (2020) 104:291–99. doi: 10.1111/jpn.13224
51. Sun B, Zhang Y, Ding M, Xi Q, Liu G, Li Y, et al. Effects of Moringa oleifera leaves as a substitute for alfalfa meal on nutrient digestibility, growth performance, carcass trait, meat quality, antioxidant capacity and biochemical parameters of rabbits. J Anim Physiol Anim Nutr. (2018) 102:194–203. doi: 10.1111/jpn.12678
52. O'Boyle N, Corl CM, Gandy JC, Sordillo LM. Relationship of body condition score and oxidant stress to tumor necrosis factor expression in dairy cattle. Vet Immunol Immunopathol. (2006) 113:297–304. doi: 10.1016/j.vetimm.2006.05.013
53. Matés JM, Sánchez-Jiménez F. Antioxidant enzymes and their implications in pathophysiologic processes. Front Biosci. (1999) 4:339–45. doi: 10.2741/A432
54. Pedernera M, Celi P, García SC, Salvin HE, Barchia I, Fulkerson WJ. Effect of diet, energy balance and milk production on oxidative stress in early-lactating dairy cows grazing pasture. Vet J. (2010) 186:352–57. doi: 10.1016/j.tvjl.2009.09.003
55. Celi P, Di Trana A, Claps S. Effects of plane of nutrition on oxidative stress in goats during the peripartum period. Vet J. (2010) 184:95–9. doi: 10.1016/j.tvjl.2009.01.014
56. Mohammed SA, Razzaque MA, Omar AE, Albert S, Al-Gallaf WM. Biochemical and hematological profile of different breeds of goat maintained under intensive production system. Afr J Biotechnol. (2016) 15:1253–57. doi: 10.5897/AJB2016.15362
57. Babeker EA, Abdalbagi YM. Effect of feeding different levels of Moringa oleifera leaves on performance, haematological, biochemical and some physiological parameters of Sudan Nubian goats. Online J Anim Feed Res. (2015) 5:50–61.
58. Janku L, Pavlata L, Mišurová L, Filípek J, Pechová A, Dvorák R. Levels of protein fractions in blood serum of periparturient goats. Acta Vet Brno. (2011) 80:185–90. doi: 10.2754/avb201180020185
59. Piccione G, Messina V, Marafioti S, Casella S, Giannetto C, Fazio F. Changes of some haematochemical parameters in dairy cows during late gestation, post-partum, lactation and dry periods. Vet Med Zoot. (2012)58:59–64.
60. Tóthová C, Nagy O, Nagyová V, Kováč G. Serum protein electrophoretic pattern in dairy cows during the periparturient period. J Appl Anim Res. (2018) 46:33–8. doi: 10.1080/09712119.2016.1256293
61. Caré S, Trevisi E, Minuti A, Ferrari A, Loor JJ, Calamari L. Plasma fructosamine during the transition period and its relationship with energy metabolism and inflammation biomarkers in dairy cows. Livest Sci. (2018) 216:138–47. doi: 10.1016/j.livsci.2018.08.003
62. Roche JR, Sheahan AJ, Chagas LM, Blache D, Berry DP, Kay JK. Long-term infusions of ghrelin and obestatin in early lactation dairy cows. J Dairy Sci. (2008) 91:4728–40. doi: 10.3168/jds.2008-1193
63. Kekana TW, Marume U, Muya CM, Nherera-Chokuda FV. Lactation performance and blood metabolites in lactating dairy cows micro-supplemented with Moringa oleifera leaf meal. S Afr J Anim Sci. (2019) 49:709–16. doi: 10.4314/sajas.v49i4.12
64. Kilany OE, Abdelrazek HM, Aldayel TS, Abdo S, Mahmoud MM. Anti-obesity potential of Moringa olifera seed extract and lycopene on high fat diet induced obesity in male Sprauge Dawely rats. Saudi J Biol Sci. (2020) 27:2733–40. doi: 10.1016/j.sjbs.2020.06.026
65. Spencer TE, Forde N, Lonergan P. The role of progesterone and conceptus-derived factors in uterine biology during early pregnancy in ruminants. J Dairy Sci. (2016) 99:5941–50. doi: 10.3168/jds.2015-10070
66. Sugino N. Roles of reactive oxygen species in the corpus luteum. Anim Sci J. (2006) 77:556–65. doi: 10.1111/j.1740-0929.2006.00386.x
67. Kleden MM, Soetanto H. Concentration of progesterone and prolactin hormones and milk production of New Zealand white rabbits doe fed moringa leaves meal. Mediterr J Soc Sci. (2017) 8:79–89. doi: 10.5901/mjss.2017.v8n3p79
68. Satish A, Sairam S, Ahmed F, Urooj A. Moringa oleifera Lam.: protease activity against blood coagulation cascade. Pharmacogn Res. (2012) 4:44–51. doi: 10.4103/0974-8490.91034
69. Hubbard RE, O'Mahony MS, Calver BL, Woodhouse KW. Plasma esterases and inflammation in ageing and frailty. Eur J Clin Pharmacol. (2008) 64:895–900. doi: 10.1007/s00228-008-0499-1
Keywords: Moringa oleifera, pregnancy stress, antioxidants status, beetal goats, biochemical indices of blood plasma
Citation: Afzal A, Hussain T and Hameed A (2021) Moringa oleifera Supplementation Improves Antioxidant Status and Biochemical Indices by Attenuating Early Pregnancy Stress in Beetal Goats. Front. Nutr. 8:700957. doi: 10.3389/fnut.2021.700957
Received: 27 April 2021; Accepted: 23 June 2021;
Published: 23 July 2021.
Edited by:
Fátima Martel, Universidade do Porto, PortugalReviewed by:
Hesham Rushed El-Seedi, Uppsala University, SwedenCopyright © 2021 Afzal, Hussain and Hameed. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tarique Hussain, ZHJ0YXJpcXVlcmFob29AZ21haWwuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.