 Jennifer N. Burns1,2
Jennifer N. Burns1,2 Aaron K. Jenkins1
Aaron K. Jenkins1 Xiangning Xue3
Xiangning Xue3 Kaitlyn A. Petersen1,2
Kaitlyn A. Petersen1,2 Kyle D. Ketchesin1,2Megan S. Perez1,4
Kyle D. Ketchesin1,2Megan S. Perez1,4 Chelsea A. Vadnie5
Chelsea A. Vadnie5 Madeline R. Scott1,2
Madeline R. Scott1,2 Marianne L. Seney1,2
Marianne L. Seney1,2 George C. Tseng3
George C. Tseng3 Colleen A. McClung1,2*
Colleen A. McClung1,2*- 1Translational Neuroscience Program, Department of Psychiatry, University of Pittsburgh School of Medicine, Pittsburgh, PA, United States
- 2Center for Neuroscience, University of Pittsburgh, Pittsburgh, PA, United States
- 3Department of Biostatistics, University of Pittsburgh, Pittsburgh, PA, United States
- 4Department of Human Genetics, School of Public Health, University of Pittsburgh, Pittsburgh, PA, United States
- 5David O. Robbins Neuroscience Program, Department of Psychology, Ohio Wesleyan University, Delaware, OH, United States
Introduction: Alterations in multiple subregions of the human prefrontal cortex (PFC) have been heavily implicated in psychiatric diseases. Moreover, emerging evidence suggests that circadian rhythms in gene expression are present across the brain, including in the PFC, and that these rhythms are altered in disease. However, investigation into the potential circadian mechanisms underlying these diseases in animal models must contend with the fact that the human PFC is highly evolved and specialized relative to that of rodents.
Methods: Here, we use RNA sequencing to lay the groundwork for translational studies of molecular rhythms through a sex-specific, cross species comparison of transcriptomic rhythms between the mouse medial PFC (mPFC) and two subregions of the human PFC, the anterior cingulate cortex (ACC) and the dorsolateral PFC (DLPFC).
Results: We find that while circadian rhythm signaling is conserved across species and subregions, there is a phase shift in the expression of core clock genes between the mouse mPFC and human PFC subregions that differs by sex. Furthermore, we find that the identity of rhythmic transcripts is largely unique between the mouse mPFC and human PFC subregions, with the most overlap (20%, 236 transcripts) between the mouse mPFC and the human ACC in females. Nevertheless, we find that basic biological processes are enriched for rhythmic transcripts across species, with key differences between regions and sexes.
Discussion: Together, this work highlights both the evolutionary conservation of transcriptomic rhythms and the advancement of the human PFC, underscoring the importance of considering cross-species differences when using animal models.
1 Introduction
The human prefrontal cortex (PFC) is a complex structure associated with many functions, including higher order cognitive function, goal directed behavior, and emotional regulation (Fuster, 2001). Two regions of particular interest are the dorsolateral prefrontal cortex (DLPFC), which is associated with cognition and plays a critical role in working memory, and the anterior cingulate cortex (ACC), which is implicated in both emotional and cognitive functions (Goldman-Rakic, 1995; Fuster, 2001; Yücel et al., 2003). Notably, alterations in the DLPFC and the ACC have been linked to psychiatric diseases such as schizophrenia and major depressive disorder (MDD; Haznedar et al., 1997; Yücel et al., 2003; Glausier and Lewis, 2018).
To investigate the role of the PFC in the pathophysiology of psychiatric diseases, many studies utilize mouse models. However, there is widespread debate about the cross-species homology between the human and mouse PFC (Carlén, 2017). Indeed, while similar functions are attributed to both the mouse medial PFC (mPFC) and the human DLPFC, such as delay period activity during a working memory task (Liu et al., 2014), the cytoarchitecture of the mouse mPFC most closely resembles that of the human ACC. Specifically, both the mouse mPFC and the human ACC lack a granular layer 4 (Carlén, 2017). Likewise, studies have suggested a role of the rodent mPFC in conflict monitoring, reminiscent of findings in the human ACC (Botvinick et al., 2004; Bissonette and Roesch, 2015). Therefore, it remains unlikely that the mouse mPFC fully encapsulates the functions of a single human PFC subregion; instead, it likely represents a spectrum of features associated with different subregions of the highly diversified human PFC.
Circadian rhythms are ~24-h rhythms present in biological processes. These rhythms are produced through the molecular clock, a transcriptional-translational feedback loop (TTFL) consisting of CLOCK and BMAL1, which dimerize as proteins and promote the transcription of the Per and Cry genes. After translation, the PER and CRY proteins then feedback and inhibit the activity of the CLOCK and BMAL1 proteins, creating an ~24 h cycle of gene expression (Buhr and Takahashi, 2013). Beyond the TTFL, the proteins encoded by core clock genes bind to E-box elements, generating rhythms in the expression of up to 80% of protein coding genes (Oishi et al., 2003; Mure et al., 2018). Interestingly, polymorphisms in core clock genes, as well as changes in rhythms, have been linked to multiple psychiatric diseases and are thought to represent a key feature of disease pathophysiology (McClung, 2007; McCarthy and Welsh, 2012; Johansson et al., 2016). Indeed, recent studies using human postmortem tissue have shown that there are broad changes in transcriptomic rhythms in the PFC of people with psychiatric diseases, including dampened rhythms in both the DLPFC and ACC of individuals with MDD and widespread circadian reprogramming in the DLPFC in schizophrenia (Li et al., 2013; Seney et al., 2019). As daily rhythms are present in functions associated with the PFC (Woodruff et al., 2018; Munnilari et al., 2023), alterations in transcriptomic rhythms may contribute to broad PFC dysfunction in psychiatric disease.
In this study, we use RNA sequencing to compare rhythms in the transcriptome of the mouse mPFC to two psychiatric disease relevant subregions of the human PFC, the DLPFC and the ACC. Our results indicate that rhythms in core molecular clock components and circadian rhythm signaling are broadly conserved. We additionally uncover species, sex, and subregion differences in the identity and associated biological functions of rhythmic transcripts.
2 Methods
2.1 Mouse tissue collection and RNA extraction
Age-matched (~12–14 weeks) adult male and female C57BL/6J (Jax ID: 000664) mice were group housed with mice of the same sex under a 12:12 light–dark cycle [lights on 0700, zeitgeber time (ZT0), lights off 1900 (ZT12)] for at least 2 weeks prior to sacrifice. Mice had access to food and water ad libitum and all experiments were performed in compliance with University of Pittsburgh Institutional Animal Care and Use Committee guidelines. The number of animals used was determined by a previous study modeling the effects of experimental design and sample size on rhythm detection (Zong et al., 2023).
Mice were sacrificed via cervical dislocation at 4-h intervals across 24 h (ZT2, 6, 10, 14, 18, 22); brains were removed and immediately placed on dry ice. Serial 150 μm coronal sections were cut on a cryostat (Leica Biosystems, Wetzlar, Germany) and tissue punches (1 mm, centered between hemispheres) of the mPFC were taken. The mPFC (containing the anterior cingulate and prelimbic cortices) was visually identified using the Allen Mouse Brain Coronal Atlas (Allen Brain Institute, Seattle, WA, USA) as a reference. RNA was extracted from mPFC tissue punches using a RNeasy Plus Micro kit (Qiagen, Hilden, Germany).
2.2 Mouse sequencing and data processing
Samples were assessed for concentration (Qubit, Thermo Fisher Scientific, Waltham, MA, USA) and RNA integrity (average: 9.1, standard deviation: 0.48; Agilent RNA 6000 Kit, Agilent, Santa Clara, CA, USA). Library preparation was performed using a SMART Stranded Total RNA kit (Takara, Kusatsu, Japan) and samples were sequenced (2x101bp; 40 million reads/sample) using a Nova-Seq S4 (Illumina, San Diego, CA, USA). One sample (female ZT10) was excluded for failure to generate a library, leaving 4–5 mice/sex/timepoint for downstream analysis (59 mice total). Samples were evaluated for read quality using FASTQC and the per base sequence quality was high (average > 34). Reads were aligned to the mouse reference genome (Mus musculus Ensembl GRCm38), converted to expression count data (HTSeq), and transformed to log2CPM (CPM = counts per million). Information on read counts can be found in Supplementary Table S1. Counts were filtered with additional mouse mPFC sequencing samples and transcripts that did not meet a criterion of log2CPM > 1 in >50% of the samples in at least one group, as well as genes on the Y-chromosome, were removed. Raw counts of the 13,102 transcripts that met the filtering criteria were normalized by the median of ratios method in DESeq2 and log2 transformed (Love et al., 2014). To identify transcriptomic rhythms, we utilized a parametric cosinor model (Cornelissen, 2014), whereby gene expression over a 24-h period was fitted to a sinusoidal curve. R2 values were calculated as a measure of goodness of fit. p-values and FDR-corrected q-values were calculated from the F-test and used to determine rhythmicity.
2.3 Human samples
Sequencing data for the human samples were obtained from the CommonMind Consortium. All subjects had a known time of death, were <65 years old, and had a postmortem interval (PMI) of <35 h. This cohort has previously been described in detail (Logan et al., 2022). Briefly, subjects were matched between sexes within subregion for age, PMI, RNA integrity, and cause of death. There were no significant differences in these variables between sexes. In total, 84 subjects (42 per sex) were used for analysis of the DLPFC, and 76 subjects (38 per sex) were used for analysis of the ACC. Transcripts were filtered based on the criteria outlined above and Y-chromosome and unidentified transcripts were removed, leaving 15,239 transcripts for downstream analysis. Rhythmicity analysis was performed as described in (Logan et al., 2022), with subject time of death normalized to sunrise.
2.4 Comparison of rhythms and downstream analysis
Due to the exploratory nature of this study and limited sample sizes, transcripts with a p < 0.05 were considered rhythmic and were used for downstream analysis. This is consistent with previous studies examining transcriptomic rhythms in the human and mouse brain (Li et al., 2013; Chen et al., 2016; Seney et al., 2019; Petersen et al., 2024). We also utilized a threshold-free approach (rank-rank hypergeometric overlap, abbreviated as RRHO) to assess differences in rhythmicity across groups (Cahill et al., 2018). Here, transcripts are ordered along the axes by -log10(p-value) and a heatmap is generated to visualize the overlap of rhythmic transcripts. To compare the phase shift in the timing of conserved canonical circadian genes (PER1-3, DBP, CIART, and NR1D1) across species by sex, differences in the peak times of these transcripts between the human PFC subregions and the mouse PFC were pooled within sex and compared across sexes using a random effects model in the metafor package (R software). Ingenuity Pathway Analysis (IPA; Qiagen) was used to assess functional pathways associated with rhythmic transcripts (p < 0.05); a user-supplied background list of all transcripts meeting our filtering criteria was used for each analysis. Pathways that contained fewer than 15 genes were excluded. To compare between groups, the top 10 pathways enriched for rhythmic transcripts in each group were used to assess significant enrichment in the opposing group. Pathways were considered enriched with a p < 0.05 (−log10p-value>1.3).
3 Results
3.1 Transcripts in the mouse mPFC show diurnal rhythms in expression
Similar to a previous study which found that ~10% of transcripts in the mouse PFC display diurnal rhythms in their expression (Yang et al., 2007), we find that ~12% of transcripts are rhythmic in the mouse mPFC (1,521 transcripts; p < 0.05 cutoff; Figure 1A; Supplementary Table S2). The top rhythmic transcripts, determined by p-value, include the core clock component Arntl, which encodes BMAL1, as well as transcripts such as Ciart, Dbp, and Tef, which have previously been shown to have conserved rhythmicity across tissues (Yan et al., 2008; Mure et al., 2018; Figure 1B). Scatterplots showing the expression of the top two rhythmic transcripts over a 24-h period, along with the fitted sinusoidal curve, are displayed in Figure 1C. Using an additional cohort of mice, RNAscope was used to confirm diurnal rhythms in the core clock gene Per2 (Supplementary materials and methods; Supplementary Figure S1).
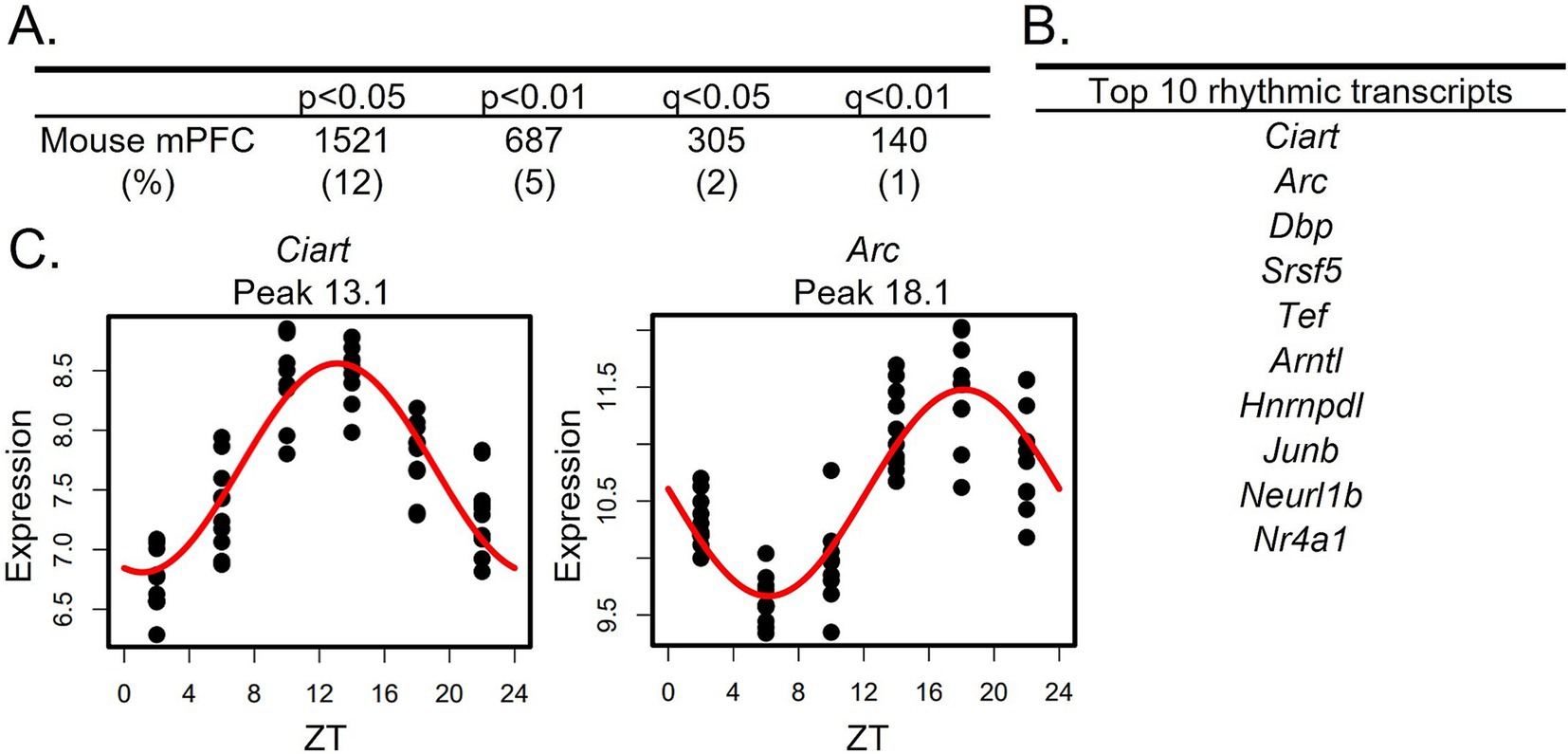
Figure 1. Rhythms in the mouse mPFC transcriptome. (A) The number (and percentage) of rhythmic transcripts detected in the mouse mPFC at different significance cutoffs. At a cutoff of p < 0.05, 12% of transcripts in the mPFC are rhythmic. (B) The top 10 rhythmic transcripts in the mouse mPFC, as determined by p-value. Top rhythmic transcripts include known circadian genes including the core molecular clock component Arntl. (C) Scatterplots depicting rhythmic expression across 24 h of the top two rhythmic transcripts. Time of death is depicted on the X-axis while expression is depicted on the Y-axis. Each point represents one subject. n = 9-10/timepoint. mPFC = medial prefrontal cortex, ZT = zeitgeber time.
3.2 Rhythms in transcripts associated with the molecular clock are conserved across species
To compare transcriptomic rhythms between the mouse mPFC and human PFC subregions, we utilized a cohort containing data from the DLPFC and the ACC that has been previously described (Logan et al., 2022). As this study found extensive sex differences in rhythmicity within the DLPFC and ACC, we performed our cross-species analysis stratified by sex. There are eight transcripts with conserved rhythms in the mouse mPFC and human PFC subregions of both sexes: CHRM4, CIART, DBP, KANSL3, NR1D1, PER1, PER2, and PER3. The majority of these transcripts (6/8) are associated with the molecular clock either directly (PER1-3) or through auxiliary loops that help to regulate the TTFL (CIART, DBP, and NR1D1; Buhr and Takahashi, 2013; Anafi et al., 2014; Figure 2). This is consistent with previous studies showing that rhythms in transcripts associated with the molecular clock are conserved between mice and humans in the brain (Li et al., 2013).
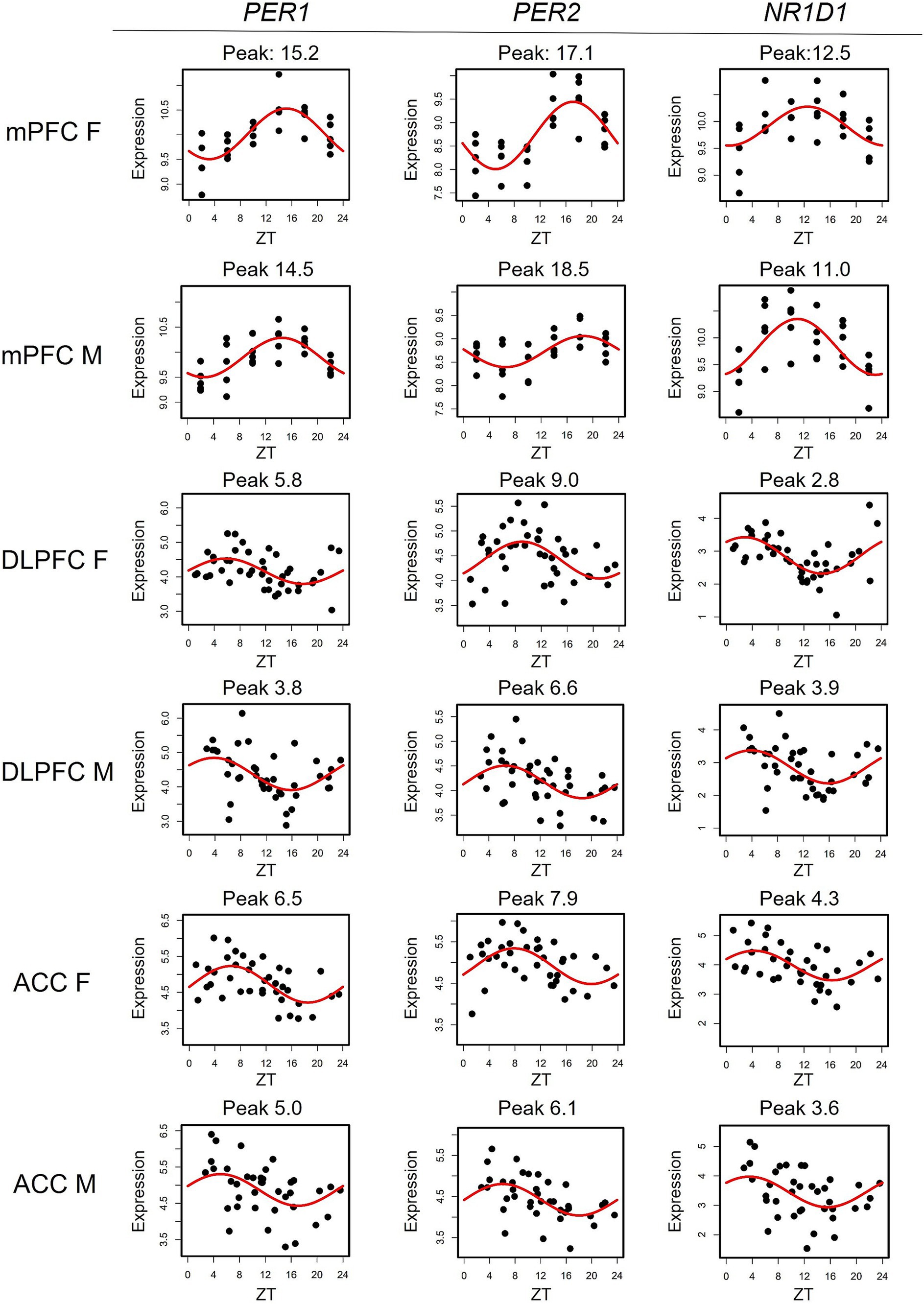
Figure 2. Rhythms in canonical circadian transcripts are conserved across species. Scatterplots depicting the expression of canonical circadian transcripts PER1, PER2, and NR1D1 in the mouse mPFC and both human PFC subregions, separated by sex. Time of death is depicted on the X-axis while expression is depicted on the Y-axis. Each point represents one subject. mPFC = medial prefrontal cortex, DLPFC = dorsolateral prefrontal cortex, ACC = anterior cingulate cortex, ZT = zeitgeber time.
3.3 Peak times of canonical circadian transcripts differ across species by sex
We next determined how canonical circadian transcripts differ in their timing between the mouse mPFC and human PFC subregions (Table 1). We find a phase shift across species that differs by sex, with known circadian transcripts largely peaking ~12 h apart between the mouse mPFC and human PFC subregions in males. However, the difference in the peak time of canonical circadian transcripts across species is significantly smaller (p = 0.002) in females, with most transcripts peaking only ~9 h apart. Moreover, previous studies have shown that in both the mouse suprachiasmatic nucleus (SCN) and the human PFC, PER1 peaks first, followed by PER3, and then PER2 (Takumi et al., 1998; Li et al., 2013), a pattern that we also observe across species in males. However, this temporal sequence is not observed in females in either the mouse mPFC or the human ACC. Therefore, while canonical circadian transcripts peak in opposing phases in the mouse mPFC and human PFC subregions, a finding that likely reflects differences in the active phase of each species, we also find sex differences in the timing of core clock components.
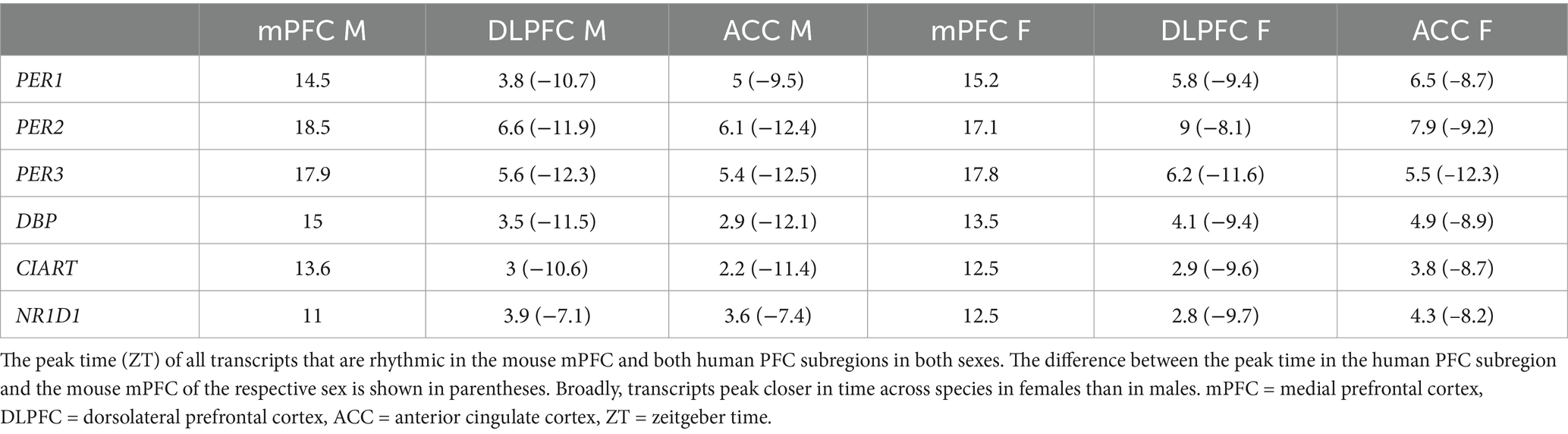
Table 1. Peak times of conserved transcripts differ across species by sex.
3.4 Largest overlap in the rhythmic transcriptome between the mouse mPFC and ACC in females
To examine transcriptome-wide similarities, or differences, in rhythms between the mouse mPFC and human PFC subregions, we next assessed the degree of overlap in rhythmic transcripts across species. When comparing rhythmic transcripts (p < 0.05) from each human PFC subregion to the mouse mPFC, we find that ~5–10% of rhythmic transcripts are shared across species (Figure 3A), except for between the mouse mPFC and the ACC in females. Here, 20% (236 transcripts) of rhythmic transcripts in the mouse mPFC are also rhythmic in the human ACC. Notably, this overlap is not reciprocal, as only 8% of rhythmic transcripts in the human ACC are also rhythmic in the mouse mPFC in females. Using a more stringent significance threshold of p < 0.01, we confirm that the greatest number of rhythmic transcripts (21 transcripts) overlaps between the mouse mPFC and human ACC in females (Supplementary Figure S2).
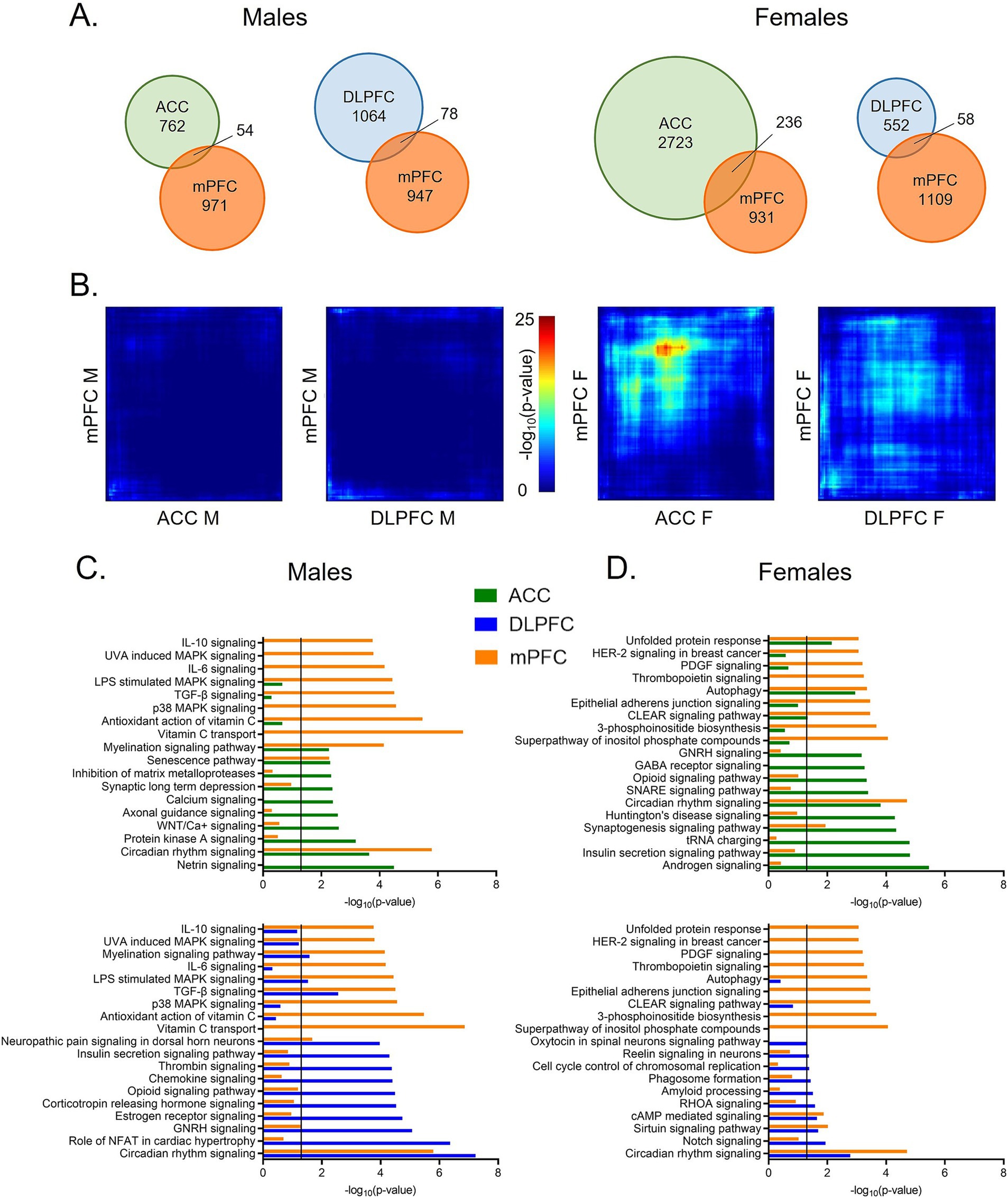
Figure 3. Greatest overlap in rhythmic transcripts between the mPFC and ACC in females. (A) Venn diagrams depicting the overlap in rhythmic transcripts (p < 0.05) between the mouse mPFC and human PFC subregions, separated by sex. Twenty percent of rhythmic transcripts in the mouse mPFC are also rhythmic in the human ACC in females. All other comparisons across species share ~5–10% of rhythmic transcripts. (B) Rank-rank hypergeometric overlap plots visualizing the overlap in rhythmic transcripts between the mouse mPFC and human PFC subregions. This threshold-free approach indicates that there is the most overlap in rhythmic transcripts between the mouse mPFC and the human ACC in females. (C) Ingenuity Pathway Analysis (IPA) was used to determine the top 10 pathways enriched for rhythmic transcripts (p < 0.05) and their overlap between the mouse mPFC and human PFC subregions in males. (D) The top 10 pathways enriched for rhythmic transcripts (p < 0.05), determined by IPA, and their overlap between the mouse mPFC and human PFC subregions in females. While the overlap in biological processes associated with rhythmic transcripts differs by region and sex, circadian rhythm signaling is among the top 10 pathways in the mouse mPFC and the human PFC subregions of both sexes. mPFC = medial prefrontal cortex, DLPFC = dorsolateral prefrontal cortex, ACC = anterior cingulate cortex.
We next used rank-rank hypergeometric overlap (RRHO) plots as a threshold-free approach. Once more, we find that among all groups, there is the most overlap in rhythmic transcripts between the human ACC and the mouse mPFC from female subjects (Figure 3B). Rhythmic transcripts in the DLPFC of female subjects and the mPFC of female mice also show slight overlap, although less than that of the ACC. Notably, the RRHO analysis only uses transcripts that are expressed in both the mouse mPFC and human PFC subregions, suggesting that there are broad cross-species differences in rhythmicity even among the same subset of transcripts.
3.5 Conservation of circadian rhythm signaling
Although the identity of rhythmic transcripts is largely distinct between the mouse mPFC and human PFC subregions, we next determined if the functions of rhythmic transcripts are conserved. Using Ingenuity Pathway Analysis (IPA), we find that circadian rhythm signaling is found among the top 10 enriched pathways in all sexes, species, and subregions (Figures 3C,D).
In males, two additional pathways (senescence signaling and myelination signaling) are enriched for rhythmic transcripts in both the human ACC and the mouse mPFC (Figure 3C-top). Myelination signaling is also enriched for rhythmic transcripts in the DLPFC in males, suggesting a sex-specific conservation of rhythms in this process (Figure 3C-bottom). Additional pathways enriched in both the human DLPFC and the mouse mPFC of males include those associated with pain (neuropathic pain signaling in dorsal horn neurons), cytokine signaling (TGF-β signaling), and mitogen-activated protein (MAP) kinase signaling (LPS stimulated MAPK signaling). Many transcripts belonging to the MAP kinase family are also found within the enriched gonadotropin-releasing hormone signaling pathway. This suggests a conserved role of rhythms in intercellular signal transduction between the mouse mPFC and the human DLPFC in males.
We next performed the same analysis in females (Figure 3D). In addition to circadian rhythm signaling, four additional pathways are significantly enriched for rhythmic transcripts in both the human ACC and the mouse mPFC (Figure 3D-top). These pathways are associated with synaptogenesis, protein folding (unfolded protein response), autophagy, and lysosomal degradation (CLEAR signaling). When pathways enriched for rhythmic transcripts in the DLPFC and the mouse mPFC of female subjects are compared, we find less overlap in enriched pathways, consistent with our finding of less overlap in the identity of rhythmic transcripts (Figure 3D-bottom). In addition to circadian rhythm signaling, there are two pathways with significant overlap between the DLPFC and mPFC in females: sirtuin signaling and cAMP-mediated signaling. Many of the rhythmic transcripts that belong to the cAMP-mediated signaling pathway are G-protein coupled receptors (GPCRs), although the type of receptor differs between the human DLPFC and the mouse mPFC. Together, this indicates that although rhythmic transcripts differ in their identity, key biological processes, including neuronal signaling and protein processing, are enriched for rhythmic transcripts across species in females. However, in both sexes, over half of the enriched pathways are distinct between the mouse mPFC and human PFC subregions.
We next performed a cross species analysis of rhythms in the mouse mPFC and an evolutionarily intermediate species, using previously published data from male baboons (Mure et al., 2018). While there are methodological differences between studies, we find that when compared to mice of the same sex, ~20% of rhythmic transcripts in the mouse mPFC are also rhythmic in the baboon PFC (Supplementary Figure S3A). This is more than twice the proportion of rhythmic transcripts that are shared between the mouse mPFC and human PFC subregions in males. Moreover, there is greater overlap in the enriched biological processes (9 pathways) between the baboon PFC and mouse mPFC than between any of the human PFC subregions and the mouse mPFC (Supplementary Figure S3B). This suggests that there is greater conservation of transcriptomic rhythms between the mouse and baboon PFC than between the mouse and human PFC.
3.6 Temporal patterns of rhythmic gene expression differ by species and sex
To examine overall patterns of rhythmic transcript expression across species, the percentage of rhythmic transcripts peaking at each timepoint (ZT) was plotted across 24 h in two-hour bins. In males, nearly 60% of all rhythmic transcripts in the mouse mPFC peak during the active (dark) phase, with peak times fairly evenly distributed across the phase (Figure 4A). Similarly, in both human PFC subregions, the majority of rhythmic transcripts peak in the active (light) phase, with ~68%, or over 80%, of rhythmic transcripts peaking during the active (light) phase in the human ACC and DLPFC, respectively.
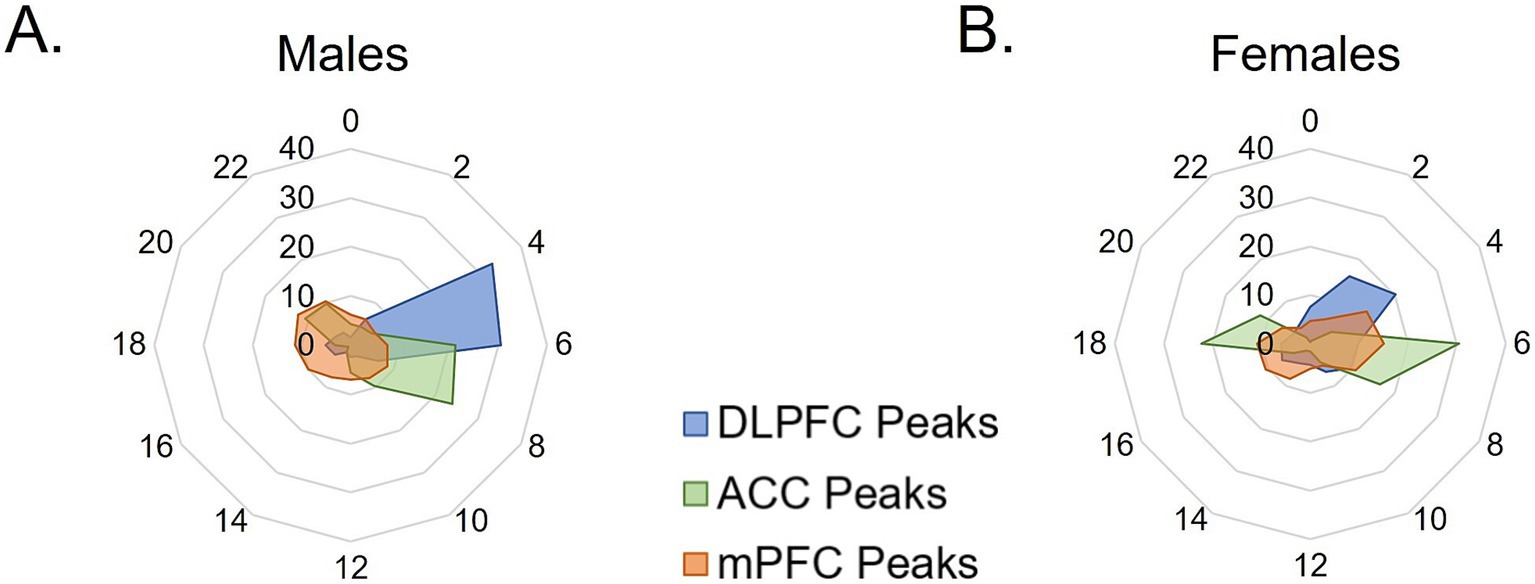
Figure 4. Temporal patterns of rhythmic expression vary by region, sex, and species. (A,B) The peak time of rhythmic transcripts (p < 0.05), plotted as the percentage of total rhythmic transcripts peaking in each 2-h bin across 24 h (ZT) in (A) males and (B) females. In males, rhythmic transcripts largely peak in the opposite phase between the mouse mPFC and human PFC subregions. In females, over half of rhythmic transcripts peak in the active phase (light) in humans, whereas about half of transcripts peak in each phase in the mouse mPFC. mPFC = medial prefrontal cortex, DLPFC = dorsolateral prefrontal cortex, ACC = anterior cingulate cortex, ZT = zeitgeber time.
In females, rhythmic transcripts in the mouse mPFC largely fall into two groups, with approximately half peaking in each phase (Figure 4B). Similarly, large groups of rhythmic transcripts in the human ACC peak in each phase, with ~57% peaking in the active (light) phase and ~ 43% peaking in the inactive (dark) phase. In contrast, in the human DLPFC of female subjects, most (~70%) of the rhythmic transcripts peak during the active (light) phase. These data demonstrate that although the expression of core clock genes can be predicted by the active phase of the species, the temporal patterns of total rhythmic transcripts are highly variable between species, PFC subregions, and even sexes.
4 Discussion
Recent literature has highlighted the importance of transcriptomic rhythms in brain health and disease (Li et al., 2013; Seney et al., 2019; Logan et al., 2022). In this study, we compared rhythms between the mouse mPFC and two subregions of the human PFC, the ACC and the DLPFC, that are heavily implicated in psychiatric disorders (Haznedar et al., 1997; Yücel et al., 2003; Glausier and Lewis, 2018). Consistent with previous studies, we find that canonical circadian genes are rhythmic in both humans and mice (Yan et al., 2008; Li et al., 2013). These conserved rhythmic transcripts largely peak in opposing phases in mice and humans, likely reflecting differences in the active phase of each species. However, we find that the difference in timing of core circadian genes between species depends on sex, with females showing a smaller shift than males (~9 h vs. ~12 h). This may be driven by sex differences in the rhythms of core clock genes within species. Indeed, previous studies found that the peak time of core clock gene expression differs by sex in the DLPFC of elderly subjects (Lim et al., 2013), whereas studies in the rodent PFC found that rhythms in core clock genes were more robust in males (Chun et al., 2015). While the mechanisms underlying sex differences in transcriptomic rhythms are currently unknown, we hypothesize that circulating hormones may have an effect. Indeed, the SCN expresses both estrogen and androgen receptors (Karatsoreos et al., 2007; Hatcher et al., 2020) and systemic estradiol administration has been shown to phase advance core clock gene expression in the SCN (Nakamura et al., 2005). Moreover, one study found that in the PFC, rhythms in Arntl differed between rats with a normal estrous cycle and non-cycling rats, while rhythms in Per1 and Per2 were unaffected (Chun et al., 2015). Although data on the estrous phase/menstrual cycle and menopausal status was not assessed/available in this study, the proportion of rhythmic transcripts is not consistently lower in female subjects. Therefore, potential variability due to changes in the levels of sex hormones across the estrous/menstrual cycle and menopause likely does not impair our ability to detect rhythmic transcripts. Nevertheless, the role of sex hormones on age-related changes in rhythmic gene expression remains an important area of future research.
While most rhythmic transcripts that are broadly conserved across species and sexes are closely associated with the molecular clock, rhythms in two transcripts, KANSL3 and CHRM4, are also broadly conserved. KANSL3, which is involved in chromatin remodeling, plays a role in regulating the transcription of housekeeping genes and facilitates the transcription of mitochondrial DNA in cells with high metabolic rates, such as neurons (Chatterjee et al., 2016; Sheikh et al., 2019). The conserved rhythmicity of this transcript suggests that molecular clock control over the transcription of housekeeping genes and mitochondrial function are evolutionarily conserved. On the other hand, CHRM4 encodes the muscarinic acetylcholine receptor M4, which has been proposed as a therapeutic target for schizophrenia (Gibbons and Dean, 2016; Gould et al., 2018), Therefore, conservation of rhythms in CHRM4 across species may be important for successful translation of these drugs into humans.
Nevertheless, we find that most of the rhythmic transcripts in the human PFC subregions are not rhythmic in the mouse mPFC of the same sex. Indeed, while 20% of rhythmic transcripts in the mouse mPFC are rhythmic in the ACC in females, only 8% of rhythmic transcripts in the human ACC are rhythmic in the mouse mPFC. This suggests that rhythms in gene expression changed as the human PFC evolved and became more specialized. Our findings are consistent with theories of PFC evolution, whereby agranular and dysgranular regions, such as the ACC, evolved earlier, while granular regions of the PFC, such as the DLPFC, evolved later and are considered to be unique to primates (Preuss and Wise, 2022). The idea that molecular rhythms diverged across evolution is further supported by our finding that there is greater overlap in rhythmic transcripts between the baboon PFC and the mouse mPFC, as well as findings from a recent paper showing that up to 38% of rhythmic transcripts are shared between mice and humans in the more evolutionarily conserved striatum (Petersen et al., 2024). Notably, however, we find that the elevated overlap between the mouse mPFC and the human ACC is specific to females, a finding perhaps driven by previously described sex differences in rhythmicity in the human ACC (Logan et al., 2022).
When the biological processes associated with rhythmic transcripts are assessed, we find that pathways associated with intercellular communication and the integration of extracellular signals are significantly enriched for rhythmic transcripts in both the mouse mPFC and human PFC subregions. The enrichment of rhythmic transcripts in these pathways suggests that control of the molecular clock over mechanisms associated with cellular signaling, including neurotransmission, are broadly conserved. Moreover, while the rhythmic transcripts belonging to each pathway are generally different between mouse and human, some are closely related. This is particularly true within the unfolded protein response pathway, whereby many rhythmic transcripts in both the human ACC and the mouse mPFC encode proteins in the DNAJ heat shock family. Of note, while this study and many others have focused on rhythms in the transcriptome, recent advances have made the measurement of rhythms in the proteome possible (Robles et al., 2014; Wang et al., 2018; Brüning et al., 2019; Noya et al., 2019). Therefore, future studies examining the relationship between transcriptomic rhythms and proteomic rhythms in the mouse and human PFC will provide further insight into the role of circadian rhythms on physiological processes in the brain.
Similar to previous studies, which found that the timing of total rhythmic transcripts varies widely even in anatomically adjacent tissues (Mure et al., 2018), we find broad differences in the temporal patterns of rhythmic gene expression across PFC subregions, sexes, and species. These patterns are much more variable than the timing of core clock genes, suggesting that they are generated downstream of the molecular clock. Indeed, studies have found that many targets of the molecular clock are transcription factors, resulting in rhythms in gene expression that are tissue specific (Miller et al., 2007). It is likely that a similar mechanism underlies the differences found in this study. Understanding temporal patterns in gene expression, and what drives them, may have clinical implications for psychiatry. For example, a recent study found that the time in which antipsychotics were administered affected the development of metabolic side effects in both humans and mice (Zapata et al., 2022). However, the timing depended on the active phase of each species, highlighting the importance of considering cross-species differences in rhythms when translating preclinical findings into humans.
Differences in rhythms between the mouse mPFC and human PFC subregions may be partially attributable to cross-species differences in the cellular makeup of these regions. Indeed, the expression of markers used to define individual cell types differs across species in the cortex and studies have shown that, even within conserved cell types, gene expression differs between mice and humans (Zeng et al., 2012; Hodge et al., 2019). Moreover, differences in the timing of transcripts across cell types may make transcripts appear non-rhythmic in homogenate tissue, as one study in the mouse SCN found that core clock genes peak earlier in neurons relative to non-neuronal cells (Wen et al., 2020). It remains unknown whether similar differences in timing exist across cell types in the PFC and whether these differences are conserved across species.
Variation in the light–dark cycle may also contribute to the observed differences in rhythmicity between the mouse mPFC and human PFC subregions. While mice in this study were housed under a strict 12:12 light–dark cycle, the light–dark cycle of the human subjects was likely more variable. While the time of death of human subjects is normalized to sunrise, we cannot eliminate the effect of artificial light or differences in behavioral rhythms. Therefore, it is likely that increased variability in human samples may result in an underestimation of true rhythmic transcripts and/or lower amplitude rhythms. Due to this variability, consistent with previous studies (Li et al., 2013; Chen et al., 2016; Seney et al., 2019; Petersen et al., 2024), we utilized a less stringent p-value for this exploratory analysis. While our percentage of rhythmic transcripts in the mouse mPFC is similar to what has been described in previous studies (Yang et al., 2007), we would undoubtedly identify additional rhythmic transcripts in humans with higher sample sizes and more statistical power.
As over 80% of proteins identified as druggable targets by the FDA show rhythms in gene expression (Mure et al., 2018), this study provides an important translational framework for understanding how these rhythms differ across species. These findings paint a complex picture, whereby molecular rhythms show distinct patterns based on sex, species, and PFC subregion, likely reflecting the unique functions of rhythmicity in the highly specialized human PFC. Given that patterns of rhythmic gene expression show extensive changes in the brains of individuals with psychiatric diseases, it is imperative to carefully consider differences in rhythms between species when mice are used for mechanistic studies into these disorders.
Data availability statement
Rhythmic transcripts in the mouse mPFC and human PFC subregions are listed in Supplementary Tables S2–S8. Sequencing data from the mouse mPFC has been shared to the Gene Expression Omnibus (GSE284053) and sequencing data from the human samples can be obtained from the CommonMind Consortium (RRID:SCR_000139).
Ethics statement
The studies involving humans were approved by the Committee for Oversight of Research and Clinical Training Involving Decedents. The studies were conducted in accordance with the local legislation and institutional requirements. The human samples used in this study were acquired from a previous study for which ethical approval was obtained. Written informed consent for participation was not required from the participants or the participants’ legal guardians/next of kin in accordance with the national legislation and institutional requirements. The animal study was approved by University of Pittsburgh IACUC. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
JB: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing. AJ: Data curation, Formal analysis, Investigation, Methodology, Writing – review & editing. XX: Data curation, Formal analysis, Investigation, Methodology, Writing – original draft. KP: Data curation, Formal analysis, Methodology, Validation, Writing – review & editing. KK: Formal analysis, Methodology, Supervision, Writing – review & editing. MP: Investigation, Validation, Writing – review & editing. CV: Investigation, Methodology, Validation, Writing – review & editing. MSc: Data curation, Formal analysis, Investigation, Methodology, Writing – review & editing. MSe: Formal analysis, Methodology, Supervision, Writing – review & editing. GT: Formal analysis, Resources, Software, Supervision, Writing – review & editing. CM: Conceptualization, Funding acquisition, Methodology, Project administration, Resources, Supervision, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This work was funded by: National Institute of Mental Health (NIMH) (MH106460), National Institute of Neurological Disorders and Stroke (NS127064), National Institute on Drug Abuse (NIDA) (DA039865, NIDA DA046346, and NIMH MH111601), and the Wood Next Foundation to CAM.
Acknowledgments
We would like to thank the CommonMind Consortium for the data from the human postmortem tissue. We would also like to thank the University of Pittsburgh Health Sciences Sequencing Core and UPMC Genome Center for assistance with library preparation and RNA-sequencing.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnins.2024.1524615/full#supplementary-material
References
Anafi, R. C., Lee, Y., Sato, T. K., Venkataraman, A., Ramanathan, C., Kavakli, I. H., et al. (2014). Machine learning helps identify CHRONO as a circadian clock component. PLoS Biol. 12:e1001840. doi: 10.1371/journal.pbio.1001840
Bissonette, G. B., and Roesch, M. R. (2015). Neural correlates of rules and conflict in medial prefrontal cortex during decision and feedback epochs. Front. Behav. Neurosci. 9:266. doi: 10.3389/fnbeh.2015.00266
Botvinick, M. M., Cohen, J. D., and Carter, C. S. (2004). Conflict monitoring and anterior cingulate cortex: an update. Trends Cogn. Sci. 8, 539–546. doi: 10.1016/j.tics.2004.10.003
Brüning, F., Noya, S. B., Bange, T., Koutsouli, S., Rudolph, J. D., Tyagarajan, S. K., et al. (2019). Sleep-wake cycles drive daily dynamics of synaptic phosphorylation. Science 366:eaav3617. doi: 10.1126/science.aav3617
Buhr, E. D., and Takahashi, J. S. (2013). Molecular components of the mammalian circadian clock. Handb. Exp. Pharmacol. 217, 3–27. doi: 10.1007/978-3-642-25950-0_1
Cahill, K. M., Huo, Z., Tseng, G. C., Logan, R. W., and Seney, M. L. (2018). Improved identification of concordant and discordant gene expression signatures using an updated rank-rank hypergeometric overlap approach. Sci. Rep. 8:9588. doi: 10.1038/s41598-018-27903-2
Carlén, M. (2017). What constitutes the prefrontal cortex? Science 358, 478–482. doi: 10.1126/science.aan8868
Chatterjee, A., Seyfferth, J., Lucci, J., Gilsbach, R., Preissl, S., Böttinger, L., et al. (2016). MOF acetyl transferase regulates transcription and respiration in mitochondria. Cell 167, 722–738.e23. doi: 10.1016/j.cell.2016.09.052
Chen, C.-Y., Logan, R. W., Ma, T., Lewis, D. A., Tseng, G. C., Sibille, E., et al. (2016). Effects of aging on circadian patterns of gene expression in the human prefrontal cortex. Proc. Natl. Acad. Sci. 113, 206–211. doi: 10.1073/pnas.1508249112
Chun, L. E., Woodruff, E. R., Morton, S., Hinds, L. R., and Spencer, R. L. (2015). Variations in phase and amplitude of rhythmic clock gene expression across prefrontal cortex, Hippocampus, amygdala, and hypothalamic paraventricular and suprachiasmatic nuclei of male and female rats. J. Biol. Rhythm. 30, 417–436. doi: 10.1177/0748730415598608
Cornelissen, G. (2014). Cosinor-based rhythmometry. Theor. Biol. Med. Model. 11:16. doi: 10.1186/1742-4682-11-16
Fuster, J. M. (2001). The prefrontal cortex—an update: time is of the essence. Neuron 30, 319–333. doi: 10.1016/S0896-6273(01)00285-9
Gibbons, A., and Dean, B. (2016). The cholinergic system: an emerging drug target for schizophrenia. Curr. Pharm. Des. 22, 2124–2133. doi: 10.2174/1381612822666160127114010
Glausier, J. R., and Lewis, D. A. (2018). “Chapter 25 - mapping pathologic circuitry in schizophrenia” in Handbook of clinical neurology. eds. I. Huitinga and M. J. Webster (Elsevier: The Handbook of Clinical Neurology), 389–417.
Goldman-Rakic, P. S. (1995). Cellular basis of working memory. Neuron 14, 477–485. doi: 10.1016/0896-6273(95)90304-6
Gould, R. W., Grannan, M. D., Gunter, B. W., Ball, J., Bubser, M., Bridges, T. M., et al. (2018). Cognitive enhancement and antipsychotic-like activity following repeated dosing with the selective M(4) PAM VU0467154. Neuropharmacology 128, 492–502. doi: 10.1016/j.neuropharm.2017.07.013
Hatcher, K. M., Royston, S. E., and Mahoney, M. M. (2020). Modulation of circadian rhythms through estrogen receptor signaling. Eur. J. Neurosci. 51, 217–228. doi: 10.1111/ejn.14184
Haznedar, M. M., Buchsbaum, M. S., Luu, C., Hazlett, E. A., Siegel, B. V. J., Lohr, J., et al. (1997). Decreased anterior cingulate gyrus metabolic rate in schizophrenia. Am. J. Psychiatry 154, 682–684. doi: 10.1176/ajp.154.5.682
Hodge, R. D., Bakken, T. E., Miller, J. A., Smith, K. A., Barkan, E. R., Graybuck, L. T., et al. (2019). Conserved cell types with divergent features in human versus mouse cortex. Nature 573, 61–68. doi: 10.1038/s41586-019-1506-7
Johansson, A.-S., Owe-Larsson, B., Hetta, J., and Lundkvist, G. B. (2016). Altered circadian clock gene expression in patients with schizophrenia. Schizophr. Res. 174, 17–23. doi: 10.1016/j.schres.2016.04.029
Karatsoreos, I. N., Wang, A., Sasanian, J., and Silver, R. (2007). A role for androgens in regulating circadian behavior and the suprachiasmatic nucleus. Endocrinology 148, 5487–5495. doi: 10.1210/en.2007-0775
Li, J. Z., Bunney, B. G., Meng, F., Hagenauer, M. H., Walsh, D. M., Vawter, M. P., et al. (2013). Circadian patterns of gene expression in the human brain and disruption in major depressive disorder. Proc. Natl. Acad. Sci. 110, 9950–9955. doi: 10.1073/pnas.1305814110
Lim, A. S. P., Myers, A. J., Yu, L., Buchman, A. S., Duffy, J. F., De Jager, P. L., et al. (2013). Sex difference in daily rhythms of clock gene expression in the aged human cerebral cortex. J. Biol. Rhythm. 28, 117–129. doi: 10.1177/0748730413478552
Liu, D., Gu, X., Zhu, J., Zhang, X., Han, Z., Yan, W., et al. (2014). Medial prefrontal activity during delay period contributes to learning of a working memory task. Science 346, 458–463. doi: 10.1126/science.1256573
Logan, R. W., Xue, X., Ketchesin, K. D., Hoffman, G., Roussos, P., Tseng, G., et al. (2022). Sex differences in molecular rhythms in the human cortex. Biol. Psychiatry 91, 152–162. doi: 10.1016/j.biopsych.2021.03.005
Love, M. I., Huber, W., and Anders, S. (2014). Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15:550. doi: 10.1186/s13059-014-0550-8
McCarthy, M. J., and Welsh, D. K. (2012). Cellular circadian clocks in mood disorders. J. Biol. Rhythm. 27, 339–352. doi: 10.1177/0748730412456367
McClung, C. A. (2007). Circadian genes, rhythms and the biology of mood disorders. Pharmacol. Ther. 114, 222–232. doi: 10.1016/j.pharmthera.2007.02.003
Miller, B. H., McDearmon, E. L., Panda, S., Hayes, K. R., Zhang, J., Andrews, J. L., et al. (2007). Circadian and CLOCK-controlled regulation of the mouse transcriptome and cell proliferation. Proc. Natl. Acad. Sci. 104, 3342–3347. doi: 10.1073/pnas.0611724104
Munnilari, M., Bommasamudram, T., Easow, J., Tod, D., Varamenti, E., Edwards, B. J., et al. (2023). Diurnal variation in variables related to cognitive performance: a systematic review. Sleep Breath. 28, 495–510. doi: 10.1007/s11325-023-02895-0
Mure, L. S., Le, H. D., Benegiamo, G., Chang, M. W., Rios, L., Jillani, N., et al. (2018). Diurnal transcriptome atlas of a primate across major neural and peripheral tissues. Science 359:eaao0318. doi: 10.1126/science.aao0318
Nakamura, T. J., Moriya, T., Inoue, S., Shimazoe, T., Watanabe, S., Ebihara, S., et al. (2005). Estrogen differentially regulates expression of Per1 and Per2 genes between central and peripheral clocks and between reproductive and nonreproductive tissues in female rats. J. Neurosci. Res. 82, 622–630. doi: 10.1002/jnr.20677
Noya, S. B., Colameo, D., Brüning, F., Spinnler, A., Mircsof, D., Opitz, L., et al. (2019). The forebrain synaptic transcriptome is organized by clocks but its proteome is driven by sleep. Science 366:eaav2642. doi: 10.1126/science.aav2642
Oishi, K., Miyazaki, K., Kadota, K., Kikuno, R., Nagase, T., Atsumi, G., et al. (2003). Genome-wide expression analysis of mouse liver reveals CLOCK-regulated circadian output genes. J. Biol. Chem. 278, 41519–41527. doi: 10.1074/jbc.M304564200
Petersen, K. A., Zong, W., Depoy, L. M., Scott, M. R., Shankar, V. G., Burns, J. N., et al. (2024). Comparative rhythmic transcriptome profiling of human and mouse striatal subregions. Neuropsychopharmacology 49, 796–805. doi: 10.1038/s41386-023-01788-w
Preuss, T. M., and Wise, S. P. (2022). Evolution of prefrontal cortex. Neuropsychopharmacology 47, 3–19. doi: 10.1038/s41386-021-01076-5
Robles, M. S., Cox, J., and Mann, M. (2014). In-vivo quantitative proteomics reveals a key contribution of post-transcriptional mechanisms to the circadian regulation of liver metabolism. PLoS Genet. 10:e1004047. doi: 10.1371/journal.pgen.1004047
Seney, M. L., Cahill, K., Enwright, J. F. 3rd, Logan, R. W., Huo, Z., Zong, W., et al. (2019). Diurnal rhythms in gene expression in the prefrontal cortex in schizophrenia. Nat. Commun. 10:3355. doi: 10.1038/s41467-019-11335-1
Sheikh, B. N., Guhathakurta, S., and Akhtar, A. (2019). The non-specific lethal (NSL) complex at the crossroads of transcriptional control and cellular homeostasis. EMBO Rep. 20:e47630. doi: 10.15252/embr.201847630
Takumi, T., Taguchi, K., Miyake, S., Sakakida, Y., Takashima, N., Matsubara, C., et al. (1998). A light-independent oscillatory gene mPer3 in mouse SCN and OVLT. EMBO J. 17, 4753–4759. doi: 10.1093/emboj/17.16.4753
Wang, Y., Song, L., Liu, M., Ge, R., Zhou, Q., Liu, W., et al. (2018). A proteomics landscape of circadian clock in mouse liver. Nat. Commun. 9:1553. doi: 10.1038/s41467-018-03898-2
Wen, S., Ma, D., Zhao, M., Xie, L., Wu, Q., Gou, L., et al. (2020). Spatiotemporal single-cell analysis of gene expression in the mouse suprachiasmatic nucleus. Nat. Neurosci. 23, 456–467. doi: 10.1038/s41593-020-0586-x
Woodruff, E. R., Chun, L. E., Hinds, L. R., Varra, N. M., Tirado, D., Morton, S. J., et al. (2018). Coordination between prefrontal cortex clock gene expression and corticosterone contributes to enhanced conditioned fear extinction recall. eNeuro 5:2018. doi: 10.1523/ENEURO.0455-18.2018
Yan, J., Wang, H., Liu, Y., and Shao, C. (2008). Analysis of gene regulatory networks in the mammalian circadian rhythm. PLoS Comput. Biol. 4:e1000193. doi: 10.1371/journal.pcbi.1000193
Yang, S., Wang, K., Valladares, O., Hannenhalli, S., and Bucan, M. (2007). Genome-wide expression profiling and bioinformatics analysis of diurnally regulated genes in the mouse prefrontal cortex. Genome Biol. 8:R247. doi: 10.1186/gb-2007-8-11-r247
Yücel, M., Wood, S. J., Fornito, A., Riffkin, J., Velakoulis, D., and Pantelis, C. (2003). Anterior cingulate dysfunction: implications for psychiatric disorders? J. Psychiatry Neurosci. 28, 350–354
Zapata, R. C., Silver, A., Yoon, D., Chaudry, B., Libster, A., McCarthy, M. J., et al. (2022). Antipsychotic-induced weight gain and metabolic effects show diurnal dependence and are reversible with time restricted feeding. Schizophrenia 8:70. doi: 10.1038/s41537-022-00276-2
Zeng, H., Shen, E. H., Hohmann, J. G., Oh, S. W., Bernard, A., Royall, J. J., et al. (2012). Large-scale cellular-resolution gene profiling in human neocortex reveals species-specific molecular signatures. Cell 149, 483–496. doi: 10.1016/j.cell.2012.02.052
Keywords: circadian rhythms, transcriptomics, prefrontal cortex, mouse, human post mortem tissue
Citation: Burns JN, Jenkins AK, Xue X, Petersen KA, Ketchesin KD, Perez MS, Vadnie CA, Scott MR, Seney ML, Tseng GC and McClung CA (2025) Comparative transcriptomic rhythms in the mouse and human prefrontal cortex. Front. Neurosci. 18:1524615. doi: 10.3389/fnins.2024.1524615
Edited by:
Rossen Donev, MicroPharm Ltd., United KingdomReviewed by:
Diou Luo, Iowa State University, United StatesHarry Pantazopoulos, University of Mississippi Medical Center, United States
Copyright © 2025 Burns, Jenkins, Xue, Petersen, Ketchesin, Perez, Vadnie, Scott, Seney, Tseng and McClung. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Colleen A. McClung, bWNjbHVuZ2NhQHVwbWMuZWR1